
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 15 January 2025
Sec. Marine Ecosystem Ecology
Volume 11 - 2024 | https://doi.org/10.3389/fmars.2024.1459935
 Mingshan Xu1,2,3*
Mingshan Xu1,2,3* Jiali Gu1
Jiali Gu1 Peng Yun1Jianjun Gan1Wenzhou Fan1Zhentao Chen1Jie Shao1
Peng Yun1Jianjun Gan1Wenzhou Fan1Zhentao Chen1Jie Shao1 Guangzhi Zhang1
Guangzhi Zhang1 Jian Zeng1*
Jian Zeng1*Globally, native saltmarshes are declining, while invasive saltmarshes are expanding rapidly. However, the underlying processes and driving mechanisms behind these trends remain poorly understood, particularly in rapidly silting coastal bays. Here, we investigated the spatiotemporal dynamics and influencing factors of native saltmarshes (Scirpus mariqueter and Phragmites australis) and invasive Spartina alterniflora in Yueqing Bay, eastern China, by interpreting Landsat series images during 1985-2023. Results showed that native saltmarshes significantly decreased from 632.24 ha to complete disappearance, while S. alterniflora expanded from zero to 2872.90 ha in Yueqing Bay during 1985-2023. Specifically, S. mariqueter experienced a notable decline from 1985 to 1995, subsequent growth between 2000 and 2005, and another reduction from 2005 to 2010. P. australis expanded from 2005 to 2010 but shrank from 2010 to 2015 as S. alterniflora encroached upon its habitats. By contrast, S. alterniflora consistently expanded annually, except when it was eradicated by human interference. Both natural factors and human activities influenced native and invasive saltmarshes; for instance, mudflat reclamation facilitated saltmarsh expansion by creating suitable habitats during a certain period, particularly for S. alterniflora. From 1984 to 2018, the coastline extended seaward, causing sedimentation and landward transformation, which contributed to the expansion of S. alterniflora. However, the 118.05 ha mangrove forests negatively impacted S. alterniflora, as planting mangroves often involves the removal of existing S. alterniflora. Additionally, maximum tidal range and mean annual temperature positively affected saltmarshes. Our study highlighted the influence of natural factors and human activities on the spatiotemporal dynamics of both native and invasive saltmarshes in rapidly silting bays, underscoring the urgent need for conservation, restoration, and management of native saltmarshes.
● The spatiotemporal dynamics of native saltmarshes and invasive Spartina alterniflora were interpreted in a rapidly silting bay.
● These dynamics were significantly influenced by a combination of natural factors and human activities.
● Mudflat reclamation, coastline changes, mangrove forests plantation, tidal range, and temperature were key determinants of saltmarsh spatiotemporal dynamics.
● Human efforts are required to protect native saltmarshes and manage invasive species in rapidly silting coastal bays.
Coastal saltmarshes play crucial social and ecological roles, encompassing carbon sequestration, shoreline protection, sedimentation promotion, landscape enhancement, and habitat provision for diverse organisms (Leonard and Croft, 2006; Campbell et al., 2022). On both global and national scales, the significant decline of native saltmarshes and expansion of invasive saltmarshes, notably S. alterniflora, may be attributed to factors such as climate change, biological invasions, rising sea levels, land reclamation, and reduced sedimentation (Boesch and Turner, 1984; Bertness, 2006). These dynamics pose serious threats to the biodiversity and ecosystem functions of coastal wetlands (Ainouche and Gray, 2016; Qi and Chmura, 2023). Despite substantial investments worldwide to control invasive saltmarshes, these efforts have proven unsatisfactory with repeated expansion (Nie et al., 2023; Qi and Chmura, 2023). Unfortunately, insufficient investigation into the processes and underlying influencing factors contributing to the spatiotemporal dynamics of native and invasive saltmarshes has hampered efforts to effectively preserve native saltmarshes and control invasive species.
In recent decades, the spatiotemporal dynamics of native saltmarsh degradation and invasive saltmarsh expansion have been extensively studied at local, regional, national, and global scales, across various time periods, and considering both single and multiple species (Campbell et al., 2022; Chen et al., 2022; Luo et al., 2024). For instance, Laengner et al. (2019) emphasized the importance of evaluating saltmarshes dynamics and influencing factors within a spatial context. A global high-resolution map revealed a net loss of 4,000 km2 of intertidal wetlands between 1999 and 2019 (Murray et al., 2022). Additionally, Campbell et al. (2022) developed an evolutionary map illustrating the status and distribution of global saltmarshes from 2000 to 2019, indicating a net loss of 1,438 km2. However, due to the coarse resolution of publicly available environmental data, quantitative studies on the driving factors behind these dynamics are often conducted at large scales. Local studies typically provide qualitative analyses of driving factors or primarily focus on land use changes (Luo et al., 2024; Yin et al., 2024). To gain a deeper understanding of the mechanisms influencing saltmarsh dynamics at the local scale, quantitative analyses are essential.
Before the introduction of S. alterniflora in 1979, China boasted a rich variety of native saltmarshes (Nie et al., 2023). The rapid growth and high reproductive capacity of S. alterniflora enable it to outcompete native saltmarshes for resources, leading to a decline in biodiversity and ecosystem functions within coastal wetlands. This transformation has shifted the local diverse community into more simplified ecosystems (Gu et al., 2018; Qi and Chmura, 2023). Zhao (2020) found that by 2018, coastal saltmarshes in China covered a total area of 106,352.5 ha, with S. alterniflora emerging as the dominant species, accounting for 61.4% of this area. A comparison between 2015 and 2018 indicated a slight decrease in saltmarshes, with a shift in dominant species from P. australis to S. alterniflora. Chen et al. (2022) created a spatiotemporal evolution map of China’s intertidal saltmarshes spanning from 1985 to 2019, outlining changes in area, species composition, and characteristics of different saltmarshes species. From 1985 to 2019, saltmarshes decreased significantly from 151,324 ha to 115,397 ha. S. glauca decreased from 77,741 ha to 3,563 ha, P. australis decreased from 60,511 ha to 33,193 ha, while S. alterniflora increased from 99 ha to 67,527 ha. These changes carry substantial ecological consequences (Luo, 2019; Nie et al., 2023). While research has delved into the processes and pathways of saltmarsh invasion (Wang et al., 2006; Li et al., 2009), the combined effects of natural factors and human activities remain unexamined.
Natural factors and human activities synergistically drive the spatiotemporal patterns of native saltmarshes decline and invasive saltmarshes expansion (Gedan et al., 2009; Kirwan et al., 2016; Nie et al., 2023). Global climate change, manifested through rising sea levels, extreme storm surges, biological invasions, and human activities, predominantly impact saltmarsh dynamics (Campbell et al., 2022; Murray et al., 2022). O’Donnell and Schalles (2016) identified significant correlations between water levels, mean sea levels, precipitation and the presence of S. alterniflora in Georgia’s coastal wetlands. Deegan et al. (2012) highlighted that coastal eutrophication driven by human activities is a global factor contributing to saltmarsh loss. In contrast, primary factors influencing the spatiotemporal evolution of saltmarshes in China include land reclamation, changes in water-sediment flux, interspecific competition, and saltmarsh succession (Chen et al., 2022). Additionally, the impacts of reclamation, S. alterniflora invasion, and sea level rise have led to rapid structural and spatial changes in coastal saltmarshes in China (Zhao, 2020). Scenario simulations predict that, with projected sea level rise, 20%-90% of global coastal wetlands are at risk of disappearing, threatening saltmarshes significantly (Schuerch et al., 2018). Furthermore, the rate of sea level rise considerably exceeds the rates of saltmarsh formation and evolution (FitzGerald and Hughes, 2019). Moreover, the global lower elevation limit for saltmarshes is influenced more by the frequency of tidal inundation than by its duration. This information is essential for accurately assessing the impacts of sea level rise and coastal changes on the lateral expansion of saltmarshes (Balke et al., 2016). Additionally, extreme weather events can alter saltmarsh distributions (Zedler, 2010). Furthermore, sediment deposition can greatly influence the growth of saltmarsh roots and shoots, which in turn affects the morphology, biomass, resilience, and blue carbon storage of these ecosystems (Tang et al., 2020; Pannozzo et al., 2023).
Natural factors and human activities have jointly led to extensive sedimentation in numerous bays, substantially modifying the growth environment and evolutionary patterns of saltmarshes (Van Geen and Luoma, 1999). In rapidly silting bays, the expansion of mudflats, along with large tidal inflows and high sedimentation rates, are compounded by human activities such as port blocking and land reclamation, further accelerating bay sedimentation (Gouleau et al., 2000). These conditions lead to sedimentation that affects tidal range and water capacity, significantly impacting the morphology of tidal channels and flats, thereby influencing the colonization, growth, and development of saltmarshes (Stumpf, 1983; Davidson-Arnott et al., 2002). Nonetheless, current research limits our understanding of the spatiotemporal dynamics and influencing factors—both natural and human— on the transition from native to invasive saltmarshes, particularly in rapidly silting coastal bays.
Yueqing Bay, one of three semi-enclosed bays in Zhejiang Province, experiences a regular semi-diurnal tide that diminishes from the outer Bay to the inner Bay. The Xuanmen reclamation project has caused significant changes in the sedimentation environment and current structure of Yueqing Bay, leading to a reduced tidal range, diminished sediment transport capacity, ongoing siltation, and annual deposition of 5 to 10 cm (Tian, 2020). Native saltmarshes in Yueqing Bay were primarily composed of P. australis and S. mariqueter. In 1983, a trial planting of invasive S. alterniflora was conducted on tidal flats to enhance sedimentation and ensure coastal stability. Unfortunately, S. alterniflora has since led to the decline of native saltmarshes (Chen et al., 2023). Currently, S. alterniflora has proliferated across the tidal flats, establishing a stable presence (Huang et al., 2016). Here, we proposed the hypothesis that mudflat reclamation and the seaward extension of coastlines can facilitate saltmarsh expansion by promoting rapid sedimentation and landward transformation (Laengner et al., 2019). Mangrove forests negatively impact S. alterniflora, as their planting often requires the removal of existing S. alterniflora. Additionally, tidal range and temperature positively influence saltmarsh expansion (Silvestri et al., 2005; Gedan and Bertness, 2010). Our research aims to investigate the spatiotemporal processes and underlying mechanisms in Yueqing Bay, focusing on two specific questions: (1) What are the spatiotemporal dynamics of native and invasive saltmarshes in rapidly silting bays? (2) How do natural factors and human activities influence these patterns in such environments? Our study seeks to identify the dominant factors affecting the spatiotemporal dynamics of native and invasive saltmarshes and to provide a theoretical basis for the restoration, conservation, and management of saltmarshes in coastal bays.
Yueqing Bay (120°57′–121°16′ E, 27°05′–28°23′ N), located in Zhejiang Province, eastern China. The bay is surrounded by hills on the east, north, and west of sides, while it opens southward with natural islands serving as barriers (Supplementary Figure S1). The bay’s shape resembles a gourd, featuring wider ends and a narrower middle, oriented roughly NNE-SSW (Supplementary Figure S1). Yueqing Bay spans approximately 42 km in length and has an average width of about 10 km. Its entrance can extend up to 21 km, while the narrowest width in the middle measures just 4.5 km. The bay contains around 30 islands and reefs larger than 1,000 m2. The bay bottom consists primarily of mud, transitioning to a mix of mud and sand at the mouth, with approximately 7 km2 of sandbanks. This region belongs to the maritime subtropical monsoon climate, characterized by mild temperatures, distinct seasons, and abundant precipitation. The mean annual temperature is 17.9°C, with a maximum of 36.7°C and a minimum of -5.8°C. The mean annual precipitation is 1,556.3 mm. Winter sediment content ranges from 0.01 to 1.37 kg/m3, while summer content ranges from 0.03 to 0.20 kg/m3. The mudflat area in Yueqing Bay covers 196.2 km2, representing around 48% of the Bay’s total area, with 103.09 km2 above sea level.
We selected two scenes of multispectral Landsat 5/8 images that fully cover Yueqing Bay, specifically from path rows 118040 and 118041 (Supplementary Table S1). Each image encompasses a maximum area of 185 km × 185 km. In total, we compiled 18 scene images from the years 1985, 1990, 1995, 2000, 2005, 2010, 2015, 2019, and 2023, approximately at five-year intervals, all featuring a spatial resolution of 30 m. To enhance the differentiation of saltmarshes from other vegetation types, we meticulously selected images based on phenological stages and tidal levels. Following the methodology of Chen et al. (2022), our image selection focused on the period between July and November, capturing saltmarshes during their peak season with the highest biomass. To improve the accuracy of saltmarshes classification, we specifically chose images taken during low tides to enhance spectral reflectance and minimize noise, as suggested by Pasquarella et al. (2016). Additionally, we manually selected images with less than 10% cloud cover and utilized FMask to eliminate pixels affected by cloud or cloud shadow, thereby reducing potential classification errors (Foga et al., 2017). In cases where suitable images were unavailable for a given year, we chose images from adjacent years. Ultimately, we processed the selected images for saltmarshes classification using the Google Earth Engine (GEE) cloud computing platform, which is frequently used for time-series studies to interpret land use and cover change (Hird et al., 2017).
We utilized Support Vector Machines (SVM) for the classification of saltmarshes, grounded in statistical learning theory and relying on training samples, as outlined by Campbell and Wang (2020). Prior to classification, we performed preprocessing and supervised classification on the selected images using the GEE platform. The SVM model categorized the Landsat 5/8 images into various saltmarsh species based on the chosen pixels (Supplementary Figure S2). To enhance the identification of saltmarsh species, the SVM model incorporated two vegetation indices and four water-related spectral indices, as recommended by Chen et al. (2022).
Field surveys and photographs captured by Unmanned Aerial Vehicles (UAV) significantly improved classification accuracy, effectively rectifying several instances of misclassification. In 2023, we comprehensively surveyed 14 sites within Yueqing Bay, these sites typically encompassed diverse saltmarshes, each covering an area of over 5 ha. We utilized UAV aerial photos to collect information on saltmarshes, and compared the classification results with these UAV images (Figure 1). The accuracy of saltmarsh classification was evaluated using the gathered data, which was divided into 70% training samples and 30% validation samples. Saltmarshes classification process with Landsat images is showed in Supplementary Figure S3. The commission error ranged from 0.09 to 0.12, while the validation accuracy varied from 0.86 to 0.95, with Kappa coefficients ranging from 0.84 to 0.96. Our saltmarsh classification achieved an accuracy exceeding 87%. The primary sources of error were misclassifications occurring in transitional zones.
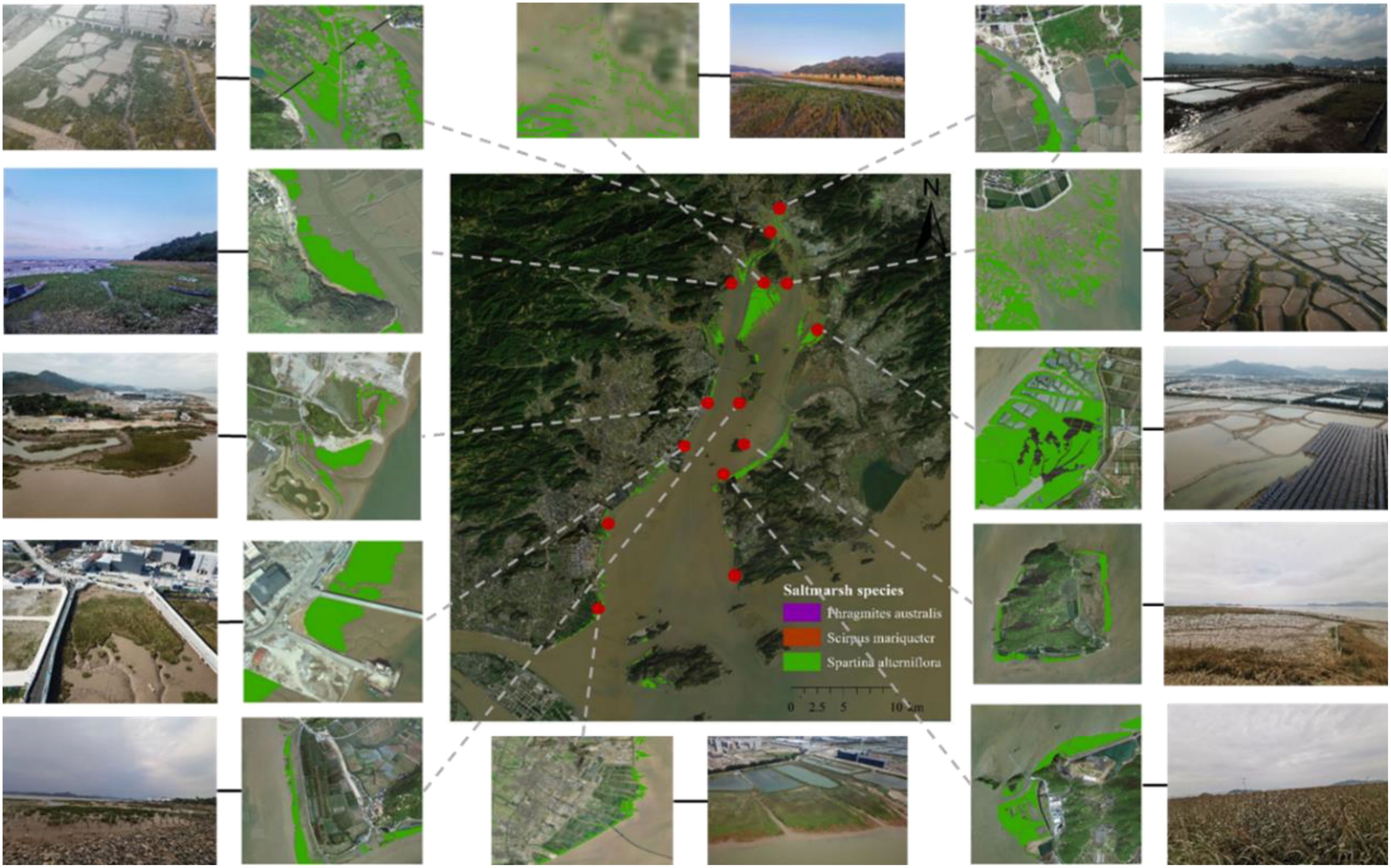
Figure 1. Geographical locations of field survey sites and classification verification of saltmarshes with aerial photos by UAV in Yueqing Bay in 2023.
We collected datasets to the best of our ability and within our authority regarding the mean sea level, maximum tidal level, mean tidal range, and maximum tidal range for each year from 1985 to 2012, utilizing the tidal gauge station of Longwan, situated in the outer Bay of Yueqing (Supplementary Table S2). This data can be compared across the years 1985, 1990, 1995, 2000, 2005, 2010, and 2012 (for 2015), with only the datasets for 2020 and 2023 missing. Consequently, the overall effects of sea level and tidal level on saltmarshes exhibit clear trends. The mean sea level recorded was 37.64 ± 8.22 cm, maximum tidal level was 421.57 ± 41.21 cm, mean tidal range was 451.18 ± 6.87 cm, and maximum tidal range was 678.82 ± 26.38 cm (Supplementary Table S2). Additionally, we compiled mean annual temperature for Yueqing Bay from the Yuhuan climate station covering the years 1970 to 2022, revealing an overall increasing trend (Supplementary Figure S4).
The largest mudflat reclamation project in Yueqing Bay was the Xuanmen project, which involved the construction of the Xuanmen Dam to connect Yuhuan Island. This project not only created over 7,000 ha of farmland but also established a reservoir with a storage capacity of 100 million m3. Other projects primarily focused on enhancing the seawall’s ability to withstand typhoons and supporting disaster prevention and mitigation efforts. Specifically, between 1985 and 1990, reclamation projects focused on the phase I of Huang’ao and Chaoyang Pond, totaling approximately 195.33 ha (Supplementary Table S3; Supplementary Figure S5). From 1990 to 2000, reclamation activities consisted of the south of Shengli Pond and Youyi Pond in Puqi, with a combined area of 393.80 ha. Between 2000 and 2005, reclamation initiatives extended to Xuanmen phase II, Wenyang, and Shashan Pond, covering a vast area of 3,910.67 ha. During the period from 2005 to 2010, reclamation efforts encompassed Puqi mudflat, Yuehai reclamation, north of Shengli Pond, Damen Island phase I, and Port reclamation, totaling 3,358.60 ha. Moving on to the period between 2010 and 2015, reclamation projects included Huangao phase II, Wenyang reclamation, and Lecheng reclamation, amounting to approximately 1,694.53 ha. Finally, from 2015 to 2023, reclamation endeavors comprised Damen port reclamation, Dianzuitou Pond, Taishan north pond, Taiping Pond, Xiaomen Island, and Lingang port in Damen Island, with a total area of 3,613.33 ha (Supplementary Table S3; Supplementary Figure S5).
The shoreline was determined by the average high tidal level during spring tide. These datasets were obtained from China Navigation Publications, which specializes in creating ocean charts. The historical development of the seabed in Yueqing Bay is prominently displayed through sediment accumulation from Puqi to Huanghua in the eastern part of Yueqing City (Supplementary Figure S6). This area serves as the primary deposition site for sediment at the southern entrance of Yueqing Bay and the branch of sediment from the Oujiang River, forming a relatively expansive plain. Analysis of the coastline of Yueqing Bay reveals substantial changes during the periods from 2002 to 2004 and from 2004 to 2011 (Supplementary Figure S6). The coastal region has undergone significant transformations due to extensive land reclamation activities, resulting in an eastward shift of the coastline caused by the development of tidal flats on the western side of the Bay. This shift led to a maximum extension of approximately 2.6 km and an average annual extension of around 108.3 m. The completion of Xuanmen phase II embankment project in 2001 notably extended the coastline from Fenshui Mountain to Xiaoqing Mountain, achieving a maximum extension of about 9.5 km. Furthermore, the coastline in the area affected by the embankment from Jianyu to Nanmian Mountain also experienced significant extension from 2005 to 2016, with a maximum extension of about 6.5 km.
In 1957, mangrove forests were introduced to Ximen Island in Yueqing Bay. Unfortunately, in the 1990s, extensive mangrove forests were destroyed by local residents for aquaculture development. By 2005, a large-scale initiative was launched to replant mangrove forests in Yueqing Bay, focusing particularly on the dominant species Kandelia obovata and Aegiceras corniculatum (L.) Blanco. The mangrove restoration process primarily involves clearing S. alterniflora before replanting the mangrove trees. We mapped the complete distribution of mangrove forests in Yueqing Bay in 2023 (Supplementary Figure S7).
Initially, we investigated the spatiotemporal distribution and area changes of native saltmarsh species (P. australis and S. mariqueter) and the invasive S. alterniflora from 1985 to 2023 in Yueqing Bay. Additionally, we analyzed the areas changes of different saltmarsh species in the Outer, Middle, and Inner Bays of Yueqing. We also highlighted four specific local regions to illustrate the spatiotemporal variations of the saltmarshes more clearly. Furthermore, we calculated the net changes in native saltmarshes and invasive S. alterniflora over five-year periods. Then, we obtained net changes for both native saltmarshes and invasive S. alterniflora during the same five-year periods across the Outer, Middle, and Inner Bays of Yueqing.
Finally, we employed a linear regression model to analyze the effects of natural factors (mean sea level, maximum tide level, mean tide range, and maximum tide range) as well as human activities (mudflat reclamation and shoreline changes) on saltmarsh dynamics. For sea level and tidal levels, we matched data from seven distinct years with saltmarsh area measurements. For mean annual temperature, we correlated saltmarsh area with mean annual temperature over nine corresponding years. Regarding mudflat reclamation, we selected reclamation areas during specific periods and examined their relationship with saltmarsh areas during those same periods.
Before performing linear regression analysis, we evaluated the normality of all influencing factors. To achieve symmetry and linearity, we applied log- transformations to mean sea level, maximum tide level, mean tide range, maximum tide range, and reclamation area. Statistical significance was determined as P < 0.05. All statistical analyses and figure creation were conducted using R version 4.3.1 (R Core Team, 2021) and ArcMap 10.8.
From 1985 to 2023, saltmarshes in Yueqing Bay exhibited an initial decline followed by a significant recovery (Table 1; Figures 2, 3). In 1985, saltmarshes covered 410.41 ha but decreased to 49.05 ha by 1995. Notably, they expanded dramatically to 2,872.90 ha by 2023. However, there was a slight decrease from 2010 to 2015, falling from 2,060.12 ha to 1,893.82 ha (Figure 2). In terms of species composition, native saltmarshes dominated by P. australis and S. mariqueter experienced a significant decline, while the invasive S. alterniflora has witnessed substantial growth (Supplementary Figure S8).

Table 1. Saltmarshes in Yueqing Bay from 1985 to 2023.
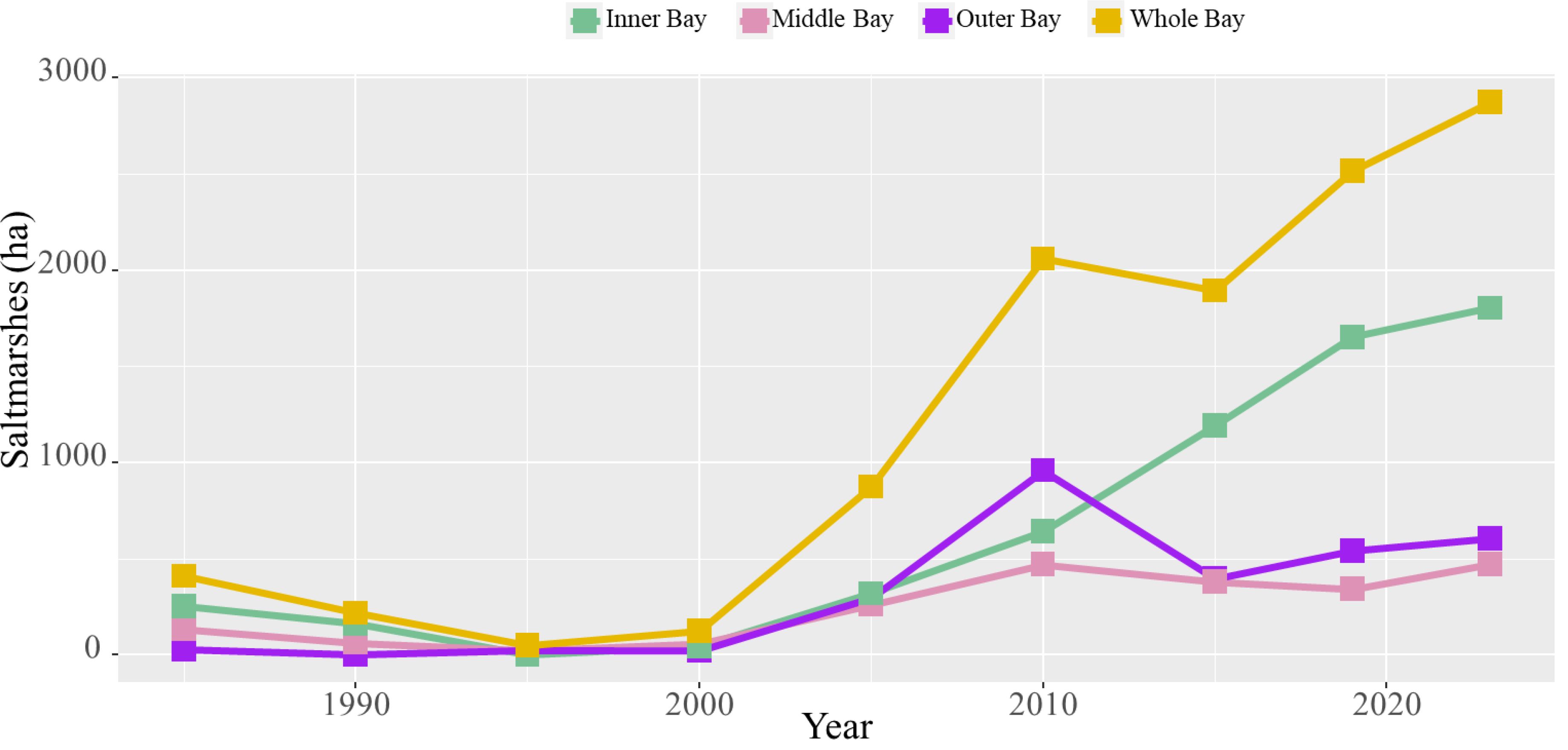
Figure 2. Saltmarshes in Yueqing Bay (including the whole, outer, middle, and inner Bays of Yueqing) during 1985-2023.

Figure 3. Spatiotemporal distribution of saltmarshes in Yueqing Bay from 1985 to 2023.
Different saltmarsh species exhibited distinct spatiotemporal dynamics in Yueqing Bay. During 1985-1990, S. mariqueter was the only species present, covering 410.41 ha in 1985 but declining to 218.52 ha by 1990, and nearly disappearing by 1995 with only 0.32 ha remaining. Brief increases were recorded in 2000 (7.19 ha) and 2005 (43.10 ha), after which S. mariqueter vanished entirely. P. australis appeared around 1995 with 26.15 ha and expanded to 60.07 ha in 2000, and 221.83 ha in 2005, but subsequently declined rapidly to 4.66 ha by 2010, ultimately disappearing thereafter. S. alterniflora was introduced to Yueqing Bay around 1995, initially covering only 22.58 ha. However, it underwent significant expansion, reaching a peak of 2,872.90 ha in 2023. During the period of 2010-2015, S. alterniflora experienced a slight decline, dropping from 2,060.12 ha in 2010 to 1,893.82 ha in 2015 (Table 1; Supplementary Figure S8). The outer, middle, and inner Bays of Yueqing exhibited unique spatiotemporal dynamics among various saltmarsh species. From 1985 to1990, S. mariqueter was distributed solely in the inner and middle Bays (Supplementary Table S4; Supplementary Figure S9). Saltmarshes in all three Bays experienced a decline from 1985 to 2000. Starting in 2000, S. alterniflora become the dominant species in Yueqing Bay, witnessing continuous growth in the inner Bay. Conversely, the outer and middle Bays experienced minor increases until 2010, followed by a downward trend with sporadic growth (Figure 2). P. australis, which first appeared in 1995 and peaked in 2005, was primarily concentrated in the middle Bay (Supplementary Table S4; Supplementary Figure S9). Additionally, land reclamation and shoreline changes had significant impact on the spatiotemporal dynamics of saltmarshes, with notable examples found in the southern of Ximen Island, Xuanmen Bay, Taiping Pond, and Shengli Pond (Figure 4).
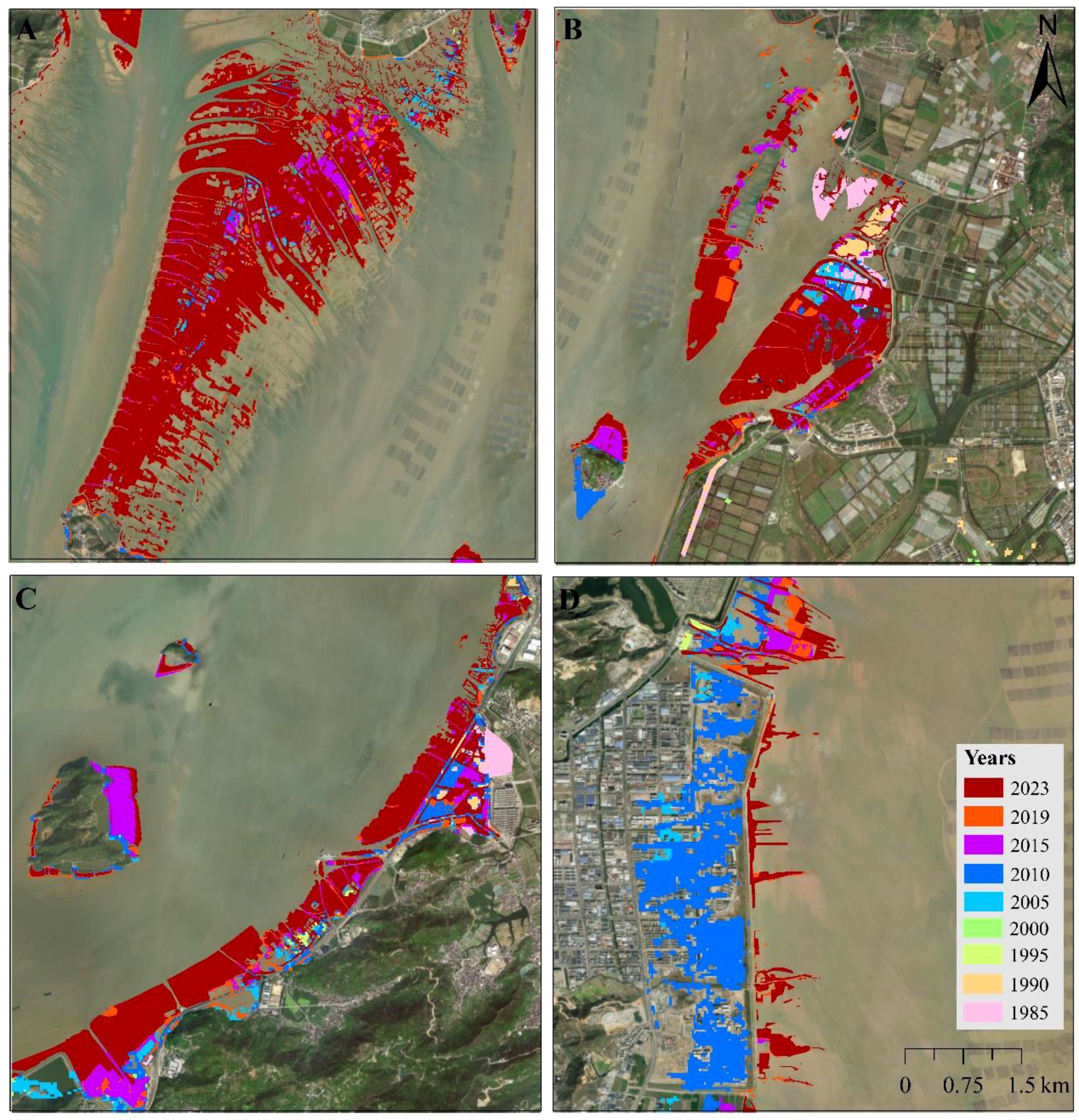
Figure 4. Distribution of saltmarshes in selected sites around Yueqing Bay from 1985 to 2023. (A), Southern of Ximen Island; (B), Xuanmen Bay; (C), Taiping Pond; (D), Shengli Pond.
In Yueqing Bay, saltmarshes experienced a decline from 1985 to 1995, followed by gradual expansion between 1995 and 2010, with the most significant growth occurring from 2005 to 2010. A slight shrinking trend emerged from 2010 to 2015, followed by consistent expansion in subsequent years. S. alterniflora demonstrated yearly increases, whereas P. australis expanded between 2005 and 2010 but contracted from 2010 to 2015. For S. mariqueter, substantial reductions were observed from 1985 to 1995, followed by growth between 2000 and 2005, and another decline from 2005 to 2010 (Figure 5; Supplementary Table S5).
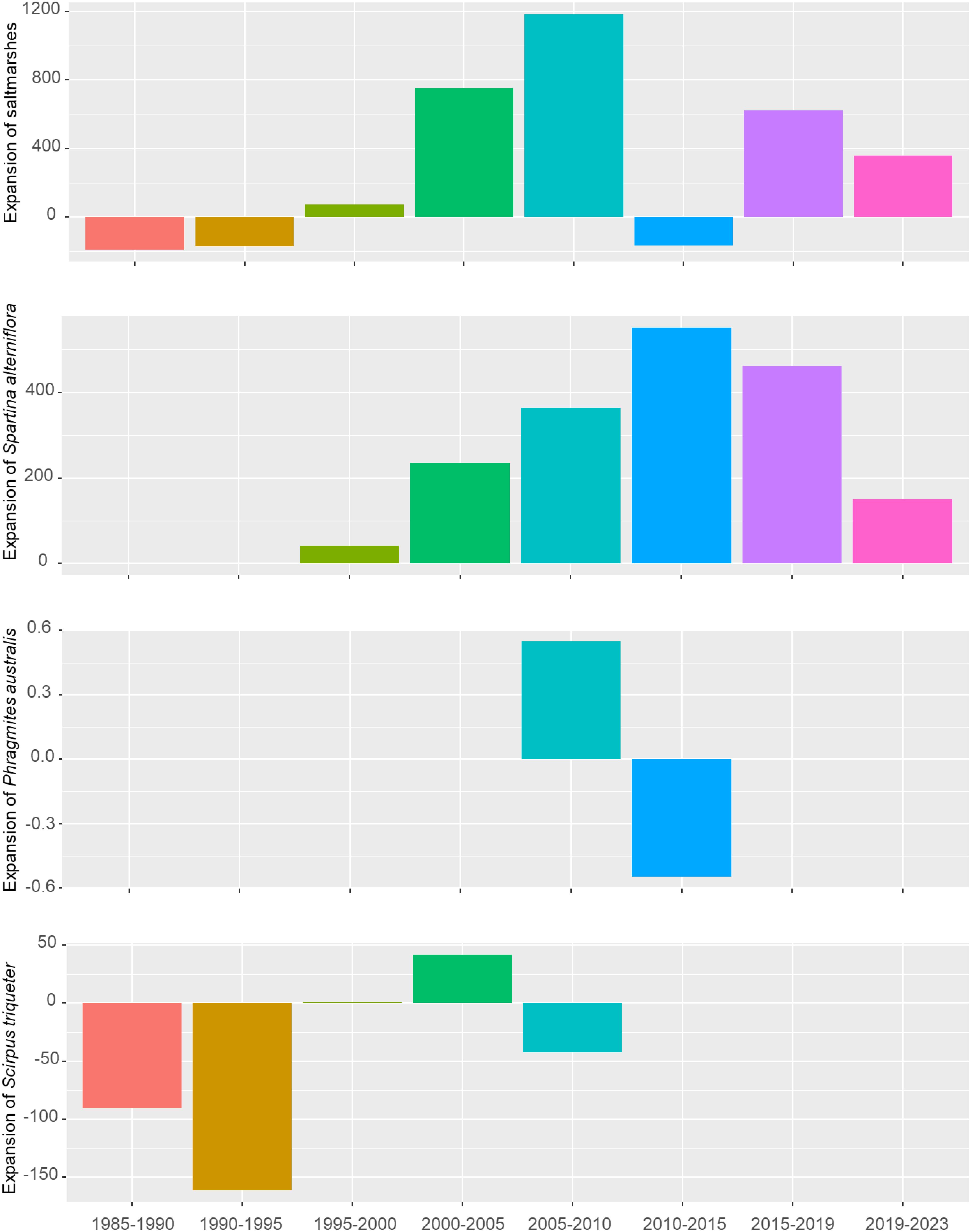
Figure 5. Changes in total saltmarshes, P. australis, S. mariqueter, and S. alterniflora in Yueqing Bay spanning from 1985 to 2023 at the interval of 5 years.
In the outer Bay of Yueqing, saltmarshes dominated by S. mariqueter experienced reductions of 27.36 ha between 1985 and 1990, and 2.56 ha between 1995 and 2000. The most significant reduction (563.7 ha) occurred between 2010 and 2015, primarily due to the spread of S. alterniflora. In other periods, saltmarshes exhibited an overall expanding trend, with an expansion of 663.91 ha between 2005 and 2010, followed by a 275.57 ha increase between 2000 and 2005, mainly attributed to the growth of S. alterniflora. In the middle Bay of Yueqing, between 1985 and 1995, as well as between 2010 and 2019, saltmarshes experienced reductions of 109.39 ha for S. mariqueter and 127.27 ha for S. alterniflora, respectively. The most severe expansion was observed for S. alterniflora between 2000-2010 and 2019-2023, with increases of 409.66 ha and 129.99 ha, respectively. P. australis expanded by 144.44 ha from 2000 to 2005 and contracted by 202.14 ha from 2005 to 2010. In the inner Bay of Yueqing, during 1985-1990 and 1990-1995, saltmarshes dominated by S. mariqueter, covering areas of 90.56 ha and 161.53 ha, respectively. During other time frames, primarily from 1995 to 2023, S. alterniflora was the main expanding species, with a total expansion of 1,803 ha. Meanwhile, S. mariqueter expanded by 41.68 ha from 2000 to 2005 but decreased by 42.63 ha from 2005 to 2010 (Figure 5; Supplementary Table S5).
Regarding natural factors, linear regression analysis showed that maximum tidal range positively influenced saltmarshes (R2 = 0.65; P = 0.021) (Figure 6D). Conversely, mean sea level showed a negative correlation with saltmarsh area, although this relationship did not reach statistical significance (P > 0.05) (Figure 6A). Neither maximum tidal level nor mean tidal range demonstrated a significant impact on saltmarsh areas (P > 0.05) (Figures 6B, C). Overall, mean annual temperature in Yueqing Bay displayed a fluctuating upward trend (Supplementary Figure S4). The saltmarsh area exhibited a significant positive correlation with the mean annual temperature (R2 = 0.49, P = 0.02) (Figure 6E).
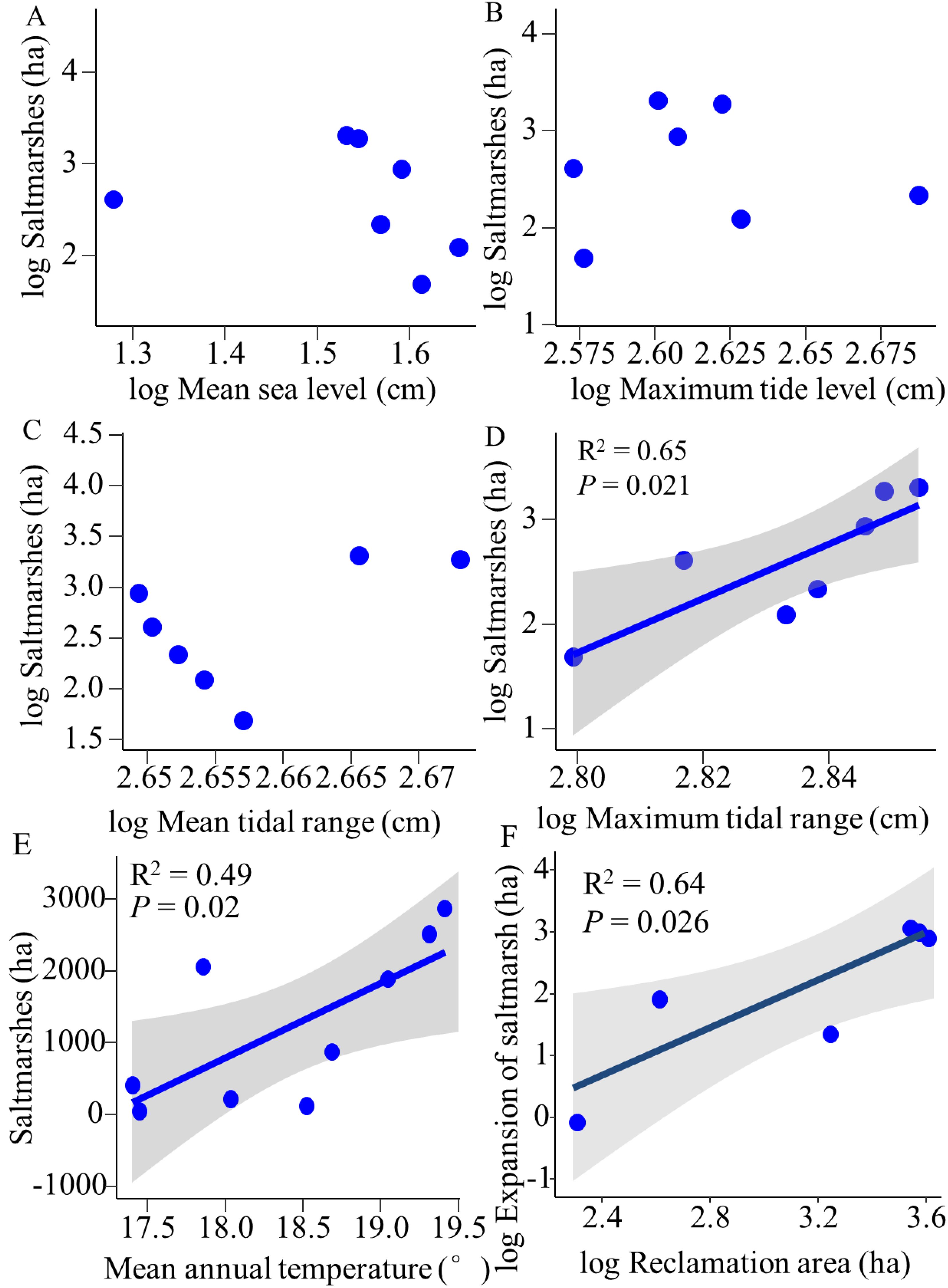
Figure 6. Linear regression of saltmarsh area versus mean sea level, maximum tide level, mean tide range, and maximum tide range (A-D), saltmarsh area and mean annual temperature (E), expansion area of saltmarshes and reclamation area (F) in Yueqing Bay from 1985 to 2023.
In terms of human activities, the overlay analysis of mudflat reclamation and saltmarshes in Yueqing Bay from 1985 to 2023 indicated that reclamation activities during specific periods contributed to the expansion of saltmarshes, thereby enhancing mudflat elevation (Supplementary Figure S10). Correlation analysis revealed a significant positive relationship between saltmarsh expansion during certain periods and the reclaimed area (R2 = 0.64; P = 0.026) (Figure 6F), suggesting that reclamation facilitates the expansion of saltmarshes, particularly S. alterniflora. Between 1984 and 2018, the coastline of Yueqing Bay predominantly extended seaward, driven by continuous coastal development and the transformation of natural coastlines into artificial ones. Substantial changes in artificial coastlines occurred from 1995 to 2002, with the smallest variations were observed between 1984 and 1995. The periods from 2002 to 2004 and from 2004 to 2011 witnessed the most significant shifts in Yueqing Bay’s coastline. The overlay analysis demonstrated that saltmarshes in Yueqing Bay expand seaward in conjunction with the coastline (Supplementary Figure S6). This seaward expansion leads to ongoing sedimentation on the seaward side, creating suitable conditions for saltmarsh growth, while continuous landward transformation rendering other areas unsuitable for saltmarsh development. Yueqing Bay currently hosts 118.05 ha of mangrove forests, primarily located on Ximen Island, Haishan Island, Jiangyan Island, Maoting Island, Daqing Island, Taiping Pond, Maotan Island, and Xiaqing Pond (Supplementary Figure S7), with Kandelia obovate being the dominant species. The mangrove forests exert a negative influence on S. alterniflora by encroached their habitats.
We observed significant spatiotemporal dynamics in native saltmarshes (P. australis and S. mariqueter), which have been declining, while invasive S. alterniflora has been expanding in the rapidly silting Bay of Yueqing from 1985 to 2023. Prior to the introduction of S. alterniflora into Yueqing Bay, the predominant native saltmarshes were P. australis and S. mariqueter, covering an area of 410.41 ha. As rapid sedimentation increases, the habitats of Yueqing Bay are altered, allowing S. alterniflora to compete with native saltmarshes for unvegetated flats, which has contributed to a decline in their expansion. Additionally, the allelopathic effect of S. alterniflora may lead to the degradation of native species (Chen et al., 2018). Simultaneously, S. alterniflora rapidly expanded, benefiting from the favorable habitat conditions, despite several manual removal attempts. Presently, S. alterniflora predominantly occupied Yueqing Bay. Research indicates that S. alterniflora covered approximately 24 km2 in 2013 and expanded to 33.5 km2 by 2023, while S. glauca and P. australis are now sporadically distributed (Huang et al., 2016; Chen et al., 2023).
Between 2005 and 2010, it was evident that S. alterniflora rapidly invaded the habitat of P. australis, leading to a notable decline in P. australis (Supplementary Figure S8). Concurrently, upon analyzing saltmarshes distribution in Zhejiang Province from 1985 to 2019 (Supplementary Table S6) noted a parallel expansion trend in both Yueqing Bay and the broader Zhejiang Province (Chen et al., 2022), supported by a notable correlation between their respective areas (R2 = 0.41; P < 0.01). Interestingly, similar expansion patterns were observed across various bays in Zhejiang Province, including Hangzhou Bay, Sanmen Bay, Xiangshan Bay, and Aojiang Bay (Lu, 2018). In contrast, the tidal flat in Yueqing Bay is broad and flat, spanning approximately 103.1 km² from the shoreline to the mean sea level, providing optimal conditions for saltmarsh growth. Over nearly four decades, S. alterniflora has spread throughout the tidal flats of Yueqing Bay, with an invasive area exceeding 3000 ha, constituting over half of the total S. alterniflora coverage in southern Zhejiang (Huang et al., 2016; Lu, 2018).
We found that both natural factors and human activities play a significant role in shaping the dynamics of saltmarshes in rapidly silting bays. This finding aligns with previous studies highlighting the impacts of coastal reclamation, sediment transport, climate change, and sea level rise on saltmarsh distribution (Chung et al., 2004). Notably, these factors are not mutually exclusive; they can interact in complex ways, further influencing saltmarsh distribution. Specifically, rising sea levels have had minimal impact on the spatiotemporal dynamics of saltmarshes in Yueqing Bay (Figure 6A), possibly because both native and invasive saltmarshes exhibit low sensitivity to sea level rise at local scales, often creating more favorable habitat conditions instead. Research indicates that sea level rise affects native saltmarshes and invasive S. alterniflora differently. While native species such as P. australis and S. mariqueter may diminish or disappear entirely, S. alterniflora could extend its range or colonize new areas in response to rising sea levels (Crosby et al., 2016; Valiela et al., 2018; Fagherazzi et al., 2020). The expansion of saltmarshes significantly increases with the maximum tidal range, as emphasized by Balke et al. (2016), who observed that the height difference between the lowest elevation of saltmarshes and mean high water increases with tidal range. Additionally, FitzGerald and Hughes (2019) identified the mean tidal range as a critical factor influencing the growth of S. alterniflora. The regular rise and fall of tides cause fluctuations in water levels, sediment composition, nutrient availability, and salinity, all of which play pivotal roles in shaping the growth and distribution of saltmarshes (Reed, 1988; Silvestri et al., 2005; Tonelli et al., 2010).
In Yueqing Bay, the saltmarsh area has expanded in response to rising temperatures (Figure 6E), this may underscore the significant impact of temperature on saltmarsh growth. Temperature directly influences the growth and survival of saltmarshes (Couto et al., 2014). Some species may struggle to adapt to warmer conditions, leading to a contraction in their distribution range, while other saltmarsh species may capitalize on the opportunity to broaden their habitats (Kirwan and Mudd, 2012). Furthermore, escalating temperatures can alter the timing and frequency of reproduction in saltmarshes, thereby influencing their distribution patterns (Gedan and Bertness, 2010).
The effects of land reclamation on saltmarshes depend on the differences in reclamation and sedimentation rates. Our research indicates that land reclamation has significantly increased the mudflats in Yueqing Bay due to rapid sedimentation, which has created more favorable habitats for saltmarshes and particularly facilitating the spread of S. alterniflora. Moreover, our findings revealed that land reclamation, particularly the Xuanmen Project, resulted in the disappearance of native saltmarshes, including P. australis and S. mariqueter (Figure 3). Conversely, numerous studies have documented that reclamation can lead to the loss of extensive saltmarsh areas, resulting in a marked decline in available habitat for these ecosystems (Chen et al., 2022; Murray et al., 2022). Land reclamation has also increased geomorphological elevation, hindering the diffusion of seawater due to tidal action and altering ecological processes (Portnoy and Giblin, 1997; Naser, 2022). These changes have led to modifications in soil physico-chemical properties and hydrological conditions in the long-term following saltmarshes reclamation (Xie et al., 2017; Lu et al., 2021). Additionally, reclamation alters water quality, soil salinity, and nutrient levels, which can enhance resources availability for saltmarshes (Gedan et al., 2009; Lu et al., 2023). Land reclamation has the potential to disrupt the natural growth and reproductive cycles of saltmarshes, thereby altering their distribution dynamics. Moreover, reclamation has indirectly promoted the growth and development of saltmarsh species (Zhang and Shao, 2013). The effects of reclamation on saltmarshes are both immediate and prolonged, with short-term fluctuations evident in the period immediately following reclamation, potentially leading to rapid and substantial changes in saltmarshes. Over the long term, these alterations may contribute to the gradual extinction of certain species within the saltmarsh ecosystem.
Changes in the coastline due to mudflat reclamation can directly affect the availability of suitable habitats for saltmarshes, thereby altering their spatiotemporal dynamics. These coastline alterations significantly impact crucial hydrological conditions such as water flow, salinity, and water availability, all of which are essential for the growth and survival of saltmarshes. Additionally, modifications to the coastline can influence sediment supply and deposition patterns, thereby impacting the soil quality and composition within saltmarshes. Research showed that between 1990 and 2017, the coastline underwent significant transformation, becoming predominantly artificial, notably during the periods of 2002-2004 and 2004-2011. The primary drivers of these changes were reclamation efforts for aquaculture and the construction of new port terminals, which reshaped the coastline (Peng et al., 2019). Specifically, from 1984 to 2018, the coastline of Yueqing Bay predominantly expanded seaward, driven by the continuous growth of artificial coastlines through activities such as aquaculture reclamation and port terminal construction. Additionally, the conversion of natural coastlines into artificial ones, exemplified by projects like the Xuanmen Project, contributed to this trend of coastal expansion. Yueqing Bay experienced increased landscape fragmentation during 1995-2010, leading to a gradual decline in patch connectivity, enhanced landscape heterogeneity, and greater landscape diversity (Sun et al., 2021). These changes in the coastline collectively influence the spatiotemporal dynamics of saltmarshes in Yueqing Bay.
Mangroves forests exerted a notably adverse effect on S. alterniflora in Yueqing Bay due to deliberate eradication efforts in conjunction with mangrove restoration. Mangroves can modify local environments by regulating factors such as water flow, salinity, sedimentation rates, and soil conditions, subsequently impacting the distribution and growth patterns of saltmarshes (Saintilan et al., 2014; Rodriguez et al., 2016; Kelleway et al., 2017). Furthermore, mangroves compete with saltmarshes for light, water, and nutrients (Rogers and Krauss, 2019; Zhang et al., 2021). Additionally, manual removal of saltmarshes to maintain mangrove forests has been observed in Yueqing Bay. Studies indicate that saltmarsh elevation is mainly influenced by its position in the tidal prism, with long-term changes linked to global sea-level trends, while short-term fluctuations result from local water level and rainfall variations (Rogers et al., 2014). Storm activity can boost local vertical accretion and minimize lateral retreat of saltmarshes (Pannozzo et al., 2023). For example, mesocosm experiments show that a 1 cm increase in sediment deposition can raise average plant height by 1.51 cm (Tang et al., 2020). Given the limited sediment data from Yuqing Bay in our study, future observations and simulations should consider sediment’s impact on the spatiotemporal dynamics of saltmarshes.
Our research indicates that mudflat reclamation, coastline changes, mangrove forest plantations, and tidal ranges significantly impact saltmarshes in rapidly silting bays. Studies have found that land reclamation, the invasion of S. alterniflora, and the rise in sea levels, water levels, and precipitation are the most significant factors affecting other wetlands (O’Donnell and Schalles, 2016; Zhao, 2020). At local scales, elevation is a comprehensive factor determining the distribution of saltmarshes, as it directly affects soil salinity and the duration of tidal flooding (He et al., 2010). At lower elevations, tidal actions determine soil salinity and oxygen content, thereby affecting saltmarshes; as elevation increases, interspecific competition and other factors become more significant (Pennings et al., 2005). Research has also shown that the upper limit of saltmarsh distribution is mainly determined by habitat conditions or interspecific competition, while the lower limit is determined by abiotic factors such as tides and flooding (He et al., 2010). Furthermore, soil nutrients play a crucial role in influencing the distribution of saltmarshes. Therefore, future studies should concentrate on the drivers of saltmarsh dynamics at local scales.
The invasive S. alterniflora has high adaptability and dissemination capabilities, occupying a large portion of China’s coastline. Although it offers high ecological service and carbon sink functions, it leads to a decline in native plants and habitats, negatively impacts migratory birds, and reduces income for aquaculture farmers. In 2022, the Chinese government initiated the S. alterniflora Prevention and Control Special Action Plan (2022-2025), it appears highly effective in the short term; however, the high recurrence rate and endless costs involved threaten the overall success of the eradication efforts. Therefore, implementing long-term monitoring and ecological restoration after the eradication of S. alterniflora, as well as protecting native saltmarsh species, has become essential.
Native saltmarshes in many coastal bays are facing significant threats from invasive species, particularly S. alterniflora. Our study has revealed notable spatiotemporal dynamics concerning the decline of native saltmarshes and the expansion of invasive S. alterniflora in Yueqing Bay. These results indicate that both natural and human-induced factors have significantly impacted these dynamics. These factors include mudflat reclamation, coastline change, mangrove forests plantation, tidal range, and temperature. Our findings highlight the importance of considering both natural and human-induced factors when managing and monitoring the dynamics of native and invasive saltmarshes in rapidly silting coastal bays. It is crucial to develop effective strategies to protect native saltmarshes and control the spread of invasive species, while ensuring the sustainable management of coastal ecosystems.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
MX: Data curation, Funding acquisition, Investigation, Software, Validation, Writing – original draft, Writing – review & editing. JLG: Data curation, Formal analysis, Investigation, Methodology, Resources, Software, Writing – original draft. PY: Data curation, Methodology, Validation, Writing – original draft. WF: Formal analysis, Methodology, Software, Writing – original draft. GZ: Investigation, Methodology, Resources, Writing – original draft. ZC: Project administration, Methodology, Writing – original draft. JJG: Investigation, Methodology, Writing – original draft. JS: Writing – original draft. JZ: Funding acquisition, Project administration, Supervision, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by the Natural Science Foundation of Zhejiang Province of China (LQ24C030006), National Natural Science Foundation of China (42301081), Zhejiang Province science and technology plan project (RB2225, RC2253, RC2255, and RB2414), Joint Funds of Natural Science Foundation of Zhejiang Province and Water Resources Department (LZJWZ23E090005), Zhejiang Basic Public Welfare Research Program (LZJWZ22E090005), and Ningbo Science and Technology Innovation 2025 Major Special Project (2023Z146, 2022Z032, and 2022Z181).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1459935/full#supplementary-material
Ainouche M., Gray A. (2016). Invasive Spartina: lessons and challenges. Biol. Invasions 18, 2119–2122. doi: 10.1007/s10530-016-1201-7
Balke T., Stock. M., Jensen. K., Bouma T. J., Kleyer M. (2016). A global analysis of the seaward salt marsh extent: The importance of tidal range. Water Resour. Res. 52, 3775–3786. doi: 10.1002/2015WR018318
Bertness M. D. (2006). Atlantic shorelines: natural history and ecology (Princeton, New Jersey: Princeton University Press).
Boesch D. F., Turner R. E. (1984). Dependence of fishery species on salt marshes-the role of food and refuge. Estuaries 7, 460–468. doi: 10.2307/1351627
Campbell A. D., Fatoyinbo L., Goldberg L., Lagomasino D. (2022). Global hotspots of salt marsh change and carbon emissions. Nature 612, 701–706. doi: 10.1038/s41586-022-05355-z
Campbell A. D., Wang Y. (2020). Salt marsh monitoring along the mid-Atlantic coast by Google Earth Engine enabled time series. PLoS One 15, e0229605. doi: 10.1371/journal.pone.0229605
Chen G., Jin R., Ye Z., Li Q., Gu J., Luo M., et al. (2022). Spatiotemporal mapping of salt marshes in the intertidal zone of China during 1985–2019. J. Remote Sens. 2022, 793626. doi: 10.34133/2022/9793626
Chen Y. T., Qiao W. H., Zeng C. S. (2018). Allelopathic effect of exotic Spartina alterniflora on native Cyperus malaccensis in Minjiang Estuary. J. Appl. Oceanogr. 37, 45–52. doi: 10.3969/J.ISSN.2095-4972.2018.01.005
Chen Y. H., Zhang S. G., Zhang Z. (2023). Estimation of organic carbon density and carbon storage of salt marsh wetland in Yueqing Bay. Mar. Environ. Sci. 42, 38–45. doi: 10.13634/j.cnki.mes.2023.01.021
Chung C. H., Zhuo R. Z., Xu G. W. (2004). Creation of Spartina plantations for reclaiming Dongtai, China, tidal flats and offshore sands. Ecol. Eng. 23, 135–150. doi: 10.1016/j.ecoleng.2004.07.004
Couto T., Martins I., Duarte B., Caçador I., Marques J. C. (2014). Modelling the effects of global temperature increase on the growth of salt marsh plants. Appl. Ecol. Env. Res. 12, 753–764. doi: 10.15666/aeer/1203_753764
Crosby S. C., Sax D. F., Palmer M. E., Booth H. S., Deegan L. A., Bertness M. D., et al. (2016). Salt marsh persistence is threatened by predicted sea-level rise. Estuar. Coast. Shelf S. 181, 93–99. doi: 10.1016/j.ecss.2016.08.018
Davidson-Arnott R. G., Van Proosdij D., Ollerhead J., Schostak L. (2002). Hydrodynamics and sedimentation in salt marshes: examples from a macrotidal marsh, Bay of Fundy. Geomorphology 48, 209–231. doi: 10.1016/S0169-555X(02)00182-4
Deegan L. A., Johnson D. S., Warren R. S., Peterson B. J., Fleeger J. W., Fagherazzi S., et al. (2012). Coastal eutrophication as a driver of salt marsh loss. Nature 490, 388–392. doi: 10.1038/nature11533
Fagherazzi S., Mariotti G., Leonardi N., Canestrelli A., Nardin W., Kearney W. S. (2020). Salt marsh dynamics in a period of accelerated sea level rise. J. Geophys. Res. 125, e2019JF005200. doi: 10.1029/2019JF005200
FitzGerald D. M., Hughes Z. (2019). Marsh processes and their response to climate change and sea-level rise. Annu. Rev. Earth Pl. Sc. 47, 481–517. doi: 10.1146/annurev-earth-082517-010255
Foga S., Scaramuzza P. L., Guo S., Zhu Z., Dilley R. D. Jr., Beckmann T., et al. (2017). Cloud detection algorithm comparison and validation for operational Landsat data products. Remote Sens. Environ. 194, 379–390. doi: 10.1016/j.rse.2017.03.026
Gedan K. B., Bertness M. D. (2010). How will warming affect the salt marsh foundation species Spartina patens and its ecological role? Oecologia 164, 479–487. doi: 10.1007/s00442-010-1661-x
Gedan K. B., Silliman B. R., Bertness M. D. (2009). Centuries of human-driven change in salt marsh ecosystems. Annu. Rev. Mar. Sci. 1, 117–141. doi: 10.1146/annurev.marine.010908.163930
Gouleau D., Jouanneau J. M., Weber O., Sauriau P. G. (2000). Short-and long-term sedimentation on Montportail–Brouage intertidal mudflat, Marennes–Oleron Bay (France). Cont. Shelf Res. 20, 1513–1530. doi: 10.1016/S0278-4343(00)00035-2
Gu J., Luo M., Zhang X., Christakos G., Agusti S., Duarte C. M., et al. (2018). Losses of salt marsh in China: Trends, threats and management. Estuar. Coast. Shelf S. 214, 98–109. doi: 10.1016/j.ecss.2018.09.015
He Q., An Y., Cui B. S. (2010). Coastal salt marshes and distribution and diversity of salt marsh plant communities. Ecol. Environ. Sci. 19, 657–664. doi: 10.16258/j.cnki.1674-5906.2010.03.024
Hird J. N., DeLancey E. R., McDermid G. J., Kariyeva J. (2017). Google Earth Engine, open-access satellite data, and machine learning in support of large-area probabilistic wetland mapping. Remote Sens. 9, 1315. doi: 10.3390/rs9121315
Huang Z. X., Shen H., Peng X., Xing C. W., Zhu D. (2016). Monitoring of Spartina Alterniflora invasion by using high resolution satellite in Yueqing Bay, Zhejiang Province. Ocean Dev. Manage. 33, 63–67. doi: 10.20016/j.cnki.hykfygl.2016.10.014
Kelleway J. J., Cavanaugh K., Rogers K., Feller I. C., Ens E., Doughty C., et al. (2017). Review of the ecosystem service implications of mangrove encroachment into salt marshes. Global Change Biol. 23, 3967–3983. doi: 10.1111/gcb.13727
Kirwan M. L., Mudd S. M. (2012). Response of salt-marsh carbon accumulation to climate change. Nature 489, 550–553. doi: 10.1038/nature11440
Kirwan M. L., Temmerman S., Skeehan E. E., Guntenspergen G. R., Fagherazzi S. (2016). Overestimation of marsh vulnerability to sea level rise. Nat. Clim. Change 6, 253–260. doi: 10.1038/NCLIMATE2909
Laengner M. L., Siteur K., van der Wal D. (2019). Trends in the seaward extent of saltmarshes across Europe from long-term satellite data. Remote Sens. 11, 1653. doi: 10.3390/rs11141653
Leonard L. A., Croft A. L. (2006). The effect of standing biomass on flow velocity and turbulence in Spartina alterniflora canopies. Estuar. Coast. Shelf S. 69, 325–336. doi: 10.1016/j.ecss.2006.05.004
Li B. O., Liao C. H., Zhang X. D., Chen H. L., Wang Q., Chen Z. Y., et al. (2009). Spartina alterniflora invasions in the Yangtze River estuary, China: an overview of current status and ecosystem effects. Ecol. Eng. 35, 511–520. doi: 10.1016/j.ecoleng.2008.05.013
Lu L. Y. (2018). Spatial distribution and influencing factors of the biomass of Spartina alterniflora in coastal wetlands of Zhejiang. Chin. Acad. Forestry.
Lu S., Lu W., Xu M., Taghizadeh-Hesary F., Tang Y. (2023). Water-energy-food security under green finance constraints in Southwest China. Energy Econ. 118, 106478. doi: 10.1016/j.eneco.2022.106478
Lu S., Zhang X., Peng H., Skitmore M., Bai X., Zheng Z. (2021). The energy-food-water nexus: Water footprint of Henan-Hubei-Hunan in China. Renew. Sust. Energ. Rev. 135, 110417. doi: 10.1016/j.rser.2020.110417
Luo M. (2019). Remote sensing monitoring and ecosystem service value assessment of coastal salt marshes in China in 2015. Zhejiang Univ.
Luo F., Dai J. C., Chen Z. P., Zhou G. H., Zeng J. W., Zhang C. (2024). Dynamic monitoring and driving force analysis of salt marsh vegetation distribution in Yancheng Nature Reserve. J. Hohai Univ. 52, 88–95. doi: 10.3876/j.issn.1000-1980.2024.03.012
Murray N. J., Worthington T. A., Bunting P., Duce S., Hagger V., Lovelock C. E., et al. (2022). High-resolution mapping of losses and gains of Earth’s tidal wetlands. Science 376, 744–749. doi: 10.1126/science.abm9583
Naser H. A. (2022). Community structures of benthic macrofauna in reclaimed and natural intertidal areas in Bahrain, Arabian Gulf. J. Mar. Sci. Eng. 10, 945. doi: 10.3390/jmse10070945
Nie M., Liu W., Pennings S. C., Li B. (2023). Lessons from the invasion of Spartina alterniflora in coastal China. Ecology 104, 1–5. doi: 10.1002/ecy.3874
O’Donnell J. P., Schalles J. F. (2016). Examination of abiotic drivers and their influence on Spartina alterniflora biomass over a twenty-eight year period using Landsat 5 TM satellite imagery of the Central Georgia Coast. Remote Sens. 8, 477. doi: 10.3390/rs8060477
Pannozzo N., Leonardi N., Carnacina I., Smedley R. K. (2023). Storm sediment contribution to salt marsh accretion and expansion. Geomorphology 430, 108670. doi: 10.1016/j.geomorph.2023.108670
Pasquarella V. J., Holden C. E., Kaufman L., Woodcock C. E. (2016). From imagery to ecology: leveraging time series of all available Landsat observations to map and monitor ecosystem state and dynamics. Remote Sens. Ecol. Conserv. 2, 152–170. doi: 10.1002/rse2.24
Peng X. J., Xu R., He Y. L. (2019). Analysis on coastline and coastal wetland changes in Yueqing Bay in recent 30 years. Mar. Environ. Sci. 38, 68–74. doi: 10.13634/j.cnki.mes.2019.01.034
Pennings S. C., Grant M. B., Bertness M. D. (2005). Plant zonation in low-latitude salt marshes: disentangling the roles of flooding, salinity and competition. J. Ecol. 93, 159–167. doi: 10.1111/j.1365-2745.2004.00959.x
Portnoy J. W., Giblin A. E. (1997). Effects of historic tidal restrictions on salt marsh sediment chemistry. Biogeochemistry 36, 275–303. doi: 10.1023/A:1005715520988
Qi X., Chmura G. L. (2023). Invasive Spartina alterniflora marshes in China: a blue carbon sink at the expense of other ecosystem services. Front. Ecol. Environ. 21, 182–190. doi: 10.1002/fee.2611
Reed D. J. (1988). Sediment dynamics and deposition in a retreating coastal salt marsh. Estuar. Coast. Shelf S. 26, 67–79. doi: 10.1016/0272-7714(88)90012-1
Rodriguez W., Feller I. C., Cavanaugh K. C. (2016). Spatio-temporal changes of a mangrove–saltmarsh ecotone in the northeastern coast of Florida, USA. Glob. Ecol. Conser. 7, 245–261. doi: 10.1016/j.gecco.2016.07.005
Rogers K., Krauss K. W. (2019). Moving from generalisations to specificity about mangrove–saltmarsh dynamics. Wetlands 39, 1155–1178. doi: 10.1007/s13157-018-1067-9
Rogers K., Saintilan N., Woodroffe C. D. (2014). Surface elevation change and vegetation distribution dynamics in a subtropical coastal wetland: implications for coastal wetland response to climate change. Estuar. Coast. Shelf S. 149, 46–56. doi: 10.1016/j.ecss.2014.07.009
Saintilan N., Wilson N. C., Rogers K., Rajkaran A., Krauss K. W. (2014). Mangrove expansion and salt marsh decline at mangrove poleward limits. Global Change Biol. 20, 147–157. doi: 10.1111/gcb.12341
Schuerch M., Spencer T., Temmerman S., Kirwan M. L., Wolff C., Lincke D., et al. (2018). Future response of global coastal wetlands to sea-level rise. Nature 561, 231–234. doi: 10.1038/s41586-019-1205-4
Silvestri S., Defina A., Marani M. (2005). Tidal regime, salinity and salt marsh plant zonation. Estuar. Coast. Shelf S. 62, 119–130. doi: 10.1016/j.ecss.2004.08.010
Stumpf R. P. (1983). The process of sedimentation on the surface of a salt marsh. Estuar. Coast. Shelf S. 17, 495–508. doi: 10.1016/0272-7714(83)90002-1
Sun X. M., Yu M. J., Wu T., Xie X. F., Jiang G. J., Zhang J. Z. (2021). Landscape pattern evolution and its response to human disturbance of Yueqing Bay in recent 20 years. J. Zhejiang Norm. Univ. 44, 319–328. doi: 10.16218/j.issn.1001-5051.2021.03.011
Tang H., Xin P., Ge Z., Gong Z., Yang Y., Zhang Y., et al. (2020). Response of a salt marsh plant to sediment deposition disturbance. Estuar. Coast. Shelf S. 237, 106695. doi: 10.1016/j.ecss.2020.106695
Tian Y. (2020). Exploration on cumulative response of hydrodynamics and water environment of Yueqing Bay to reclamation works. North China Univ. Water Resour. Electric Power. doi: 10.27144/d.cnki.ghbsc.2020.000120
Tonelli M., Fagherazzi S., Petti M. (2010). Modeling wave impact on salt marsh boundaries. J. Geophys. Res-Oceans 115, C09028. doi: 10.1029/2009JC006026
Valiela I., Lloret J., Bowyer T., Miner S., Remsen D., Elmstrom E., et al. (2018). Transient coastal landscapes: rising sea level threatens salt marshes. Sci. Total Environ. 640, 1148–1156. doi: 10.1016/j.scitotenv.2018.05.235
Van Geen A., Luoma S. N. (1999). The impact of human activities on sediments of San Francisco Bay, California: an overview. Mar. Chem. 64, 1–6. doi: 10.1016/S0304-4203(98)00080-2
Wang Q., An S. Q., Ma Z. J., Zhao B., Chen J. K., Li B. (2006). Invasive Spartina alterniflora: biology, ecology and management. J. Syst. Evol. 44, 559. doi: 10.1360/aps06044
Xie X. F., Pu L. J., Wang Q. Q., Zhu M. G., Xu Y., Zhang M. (2017). Response of soil physicochemical properties and enzyme activities to long-term reclamation of coastal saline soil, Eastern China. Sci. Total Environ. 607-608, 1419–1427. doi: 10.1016/j.scitotenv.2017.05.185
Yin X. L., Tan C. Y., Ke Y. H., Zhou D. M. (2024). Evolution and driving factors of salt marsh wetland landscape pattern in the Yellow River Delta in 1973-2020. Acta Ecol. Sin. 44, 67–80. doi: 10.20103/j.stxb.202211203355
Zedler J. B. (2010). How frequent storms affect wetland vegetation: a preview of climate-change impacts. Front. Ecol. Environ. 8, 540–547. doi: 10.1890/090109
Zhang Z., Li J., Li Y., Liu W. W., Li Y. F. (2021). Spatially discontinuous relationships between salt marsh invasion and mangrove forest fragmentation. For. Ecol. Manage. 499, 119611. doi: 10.1016/j.foreco.2021.119611
Zhang L., Shao H. (2013). Direct plant-plant facilitation in coastal wetlands: A review. Estuar. Coast. Shelf S. 119, 1–6. doi: 10.1016/j.ecss.2013.01.002
Keywords: coastline change, Google Earth Engine, invasive Spartina alterniflora, mangrove forests, mudflat reclamation, tidal range, Yueqing Bay
Citation: Xu M, Gu J, Yun P, Gan J, Fan W, Chen Z, Shao J, Zhang G and Zeng J (2025) Spatiotemporal dynamics and influencing factors of native and invasive saltmarshes in a rapidly silting bay during 1985–2023. Front. Mar. Sci. 11:1459935. doi: 10.3389/fmars.2024.1459935
Received: 05 July 2024; Accepted: 19 December 2024;
Published: 15 January 2025.
Edited by:
Alberto Basset, University of Salento, ItalyCopyright © 2025 Xu, Gu, Yun, Gan, Fan, Chen, Shao, Zhang and Zeng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mingshan Xu, eHVtczAxMjNAMTYzLmNvbQ==; Jian Zeng, emVuZ2ppYW5Aemp3YXRlci5nb3YuY24=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.