 Leonardo D. Mendoza-González†
Leonardo D. Mendoza-González† Lucia Suárez-López
Lucia Suárez-López Carmen G. Paniagua-Chávez
Carmen G. Paniagua-Chávez- Department of Aquaculture, Centro de Investigación Científicia y de Educación Superior de Ensenada, B. C. (CICESE), Ensenada, Baja California, Mexico
The cryopreservation of cell lines such as primordial germ cells and germ cells is a promising strategy to conserve and reconstitute endangered or commercially important species in aquaculture. In Mexico, the northwest region is the center of the country’s most significant fishing and aquaculture production. However, most of the species used in capture fishing are overexploited. Despite this, protocols for the cryopreservation of germ cells are non-existent. Therefore, this work aimed to establish a protocol of isolation, identification, and cryopreservation of germ cells in two species, totoaba (Totoaba macdonaldi) and yellowtail amberjack (Seriola lalandi). Three concentrations of trypsin (0.25%, 0.3%, and 0.5%) were tested for gonadal dissociation. The 0.3% trypsin concentration was the best because it presented the most significant number of viable cells, with 14.35 × 105 for totoaba and 2.96 × 105 for yellowtail amberjack. The immunohistochemistry identification of germ cells in both species was positive for vasa, with 33.30% for totoaba and 34.20% for yellowtail amberjack. The cryoprotectant used was ethylene glycol (1.5 M or 2 M). The ideal temperature for the cryopreservation of gonadal tissue was different for each species, −1°C/min for totoaba and −5°C/min for yellowtail amberjack with 58.42% and 63.48% viable cells after thawing, respectively, with ethylene glycol 1.5 M being the best for both species. The non-controlled rate was the most effective technique to freeze the cell suspension, with 4.20 ± 1.09 × 105/mL viable cells for totoaba and 7.31 ± 2.25 × 105/mL for yellowtail amberjack. In conclusion, the results of the isolation, identification, and cryopreservation protocols for germ cells in totoaba and yellowtail amberjack obtained in this work are the first report for fish species from northwest Mexico, opening the door for the generation of cryobanking of germ cells. Finally, this work would help conserve endangered species and be an alternative to conserving species of commercial importance in aquaculture.
1 Introduction
Aquaculture is the food production activity that has expanded the most in recent decades, with the aim of ensuring the growing demand for food that exists worldwide (FAO, 2022). For this reason, the development of research, reproductive biotechnology, and conservation of genetic resources plays an important role in the development of aquaculture. One of the cutting-edge research projects related to reproductive biotechnology has been the cryopreservation of germplasm of species of high commercial value, endangered species, and model genetic lines for research or zootechnical potential (Cabrita et al., 2010; Martínez-Páramo et al., 2017). The cryopreservation of genetic material has reached great importance since this technique allows it to indefinitely safeguard material of high genetic value (Labbé et al., 2013; Marinović et al., 2019). In fish, the number of protocols for the cryopreservation of sperm, germ cell lines, and somatic cells has increased due to the high demand and use of fishing resources (FAO, 2022; Mayer and Pšenička, 2024). In fish, there are innumerable protocols for sperm cryopreservation (Asturiano et al., 2017; Martínez-Páramo et al., 2017). However, species reconstitution through androgenesis using cryopreserved sperm is not the ideal technique to be used in species conservation programs since, through this cell type, it is not possible to transfer genetic material such as mitochondrial DNA (Yasui et al., 2010; Martínez-Páramo et al., 2017).
Consequently, the cryopreservation of cell lines, such as primordial germ cells (PGCs) and germ cells (spermatogonia or oogonia), is used in the conservation and reconstitution of species since they are precursors of gametes, and through them, it is possible to transmit genetic information to subsequent generations (Braat et al., 1999; Cinalli et al., 2008). Furthermore, these cells have high sexual plasticity (differentiating eggs and sperm) and the ability to colonize and proliferate the gonad of allogeneic or xenogeneic recipients (Okutsu et al., 2007; Nóbrega et al., 2010; Hattori et al., 2019). The protocols for the cryopreservation of PGCs and germ cells (GCs) from different fish species have been standardized to safeguard genetic material, allowing the future reconstitution of a species in the event of extinction (Table 1). The results so far have shown that the protocols are species-specific and can often also be cell-specific (Table 1).
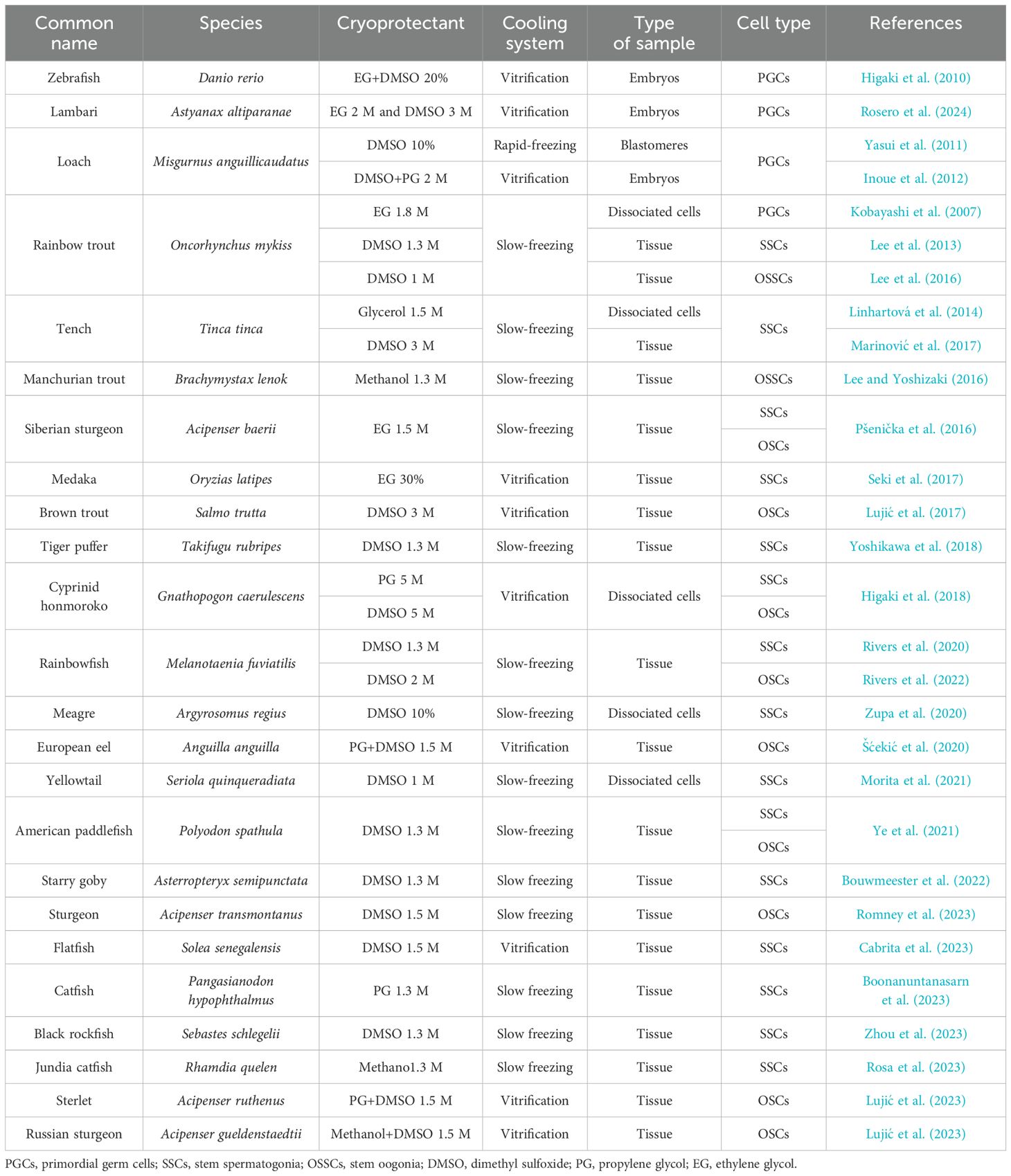
Table 1. The following list describes some cryopreservation protocols for teleost fish established in the last 15 years, indicating the types of cryoprotectants, freezing method, type of sample, and type of cells cryopreserved.
These properties make the GCs suitable for cryopreservation either as male or female germplasm and open the door to new biotechnologies: breeding programs such as GC transplantation and GC culture. The first genetic cryobanks were established in model species such as trout (Oncorhynchus mykiss) (Yoshizaki et al., 2011), tilapia (Oreochromis niloticus) (Lacerda et al., 2010), and zebrafish (Danio rerio) (Higaki et al., 2010). In these species, cryopreservation of GCs and cell transplantation for sterile recipients were associated, resulting in a new method for the preservation and reconstitution of species. The success obtained in these species served as a starting point to establish genetic cryobanks of endangered species such as the Manchurian trout (Brachymystax lenok) (Lee and Yoshizaki, 2016), sturgeon (Acipenser transmontanus) (Romney et al., 2023), and cyprinid honmoroko (Gnathopogon caerulescens) (Higaki et al., 2017), obtaining offspring from sterile recipient animals or through the production of functional spermatozoa by in-vitro culture.
In Mexico, the northwest region plays a very important role in the country’s fishing and aquaculture production, with 82% of the total national production (SADER, 2022). Therefore, it is necessary to start developing biotechnological conservation strategies such as the creation of genetic cryobanks. According to the Official Gazette of the Federation, 13.9% of the species used in capture fishing are overexploited (SADER, 2022; Oficial, 2024). That is why in this work cryopreservation protocols for GCs were developed and standardized in totoaba (Totoaba macdonaldi) and yellowtail amberjack (Seriola lalandi), two species of great economic importance in aquaculture in northwest Mexico. The totoaba, a species endemic to the Gulf of California, is critically endangered due to illegal overfishing, as the fish’s swim bladder is considered an aphrodisiac dish (Cisneros-Mata et al., 2021). The totoaba has a high productive potential in aquaculture due to its large size and rapid growth, but its sexual maturity takes up to 3 years in captivity and 5 years in the wild (Findley, 2010). On the other hand, a circumglobal species, yellowtail amberjack, is a highly farmed fish in México and is wanted by commercial fisheries and sport fishers around the world (Smith-Vaniz and Williams, 2015).
Therefore, the main objective of this study was to develop a method to dissociate gonadal tissue, identify GCs, and subsequently cryopreserve them using different freezing methods. In this work, two species were selected: the totoaba represents an endemic organism in danger of extinction and with high productive potential, while the yellowtail amberjack is a species of high economic importance at the international level. Conserving their GCs and standardizing the protocols for this purpose will allow for the establishment of new conservation, reproduction, and genetic improvement programs, as well as establishing the bases for standardizing protocols in other phylogenetically close species.
2 Materials and methods
2.1 Ethics
The experiments were carried out following the guide for the use of fish in research by Jenkins et al. (2014) in the chapter “5.4.1. Drugs Approved for Use on Fish.” The animals used for blood collection were anesthetized using a concentration of 50 mg/L of tricaine metasulfonate (MS-222) diluted in 90 L of seawater and constant oxygenation. After 2 to 5 min, the animals lost balance and the flight reflex, and the experimental procedures were performed immediately. The animals used for extraction of gonadal tissue were subjected to euthanasia by the physical method, performing a craniocerebral puncture, and subsequently decapitation was performed. All procedures were approved by the Bioethics Committee of Centro de Investigación Científica y de Educación Superior de Ensenada, BC (CICESE) opinion No. ORGA_ACUA_2020.3. The animals used in this work were obtained from the Marine Fish Laboratory of the Aquaculture Department of CICESE.
2.2 Fish husbandry
In this work, the number of fish used was as follows: n = 35 for totoaba (955 ± 300 g) and n = 45 for yellowtail amberjack (399 ± 147 g). All animals were sexually immature juveniles (1 year). The animals were maintained in a recirculating aquaculture system (RAS), which consisted of a 3,000-L culture tank coupled to a 640-L surge tank and a bubble-washed bead filter with a capacity of 1.13 L of medium composed of plastic beads (BBF-XS-8000, Aquaculture Systems Technologies, Louisiana, EUA). The water flow (160 L/min) was given by a centrifugal pump (0.5 HP, Sweetwater). The maintenance parameters were controlled under the following levels: temperature of 26°C for totoaba (T. macdonaldi) and 22°C for yellowtail amberjack (S. lalandi), salinity of 35 ppm, total ammonia nitrogen <1.0 mg/L, nitrites <1.0 mg/L, nitrates <400 mg/L, pH 7.6–8.5, and photoperiod 12:12. The feeding of the organisms was carried out with 3% of the total biomass with dry matter per day, with a commercial diet (Skretting and Ewos Cargill). Feeding was stopped 24 h prior to each experiment.
2.3 Blood sampling
Fish were anesthetized using 50 mg/L of MS-222 until loss of escape reflex, as seen in Section 2.1. Samples were obtained from the caudal vein, and the blood plasma was separated by centrifugation (Centrifuge Eppendorf 5430, Hamburg, Germany) at 500×g for 30 min. Osmolality was measured using an osmometer (Vapro® 5520, Wescor, Logan, UT, EUA). All suspensions were adjusted to the mean value of osmolality for each species.
2.4 Gonadal extraction
Three fish were euthanized before each experiment (Section 2.1). The gonad was obtained under aseptic conditions according to the methodology described by Lacerda et al. (2018). The extraction was performed aseptically and rinsed with 0.1% antibiotic and antimycotic (Sigma-Aldrich #A5955, St. Louis, USA) in Hank’s Balanced Saline Solution (HBSS). Then, the gonads were weighted and cut into small pieces (~1 mm3).
2.5 Enzymatic dissociation
Enzymatic dissociation was performed following the protocol of Pšenička et al. (2015) with some modifications. Each gonad fraction was submerged in 900 μL of enzymatic media and incubated for 2.5 h in a bioshaker (Thermo Scientific, Waltham, Massachusetts, EUA #4625) at 200 rpm. The enzymatic media consisted of 0.25%, 0.3%, or 0.5% trypsin (Sigma-Aldrich #59427C, St. Louis, USA) and 0.05% DNAse in HBSS. In the case of control, HBSS without enzymes was used. One hundred microliters of 10% bovine serum albumin (BSA) (Sigma-Aldrich #A-7906, St. Louis, USA) was added to stop the trypsin activity. Then, the gonads were filtered through a 40-μm cell strainer, pushing the tissue rest with HBSS before centrifugation at 500×g for 15 min at 4°C. The pellets were resuspended in 100 μL of 0.75% BSA in HBSS.
2.6 Discontinuous Percoll gradient
The densities were made including the Percoll concentrations that have been used to enrich CGs in a gonad suspension from other fish (Lacerda et al., 2018; Pšenička et al., 2015). To enrich GCs in suspension, 40% and 10% discontinuous Percoll® gradients in HBSS were prepared and put in that order at 3 mL each. The pellet recovered from the enzymatic dissociation was carefully added on the top discontinuous Percoll® gradient and centrifugated at 500×g and 4°C with slow rotor acceleration for 30 min. The 10% layer was recovered and washed, adding HBSS 1:10 and centrifuging for 30 min. The pellet of the best treatment was used for identification by immunocytochemistry and cryopreservation.
2.7 Immunocytochemistry
The identification of GCs was performed by immunocytochemistry using vasa. The protocol used was modified from Pšenička et al. (2015) and Escuredo-Vielba et al. (2018) for the purposes of this study. The 10% Percoll® layer from Section 2.6 was fixed in 1:1 of 4% paraformaldehyde (PFA) (Sigma-Aldrich #158127, St. Louis, USA) in PBS (Bio Basic #PD8117, Canada INC) and put on antibody slides with two 15-mm rings, previously incubated in 0.1% poly-L-lysine (Sigma-Aldrich #P8920, St. Louis, USA). After 3 h of cell sedimentation, the slides were washed once with PBS. The cell permeabilization was performed by adding 3.0% Triton®X-100 (Sigma-Aldrich, St. Louis, USA) in PBS at 4°C for 15 min and then washed three times in PBS. Samples were blocked with 1% BSA and 0.05% Tween®20 (Sigma-Aldrich #P2287, St. Louis, USA) in PBS for 1 h at 4°C, and rabbit anti-vasa primary antibody (Anti-DDX4/MVH, Abcam, ab13840) (1:300) was added during 18 h, except the negative control group, and washed three times for 5 min with blocking solution before incubation with a secondary goat anti-rabbit Alexa Fluor 488 antibody (Molecular Probes, Carlsbad, CA, USA) (1:800). Samples were washed three times for 5 min with PBSS, counterstained with 10 μg/mL of nucleus staining, 4′,6-diamidino-2-fenilindol DAPI (Sigma-Aldrich #D9542, St. Louis, USA), washed five times, and rinsed. Negative control samples were mounted with Sigma Mountain media. All samples were visualized after drying with an Olympus FV1000 confocal microscope. Vasa-positive cells were counted following the protocol described below (Section 2.8).
2.8 Viability
Cell viability was performed using the kit fluorescein diacetate (FDA)/propidium iodide (PI) staining (Logos Biosystems, EUA #F23214). Double staining was then performed with 2.4 mM of FDA for 5 min and then 7.4 mM of PI for 5 min. The evaluation of viable and non-viable cells was performed using a hemocytometer. Cells were counted in 8 squares of the hematocytometer in triplicate in a fluorescence microscope (Nikon H600L Eclipse 80i Nikon Co. Tokyo, Japan) using a blue 480/30 nm excitation filter.
2.9 Whole tissue cryopreservation
The gonad of three fish of each species was removed (Section 2.4). Each gonad (n = 3) was washed in PBS (383.33 ± 5.13 mOsm/kg for totoaba and 417.66 ± 11.93 mOsm/kg for yellowtail amberjack) and sliced into four equal portions (~1 mm3). Then, gonad portions were submerged in an extender solution composed of 0.5% BSA, 0.1% antibiotic-antimycotic (10,000 UI of penicillin and 10 mg/mL of streptomycin), 50 mM of glucose (Sigma-Aldrich #S0389, St. Louis, USA), and as a cryoprotectant 1.5 M or 2 M of ethylene glycol (EG) (Sigma-Aldrich #102466, St. Louis, USA) during 10 min in 1.5 mL cryovials. The freezing protocol, according to Pšenička et al. (2016), started at 10°C for 10 min and then frozen at −1°C/min or −5°C/min per minute cooling rate (CR) in a programmable freezer (Kryo 560-16, Planer PLC) until reaching −80°C and plunged into liquid nitrogen (−196°C) for 7 days. As a control group, the gonad of an animal was cryopreserved without any cryoprotectant. Cryovials were thawed in a water bath (Memmert #WNB-22, Germany) at 25°C for 1 min. The tissue portions were washed (two times), subsequently subjected to the enzymatic dissociated (Section 2.5), and enriched by Percoll® density gradient (Section 2.6). The viability was evaluated following the procedure in Section 2.8.
2.10 Cell suspension cryopreservation
The gonad of three fish of each species was removed (Section 2.4), dissociated (Section 2.5), and enriched by Percoll® density gradient (Section 2.6). The pellet obtained after CG enrichment was suspended in an extender solution with 1.5 M of EG as cryoprotectant and incubated for 10 min in 250 µL French straws (3 straws per repetition). The cryoprotectant solution was selected because it showed the best results in tissue cryopreservation. The straws were sealed with blue sealing powder (IMV Technologies, #018818, L´Aigle, France) and placed in the programmable freezer chamber for controlled cooling rates (−1°C/min or −5°C/min) until reaching −80°C (Pšenička et al., 2016) or in a polystyrene box (27.5 cm × 20.5 cm) for a non-controlled cooling rate (Lacerda et al., 2010). The straws were placed in a net 15 cm above liquid nitrogen and moved 1 cm toward the liquid nitrogen until reaching −80°C. As a negative control, the samples were directly plunged into liquid nitrogen, and as a positive control, the viability of the cell suspension was evaluated before freezing. After cooling, the straws were plunged into liquid nitrogen (−196°C) for 7 days. Thawing of the samples was performed as described in Section 2.9. The straws were emptied into 2.0 mL conical tubes, and cells were suspended 1:4 in HBSS. The viability was evaluated following the procedure in Section 2.8. The stages of enzymatic dissociation, identification, and freezing of germ cells in totoaba and yellowtail amberjack are represented in Figure 1.
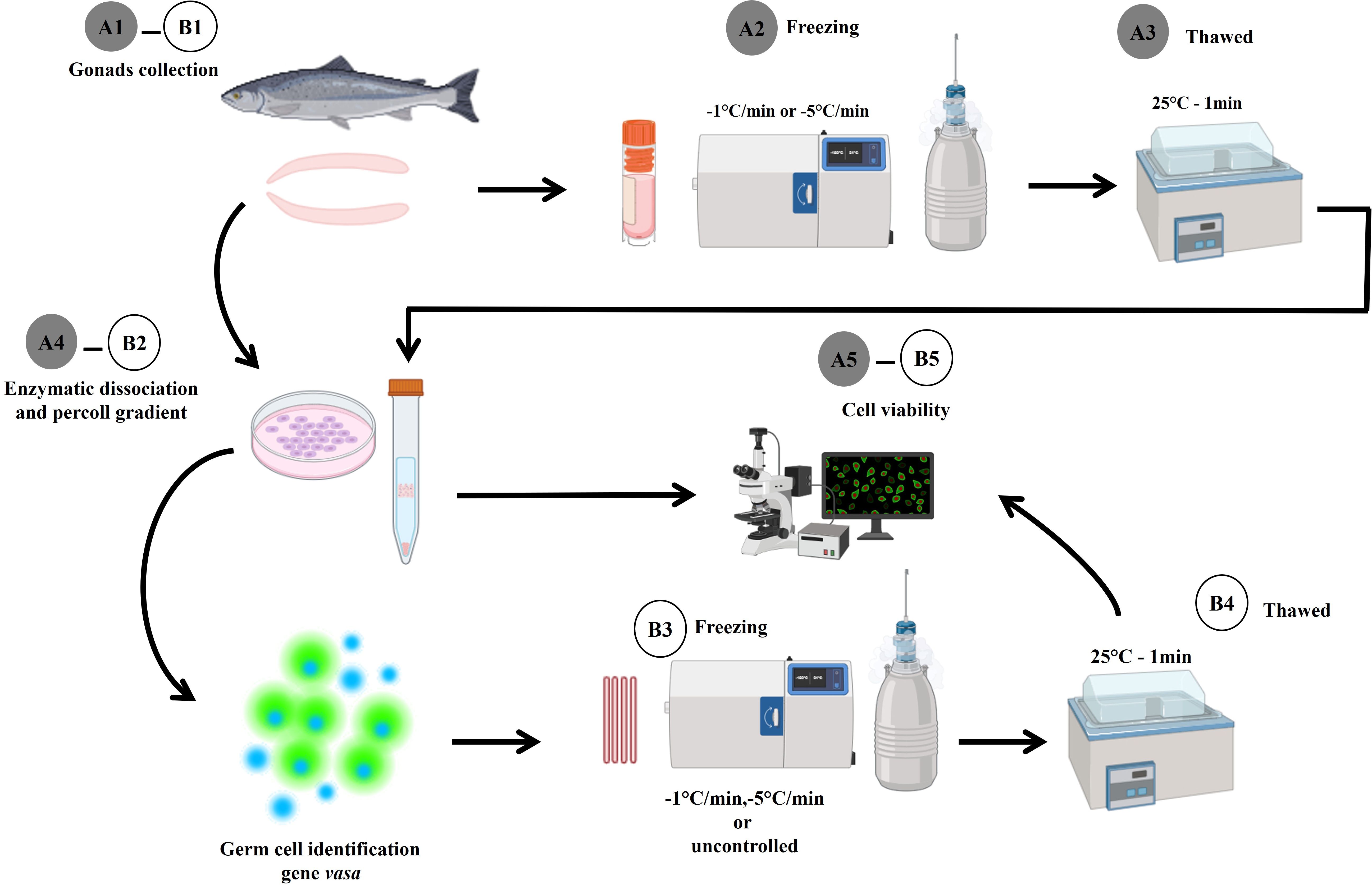
Figure 1. An illustrated methodology scheme of how the cryopreservation of germ cells in Totoaba macdonaldi and Seriola lalandi was carried out. Cryopreservation of gonadal tissue. (A1) Collection and fraction of gonadal tissue for cryopreservation. (A2) Exposure to cryoprotectant and controlled freezing. (A3) Thawing of gonadal tissue. (A4) Enzymatic dissociation and cell separation by Percoll® gradient. (A5) Cell viability of previously cryopreserved gonadal tissue cells. Cryopreservation of enriched germ cell suspension. (B1) Gonad collection. (B2) Enzymatic disintegration and cell separation by Percoll® density gradient. (B3) Cell purification and subsequent cryopreservation of germ cells using 1.5 M of ethylene glycol and controlled (−1°C/min and −5°C/min) and non-controlled cooling rates. (B4) Thawing of germ cells. (B5) Cell viability after thawing.
2.11 Data analysis
Statistical analysis was performed using Minitab v 18 software for Windows®. Percentage data were transformed into arcsine square roots before data analysis. A one-way ANOVA was used to test the effect of enzymatic dissociation or the cooling rates (controlled or non-controlled) on cell suspension. Two-way ANOVA was used to assess the viability of whole tissue suspended in two different cryoprotectants and cooling rates. Specific differences were tested with Tukey’s post-hoc test. A P-value <0.05 was used as the significance level.
3 Results
The physicochemical characteristics of the blood plasma in the species were as follows: osmolality of the blood plasma of 383.33 ± 5.13 mOsm/kg (mean ± SD) with a pH of 7.07 ± 0.34 for totoaba and that for yellowtail amberjack was 417.66 ± 11.93 mOsm/kg with pH 6.90 ± 0.05. The viability of the GCs did not show statistical differences between trypsin concentrations for both species (Figures 2, 3). The highest disaggregated cell viability for totoaba and yellowtail amberjack was found using 0.25% trypsin, with 78.42% and 64.23% cell viability, respectively (Figure 3). Nevertheless, all trypsin treatments were different from the control. Significant differences were found in the concentration of viable cells disaggregated from totoaba to the control (P = 0.00). The highest number of viable cells 14.35 × 105 was found in the 0.3% trypsin treatment (Figure 3). The concentration of viable cells in yellowtail amberjack had no significant differences (P = 0.11) between treatments to dissociate the tissue; the number of viable cells was 2.96 × 105 (Figure 3). The 0.3% concentration showed less debris quantity in both species. The observed vasa signal-positive cells were 33.33% for totoaba and 34.16% for yellowtail amberjack in the 10% Percoll® layer (Figure 4).
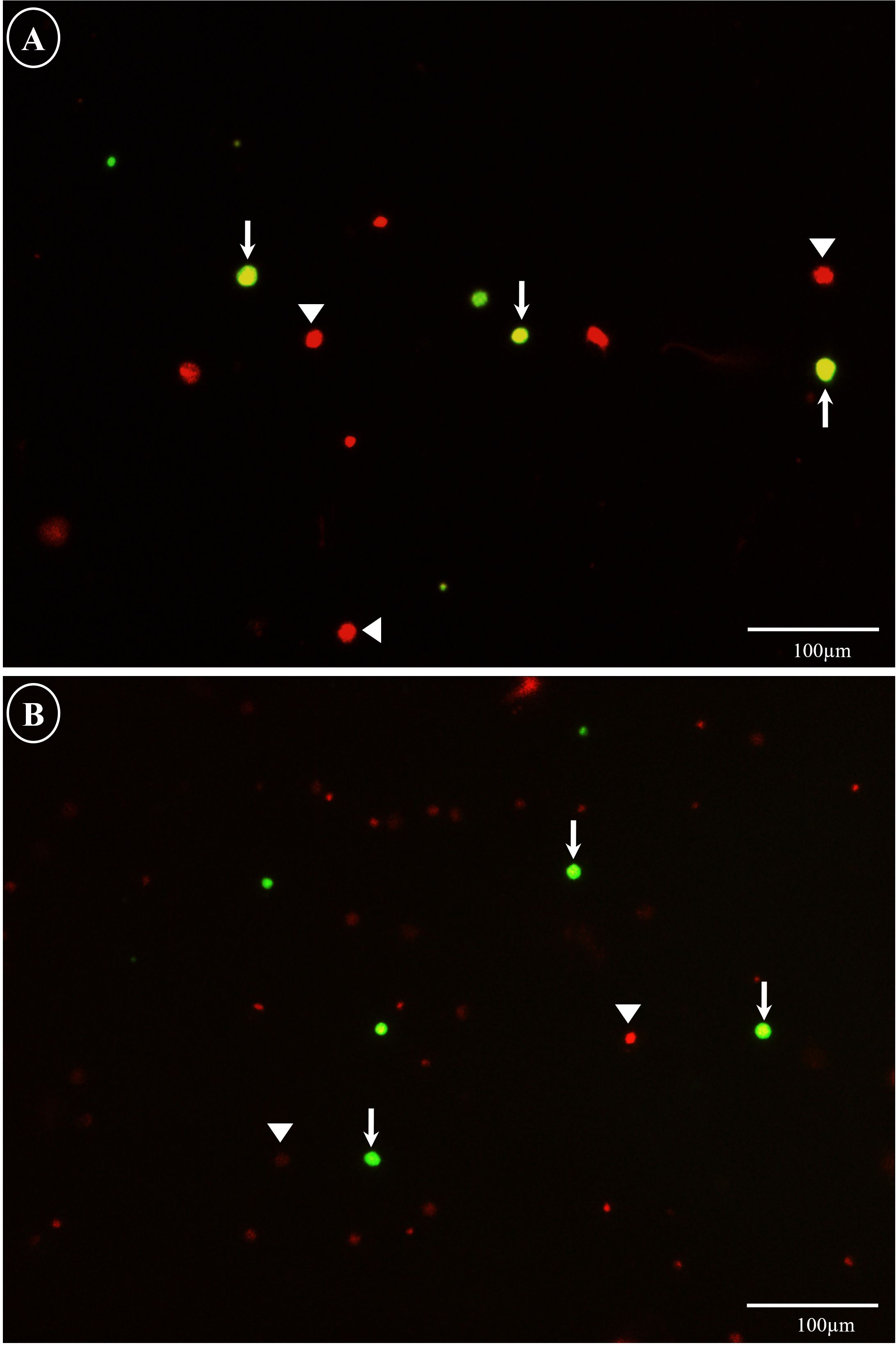
Figure 2. Cell viability analysis of germ cells in Totoaba macdonaldi (A) and Seriola lalandi (B). Viability was assessed using a double stain of fluorescein diacetate (FDA) and propidium iodide (PI). Arrows indicate viable cells (green) and triangles indicate non-viable cells (red). Scale: (A) 100 µm.
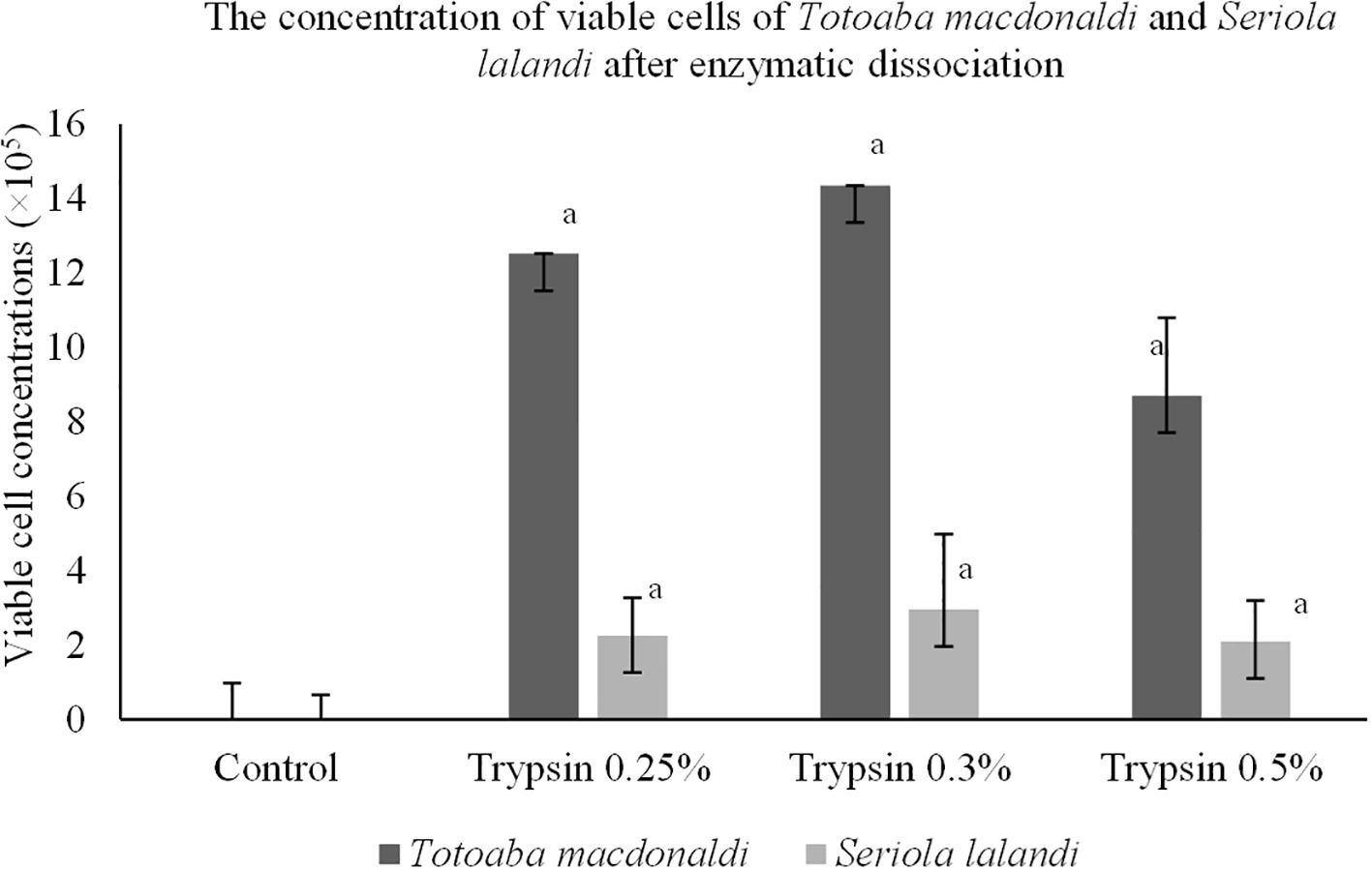
Figure 3. The concentration of viable cells of Totoaba macdonaldi and Seriola lalandi after enzymatic dissociation using different concentrations of trypsin. The values correspond to the mean ± standard deviation. Letters of statistical significance are grouped based on Tukey’s post-hoc analysis (P = 0.05). The same letter means that there are no statistically significant differences.
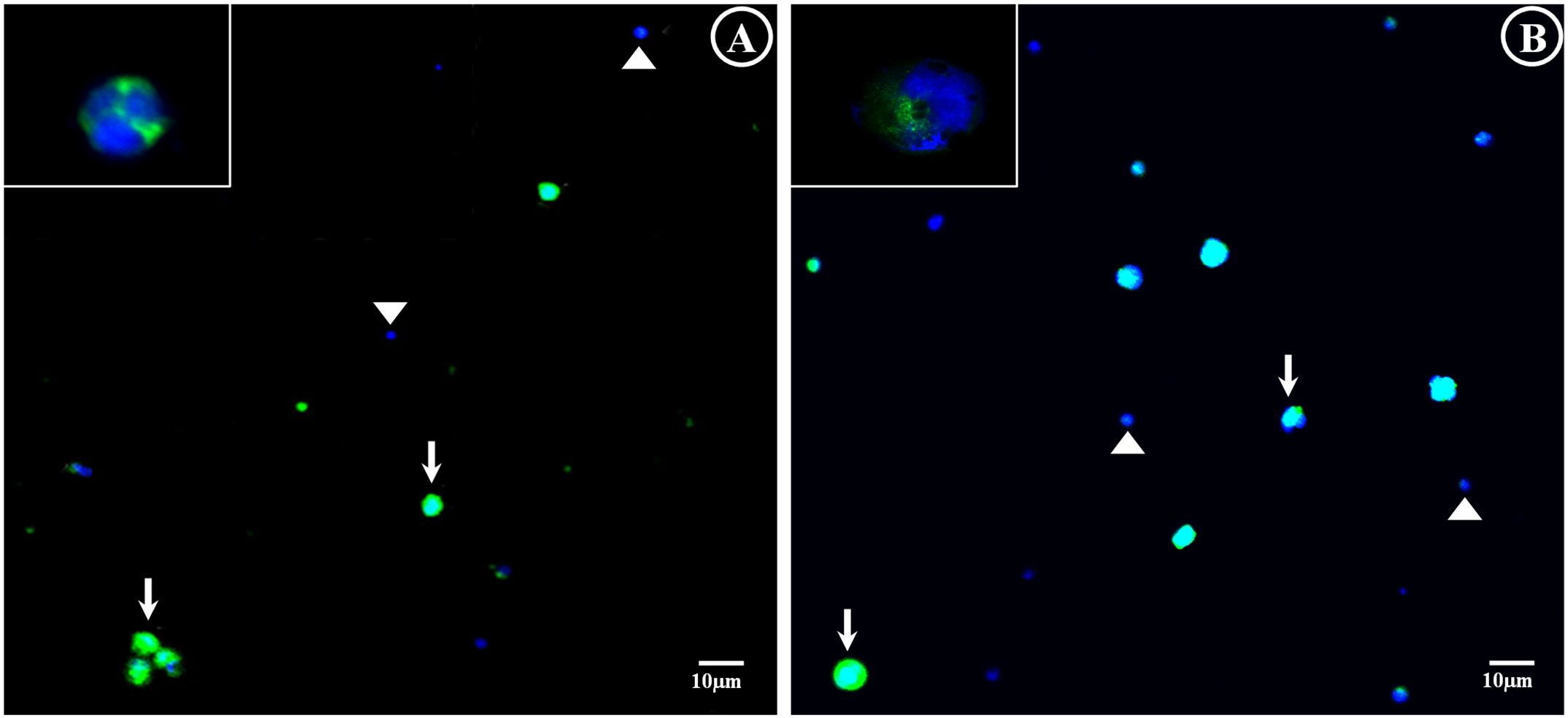
Figure 4. Fluorescent micrograph of germ cells stained with anti-vasa conjugated with Alexa Fluor 488 and DAPI. (A) Totoaba macdonaldi and (B) Seriola lalandi. Cells positive for vasa expression stained green in their cytoplasm and blue in their nuclei (arrow). Somatic cells can be seen with the blue staining of their nuclei (triangle). A vasa-positive cell is observed at higher magnification in the upper left part. Scale: (A) 10 µm.
Two-way ANOVA revealed no significant differences in the percentage of viable cells (>50%) for thawed gonadal tissue regarding EG concentration (P = 0.67 for totoaba and P = 0.61 for yellowtail amberjack) or CR (P = 0.10 for totoaba and P = 0.14 for yellowtail amberjack). The best CR was −1°C/min in totoaba, which allows 58.42% of viable cells, opposite with yellowtail amberjack, whose best CR was −5°C/min with 63.48% of viable cells (Figure 5A).
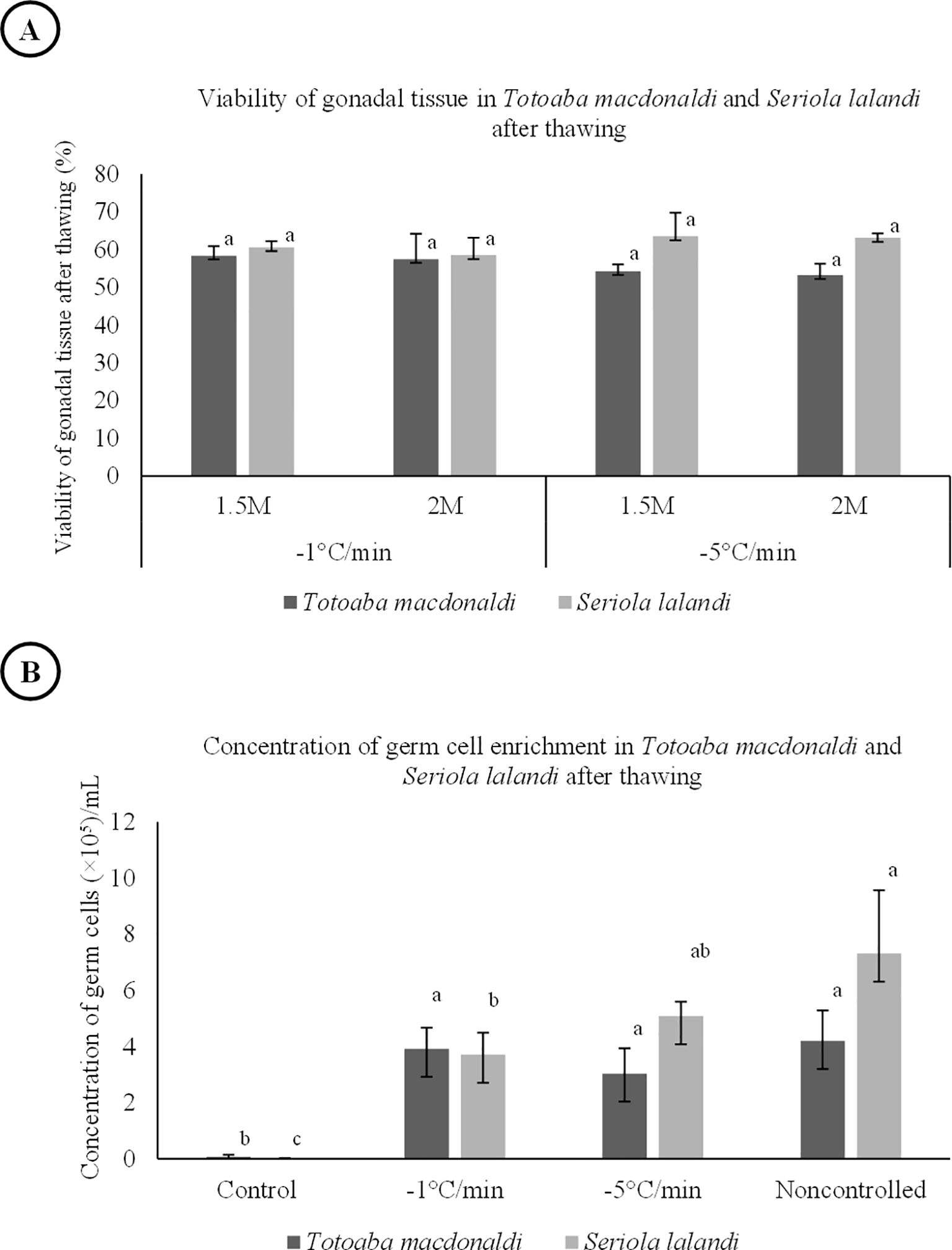
Figure 5. Viability of gonadal tissue in Totoaba macdonaldi and Seriola lalandi after thawing (A). Concentration of germ cell enrichment in Totoaba macdonaldi and Seriola lalandi after thawing (B). The values correspond to the mean ± standard deviation. Letters of statistical significance are grouped based on Tukey’s post-hoc analysis (P = 0.05). The same letter means that there are no statistically significant differences.
The enriched suspension of germ cells in both species shows high viability after thawing, between 38.50% and 47.37% viable cells for totoaba and 18.30% and 39.77% for yellowtail amberjack. Cell contraction was 4.20 ± 1.09 × 105 cel/mL for totoaba and 7.31 ± 2.25 × 105 cel/mL for yellowtail amberjack, obtaining better results with the non-controlled treatment. However, the ANOVA did not show significant differences among treatments except for the control (P = 0.00) (Figure 5B).
4 Discussion
In the present work, a successful protocol for enzymatic dissociation, identification, and cryopreservation of GCs was established for the first time in two species of ecological and economic importance from Mexico, the totoaba and yellowtail amberjack. The 0.3% trypsin concentration was the best for carrying out both species’ enzymatic dissociation of the gonadal tissue, obtaining the highest purity, concentration, and viability of GCs. Gonadal dissociation using trypsin has already been used in other species such as Siberian sturgeon (Acipenser baerii) (Pšenička et al., 2015), pejerrey (Odontesthes bonariensis) (Majhi et al., 2009), and goldfish (Carassius auratus) (Majhi, 2023), managing to maintain the proliferation capacity in exogenous gonads after germ cell transplant. In Siberian sturgeon (A. baerii), cell proliferation was observed up to 90 days post-transplant in 60% of the recipients. Meanwhile, in pejerrey (O. bonariensis) and goldfish (C. auratus), it was possible to obtain offspring from the donor species of cells with 13.3% and 100% of the recipients, respectively. However, incubation with trypsin in-vitro culture in rainbow trout (O. mykiss) spermatogonia revealed a rupture of membrane proteins, causing a reversible decrease in the mitotic activity of the cells (Shikina et al., 2013). Nevertheless, in this study, incubation with trypsin was shown to be efficient for gonadal dissociation in both species. However, further studies are needed regarding the effect of trypsin treatment on GCs in totoaba and yellowtail amberjack and their possible in-vitro recovery as well as transplantation experiments.
Enrichment of a suspension with GCs, by using a discontinuous density gradient, proved to be a low-cost and efficient technique. In this study, more than 30% GCs were obtained in the 10% Percoll concentration layer in the two species used in this study. This same concentration of Percoll is recommended by various authors because the cells with the highest density, usually blood cells, are found in the precipitate with the highest concentration, which in this study was 40%, and it has been observed that the supernatant consists mainly of cellular debris and tissues (Lacerda et al., 2018, 2010; Linhartová et al., 2014; Pšenička et al., 2015). For this reason, the 10% Percoll precipitate was used to perform the identification with immunocytochemistry.
The vasa gene has been used as a germ cell line labeling in many teleost species (Higuchi et al., 2011; Ricci et al., 2018; Wu et al., 2018; Rosero et al., 2023), allowing the identification of cells during embryonic development, in-vitro cultures, or cell transplants. Using immunocytochemistry, we demonstrated that differentiated GCs of totoaba and yellowtail amberjack are signal-positive to vasa in non-mature organisms. Escuredo-Vielba et al. (2018) conducted immunostaining using the whole-mount technique and found the presence of vasa-positive signal PGCs in two- and four-cell stage totoaba embryos using the same labeling. In species such as yellowtail (Seriola quinqueradiata), sturgeon (A. baerii), and tench (Tinca tinca), positive expression of the vasa gene has been used to confirm the presence of GCs in the isolated cell suspension, obtaining success in the transplantation of germ cells (Linhartová et al., 2014; Pšenička et al., 2015; Morita et al., 2021).
The results obtained in the cryopreservation protocols are species-specific since the ideal temperature for cryopreservation of gonadal tissue was different for each species, −1°C/min for totoaba and −5°C/min for yellowtail amberjack. However, for cryopreserving cell suspension, the non-controlled rate was the most effective technique. In fact, the results indicate that after each critical process, totoaba and yellowtail amberjack GCs have positive esterase activity, as well as integrity in their cytoplasmic membranes. These properties suggest that the use of germ cells can be further studied with future work based on the present, such as in-vitro cell culture and development of transplant protocols. Using cell culture, it would be possible to assess whether cells from previously frozen tissue or cell suspension have the capacity to perform mitosis in vitro (Lacerda et al., 2010) or proliferate the gonads of sterile recipients by cell transplantation, successfully achieving offspring from cell donors (Yoshizaki and Lee, 2018).
Cryopreservation of gonadal tissue and cell suspension in other species, such as sturgeon (A. baerii), goldfish (C. auratus), and tench (T. tinca), was achieved successfully, and no significant differences were found between both techniques (Pšenička et al., 2016; Marinović et al., 2017). However, the results were different in neotropical catfish (Rhamdia quelen). Tissue cryopreservation yields better viable cells (67%) after thawing using 1.4 M of DMSO as a cryoprotectant when compared to cell suspension cryopreservation (27% viable cells) after thawing in 1.3 M of DMSO (Rosa et al., 2023). These results suggest that it is fundamental to develop species-specific protocols, in addition to specific techniques, from dissociation to cryopreservation, as well as thawing and viability tests (Pšenička et al., 2016). However, protocols for gonadal tissue cryopreservation have increased (Table 1) due to the advantages it can offer, such as 1) it can be carried out under field conditions using vitrification as a freezing method; 2) there is better purification of viable cells since, when the tissue is thawed, it goes through the enzymatic disintegration process and the activity of enzymes that dissociate gonadal tissue helps eliminate weakened cells, sometimes observing agglutinations during the dissociation process (Marinović et al., 2017); and finally, 3) the solution supplemented during the dissociation process would help eliminate the remnants of the cryoprotective solution.
In conclusion, the results of the isolation, identification, and cryopreservation protocols for GCs in totoaba and yellowtail amberjack obtained in the present work would be the first report for fish species from northwest Mexico, opening the door for the generation of cryobanking of GCs. However, new work is necessary to confirm the proliferative and differentiation capacity of GCs after the thawing process. Finally, this work would help conserve high genetic value material from species in danger of extinction or of commercial importance in aquaculture because our results indicate that cryopreservation of GCs can be a valuable tool to implement future species reconstitution actions such as in-vitro culture and germ cell transplantation.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by the Bioethics Committee of Centro de Investigación Científica y de Educación Superior de Ensenada, BC (CICESE) opinion No. ORGA_ACUA_2020.3. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
LM: Writing – original draft, Methodology, Investigation, Formal analysis, Data curation, Conceptualization. LS: Writing – review & editing, Writing – original draft. CP: Writing – review & editing, Writing – original draft, Validation, Supervision, Resources, Project administration, Funding acquisition.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by CICESE project No. 623142 and by a master’s degree grant from the National Council for Humanities, Science and Technology (CONAHCYT).
Acknowledgments
We thank C. M. Ochoa-Saloma, P. González, and S. Villasuso for their technical assistance.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Asturiano J. F., Cabrita E., Horváth (2017). Progress, challenges and perspectives on fish gamete cryopreservation: A mini-review. Gen. Comp. Endocrinol. 245, 69–76. doi: 10.1016/j.ygcen.2016.06.019
Boonanuntanasarn S., Sreebun S., Booncherd K., Khaosa-art P., Sooksawat T., Ichida K., et al. (2023). Cryopreservation of testicular cell in striped catfish (Pangasianodon hypophthalmus) and its effects on apoptosis, germ-cell specific gene expression and germ cell transplantability. Aquaculture 570. doi: 10.1016/j.aquaculture.2023.739370
Bouwmeester J., Daly J., Henley E. M., Parenti L. R., Pitassy D. E., Hagedorn M. (2022). Conservation of coral reef fishes: a field-hardy method to cryopreserve spermatogonial cells. Coral Reefs 41, 855–861. doi: 10.1007/s00338-022-02268-1
Braat A. K., Speksnijder J. E., Zivkovic D. (1999). Fish germline.Pdf. Int. J. Dev. Biol. 760, 745–760. Available at: www.ehu.es/ijdb.
Cabrita E., Pacchiarini T., Fatsini E., Sarasquete C., Herráez M. P. (2023). Post-thaw quality assessment of testicular fragments as a source of spermatogonial cells for surrogate production in the flatfish Solea Senegalensis. Fish Physiol. Biochem. doi: 10.1007/s10695-023-01232-2
Cabrita E., Pacchiarini T., Sarasquete C., Herraez M. P. (2010). 141. Development of a cryopreservation protocol for Senegalese sole testicular germ cells. Cryobiology 61, 405. doi: 10.1016/j.cryobiol.2010.10.145
Cinalli R. M., Rangan P., Lehmann R. (2008). Germ cells are forever. Cell 132, 559–562. doi: 10.1016/j.cell.2008.02.003
Cisneros-Mata M. Á., True C., Enriquez-Paredes L. M., Sadovy Y., Liu M. (2021). Totoaba macdonaldi. The IUCN Red List of Threatened Species 2021: e.T22003A2780880. doi: 10.2305/IUCN.UK.2021-2.RLTS.T22003A2780880.en (accessed on 27 September 2024).
Escuredo-Vielba R., Del Río-Portilla M. A., Mata-Sotres J. A., Barón-Sevilla B., Guerrero-Rentería Y., Paniagua-Chávez C. G. (2018). Characterization and localization of primordial germ cells in Totoaba macdonaldi. Comp. Biochem. Physiol. Part - B: Biochem. Mol. Biol. 225, 29–37. doi: 10.1016/j.cbpb.2018.07.001
FAO. (2022). The state of world fisheries and aquaculture 2022: towards blue transformation. (Rome: FAO). doi: 10.4060/cc0461en
Findley L. (2010). Totoaba macdonaldi. IUCN Red List Threatened Species 8235, 8. doi: 10.2305/IUCN.UK.2010-3.RLTS.T22003A9346099.en
Hattori R. S., Yoshinaga T. T., Katayama N., Hattori-Ihara S., Tsukamoto R. Y., Takahashi N. S., et al. (2019). Surrogate production of Salmo salar oocytes and sperm in triploid Oncorhynchus mykiss by germ cell transplantation technology. Aquaculture 506, 238–245. doi: 10.1016/j.aquaculture.2019.03.037
Higaki S., Eto Y., Kawakami Y., Yamaha E., Kagawa N., Kuwayama M., et al. (2010). Production of fertile zebrafish (Danio rerio) possessing germ cells (gametes) originated from primordial germ cells recovered from vitrified embryos. REPRODUCTION 139, 733–740. doi: 10.1530/REP-09-0549
Higaki S., Shimada M., Kawamoto K., Todo T., Kawasaki T., Tooyama I., et al. (2017). In vitro differentiation of fertile sperm from cryopreserved spermatogonia of the endangered endemic cyprinid honmoroko (Gnathopogon caerulescens). Sci. Rep. 7, 1–14. doi: 10.1038/srep42852
Higaki S., Todo T., Teshima R., Tooyama I., Fujioka Y., Sakai N., et al. (2018). Cryopreservation of male and female gonial cells by vitrification in the critically endangered cyprinid honmoroko Gnathopogon caerulescens. Fish Physiol. Biochem. 44, 503–513. doi: 10.1007/s10695-017-0449-x
Higuchi K., Takeuchi Y., Miwa M., Yamamoto Y., Tsunemoto K., Yoshizaki G. (2011). Colonization, proliferation, and survival of intraperitoneally transplanted yellowtail Seriola quinqueradiata spermatogonia in nibe croaker Nibea mitsukurii recipient. Fisheries Sci. 77, 69–77. doi: 10.1007/s12562-010-0314-7
Inoue D., Fujimoto T., Kawakami Y., Yasui G. S., Yamaha E., Arai K. (2012). Vitrification of primordial germ cells using whole embryos for gene-banking in loach, Misgurnus anguillicaudatus. J. Appl. Ichthyology 28, 919–924. doi: 10.1111/jai.12058
Jenkins J. A., Bart H. L., Bowker J. D., Bowser P. R., MacMillan J. R., Nickum J. G., et al. (2014). Guidelines for Use of Fishes in Research—Revised and Expanded, 39. doi: 10.1080/03632415.2014.924408
Kobayashi T., Takeuchi Y., Takeuchi T., Yoshizaki G. (2007). Generation of viable fish from cryopreserved primordial germ cells. Mol. Reprod. Dev. 74, 207–213. doi: 10.1002/mrd.20577
Labbé C., Robles V., Herraez M. P. (2013). “Cryopreservation of gametes for aquaculture and alternative cell sources for genome preservation,” in Advances in aquaculture hatchery technology. (Elsevier), 76–116. doi: 10.1533/9780857097460.1.76
Lacerda S. M. S. N., Batlouni S. R., Costa G. M. J., Segatelli T. M., Quirino B. R., Queiroz B. M., et al. (2010). A new and fast technique to generate offspring after germ cells transplantation in adult fish: The nile tilapia (Oreochromis niloticus) model. PloS One 5, 1–9. doi: 10.1371/journal.pone.0010740
Lacerda S. M. S. N., Batlouni S. R., Silva S. B. G., Homem C. S. P., França L. R. (2018). Germ cells transplantation in fish: the Nile-tilapia model. Anim. Reprod. 3 (2), 146–159. Available online at: https://animal-reproduction.org/article/5b5a607ff7783717068b47c8/pdf/animreprod-3-2-146.pdf.
Lee S., Iwasaki Y., Shikina S., Yoshizaki G. (2013). Generation of functional eggs and sperm from cryopreserved whole testes. Proc. Natl. Acad. Sci. U. S. A. 110, 1640–1645. doi: 10.1073/pnas.1218468110
Lee S., Katayama N., Yoshizaki G. (2016). Generation of juvenile rainbow trout derived from cryopreserved whole ovaries by intraperitoneal transplantation of ovarian germ cells. Biochem. Biophys. Res. Commun. 478, 1478–1483. doi: 10.1016/j.bbrc.2016.08.156
Lee S., Yoshizaki G. (2016). Successful cryopreservation of spermatogonia in critically endangered Manchurian trout (Brachymystax lenok). Cryobiology 72, 165–168. doi: 10.1016/j.cryobiol.2016.01.004
Linhartová Z., Rodina M., Guralp H., Gazo I., Saito T., Pšenička M. (2014). Isolation and cryopreservation of early stages of germ cells of tench (Tinca tinca). Czech J. Anim. Sci. 59, 381–390. doi: 10.17221/7589-cjas
Lujić J., Franěk R., Marinović Z., Kašpar V., Xie X., Horváth Á., et al. (2023). Vitrification of the ovarian tissue in sturgeons. Theriogenology 196, 18–24. doi: 10.1016/j.theriogenology.2022.11.009
Lujić J., Marinović Z., Sušnik Bajec S., Djurdjevič I., Kása E., Urbányi B., et al. (2017). First successful vitrification of salmonid ovarian tissue. Cryobiology 76, 154–157. doi: 10.1016/j.cryobiol.2017.04.005
Majhi S. K. (2023). Generation of surrogate goldfish Carassius auratus progeny from common carp Cyprinus carpio parents. 3 Biotech. 13, 1–14. doi: 10.1007/s13205-022-03424-8
Majhi S. K., Hattori R. S., Yokota M., Watanabe S., Strüssmann C. A. (2009). Germ cell transplantation using sexually competent fish: An approach for rapid propagation of endangered and valuable germlines. PloS One 4, 1–8. doi: 10.1371/journal.pone.0006132
Marinović Z., Li Q., Lujić J., Iwasaki Y., Csenki Z., Urbányi B., et al. (2019). Preservation of zebrafish genetic resources through testis cryopreservation and spermatogonia transplantation. Sci. Rep. 9, 1–10. doi: 10.1038/s41598-019-50169-1
Marinović Z., Lujić J., Kása E., Bernáth G., Urbányi B., Horváth Á. (2017). Cryosurvival of isolated testicular cells and testicular tissue of tench Tinca tinca and goldfish Carassius auratus following slow-rate freezing. Gen. Comp. Endocrinol. 245, 77–83. doi: 10.1016/j.ygcen.2016.07.005
Martínez-Páramo S., Horváth Á., Labbé C., Zhang T., Robles V., Herráez P., et al. (2017). Cryobanking of aquatic species. Aquaculture 472, 156–177. doi: 10.1016/j.aquaculture.2016.05.042
Mayer I., Pšenička M. (2024). Conservation of teleost fishes: Application of reproductive technologies. Theriogenology Wild 4, 100078. doi: 10.1016/j.therwi.2024.100078
Morita T., Miwa M., Kumakura N., Morishima K., Miki T., Takeuchi Y., et al. (2021). Production of functional sperm from cryopreserved testicular germ cells following intraperitoneal transplantation into allogeneic surrogate in yellowtail (Seriola quinqueradiata). Cryobiology 100, 32–39. doi: 10.1016/j.cryobiol.2021.04.001
Nóbrega R. H., Greebe C. D., van de Kant H., Bogerd J., de França L. R., Schulz R. W. (2010). Spermatogonial stem cell niche and spermatogonial stem cell transplantation in zebrafish. PloS One 5, 1–16. doi: 10.1371/journal.pone.0012808
Oficial D. (2024). https://www.dof.gob.mx/nota_detalle_popup.php?codigo=56091941/39. Available online at: https://www.dof.gob.mx/nota_detalle_popup.php?codigo=5609194 (accessed May 20, 2024).
Okutsu T., Shikina S., Kanno M., Takeuchi Y., Yoshizaki1 G. (2007). Production of trout offspring from triploid salmon parents. Science 317, 50. doi: 10.1126/science.1145626
Pšenička M., Saito T., Linhartová Z., Gazo I. (2015). Isolation and transplantation of sturgeon early-stage germ cells. Theriogenology 83, 1085–1092. doi: 10.1016/j.theriogenology.2014.12.010
Pšenička M., Saito T., Rodina M., Dzyuba B. (2016). Cryopreservation of early stage Siberian sturgeon Acipenser baerii germ cells, comparison of whole tissue and dissociated cells. Cryobiology 72, 119–122. doi: 10.1016/j.cryobiol.2016.02.005
Ricci J. M. B., Martinez E. R. M., Butzge A. J., Doretto L. B., Oliveira M. A., Bombardelli R. A., et al. (2018). Characterization of vasa homolog in a neotropical catfish, Jundiá (Rhamdia quelen): Molecular cloning and expression analysis during embryonic and larval development. Gene 654, 116–126. doi: 10.1016/j.gene.2018.02.029
Rivers N., Daly J., Jones R., Currie P. D., Temple-Smith P. (2022). Cryopreservation and flow cytometric analysis of ovarian tissue in murray river rainbowfish, melanotaenia fluviatilis. Animals 12, 1–13. doi: 10.3390/ani12060794
Rivers N., Daly J., Jones R., Temple-Smith P. (2020). Cryopreservation of testicular tissue from Murray River Rainbowfish, Melanotaenia fluviatilis. Sci. Rep. 10, 1–9. doi: 10.1038/s41598-020-76378-7
Romney A. L. T., Myers D. M., Martin F. R., Scanlan T. N., Meyers S. A. (2023). Germ cell recovery, cryopreservation and transplantation in the California white sturgeon, Acipenser transmontanus. Sci. Rep. 13, 1–12. doi: 10.1038/s41598-023-44079-6
Rosa I. F., Martinez E. R. M., Digmayer M., Doretto L. B., Nóbrega R. H. (2023). Successful Cryopreservation of Spermatogonia Stem Cells of Neotropical Catfish (Rhamdia quelen) and Enriched Germ Cell Transplantation into Common Carp (Cyprinus carpio) Testes. Fishes 8, 1–13. doi: 10.3390/fishes8100478
Rosero J., Monzani P. S., Pessoa G. P., Coelho G. C. Z., Carvalho G. B., López L. S., et al. (2023). Traceability of primordial germ cells in three neotropical fish species aiming genetic conservation actions. Fish Physiol. Biochem. doi: 10.1007/s10695-023-01279-1
Rosero J., Pessoa G. P., Carvalho G. B., López L. S., dos Santos S. C. A., Bressan F. F., et al. (2024). Primordial germ cells of Astyanax altiparanae, isolated and recovered intact after vitrification: A preliminary study for potential cryopreservation of Neotropical fish germplasm. Cryobiology 116, 104929. doi: 10.1016/j.cryobiol.2024.104929
SADER. (2022). Expectativas agroalimentarias. Available online at: https://www.gob.mx/cms/uploads/attachment/file/723488/Expectativas_Agroalimentarias_2022.pdf (accessed 26 May, 2024).
Šćekić I., Marinović Z., Lujić J., Müller T., Kitanović N., Urbányi B., et al. (2020). A novel strategy for conservation of European eel (Anguilla Anguilla) genetic resources: Cryopreservation of ovarian stem cells. Cryobiology. doi: 10.1016/j.cryobiol.2020.03.009
Seki S., Kusano K., Lee S., Iwasaki Y., Yagisawa M., Ishida M., et al. (2017). Production of the medaka derived from vitrified whole testes by germ cell transplantation. Sci. Rep. 7, 1–11. doi: 10.1038/srep43185
Shikina S., Nagasawa K., Hayashi M., Furuya M., Iwasaki Y., Yoshizaki G. (2013). Short-term in vitro culturing improves transplantability of type A spermatogonia in rainbow trout (Oncorhynchus mykiss). Mol. Reprod. Dev. 80, 763–773. doi: 10.1002/mrd.22208
Smith-Vaniz W., Williams I. (2015). “Seriola lalandi,” in The IUCN Red List of Threatened Species 2015, Vol. 8235. 10. e.T195097A43155921
Wu X. L., Tang Z. K., Li W., Chu Z. J., Hong X. Y., Zhu X. P., et al. (2018). Identifying the germ cells during embryogenesis and gametogenesis by germ-line gene vasa in an anadromous fish, American shad Alosa sapidissima. J. Fish Biol. 92, 1422–1434. doi: 10.1111/jfb.13595
Yasui G. S., Fujimoto T., Arai K. (2010). Restoration of the loach, Misgurnus anguillicaudatus, from cryopreserved diploid sperm and induced androgenesis. Aquaculture 308, S140–S144. doi: 10.1016/j.aquaculture.2010.05.041
Yasui G. S., Fujimoto T., Sakao S., Yamaha E., Arai K. (2011). Production of loach (Misgurnus anguillicaudatus) germ-line chimera using transplantation of primordial germ cells isolated from cryopreserved blastomeres. J. Anim. Sci. 89, 2380–2388. doi: 10.2527/jas.2010-3633
Ye H., Zhou C., Yue H., Wu M., Ruan R., Du H., et al. (2021). Cryopreservation of germline stem cells in American paddlefish (Polyodon spathula). Anim. Reprod. Sci. 224. doi: 10.1016/j.anireprosci.2020.106667
Yoshikawa H., Ino Y., Shigenaga K., Katayama T., Kuroyanagi M., Yoshiura Y. (2018). Production of tiger puffer Takifugu rubripes from cryopreserved testicular germ cells using surrogate broodstock technology. Aquaculture 493, 302–313. doi: 10.1016/j.aquaculture.2018.05.016
Yoshizaki G., Fujinuma K., Iwasaki Y., Okutsu T., Shikina S., Yazawa R., et al. (2011). Spermatogonial transplantation in fish: A novel method for the preservation of genetic resources. Comp. Biochem. Physiol. - Part D: Genomics Proteomics 6, 55–61. doi: 10.1016/j.cbd.2010.05.003
Yoshizaki G., Lee S. (2018). Production of live fish derived from frozen germ cells via germ cell transplantation. Stem Cell Res. 29, 103–110. doi: 10.1016/j.scr.2018.03.015
Zhou L., Wang X., Du S., Xu S., Wang Y., Song Z., et al. (2023). Successful transplantation of cryopreserved spermatogonia in Sebastes schlegelii: A simple and suitable alternative approach for conservation of viviparous fish. Water Biol. Secur. 2. doi: 10.1016/j.watbs.2023.100142
Keywords: conservation, DDX4, endangered species, gene bank, reproduction
Citation: Mendoza-González LD, Suárez-López L and Paniagua-Chávez CG (2024) Cryopreservation of germ cells as a conservation strategy for two valuable species in Mexico: Totoaba macdonaldi and Seriola lalandi. Front. Mar. Sci. 11:1454409. doi: 10.3389/fmars.2024.1454409
Received: 25 June 2024; Accepted: 04 September 2024;
Published: 04 October 2024.
Edited by:
Yusuf Bozkurt, Iskenderun Technical University, TürkiyeReviewed by:
Tan Suet May Amelia, Chang Gung University, TaiwanShaharior Hossen, Chonnam National University, Republic of Korea
Copyright © 2024 Mendoza-González, Suárez-López and Paniagua-Chávez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carmen G. Paniagua-Chávez, Y3BhbmlhZ3VAY2ljZXNlLm14; Lucia Suárez-López, bHN1YXJlekBjaWNlc2UubXg=
†These authors have contributed equally to this work