 Filippo Drigo
Filippo Drigo Chiara Stefanelli
Chiara Stefanelli Isabella Moro*
Isabella Moro* Cristiano De Pittà
Cristiano De Pittà- Department of Biology, University of Padua, Padua, Italy
The Venice Lagoon is the largest transitional environment within the Mediterranean Sea (almost 550 km2). Being a transitional environment, it contains many different ecological niches, hence the biodiversity in this environment is very high. However, at the same time, this lagoon system is under constant pressure from human activities, and it has been greatly modified by humans during the centuries. Examples are the construction of the city of Venice, the digging of the canals for commercial ships, and the modifications of the three inlets to the lagoon. All these activities altered the circulation inside the lagoon, modifying the equilibrium between river and sea inputs, also influencing the sediment intake, fundamental for saltmarshes stability. During the centuries, many authors monitored the photosynthetic taxa (seaweeds, halophytes, and seagrasses) inhabiting the Venice Lagoon. In this study, we reviewed works from the nineteenth century to the present time to obtain a complete and updated view of the photosynthetic species that inhabit this environment, observing a decrease in the presence of charophytes and heterokontophytes and an increase in rhodophytes in the analyzed period. Moreover, we also considered the ecological valence of the species reported in this environment through two different ecological indexes (Macroalgae Quality Index – MaQI, Ecological Evaluation Index – EEI), observing a decrease in the relative contribution of the species with high ecological valence along the centuries. We also reviewed the presence of threatened and non-indigenous species, observing some differences in the considered works. Finally, we also considered the molecular resources available on online databases, finding only half macroalgal species with a reference sequence, compared to almost 90% for higher plants.
1 Introduction
1.1 The Venice Lagoon: historical background and anthropogenic impact
The Venice Lagoon, situated in the north of the Adriatic Sea (Figure 1), represents the largest transitional environment within the Mediterranean Sea, covering an area of almost 550 km2 (55000 ha) (Sfriso et al., 2009; Madricardo et al., 2019). Characterized by salt marshes and muddy areas in the upper intertidal zone, it undergoes regular tidal flooding. This microtidal environment experiences a mean tidal range of 1 m and is categorized as a restricted lagoon, where tidal movements and wind primarily drive its circulation (Fogarin et al., 2019).
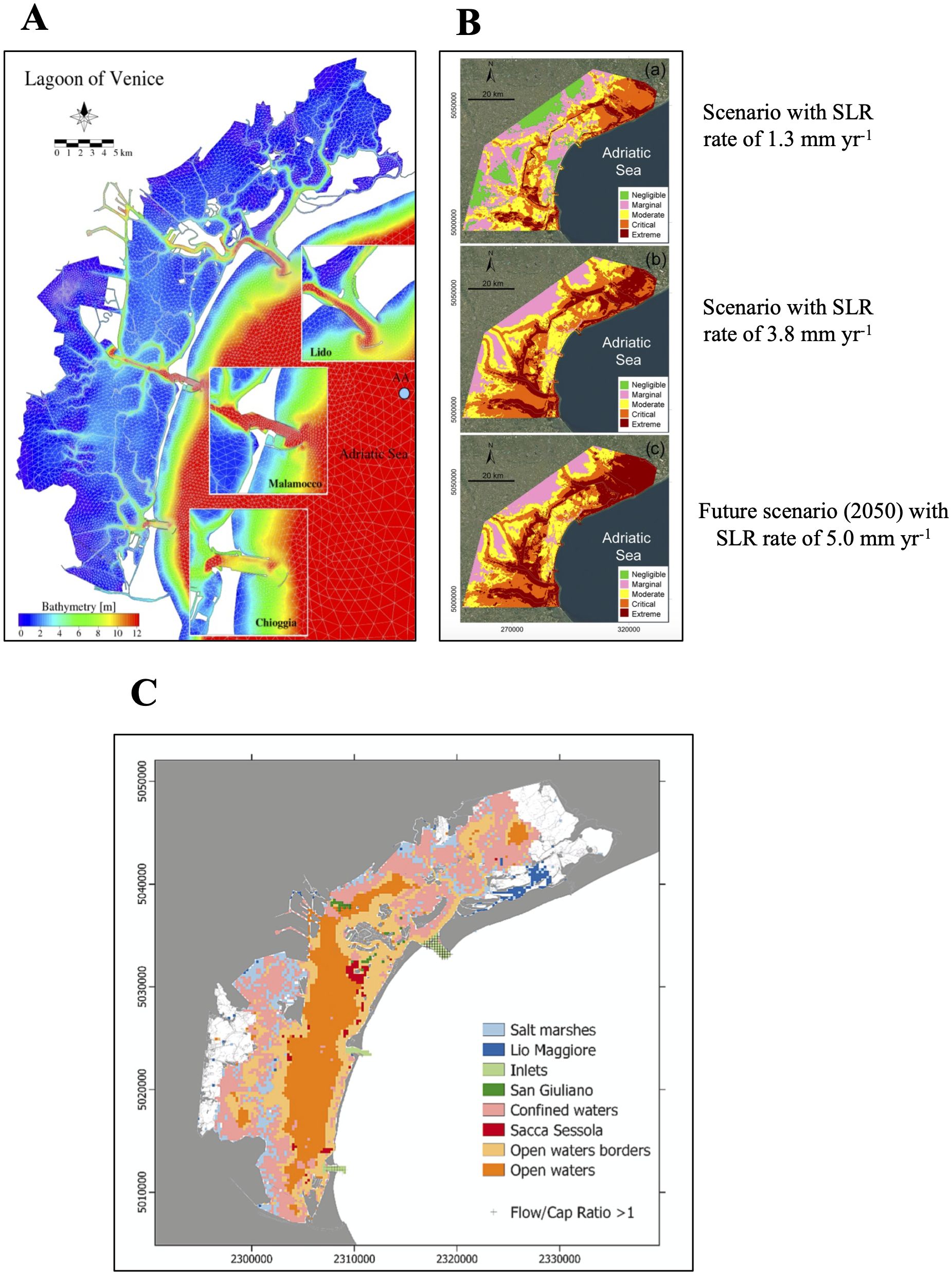
Figure 1. Description of the Venice Lagoon. (A) Representation of the bathymetric profile of the Venice Lagoon (Reproduced from Ferrarin et al., 2013, licensed under CC BY 3.0). (B) Simulation of the vulnerability of various regions in the Venice Lagoon to Relative Sea Level Rise (RSLR) under different scenarios (Reproduced from Tosi et al., 2020, licensed under CC BY 4.0). (C) Spatial distribution of various habitat types in the Venice Lagoon. Details regarding the definition of these zones can be found in the source article (Reproduced from Rova et al., 2023, licensed under CC BY-NC-ND 4.0).
Its origin dates to 12’000 years ago deriving from the combined effect of marine currents on river born sediments and sea-level lowering during the Würm glaciation (Pavoni et al., 1992). At the end of the glacial period, during the Flandrian transgression, sea level rise flooded the Upper Adriatic Würmian paleoplain shaping the coast to approximately the present condition (Brambati et al., 2003). Similar to most lagoons, the Venice Lagoon is a complex and dynamic system, greatly influenced by the Adriatic Sea, river inputs, and by the atmospheric compartment (winds and air temperature) due to its shallow depth (Ravera, 2000). This instability can lead the lagoon system to either evolve towards a marine-bay system or towards a dry-land system (Ravera, 2000). Moreover, this instability shortens the life span of lagoons, considering a geological time scale (Ravera, 2000). On the other hand, the establishment of the Serenissima Republic of Venice slowed down the natural succession by continuously preserving its integrity (Pavoni et al., 1992; Ravera, 2000). Before the nineteenth century, human activities primarily focused on mitigating silting from tributaries. Left unchecked, this silting process could gradually transform the lagoon into dry land. Additionally, efforts were made to safeguard the lagoon from erosion by the sea, which could otherwise lead to the conversion of the area into a marine-bay system (Ravera, 2000). These actions were implemented to ensure the maintenance of a navigable waterway for commercial purposes and to safeguard Venice from the erosive forces of the sea and potential threats from enemies (Ravera, 2000). Hence, in response to the escalating solid intake from tributaries caused by erosion resulting from agricultural and husbandry activities, the Republic took measures between the fifteenth and seventeenth centuries to redirect the courses of the main tributaries (e.g., Bacchiglione, Brenta, Piave, Sile) directly to the sea (Pavoni et al., 1992; Ravera, 2000; Solidoro et al., 2010; Madricardo et al., 2019). Currently, only small rivers discharge waters directly in the lagoon (Day et al., 2019).
These actions led to a decrease in nutrients of lagoon waters (mainly phosphorous), and a progressive increase in salinity. Consequently, there was a flattening of the lagoon bottom and an increase in mean depth, as natural subsidence was no longer offset (Ravera, 2000). Figure 1A displays a recent bathymetry map of the Venice Lagoon.
Conversely, various measures have been implemented since the thirteenth century to prevent sandbar erosion. It is only in the 1740 that murazzi (walls of Istrian rocks cemented together to form a barrier against the violence of the sea) were built at Malamocco and Pellestrina (Ravera, 2000; Solidoro et al., 2010; Madricardo et al., 2019).
It was during the nineteenth century that human intervention became even more pronounced. Specifically, the construction of outside dikes at the three inlets (Malamocco between 1838-1853, Lido between 1881-1905 and Chioggia in 1934) (Figure 1) had a profound impact on water circulation patterns (Ravera, 2000). Notably, the velocity of tidal currents at the Malamocco inlet increased, resulting in a deepening of the inlet, which is now the deepest point of the North-Adriatic Sea (-50 m) (Sfriso et al., 2009). Furthermore, the construction of dikes outside the inlets diminished the influx of marine sediments into the lagoon (Carniello et al., 2009; Day et al., 2019), leading to a net sediment export from the Venice Lagoon (Sarretta et al., 2010).
Additionally, the excavation of the Vittorio Emanuele III Canal (1920-1930) and the digging of Canale dei Petroli (1962-1969) amplified the influence of tidal currents, leading to an increase in deepening and land loss (with 50% reduction in salt marshes surface area) (Madricardo et al., 2019) and further transformation of the lagoon in a water-bay system (Fogarin et al., 2019). These activities, combined with land reclamation for agriculture purposes and construction of industrial areas, contributed to a reduction of nearly 7’280 hectares of the lagoon area between 1924 and 1960 (Ravera, 2000). Additionally, from the 1900 to 2019, the coverage of saltmarshes decreased from 12’000 to 3’500 hectares respectively, due to the same human activities (Day et al., 2019).
Anthropogenic activities have also impacted the subsidence rate of the lagoon. With reduced sediment input from diverted rivers, the balance between natural subsidence and the natural increase in the lagoon bottom has been disrupted (Ravera, 2000; Day et al., 2019). Furthermore, industrial activities involving groundwater and natural gas extraction in the Venice Lagoon accelerated subsidence in this area from 1930 to 1970 until management measures were implemented (Ravera, 2000; Solidoro et al., 2010; Madricardo et al., 2019). This further deteriorated the integrity of the Venice Lagoon ecosystem, particularly affecting the integrity and persistence of saltmarshes.
As a result, the reduction of sediment input from rivers (caused by dikes preventing riverine sediments intake), and the increased outflow of sediments from the inlets (due to increased impact of tidal currents), reduced the availability of sediments for saltmarsh accretion, shifting the balance towards their erosion (Carniello et al., 2009; Day et al., 2019). Moreover, the presence of waves induced by boat traffic, poses a continuous threat to the integrity of salt marshes (Madricardo et al., 2019).
Since the 1980s, some actions have been made to preserve the natural environment of the Venice Lagoon. The governance initiated a restoration project using sediments dredged from lagoon channels, resulting in the restoration of 1’300 hectares of salt marshes by 2019 (Day et al., 2019).
On the contrary, the presence of the MOSE (MOdulo Sperimentiale Elettromeccanico - Electromechanical experimental module) infrastructure may undermine the restoration efforts. The presence of this infrastructure has increased the dominance of the ebb in lagoon circulation, favoring the phase of the tidal cycle where water levels are decreasing (Ferrarin et al., 2015). Consequently, there is a prevalence of currents flowing seaward. Moreover, the closure of the MOSE during high tides prevents the inflow of elevated water levels which would otherwise flood the marshes and facilitate high rates of mineral sediment deposition, especially during the Sirocco wind (Day et al., 2019).
Additionally, increasing intensity of weather phenomena (attributed to the global climate change) and Sea Level Rise (SLR) are two other factors to consider when assessing the survival of saltmarshes in the coming years. The decline in sediments input into the lagoon for natural saltmarsh accretion, combined with a projected SLR ranging from 0.28-0.55 meters by 2100 (relative to mean sea levels from 1995-2014, Calvin et al., 2023), also diminishes the likelihood of these habitats surviving in the future due to submersion (Day et al., 2019). Figure 1B illustrates a model of the lagoon ecosystem’s vulnerability under three different rates of sea level rise (SLR).
1.2 Ecosystem services and importance of photosynthetic taxa
Coastal lagoons are important ecosystems for the different habitats they host (Figure 1C), serving as a link between the marine with freshwater and land ecosystems. Categorized as Transitional Water Systems (TWS), these environments are rich in biodiversity, further enhancing the ecosystem services that a lagoon can provide. Researchers are still debating on a common accepted definition of ecosystem services. Nonetheless, they can be defined as “the functions and products of ecosystem that benefit humans, or yield welfare to society” (Newton et al., 2018). These services can be categorized into provisioning, cultural, supporting and regulating services. The latter two categories encompass carbon sequestration, climate regulation, nursery, nutrient cycling, and more, which are essential for the health and functioning of the ecosystem, not solely for human benefit.
In the Venice Lagoon we can find three kinds of macroscopic photosynthetic taxa: seaweeds, seagrasses, and halophytic vegetation (which mainly colonize saltmarshes). As primary producers, they sustain the whole trophic web of the Venice Lagoon. These organisms serve as habitats and foraging areas for numerous faunal species, thereby supporting substantial populations of both commercial and recreational fish species (Macreadie et al., 2017). Moreover, higher plants, like seagrasses and halophytes, with their root system, compact and stabilize the soil (Macreadie et al., 2017; Sfriso et al., 2017), also providing oxygen and nutrients to the environment (Macreadie et al., 2017). This aspect is particularly significant in the Venice Lagoon, characterized by the prevalence of medium-fine sediments (Taramelli et al., 2021) and high anthropogenic impact, which, as previously mentioned, significantly enhance sediment resuspension and erosion of saltmarshes. These natural structures are characteristic of the Venice Lagoon and contribute to the area’s heterogeneity.
Increased sediment resuspension also increases the turbidity of the water. This can impact the survival of the phytobenthos, particularly affecting sensitive species, such as seagrasses, consequently further degrading the integrity of the benthic compartments (Sfriso et al., 2017). Moreover, seagrass loss may result in a decline in nursery habitats crucial for many species, including commercially important fish species, potentially leading to significant economic impacts on fisheries operating in these areas (Solidoro et al., 2010).
1.3 Biological invasion
Transitional water ecosystems are also threated by invasive species, posing a significant risk to their delicate balance. These environments boast unique gradients of environmental conditions, finely tuned to suit the needs of indigenous species. However, this specificity renders them vulnerable to sudden and strong changes. In the case of the Venice Lagoon, human activity has substantially altered the environmental dynamics of this ecosystem, posing a serious threat to the native flora and fauna. As indigenous species weaken in the face of these changes, the Venice Lagoon becomes increasingly vulnerable to biological invasions by non-native species (Cuthbert et al., 2021). These invaders, often tolerant to a wide range of conditions, can take advantage from less competitive autochthonous species, exploiting vacant ecological niches with less competition (Marchini et al., 2015).
The Mediterranean Sea stands as a prominent global hotspot for biological invasion (Katsanevakis et al., 2014; Petrocelli et al., 2019). Within this context, the Venice Lagoon emerges as a significant contributor, harboring at least 34.6% of the total non-indigenous species (NIS) found in the Mediterranean basin (Manghisi et al., 2011). This designation makes the Venice Lagoon a key Mediterranean focal point for the introduction of alien species (Sfriso, 2020), with documented occurrences including 33 seaweed species (Sfriso et al., 2023) and 2 halophytic species (Sporobolus x townsendii (H.Groves & J.Groves) P.M.Peterson & Saarela and Sporobolus anglicus (C.E.Hubb.) P.M.Peterson & Saarela) (Wong et al., 2018).
The main vectors of introduction of alien species in the Venice Lagoon are aquaculture (for example, imported shellfish contamination by allochthonous macrophytes) and commercial activities (hull fouling and ballast waters) (Manghisi et al., 2011; Katsanevakis et al., 2014; Petrocelli et al., 2019; Sfriso et al., 2023).
Non-indigenous species (NIS) pose a dual threat: they can lead to the extinction of native, vulnerable species, while also disrupting entire ecosystems by altering biogeochemical cycles and trophic chains, with consequential impacts on human well-being (Katsanevakis et al., 2014; Gallardo et al., 2016; Cuthbert et al., 2021; Sfriso et al., 2023). However, it is noteworthy that Sfriso et al. (2023) have identified some positive aspects of invasive seaweed species. For instance, Gracilariopsis vermiculophylla (Ohmi) Papenfuss 1967, considered a pioneer species, thrives in turbid environments with barren bottoms, effectively inhibiting the growth of Ulva species and thereby reducing the risk of hypoxic crises (Sfriso, 2020; Sfriso et al., 2023). Nevertheless, assessing the impact of NIS is a highly intricate task that cannot be reduced to a singular dimension (Flood et al., 2020). The consequences of NIS presence extend beyond direct ecosystem effects to include indirect impacts, such as trophic cascade (Gallardo et al., 2016), resulting in profound alterations to ecosystems functions (Flood et al., 2020).
2 Photosynthetic taxa in the Venice Lagoon
2.1 Examined literature and temporal trends
In order to obtain a comprehensive view of the photosynthetic taxa in the Venice Lagoon, we examined the literature reported in Table 1, dating back to the early nineteenth century.

Table 1. List of the analyzed literature.
The oldest records analyzed were available only in the libraries of the University of Padua, and accessing every old record was not always easy. Additionally, analyzing these catalogues proved challenging, as many entries lacked clear geographical indications and were therefore not included in the final list. Lastly, we faced difficulties during the reclassification of old species lists. Most entries have been updated according to contemporary databases (Guiry and Guiry, 2024; POWO, 2024). However, a minority could not be found in those databases and were excluded from the list presented in this work.
The most recent comprehensive catalogues of seaweeds and seagrasses species in the Venice Lagoon were compiled by Sfriso and Curiel (2007) and Sfriso et al. (2009), covering data from the 1983 up to the early 21st century. Conversely, for halophytes, the most recent comprehensive catalogue available is the work of Pignatti (1966). Subsequent studies have been more focused on specific areas and species. Additionally, the literature concerning halophytes in the Venice Lagoon is comparatively scarce compared to macroalgae; we identified only 7 works that reported halophytes species out of a total of 86 works analyzed.
In total, we identified 902 species, belonging to the Charophyta, Chlorophyta, Heterokontophyta, Rhodophyta and Tracheophyta phyla. Among these, 460 species were documented in the 1800s, 589 in the 1900s and 375 species in the first two decades of the twenty-first century. The complete list of the species is reported in the Supplementary Table S1 of Supplementary Material, where the nomenclature has been meticulously revised and updated in accordance with Guiry and Guiry (2024) for seaweeds and POWO (2024) for higher plants, including seagrasses.
In Figure 2A, a heatmap-tree illustrates the distribution of families among higher plants and macroalgae documented over centuries. The data reveal that red algae (Rhodophyta) dominate with 394 species, followed by green algae (Chlorophyta) with 224 species and Heterokontophyta with 167 species. Tracheophyta (higher plants) are represented by 100 species, while Charophyta are the least represented with only 17 species.
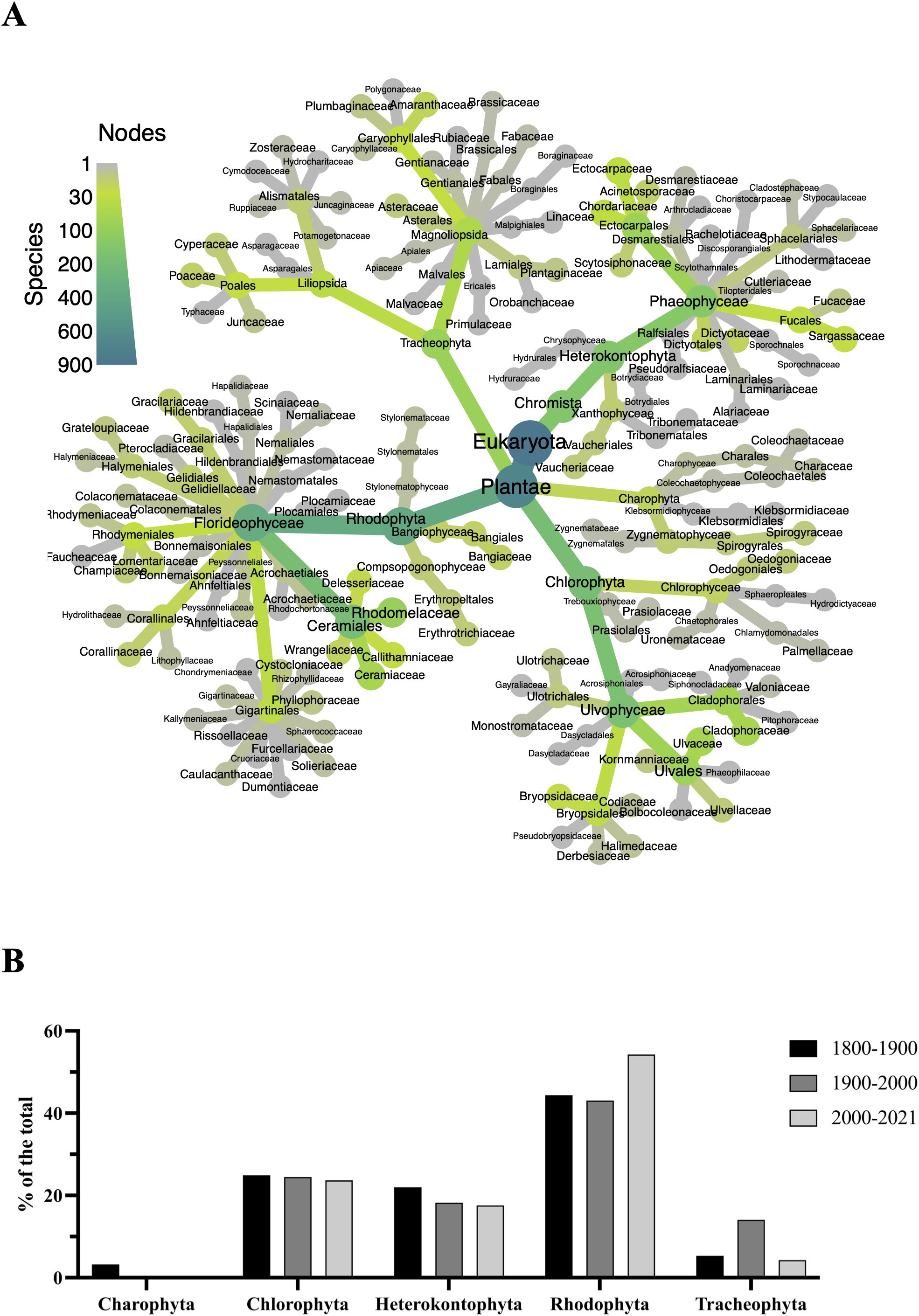
Figure 2. Description of photosynthetic taxa in the Venice Lagoon. (A) Heatmap-tree created using the R package metacoder (Foster et al., 2017) with all the species ever documented in the Venice Lagoon. The size and the color of the lines and nodes reflect the number of species reported for each taxon. (B) The temporal dynamics of photosynthetic taxa in the Venice Lagoon. The x-axis delineates the major taxonomic divisions of species found in these environments, while the y-axis represents the percentage of different taxa relative to the total across different time intervals (457 in the 1800-1900 series, 586 in the 1900-2000 series, and 375 in the 2000-2023 series).
Moreover, certain families exhibit higher biodiversity within their respective orders (e.g., the Cladophoraceae in Cladophorales and Sargassaceae in Fucales), indicating variability in species distribution among families. Despite the diversity of families, some are represented by a limited number of species, such as Charophyta overall and Malvaceae within the order Malvales for the class Magnoliposida.
In addition, considering the variation over time of the four phyla in the Venice Lagoon (Figure 2B), it is possible to notice that Charophyta and Heterokontophyta declined from the 1800s until the first two decades of this century, whereas Rhodophyta increased during the same period (in percentage terms). Supplementary Table S1 provides additional details on the temporal distribution of individual species.
2.2 Ecological status
The ecological status of an environment is not strictly correlated to the number of species, since a low number of species is not always an index of poor environmental quality, especially in oligo-mesohaline or chocked basins (Sfriso et al., 2017). The ecological value of species holds greater importance in assessing ecological status (Sfriso et al., 2017). Consequently, we focused on the ecological significance associated with various species and genera using two distinct indices: the Macroalgae Quality Index (MaQI) (Sfriso et al., 2014) and the Ecological Evaluation Index (EEI) (Orfanidis et al., 2001; Orfanidis et al., 2003; Orfanidis et al., 2011; Bermejo et al., 2012; Neto et al., 2012).
The MaQI categorizes species into three groups: (1)opportunistic species with a score of 0; (2) indifferent species with a score of 1; (3) sensitive species with a score of 2 (Sfriso et al., 2014). Meanwhile, the Ecological Evaluation Index distinguishes between two categories: (1) slow growing, late successional or perennial taxa classified under Ecological State Group I (ESG I); (2) fast-growing, opportunistic, or annual taxa classified under Ecological State Group II (ESG II) (Orfanidis et al., 2011; Neto et al., 2012).
Figures 3A, B depicts the correlation results between the ecological score and temporal changes. A noticeable trend is the decline in the proportion of species classified as sensitive (Score 2) and slow growing, late successional species (ESG I) from the 19th century through the first two decades of this century.
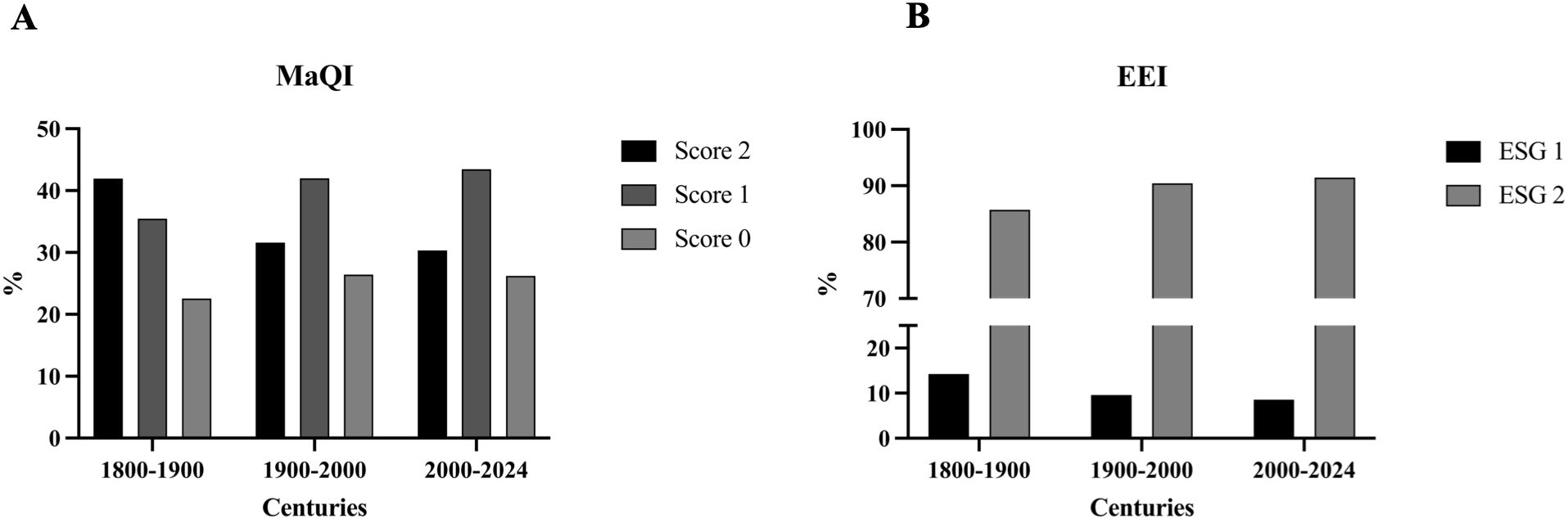
Figure 3. Ecological value of the species across different time series. (A) Temporal variation of species categorized into three ecological categories of the MaQI (Macrophyte Quality Index), showed as percentage (%). (B) Temporal variation of species categorized into two ecological categories of the EEI (Ecological Evaluation Index), represented as percentage (%).
In addition, focusing solely on the Rhodophyta (red algae) and Phaeophyceae (brown algae), Figure 2B clearly shows a decline in Phaeophyceae compared to Rhodophyta. Between 1800 and 1900, the Rhodophyta/Phaeophyta ratio was 2.19; between 1900 and 2000, it rose to 2.48; and in the first two decades of the 21st century, it increased further to 3.34. Since brown algae are often indicators of higher water quality, a consistent decline in this group compared to red algae may serve as a proxy for declining ecological conditions (Marzocchi et al., 2001).”
2.3 Threatened and non-indigenous species
As previously mentioned, the Venice Lagoon is a hotspot of biological invasion, underscoring the need for a comprehensive catalogue of both threatened and non-indigenous species for a correct management of this complex environment.
Although the IUCN – Red List stands as a global reference for threatened species, it predominantly features terrestrial organisms, with limited occurrence of macroalgae and halophytes, leaving many categorized as non-evaluated (NE) (IUCN, 2024). Considering species documented in the Venice Lagoon, only some halophytes and all seagrasses are included in the list, with many designated as Low-Concern (LC) (Table 2).
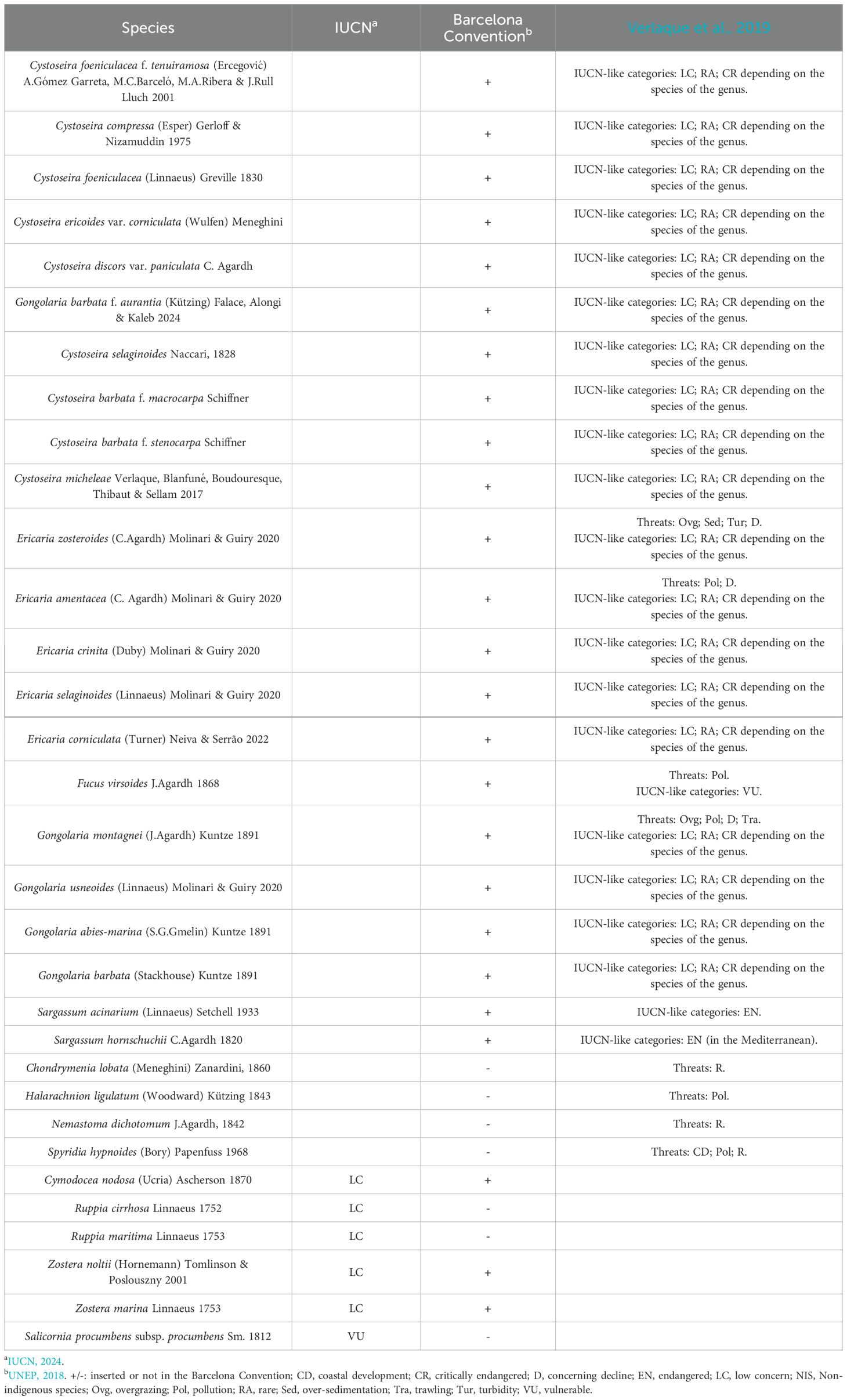
Table 2. List of the species included in the IUCN - Red List, the Barcelona Convention, and the threats and IUCN-like categories reported by Verlaque et al. (2019) for each species.
For the Mediterranean Sea, the SPA/BD Protocol Annex II (UNEP, 2018) provides a listing of threatened and endangered species, extensively reviewed by Verlaque et al. (2019). These authors assigned IUCN-like categories to species listed in the Barcelona Convention (as reported in Table 2) and raised queries regarding the inclusion of some species, such as Caulerpa ollivieri Dostàl which pose threats to indigenous species like Posidonia oceanica (L.) Delile.
Nevertheless, the SPA/BD Protocol – Annex II and the work of Verlaque et al. (2019) encompass only a limited number of macroalgal species, leaving many species found in the Venice Lagoon without a conservation status classification (Table 2). Concerning non-indigenous species (NIS) in the Venice Lagoon, recent studies by Sfriso et al. (2020; 2023) have documented 33 NIS macroalgal species. For halophytes, reports are limited, with observations primarily focused on Sporobolus anglicus and Sporobolus x townsendii, even though the latter’s presence remains uncertain (Scarton et al., 2003; Cazzin et al., 2009; Wong et al., 2018).
Additional research and databases report NIS macroalgal species at various geographic levels, including European (EASIN Catalogue; Roy et al., 2020), Mediterranean (Galil et al., 2018; Zenetos et al., 2017; Zenetos and Galanidi, 2020), and Italian areas (Pederson et al., 2017). Table 3 shows the list of NIS species reported by the different authors and databases. Notably, discrepancies exist, while 26 species are identified as NIS at the Mediterranean or European level, they are not documented in the Venice Lagoon. Zenetos et al. (2017) advise removing three species from the non-indigenous species lists: Fucus spiralis (known only from original records), Antithamnionella elegans and A. spirographidis (now considered as cryptogenic). Among the other 23 species, three (Codium vermilara, Ulva lacinulata and Cutleria multifida) were reported in the Venice Lagoon only in the 19th century, while Hypnea spinella was reported only in the 20th century.
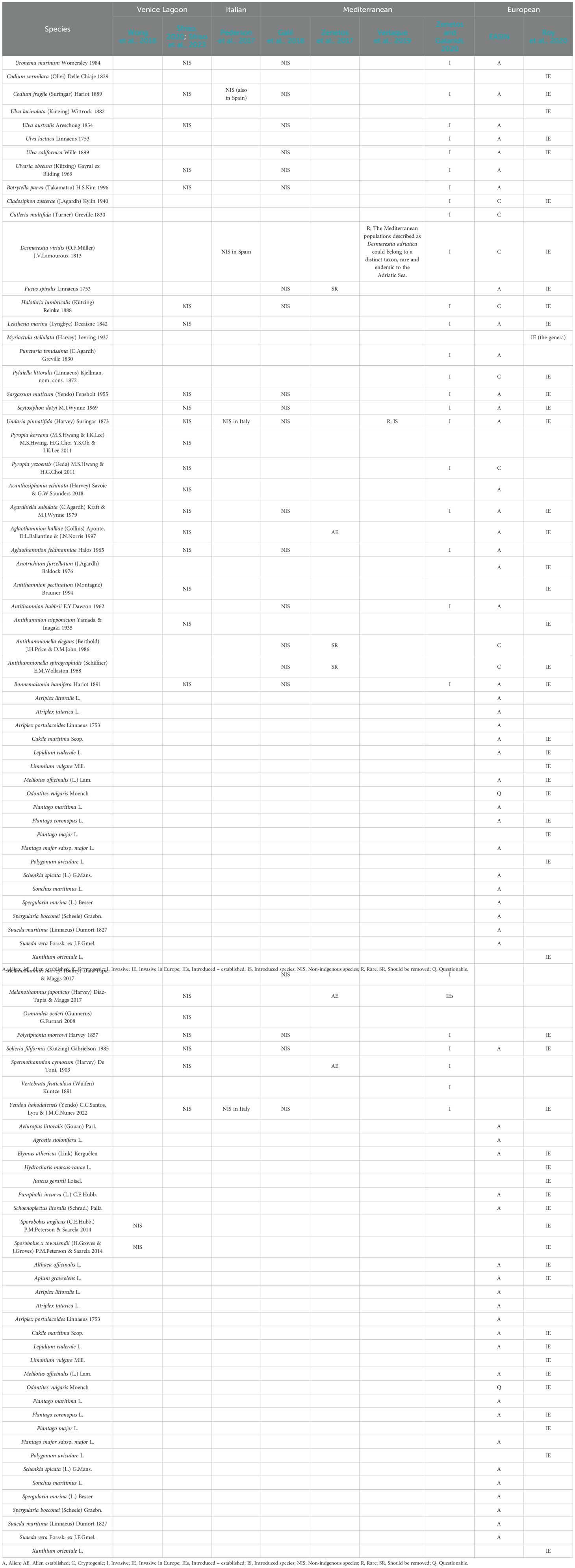
Table 3. List of non-indigenous species reported in various articles and databases: Venice Lagoon level (Wong et al., 2018; Sfriso, 2020; Sfriso et al., 2023), Italian level (Pederson et al., 2017), Mediterranean level (Galil et al., 2018; Zenetos et al., 2017; Verlaque et al., 2019; Zenetos and Galanidi, 2020), European level (EASIN; Roy et al., 2020).
The status of all other species should be monitored to ascertain their presence and impact in this environment.
Considering higher plants, Table 3 presents species listed as non-indigenous (Roy et al., 2020; EASIN Catalogue), totaling 30 species, although disparities exist in the two catalogues (e.g., Odontites vulgaris is considered invasive by Roy et al. (2020), but questionable according to EASIN Catalogue). Moreover, two of these species are listed in the IUCN – Red List and categorized as of Low concern (LC) (Apium graveolense and Spergularia marina) (IUCN, 2024).
Six species are included in the IUCN – Red List, with 5 categorized as LC (Cymodocea nodosa, Ruppia cirrhosa, Ruppia maritima, Zostera noltii and Zostera marina, all seagrasses reported in the Venice Lagoon over the centuries), and one listed as Vulnerable (VU) (Salicornia procumbens subsp. procumbens, ex. Salicornia veneta Pignatti & Lausi) (IUCN, 2024).
Cymodocea nodosa, Zostera noltii and Zostera marina are also included in the Barcelona Convention (UNEP, 2018).
Lastly, some species are reported as cryptogenic in the EASIN Catalogue (Ectocarpus siliculosus, Ectocarpus siliculosus var. hiemalis, Pyropia leucosticta, Sphaerococcus coronopifolius) and one is labelled as questionable (Limonium narbonense). Further investigations are necessary to correctly define their status.
2.4 Modelling
The detection of species in a specific study area over time can inform models of species distribution, often referred to as “biodiversity models,” which predict patterns of biodiversity (Pollock et al., 2020). Among these, Spatial Distribution Models (SDMs) are particularly noteworthy (Feeley and Silman, 2011). SDMs integrate species occurrence data with environmental factors to estimate the likelihood of species presence in unsurveyed locations or times (Milanesi et al., 2020). However, SDMs typically assume a static environment, which fails to capture the dynamic nature of real ecosystems (Milanesi et al., 2020). Species distribution evolves over both time and space, prompting recent research to focus on developing dynamic SDMs (Oliver et al., 2012; Thorson et al., 2016; Milanesi et al., 2020).
According to Milanesi et al. (2020), these models can incorporate temporal changes in species distribution and relevant ecological characteristics, leveraging time series data from various sources (some examples are reported in Milanesi et al., 2020). This results in robust models capable of predicting species occurrence across seasonal and annual variations which is crucial for conservation efforts. Additionally, SDMs can estimate two key parameters used by the IUCN for the Red List: the Extent of Occurrence (EOO) and the Area of Occupancy (AOO) (Fivaz and Gonseth, 2014). As noted in Section 2.3, there is a significant data gap for many species reported in the Venice Lagoon. Therefore, employing the species list from this study alongside the methodology proposed by Fivaz and Gonseth (2014) can assist in assessing the conservation status of the photosynthetic biodiversity in the Venice Lagoon. Moreover, if time series data on species and ecological parameters become available for other regions (e.g., coastal systems), similar models could be developed for those areas as well.
2.5 Molecular resources for species classification
Species identification may encounter several limitations: (1) phenotypic plasticity and genetic variation can cause mistakes in the identification; (2) cryptic species, characterized by similarities in size, life stage and/or morphology similar to other different species, are prevalent in various taxonomic groups (Bartolo et al., 2020); (3) morphological keys are useful during specific life stages of the species (e.g. the presence of the flower for identifying higher plant species); (4) utilizing morphological keys require a high level of expertise; however, they still pose the risk of misidentification (Kowalska et al., 2019).
The integration of molecular tools with the use of morphological keys can speed up and facilitate the identification process for photosynthetic cryptic species, as well as aiding in the control and detection of non-indigenous species (Kowalska et al., 2019). One such tool is DNA barcoding, which relies on sequencing one or a few relatively short loci standardized across many taxa, providing unequivocal identification of the targeted taxa (CBOL et al., 2009; Kowalska et al., 2019; Bartolo et al., 2020). Commonly used markers for photosynthetic taxa include rbcL and matK for plants (CBOL et al., 2009); cox-1 and rbcL for Rhodophyta and Heterokontophyta, and tufA for Chlorophyta (Saunders and McDevit, 2012).
The sequences obtained from the amplification of these specific markers are subsequently compared against public accessible databases. Therefore, if the sequence is not available, identification becomes more challenging.
On this regard, after a search of the available sequences on NCBI (last accessed on 20th February 2024) for the species recorded in the Venice Lagoon, 50.56% of the macroalgal species ever reported lack any available sequences, whereas for plants, this value stands at only 11.83%. Additionally, considering other online databases, as reported by Bartolo et al. (2020), the availability of macroalgal sequences in the Barcode of Life Data System (BOLD) for the Mediterranean Sea is limited. This underscores that without a morphological description of the organisms, identification becomes impossible for many species when only genomic data are accessible.
Nevertheless, even if molecular references are available, the species associated to that sequence can sometime be not reliable, for example due to the taxonomic impediment (lack of taxonomy experts) (Robuchon et al., 2015). Therefore, the integration of both fields (morphology and molecular) can help in the creation of a database of sequences named after a reliable morphological identification. This would be an important support in the discovery of phenotypic plasticity, in particular in seaweeds.
3 Conclusions and potential implications
Coastal systems are particularly vulnerable ecosystems, heavily impacted by high concentrations of human activities such as tourism, aquaculture, and urban development. These disturbances and the stresses from anthropogenic activities alter the ecosystem, making it more susceptible to biological invasions and sudden environmental changes.
From the 19th century to the first two decades of the 21st century, human activities have significantly altered the Venice Lagoon, exerting a profound influence on its environmental dynamics. At the same time, analysis of past record of the photosynthetic taxa reveals shifts in the contribution of certain taxa to the total photosynthetic biodiversity, for instance, an increase in the percentage of Rhodophyta and a decrease of Heterokontophyta in more recent records. Moreover, considering the ecological value of the species reported at various times since the 19th century, there has been a rise in opportunistic and short-lived species, potentially compromising the stability and health of the ecosystem. Compounded by the presence of numerous non-indigenous species, all these factors collectively represent a serious threat for the lagoon. To address these challenges, comprehensive monitoring initiatives must be implemented, particularly focusing on mapping species distribution throughout the entire lagoon. Such efforts are crucial for early detection of invasive species and declines in sensitive species, facilitating timely intervention to mitigate further ecological degradation. Moreover, comparing historical species lists with recent monitoring campaigns is crucial for understanding the changes that have occurred over the centuries (Willis et al., 2007). These findings can guide the prioritization of monitoring efforts by highlighting areas of past distribution that warrant closer examination. Additionally, about potential restoration activities, this work can serve as a baseline for planning efforts by focusing on areas that retain characteristics more similar to the past (Yoshioka et al., 2014). It also provides a reference for the historical ‘biological status’ of the lagoon system under study, which can aid in planning more effective restoration initiatives (Willis et al., 2007). In conclusion, historical reviews of species are very important tools for addressing the challenges posed by anthropogenic impacts and global climate change, particularly in lagoon ecosystems that are highly sensitive to disturbances. Understanding how these systems have changed over the centuries, both in terms of species composition and significant events, can inform the development of biodiversity models that forecast future scenarios. By integrating these historical data with the ecological characteristics of the environment being studied, researchers can enhance the accuracy of their predictions (Milanesi et al., 2020).
Author contributions
FD: Conceptualization, Data curation, Investigation, Writing – original draft, Methodology. CS: Writing – review & editing, Investigation. IM: Conceptualization, Data curation, Funding acquisition, Project administration, Supervision, Validation, Writing – review & editing. CD: Conceptualization, Data curation, Funding acquisition, Project administration, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was funded under the National Recovery and Resilience Plan (NRRP), Mission 4 Component 2 Investment 1.4 - Call for tender No. 3138 of 16 December 2021, rectified by Decree n.3175 of 18 December 2021 of Italian Ministry of University and Research funded by the European Union – NextGenerationEU. Project code CN_00000033, Concession Decree No. 1034 of 17 June 2022 adopted by the Italian Ministry of University and Research, CUP C93C22002810006, Project title “National Biodiversity Future Center - NBFC” to CD and IM. It was also funded by Interreg Italy-Croatia Programme 2021-2027 “BlueDiversity-ITHR0200404” to CD. Open Access funding provided by Università degli Studi di Padova | University of Padua, Open Science Committee.
Acknowledgments
We are grateful to the libraries of the University of Padua for the support in the retrieval of the historical books and Federico Merlin for the analysis of part of the historical records.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1433680/full#supplementary-material
References
Agardh C. A. (1824). Systema Algarum Vol. 1, I-XXXVIII (Sweden: Lundae: Literis Berlingianis), 1–312.
Agardh J. G. (1842). Algae maris Mediterranei et Adriatici, observationes in diagnosin specierum et dispositionem generum Vol. I-X (Parisiis: Apud fortin, Masson et Cie), 1–164.
Ardissone F. (1871). Rivista dei ceramii della flora italiana. Nuovo giornale botanico italiano 3, 32–50.
Ardissone F. (1883). Phycologia Mediterranea. Parte prima, Floridee Vol. I-X (Varese: Antica tipografia ferri di Maj e Malnati), 1–516.
Ardissone F. (1886). Phycologia Mediterranea. Parte seconda, Oospore-Zoospore-Schizospore Vol. 1 (Varese: Antica tipografia ferri di Maj e Malnati), 1–325.
ARPAV (2011). Laguna di Venezia – dati macrofite – anno 2011. Available online at: https://www.arpa.veneto.it/dati-ambientali/open-data/file-e-allegati/soaml/laguna-di-venezia/macrofite/actrmo0044_dati-macrofite-venezia.csv/@@download/file (Accessed October 19, 2023).
ARPAV (2014). Laguna di Venezia – dati macrofite – anno 2014. Available online at: https://www.arpa.veneto.it/dati-ambientali/open-data/file-e-allegati/soaml/laguna-di-venezia/macrofite/macrofite_venezia_2014.csv/@@download/file (Accessed October 19, 2023).
ARPAV (2018). Laguna di Venezia – dati macrofite – anno 2018. Available online at: https://www.arpa.veneto.it/dati-ambientali/open-data/file-e-allegati/soaml/laguna-di-venezia/macrofite/dati_macrofite_venezia_2018.csv/@@download/file (Accessed October 19, 2023).
ARPAV (2021). Laguna di Venezia – dati macrofite – anno 2021. Available online at: https://www.arpa.veneto.it/dati-ambientali/open-data/file-e-allegati/soaml/laguna-di-venezia/macrofite/macrofite-venezia-2021.csv/@@download/file (Accessed October 19, 2023).
Bartolo A. G., Zammit G., Peters A. F., Küpper F. C. (2020). The current state of DNA barcoding of macroalgae in the Mediterranean Sea: presently lacking but urgently required. Botanica Marina 63, 253–272. doi: 10.1515/bot-2019-0041
Béguinot A. (1907). Le attuali conoscenze sulla Flora Lagunare ed i problemi che ad essa si collegano. Venezia: Officine grafiche di C. Ferrari. 1, 1–321.
Béguinot A. (1938). La vita delle piate nella laguna e nei lidi di Venezia. Premiate officine grafiche di C. Ferrari. 1, 1–18.
Bellemo G., Curiel D., Marzocchi M., Iuri M., Pavan M., Vecchiato N., et al. (1999). Aggiornamento di nuove specie algali per la laguna di Venezia. Lavori – Soc Ven. Sc. Nat. 24, 55–66.
Bellemo G., Curiel D., Rocca B. L., Scattolin M., Marzocchi M. (2001). First report of a filamentous species of desmarestia (Desmarestiaceae, fucophyceae) in the lagoon of venice (Italy, Mediterranean sea). Botanica Marina 44, 541–545. doi: 10.1515/BOT.2001.060
Benacchio N. (1938). Osservazioni sistematiche e biologiche sulle Zosteracee dell’Alto Adriatico. Venezia: Carlo Ferrari. 1, 1–38.
Bermejo R., Vergara J. J., Hernández I. (2012). Application and reassessment of the reduced species list index for macroalgae to assess the ecological status under the Water Framework Directive in the Atlantic coast of Southern Spain. Ecol. Indic. 12, 46–57. doi: 10.1016/j.ecolind.2011.04.008
Bertoloni A. (1862b). Flora Italica Cryptogama – Algae Vol. 2 (Bologna: Gisueppe Cenerelli), 129–256.
Bertoloni A. (1867). Flora Italica Cryptogama – Algae Vol. 2 (Bologna: Gisueppe Cenerelli), 257–338.
Brambati A., Carbognin L., Quaia T., Teatini P., Tosi L. (2003). The Lagoon of Venice: geological setting, evolution and land subsidence. Episodes J. Int. Geosci. 26, 264–268. doi: 10.18814/epiiugs/2003/v26i3/020
Brunelli G., Magrini G., Miliani L., Orsi P. (1938). La Laguna di Venezia - Monografia. Colume III, Parte V, Tomo IX, Fascicolo I (Venezia: Officine Grafiche Carlo Ferrari).
Calvin K., Dasgupta D., Krinner G., Mukherji A., Thorne P. W., Trisos C., et al. (2023). “IPCC 2023: Climate Change 2023: Synthesis Report,” in Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Eds. Lee H., Romero J. (Intergovernmental Panel on Climate Change (IPCC, Geneva, Switzerland). doi: 10.59327/IPCC/AR6-9789291691647
Carniello L., Defina A., D’Alpaos L. (2009). Morphological evolution of the Venice lagoon: Evidence from the past and trend for the future. J. Geophys. Res. 114, 2008JF001157. doi: 10.1029/2008JF001157
Cazzin M., Ghirelli L., Mion D., Scarton F. (2009). Completamento della cartografia della vegetazione e degli habitat della laguna di Venezia: anni 2005-2007. Lavori Soc. Ven. Sc. Nat. 34, 81–89.
CBOL Plant Working Group1, Hollingsworth P. M., Forrest L. L., Spouge J. L., Hajibabaei M., Ratnasingham S., et al. (2009). A DNA barcode for land plants. Proc. Natl. Acad. Sci. U.S.A. 106, 12794–12797. doi: 10.1073/pnas.0905845106
Curiel D. (2008-2009). LA LAGUNA DI VENEZIA: genesi, evoluzione, naturalità e salvaguardia: Presenza e distribuzione di specie alloctone. [Course Material]. CITTÀ DI VENEZIA. Available online at: https://www.comune.venezia.it/sites/comune.venezia.it/files/immagini/Ambiente/CL_Alghe_aliene_Daniele_Curiel.pdf
Curiel D., Bellato A., Solazzi A., Marzocchi M. (1994). Radicilingua thysanorhizans (Holmes) Papenfuss (Rhodophyta, Ceramiales): nuova specie per la laguna di Venezia. Informatore botanico italiano – note scientifiche 26, 171–175.
Curiel D., Bellemo G., Checchin E., Dri C., Miotti C., Marzocchi M. (2005). Segnalazione di nuove macroalghe per la laguna di Venezia. Lavori – Soc Ven. Sc. Nat. 30, 41–44.
Curiel D., Bellemo G., Marzocchi M. (1996a). New records of marine algae in the lagoon of Venice. Giornale botanico italiano 130, 352–352. doi: 10.1080/11263509609439592
Curiel D., Bellemo G., Marzocchi M., Iuri M. (1997a). Il macrofitobenthos algale di superficie e di profondità dei moli foranei della laguna di Venezia. Biol. Mar. Medit. 4, 44–50.
Curiel D., Bellemo G., Marzocchi M., Scattolin M., Parisi G. (1998a). Distribution of introduced Japanese macroalgae Undaria pinnatifida, Sargassum muticum (Phaeophyta) and Antithamnion pectinatum (Rhodophyta) in the Lagoon of Venice. Hydrobiologia 385, 17–22. doi: 10.1023/A:1003437105147
Curiel D., Bellemo G., Rocca B. L., Scattolin M., Marzocchi M. (2002). First report of polysiphonia morrowii harvey (Ceramiales, Rhodophyta) in the Mediterranean sea. Botanica Marina 45, 66–70. doi: 10.1515/BOT.2002.008
Curiel D., Bellemo G., Scattolin M. (2003a). Ritrovamento in Laguna di Venezia di specie algali nuove per l’Adriatico e rare per il Mediterraneo. Informatore Botanico Italiano 35, 7–11.
Curiel D., Bellemo G., Scattolin M., Marzocchi M. (2001). Variazioni della flora bentonica macroalgale nei substrati duri della laguna di Venezia: confronto con rilievi del 1938. Lavori – Soc Ven. Sc. Nat. 26, 71–83.
Curiel D., Bellemo G., Scattolin M., Marzocchi M. (2006). First report of Lomentaria hakodatensis (Lomentariaceae, Rhodophyta) from the lagoon of Venice (Adriatic Sea, Mediterranean). Acta ADRIATICA 47, 65–72.
Curiel D., Bellemo G., Scattolin M., Marzochi M. (2000). Le macroalghe dei substrati duri della laguna di Venezia: analisi di un gradiente ecologico tra il mare e la terraferma. Boll. Mus. Civ. St. Nat. 51, 11–30.
Curiel D., Boscolo N., Marzocchi M. (2008). Il macrofitobenthos delle valli da pesca della laguna di Venezia. Lavori – Soc Ven. Sc. Nat. 33, 59–70.
Curiel D., Marzocchi M., Bellemo G. (1996b). First report of fertile antithamnion pectinatum (Ceramiales, rhodophyceae) in the north Adriatic sea (Lagoon of Venice, Italy). Botanica Marina 39, 19–22. doi: 10.1515/botm.1996.39.1-6.19
Curiel D., Pranovi F., Marozcchi M. (1996c). I popolamenti macrobentonici di una valle da pesca. La Valle Averto nella Laguna veneta. Ambiente Risorse Salute 43, 25–30.
Curiel D., Rismondo A., Bellemo G., Marzocchi M. (2004). Macroalgal biomass and species variations in the Lagoon of Venice (Northern Adriatic Sea, Italy): 1981-1998. Sci. Mar. 68, 57–67. doi: 10.3989/scimar.2004.68n157
Curiel D., Rismondo A., Marzocchi M., Scattolin M. (1998b). Variazioni Strutturali nella comunità fouling dei canali della laguna di Venezia (centro storico e isole). Lavori – Soc Ven. Sc. Nat. 23, 25–36.
Curiel D., Rismondo A., Marzocchi M., Solazzi A. (1995). Distribuzione di Sargassum muticum (Yendo) Fensholt (Phaeophyta) in laguna di Venezia. Acqua Aria 8, 831–834.
Curiel D., Scattolin M., Marzocchi M. (2003b). Modificazioni dei popolamenti fitobentonici di substrato duro nella laguna di Venezia in seguito all’introduzione di Undaria Pinnatifida (Harvey) Suringar. Lavori – Soc Ven. Sc. Nat. 28, 25–31.
Curiel D., Solazzi A., Marzocchi M., Scattolin M. (1997b). Il macrofitbenthos della “Palude Maggiore” (Laguna di Venezia). Lavori – Soc Ven. Sc. Nat. 22, 81–89.
Cuthbert R. N., Pattison Z., Taylor N. G., Verbrugge L., Diagne C., Ahmed D. A., et al. (2021). Global economic costs of aquatic invasive alien species. Sci. Total Environ. 775, 145238. doi: 10.1016/j.scitotenv.2021.145238
Day J. W., Ibáñez C., Pont D., Scarton F. (2019). Status and sustainability of mediterranean deltas: the case of the Ebro, Rhône, and Po deltas and Venice lagoon. Coasts Estuaries, 237–249. doi: 10.1016/B978-0-12-814003-1.00014-9
De Toni J. B. (1889). Sylloge Algarum Vol. I (Patavii: Sumptibus Auctoris Typis Seminarii), 1–1315. Chlorophyceae 1.
De Toni J. B. (1895). Sylloge Algarum Vol. III (Patavii: Sumptibus Auctoris Typis Seminarii), 1–638. Fucoideae 1.
De Toni J. B. (1897). Sylloge Algarum Vol. IV (Patavii: Sumptibus Auctoris Typis Seminarii), 1–338. Florideae 1, Sectio I.
De Toni J. B. (1900). Sylloge Algarum Vol. IV (Patavii: Sumptibus Auctoris Typis Seminarii), 387–776. Florideae 1, Sectio II.
De Toni J. B. (1903). Sylloge Algarum Vol. IV (Patavii: Sumptibus Auctoris Typis Seminarii), 775–1525. Florideae 1, Sectio III.
De Toni J. B. (1905). Sylloge Algarum Vol. IV (Patavii: Sumptibus Auctoris Typis Seminarii), 1523–1973. Florideae 1, Sectio IV.
De Toni G. B. (1923). Un’aggiunta all’algarium zanardini. Atti del reale istituto veneto di scienze lettere ed arti 82, 475–485.
De Toni J. B. (1924). Sylloge Algarum Vol. IV (Patavii: Sumptibus Auctoris Typis Seminarii), 1–767. Florideae 1, Sectio V.
De Toni G. B., Levi D. (1885). Flora algologica della Venezia. Parte prima: Le Floridee Vol. 1 (Venezie: Tipografia di G. Antonelli), 1–182.
De Toni G. B., Levi D. (1886). Flora algologica della Venezia. Parte seconda: Le Melanoficee Vol. 1 (Venezie: Tipografia di G. Antonelli), 1–109.
De Toni G. B., Levi D. (1888a). “Collezioni di storia naturale. I. Collezioni botaniche,” in L’algarium Zanardini, vol. 1. (Civico Museo e raccolta Correr, Venezia), 1–144.
De Toni G. B., Levi D. (1888b). Flora algologica della Venezia. Parte terza: Le Cloroficee Vol. 1 (Venezie: Tipografia di G. Antonelli), 1–206.
EASIN Catalogue. Available online at: https://easin.jrc.ec.europa.eu/apixg/catxg/getall/skip/0/take/15000 (Accessed February 15, 2024).
Feeley K. J., Silman M. R. (2011). Keep collecting: accurate species distribution modelling requires more collections than previously thought: Temporal autocorrelated biases necessitate more collections. Diversity Distrib. 17, 1132–1140. doi: 10.1111/j.1472-4642.2011.00813.x
Ferrarin C., Ghezzo M., Umgiesser G., Tagliapietra D., Camatti E., Zaggia L., et al. (2013). Assessing hydrological effects of human interventions on coastal systems: numerical applications to the Venice Lagoon. Hydrol. Earth Syst. Sci. 17, 1733–1748. doi: 10.5194/hess-17-1733-2013
Ferrarin C., Tomasin A., Bajo M., Petrizzo A., Umgiesser G. (2015). Tidal changes in a heavily modified coastal wetland. Continent. Shelf Res. 101, 22–33. doi: 10.1016/j.csr.2015.04.002
Fivaz F. P., Gonseth Y. (2014). Using species distribution models for IUCN Red Lists of threatened species. J. Insect Conserv. 18, 427–436. doi: 10.1007/s10841-014-9652-6
Flood P. J., Duran A., Barton M., Mercado-Molina A. E., Trexler J. C. (2020). Invasion impacts on functions and services of aquatic ecosystems. Hydrobiologia 847, 1571–1586. doi: 10.1007/s10750-020-04211-3
Fogarin S., Madricardo F., Zaggia L., Sigovini M., Montereale-Gavazzi G., Kruss A., et al. (2019). Tidal inlets in the Anthropocene: Geomorphology and benthic habitats of the Chioggia inlet, Venice Lagoon (Italy). Earth Surf Process. Landf 44, 2297–2315. doi: 10.1002/esp.4642
Foster Z., Sharpton T., Grunwald N. (2017). Metacoder: An R package for visualization and manipulation of community taxonomic diversity data. PloS Comput. Biol. 13, 1–15. doi: 10.1371/journal.pcbi.1005404
Furnari G., Cormaci M., Serio D. (1999). Catalogue of the benthic marine macroalgae of the Italian coast of the Adriatic Sea. Bocconea 12, 5–214.
Gargiulo M. G., De Masi F., Tripodi G. (2000). Problemi nomenclaturali, tassonomici e ambientali relativi alle macroalghe della laguna di Venezia. Ist. Ven. Sci. Lett. Arti, ed) La Ricerca Scientifica per Venezia Vol. 2 (Il progetto Sistema Lagunare Veneziano), 1116–1135.
Galil B. S., Marchini A., Occhipinti-Ambrogi A. (2018). East is east and West is west? Management of marine bioinvasions in the Mediterranean Sea. Estuarine Coast. Shelf Sci. 201, 7–16. doi: 10.1016/j.ecss.2015.12.021
Gallardo B., Clavero M., Sánchez M. I., Vilà M. (2016). Global ecological impacts of invasive species in aquatic ecosystems. Global Change Biol. 22, 151–163. doi: 10.1111/gcb.13004
Guiry M. D., Guiry G. M. (2024). AlgaeBase (World-wide electronic publication, University of Galway). Available at: https://www.algaebase.org.
IUCN (2024). The IUCN Red List of Threatened Species. Version 2024-1. Available online at: https://www.iucnredlist.org (Accessed March 8, 2024).
Katsanevakis S., Coll M., Piroddi C., Steenbeek J., Ben Rais Lasram F., Zenetos A., et al. (2014). Invading the Mediterranean Sea: biodiversity patterns shaped by human activities. Front. Mar. Sci. 1. doi: 10.3389/fmars.2014.00032
Kowalska Z., Pniewski F., Latała A. (2019). DNA barcoding – A new device in phycologist’s toolbox. Ecohydrol. Hydrobiol. 19, 417–427. doi: 10.1016/j.ecohyd.2019.01.002
Macreadie P. I., Jarvis J., Trevathan-Tackett S. M., Bellgrove A. (2017). “Seagrasses and Macroalgae: Importance, Vulnerability and Impacts,” in Climate Change Impacts on Fisheries and Aquaculture. Eds. Phillips B. F., Pérez-Ramírez M. (Hoboken, NJ, USA: Wiley), 729–770. doi: 10.1002/9781119154051.ch22
Madricardo F., Foglini F., Campiani E., Grande V., Catenacci E., Petrizzo A., et al. (2019). Assessing the human footprint on the sea-floor of coastal systems: the case of the Venice Lagoon, Italy. Sci. Rep. 9, 6615. doi: 10.1038/s41598-019-43027-7
Manghisi A., Minicante S. A., Bertuccio C., Morabito M., Genovese G. (2011). A cryptic alien seaweed spreading in Mediterranean coastal lagoons. Transitional Waters Bulletin 5 (1), 1–7.
Marchini A., Ferrario J., Sfriso A., Occhipinti-Ambrogi A. (2015). Current status and trends of biological invasions in the Lagoon of Venice, a hotspot of marine NIS introductions in the Mediterranean Sea. Biol. Invasions 17, 2943–2962. doi: 10.1007/s10530-015-0922-3
Marzocchi M., Bellemo G., Miotti C., Curiel D., Scattolin M. (2001). Le macroalghe dei substrati duri del canal grande (centro storico di venezia): prime considerazioni. Boll. Mus. Civ. St. Nat. 52, 25–39.
Marzocchi M., Curiel D., Scattolin M. (2003). Variazioni del fitobenthos di substrato duro della laguna di Venezia tra il mare e la terraferma. Boll. Mus. Civ. St. Nat. 54, 5–17.
Meneghini G. (1842a). Alghe Italiane e Dalamtiche. Fasciolo 1 Vol. 1 (Padova: Tipografia di Angelo Sicca), 1–80.
Meneghini G. (1842b). Alghe Italiane e Dalamtiche. Fasciolo 2 Vol. 1 (Padova: Tipografia di Angelo Sicca), 81–160.
Meneghini G. (1842c). Alghe Italiane e Dalamtiche. Fasciolo 3 Vol. 1 (Padova: Tipografia di Angelo Sicca), 161–255.
Meneghini G. (1843). Alghe Italiane e Dalamtiche. Fasciolo 4 Vol. 1 (Padova: Tipografia di Angelo Sicca), 256–352.
Meneghini G. (1846). Alghe Italiane e Dalamtiche. Fasciolo 5 Vol. 1 (Padova: Tipografia di Angelo Sicca), 353–384.
Milanesi P., Della Rocca F., Robinson R. A. (2020). Integrating dynamic environmental predictors and species occurrences: Toward true dynamic species distribution models. Ecol. Evol. 10, 1087–1092. doi: 10.1002/ece3.5938
Miotti C., Curiel D., Rismondo A., Bellemo G., Dri C., Checchin E., et al. (2005). First report of a species of Prasiola (Chlorophyta: Prasiolaceae) from the Mediterranean Sea (Lagoon of Venice). Sci. Mar. 69, 343–346.
Neto J. M., Gaspar R., Pereira L., Marques J. C. (2012). Marine Macroalgae Assessment Tool (MarMAT) for intertidal rocky shores. Quality assessment under the scope of the European Water Framework Directive. Ecol. Indic. 19, 39–47. doi: 10.1016/j.ecolind.2011.09.006
Newton A., Brito A. C., Icely J. D., Derolez V., Clara I., Angus S., et al. (2018). Assessing, quantifying and valuing the ecosystem services of coastal lagoons. J. Nat. Conserv. 44, 50–65. doi: 10.1016/j.jnc.2018.02.009
Oliver T. H., Gillings S., Girardello M., Rapacciuolo G., Brereton T. M., Siriwardena G. M., et al. (2012). Population density but not stability can be predicted from species distribution models. J. Appl. Ecol. 49, 581–590. doi: 10.1111/j.1365-2664.2012.02138.x
Orfanidis S., Panayotidis P., Stamatis N. (2001). Ecological evaluation of transitional and coastal waters: A marine benthic macrophytes-based model. Medit. Mar. Sci. 2, 45. doi: 10.12681/mms.266
Orfanidis S., Panayotidis P., Stamatis N. (2003). An insight to the ecological evaluation index (EEI). Ecol. Indic. 3, 27–33. doi: 10.1016/S1470-160X(03)00008-6
Orfanidis S., Panayotidis P., Ugland K. (2011). Ecological Evaluation Index continuous formula (EEI-c) application: a step forward for functional groups, the formula and reference condition values. Medit. Mar. Sci. 12, 199. doi: 10.12681/mms.60
Pavoni B., Marcomini A., Sfriso A., Donazzolo R., Orio A. A. (1992). “Changes in an Estuarine ecosystem. The Lagoon of Venice as a case study,” in The Science of Global Change: The Impact of Human Activities on the Environment, ACS Symposium Series. Eds. Dunnette D. A., O’Brien R. J. (American Chemical Society, Washington, DC). doi: 10.1021/bk-1992-0483
Pederson J. A., Gollasch S., Laing I., McCollin T., Miossec L., Occhipinti-Ambrogi A., et al. (2017). Status of introductions of non-indigenous marine species in the North Atlantic and adjacent waters 2003-2007. ICES Cooperative Research Report (CRR) 334, 151 pp. doi: 10.17895/ICES.PUB.1977
Petrocelli A., Antolić B., Bolognini L., Cecere E., Cvitković I., Despalatović M., et al. (2019). Port Baseline Biological Surveys and seaweed bioinvasions in port areas: What’s the matter in the Adriatic Sea? Mar. pollut. Bull. 147, 98–116. doi: 10.1016/j.marpolbul.2018.04.004
Pignatti S. (1962). Associazioni di alghe marine sulla costa veneziana (Venezia: Istituto Veneto di scienze lettere ed arti).
Pignatti S. (1966). La vegetazione alofila della laguna veneta (Venezia: Istituto Veneto di scienze, lettere ed arti).
Pollock L. J., O’Connor L. M. J., Mokany K., Rosauer D. F., Talluto L., Thuiller W. (2020). Protecting biodiversity (in all its complexity): new models and methods. Trends Ecol. Evol. 35, 1119–1128. doi: 10.1016/j.tree.2020.08.015
POWO (2024). Plants of the World Online (Facilitated by the Royal Botanic Gardens, Kew). Available at: http://www.plantsoftheworldonline.org/.
Ravera O. (2000). The Lagoon of Venice: the result of both natural factors and human influence. J. Limnol. 59, 19. doi: 10.4081/jlimnol.2000.19
Robuchon M., Valero M., Gey D., Le Gall L. (2015). How does molecular-assisted identification affect our estimation of α, β and γ biodiversity? An example from understory red seaweeds (Rhodophyta) of Laminaria kelp forests in Brittany, France. Genetica 143, 207–223. doi: 10.1007/s10709-014-9796-z
Rova S., Stocco A., Pranovi F. (2023). Sustainability threshold for multiple ecosystem services in the Venice lagoon, Italy. Ecosys. Serv. 64, 101568. doi: 10.1016/j.ecoser.2023.101568
Roy D., Alderman D., Anastasiu P., Arianoutsou M., Augustin S., Bacher S., et al. (2020). DAISIE - Inventory of alien invasive species in Europe. Version 1.7 (Research Institute for Nature and Forest (INBO) (Accessed March 8, 2024).
Ruchinger G. (1818). Flora dei lidi Veneti Vol. 1 (Venezia: Presso Gio. Giacomo Fuchus. Dalla Tip. Picotti), 1–281.
Sarretta A., Pillon S., Molinaroli E., Guerzoni S., Fontolan G. (2010). Sediment budget in the lagoon of Venice, Italy. Continent. Shelf Res. 30, 934–949. doi: 10.1016/j.csr.2009.07.002
Saunders G. W., McDevit D. C. (2012). “Methods for DNA Barcoding Photosynthetic Protists Emphasizing the Macroalgae and Diatoms,” in DNA Barcodes, Methods in Molecular Biology. Eds. Kress W. J., Erickson D. L. (Humana Press, Totowa, NJ), 207–222. doi: 10.1007/978-1-61779-591-6_10
Scarton F., Ghirelli L., Curiel D., Rismondo A. (2003). First data on Spartina× townsendii in the lagoon of Venice (Italy). In Proceedings of the sixth international conference on the Mediterranean coastal environment, MEDCOAST, Ravenna, Italy, vol. 3, pp. 787–792.
Schiffner V., Vatova A. (1937). “Le alghe della Laguna di Venezia. Estratto della monografia. La laguna di Venezia- Volume III, Parte V, Tomo IX,” in La Laguna di Venezia - Monografia. Eds. Brunelli G., Magrini G., Miliani L., Orsi P. (Officine Grafiche Carlo Ferrari, Venezia).
Sfriso A. (1987). Flora and vertical distribution of macroalgae in the lagoon of Venice: a comparison with previous studies. Giornale botanico italiano 121, 69–85. doi: 10.1080/11263508709431647
Sfriso A. (2020). Invasion of alien macroalgae in the Venice Lagoon, a pest or a resource? AI 15, 245–270. doi: 10.3391/ai.2020.15.2.03
Sfriso A., Buosi A., Facca C., Sfriso A. A. (2017). Role of environmental factors in affecting macrophyte dominance in transitional environments: The Italian Lagoons as a study case. Mar. Ecol. 38, e12414. doi: 10.1111/maec.12414
Sfriso A., Curiel D. (2007). Check-list of seaweeds recorded in the last 20 years in Venice lagoon, and a comparison with the previous records. botm 50, 22–58. doi: 10.1515/BOT.2007.004
Sfriso A., Curiel D., Rismondo A. (2009). “The Lagoon of Venice,” in Flora and Vegetation of the Italian Transitional Water Systems, 1st ed. Eds. Cecere E., Petrocelli A., Izzo G., Sfriso A. (Stampa “Multigraf”, Spinea (VE), 17–80.
Sfriso A., Facca C., Bonometto A., Boscolo R. (2014). Compliance of the macrophyte quality index (MaQI) with the WFD, (2000/60/EC) and ecological status assessment in transitional areas: The Venice lagoon as study case. Ecol. Indic. 46, 536–547. doi: 10.1016/j.ecolind.2014.07.012
Sfriso A., La Rocca B. (2005). Aggiornamento sulle macroalghe presenti lungo i litorali e sui bassifondi della laguna di Venezia. Lavori – Soc Ven. Sc. Nat. 30, 45–56.
Sfriso A., La Rocca B., Godini E. (2002). Inventario di taxa macroalgali in tre aree della laguna di Venezia a differente livello di trofia. Lavori – Soc Ven. Sc. Nat. 27, 85–99.
Sfriso A., Wolf M. A., Buosi A., Sciuto K., Sfriso A. A. (2023). Alien macroalgal rearrangement in the soft substrata of the Venice lagoon (Italy): impacts, threats, time and future trends. Sustainability 15, 8256. doi: 10.3390/su15108256
Silvestri S., Defina A., Marani M. (2005). Tidal regime, salinity and salt marsh plant zonation. Estuarine Coast. Shelf Sci. 62, 119–130. doi: 10.1016/j.ecss.2004.08.010
Solazzi A., Curiel D., Chiozzotto E. (1991-1994). Flora macroalgale di fondali mobili (paludi e velme) della laguna di Venezia. Nuova Thalassia 12, 59–68.
Solidoro C., Bandelj V., Bernardi F., Camatti E., Ciavatta S., Cossarini G., et al. (2010). “Response of the Venice Lagoon Ecosystem to Natural and Anthropogenic Pressures over the Last 50 Years,” in Coastal Lagoons, Marine Science. Eds. Kennish M., Paerl H. (Boca Raton, FL: CRC Press), 483–511. doi: 10.1201/EBK1420088304-c19
Taramelli A., Valentini E., Piedelobo L., Righini M., Cappucci S. (2021). Assessment of state transition dynamics of coastal wetlands in Northern Venice lagoon, Italy. Sustainability 13, 4102. doi: 10.3390/su13084102
Thorson J. T., Ianelli J. N., Larsen E. A., Ries L., Scheuerell M. D., Szuwalski C., et al. (2016). Joint dynamic species distribution models: a tool for community ordination and spatio-temporal monitoring. Global Ecol. Biogeogr. 25, 1144–1158. doi: 10.1111/geb.12464
Tolomio C. (1993). Prima segnalazione di Grateloupia doryphora (Mont.) Howe (Rhodophyceae) nella laguna di Venezia. Lavori – Soc Ven. Sc. Nat. 18, 215–220.
Tosi L., Da Lio C., Donnici S., Strozzi T., Teatini P. (2020). Vulnerability of Venice’s coastland to relative sea-level rise. Proc. IAHS 382, 689–695. doi: 10.5194/piahs-382-689-2020
UNEP/MAP-SPA/RAC (2018). SAP/RAC: SPA-BD Protocol - Annex II: List of endangered or threatened species.
Verlaque M., Boudouresque C.-F., Perret-Boudouresque M. (2019). Mediterranean seaweeds listed as threatened under the Barcelona Convention: A critical analysis. Sci. Rep. Port-Cros Natl. Park 33, 179–214.
Willis K. J., Araújo M. B., Bennett K. D., Figueroa-Rangel B., Froyd C. A., Myers N. (2007). How can a knowledge of the past help to conserve the future? Biodiversity conservation and the relevance of long-term ecological studies. Phil. Trans. R. Soc B 362, 175–187. doi: 10.1098/rstb.2006.1977
Wolf M. A., Buosi A., Sfriso A. (2020). First record of Acanthosiphonia eChinata (Rhodomelaceae, Rhodophyta) in the Mediterranean Sea, molecular and morphological characterization. Botanica Marina 63, 241–245. doi: 10.1515/bot-2019-0072
Wong J. X. W., Costantini F., Merloni N., Savelli L., Geelen D., Airoldi L. (2018). The widespread and overlooked replacement of Spartina maritima by non-indigenous S. anglica and S. townsendii in north-western Adriatic saltmarshes. Biol. Invasions 20, 1687–1702. doi: 10.1007/s10530-017-1654-3
Yoshioka A., Akasaka M., Kadoya T. (2014). Spatial prioritization for biodiversity restoration: A simple framework referencing past species distributions. Restor. Ecol. 22, 185–195. doi: 10.1111/rec.12075
Zanardini G. (1841). Synopsis algarum in mari Adriatico hucusque collectarum, cui accedunt monographia siphonearum nec non generales de algarum vita et structura disquisitiones cum tabulis auctoris manu ad vivum depictis (Turin: Tome) Vol. 4, 1–153.
Zanardini G. (1843). Saggio di classificazione naturale delle Ficee del Dottore Giovanni Zanardini. Aggiunti nuovi studi sopra l’androsace degli antichi con tavola miniata ed enumerazione di tutte le specie scoperte e raccolte dall’autore in Dalmazia Vol. 1 (Venezie: Dallo stabilimento tipografico enciclopedico di Girolamo Tasso), 1–64.Available online at: https://www.algaebase.org/search/bibliography/detail/?biblio_id=13496&token=AC100CF31ae1e209B3MGQ14673D6.
Zanardini G. (1847). “Notizie intorno alle cellulari marine delle lagune e de litorali di Venezia (1),” in Atti del Reale Istituto Veneto di Scienze, Lettere ed Arti, Vol. 6, 185–262.
Zanardini G. (1860). Iconographia phycologica adriatica ossia scelta di Ficee nuove o più rare del mare adriatico figurate, descritte ed illustrate da G. Zanardini Vol. I-VIII (Venezia), 1–175. nel priv. stabil. di G. Antonelli.
Zanardini G. (1863). Scelta di Ficee nuove o più rare del mare Adriatico. Decade quarta(Venezia: Memorie del Reale Istituto Veneto di Scienze, Lettere ed Arti) Vol. 11, 269–306.
Zanardini G. (1865). Iconographia phycologica adriatica ossia scelta di Ficee nuove o più rare del mare adriatico figurate, descritte ed illustrate da G. Zanardini. edit. 2 Vol. I-VIII (Venezia), 1–168. nel priv. stabil. di G. Antonelli.
Zanardini G. (1871). Scelta di Ficee nuove o più rare del mare Adriatico Vol. 15 (Venezia: Memorie del Reale Istituto Veneto di Scienze, Lettere ed Arti), 427–460.
Zanardini G. (1874). Iconographia Phycologica Adriatica ossia scelta di Ficee nuove o più rare del Mare Adriatico figurate, descritte ed illustrate da G. Zanardini Vol. 3 (Venezia: Nello Stabil. Tip. G. Antonelli), 1–132.
Zanardini G. (1876). Scelta di Ficee nuove o più rare dei mari Mediterraneo ed Adriatico Vol. 19 (Venezia: Memorie del Reale Istituto Veneto di Scienze, Lettere ed Arti Col), 509–544.
Zenetos A., Çinar M. E., Crocetta F., Golani D., Rosso A., Servello G., et al. (2017). Uncertainties and validation of alien species catalogues: The Mediterranean as an example. Estuarine Coast. Shelf Sci. 191, 171–187. doi: 10.1016/j.ecss.2017.03.031
Keywords: photosynthetic taxa, Venice Lagoon, nineteenth century, biodiversity, ecological status, threatened and non-indigenous species
Citation: Drigo F, Stefanelli C, Moro I and De Pittà C (2024) Evaluation of photosynthetic Taxa in the Venice Lagoon from the nineteenth century to present day. Front. Mar. Sci. 11:1433680. doi: 10.3389/fmars.2024.1433680
Received: 16 May 2024; Accepted: 12 November 2024;
Published: 11 December 2024.
Edited by:
Emilio Fernández, University of Vigo, SpainReviewed by:
Mirta Teichberg, Marine Biological Laboratory (MBL), United StatesAbdul-Salam Fayiz Juhmani, Ca’ Foscari University of Venice, Italy
Copyright © 2024 Drigo, Stefanelli, Moro and De Pittà. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Isabella Moro, aXNhYmVsbGEubW9yb0B1bmlwZC5pdA==; Cristiano De Pittà, Y3Jpc3RpYW5vLmRlcGl0dGFAdW5pcGQuaXQ=