
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Mar. Sci., 19 July 2024
Sec. Marine Megafauna
Volume 11 - 2024 | https://doi.org/10.3389/fmars.2024.1427334
 Ana M. Correia1,2*
Ana M. Correia1,2* Marieta Mihova1,2
Marieta Mihova1,2 Ágatha Gil1,2
Ágatha Gil1,2 Cláudia Oliveira-Rodrigues1,2
Cláudia Oliveira-Rodrigues1,2 Luís Afonso1
Luís Afonso1 Raul Valente1,2
Raul Valente1,2 Massimiliano Rosso1,3
Massimiliano Rosso1,3 Isabel Sousa-Pinto1,2
Isabel Sousa-Pinto1,2 Graham J. Pierce4,5
Graham J. Pierce4,5The north-western coast of Africa is one of the most productive marine systems in the world, where a high diversity of cetacean species has been recorded. However, management of human activities that impact cetacean populations in the area is inadequate, in part due to poor knowledge on cetacean occurrence and the lack of systematic surveys. We reviewed existing knowledge on cetacean distribution between Morocco and Liberia and analyzed the spatial and temporal distribution of records, as well as species occurrence, further discussing the reported distribution ranges. The systematic literature review included 39 sources reporting on 36 species in the area, with a total of 2241 sightings and 2178 non-sighting records. Delphinus delphis (common dolphin) and Tursiops truncatus (bottlenose dolphin) are the two species with the highest number of sources reporting on them. This review reveals a spatially and temporally heterogeneous coverage of the area in the literature and published data on cetacean occurrence. In summary, although most of the compiled sources are recently published (2011-2020), the majority of records are from prior to 2000. In several range states, especially south of Senegal, little information is available. Therefore, the available information is unlikely to represent the current status of cetacean communities in the region. It is essential to fill knowledge gaps by increasing the collection of cetacean distribution data in the area, in order to support effective conservation measures and ensure sustainable utilization of the living marine resources in this rapidly developing region.
The Canary Current Large Marine Ecosystem off north-west Africa (NWA) is one of the most productive in the world (Caldeira et al., 2002; UNEP-CMS, 2008; Mason, 2009; UNEP-CMS, 2012; Sala et al., 2013; Satia, 2016), with a strong upwelling system supported by the Canary Current System and extremely dynamic oceanographic processes. In this region, the Islamic Republic of Mauritania’s Banc d’Arguin National Park, established between Mauritania and Guinea, is the largest Marine Protected Area in western Africa. It has some of the richest fish resources in the world (FAO, 2013), sustaining high marine biodiversity, including several species of cetaceans (Subregional Fisheries Commission, 2003; IUCN World Heritage Outlook, 2020). Some cetaceans pass through the region during their yearly migrations while others may remain throughout the year (Jefferson et al., 1997; Robineau and Vely, 1998; Van Waerebeek et al., 2000; Perrin and Van Waerebeek, 2012; Weir and Pierce, 2013; Baines and Reichelt, 2014; Djiba et al., 2015; Masski and de Stéphanis, 2015; Russell et al., 2018; Correia et al., 2019). Therefore, the marine management of the region is fundamental for the conservation of cetacean populations in the NWA (UNEP-CMS, 2012).
The Memorandum of Understanding (MoU) Concerning the Conservation of the West African Manatee and Small Cetaceans of Western Africa and Macaronesia was signed in 2008, under the Convention on Migratory Species of Wild Animals (CMS) of the United Nations Environmental Programme (CMS, www.cms.int/aquatic-mammals/en). This was the first regional effort to address the conservation of cetaceans in these waters. However, to avoid overlapping with the competences of the International Whaling Commission, large cetaceans were not included in the MoU (Caddell, 2009). Moreover, the Memorandum is not legally binding and it is still not signed by all west African range states (i.e. nations). The MoU highlights the fact that “knowledge of the biology, ecology, migration, population abundance, and the conservation status of manatees and small cetaceans is insufficient”, and that there is urgency for sustainable management of “oil and gas exploration, extraction and other mining activities, fishing and tourism” (CMS, www.cms.int/aquatic-mammals/en). The CMS listed numerous threats to small cetaceans on the African coast due to human activities, including those related to fisheries: bycatch, direct take, trophic interaction with fisheries, and ship strikes (UNEP, 2008).
The fishing sector represents an important economic activity along the west African coast, and there has been increasing overexploitation of fishing resources by the intensive activity of both legal and illegal industrial fishing fleets (FAO, 2013; Belhabib et al., 2017; Englander and Costello, 2023). Both inside and outside the Banc d’Arguin National Park, the overexploitation of marine resources, including illegal and/or unsustainable fishing (large scale, commercial, and artisanal), is still a major pressure impacting biodiversity in NWA (UNEP-CMS, 2008; Trouillet et al., 2011; UNEP-CMS, 2012; Weir and Pierce, 2013; Diedhiou and Yang, 2018; IUCN World Heritage Outlook, 2020).
Among the known impacts (e.g., prey depletion, habitat degradation), fishing activity in the area is linked to the decline of marine megafauna of conservation concern due to bycatch. Since there is a high amount of fishing effort in the NWA by European fleets, to support the high domestic demand for fish resources (Nagel and Gray, 2012; Tickler et al., 2018), Europe also has a major responsibility in terms of reducing the regional bycatch of cetaceans.
With the aim of reducing the overexploitation of resources, partnerships between the European Union (EU) and west African states have been established, specifically the Sustainable Fisheries Partnerships Agreements (SFPAs). SFPAs are agreements with partner third countries “for the purpose of obtaining access to their waters and resources in order to sustainably exploit a share of surplus in exchange for financial compensation from the EU, which includes sectoral support” (European Commission, 2023). However, overall, it seems these agreements have so far failed to accomplish their goal of achieving sustainable use of fishery resources through shared decision-making between EU and African countries (Nagel and Gray, 2012; Corten, 2014; IUCN World Heritage Outlook, 2020). In 2023, the European Commission identified several areas of improvement, which would be necessary for the SFPAs. The report emphasizes the need to strengthen the “scientific inputs to the design of ecosystem measures”, justifying that the lack of scientific information is preventing the implementation of measures to reduce bycatch and fisheries interaction with protected fauna (European Commission, 2023).
Weir and Pierce (2013) reviewed the impact of human activities on cetaceans in the eastern tropical Atlantic but highlighted that it is challenging to estimate the magnitude of the impacts due to a lack of data from regular monitoring programs. Baseline knowledge on cetacean distribution in the NWA is lacking and existing records come from occasional surveys, which often covered only small parts of the area or were dedicated to certain species (e.g., Expósito and Qninba, 2010; Masski and de Stéphanis, 2015; Weir and Collins, 2015), and/or derive from “grey literature” (e.g., Bowman Bishaw Gorham, 2003). To date, the most complete reviews compiling cetacean occurrence for the NWA are Jefferson et al. (1997), which is based on what are now historical records, and Perrin and Van Waerebeek (2012), which does not include quantitative data.
For reliable assessments of the impacts of anthropogenic threats, and to promote effective marine management, it is crucial to create and maintain an updated cetacean species inventory in the area through non-lethal research (UNEP-CMS, 2008, 2012; Weir and Pierce, 2013). To contribute to this objective, we assessed published cetacean records throughout the NWA, from Morocco to Liberia, to create a comprehensive and quantitative inventory of cetacean species and to identify data gaps.
We collated published documents on cetacean occurrence in the NWA, from Morocco to Liberia [using the boundaries for the NWA as defined in Weir et al. (2014)]. The review followed the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA, McIvor et al., 2022). First, we retrieved the sources by searching the Web of Science platform for documents, published up to 2020, using the words: “cetacean(s)/marine mammal(s)/dolphin(s)/whale(s)” plus “Africa/Morocco/Western Sahara/Mauritania/Senegal/The Gambia/Guinea Bissau/Guinea/Sierra Leone/Liberia”. The titles and abstracts of the documents were then screened, removing any that were clearly unrelated to the topic (e.g., not on cetaceans or not providing information for the study area). Additional documents were removed when reviewing the complete documents for data collation, following the same criteria. The search yielded many sources in which the records reported derived from previous publications. Often, these previous publications were not retrieved by the search and, despite further efforts to trace them, many could not be located (mainly due to being historical/grey/regional documents not available online). In all cases, we cited the original reporting sources in the compiled dataset.
The process for data collation is represented schematically in Appendix A and the resulting full dataset is presented in Appendix B. In summary, the set of selected documents resulting from the PRISMA process was rigorously revised to compile cetacean occurrence records. Occurrence records were categorized into sightings and non-sightings. Sightings corresponded to records obtained at-sea from visual monitoring, from which group size could often be assessed (each record is a single sighting, regardless of the number of individuals). Non-sighting records corresponded to other sources of data such as strandings, bycatch, direct takes, cetacean collections, and acoustic data, from which quantitative information (i.e. the number of individuals) could sometimes be extracted. The specification of non-sighting typology was not provided in the dataset as, in many cases, the origin of a record was ambiguous, or it fitted under more than one category.
We collated both qualitative and quantitative information. The latter included number of sightings and/or non-sightings, group size (for sightings only), location of the occurrence record (range state and region), date (or time interval), and source. For qualitative data (i.e., documentation of the occurrence of a species in the area, without further associated quantitative information), we retrieved the species, range states of occurrence, and sources. For cases with unconfirmed species identification, and/or uncertain state of occurrence (i.e., the location was not fully specified, but the description is sufficient to place it within the study area), we compiled records under “unconf. ID” and/or “unconf. state of occurrence”, respectively. When the documents contained information on a larger region, we attempted to gather only the records within NWA (i.e., as defined between Morocco and Liberia). If this was not possible (i.e., the geographic location of the records was poorly specified, or unspecified), and for records likely located in the study area but without absolute certainty, these were classified with “unconf. location”. In the case of occurrences reported for the Canary Islands, as presented in Djiba et al. (2015) and Camphuysen (2000), the range state was considered as “Morocco and/or Western Sahara” (under the classification of “unconf. state of occurrence”), because the surveys were conducted between the west coast of Africa and the Canary Islands, crossing both EEZs (Spanish and African) and, therefore, it was reasonable to assume that sightings were within (or at the limit of) the study area.
All data were cross-checked between sources and all efforts were made to avoid repeated records. Nevertheless, when insufficient data were provided (date and region), although this rarely happened, cross-checking may have failed to detect duplicated records. When the same record was repeated in more than one source, all the sources documenting the record were noted in the dataset.
Two of the authors (Ana M. Correia and Ágatha Gil) oversaw the initial compilation process and a third author (Marieta Mihova) verified the resulting dataset.
First, for each source, we compiled information on the type of data presented, study area, period covered, number of species, and number of records documented (sightings and/or non-sightings). For the number of records, confirmed and unconfirmed identifications from sighting and non-sighting records were included (i.e., records with “unconf. ID” were included), as long as the locations were unquestionably inside the study area (i.e., excluding “unconf. location”). For the number of species, we considered only identifications confirmed to the species level (i.e., excluding “unconf. ID” and identifications made to a higher taxonomic level). For repeated records (present in multiple sources), the numbers of sightings and non-sightings were usually taken from the source with the earliest year of publication. An exception was made for Sousa teuszii (Atlantic humpback dolphin), for which a very complete and recent review was conducted by Weir and Collins (2015), with new data included. In this case, we used Weir and Collins (2015) as our first source (if they cited the other sources), because it was impossible to track all these records back to their original sources (we retrieved as many records as possible, avoiding duplications). Publication dates ranged from 1988 to 2020, while occurrence records were documented from 1882 to 2019. Thus, for temporal analysis of the dataset, three time-periods were defined: before 2000, 2000-2010, and 2011-2020. Sources were then summarized by the three time-periods, after being classified according to the type of publication (reviews, inventories/reports/surveys, and case reports), the area covered (e.g. the whole area, part of the area, or a single range state), and the number of species documented (single or multiple).
Data were then organized by species (excluding “unconf. ID” and “unconf. location”), recording the number of occurrence records (sightings and non-sightings), group size (in the case of sightings), and the sources (all sources reporting on a species were identified, even in cases of repeated records). The number of records by species was also summarized for each of the three time-periods, based on the date of the record (not the publication date). Records for which the information was insufficient to assign it to a specific time-period were excluded for this temporal analysis and were classified as “unspec. time period” in the dataset.
To assess literature coverage by time-period, for each range state and for the whole study area (records with “unconf. location” excluded), we calculated the percentage of publications, species, and sighting and non-sighting records in relation to the respective total. Although the date of the record was used for species, sightings, and non-sightings, the publication date was used to derive the number publications by time-period. To assess species coverage, only confirmed identifications to the species level were considered. Finally, distribution range was assessed for each species, excluding records with “unconf. ID”, identity confirmed only to a higher taxonomic level, “unconf. location”, and/or “unconf. state of occurrence”.
A detailed analysis of the literature coverage is presented in Appendix C and Figure 1. Of the 39 sources compiled, 10 are reviews, of which 4 document the occurrence of a single species and 6 document the occurrence of multiple species. In two of the reviews, Jefferson et al. (1997) and Perrin and Van Waerebeek (2012), the period covered by the occurrence records is not specified, and the latter presents qualitative data only (Appendix C.1). A majority (6) of the 10 reviews were published between 2011 and 2020, although they cover only records up to 2000 (Appendix C.2).
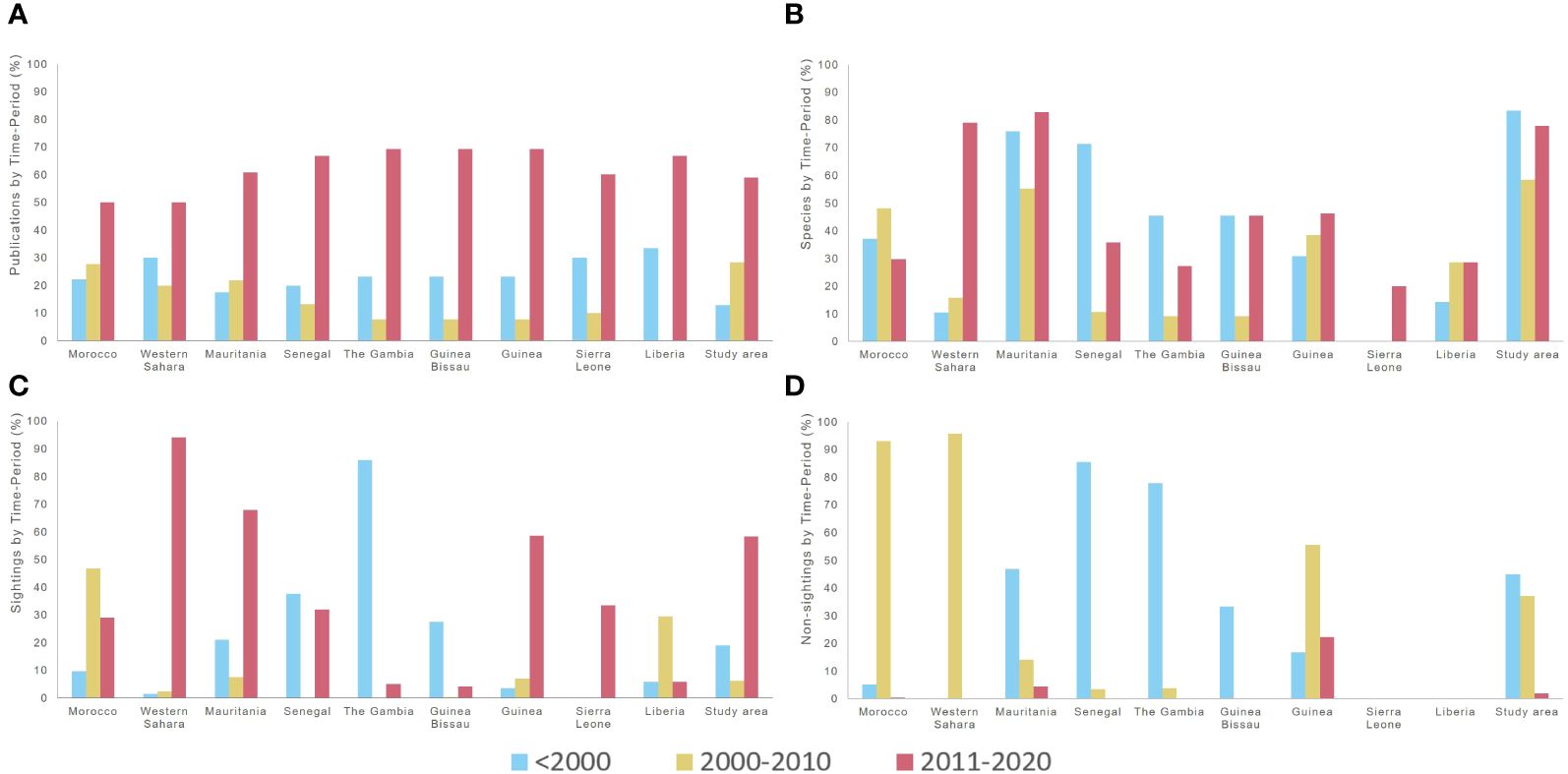
Figure 1 Literature coverage for each range state, and overall, by time-period, regarding the percentage of publications (A), species (B), sightings (C), and non-sightings (D). The year of publication and the year of records were considered to determine the time-period. Percentages were calculated for each time-period (records with non-specified time-period excluded), in relation to the total data gathered (including records with non-specified time-period). The percentages of species, sightings, and non-sightings, reported per time-period, do not sum up to 100%, since several species are repeatedly reported across time-periods, and records with non-specified time-periods were accounted for only in the denominator (i.e., total data gathered). For percentages by range state, only records with confirmed locations (at least to state level) were included; while for the study area, all records were considered (if definitely within the study area).
There are 24 inventories, reports, or based on survey data. Of these, 11 were published between 2001 and 2010, and the other 13 between 2011 and 2020; and 19 sources include data on multiple species. Finally, there are five case reports (i.e., reporting on a single species in a single range state), four of which were published between 2011 and 2020 and one before 2000 (Appendix C.2). Out of the 39 sources, 23 sources were published during 2011-2020 (Appendix C.2). Most publications are from this time-period, overall and within each range state (Figure 1A).
In total, 36 species are reported for the study area, and we retrieved quantitative data for 34 of them, summing 2241 sightings and 2178 non-sightings (Table 1). Robineau and Vely (1998) report the highest number of species, with 29 species recorded, including quantitative data (mostly from beach census data, i.e., strandings) for 21 of them. Nevertheless, this source refers only to the state of Mauritania. Correia et al. (2019) report the greatest number of sighting records (456 observations of 20 species), albeit only for a sub-region of the study area (Morocco to Senegal). Regarding non-sightings, Van Waerebeek et al. (2000), in a report for UNEP, presents the highest number, with 550 records from Senegal to Guinea-Bissau (Appendix C.1).
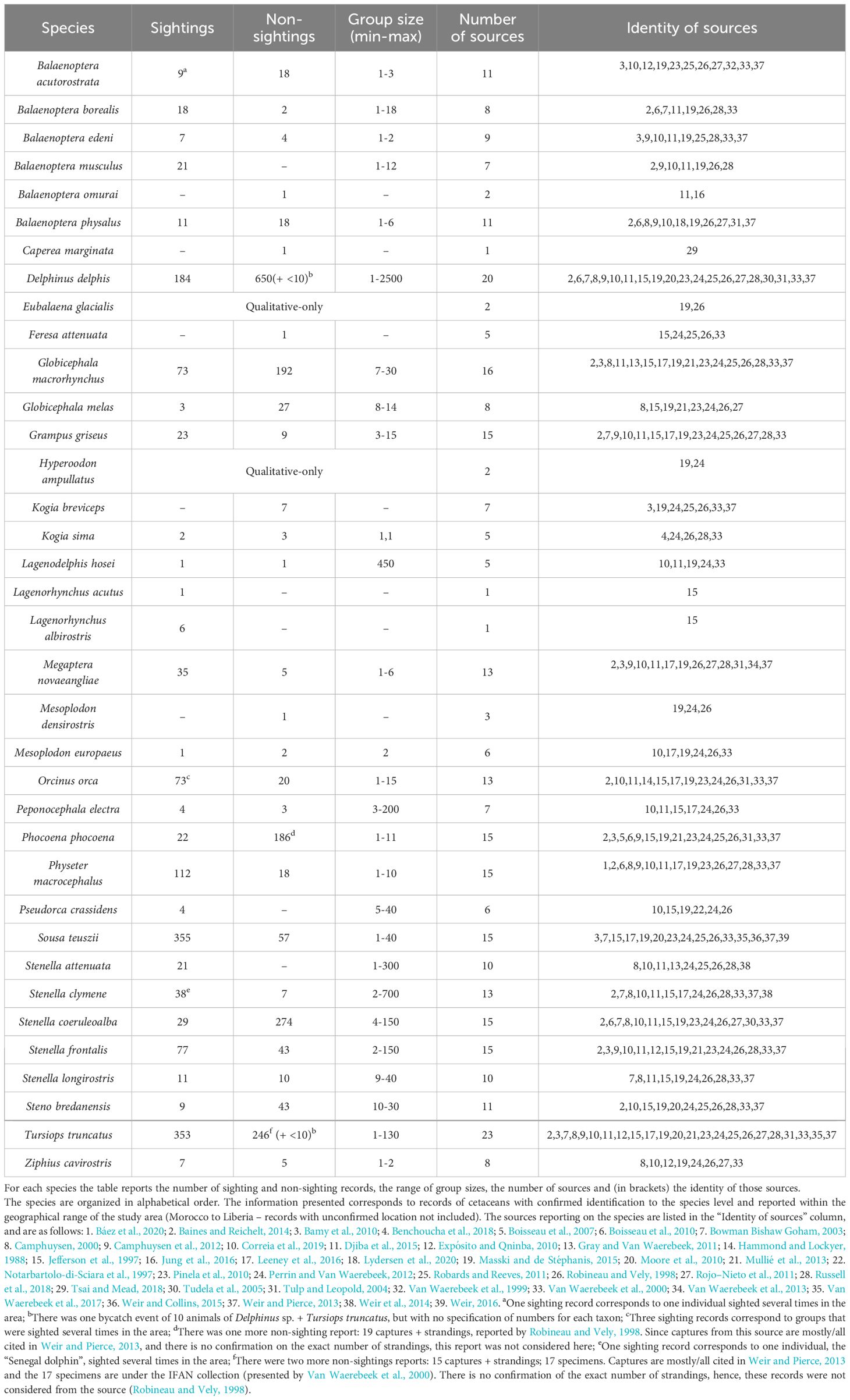
Table 1 Summary of the number of cetacean records, with confirmed identification to species level, found in the literature for north-western Africa, from Morocco to Liberia.
Only five sources — all reviews — provide data for the entire NWA (Morocco to Liberia), three of them published before 2000 (two of these on a single species) and two published between 2011 and 2020 (one for a single species). Among the sources covering the entire study area, Jefferson et al. (1997) reported the highest number of species (18 species, of which there are quantitative data on only 5), Weir and Collins (2015) reported the highest number of records (321 sightings), although targeting only S. teuszii, and only Jefferson et al. (1997) compiled non-sightings from multiple species, although reporting only 9 of such records (Appendix C.1). In the entire study area, the time-period with the highest percentages of the total number of species and the total number of non-sightings is the period before 2000 (Figures 1B, D, respectively), while the time-period with the highest percentage of sightings is 2011-2020 (Figure 1C).
Among 24 sources with data from a single state, 8 reported data from Morocco and 9 reported data from Mauritania (Appendix C.1). For both Senegal and The Gambia, the time-period with the highest percentage of species reported was prior to 2000. The same percentage of reported species was observed in Guinea Bissau for the time-periods <2000 and 2011-2020. In Western Sahara, the highest percentage of species, by far, was reported in 2011-2020; and in Sierra Leone, time-period assignment was only possible for a single species, which was reported in 2011-2020. Morocco is the only state for which the largest percentage of species was reported in the decade 2000 to 2010. In Liberia, the percentage of species reported was the same for 2000-2010 and 2011-2020 (Figure 1B).
In relation to sighting and non-sighting records (Figures 1C, D), in Morocco, most non-sightings were reported in 2000-2010; in Western Sahara, almost all sighting records were registered between 2011 and 2020, while all non-sighting records were gathered between 2000 and 2010. Mauritania is the state with the highest total number of records (sightings and non-sightings), with most sightings being from recent years (2011-2020) and most non-sightings having been recorded prior to 2000. In Senegal, the majority of non-sighting records are dated prior to 2000. In The Gambia and Guinea Bissau, nearly all data (sightings and non-sightings) were registered before 2000. In Guinea, while sightings were mostly for 2011-2020, most non-sightings were registered between 2000 and 2010. For Sierra Leone, all sighting records were collected between 2011 and 2020, while most sightings from Liberia are from 2000 to 2010; no non-sighting records were located for either country.
Of the 36 species reported, Delphinus delphis (common dolphin) and Tursiops truncatus (bottlenose dolphin) were reported by the highest number of sources (by 23 and 20 sources, respectively). S. teuszii is the species with the highest number of sightings (355), closely followed by T. truncatus (353). While many of these sightings were reported prior to 2000 (197 and 140, respectively), these two species were also the two most frequently sighted species prior to 2000.
Considering non-sightings, D. delphis has the highest number of records overall (approximately 660), most of them reported before 2000 (310 non-sighting records – the highest number in this time-period for any species). It is also the species with the most sightings in the time-periods 2000-2010 and 2011-2020 (39 and 140, respectively). For the period between 2000 and 2010, Stenella coeruleoalba (striped dolphin) is the species with the most non-sighting records (263) and it is also, by far, the species with highest total number of records (sightings and non-sightings) across the three time-periods. In the time-period 2011-2020, with 12 records, Phocoena phocoena (harbor porpoise) was the species with the most non-sightings (Tables 1, 2).

Table 2 Summary of the occurrence (number of records) of cetacean species in different time-periods, according to data found in the literature, reported for north-western Africa (Morocco to Liberia), with confirmed identifications to species level.
Both Balaenoptera musculus (blue whale) and Stenella attenuata (pantropical spotted dolphin) were reported 21 times, with the majority of records occurring between 2011 and 2020. Several species are represented by fewer than 10 sightings and fewer than 10 non-sightings: Balaenoptera edeni (Bryde’s whale), Kogia breviceps (pygmy sperm whale), Kogia sima (dwarf sperm whale), Lagenodelphis hosei (Fraser’s dolphin), Lagenorhynchus albirostris (white-beaked dolphin), Mesoplodon europaeus (Gervais’s beaked whale), Peponocephala electra (melon-headed whale), Pseudorca crassidens (false killer whale), and Ziphius cavirostris (Cuvier’s beaked whale) (Table 1).
Five species are represented by only one record in the entire NWA. Balaenoptera omurai (Omura’s whale) is represented by a stranding of a juvenile in Mauritania in 2013 (Djiba et al., 2015; Jung et al., 2016). A single individual of Carepea marginata (pygmy right whale) was stranded in 1995 in The Gambia (Tsai and Mead, 2018). An individual of Feresa attenuata (pygmy killer whale) was captured off Senegal in 1958 (Van Waerebeek et al., 2000), although qualitative information in this species is also available for Senegal (Jefferson et al., 1997; Robineau and Vely, 1998; Robards and Reeves, 2011; Perrin and Van Waerebeek, 2012). There was a sighting of Lagenorhynchus acutus (Atlantic white-sided dolphin) off Gibraltar (Jefferson et al., 1997). Mesoplodon densirostris (Blainville’s beaked whale) is represented by a stranding of a single individual in Mauritania in 1992 (Robineau and Vely, 1998) plus qualitative data from Morocco or Western Sahara and between Mauritania and Senegal (Robineau and Vely, 1998; Perrin and Van Waerebeek, 2012; Masski and de Stéphanis, 2015) (Table 1; Appendix C).
Two species lack quantitative data, Eubalaena glacialis (North Atlantic right whale) and Hyperoodon ampullatus (northern bottlenose dolphin), and are documented only in the study area’s northernmost countries: E. glacialis in Morocco and Western Sahara (Robineau and Vely, 1998; Masski and de Stéphanis, 2015) and H. ampullatus in Morocco (Perrin and Van Waerebeek, 2012; Masski and de Stéphanis, 2015).
For nine species, the distribution range covers the entire study area, with missing data in just a few countries: Globicephala macrorhynchus (short-finned pilot whale), Orcinus orca (orca), Physeter microcephalus (sperm whale), S. teuszii, S. attenuata, S. clymene, Stenella longirostris (spinner dolphin), Steno bredanensis (rough-toothed dolphin), and T. truncatus. D. delphis is the only species documented in all range states of the study area (Figure 2).
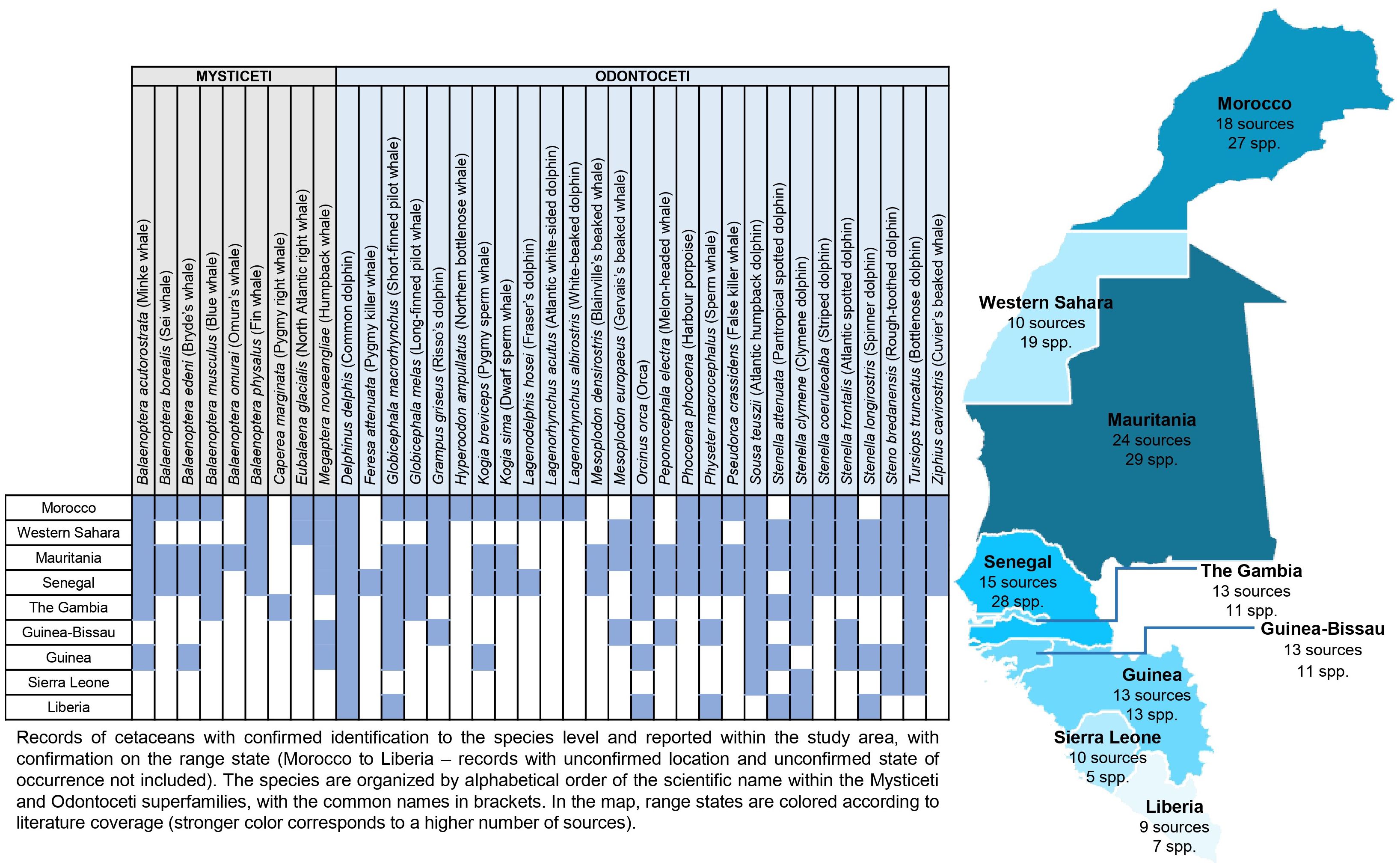
Figure 2 Species’ distribution ranges in the study area and literature coverage by range state (number of sources and species reported).
Senegal and The Gambia are the southern limits for the distribution range of 11 species: Balaenoptera borealis (Sei whale), B. musculus, Globicephala melas (long-finned pilot whale), Grampus griseus (Risso’s dolphin), K. sima, L. hosei, M. densirostris, P. phocoena, P. crassidens, S. coeruleoalba, and Z. cavirostris. For M. europaeus and P. electra, the southern limit is Guinea-Bissau.
Six species have their distribution range extending up to Guinea, albeit with missing data in the two countries with the lowest number of sources (Sierra Leone and Liberia, with 10 sources): Balaenoptera acutorostrata (minke whale), B. edeni, Balaenoptera physalus (fin whale), K. breviceps, Megaptera novaeangliae (humpback whale), and Stenella frontalis (Atlantic spotted dolphin).
Three species have their southern distribution limit at the northern boundary of the study area (Morocco/Western Sahara): E. glacialis, H. ampullatus, L. acutus, and L. albirostris.
In the compiled bibliography, it is clear that most publications on cetacean occurrence and distribution on the NWA consist of inventories, reports, or surveys, published between 2011 and 2020. However, much of the information relates to non-sighting records, notably of strandings and museum specimens, and was mostly compiled from reviews reporting on data from prior to the year 2000 (even though 6 out of 10 were published during 2011-2020). Although useful, this information may not represent the current status of cetacean communities in the region. In fact, as well as making clear the high variation in both the type and amount of information collected, both between countries and over time, reflecting a general lack of systematic monitoring, this study reveals a particular lack of monitoring effort in certain states, especially south of Senegal, from where there are few records.
Since the time-period covered here (up to 2020), to date (by June 2023), three new reviews covering part of the north of the study area were published. Cartagena-Matos et al. (2021) analyze the research topics in scientific publications published up to 2018, focusing on the Macaronesia area and the North African coast, from Morocco to Senegal. Bilal et al. (2023) reported the occurrence of L. hosei off the coast of Mauritania (a sighting of a group in November 2021). In our review, we found no records of this species in Mauritania, although it was recorded in the adjacent range states (Morocco and Senegal). Kaddouri et al. (2023) reported on trends in cetacean strandings in and around the Strait of Gibraltar, Morocco, until 2021. All the species reported in the latter article are already described for Morocco in the present review.
The results presented here highlight the need to increase the data available on cetacean occurrence (from both sightings and non-sighting records) in the NWA, and to update the inventory of cetacean species in the region. Furthermore, there is an evident urgency to strengthen monitoring/research efforts in the southernmost part of the NWA. However, in addition to the need for new monitoring and research to address the data gaps, it should be noted that the present review depended entirely on available literature and it would be useful to assemble data and information from local knowledge, e.g., via direct contacts with communities, fishers, stranding networks, museums, local authorities, NGOs and researchers.
As expected, D. delphis was the most frequently reported species from records throughout the region, consistent with the global assessment of this species in the IUCN Red List for Threatened Species (Braulik et al., 2021). Similarly, T. truncatus was recorded across most of the entire study area, consistent with wide distribution range of the species (Wells et al., 2019). Sousa teuszii was also one of the most frequently recorded species, occurring from Morocco to Sierra Leone. There are major concerns about the conservation status of this endemic species, due to its distribution being restricted to coastal areas and its exposure to threats from human activities such as overfishing, consumption of marine bushmeat (documented in Senegal; see Cosentino and Fisher, 2016), coastal development (e.g, over-exploitation of mangroves, coastal construction, aquaculture, oil and gas exploration and extraction, accidental spills, shipping, tourism, and effluents), and climate change. These increasing pressures mainly arise because of the strong human population growth in the region (Van Waerebeek et al., 2003; UNEP-CMS, 2007; Weir, 2010).
By analyzing cetacean distributions in the study area, it is possible to delimit probable boundaries of distribution range for several species in the study area. Firstly, dolphins of the genus Lagenorhynchus (L. acutus and L. albirostris), as well as E. glacialis and H. ampullatus occurred in temperate zones, not being reported south of Western Sahara. In the case of the species E. glacialis and H. ampullatus, this is consistent with the IUCN global assessment (Cooke, 2020; Whitehead et al., 2021). However, the distributions of both species of the genus Lagenorhynchus, their distribution in the northeast Atlantic Ocean have been described as being restricted to the north of the Iberian Peninsula (Kiszka and Braulik, 2018a; Braulik, 2019a). For these species, very few records were compiled within the present review and they were all from Morocco, prior to the year 2000 (Jefferson et al., 1997), at least when considering only confirmed identifications. It is possible that these records are extra-limital and/or that there has been a northward shift of these species’ distributions over the last few decades.
Secondly, there is a large number of species for which the range observed in the present study extends as far south as Senegal/The Gambia/Guinea-Bissau, thus extending to the tropical zone. This was the case for several baleen whales (B. borealis, B. musculus, and B. physalus), G. griseus, G. melas, K. sima, L. hosei, several beaked whales (M. densirostris, M. europaeus, and Z. cavirostris), P. electra, P. phocoena, P. crassidens, and S. coeruleoalba. Aside from G. melas, for which the southern limit of distribution in The Gambia (although there are quantitative data only for Morocco and Mauritania), which is further south than the limit reported in the IUCN Red List assessment (in Mauritania, Minton et al., 2018), and P. phocoena for which the distribution range is consistent with that reported by IUCN (Braulik et al., 2020), all other species are previously described as also occurring further south than was found in the present review. The aforementioned species of baleen whales, with the addition of B. edeni and B. acutorostrata, are described as being widely distributed throughout the Atlantic Ocean (Cooke and Brownell, 2018; Cooke, 2018a, b, c, d). For G. griseus, K. sima (and also K. breviceps), L. hosei, P. crassidens, and S. coeruleoalba, the IUCN reports a distribution extending along the whole the West African coast (Baird, 2018; Kiszka and Braulik, 2018b, c; Braulik, 2019b; Kiszka and Braulik, 2020). Similarly, P. electra is described across the West African coast, but with the northern limit of distribution in Mauritania (Kiszka and Brownell, 2019) which is consistent with the present review. As for the three species of beaked whales, the described distributions extend across the NWA (Baird et al., 2020; Pitman and Brownell, 2020a, b). For all these species, the apparent southern distribution limit obtained with the data collected in this review may be a strong indication of the lack of monitoring effort south of The Gambia.
In relation to P. phocoena, the distribution range in the study area extends from Morocco to Senegal. Fontaine (2016) points to the need for an assessment of the status and threats faced by the southern ecotype, and putative subspecies, of P. phocoena in the eastern North Atlantic – which he refers to as P. phocoena meridionalis. This is a unique ecotype with small populations and low genetic diversity, inhabiting the coasts of Iberian Peninsula and NWA. Therefore, there is a need for further research on the impacts of exploitation and adaptation in the face of climate change (Fontaine, 2016). P. phocoena is the smallest cetacean species in the North Atlantic and generally has a coastal distribution. Nevertheless, occurrences beyond the continental shelf are also documented, for example off Greenland (Nielsen et al., 2018; Braulik et al., 2020).
In addition to D. delphis, T. truncatus, and S. teuszii, the apparent distribution range of several other species extended from the northern states (Morocco/Western Sahara) to the southern limit of the study area (Guinea to Liberia): G. macrorhynchus, O. orca, M. novaeangliae, P. macrocephalus, S. bredensis, and several species of the genus Stenella (S. attenuata, S. longirostris, S. frontalis, and S. clymene). For M. novaeangliae, the Cape Verde islands are a well-known wintering ground in the northeast Atlantic (Hazevoet and Wenzel, 2000; Hazevoet et al., 2010, 2011) but the distribution of the species in the coastal areas of the NWA is poorly studied. Nonetheless, Van Waerebeek et al. (2013) presents strong evidence that the area between Senegal and Guinea may also be a wintering and nursing ground for a south Atlantic stock.
Lastly, there are also single non-sighting records of B. omurai, C. marginata, and F. attenuata. In the case of Omura’s whale, the single record refers to the first identification of the species in African waters, through genetic methods, from a stranding near the Mauritania-Senegal border in 2013 (Djiba et al., 2015; Jung et al., 2016) – it is worth noting that the species was first discovered only 20 years ago (Wada et al., 2003) and it is possible that specimens were previously recorded as unidentified rorquals. The C. marginata was reported by Tsai and Mead (2018), who refer to a stranding in 1995 that constitutes the first record (and likely, the only known record) of the species north of the equator. The known distribution range in African waters is limited to the south of Africa (Cooke, 2018e). This stranding most likely represents an extra-limital record. Regarding F. attenuata, there was a single capture (either a directed take or a bycatch) of an individual in Senegal (Van Waerebeek et al., 2000) in 1958, although the IUCN reports a distribution range across the entire African coast (Braulik, 2018).
This review assembles an important body of knowledge on cetacean occurrence and distribution in the NWA, contributing to baseline knowledge to inform marine management strategies in the area. In particular, available data could be useful in spatial management, allowing for the identification of important areas for future protection, e.g. areas with high biological and ecological value or a high risk of anthropogenic impact on populations (e.g., through mapping of bycatch risk).
However, it is clear that the available information remains patchy, and it may not characterize the real diversity and distribution of the cetacean species inhabiting NW African waters. A substantial increase in the monitoring effort along the NWA coast is essential, especially addressing areas where there is least information (such as the south part of this region), to facilitate assessment of status and enable effective conservation measures. Besides increasing research/monitoring effort, relevant information should be acquired directly through consultation of local knowledge/expertise (local communities, fishers, stranding networks, researchers).
Finally, more effective management of industrial fishing is crucial, for example through improvements of the SFPAs, as recognized by the European Commission (2023). It is worth highlighting that Atlantic cetacean populations depend on the appropriate management of the NWA, where the fishing activity of European fleets contributes to overexploitation of fishing resources. Therefore, EU could play a major role in sustainable fishery management in the area. It is necessary to reconcile the rapid socio-economic development occurring on the coast – and the imperative to provide fish for human consumption – with the need to ensure sustainable utilization of the area´s living marine resources and to reduce or eliminate the increasing threats to cetaceans and their habitats.
AC: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. MM: Data curation, Writing – original draft, Writing – review & editing, Formal analysis, Investigation. ÁG: Writing – original draft, Writing – review & editing, Data curation, Investigation. CO-R: Writing – original draft, Writing – review & editing, Data curation, Formal analysis, Investigation. LA: Investigation, Writing – original draft, Writing – review & editing. RV: Investigation, Writing – original draft, Writing – review & editing, Data curation. MR: Supervision, Writing – original draft, Writing – review & editing. IS-P: Resources, Writing – original draft, Writing – review & editing, Supervision. GP-P: Resources, Supervision, Writing – original draft, Writing – review & editing, Formal analysis, Validation.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Ph.D. fellowships for authors AC (SFRH/BD/100606/2014), RV (SFRH/BD/144786/2019) and ÁG (PD/BD/150603/2020) were granted by Fundação para a Ciência e Tecnologia (FCT, Portugal) under the auspices of Programa Operacional Regional Norte, supported by the European Social Fund (ESF) and Portuguese funds (MECTES). This work is also partially within the REDUCE project (Horizon Europe, Project 101135583), funded by the European Union.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1427334/full#supplementary-material
Datasheet 1 | Appendix A
Table 1 | Appendix B
Datasheet 2 | Appendix C
Báez J. C., de Stephanis R., García-Ramos M. C., Ramos M. L., Pascual P. (2020). Differential space distribution of the genus Balaenoptera in the eastern tropical Atlantic Ocean. Regional Stud. Mar. Sci. 37, 101346. doi: 10.1016/j.rsma.2020.101346
Baines M. E., Reichelt M. (2014). Upwellings, canyons and whales: An important winter habitat for balaenopterid whales off Mauritania, northwest Africa. J. Cetacean Res. Manage. 14, 57–67. doi: 10.47536/jcrm.v14i0.30
Baird R. W. (2018). Pseudorca crassidens. In: The IUCN Red List of Threatened Species 2018 (Accessed 24 June 2023).
Baird R. W., Brownell R. L. Jr., Taylor B. L. (2020). Ziphius cavirostris. In: The IUCN Red List of Threatened Species 2020 (Accessed 24 June 2023).
Bamy I. L., Van Waerebeek K., Bah S. S., Dia M., Kaba B., Keita N., et al. (2010). Species occurrence of cetaceans in Guinea, including humpback whales with southern hemisphere seasonality. Mar. Biodiversity Records 3, e48. doi: 10.1017/S1755267210000427
Belhabib D., Greer K., Pauly D. (2017). Trends in industrial and artisanal catch per effort in West African fisheries. Conserv. Lett. 11, e12360. doi: 10.1111/conl.12360
Benchoucha S., Tamsouri M. N., El Aamri F. (2018). First stranding record of Kogia sima (Owen 1866) in Morocco (Strait of Gibraltar). J. Black Sea 24, 175–179.
Bilal A. S., Wagner M., Wagué A., Dia A., Van Waerebeek K. (2023). The Cetaceans of Mauritania, West Africa: a concise zoogeographical review with two new species records. J. Anim. Diversity 5, 1–35. doi: 10.3390/jan5010001
Boisseau O., Lacey C., Lewis T., Moscrop A., Danbolt M., McLanaghan R. (2010). Encounter rates of cetaceans in the Mediterranean Sea and contiguous Atlantic area. J. Mar. Biol. Assoc. United Kingdom 90, 1589–1599. doi: 10.1017/S0025315409990739
Boisseau O., Matthews J., Gillespie D., Lacey C., Moscrop A., El Ouamari N. (2007). A visual and acoustic survey for harbour porpoises off North-West Africa: further evidence of a discrete population. Afr. J. Mar. Sci. 29, 403–410. doi: 10.2989/AJMS.2007.29.3.8.339
Bowman Bishaw Gorham Chinguetti Development Project (2003). “Seabird and cetacean surveys in the vicinity of the Chinguetti oil field, offshore Mauritania,” in Woodside Australian Energy, 66pp. (Australia: Woodside Energy Pty Ltd).
Braulik G. (2018). Feresa attenuata. In: The IUCN Red List of Threatened Species 2018 (Accessed 27 June 2023).
Braulik G. (2019a). Lagenorhynchus acutus. In: The IUCN Red List of Threatened Species 2019 (Accessed 24 June 2023).
Braulik G. (2019b). Stenella coeruleoalba. In: The IUCN Red List of Threatened Species 2019 (Accessed 24 June 2023).
Braulik G., Jefferson T. A., Bearzi G. (2021). Delphinus delphis. In: The IUCN Red List of Threatened Species 2021 (Accessed 24 June 2023).
Braulik G., Minton G., Amano M., Bjørge A. (2020). Phocoena phocoena. In: The IUCN Red List of Threatened Species 2020 (Accessed 24 June 2023).
Caddell J. R. (2009). International Law and the Regulation of Cetaceans: An Analysis of the Role of Global and Regional Instruments and Institutions in the Conservation of Marine Living Resources. Cardiff University, Cardiff, Wales. PhD thesis.
Caldeira R. M. A., Groom S., Miller P., Pilgrim D., Nezlin N. P. (2002). Sea-surface signatures of the island mass effect phenomena around Madeira Island, Northeast Atlantic. Remote Sens. Environ. 80, 336–360. doi: 10.1016/S0034-4257(01)00325-1
Camphuysen C. J. (2000). Seabirds and marine mammals off West Africa – Responses 2000 cruise report (Netherlands: Netherlands Institute for Sea Research), 50pp.
Camphuysen C. J., van Spanje T. M., Verdaat H. (2012). Ship based seabird and marine mammal surveys off Mauritania, Nov-Dez 2012 – cruise report (Mauritania: Mauritanian Institute for oceanographic research and fisheries - IMROP), 73pp.
Cartagena-Matos B., Lugué K., Fonseca P., Marques T. A., Prieto R., Alves F. (2021). Trends in cetacean research in the Eastern North Atlantic. Mammal Rev. 51, 436–453. doi: 10.1111/mam.12238
Cooke J. G. (2018a). Balaenoptera acutorostrata. In: The IUCN Red List of Threatened Species 2018 (Accessed 24 June 2023).
Cooke J. G. (2018b). Balaenoptera borealis. In: The IUCN Red List of Threatened Species 2018 (Accessed 24 June 2023).
Cooke J. G. (2018c). Balaenoptera musculus. In: The IUCN Red List of Threatened Species 2018 (Accessed 24 June 2023).
Cooke J. G. (2018d). Balaenoptera physalus. In: The IUCN Red List of Threatened Species 2018 (Accessed 24 June 2023).
Cooke J. G. (2018e). Caperea marginata. In: The IUCN Red List of Threatened Species 2018 (Accessed 24 June 2023).
Cooke J. G. (2020). Eubalaena glacialis. In: The IUCN Red List of Threatened Species 2020 (Accessed 24 June 2023).
Cooke J. G., Brownell R. L. Jr. (2018). Balaenoptera edeni. In: The IUCN Red List of Threatened Species 2018 (Accessed 24 June 2023).
Correia A. M., Gandra M., Valente R., Gil Á., Rosso M., Pierce G. J., et al. (2019). A dataset of cetacean occurrences in the Eastern North Atlantic. Sci. Data 6, 177. doi: 10.1038/s41597-019-0192-1
Corten A. (2014). EU–Mauritania fisheries partnership in need of more transparency. Mar. Policy 49, 1–11. doi: 10.1016/j.marpol.2013.12.014
Cosentino A. M., Fisher S. (2016). The utilization of aquatic bushmeat from small cetaceans and manatees in South America and West Africa. Front. Mar. Sci. 3. doi: 10.3389/fmars.2016.00163
Diedhiou I., Yang Z. (2018). Senegal’s fisheries policies: Evolution and performance. Ocean Coast. Manage. 165, 1–8. doi: 10.1016/j.ocecoaman.2018.08.003
Djiba A., Bamy I. L., Bilal A. S. O., Van Waerebeek K. (2015). Biodiversity of cetaceans in coastal waters of northwest Africa: new insights through platform-of-opportunity visual surveying in 2011-2013. IOC Tech. Ser. 115, 283–297. doi: 10.13140/RG.2.1.4820.3929
Englander G., Costello C. (2023). A fish cartel for Africa. Nat. Commun. 14, 7124. doi: 10.1038/s41467-023-42886-z
European Commission. (2023). Evaluation and analysis of the Sustainable Fisheries Partnership Agreements (SFPAs) between the EU and third countries including an in-depth analysis of the sectoral support component of the SFPAs – Final report. Publications Office of the European Union, 293pp. doi: 10.2771/52188
Expósito C. G., Qninba A. (2010). Observations de Tortues, d’Oiseaux et de Mammifères marins au large des côtes atlantiques marocaines durant l’automne 2010. Bull. l’Institut Scientifique Rabat section Sci. la Vie 32, 129–132.
FAO. (2013). Marine protected areas - Country case studies on policy, governance and institutional issues. In: FAO Fisheries and Aquaculture Technical Paper (Food and Agriculture Organization of the United Nations). Available online at: http://www.fao.org/publications/card/en/c/452ece29-e667-5f47-92a5-8a0e8eb5cbab/ (Accessed 8 November 2018).
Fontaine M. C. (2016). Harbour porpoises, Phocoena phocoena, in the Mediterranean Sea and adjacent regions: biogeographic relicts of the last glacial period. Adv. Mar. Biol. 75, 333–358. doi: 10.1016/bs.amb.2016.08.006
Gray H., Van Waerebeek K. V. (2011). Postural instability and akinesia in a pantropical spotted dolphin, Stenella attenuata, in proximity to operating airguns of a geophysical seismic vessel. J. Nat. Conserv. 19, 363–367. doi: 10.1016/j.jnc.2011.06.005
Hammond P. S., Lockyer C. (1988). Distribution of killer whales in the eastern North Atlantic. Rit Fiskideildar 11, 24–41.
Hazevoet C. J., Gravanita B., López Suárez P., Wenzel F. W. (2011). Seasonality of humpback whale Megaptera novaeangliae (Borowski 1871) records in Cape Verde seas: evidence for the occurrence of stocks from both hemispheres. Zoologia Caboverdiana 2, 25–29.
Hazevoet C. J., Monteiro V., López P., Varo N., Torda G., Berrow S., et al. (2010). Recent data on whales and dolphins (Mammalia: Cetacea) from the Cape Verde Islands, including records of four taxa new to the archipelago. Zoologia Caboverdiana 1, 75–99.
Hazevoet C. J., Wenzel W. (2000). Whales and dolphins (Mammalia, Cetacea) of the Cape Verde Islands with special reference to the Humpback Whale Megaptera novaeangliae (Borowski 1871). Contributions to Zool. 69, 197–211. doi: 10.1163/18759866-06903004
IUCN World Heritage Outlook. (2020). Banc d’Arguin National Park – 2020 Conservation Outlook Assessment (International Union for Conservation of Nature’s World Heritage Outlook). Available online at: https://worldheritageoutlook.iucn.org/explore-sites/wdpaid/20388 (Accessed 19 December 2022).
Jefferson T. A., Curry B. E., Leatherwood S., Powell J. A. (1997). Dolphins and porpoises of West Africa: a review of records (Cetacea: Delphinidae, Phocoenidae). Mammalia 61, 87–108. doi: 10.1515/mamm.1997.61.1.87
Jung J., Mullié W. C., Van Waerebeek K., Wagne M. M., Bilal A. S. O., Sidaty Z. E. A. O., et al. (2016). Omura’s whale off West Africa: autochthonous population or inter-oceanic vagrant in the Atlantic Ocean? Mar. Biol. Res. 12, 66–75. doi: 10.1080/17451000.2015.1076105
Kaddouri A., Analla M., Tamsouri N., Aksissou M., Awadh H., Benchoucha S. (2023). Evaluation of cetacean strandings on the north-western coast of Morocco from 2016 to 2021. Mamm. Biol. 103, 317–328. doi: 10.1007/s42991-023-00359-2
Kiszka J., Braulik G. (2018c). Lagenodelphis hosei. In: The IUCN Red List of Threatened Species 2018 (Accessed 24 June 2023).
Kiszka J., Braulik G. (2018a). Lagenorhynchus albirostris. In: The IUCN Red List of Threatened Species 2018 (Accessed 24 June 2023).
Kiszka J., Braulik G. (2018b). Grampus griseus. In: The IUCN Red List of Threatened Species 2018 (Accessed 24 June 2023).
Kiszka J., Braulik G. (2020). Kogia sima. In: The IUCN Red List of Threatened Species 2020 (Accessed 24 June 2023).
Kiszka J., Brownell R. L. Jr. (2019). Peponocephala electra. In: The IUCN Red List of Threatened Species 2019 (Accessed 27 June 2023).
Leeney R. H., Weir C. R., Campredon P., Regalla A., Foster J. (2016). Occurrence of Atlantic humpback (Sousa teuszii) and bottlenose (Tursiops truncatus) dolphins in the coastal waters of Guinea-Bissau, with an updated cetacean species checklist. J. Mar. Biol. Assoc. United Kingdom 96, 933–941. doi: 10.1017/S0025315415000661
Lydersen C., Vacquié-Garcia J., Heide-Jørgensen M. P., Øien N., Guinet C., Kovacs K. M. (2020). Autumn movements of fin whales (Balaenoptera physalus) from Svalbard, Norway, revealed by satellite tracking. Sci. Rep. 10, 16966. doi: 10.1038/s41598-020-73996-z
Mason E. (2009). High-resolution Modelling of the Canary Basin Oceanic Circulation. Universidad de Las Palmas de Gran Canaria, Las Palmas de Gran Canaria. PhD thesis.
Masski H., de Stéphanis R. (2015). Cetaceans of the Moroccan coast: information from a reconstructed strandings database. J. Mar. Biol. Assoc. United Kingdom 98, 1029–1037. doi: 10.1017/S0025315415001563
McIvor A. J., Williams C. T., Alves F., Dinis A., Pais M. P., Canning-Clode J. (2022). The status of marine megafauna research in Macaronesia: A systematic review. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.819581
Minton G., Reeves R., Braulik G. (2018).Globicephala melas. In: The IUCN Red List of Threatened Species 2018 (Accessed 27 June 2023).
Moore J. E., Cox T. M., Lewison R. L., Read A. J., Bjorkland R., McDonald S. L., et al. (2010). An interview-based approach to assess marine mammal and sea turtle captures in artisanal fisheries. Biol. Conserv. 143, 795–805. doi: 10.1016/j.biocon.2009.12.023
Mullié W. C., Wagne M. M., Elmamy C. A. A., Yahya F. M., Veen J., van Waerebeek K. (2013). Large number of stranded harbour porpoises Phocoena as by-catch victims in Mauritania (Jeju, Korea: Scientific Committee Document of International Whaling Commission), 5pp. June 2013. SC/65a/HIM03.
Nagel P., Gray T. (2012). Is the EU’s Fisheries Partnership Agreement (FPA) with Mauritania a genuine partnership or exploitation by the EU? Ocean Coast. Manage. 56, 26–34. doi: 10.1016/j.ocecoaman.2011.11.002
Nielsen N. H., Teilmann J., Sveegaard S., Hansen R. G., Sinding M. H. S., Dietz R., et al. (2018). Oceanic movements, site fidelity and deep diving in harbour porpoises from Greenland show limited similarities to animals from the North Sea. Mar. Ecol. Prog. Ser. 597, 259–272. doi: 10.3354/meps12588
Notarbartolo-di-Sciara G., Barbaccia G., Azzellino A. (1997). Birth at sea of a false killer whale, Pseudorca crassidens. Mar. Mammal Sci. 13, 508–511. doi: 10.1111/j.1748-7692.1997.tb00660.x
Perrin W. F., Van Waerebeek K. (2012). The small-cetacean fauna of the west coast of Africa and Macaronesia: diversity and distribution (Bonn, Germany: Convention on the Conservation of Migratory Species of Wild Animals), 7–17.
Pinela A. M., Borrel A., Cardona L., Aguilar A. (2010). Stable isotope analysis reveals habitat partitioning among marine mammals off the NW African coast and unique trophic niches for two globally threatened species. Mar. Ecol. Prog. Ser. 416, 295–306. doi: 10.3354/meps08790
Pitman R. L., Brownell R. L. Jr. (2020a).Mesoplodon densirostris. In: The IUCN Red List of Threatened Species 2020 (Accessed 24 June 2023).
Pitman R. L., Brownell R. L. Jr. (2020b).Mesoplodon europaeus. In: The IUCN Red List of Threatened Species 2020 (Accessed 24 June 2023).
Robards M. D., Reeves R. R. (2011). The global extent and character of marine mammal consumption by humans, (1970-2009). Biol. Conserv. 144, 2770–2786. doi: 10.1016/j.biocon.2011.07.034
Robineau D., Vely M. (1998). Ces cétacés des côtes de Mauritanie (Afrique du Nordouest). Particularltés et variations spatio-temporelles de repartition: role des facteurs oceanographiques. Rev. d Ecologie (Terre Vie) 53, 123–152.
Rojo–Nieto E., Álvarez–Díaz P. D., Morote E., Burgos–Martín M., Montoto–Martínez T., Sáez–Jiménez J., et al. (2011). Strandings of cetaceans and sea turtles in the Alboran Sea and Strait of Gibraltar: a long–term glimpse at the north coast (Spain) and the south coast (Morocco). Anim. Biodiversity Conserv. 34, 151–163. doi: 10.32800/abc.2011.34.0151
Russell G., Sánchez-Cabanes A., Nimak-Wood M. (2018). The autumn occurrence and spatial distribution of cetaceans in the waters of Mauritania during a geophysical survey in 2012. Afr. J. Mar. Sci. 40, 371–381. doi: 10.2989/1814232X.2018.1531786
Sala I., Caldeira R. M. A., Estrada-Allis S. N., Froufe E., Couvelard X. (2013). Lagrangian transport pathways in the northeast Atlantic and their environmental impact. Limnol. Oceanogr. – Fluids Environ. 3, 40–60. doi: 10.1215/21573689-2152611
Satia B. P. (2016). An overview of the large marine ecosystem programs at work in Africa today. Environ. Dev. 17, 11–19. doi: 10.1016/j.envdev.2015.06.007
Subregional Fisheries Commission (2003). Regional Strategy for Marine Protected Areas in West Africa. Available online at: http://www.fibarguin.org/var/plain/storage/original/application/b5cbded23840211ccdf9d60838afb302.pdf (Accessed 8 November 2018).
Tickler D., Meeuwig J. J., Palomares M. L., Pauly D., Zeller D. (2018). Far from home: Distance patterns of global fishing fleets. Environ. Stud. 4 (8), eaar3279. doi: 10.1126/sciadv.aar3279
Trouillet B., Guineberteau T., Bernardon M., Le Roux S. (2011). Key challenges for maritime governance in West Africa: Fishery-based lessons from Guinea and Mauritania. Mar. Policy 35, 155–162. doi: 10.1016/j.marpol.2010.09.001
Tsai C. H., Mead J. G. (2018). Crossing the equator: a northern occurrence of the pygmy right whale. Zoological Lett. 4, 30. doi: 10.1186/s40851-018-0104-7
Tudela S., Kai A., Maynou F., El Andalossi M., Guglielmi P. (2005). Driftnet fishing and biodiversity conservation: the case study of the large-scale Moroccan driftnet fleet operating in the Alboran Sea (SW Mediterranean). Biol. Conserv. 121, 65–78. doi: 10.1016/j.biocon.2004.04.010
Tulp I., Leopold M. F. (2004). Marine mammals and seabirds in Mauritanian waters. Pilot study April 2004 - Internal Report (Wageningen UR: RIVO-Netherlands Institute for Fisheries Research - Animal Sciences Group), 42 pp.
UNEP-CMS. (2007). Conservation Status of the Atlantic Humpback Dolphin, a Compromised Future? (Bonn, Germany: Convention on the Conservation of Migratory Species of Wild Animals), 10pp.
UNEP-CMS. (2008).Action plan for the conservation of small cetaceans of Western Africa and Macaronesia. In: Annex II to the Memorandum of Understanding Concerning the Conservation of the Manatee and Small Cetaceans of Western Africa and Macaronesia (CMS - Convention on the Conservation of Migratory Species of Wild Animals). Available online at: http://www.cms.int/en/document/action-plan-conservation-small-cetaceans-western-africa-and-macaronesia (Accessed 8 November 2018).
UNEP-CMS. (2012).Conserving cetaceans and manatees in the western African region. In: CMS Technical Series No. 26 (CMS - Convention on the Conservation of Migratory Species of Wild Animals). Available online at: http://www.cms.int/en/publication/conserving-cetaceans-and-manatees-western-african-region-ts-no-26 (Accessed 8 November 2018).
Van Waerebeek K., André M., Sequeira M., Martín V., Robineau D., Collet A., et al. (1999). Spatial and temporal distribution of the minke whale, Balaenoptera acutorostrata (Lacépède 1804), in the southern northeast Atlantic Ocean and the Mediterranean Sea, with reference to stock identity. J. Cetacean Res. Manage. 1, 223–237. doi: 10.47536/jcrm.v1i3.470
Van Waerebeek K., Barnett L., Camara A., Cham A., Diallo M., Djiba A., et al. (2003). Conservation of Cetaceans in The Gambia and Senegal 1999-2001, and Status of the Atlantic humpback dolphin (Bonn, Germany: UNEP/CMS Secretariat), 56 pp.
Van Waerebeek K., Djiba A., Krastad J. O., Samba Ould Bilal A., Almeida A., Mass Mbye E. (2013). New evidence for a South Atlantic stock of humpback whales wintering on the Northwest African continental shelf. Afr. Zool. 48, 177–186. doi: 10.3377/004.048.0120
Van Waerebeek K., Ndiaye E., Djiba A., Diallo M., Murphy P., Jallow A., et al. (2000). A survey of the conservation status of cetaceans in Senegal, The Gambia and Guinea-Bissau (Bonn, Germany: UNEP/CMS Secretariat), 80pp.
Van Waerebeek K., Uwagbae M., Segniagbeto G., Bamy I. L., Ayissi I. (2017). New records of Atlantic humpback dolphin (Sousa teuszii) in Guinea, Nigeria, Cameroon and Togo underscore pressure from fisheries and marine bushmeat demand. Rev. d’Ecologie (Terre Vie) 72, 192–205. doi: 10.3406/revec.2017.1885
Wada S., Oishi M., Yamada K. (2003). A newly discovered species of living baleen whale. Nature 426, 278–281. doi: 10.1038/nature02103
Weir C. R. (2010). A review of cetacean occurrence in West African waters from the Gulf of Guinea to Angola. Mammal Rev. 40, 2–39. doi: 10.1111/j.1365-2907.2009.00153.x
Weir C. R. (2016). Atlantic humpback dolphins Sousa teuszii in the Saloum Delta (Senegal): distribution, relative abundance and photo-identification. Afr. J. Mar. Sci. 38, 385–394. doi: 10.2989/1814232X.2016.1216893
Weir C. R., Coles P., Ferguson A., May D., Baines M., Figueirdo I., et al. (2014). Clymene dolphins (Stenella clymene) in the eastern tropical Atlantic: distribution, group size, and pigmentation pattern. J. Mammal. 95, 1289–1298. doi: 10.1644/14-MAMM-A-115
Weir C., Collins T. (2015). A review of the geographical distribution and habitat of the Atlantic humpback dolphin (Sousa teuszii). Adv. Mar. Biol. 72, 79–117. doi: 10.1016/bs.amb.2015.08.001
Weir C. R., Pierce G. J. (2013). A review of the human activities impacting cetaceans in the eastern tropical Atlantic. Mammal Rev. 43, 258–274. doi: 10.1111/j.1365-2907.2012.00222.x
Wells R. S., Natoli A., Braulik G. (2019). Tursiops truncatus. In: The IUCN Red List of Threatened Species 2019 (Accessed 24 June 2023).
Keywords: species inventory, species richness, range, occurrence data, spatiotemporal, literature coverage
Citation: Correia AM, Mihova M, Gil Á, Oliveira-Rodrigues C, Afonso L, Valente R, Rosso M, Sousa-Pinto I and Pierce GJ (2024) Cetaceans of north-western continental Africa (Morocco to Liberia): diversity and distribution. Front. Mar. Sci. 11:1427334. doi: 10.3389/fmars.2024.1427334
Received: 03 May 2024; Accepted: 01 July 2024;
Published: 19 July 2024.
Edited by:
Roberto Carlucci, University of Bari Aldo Moro, ItalyReviewed by:
Angela R. Szesciorka, Pacific Northwest National Laboratory (DOE), United StatesCopyright © 2024 Correia, Mihova, Gil, Oliveira-Rodrigues, Afonso, Valente, Rosso, Sousa-Pinto and Pierce. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana M. Correia, YW5hbWFmYWxkYWNvcnJlaWFAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.