
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Mar. Sci. , 22 October 2024
Sec. Discoveries
Volume 11 - 2024 | https://doi.org/10.3389/fmars.2024.1425511
This article is part of the Research Topic New Observations on the Behavior, Ecology, and Biology of Sharks and Rays View all 18 articles
 Francesco Ferretti1*
Francesco Ferretti1* Brendan D. Shea1,2
Brendan D. Shea1,2 Chiara Gambardella3,4
Chiara Gambardella3,4 Jeremy F. Jenrette1
Jeremy F. Jenrette1 Stefano Moro4
Stefano Moro4 Khaled Echwikhi5
Khaled Echwikhi5 Robert J. Schallert6,7
Robert J. Schallert6,7 Austin J. Gallagher2
Austin J. Gallagher2 Barbara A. Block6
Barbara A. Block6 Taylor K. Chapple8
Taylor K. Chapple8White sharks (Carcharodon carcharias) are among the most widespread, charismatic, and studied predators in the ocean. However, their conservation status is concerning in many ocean sectors, most notably the Mediterranean Sea, which hosts one of the least known and most endangered populations globally. Though they were historically abundant and widely distributed in the region, Mediterranean white sharks have declined to dangerously low abundance levels, impacted by centuries of coastal and, more recently, industrial fishing. The IUCN lists this species as Critically Endangered in the region, but information about its current abundance and ecology is scarce, hindering effective management and conservation. Here, we describe our initial effort to find and track the remaining Mediterranean white sharks and report what we have learned from these activities. In 2021-2023, we conducted three pilot expeditions in the Sicilian Channel, covering four major sites, collecting 159 eDNA samples, and carrying out 359 hrs of pelagic mid-water baited video surveys, 43 hours of deep-water benthic baited video surveys, and 111 hrs of fishing. Baited video surveys detected 42 species of bony fishes, elasmobranchs, marine mammals, and turtles. We detected white sharks at four sites from eDNA samples. Though we did not observe white sharks directly, these activities supported the identification of one of the last strongholds of this population in the region and started a multi-institutional white shark conservation program in the Mediterranean Sea, aiming to track the last white sharks in the region, estimate their abundance and extinction risk, characterize the species’ ecology and inform management and conservation.
White sharks (Carcharodon carcharias) have been sporadically but regularly detected in the Mediterranean Sea throughout history (Fergusson, 1996; Gubili et al., 2011; Boldrocchi et al., 2017; Tiralongo et al., 2020). They are part of a genetically distinct population for which we know very little about their ecology and biology. Recent analyses of incidental catches and sightings indicate these sharks are some of the largest on the planet, reaching sizes up to 7 meters (Moro et al., 2020; De Maddalena and Heim, 2012); however, these data also suggest that this population is one of the most endangered globally. The IUCN has classified the Mediterranean white shark as Critically Endangered (Dulvy et al., 2016). Historically abundant and widely distributed, Mediterranean white sharks have declined to dangerously low abundance levels in the region, impacted by centuries of coastal and, more recently, industrial fishing (Moro et al., 2020). Despite the pressing need to effectively protect these species, we know very little about them outside of incidental and fisheries interactions, inhibiting their effective management and conservation. Therefore, it is now crucial to facilitate more research and exploration to advance knowledge and conservation of this population. Specifically, we need to increase our understanding of the Mediterranean white sharks’ population size and structure, as well as their distribution, migration patterns, and potential connectivity with the Atlantic populations. This information is crucial to plan effective conservation and recovery programs.
White sharks are present in the Mediterranean Sea, but the very low population density and the absence of conventional aggregation sites, such as pinniped colonies (Klimley and Anderson, 1996), have limited monitoring and exploration of this population and consequently hindered research and conservation. Previous monitoring studies that attempted to deploy electronic tags on Mediterranean white sharks were hampered by limited information on the species’ seasonality and distribution and were not able to find live individuals (Soldo and Pierce, 2005; Micarelli et al., 2023). The limited Mediterranean white shark samples available in zoological and osteological museum collections have been leveraged in genetic and isotopic analyses to characterize the population’s phylogenetics, biogeography, and connectivity and reconstruct important ecological and evolutionary aspects of this population (Leone et al., 2020; Gubili et al., 2011; Wagner et al., 2024). However, while these valuable efforts continue, detecting individuals in the wild remains a core challenge.
Given the importance of interacting with live animals in the Mediterranean to characterize the ecology of the species there and worldwide, and the established challenge this presents, we recognized the need to develop a broader, more holistic suite of approaches to interact with these animals in the wild. Here, we report on the planning and execution of three multifaceted research expeditions in the Mediterranean Sea in search of the white shark.
Expeditions were prepared throughout the year, and involved securing funds for field operations, developing occurrence models for predicting shark occurrence, and establishing partnerships with local researchers and operators able to provide local expertise and infrastructure and facilitate the on-site operations.
With an integrative and historical approach, we analyzed all opportunistic white shark occurrence records in the Mediterranean Sea, building on the work of Moro et al. (2020), and produced high-resolution species occurrence predictions (HRM) (Jenrette et al., 2023; Panunzi et al., 2024) for the region. These models, Jenrette et al. (2023) for 2021-2022 and Panunzi et al. (2024) for 2023, were used to estimate the relative probability of observing white sharks in different Mediterranean sectors and seasons and informed expedition periods and locations for search and survey operations.
Our spatial analyses, updated with real-time sightings and environmental information, identified the Sicilian Channel as a seasonal white shark occurrence hot spot in the early summer. These findings corroborated anecdotal evidence that this region may be a nursery area for small sharks (Fergusson, 2002; De Maddalena and Heim, 2012), and an important feeding ground for adults following annual spawning aggregations of bluefin tuna (Thunnus thynnus) between May and June.
We then developed the on-site capacity and local infrastructure to support expeditions and physically prepared for the field operations (e.g., permit applications, bait acquisition, gear preparation, etc.). In each expedition, we identified a primary port as the base of our onshore operations. In 2021, we based our expedition in Marsala, Sicily, the most important longline fishing port in the Sicilian Channel (Borsellino et al., 2006; Pignalosa et al., 2019) and a major hub for the bluefin tuna fishery (Cermeño et al., 2015). Bluefin tuna are expected to be a major white shark prey source in the Mediterranean (Moro et al., 2020), and, thus, likely attract white sharks together with other shark predators during their spawning migration through the Sicilian channel. In 2022, we focused our operations on Lampedusa, a small island (20 km2 surface) occupying a strategic position in the middle of the Sicilian Channel. In 2023, we worked out of Monastir (Tunisia), and Lampedusa. Monastir, located across the Sicilian Channel from Lampedusa, is a convenient touristic marina near an important fishing port, Teboulba, in the northern part of the Tunisian plateau.
We began on-the-ground work by first conducting interview surveys throughout the year in the above sectors to collect additional historical data on local white shark catches and sightings, focusing on the main ports of interest in the Sicilian Channel, including ports in Sicily, Lampedusa, Tunisia, and Malta. Specifically, we conducted interview surveys to collect additional historical data on local white shark catches and sightings in the Italian fishing ports of: Sciacca, Mazara del Vallo, Marsala, Trapani, and Favignana in 2021; and Lampedusa in 2022. In 2023, we began an systematic monitoring of the main Tunisian fishing ports (Monastir, Teboulba, Djerba, Zarzis, Kelibia). A detailed description of these surveys and monitoring efforts will be included in a separate contribution. For the scope of our field expeditions, we used the most immediate information about historical interaction hotspots, i.e., sectors fishers indicated as likely to result in white shark encounters, and additional photographs to update our white shark record database feeding the species occurrence models (Panunzi et al., 2024). Finally, local collaborators and our global team visited the primary ports in the weeks leading up to the expedition to further strengthen our partnerships with local fishers and their communities.
From June 11th through June 23rd, 2021, we spent a total of 10 days sampling. We sampled for five days around the Egadi Islands, primarily Marettimo and Favignana. We then spent one day each at Pantelleria Shoal and “Banco Murena” (Moray Eel Bank), before traveling to Pantelleria and then Lampedusa. We spent three days sampling around Lampedusa, including the area surrounding the island of Lampione, a marine protected area located 13 miles west of the island, within the Isole Pelagie Marine Reserve. During this expedition, we worked from a number of vessels-for-rent, including a 15 m catamaran and two 5 m, 40HP, zodiacs (Figure 1). From June 1st through June 8th, 2022, we focused on the area surrounding Lampedusa, including Lampione and the Secca di Levante banks to the east, executing our work from a 33-foot ARS Mare yacht with 2 inboard 110 HP Caterpillar engines. From May 15th to June 1st, 2023, we spent a total of 18 days, including three overnight longline trips, sampling in the region from a mixture of Tunisian and Italian fishing vessels. We first surveyed the waters off Monastir before moving our base of operations to Lampedusa. We also completed two eDNA transects from Monastir to Lampedusa on May 21st and from Lampedusa to Malta on June 1st. Operations were supported by a 27 m sail yacht ‘Blue Titan’ which acted as a floating accommodation, cruising vessel, support boat for zodiac operations, and platform to conduct eDNA sampling and analysis. Fishing operations were conducted from two professional longlining fishing boats in Tunisia and Lampedusa. BRUV deployments were carried out from a 7 m, 225HP zodiac rented in Lampedusa.

Figure 1. (A) Jeremy Jenrette and Brendan Shea deploying a drop camera off Marettimo (left) from a 5 m Zodiac rented in Marsala in 2021; (B) 42 foot catamaran chartered from Sailing Sicily in 2021; (C) 33-foot ARS Mare yacht used in 2022; (D) Blue Titan, sailboat used as a survey support boat; (E) 7 m zodiac rented in Lampedusa for BRUV deployments; (F) Amir el Bahr, Tunisian fishing vessel used for fishing and tagging operations.
At each survey location, we sampled water to detect white shark DNA and confirm their presence (Supplementary Figure S1). At each station, we collected water from 0-100 m depth. We used filtered mitochondrial DNA, amplified using white shark specific primers (Lafferty et al., 2018), and visualized the target gene via gel electrophoresis (Jenrette et al., 2023). This process, from water collection to visualization of white shark eDNA, could be completed in as little as 4.5 hours. This quasi-real-time knowledge of species presence allowed us to hindcast where sharks likely shed their DNA (Dagestad et al., 2018) and adapt sampling and field operations as we proceeded with the surveys. Later onshore, we performed library preparation and sequencing of positive samples to confirm white shark presence (Johri et al., 2019; Truelove et al., 2019) using the approach detailed in Jenrette et al. (2023).
After DNA sampling, we deployed a line of pelagic-baited remote underwater cameras (Pelagic-BRUVs) to look for the presence of sharks in our sampling sectors. We used a set of 4 BRUVs spaced every 200 meters and deployed at 10 meters depth. Each BRUV unit consisted of a metal frame with one or two GoPro Hero 8 cameras pointed toward a bait canister extending from the mainframe. In 2021, the BRUV longline had units with a 5° convergence stereo-camera set-up, which were tethered in-line to the research vessel as we drifted, allowing the team to deploy additional chum consistently. In 2022, we used a single camera on each BRUV and fixed the longline using an anchor tethered to the buoy associated with the most up-current BRUV. In 2023, we used a stereo-camera setup again, tethering the longline to a zodiac, with the entire line left to drift while the team on-board chummed continuously. BRUVs recorded videos for 1-5 hours per deployment (Figure 2).
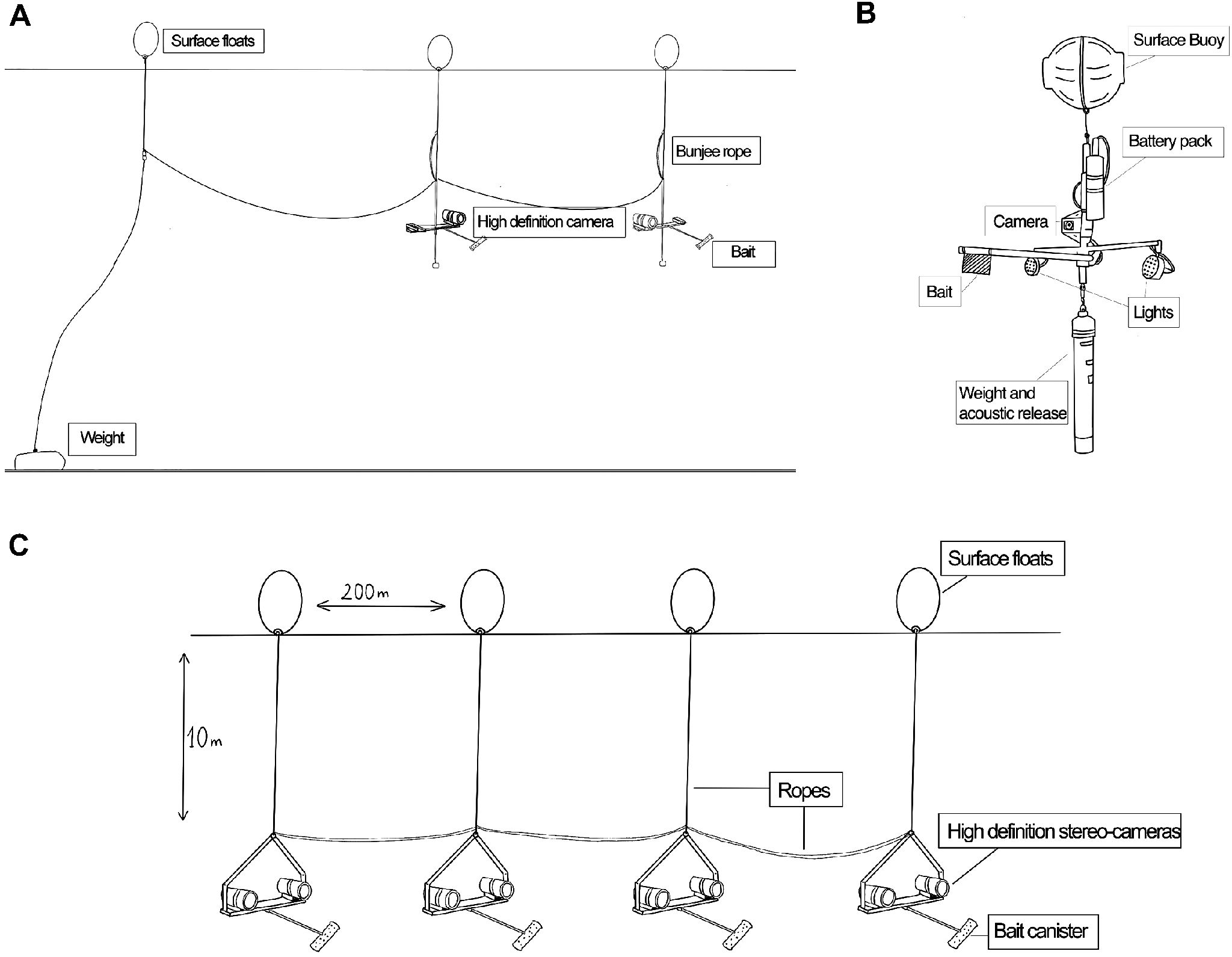
Figure 2. Schemes of longline BRUVs and drop-cams used during the 3 expeditions. (A) Longline BRUV rigging used in 2022 with mono-camera setup; (B) drop-cams used in 2021; and (C) Longline BRUV rigging used in 2021 and 2023 with stereo-camera setup.
In 2021, we also deployed deep-water BRUVs (or drop cams) to test for the presence of white sharks in deeper waters close to the seafloor. The deep-water system consisted of a vertical frame, 20 kg of weights, floats, and a housed, pressure-rated GoPro camera (to record up to 6 hours in HD) mounted 1 meter above the seafloor. Integrated lights illuminated the seafloor in the area, and a bait bag was attached to the main frame to attract white sharks. The units were recovered via acoustic release. All BRUVs were baited with tuna remains, sardines, or general fish scraps.
BRUV videos were viewed and annotated daily. In addition to being viewed by a team member for annotation, videos were processed with a shark detection software for automatic shark identification and classification (Jenrette et al., 2022) as a cross-check and for testing a novel autonomous BRUV postprocessing system. Videos were played back using VLC media player (Version 3.0.6) or QuickTime Player (Version 10.5). We collected information for any identifiable species observed. Annotation data consisted of the date, position of the camera in the longline, species name, the timestamps of an individual’s entry and exit from the camera frame, and MaxN - a relative abundance index equal to the maximum number of conspecifics observed in any single video frame during a deployment (Langlois et al., 2020; Whitmarsh et al., 2017)
While BRUVs were deployed, we continuously deployed additional attractants, including blood, oil, and fish pieces (preferentially bluefin tuna), to draw individuals close to the boat and the BRUV longline. In 2021, we almost exclusively chummed with tuna carcasses. In 2022, we used bluefin tuna, bottlenose dolphin, and a range of other pelagic and demersal fish scraps available in the local fish markets. The bottlenose dolphin was sourced from the National Research Council in Mazara del Vallo, Sicily, which systematically recovers and necropsies carcasses stranded in the Sicilian Channel. In 2023, we used a diverse array of species, preferentially scombrids. We generally deployed between 70 and 240 kg of attractant each day.
Several different fishing methods were employed over the years based on gear availability and location. Free-drifting longline deployments were deemed hazardous in the surveyed sectors, given the high marine traffic characteristic of the region and the resultant risk of losing gear or damaging vessels. Therefore, in 2021 we used baited handlines terminating at 14/0 circle hooks that were deployed from the research vessel and actively fished by team members. We spent approximately 24 hours fishing in 2021. In 2022, we used stand-up rod-and-reel set-ups, aiming to cover a greater portion of the water column while fishing. For stand-up rod-and-reel fishing, we used Penn 130ST International II reels mounted on curved rod butts, which were attached to ∼150 cm rod tips and rigged with 130–250 lb test monofilament fishing line and a leader consisting of a 152-cm long, 480 lb test braided stainless steel wire terminating in either a 16/0, 18/0, or 20/0 circle hook. We fished for approximately 35 hours in 2022. In 2023, we worked with commercial longline vessels to deploy short bottom longline sets targeted to catch juvenile white sharks. We conducted three longline trips, deploying four bottom longline sets and one meso-pelagic longline set. One fishing trip was completed off Monastir and two off Lampedusa (Figure 3; Supplementary Table S1). In total, we spent 52 hours fishing in 2023, deploying 4,350 hooks.
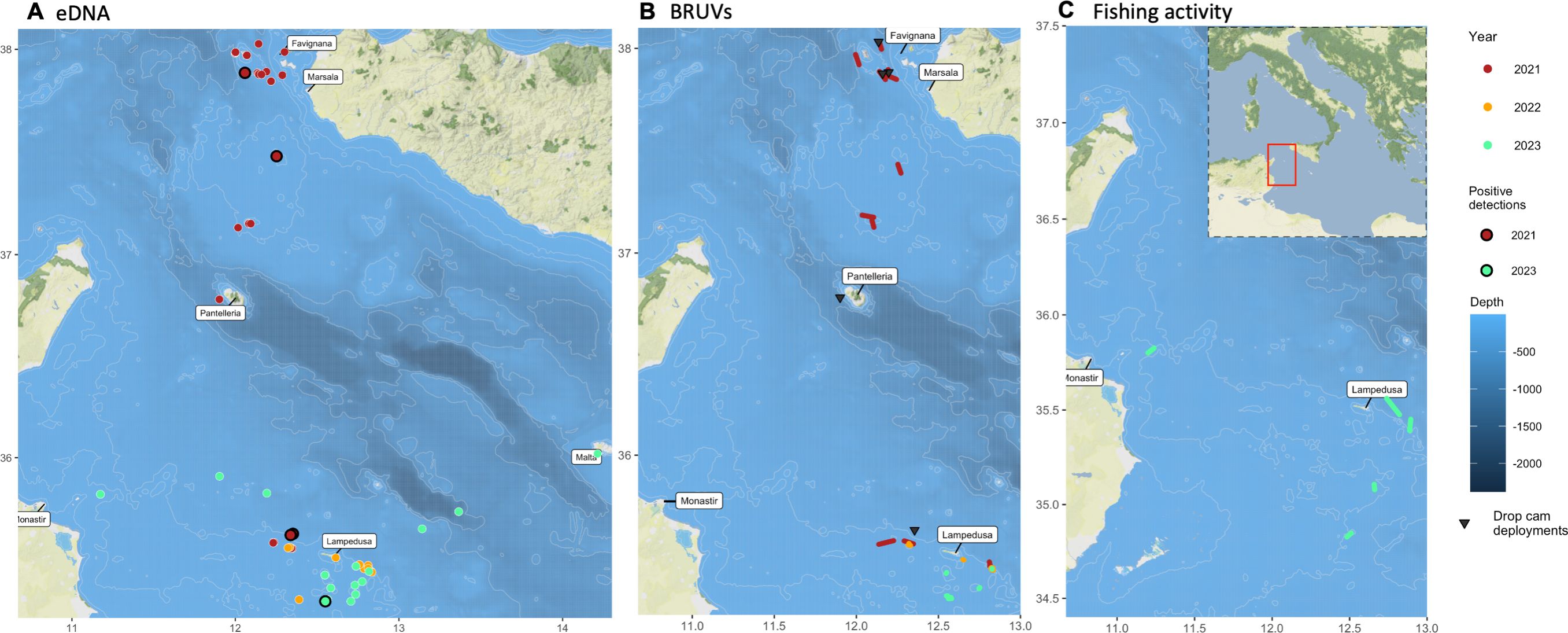
Figure 3. Map of the sampling stations surveyed during the expedition. (A) indicates locations where we sampled eDNA. Dots are color-coded by collection years. Positive detections are indicated with black borders. (B) shows the pelagic BRUV (colored segments) and drop-cam deployments (triangles). (C) indicates longline deployments.
We were prepared with a variety of electronic tags in the event we encountered a white shark, including both satellite and archival tag types. Large sharks that were attracted to the chum but not physically captured were to be tagged using pop-off satellite archival tags (PSATs; Model: MiniPAT; Manufacturer: Wildlife Computers, Redmond, WA, USA) deployed from a tagging pole following Jorgensen et al. (2010). PSATs were programmed with a range of deployment times 180-270 days, with the mortality switch and depth-threshold release (set to 1800 m) both enabled. In the event we physically captured an animal, we also had non-satellite linked archival tags (model: TDR-Mk9-286F, Manufacturer: Wildlife Computers, Redmond, WA, USA) and fin-mounted, satellite-linked Smart Positioning and Temperature (SPOT) tags (model: SPOT5; Manufacturer: Wildlife Computers) ready for deployment.
In all expeditions together, we conducted 359 hours of BRUV deployments, 43 hours of drop cam deployments, 52 hours of longline fishing, ∼35 hours of rod-and-reel fishing, and ∼24 hours of handline fishing, and collected 159 water samples for eDNA analysis. Using these methods, we identified multiple megafauna species, such as bluefin tunas, mako sharks, turtles, dolphins, mobulids, and other species (Supplementary Table S2).
Mid-water BRUVs detected 16 species of bony fishes, elasmobranchs, and marine mammals (Supplementary Table S2). We deployed 7 drop cams in 2021 at depths ranging from 73 to 651 meters (Figure 3). Drop cams detected 26 species, including 7 elasmobranchs and 19 bony fishes (Supplementary Table S3).
No animals were encountered while fishing with handlines or rod-and-reel. However, our 2023 longlining operations in Tunisian and Lampedusa were more productive, catching 15 species of fish, and invertebrates (Supplementary Table S1), and allowed us to tag a juvenile shortfin mako shark (Isurus oxyrinchus) for the first time in the Mediterranean Sea (Supplementary Table S1, Shea et al. submitted manuscript1). The average catch rate was 18.81 individuals (any species) per thousand hooks (range: 7.5 - 54.17). The highest catch rates (54.17) were recorded when catching with mesopelagic longlines, while the bottom sets recorded an average of 10 individuals per thousand hooks. Meso-pelagic longlines were expected to have a lower chance of catching juvenile white sharks but also to increase the range of habitats to be sampled by the gear, which was slightly modified to probe pelagic and demersal environments, i.e. increasing the probability of a diverse shark by-catch.
Although no white sharks were observed from the boat at any time, we successfully detected the presence of white shark DNA in 5 samples out of 159 from 2021-2023, across the Sicilian Channel; four in 2021, which included a false-negative detection while on board (Jenrette et al., 2023), and one in 2023. Positive samples were collected at the Pantelleria Banks, northern and southern ends of Lampedusa, and the Egadi Islands (Figure 3). These results confirmed that white sharks were in the area, spatially (within 25 km) and temporally (within 48 hrs) close to at least four sampling locations during our expedition (Jenrette et al., 2023; Collins et al., 2018).
We carried out three white shark research expeditions in the Mediterranean Sea, one of the world’s most heavily utilized and inhabited ocean regions, where this species is rare and sparse (Moro et al., 2020; Coll et al., 2010). When we started this program, we did not know where to systematically encounter white sharks in the region and previous work (i.e., Soldo and Pierce, 2005) suggested interacting with live animals would be difficult. This led to the planning and execution of a series of pilot expeditions to expand the tools and approaches conventionally used to find these animals (Soldo and Pierce, 2005; Micarelli et al., 2023) and increase our understanding of their ecology and biology. Over these three years, we did not directly interact with live white sharks, but collected novel information on their current population density, catches, historical hotspots, and current presence and interactions with fisheries, leading us to identify the remaining stronghold of this population in the Mediterranean Sea.
Detecting white shark eDNA supported our predictions of where and when finding the animals in the region. However, our cruises, combined with year-round investigations over the last three years, suggest that the Mediterranean White shark population may be smaller and more fragmented than previously thought (Moro et al., 2020; Panunzi et al., 2024). Therefore, a more focused effort is required to document and tag these elusive animals. The eDNA assay results combined with particle drift models suggested that white sharks were likely within a radius of 13.5 nautical miles from our sampling sites within a period of two days (Jenrette et al., 2023) (Figure 3). Despite the population being seemingly sparse and elusive, the rapid onboard sampling and processing gave us an early qualitative alert to the presence of individuals. Follow-up analyses with highly sensitive detection assays refined these results, leading us toward sectors where these animals may have a systematic presence. Continued development of quasi-real-time eDNA detection approaches and the use of specific primers for the Mediterranean population can help future expeditions to more rapidly adapt sampling strategies in response to positive detections.
We did not detect white sharks on pelagic BRUVs or drop cams; this is not surprising given their scarcity and typical sighting rates at other robust white shark hotspots. In Cape Cod (Northwest Atlantic), 1-hour deployments recorded an average of 0.29 individuals hr−1 (Shea et al., 2020) and in Port Stephens, New South Wales (Australia), 0.07 sharks hr−1 were recorded from BRUV deployments (Harasti et al., 2016). These locations are small hotspots with large numbers of adults and juveniles, respectively (Harasti et al., 2016; Bruce and Bradford, 2012; Bruce et al., 2013). Additionally, pelagic BRUVs have a lower detection capability than benthic ones as they probe habitats with a sparser animal density. Yet, they are as efficient as scientific longline surveys to detect changes in relative abundance (Santana-Garcon et al., 2014). Our pelagic BRUVs recorded a MaxN hr−1of 0.0057. A similar study in Western Australia recorded a combined MaxN hr−1 of 0.311 for nine shark species detected (Santana-Garcon et al., 2014). These estimates underscore the lack of observations in the present study, which, however, is in line with other indices detected in the Mediterranean Sea. A recent pelagic BRUV survey around the Balearic islands (northwest Mediterranean) successfully recorded 11 blue sharks (Prionace glauca), and a bluntnose sixgill shark (Hexanchus griseus), generating a comparable shark detection rate (0.0064 sharks hr−1). This study employed a modified BRUV system capable of 24-hour continuous deployment for 1,884 hours (no MaxN reported) (Prat-Varela et al., 2023). Given the depleted state of the Mediterranean and the remote nature of the pelagic realm, longer deployments such as these, or marine-adapted models of camera traps widely used in terrestrial ecology (Bicknell et al., 2016), may increase the potential for observing white sharks.
In general, our BRUV results revealed a scantness of marine megafauna and ichthyofauna in one of the most biodiverse sectors of the Mediterranean Sea (Coll et al., 2010). After over a decade, large predatory sharks remain heavily depleted in the Mediterranean Sea due to intensive coastal and industrial fishing (Ferretti et al., 2008). The Mediterranean Sea is one of the most exploited large marine ecosystems globally (Kroodsma et al., 2018), and the central Sicilian Channel’s offshore banks (i.e., Pantelleria shoal), were considered among the best spots for fishing in the past (Altobelli et al., 2017). In this sector, at least 13 nations fish with longlines, purse seines, bottom trawls, and small artisanal fishing gears in coastal and offshore waters (Figures 4A, B). Here, deploying our gear was sometimes challenging as there was a high risk of entanglement with other set fishing nets and interfering with the operations of other fishing boats in the surrounding waters. Remote sensing with Automatic Identification System (AIS) and Synthetic Aperture Radar (SAR) indicates that fishing is intense and often unmonitored throughout the Sicilian Channel and Tunisian plateau. In Tunisian and Libyan waters, where virtually no fishing boats are equipped with AIS transmitters, SAR reveals comparably intense fishing activities, even with its limitations to efficiently capture small-scale coastal fishing (Paolo et al., 2024), which is prevalent in the region.
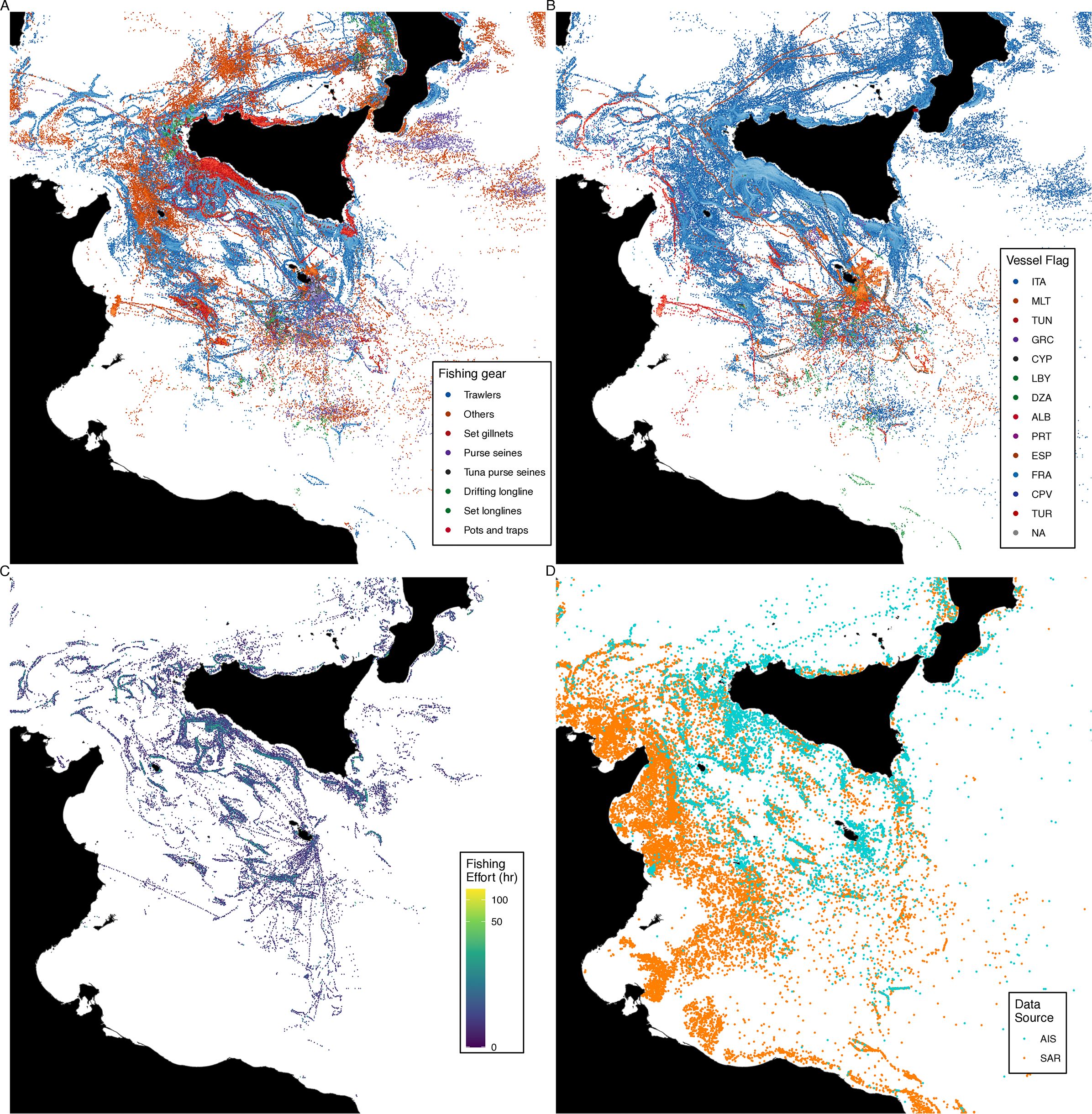
Figure 4. Fishing effort detected through AIS, by nation (A) and fishing method (B) recorded in the Sicilian Channel in 2019. (C) AIS data recorded in May 2023. (D) SAR detections matched (Green) and unmatched with AIS detections (Orange). Data are from the Global Fishing Watch database (globalfishingwatch.org).
While we could not visually detect a white shark, these pilot expeditions were essential to gather information on the challenges, available infrastructures, and logistical problems associated with successfully detecting free-living white sharks in the Mediterranean Sea. We devised multiple solutions and established a network of collaborators, building infrastructure to support a more programmatic future effort. A major challenge was not having an adequate research vessel to host scientists, equipment, needed materials, and tenders. This affected methodology and limited the operations’ geographic range. The type of vessels required for this work is expensive and rarely available through the networks of yacht owners we used for our surveys. Fishing vessels often have restrictions on their availability. Sailboats and catamarans are often available and affordable options, but these have limited cruising speed and space and inadequate auxiliary tenders or workboats. A large, refrigerated cell for bait storage (up to 240 kg day−1), allowing for continuous sampling over longer periods without the need to visit shore, and a wide deck to conduct operations safely and without damaging the boat are crucial for conducting these operations seamlessly. Due to the boat's limited storage capacity, we often faced the additional challenge of needing to continuously source bait. Through agreements with local seafood wholesalers, we stored and refrigerated bluefin tuna remains in their facilities when tuna catches were processed and accessed as needed. Still, our onboard transport capacity remained a limiting factor, forcing us to adapt at times and use different baits. The fishing vessels we used in 2023 (Figure 1) increased our chances of tagging sharks and allowed us to operate more flexibly with multiple survey approaches. However, finding fishing vessels can be difficult when survey timing overlaps with peak fishery seasons.
The three pilot expeditions we completed have evolved into a multi-institutional monitoring program on White Sharks in the Mediterranean Sea, aimed at saving this population from extinction. The data and information collected during these three years of fieldwork and activities on the ground were crucial to refining our search strategy and selecting focal areas for the following years. Our ongoing monitoring of sightings and catches continues to update habitat and distribution models, increasing our chances of interacting with white sharks in upcoming expeditions. What began as a broad search in the Sicilian Channel has narrowed as we have constantly updated search strategies in response to our data, initially focusing on the southern SC (e.g., Lampedusa) and eventually moving our effort toward Tunisia in 2023. While we sporadically recorded sightings of white sharks from other Mediterranean regions and northern sectors of the Sicilian channel (e.g., Tiralongo et al., 2020), Tunisia consistently appeared on social media and other channels with reported catches, contrasting the patterns of fishing detected in the region (Figure 4). However, a deeper investigation of fishing activities visible from SAR and AIS revealed that Tunisia’s fishing intensity was comparable to, if not greater than, the SC’s northern sectors, leading us to deepen our investigations of the interaction between fisheries and sharks in Tunisia.
White Sharks are still in the Mediterranean Sea, but we do not know how many individuals are left and where and whether they occur year-round. Therefore, it is now imperative to leverage the infrastructure and experience from our first three trips to unfold a more articulated monitoring program with year-round activities on the ground and follow-up expeditions. We will focus our efforts on Tunisia, which appears to be one of the population’s last strongholds. Here, we will intensify monitoring and sampling efforts for biological material and fisheries data. Meanwhile, we will expand our search to other Mediterranean sectors, scaling up with eDNA sampling across the region and planning explorations in other hotspots identified by our habitat and distribution models.
The original contributions presented in the study are included in the article/Supplementary Material. DNA sequences of cytochrome b gene PCR amplicons from white shark detections are accessible through the Virginia Tech Data Repository at https://doi.org/10.7294/27048106. Further inquiries can be directed to the corresponding author.
The animal study was approved by Virginia Tech Institutional Animal Care and Use Committee (IACUC) protocol for project #21-208. The study was conducted in accordance with the local legislation and institutional requirements. Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article.
FF: Writing – review & editing, Writing – original draft, Visualization, Validation, Supervision, Resources, Project administration, Methodology, Investigation, Conceptualization. BDS: Writing – review & editing, Validation, Methodology, Investigation, Formal analysis, Data curation. CG: Writing – review & editing, Visualization, Methodology, Investigation, Formal analysis. JFJ: Writing – review & editing, Visualization, Methodology, Investigation, Formal analysis. SM: Writing – review & editing, Visualization, Methodology, Investigation, Formal analysis. KE: Writing – review & editing, Resources, Investigation. RJS: Writing – review & editing, Methodology, Investigation. AJG: Writing – review & editing, Resources, Investigation. BAB: Writing – review & editing, Supervision, Resources, Methodology. TKC: Writing – review & editing, Resources, Methodology, Investigation, Funding acquisition.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Funding was provided by The Explorers Club, Discovery Channel, Sharkproject, the Bertarelli Foundation, the Augmentum platform, and individual donors. The individual donors were not involved in the study design, collection, analysis, interpretation of data, the writing of this article, or the decision to submit it for publication.
We would like to thank Gaspare Buffa for providing dolphin bait; Hani Barriche, Chiheb Lemsi and Naomi Massaccesi for conducting interviews in Malta and Tunisia; Pasquale Baiata, Andreas Barreca, and Giovanni Tarantino for their help and support during our operations in Sicily and Lampedusa. Rosie O’Donnell and Yacht for Science for their support in securing boats and funding from private donors; Luca Marsaglia and David Kroodsma of the Global Fishing Watch for sharing non-public SAR and AIS data. Blue Titan's owner Frank Peeters, Captain Jeff Doolan, and crew member Romy Hunt for their in-kind support. Captain Ugo Carsana and Marco Spoto for their skippering service in 2021.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1425511/full#supplementary-material
Altobelli C., Perzia P., Falautano M., Consoli P., Canese S., Romeo T., et al. (2017). Mediterranean banks in ebsa area: Hotspots of biodiversity under threat. Mar. Environ. Res. 131, 57–68. doi: 10.1016/j.marenvres.2017.09.005
Bicknell A. W., Godley B. J., Sheehan E. V., Votier S. C., Witt M. J. (2016). Camera technology for monitoring marine biodiversity and human impact. Front. Ecol. Environ. 14, 424–432. doi: 10.1002/fee.1322
Boldrocchi G., Kiszka J., Purkis S., Storai T., Zinzula L., Burkholder D. (2017). Distribution, ecology, and status of the white shark, Carcharodon carcharias, in the Mediterranean Sea. Reviews in fish biology and fisheries 27, 515–534.
Borsellino V., Crescimanno M., De Stefano V. (2006). Political and economic fishery management in the Channel of Sicily. Chem. Ecol. 22, S411–S432. doi: 10:1080/02757540600572214
Bruce B., Bradford R., Hughes B., Carraro R., Gallen C., Harasti D., et al. (2013). Acoustic tracking and aerial surveys of juvenile white sharks in the Hunter Catchment Management Authority region. Final Rep. Tech. Rep. 50.
Bruce B. D., Bradford R. W. (2012). “Habitat use and spatial dynamics of juvenile white sharks, Carcharodon carcharias, in eastern Australia,” in Global perspectives on the biology and life history of the white shark. Ed. Domeier M. L. (CRC Press, Boca Raton, FL), 225–254.
Cermeño P., Quílez-Badia G., Ospina-Alvarez A., Sainz-Trapaga S., Boustany A. M., Seitz A. C., et al. (2015). Electronic tagging of Atlantic bluefin tuna (Thunnus thynnus, L.) reveals habitat use and behaviors in the Mediterranean Sea. PloS One 10, e0116638. doi: 10.1371/journal.pone.0116638
Coll M., Piroddi C., Steenbeek J., Kaschner K., Ben Rais Lasram F., Aguzzi J., et al. (2010). The biodiversity of the Mediterranean Sea: estimates, patterns, and threats. PloS One 5, e11842. doi: 10.1371/journal.pone.0011842
Collins R. A., Wangensteen O. S., O’Gorman E. J., Mariani S., Sims D. W., Genner M. J. (2018). Persistence of environmental DNA in marine systems. Commun. Biol. 1, 185. doi: 10.1038/s42003-018-0192-6
Dagestad K.-F., Röhrs J., Breivik Ø., Ådlandsvik B. (2018). Opendrift v1.0: a generic framework for trajectory modelling. Geoscientific Model. Dev. 11, 1405–1420. doi: 10.5194/gmd-11-1405-2018
De Maddalena A., Heim W. (2012). Mediterranean great white sharks: a comprehensive study including all recorded sightings (McFarland).
Dulvy N., Allen D., Ralph G., Walls R. (2016). The conservation status of sharks, rays and chimaeras in the Mediterranean Sea. IUCN Centre Mediterr. Cooperation Malaga Spain 14pp.
Fergusson I. K. (1996). “Distribution and autecology of the white shark in the eastern north Atlantic Ocean and the Mediterranean Sea,” in Great White Sharks: The biology of Carcharodon carcharias. Eds. Klimley A. P., Ainley D. G. (Academic Press, Inc, San Diego, CA), 321–345.
Fergusson I. K. (2002). “Occurrence and biology of the great white shark, Carcharodon carcharias, in the Central Mediterranean Sea: A review,” in Proceedings of the 4th European Elasmobranch Association Meeting, Livorno, Italy. 7–23.
Ferretti F., Myers R. A., Serena F., Lotze H. K. (2008). Loss of large predatory sharks from the Mediterranean Sea. Conserv. Biol. 22, 952–964. doi: 10.1111/j.1523-1739.2008.00938.x
Gubili C., Bilgin R., Kalkan E., Karhan S., Jones C., Sims D., et al. (2011). Antipodean white sharks on a Mediterranean walkabout? Historical dispersal leads to genetic discontinuity and an endangered anomalous population. Proc. R. Soc. London B: Biol. Sci. 278, 1679–1686. doi: 10.1098/rspb20101856
Harasti D., Lee K., Laird R., Bradford R., Bruce B. (2016). Use of stereo baited remote underwater video systems to estimate the presence and size of white sharks (Carcharodon carcharias). Mar. Freshw. Res. 68, 1391–1396. doi: 10.1071/MF16184
Jenrette J. F., Jenrette J. L., Truelove N. K., Moro S., Dunn N. I., Chapple T. K., et al. (2023). Detecting Mediterranean white sharks with environmental DNA. Oceanography 36, 87–89. doi: 10.5670/oceanog.2023.s1.28
Jenrette J., Liu Z., Chimote P., Hastie T., Fox E., Ferretti F. (2022). Shark detection and classification with machine learning. Ecol. Inf. 69, 101673. doi: 10.1016/j.ecoinf.2022.101673
Johri S., Doane M., Allen L., Dinsdale E. (2019). Taking advantage of the genomics revolution for monitoring and conservation of chondrichthyan populations. Diversity 11, 49. doi: 10.3390/d11040049
Jorgensen S. J., Reeb C. A., Chapple T. K., Anderson S., Perle C., Van Sommeran S. R., et al. (2010). Philopatry and migration of Pacific white sharks. Proc. Biol. Sci. 277, 679–688. doi: 10.1098/rspb.2009.1155
Klimley A. P., Anderson S. D. (1996). “Residency patterns of white sharks at the South Farallon Islands, California,” in Great White Sharks (San Diego, CA: Elsevier).
Kroodsma D. A., Mayorga J., Hochberg T., Miller N. A., Boerder K., Ferretti F., et al. (2018). Tracking the global footprint of fisheries. Science 359, 904–908. doi: 10.1126/science.aao5646
Lafferty K. D., Benesh K. C., Mahon A. R., Jerde C. L., Lowe C. G. (2018). Detecting southern California’s white sharks with environmental DNA. Front. Mar. Sci. 5, 355. doi: 10.3389/fmars.2018.00355
Langlois T., Goetze J., Bond T., Monk J., Abesamis R. A., Asher J., et al. (2020). A field and video annotation guide for baited remote underwater stereo-video surveys of demersal fish assemblages. Methods Ecol. Evol. 11, 1401–1409. doi: 10.1111/2041-210X.13470
Leone A., Puncher G., Ferretti F., Sperone E., Tripepi S., Micarelli P., et al. (2020). Pliocene colonization of the Mediterranean by Great White Shark inferred from fossil records, historical jaws, phylogeographic and divergence time analyses. J. Biogeography 47, 1119–1129. doi: 10.1111/jbi.13794
Micarelli P., Reinero F. R., Marsella A., Vernelli E., Vittorini E., Monteleone L., et al. (2023). Attempts to locate and sample the white shark, Carcharodon carcharias (lamniformes: Lamnidae), along the italian coasts in the mediterranean sea. Acta Adriatica 64, 181–186. doi: 10.32582/aa.64.2.6
Moro S., Jona-Lasinio G., Block B., Micheli F., De Leo G., Serena F., et al. (2020). Abundance and distribution of the white shark in the Mediterranean Sea. Fish Fisheries 21, 338–349. doi: 10.1111/faf.12432
Panunzi G., Moro S., Marques I., Martino S., Colloca F., Ferretti F., et al. (2024). Estimating the spatial distribution of the white shark in the Mediterranean Sea via an integrated species distribution model accounting for physical barriers. Environmetrics, e2876. doi: 10.1002/env.2876
Paolo F., Kroodsma D., Raynor J., Hochberg T., Davis P., Cleary J., et al. (2024). Satellite mapping reveals extensive industrial activity at sea. Nature 625, 85–91. doi: 10.1038/s41586-023-06825-8
Pignalosa P., Pappalardo L., Gioacchini G., Carnevali O. (2019). Length-weight relationships and size distributions of Mediterranean swordfish (Xiphias Gladius L.) caught by longliners in the Mediterranean Sea. Coll. Vol. Sci. Pap. ICCAT 76, 95–106.
Prat-Varela A., Torres A., Cervantes D., Aquino-Baleytó M., Abril A.-M., Clua E. E. (2023). Improved baited remote underwater video (BRUV) for 24 h real-time monitoring of pelagic and demersal marine species from the epipelagic zone. J. Mar. Sci. Eng. 11, 1182. doi: 10.3390/jmse11061182
Santana-Garcon J., Braccini M., Langlois T. J., Newman S. J., McAuley R. B., Harvey E. S. (2014). Calibration of pelagic stereo-bruvs and scientific longline surveys for sampling sharks. Methods Ecol. Evol. 5, 824–833. doi: 10.1111/2041-210x.12216
Shea B. D., Benson C. W., de Silva C., Donovan D., Romeiro J., Bond M. E., et al. (2020). Effects of exposure to large sharks on the abundance and behavior of mobile prey fishes along a temperate coastal gradient. PloS One 15, e0230308. doi: 10.1371/journal.pone.0230308
Soldo A., Pierce R. (2005). Shark chumming in the eastern Adriatic. Annales Ser. Ist. Nat. 15, 203–208.
Tiralongo F., Monaco C., de Maddalena A. (2020). Report on a great white shark carcharodon carcharias observed off lampedusa, Italy. Annales: Ser. Hist. Naturalis 30, 181–186. doi: 10.19233/ASHN.2020.21
Truelove N., Andruszkiewicz E., Block B. (2019). A rapid environmental DNA method for detecting white sharks in the open ocean. Methods Ecol. Evol. 10, 1128–1135. doi: 10.1111/2041-210X.13201
Wagner I., Smolina I., Koop M. E. L., Bal T., Lizano A. M., Choo L. Q., et al. (2024). Genome analysis reveals three distinct lineages of the cosmopolitan white shark. Curr. Biol. 34 (15), 3582–3590.e4. doi: 10.1016/j.cub.2024.06.076
Keywords: white shark, BRUV, eDNA, field expedition, distribution models, elusive species, fishing impact, extinction risk
Citation: Ferretti F, Shea BD, Gambardella C, Jenrette JF, Moro S, Echwikhi K, Schallert RJ, Gallagher AJ, Block BA and Chapple TK (2024) On the tracks of white sharks in the Mediterranean Sea. Front. Mar. Sci. 11:1425511. doi: 10.3389/fmars.2024.1425511
Received: 29 April 2024; Accepted: 10 September 2024;
Published: 22 October 2024.
Edited by:
Carlotta Mazzoldi, University of Padua, ItalyReviewed by:
Francesco Tiralongo, University of Catania, ItalyCopyright © 2024 Ferretti, Shea, Gambardella, Jenrette, Moro, Echwikhi, Schallert, Gallagher, Block and Chapple. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Francesco Ferretti, ZmVycmV0dGlAdnQuZWR1
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.