 José Carlos Báez
José Carlos Báez Laura Fuentes Tejada
Laura Fuentes Tejada Davinia Torreblanca
Davinia Torreblanca- 1Centro Oceanográfico de Málaga, Instituto Español de Oceanografía (IEO), Consejo Superior de Investigación Científica (CSIC), Fuengirola, Spain
- 2Departament of Biodiversidad, Ecología y Evolución, Universidad Complutense de Madrid, Madrid, Spain
Introduction
The first comprehensive extinction-risk assessment of reptiles conducted by Cox et al. found that at least 1,829 out of 10,196 species are threatened (i.e. 21.1% of reptile species, excluding data-deficient species from the assessment process) (Cox et al., 2022). The study also showed that the main factors that threaten non-marine reptiles are similar to threats faced by other tetrapods: agriculture, logging, urban development, and invasive species. However, marine reptiles, a subset of the reptile group, are threatened by other factors such as fisheries bycatch (Camiñas et al., 2021; Rao et al., 2021), habitat loss (Lane & Guinea, 2010; Lukoschek et al., 2010), and climate change (Patrício et al., 2021; Hochscheid et al., 2022).
Marine reptiles are a non-taxonomic group of reptile species which share similar ecological requirements but have different adaptations regarding their return to the sea (Rasmussen et al., 2011). These adaptations involve physiological changes (e.g. the lachrymal glands of sea turtles), anatomical changes (e.g. the vertically flattened paddle-like tail of sea snakes), and reproductive adaptations (e.g. the viviparity of true sea snakes). Currently, there are 91 species of marine reptiles which form just 0.89% of all extant reptile species (Table 1). These 91 species include the following: 79 sea snake species (including Elapids [67], and aquatic/estuarine snakes Homalopsids [10] and Acrochordids [2]), 9 sea turtle species (in four different families including 7 sea turtles species, and two aquatic/estuarine turtles species), 2 saltwater crocodile species (Crocodylidae), and 1 marine iguana species (Iguanidae). In relation to aquatic/estuarine turtles there are a lot of species potentially to be included as “marine” (Rasmussen et al., 2011), however, we include those estuarine species that have been bycatches in marine fisheries (Taskavak et al., 1999; Eisemberg et al., 2011).
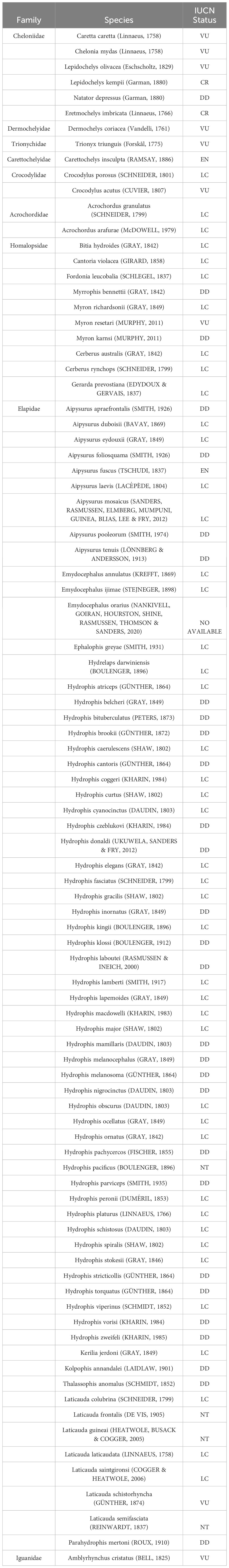
Table 1 Species of marine reptiles and their status according to the Red List of International Union for Conservation of Nature (IUCN).
Marine reptiles occupy a wide variety of marine habitats with very heterogeneous geographical range distributions between groups. For example, marine iguanas (Amblyrhynchus cristatus) and many species of sea snakes have very limited ranges and occupy shallow waters. However, sea turtles, with the exception of the Australian flatback turtles (Natator depressus), have life histories that span vast ocean areas, including feeding areas that are very far from their nesting beaches. Historically, saltwater crocodiles (Crocodylus porosus), had a much wider distribution that included the Seychelle Islands and the coast of China (Webb et al., 2010). Although they occupy mangrove forests and river systems close to the coast, it is possible to see them swimming in open waters.
The aim of this study was to review available information on the population statuses of marine reptiles as well as on the threats and impacts affecting this group. We also discuss possible future measures to prevent the extinction of marine reptiles.
Status of marine reptiles in relation to other vertebrate groups
It is relevant to note that of the 91 species of marine reptiles, 67 are within the group of sea snakes (Elapidae). This group is of uncertain taxonomy and is still under review (Sanders et al., 2013). In addition to the genetic issues such as low variation at all five nuclear markers and conflicting relationships supported by mitochondrial and nuclear trees (Sanders et al., 2013), there are further complications stemming from the limited attention this group receives compared to more charismatic species like sea turtles. This disparity in attention results in inadequate funding and research efforts, leading to gaps in data collection and conservation initiatives (McClenachan et al., 2012; Udyawer et al., 2018).
Among the marine reptiles, 33.0% (including 29 species of sea snakes and the Australian flatback sea turtle, Natator depressus) are considered to be data-deficient such that their extinction-risk assessment cannot be conducted (to see Table 1). This relative percentage is more than the percentage of species for which extinction-risk assessments have not been conducted (reptiles, 14%; amphibians, 20.4%; mammals, 15.1%; birds, 0.5%) (Cox et al., 2022). This could be indicative of the lack of attention that these species have had among specialists in recent decades, as well as of the difficulty of researching in marine habitats.
In relation to their extinction-risk assessment, 14.3% of marine reptile species could be considered threatened of extinction (i.e. the percentage of species relative to the total number of marine reptile species adding the categories Critical Endangered, Endangered, and Vulnerable) (Cox et al., 2022). However, while most sea turtle species, marine crocodiles, and the marine iguana are included in an International Union for Conservation of Nature (IUCN) Red List threatened category, only 3 species out of the 79 sea snakes are included in a threatened category; even though there are 29 species of sea snakes included in the “Data Deficient” category, which could indicate that this group needs more research and monitoring (Elfes et al., 2013).
In the case of sea turtles, many specialists have questioned the suitability of the IUCN Red List for estimating the true conservation status of a species (Godfrey & Godley, 2008; Campbell, 2012). Indeed, it is debatable whether a single conservation category can summarise the multiple fluctuations that different populations of the same species can undergo. For example, marine turtles have wide distribution ranges, and so different responses could be expected from different populations in the face of similar unique threats, such as climate change.
Threatening processes affecting marine reptiles
As discussed, marine reptiles have wide geographical ranges and occupy multiple areas. Nevertheless, they are all affected by the same three threats: bycatch, habitat loss, and climate change.
Bycatch in fisheries is a worrying threat that mainly affects sea turtles (Wallace et al., 2010; Camiñas et al., 2021) and sea snakes (Van Cao et al., 2014; Rao et al., 2021). In the case of sea snakes, fisheries from Asia need better monitoring and tracking to better understand the extent of the problem (Van Cao et al., 2014). Regarding both groups, Ecological Risk Assessments (e.g. see Angel et al., 2014) of the fisheries that may be susceptible to reptile bycatch should be conducted. Courses on how to handle these animals for their healthy and correct release to increase their prospects of post-release survival (e.g. see Parga, 2012) must be implemented. Such courses should also safeguard the physical safety of fishermen, especially when handling sea snakes which have a potentially lethal bite (Van Cao et al., 2014). Moreover, it is a priority to engage both the fishing industry as a whole and the artisanal fishing industry—especially those working in the tropical belt—in marine reptile awareness and conservation programs.
Habitat loss is another important threat that affects favourable areas for feeding and reproduction (Udyawer et al., 2020). It should be noted that most marine reptile species (except for some species of sea snakes) currently require emergent terrain for nesting and laying their eggs. They are therefore particularly vulnerable to the loss of favourable nesting sites. Moreover, aquatic/estuarine species spend their entire lifecycle in these environments, whereas amphibious species that use these habitats only for nesting or resting may be less impacted by coastal threats.
Although there is currently insufficient evidence to be able to assess the short- and medium-term threats of climate change to reptiles (Cox et al., 2022), marine reptiles can be impacted by climate change in two ways: (a) by limiting suitable habitats and the resources provided by these habitats (as in the case of true sea snakes, sea turtles, and marine iguanas); and (b) by causing a bias in sex ratios of populations due to alterations in ecological sex determination (as in the case of sea turtles and saltwater crocodiles). Concerning the former threat, climate-change-driven temperature rises affect coral reefs, which are inhabited by true marine snakes (Lane & Guinea, 2010). Moreover, increasing sea levels and severe storms due to climate change could reduce the number of available nesting beaches (Torres et al., 2021) used by sea turtles and some species of oviparous sea snakes for egg laying (e.g. Lane and Guinea, 2010; Maneja et al., 2021). Regarding iguanas, rising sea surface temperatures driven by El Niño events—which are increasing in the context of climate change (Cai et al., 2014)—disrupt the supply of seaweed. Since seaweed is the main food source for iguanas, its loss can cause up to 90% of the population to die from starvation during these events (MacLeod et al., 2020). Concerning the latter threat, sea turtles and saltwater crocodiles have temperature-dependent sex determination (Woodward & Dickson, 1993; Tezak et al., 2020; Patrício et al., 2021), and therefore climate-change-driven temperature rises could affect sex bias within these populations. Hochscheid et al. found that the nesting range of loggerhead turtles had expanded in the Mediterranean despite their philopatry, which has been interpreted as a response to climate change. Predictive models could help identify new potential nesting areas and implement proactive conservation actions to establish new rookeries (Hochscheid et al., 2022). Regarding sea snakes, predictive models could help in preventing encounters with the human population (Needleman et al., 2018).
In addition, the marine reptiles as ectothermic air-breathing marine animals, increases in water temperatures will increase metabolic demand, and subsequently reduce their diving capacities (Rodgers et al., 2021). This has large implications on foraging efficiencies, diving capabilities and survival of this whole group, depending on the species ability to thermally acclimate to changing temperatures (Rodgers et al., 2021).
Discussion
Other important threats also exist such as contamination, emerging diseases, and hybridization between species. However, these are common for all marine reptile species, and we therefore do not know the degree of prevalence per species. Many conservation policies described here cannot be considered specific to marine reptiles. However, in the framework of marine megafauna, marine reptiles are especially affected by the same three threats described above (i.e. bycatch, habitat loss, and climate change). Thus, new tools are needed to delimit populations and improve knowledge on their fluctuations and trends. Wallace et al. proposed the use of Regional Marine Turtle Management Units (RMUs) as a framework for prioritising conservation of sea turtles across multiple scales (Wallace et al., 2010). In this regard, Regional Tuna Fisheries Organisations have invested great efforts in assessing stocks of different turtle RMUs (Camiñas et al., 2021). To prevent habitat loss, it may be of interest to first identify favourable areas for marine reptiles (Udyawer et al., 2020). A second step would be to identify Important Marine Reptile Areas in the same way that Important Marine Mammal Areas (IMMAs) have been identified (Tetley et al., 2022), while including the nesting and foraging zones of different populations.
It is worth noting that marine mammals and highly migratory species are primarily governed by the United Nations Convention on the Law of the Sea (UNCLOS). UNCLOS serves as the primary legal instrument governing activities in the world’s oceans by setting out general principles and obligations aimed at ensuring the sustainable use and conservation of marine resources. Article 65 of the treaty specifically addresses the conservation and management of highly migratory species, emphasizing the need for states to cooperate both within and beyond national jurisdiction. In addition to highly migratory species, UNCLOS also imposes obligations on states to protect and preserve marine mammals. Moreover, UNCLOS establishes mechanisms for international cooperation and dispute resolution to address issues concerning the conservation and management of marine mammals and highly migratory species. This includes provisions for regional fisheries management organizations (RFMOs) and other relevant bodies to coordinate conservation measures and regulate activities impacting these species across different maritime zones.
However, marine reptiles are not currently included in UNCLOS. Including marine reptiles in UNCLOS could help create specific international policies which would increase the effectiveness of conservation efforts for this group. Other international treaties such as the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES), include all non-snake marine reptiles in their annexes. In fact, CITES does not have any species of sea snake in their appendices (Davies, 2023). CITES plays a crucial role in regulating the international trade of endangered species, regulates international trade to prevent over-exploitation, and fosters global cooperation to avoid illegal exploitation.
On the other hand, for these taxa country level policies are likely to be important (e.g., such as the Endangered Species Act provided by the United States Environmental Protection Agency, or the legislation Australian Environment Protection and Biodiversity Conservation).
Finally, it should be a priority to integrate fishermen, especially from the tropical belt, in awareness and conservation programs for marine reptiles which are an ancient component of marine biodiversity. As a threatened and distinct group, marine reptiles require specific management to ensure their correct release and to increase their survival post-release. Because of this, management guides could help when working with sea snakes. There already exists a multitude of these guides for sea turtles that could be used as examples when creating them for sea snakes.
Author contributions
JB: Conceptualization, Funding acquisition, Writing – original draft. LF: Data curation, Investigation, Resources, Writing – review & editing. DT: Conceptualization, Data curation, Investigation, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study has been funded by the Spanish Herpetological Association (AHE).
Acknowledgments
We are grateful the suggestions and recommendations provided by the reviewer.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Angel A., Nel R., Wanless R. M., Mellet B., Harris L., Wilson I. (2014). Ecological Risk Assessment of sea turtles to tuna fishing in the ICCAT region. Collect Vol. Sci. Pap. ICCAT 70, 2226–2259.
Cai W., Borlace S., Lengaigne M., Van Rensch P., Collins M., Vecchi G., et al. (2014). Increasing frequency of extreme El Niño events due to greenhouse warming. Nat. Clim. Change 4, 111–116. doi: 10.1038/nclimate2100
Camiñas J. A., Domingo A., Coelho R., Bruyn P., Abascal F. J., Báez J. C. (2021). Tuna regional fisheries management organizations and the conservation of sea turtles: A reply to Godley et al. Oryx 55, 12. doi: 10.1017/S0030605320000708
Campbell L. M. (2012). Seeing red: inside the science and politics of the IUCN red list. Conserv. Soc 10, 367–380. doi: 10.4103/0972-4923.105560
Cox N., Young B. E., Bowles P., Fernandez M., Marin J., Rapacciuolo G., et al. (2022). A global reptile assessment highlights shared conservation needs of tetrapods. Nature 605, 285–290. doi: 10.1038/s41586-022-04664-7
Davies E. (2023). A deep dive into sea snakes, sea kraits and their aquatic adaptations (London: The Trustees of The Natural History Museum). Available at: https://www.nhm.ac.uk/discover/sea-snakes-sea-kraits-and-their-aquatic-adaptations.html.
Eisemberg C. C., Rose M., Yaru B., Georges A. (2011). Demonstrating decline of an iconic species under sustained indigenous harvest – The pig-nosed turtle (Carettochelys insculpta) in Papua New Guinea. Biol. Conserv. 144, 2282–2288. doi: 10.1016/j.biocon.2011.06.005
Elfes C. T., Livingstone S. R., Lane A., Lukosche V., Sanders K. L., Courtney A. J., et al. (2013). Fascinating and forgotten: The conservation status of marine elapid snakes. Herpetol. Conserv. Biol. 8, 37–52.
Godfrey M. H., Godley B. J. (2008). Seeing past the red: flawed IUCN global listings for sea turtles. Endanger Species Res. 6, 155–159. doi: 10.3354/esr00071
Hochscheid S., Maffucci F., Abella E., Bradai M. N., Camedda A., Carreras C., et al. (2022). Nesting range expansion of loggerhead turtles in the Mediterranean: phenology, spatial distribution, and conservation implications. Glob. Ecol. Conserv. 38, e02194. doi: 10.1016/j.gecco.2022.e02194
Lane A., Guinea M. (2010). Laticauda schistorhynchus. The IUCN Red List of Threatened Species 2010. doi: 10.2305/IUCN.UK.2010–4.RLTS.T176740A7294814.en
Lukoschek V., Guinea M., Rasmussen A. R. (2010). Aipysurus fuscus. The IUCN Red List of Threatened Species 2010. doi: 10.2305/IUCN.UK.2010–4.RLTS.T176763A7299535.en
MacLeod A., Nelson K. N., Grant T. D. (2020). Amblyrhynchus cristatus The IUCN Red ListofThreatenedSpecies2020. doi: 10.2305/IUCN.UK.2020–2.RLTS.T1086A177552193.en
Maneja R., Miller J. D., Li W., Rejoice T., El-Askary H., Perera S., et al. (2021). Multidecadal analysis of beach loss at the major offshore sea turtle nesting islands in the Northern Arabian gulf. Ecol. Indic. 121, 107146. doi: 10.1016/j.ecolind.2020.107146
McClenachan L., Cooper A. B., Carpenter K. E., Dulvy N. K. (2012). Extinction risk and bottlenecks in the conservation of charismatic marine species. Conserv. Lett. 5, 73–80. doi: 10.1111/j.1755-263X.2011.00206.x
Needleman R. K., Neylan I. P., Erickson T. B. (2018). Environmental and ecological effects of climate change on venomous marine and amphibious species in the wilderness. Wilderness Environ. Med. 29, 343–356. doi: 10.1016/j.wem.2018.04.003
Parga M. L. (2012). Hooks and sea turtles: a veterinarian’s perspective. Bull. Mar. Sci. 88, 731–741. doi: 10.5343/bms.2011.1063
Patrício A. R., Hawkes L. A., Monsinjon J. R., Godley B. J., Fuentes M. M. P. B. (2021). Climate change and marine turtles: recent advances and future directions. Endanger Species Res. 44, 363–395. doi: 10.3354/esr01110
Rao C., Dsouza S., Gupta T., Manoharakrishnan M., Lobo A. S. (2021). Fisheries induced shift in sea snake community assemblages along the Konkan coast, India. Aquat. Conserv. 31, 2402–2411. doi: 10.1002/aqc.3658
Rasmussen A. R., Murphy J. C., Ompi M., Gibbons J. W., Uetz P. (2011). Marine reptiles. PloS One 6, e27373. doi: 10.1371/journal.pone.0027373
Rodgers M. E., Franklin C. E., Noble D. W. A. (2021). Diving in hot water: a meta-analytic review of how diving vertebrate ectotherms will fare in a warmer world. J. Exp. Biol. 224, jeb228213. doi: 10.1242/jeb.228213
Sanders K. L., Lee M. S. Y., Mumpuni M., Bertozzi T., Rasmussen A. R. (2013). Multilocus phylogeny and recent rapid radiation of the viviparous sea snakes (Elapidae: Hydrophiinae). Mol. Phylogenet. Evol. 66, 575–591. doi: 10.1016/j.ympev.2012.09.021
Taskavak E., Reimann M. J., Polder W. N. (1999). First Record of the Nile Soft-Shelled Turtle, Trionyx triunguis, from Kos Island, Greece, with Comments on its Occurrence in the Eastern Mediterranean. Chelonian Conserv. Biol. 3, 510–512.
Tetley M. J., Braulik G. T., Lanfredi C., Minton G., Panigada S., Politi E., et al. (2022). The important marine mammal area network: a tool for systematic spatial planning in response to the marine mammal habitat conservation crisis. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.841789
Tezak B., Sifuentes-Romero I., Milton S., Wyneken J. (2020). Identifying sex of neonate turtles with temperature-dependent sex determination via small blood samples. Sci. Rep. 10, 5012. doi: 10.1038/s41598-020-61984-2
Torres A., Simoni M. U., Keiding J. K., Müller D. B., Ermgassen S., Liu J., et al. (2021). Sustainability of the global sand system in the Anthropocene. OneEarth 4, 639–650. doi: 10.1016/j.oneear.2021.04.011
Udyawer V., Barnes P., Bonnet X., Brischoux F., Crowe-Riddell J. M., D’Anastasi B., et al. (2018). Future directions in the research and management of marine snakes. Front. Mar. Sci. 5. doi: 10.3389/fmars.2018.00399
Udyawer V., Somaweera R., Nitschke C., d’Anastasi B., Sanders K., Webber B. L., et al. (2020). Prioritising search effort to locate previously unknown populations of endangered marine reptiles. Glob. Ecol. Conserv. 22, e01013. doi: 10.1016/j.gecco.2020.e01013
Van Cao N., Thien Tao N., Moore A., Montoya A., Rasmussen A. R., Broad K., et al. (2014). Sea snake harvest in the gulf of Thailand. Conserv. Biol. 28, 1677–1687. doi: 10.1111/cobi.12387
Wallace B. P., Lewison R. L., McDonald S. L., McDonald R. K., Kot C. Y., Kelez S., et al. (2010). Global patterns of marine turtle bycatch. Conserv. Lett. 3, 369–369. doi: 10.1111/j.1755-263X.2010.00105.x
Webb G. J. W., Manolis S. C., Brien M. L. (2010). “Saltwater Crocodile Crocodylus porosus,” in Crocodiles. Status Survey and Conservation Action Plan, 3rd ed. Eds. Manolis S. C., Stevenson C. (Gland, Switzerland: IUCN), 99–113.
Keywords: snake, turtle, crocodile, marine reptiles, conservation
Citation: Báez JC, Fuentes Tejada L and Torreblanca D (2024) Specific marine policies are needed to prevent the extinction of marine reptiles. Front. Mar. Sci. 11:1416178. doi: 10.3389/fmars.2024.1416178
Received: 11 April 2024; Accepted: 05 June 2024;
Published: 17 June 2024.
Edited by:
Salvatore Siciliano, Fundação Oswaldo Cruz (Fiocruz), BrazilReviewed by:
Vinay Udyawer, Australian Institute of Marine Science (AIMS), AustraliaCopyright © 2024 Báez, Fuentes Tejada and Torreblanca. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José Carlos Báez, am9zZWNhcmxvcy5iYWV6QGllby5jc2ljLmVz
†ORCID: José Carlos Báez, orcid.org//0000-0003-2049-0409