
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Mar. Sci. , 28 May 2024
Sec. Marine Megafauna
Volume 11 - 2024 | https://doi.org/10.3389/fmars.2024.1407575
 Michelle María Early-Capistrán1*
Michelle María Early-Capistrán1* Nicole L. Crane2Larry B. Crowder1Gerardo Garibay-Melo3
Nicole L. Crane2Larry B. Crowder1Gerardo Garibay-Melo3 Jeffrey A. Seminoff4David Johnston5
Jeffrey A. Seminoff4David Johnston5The accelerated pace of climate-driven shifts is surpassing the temporal scope of conventional field research, potentially leading to a disconnect between ecosystem changes and scientific data collection. Climate change is producing rapid transformations within dynamic marine ecosystems, with a pronounced effect on high trophic-level species such as loggerhead sea turtles (Caretta caretta). We present a new model for data collection using ethnobiological methods, emphasizing how local community members can contribute to expanding scientific knowledge via context-informed observations, to document species occurrences beyond their anticipated habitats during climatic anomalies. In rapidly changing conditions, local expert knowledge can complement conventional scientific methods, providing high-quality data with extensive coverage—especially for elusive species—and yielding insight into potential emerging phenomena that may otherwise go unnoticed. Conventional methods for predicting distribution shifts in rare species are vulnerable to spatial biases, favoring predictions based on the most probable habitats. We present the case study of a live sea turtle sighting by a local expert in Monterey Bay, California, USA, identified post hoc as a loggerhead, to illustrate methods which can be transferred and applied to other rare and highly migratory marine species such as marine mammals, sharks, and seabirds. This emerging framework incorporates diverse knowledge sources and methodologies in monitoring climate-driven ecological shifts, enriching conservation strategies, enhancing our understanding of complex ecosystems, and contributing to robust evidentiary standards for rare species observations.
Climate change is leading to rapid and unprecedented biological shifts, particularly in dynamic marine environments (Beaugrand et al., 2019). Climate change is rapidly shifting species distributions, and may cause ecosystems and abiotic drivers to vary more rapidly than time-frames required for scientific field research, which often range from years to decades (Poloczanska et al., 2013; Molinos et al., 2015; Taheri et al., 2021). These difficulties are amplified for highly-migratory endangered species, which (i) face a heightened risk of extinction due to climate change, and (ii) are challenging to sample and model, especially outside established habitats, due to their rarity (Jeliazkov et al., 2022). Observations by community members and the general public can be valuable for locating sentinel species in unexpected locations: while scientists may be in the field intermittently, ocean users such as fishers, surfers, kayakers, inter alia are consistently observing the ocean with direct exposure and repeat observations (Jones et al., 2018; Hanna et al., 2021; Tengö et al., 2021). We use the term “community-member reports” to define reports made to scientists by community members outside structured programs, in contrast with community or citizen science, in which community members gather data for scientific research through outreach or monitoring programs, or digital platforms (e.g., eBird, iNaturalist, etc.) (Gray et al., 2017). Integrating community science and community-member reports into scientific research is critical, especially for highly migratory and rare species (Jambura et al., 2021; Fontaine et al., 2022). We posit that transdisciplinary research—integrating rigorous methods from ecology and ethnobiology with non-academic stakeholders—can yield important insights without compromising the integrity of the scientific process or the results, and is needed to more effectively document changes in species distribution.
In this context, we present a community-member report, corroborated using established methods from ethnobiology, that documents the first sighting of a live loggerhead sea turtle (Caretta caretta, henceforth “loggerhead”) in Monterey Bay, California, U.S.A., ~1,500 km north of the nearest resident population. North Pacific loggerhead sea turtles nest exclusively in Japan, and undertake basin-wide developmental migrations to forage in highly productive pelagic zones in the Central North Pacific and northwest Mexico, with the epicenter of distribution in the Eastern Pacific in the Gulf of Ulloa along the Baja California peninsula, Mexico (Polovina et al., 2017; Okuyama et al., 2022; Figure 1). Upon arrival to foraging habitats, they establish long-term (~20 year) residency (Turner Tomaszewicz et al., 2017). Loggerheads are listed globally as Vulnerable by the International Union for the Conservation of Nature (IUCN) Red List of Threatened Species (Casale and Tucker, 2017), and the North Pacific Distinct Population Segment is listed as Endangered under the U.S. Endangered Species Act (National Marine Fisheries Service and U.S. Fish and Wildlife Service, 2020).
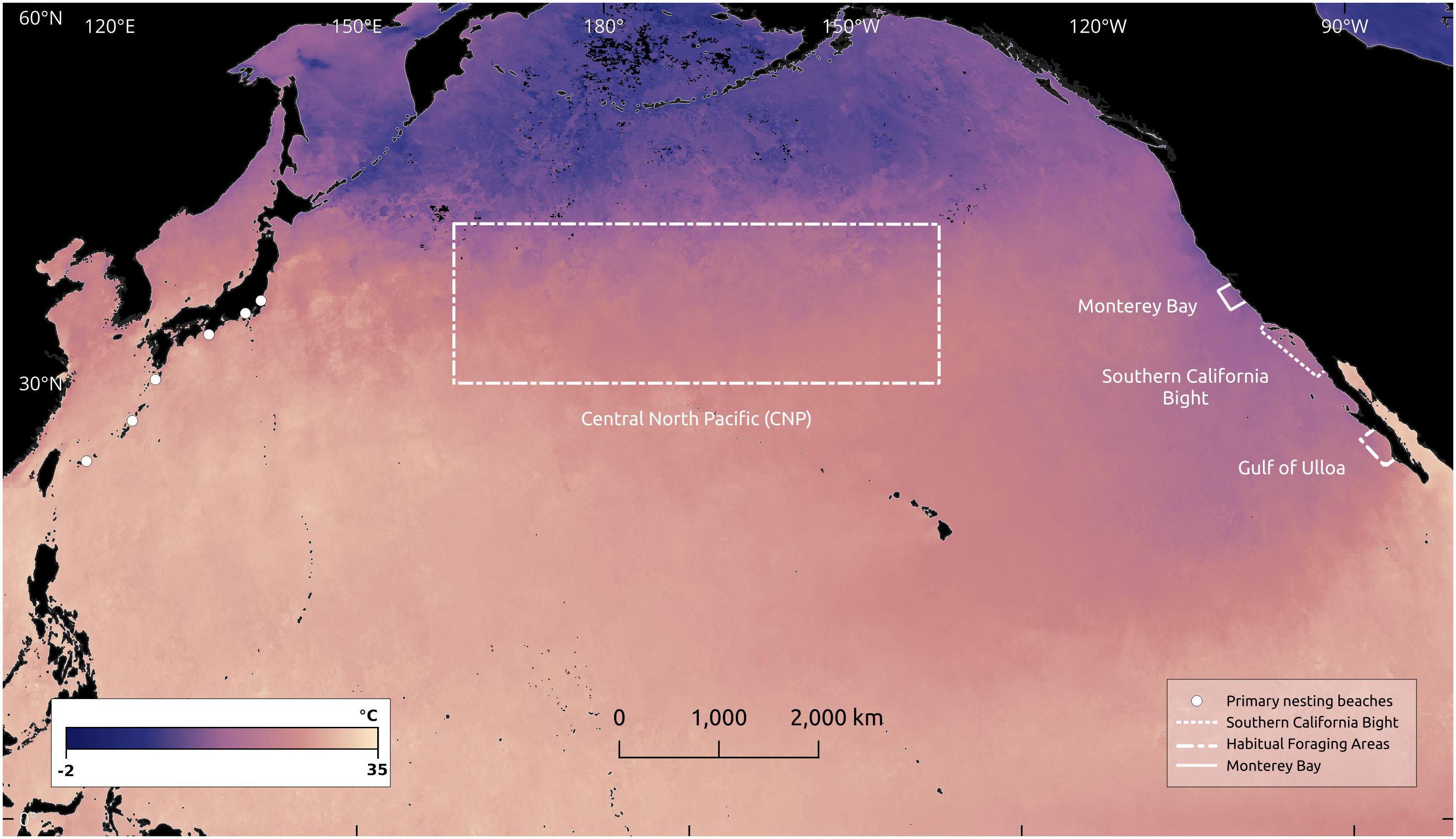
Figure 1 Locations of loggerhead sea turtle (Caretta caretta) primary nesting beaches (white dots) and foraging habitats (dashed-dot line), along with Monterey Bay (solid line) and the Southern California Bight (dotted line). Sea Surface Temperature (SST) for July, 2023 is shown at 0.1° resolution. Imagery processed by the NASA Earth Observations (NEO) team in collaboration with Gene Feldman and Norman Kuring, NASA OceanColor Group (NASA Earth Observatory, 2023).
High trophic-level marine species can be important “climate sentinels”, defined as species which provide insights into unobserved components of the ecosystem and respond specifically to climate variability or change (Sydeman et al., 2015; Hazen et al., 2019). Loggerheads’ conspicuousness, sensitivity to ecosystem processes, and high trophic level make them valuable climate sentinels (Hazen et al., 2019; Mallard and Couderchet, 2019). As ectotherms, loggerheads rely on thermal cues for habitat selection and avoidance, and temperature is a critical driver of their distribution and migratory patterns (Abecassis et al., 2013; Briscoe et al., 2016). Importantly, Pacific loggerheads exhibit strong fidelity to the 18°C isotherm, a widely-used proxy for the highly-productive Transition Zone Chlorophyll Front (Polovina et al., 2017; Briscoe et al., 2021; Figure 1). Thermal cues and patterns are closely linked to ocean temperature, which is dynamic and operates on various temporal scales including annual variations, irregular events like the El Niño Southern Oscillation (ENSO). These oceanic temperature dynamics are themselves subject to alterations driven by changing climatic conditions (Patrício et al., 2011; Almpanidou et al., 2019; Webb and Magi, 2022). Importantly, ENSO dynamics have important impacts on loggerhead spatial dynamics, and may drive the periodic occurrence of large aggregations (>15,000) of loggerheads in the southern California Bight during El Niño events (Eguchi et al., 2018).
We recognize that a single sighting, at present, is insufficient to establish a range expansion. However, this finding demonstrates the pivotal role that transdisciplinary approaches and community-engaged science can play in documenting rare species occurrences beyond their anticipated selected habitats, and the need for continued observation to establish if unexpected sightings are singular events or linked to broader trends. Thus, we aim to (i) provide rigorous, transferable methods that can be applied to other rare and highly migratory marine species, such as sharks, marine mammals, or seabirds; and (ii) highlight the need for community-engaged research to facilitate rare species documentation outside predicted ranges in the fast-changing context of climate change. Likewise, we emphasize the need for expanding our reporting criteria and methods to encompass platforms for community-member reports and implementing rigorous methods for evaluating reports and identifying species when photographic or video evidence is lacking.
Hard-shelled turtles are primarily tropical and sub-tropical. Low sea surface temperatures (SST) in Monterey Bay, which oscillate from ~10–15°C (NOAA, 2023c) make the location a suboptimal habitat with a high risk of cold-stunning, a hypothermic reaction which causes debilitation and lethargy, often leading to death (Innis et al., 2007; Briscoe et al., 2016; Griffin et al., 2019). To date, only two hard-shelled sea turtle sightings have been recorded in Monterey Bay. These include one live green turtle (Chelonia mydas) in 2017 (NOAA, unpublished data) and a live, stranded olive ridley (Lepidochelys olivacea) in 2011 which was rescued and rehabilitated (Monterey Bay Aquarium, 2011). A live olive ridley was also recorded in August, 2023, after the sighting which is the subject of this paper (The Marine Mammal Center, 2023). Crucially, the sea turtle occurrences 2017 and August 2023 were registered through community-member reports.
Moss Landing, California, U.S.A. (36.804444°, -121.786944°) is located in Monterey Bay within the Monterey Bay Marine National Marine Sanctuary, a marine vertebrate biodiversity hotspot characterized by seasonal upwelling and high productivity (Chavez and Messié, 2009; Paduan et al., 2018; NOAA, 2023a). Specifically, Moss Landing is located at the head of the Monterey Submarine Canyon, one of the largest submarine canyons on the Pacific coast of the United States, with maximum depths >2300m (Paull et al., 2011). It is a critical habitat for migratory shorebirds and marine mammals such as sea otters (Enhydra lutris), harbor seals (Phoca vitulina), and humpback whales (Megaptera novaeangliae) (Palumbi and Sotka, 2012).
The presumptive loggerhead sea turtle sighting was reported by senior author D. Johnston, an expert naturalist and professional kayak guide with 40 years of experience and an estimated 16,000 hours of observation in Monterey Bay. Mr. Johnston spends over 50 days per year on the water, leading wildlife kayaking tours and educating recreational kayakers about the marine and coastal fauna and environment Monterey Bay Marine National Marine Sanctuary. We employed participatory mapping, which uses collaborative mapping exercises to capture and co-produce spatial representations of LEK regarding the sightings and related environmental conditions (Gray et al., 2017; Wedemeyer-Strombel et al., 2019), to understand the biophysical conditions related to the sighting. Interviews were carried out following ethical protocols of the International Society of Ethnobiology (2008). The interview and participatory mapping exercise were recorded on audio and video with D. Johnston’s informed consent, and transcribed.
M.M. Early-Capistrán conducted a brief (~10 minute) telephone interview on the day of the sighting to gather preliminary information and develop a guide for the participatory mapping exercise, with primary topic areas to be discussed while allowing for emerging topics to be followed as they emerged during the mapping process (Supplementary Material 1). D. Johnston, M.M. Early-Capistrán and G. Garibay-Melo then conducted a mapping exercise in Google Earth Pro 7.3.6. We first oriented ourselves on the map by pointing out key landmarks. We discussed the activities and events on the day of the sighting, and asked D. Johnston to label on the map where the sightings occurred (Albuquerque et al., 2014; Wedemeyer-Strombel et al., 2019). D. Johnston identified locations through his knowledge of the local bathymetry and coastline. We transferred coordinates to GeoMapApp 3.7.1 (Lamont-Doherty Earth Observatory, 2023) to access Global Multi-Resolution Topography synthesis (Ryan et al., 2009) and high-resolution (10m) bathymetric data of Monterey Canyon (Paull and Caress, 2019). We included 500-m bounding boxes around each sighting location to account for uncertainty. Additionally, we used NASA monthly sea surface temperature (SST) products for July, 2023 collected with Moderate Resolution Imaging Spectroradiometer (MODIS) instruments at 0.1-° resolution based on MODIS-calibrated mid- and far-infrared (IR) radiances (Bands 20, 22, 23, 31, and 32 from MOD02) to map the sighting location in relation to frequently used foraging habitats (NASA Earth Observatory, 2023). All maps were generated with QGIS 3.32.
We followed established ethnobiological techniques for species identification via visual stimuli (Albuquerque et al., 2014), which uses images, photographs or films as an auxiliary strategy to recall information of interest or to contextually orient the interviewee by discussing specific details of the species of interest (Albuquerque et al., 2008; Pozzi and Ladio, 2023). We used two sets of stimuli images along with a semi-structured interview guide (Supplementary Material 2) and prompts to ascertain a species ID. The interview guide provided a set of topics while allowing flexibility to follow leads as the interview progressed (Bernard, 2011).
First, we viewed three published sea turtle identification guides, one of which was designed for community scientists observing sea turtles from above-water (e.g., cruise ships, tour boats, and other vessels) (Pritchard and Mortimer, 1999; Secretariat of the Pacific Community, 2003; Upwell and Grupo Tortuguero de las Californias, 2023), and discussed (i) which species had morphological characteristics most similar to the observed turtle (e.g., which characteristics were similar or different) and (ii) the habitat preferences and natural history of each species (e.g., where each species is habitually found, dietary preferences, migratory patterns, etc.) in comparison to the environmental conditions and behavioral observations of the sighting.
We reviewed composites of photos of the three potential species (L. olivacea, C. mydas, and C. caretta) (Figure 2). Leatherback sea turtles (Dermochelys coriacea), which forage in Monterey Bay, were ruled out due to their distinctive morphology (Benson et al., 2020). Hawksbill sea turtles (Eretmochelys imbricata) were also ruled out due to their restricted range (e.g., tropical coral reefs and mangroves) and restricted movements (Martínez-Estévez et al., 2022). Each composite image contained photographs of a single species from multiple angles, with clearly visible distinctive features to aid in their characterization (Pozzi and Ladio, 2023). Species names were not provided during the discussion to avoid biasing responses. We discussed each composite image in-depth, comparing the features in the image with the features of the observed turtle such as the size and number of scutes, facial scaling, pigmentation, cranial morphology, and the size and proportions of the axial body and appendages.

Figure 2 Visual stimuli. Images of three hard-shelled turtle species found in the temperate and sub-tropical northeast Pacific. Column (A) loggerhead sea turtle (Caretta caretta); Column (C) olive Ridley (Lepidochelys olivacea); Column (C) green turtle (Chelonia mydas). Images of each species were reviewed separately during the interviews. All images from Wikimedia Commons - CC-BY.
D. Johnston observed the turtle from a kayak on July 9, 2023 at approximately 10:30 AM near the drop-off of the Monterey Submarine Canyon. Sea surface temperature was 12.5°C, with an ambient temperature of 15°C (NOAA, 2023b). Loggerheads in the North Pacific have reported thermal tolerance ranging from 10–27.84°C, with reported likelihood of cold-stunning at <8°C (Briscoe et al., 2016; Saito et al., 2018). In the North Pacific, active loggerheads have been observed at 10.5–10.8°C (Halpin et al., 2018). Local SST at the time of the sighting was within the low range of reported thermal tolerance levels (12.5°C).
During the first sighting (approximately 36.808711°, -121.805436°), the turtle came up to breathe and then swam southward. The second sighting (approximately 36.803556°, -121.804919°) occurred approximately five minutes later when the turtle surfaced to breathe again (Figure 3). The turtle was active and did not exhibit visible signs of cold-stunning (e.g., debilitation, lethargy) (Griffin et al., 2019). Both sightings occurred in a pelagic location with possible topographic upwelling and potential foraging species, which correspond with suitable foraging conditions for loggerheads (Figure 3; Table 1). “It was straight off of Moss Landing […] where the submarine canyon initiates about a mile from shore […]. There could have been some upwelling right there.” Invertebrates observed at the sighting locations include likely prey species, “a lot of pieces of jellyfish around […] Purple striped jellyfish, brown sea nettles, moon jellies”. The turtle was not observed eating, but the presence of pieces of jellyfish suggest possible foraging activity (Table 1).
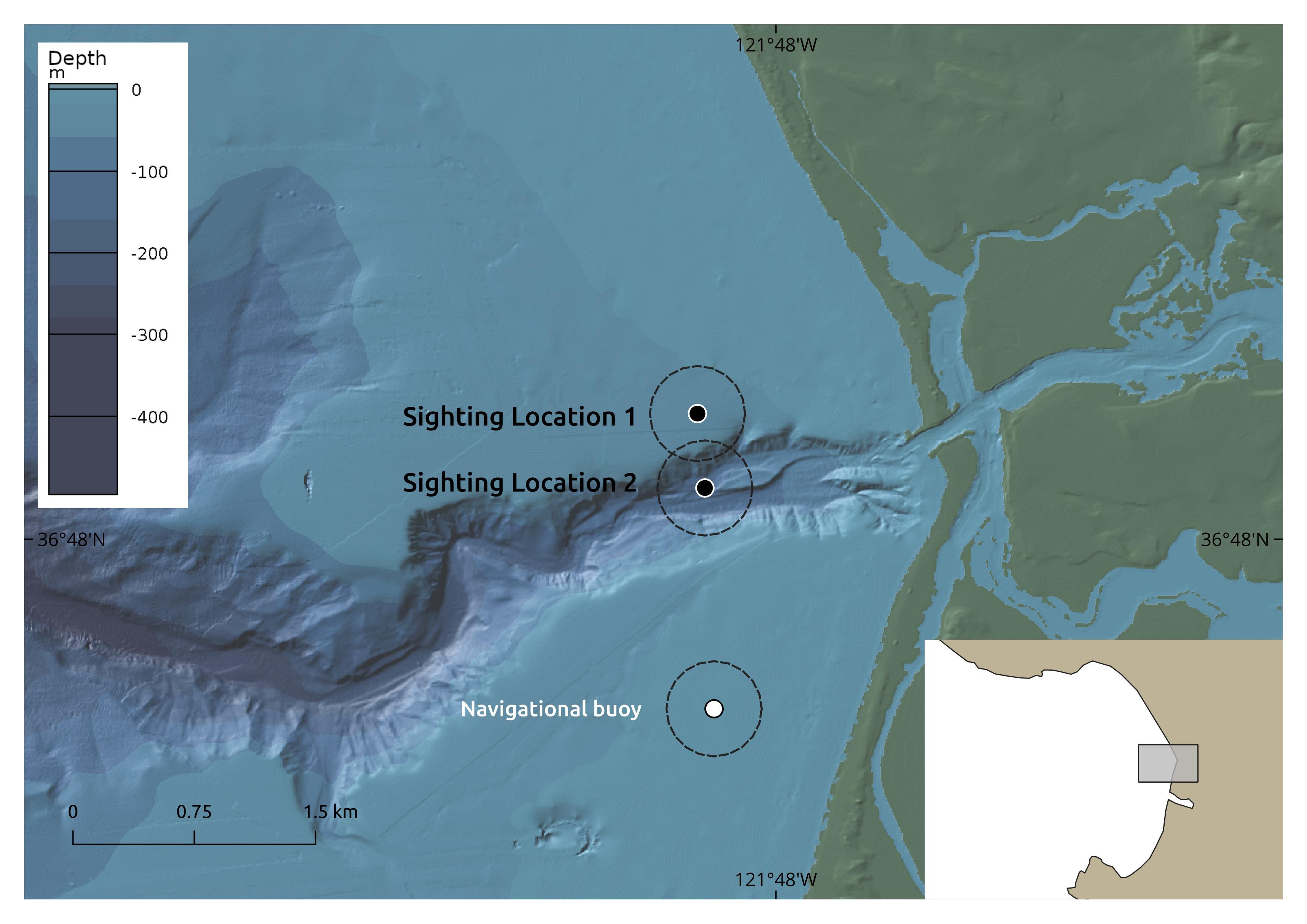
Figure 3 The locations of loggerhead (Caretta caretta) sighting reported on July 9, 2023 in Monterey Bay, along with the location of a navigational buoy used as a spatial reference and high-resolution (10m) bathymetry of Monterey Canyon (Paull and Caress, 2019). Dashed lines show 500m bounding for uncertainty. Map created with GeoMapApp - CC-BY.
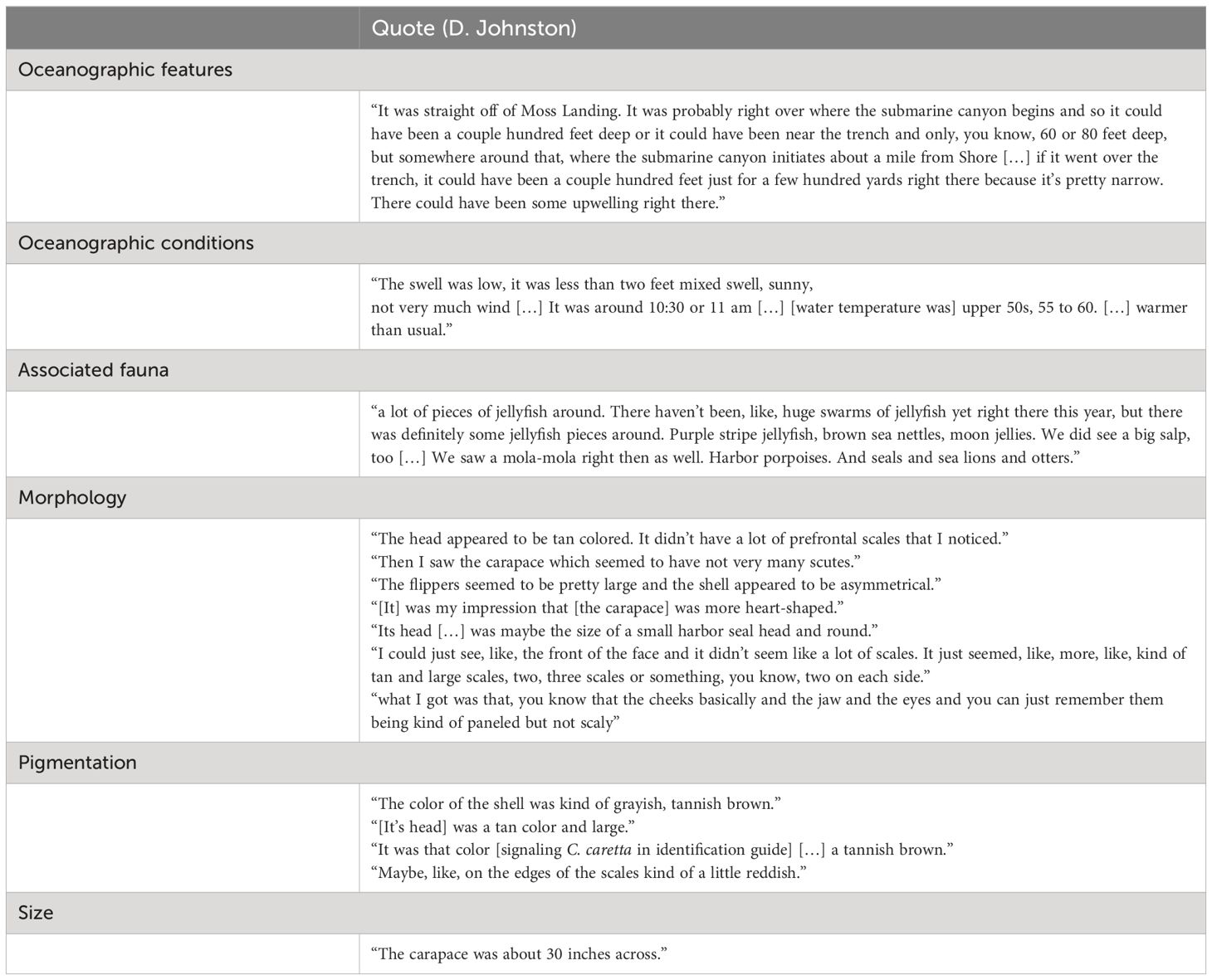
Table 1 Key observations of putative loggerhead sea turtle sightings from Local Ecological Knowledge interview.
Morphological characteristics are consistent with loggerheads. During the initial phone interview, Mr. Johnston described the carapace as “not circular, kind of heart-shaped […]”, and referred to “a big head with a tan-colored face” and “tan, light brownish-grey” coloration. In relation to visual stimuli, Mr. Johnston immediately ruled out green turtles (Chelonia mydas) which he had encountered previously while snorkeling in Hawai’i. While viewing published ID guides, he immediately selected C. Caretta based on coloration (“It was that color [signaling C. caretta in identification guide] […] a tannish brown”), and reiterated the description of the carapace shape (“the shell appeared to be asymmetrical […] [It] was my impression that [the carapace] was more heart-shaped”). Descriptions of pigmentation in relation to visual stimuli highlighted distinctive coloration of loggerheads which ranges from yellow-orange to reddish brown or dark brown (“The head appeared to be tan colored”, “The color of the shell was kind of greyish, tannish brown”, “on the edges of the scales kind of a little reddish”; Table 1). Importantly, “its head […] was maybe the size of a small harbor seal head and round”. A proportionally large head is a distinctive characteristic of C. caretta (Pritchard and Mortimer, 1999). The limited number of prefrontal scales (“the front of the face and it didn’t seem like a lot of scale […] kind of tan and large scales”), tympanic scales (“the cheeks […] [were] kind of paneled but not scaly”), and scutes (“I saw the carapace which seemed to have not very many scutes.”) are also consistent with loggerheads (Pritchard and Mortimer, 1999; Lee et al., 2014) (Table 1). Thus, we concluded that this individual was the 1st confirmed C. caretta for Monterey Bay.
This sighting highlights the value of ethnobiological methods to document community-member reports and generate rigorous post-hoc evaluations of rare species occurrence outside anticipated habitats. Furthermore, such rigor can contribute to developing robust evidentiary standards for rare species observations (McKelvey et al., 2008), shifting from “anecdotes” to reliable data (Early-Capistrán et al., 2020).Non-traditional data sources such as community-member reports, Local Ecological Knowledge (LEK), and community or citizen science can complement conventional science methods in fast-changing systems by substantially increasing the reporting of important data (Chandler et al., 2017; Eguchi et al., 2018; Hanna et al., 2021). Rigorously documented observations by members of the public—especially local experts like naturalists, tour operators, guide outfitters, hunters and fishers, indigenous or local knowledge keepers, etc.—can generate high-quality data with broad coverage for rare or elusive species (Beaudreau and Levin, 2014; McKinley et al., 2017; Aswani et al., 2018).
We recognize that community-member reports and experience-based knowledge are epistemologically distinct from data gathered under experimental conditions. However, these differences are more often reflections of epistemological differences or methods of collection than inherent unreliability (Brook and McLachlan, 2005). Researchers can elicit and corroborate qualitative data derived from empirically-lived situations using established and robust methods from fields such as ethnobiology and environmental anthropology to gather and catalog data, as well as assure data quality, clarity, validity, and reliability. By synthesizing this data, submitting it to rigorous analysis and validation; and explicitly recognizing the associated limitations and uncertainty, it can reliably be integrated into scientific research (Palmer and Wadley, 2007; Early-Capistrán et al., 2020). Specifically, the methods of participatory mapping and identification through visual stimuli can provide spatially-explicit data and species identification for rare species sightings. Future applications of these methods can benefit, when possible, from establishing consensus and validation across multiple observers (e.g., Early-Capistrán et al., 2020). Such approaches can contribute substantially to expanding data availability for rare species, climate sentinels, and marine species undergoing rapid climate-driven shifts.
Loggerheads’ close connection to temperature variability makes them valuable climate sentinels, either as indicators of temperature anomalies or expanding species range as a result of climate change (Hazen et al., 2013, 2019). The close association between loggerheads and SST changes has led to policies based on dynamic-fisheries management to prevent increased loggerhead bycatch in the California Current Large Marine Ecosystem during anomalous warm-water periods thorough a time-area closure enacted when an El Niño event is forecasted or declared (NOAA, 2001; Eguchi et al., 2018; Briscoe et al., 2021). ENSO dynamics—particularly El Niño events—may drive periodic loggerhead presence in the Southern California Bight. Importantly, 94% of bycatch in the region have occurred during El Niño years, and high density aggregations of loggerheads (>15,000 individuals) were observed in the Southern California Bight in 2015, when anomalous warming of the North Pacific co-occurred with El Niño (Eguchi et al., 2018). Importantly, the sightings occurred in the first months of an El Niño event (NOAA, 2024a): the June-August 2023 Oceanic Niño Index (ONI)—a 3-month running mean of SST anomalies in the Niño 3.4 region—was 1.1°C above the long-term average (Becker, 2023; NOAA, 2024b). The connection between loggerhead presence in California waters underscores the need for regular surveys throughout their foraging areas along the west coast of North America (Eguchi et al., 2018). While a single sighting does not yet establish Monterey Bay as a loggerhead habitat, it demonstrates the need for continued community-engaged observation, particularly during climatic anomalies and in the context of fast-changing environmental conditions under climate change.
The datasets presented in this article are not readily available because due to the confidential nature of interview data, primary data are stored in encrypted archives and may only be accessed by the core research team, in accordance with the Code of Ethics of the International Society of Ethnobiology. Requests to access the datasets should be directed toZWFybHljQHN0YW5mb3JkLmVk.
ME: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. NC: Conceptualization, Funding acquisition, Investigation, Writing – original draft, Writing – review & editing. LC: Funding acquisition, Resources, Writing – original draft, Writing – review & editing. GG: Conceptualization, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. JS: Funding acquisition, Resources, Writing – original draft, Writing – review & editing. DJ: Conceptualization, Investigation, Methodology, Resources, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. Research was funded through a David H. Smith Conservation Research Fellowship awarded to MMEC.
We thank the Society for Conservation Biology and the Cedar Tree Foundation. We thank the two reviewers whose comments greatly improved the quality of this manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1407575/full#supplementary-material
Abecassis M., Senina I., Lehodey P., Gaspar P., Parker D., Balazs G., et al. (2013). A model of loggerhead sea turtle (Caretta caretta) habitat and movement in the oceanic North Pacific. PloS One 8, e73274. doi: 10.1371/journal.pone.0073274
Albuquerque U. P., Cruz Da Cunha L. V. F., De Lucena R. F. P., Alves R. R. N. (Eds.) (2014). Methods and Techniques in Ethnobiology and Ethnoecology (New York, NY: Springer New York). doi: 10.1007/978–1-4614–8636-7
Albuquerque U. P. D., Ramos M. A., Medeiros P. M. D., De A. L. S., Almeida Reinaldo, Farias Paiva De Lucena (2008). “The role of visual stimuli in ethnobotanical surveys: An overview,”. in Current Topics in Ethnobotany, eds. Albuquerque U. P., Alves Ramos M. (Kerala, India: Research Signpost), 125–137.
Almpanidou V., Markantonatou V., Mazaris A. D. (2019). Thermal heterogeneity along the migration corridors of sea turtles: Implications for climate change ecology. J. Exp. Mar. Biol. Ecol. 520, 151223. doi: 10.1016/j.jembe.2019.151223
Aswani S., Lemahieu A., Sauer W. H. H. (2018). Global trends of local ecological knowledge and future implications. PloS One 13, e0195440. doi: 10.1371/journal.pone.0195440
Beaudreau A. H., Levin P. S. (2014). Advancing the use of local ecological knowledge for assessing data-poor species in coastal ecosystems. Ecol. Appl. 24, 244–256. doi: 10.1890/13-0817.1
Beaugrand G., Conversi A., Atkinson A., Cloern J., Chiba S., Fonda-Umani S., et al. (2019). Prediction of unprecedented biological shifts in the global ocean. Nat. Clim. Change 9, 237–243. doi: 10.1038/s41558-019-0420-1
Becker E. (2023) September 2023 ENSO Outlook: An El Niño convo (NOAA Climate.gov. NOAA Clim). Available online at: http://www.climate.gov/news-features/blogs/september-2023-enso-outlook-el-nino-convo (Accessed February 28, 2024).
Benson S. R., Forney K. A., Moore J. E., LaCasella E. L., Harvey J. T., Carretta J. V. (2020). A long-term decline in the abundance of endangered leatherback turtles, Dermochelys coriacea, at a foraging ground in the California Current Ecosystem. Glob. Ecol. Conserv. 24, e01371. doi: 10.1016/j.gecco.2020.e01371
Bernard H. R. (2011). Research Methods in Anthropology: Qualitative and Quantitative Approaches., 5th Edn (New York: AltaMira Press).
Briscoe D. K., Parker D. M., Bograd S., Hazen E., Scales K., Balazs G. H., et al. (2016). Multi-year tracking reveals extensive pelagic phase of juvenile loggerhead sea turtles in the North Pacific. Mov. Ecol. 4, 23. doi: 10.1186/s40462-016-0087-4
Briscoe D. K., Turner Tomaszewicz C. N., Seminoff J. A., Parker D. M., Balazs G. H., Polovina J. J., et al. (2021). Dynamic thermal corridor may connect endangered loggerhead sea turtles across the Pacific Ocean. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.630590
Brook R. K., McLachlan S. M. (2005). On Using Expert-Based Science to “Test” Local Ecological Knowledge. E&S 10, resp3. doi: 10.5751/ES-01478-1002r03
Casale P., Tucker A. D. (2017). Caretta caretta: the IUCN red list of threatened species. doi: 10.2305/IUCN.UK.2017–2.RLTS.T3897A119333622.en
Chandler M., See L., Copas K., Bonde A. M. Z., López B. C., Danielsen F., et al. (2017). Contribution of citizen science towards international biodiversity monitoring. Biol. Conserv. 213, 280–294. doi: 10.1016/j.biocon.2016.09.004
Chavez F. P., Messié M. (2009). A comparison of eastern boundary upwelling ecosystems. Prog. Oceanogr. 83, 80–96. doi: 10.1016/j.pocean.2009.07.032
Early-Capistrán M.-M., Solana-Arellano E., Abreu-Grobois F. A., Narchi N. E., Garibay-Melo G., Seminoff J. A., et al. (2020). Quantitative datasets and R Code: Quantifying local ecological knowledge to model historical abundance of long-lived, heavily-exploited fauna. PeerJ 8, e9494. doi: 10.7717/peerj.9494/supp-16
Eguchi T., McClatchie S., Wilson C., Benson S. R., LeRoux R. A., Seminoff J. A. (2018). Loggerhead turtles (Caretta caretta) in the California Current: Abundance, distribution, and anomalous warming of the North Pacific. Front. Mar. Sci. 5. doi: 10.3389/fmars.2018.00452
Fontaine A., Simard A., Brunet N., Elliott K. H. (2022). Scientific contributions of citizen science applied to rare or threatened animals. Conserv. Biol. 36, e13976. doi: 10.1111/cobi.13976
Gray S., Jordan R., Crall A., Newman G., Hmelo-Silver C., Huang J., et al. (2017). Combining participatory modelling and citizen science to support volunteer conservation action. Biol. Conserv. 208, 76–86. doi: 10.1016/j.biocon.2016.07.037
Griffin L. P., Griffin C. R., Finn J. T., Prescott R. L., Faherty M., Still B. M., et al. (2019). Warming seas increase cold-stunning events for Kemp’s ridley sea turtles in the northwest Atlantic. PloS One 14, e0211503. doi: 10.1371/journal.pone.0211503
Halpin L. R., Seminoff J. A., Hanke G. F. (2018). First photographic evidence of a loggerhead sea turtle (Caretta caretta) in British Columbia. Northwest. Nat. 99, 73–75. doi: 10.1898/NWN17-26.1
Hanna M. E., Chandler E. M., Semmens B. X., Eguchi T., Lemons G. E., Seminoff J. A. (2021). Citizen-sourced sightings and underwater photography reveal novel insights about green sea turtle distribution and ecology in southern California. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.671061
Hazen E. L., Abrahms B., Brodie S., Carroll G., Jacox M. G., Savoca M. S., et al. (2019). Marine top predators as climate and ecosystem sentinels. Front. Ecol. Environ. 17, 565–574. doi: 10.1002/fee.2125
Hazen E. L., Jorgensen S., Rykaczewski R. R., Bograd S. J., Foley D. G., Jonsen I. D., et al. (2013). Predicted habitat shifts of Pacific top predators in a changing climate. Nat. Clim. Change 3, 234–238. doi: 10.1038/nclimate1686
Innis C. J., Tlusty M., Merigo C., Weber E. S. (2007). Metabolic and respiratory status of cold-stunned Kemp’s ridley sea turtles (Lepidochelys kempii). J. Comp. Physiol. B 177, 623–630. doi: 10.1007/s00360-007-0160-9
International Society of Ethnobiology (2008) ISE Code of Ethics (with 2008 additions). Available online at: http://www.ethnobiology.net/wp-content/uploads/ISE-COE_Eng_rev_24Nov08.pdf (Accessed May 29, 2019).
Jambura P. L., Türtsche J., De Maddalena A., Giovos I., Kriwer J., Rizgalla J., et al. (2021). Using citized science to detect rare and endagered speices: new records of the great white shark (Cacharodon carcharias) off the Libyan coast. Annales : Ann. Istrian Mediterr. Stud. Ser. Hist. Naturalis 31, 51. doi: 10.19233/ASHN.2021.08
Jeliazkov A., Gavish Y., Marsh C. J., Geschke J., Brummitt N., Rocchini D., et al. (2022). Sampling and modelling rare species: Conceptual guidelines for the neglected majority. Glob. Change Biol. 28, 3754–3777. doi: 10.1111/gcb.16114
Jones B. L., Unsworth R. K. F., McKenzie L. J., Yoshida R. L., Cullen-Unsworth L. C. (2018). Crowdsourcing conservation: The role of citizen science in securing a future for seagrass. Mar. pollut. Bull. 134, 210–215. doi: 10.1016/j.marpolbul.2017.11.005
Lamont-Doherty Earth Observatory (2023) GeoMapApp 3.7.1 (GeoMapApp). Available online at: http://www.geomapapp.org/ (Accessed March 14, 2014).
Lee H.-J., Kim I.-H., Kim J.-K., Jeong S., Park D. (2014). First detailed morphological description of the loggerhead sea turtle (Caretta caretta) caught from the Yellow Sea of Korea. J. Ecol. Environ. 37, 201–208. doi: 10.5141/ecoenv.2014.024
Mallard F., Couderchet L. (2019). “Climate Sentinels Research Program: Developing indicators of the effects of climate change on biodiversity in the region of New Aquitaine (south west, France),” in Handbook of Climate Change and Biodiversity. Eds. Leal Filho W., Barbir J., Preziosi R. (Springer International Publishing, Cham), 223–241. doi: 10.1007/978–3-319–98681-4_14
Martínez-Estévez L., Angulo Angulo A., Astorga M. E., Becerra C. D., Leyva N. C., Amador F. C., et al. (2022). Exploring the demography and conservation needs of hawksbill sea turtles Eretmochelys imbricata in north-west Mexico. Oryx 57 (3), 1–9. doi: 10.1017/S0030605322000709
McKelvey K. S., Aubry K. B., Schwartz M. K. (2008). Using anecdotal occurrence data for rare or elusive species: the illusion of reality and a call for evidentiary standards. BioScience 58, 549–555. doi: 10.1641/B580611
McKinley D. C., Miller-Rushing A. J., Ballard H. L., Bonney R., Brown H., Cook-Patton S. C., et al. (2017). Citizen science can improve conservation science, natural resource management, and environmental protection. Biol. Conserv. 208, 15–28. doi: 10.1016/j.biocon.2016.05.015
Molinos J. G., Halpern B. S., Schoeman D. S., Brown C. J., Kiessling W., Moore P. J., et al. (2015). Climate velocity and the future global redistribution of marine biodiversity. Nat. Clim. Change 6, 83–88. doi: 10.1038/nclimate2769
Monterey Bay Aquarium (2011) Rare Stranding of Olive Ridley Sea Turtle in Pacific Grove (Sanctuary Integr. Monit. Netw). Available online at: https://sanctuarysimon.org/2011/10/rare-stranding-of-olive-ridley-sea-turtle-in-pacific-grove/ (Accessed August 11, 2023).
NASA Earth Observatory (2023) Sea Surface Temperature (1 month - Aqua/MODIS) (Sea Surf. Temp. 1 Mon. - AquaMODIS). Available online at: https://neo.gsfc.nasa.gov/view.php?datasetId=MYD28M (Accessed August 15, 2023).
National Marine Fisheries Service and U.S. Fish and Wildlife Service (2020). Loggerhead Sea Turtle (Caretta caretta) North Pacific Ocean DPS 5-Year Review: Summary and Evaluation (Washington D.C: NOAA & USFWS). Available at: https://www.fws.gov/node/1729941.
NOAA (2001) Endangered and Threatened Wildlife; Sea Turtle Conservation Requirements; Taking of Threatened or Endangered Species Incidental to Commercial Fishing Operations. Available at: https://www.federalregister.gov/documents/2001/08/24/01-21512/endangered-and-threatened-wildlife-sea-turtle-conservation-requirements-taking-of-threatened-or.
NOAA (2023a) Monterey Bay National Marine Sanctuary (Monterey Bay Natl. Mar. Sanctuary). Available online at: https://montereybay.noaa.gov/ (Accessed August 25, 2023).
NOAA (2023b) National Data Buoy Center: Station MLSC1 (Natl. Data Buoy Cent). Available online at: https://www.ndbc.noaa.gov/station_history.php?station=mlsc1 (Accessed August 11, 2023).
NOAA (2023c) NCEI Coastal Water Temperature Guide (Mon. Aver. Temp. °F Cent. Pac. Coast). Available online at: https://www.ncei.noaa.gov/access/coastal-water-temperature-guide/all_table.html (Accessed February 13, 2024).
NOAA (2024a) Climate Prediction Center: ENSO Diagnostic Discussion (NOAA Clim. Predict. Cent). Available online at: https://www.cpc.ncep.noaa.gov/products/analysis_monitoring/enso_advisory/ensodisc.html (Accessed August 24, 2023).
NOAA (2024b) NOAA Climate Prediction Center: Historical El Nino/La Nina episodes, (1950-present). Available online at: https://www.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v5.php (Accessed February 28, 2024).
Okuyama J., Watabe A., Takuma S., Tanaka K., Shirai K., Murakami-Sugihara N., et al. (2022). Latitudinal cline in the foraging dichotomy of loggerhead sea turtles reveals the importance of East China Sea for priority conservation. Divers. Distrib. 28, 1568–1581. doi: 10.1111/ddi.13531
Paduan J. D., Cook M. S., Tapia V. M. (2018). Patterns of upwelling and relaxation around Monterey Bay based on long-term observations of surface currents from high frequency radar. Deep Sea Res. Part II Top. Stud. Oceanogr. 151, 129–136. doi: 10.1016/j.dsr2.2016.10.007
Palmer C. T., Wadley R. L. (2007). RLocal Environmental Knowledge, Talk, and Skepticism: Using ‘LES’ to Distinguish ‘LEK’ from ‘LET’ in Newfoundland. L. Hum. Ecol. 35, 749–760. doi: 10.1007/s10745-006-9108-z
Palumbi S., Sotka C. (2012). The Death and Life of Monterey Bay: A Story of Revival (Washington D.C: Island Press).
Patrício A., Velez-Zuazo X., Diez C., Dam R., Sabat A. (2011). Survival probability of immature green turtles in two foraging grounds at Culebra, Puerto Rico. Mar. Ecol. Prog. Ser. 440, 217–227. doi: 10.3354/meps09337
Paull C., Caress D. (2019). High-resolution (1m) bathymetric data from Monterey Canyon, offshore California. doi: 10.1594/IEDA/324756
Paull C. K., Caress D. W., Iii W. U., Lundsten E., Meiner-Johnson M. (2011). High-resolution bathymetry of the axial channels within Monterey and Soquel submarine canyons, offshore central California. Geosphere 7, 1077–1101. doi: 10.1130/GES00636.1
Poloczanska E. S., Brown C. J., Sydeman W. J., Kiessling W., Schoeman D. S., Moore P. J., et al. (2013). Global imprint of climate change on marine life. Nat. Clim. Change 3, 919–925. doi: 10.1038/nclimate1958
Polovina J. J., Howell E. A., Kobayashi D. R., Seki M. P. (2017). The Transition Zone Chlorophyll Front updated: Advances from a decade of research. Prog. Oceanogr. 150, 79–85. doi: 10.1016/j.pocean.2015.01.006
Pozzi C. M., Ladio A. H. (2023). Variation of local zoological knowledge about Southern river otter and other semi-aquatic mammals in Nahuel Huapi National Park (Argentina). J. Ethnobiol. Ethnomedicine 19, 15. doi: 10.1186/s13002-023-00590-8
Pritchard P. C. H., Mortimer J. A. (1999)Taxonomy, external morphology, and species identification. In: Research and Management Techniques for the Conservation of Sea Turtles (Gland, Switzerland: IUCN/SSC Marine Turtle Specialist Group), 21–38. Available online at: https://rollingharbour.files.wordpress.com/2018/06/sea-turtle-id-sheet.jpg (Accessed August 25, 2023).
Ryan W. B. F., Carbotte S. M., Coplan J. O., O’Hara S., Melkonian A., Arko R., et al. (2009). Global multi-resolution topography synthesis. Geochem. Geophys. Geosystems 10, n/a–n/a. doi: 10.1029/2008GC002332
Saito T., Kurita M., Okamoto H., Kakizoe Y., Parker D., Briscoe D., et al. (2018). Satellite tracking immature loggerhead turtles in temperate and subarctic ocean habitats around the Sea of Japan. Micronesica 2018, 21.
Secretariat of the Pacific Community (2003) Marine turtle identification cards (Pac. Data Hub). Available online at: https://pacificdata.org/dataset/oai-www-spc-int-d8cde888–2ca3–460d-87a6–19fd2ef8c02b (Accessed August 25, 2023).
Sydeman W. J., Poloczanska E., Reed T. E., Thompson S. A. (2015). Climate change and marine vertebrates. Science 350, 772–777. doi: 10.1126/science.aac9874
Taheri S., Naimi B., Rahbek C., Araújo M. B. (2021). Improvements in reports of species redistribution under climate change are required. Sci. Adv. 7, eabe1110. doi: 10.1126/sciadv.abe1110
Tengö M., Austin B. J., Danielsen F., Fernández-Llamazares Á. (2021). Creating synergies between citizen science and indigenous and local knowledge. BioScience 71, 503–518. doi: 10.1093/biosci/biab023
The Marine Mammal Center (2023) Sea Turtle Reported in Monterey Harbor. Available online at: https://mail.google.com/mail/u/0/#search/ridley/FMfcgzGtwqGQMHBcBhqsttmCnchxpDWT (Accessed September 1, 2023).
Turner Tomaszewicz C. N., Seminoff J. A., Peckham S. H., Avens L., Kurle C. M. (2017). Intrapopulation variability in the timing of ontogenetic habitat shifts in sea turtles revealed using δ 15 N values from bone growth rings. J. Anim. Ecol. 86, 694–704. doi: 10.1111/1365-2656.12618
Upwell, Grupo Tortuguero de las Californias (2023) East Pacific Sea Turtles Field Guide. Available online at: https://static1.squarespace.com/static/59cae0d6be42d63f64cf6dd2/t/5cb928d4f4e1fc095efbf7dc/1555638522637/East+Pacific+Turtles+Field+Guide (Accessed August 25, 2023).
Webb E. J., Magi B. I. (2022). The ensemble oceanic niño index. Int. J. Climatol. 42, 5321–5341. doi: 10.1002/joc.7535
Wedemeyer-Strombel K. R., Peterson M. J., Sanchez R. N., Chavarría S., Valle M., Altamirano E., et al. (2019). Engaging fishers’ ecological knowledge for endangered species conservation: Four advantages to emphasizing voice in Participatory Action Research. Front. Commun. 4. doi: 10.3389/fcomm.2019.00030
Keywords: climate change, sea turtles, marine conservation, ethnobiology, transdisciplinary
Citation: Early-Capistrán MM, Crane NL, Crowder LB, Garibay-Melo G, Seminoff JA and Johnston D (2024) Ethnobiological methods enhance our capacity to document potential climate sentinels: a loggerhead sea turtle (Caretta caretta) case study. Front. Mar. Sci. 11:1407575. doi: 10.3389/fmars.2024.1407575
Received: 26 March 2024; Accepted: 14 May 2024;
Published: 28 May 2024.
Edited by:
Xuelei Zhang, Ministry of Natural Resources, ChinaReviewed by:
Juanita Joseph, Universiti Malaysia Sabah, MalaysiaCopyright © 2024 Early-Capistrán, Crane, Crowder, Garibay-Melo, Seminoff and Johnston. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michelle María Early-Capistrán, ZWFybHljQHN0YW5mb3JkLmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.