 Brooke N. Anderson
Brooke N. Anderson Lauren Horstmyer2
Lauren Horstmyer2 Jon F. Dodd
Jon F. Dodd- 1School of Life Sciences, Arizona State University, Tempe, AZ, United States
- 2Coastal Oregon Marine Experiment Station, Oregon State University, Newport, OR, United States
- 3Atlantic Shark Institute, Wakefield, RI, United States
Pop-off satellite archival tags (PSATs) have been increasingly deployed on marine megafauna to remotely monitor their movements, behavior, and environmental preferences. These tags have also allowed the occasional novel documentation of ecological interactions, such as predation on tagged animals. A PSAT deployed on a pregnant porbeagle Lamna nasus in the Northwest Atlantic suggests the shark was predated upon in mesopelagic waters near Bermuda. Predation was evident approximately 5 months after tagging based on depth and temperature data transmitted by the PSAT. Four days prior to PSAT pop-off, depth data indicated that the tag continued to descend and ascend in the water column while the temperature remained approximately 5°C above ambient levels, even at several hundred meters in depth, indicating ingestion. Given the location of predation and elevated temperature at depth recorded by the pregnant porbeagle’s tag, potential predators include endothermic shark species such as the white shark Carcharodon carcharias and shortfin mako Isurus oxyrhinchus. This is the first evidence of predation on a porbeagle globally and provides novel insight into inter-specific interactions for this large, threatened shark species.
1 Introduction
Pop-off satellite archival tags (PSATs) have been traditionally utilized to examine the three-dimensional movements of elasmobranchs over varying temporal and spatial scales (e.g., Skomal et al., 2021). PSATs have also been used to document post-release mortality (PRM) associated with capture and handling (e.g., Goldsmith et al., 2017), and have opportunistically documented predation events in species such as Atlantic salmon Salmo salar (Lacroix, 2014) and white marlin Tetrapturus albidus (Kerstetter et al., 2004). To discern between the two types of mortality, several characteristic changes can often be observed in the PSAT data to indicate tag ingestion by a predator, including the sudden absence of light (e.g., Béguer-Pon et al., 2012), temperature increases (coinciding with the stomach temperature of predatory species), and alterations in depth range or diving behavior (e.g., Kerstetter et al., 2004).
Porbeagle Lamna nasus are large-bodied, highly mobile sharks documented in cold-temperate waters of the Northwest Atlantic through previous PSAT studies (Skomal et al., 2021). The Northwest Atlantic populations’ life history is characterized by late ages at maturity (Natanson et al., 2002), low reproductive output (Jensen et al., 2002; Natanson et al., 2019), and low estimated annual natural mortality (Cortés and Semba, 2020). We present the first documented predation of an adult female porbeagle affixed with a PSAT that migrated off the continental shelf waters of New England to the Sargasso Sea, and suggest the predators likely responsible for the observed mortality.
2 Methods
Porbeagles were caught via rod-and-reel (following Anderson et al., 2021) southeast of Cape Cod, Massachusetts, US in late October of 2020 and 2022. Captured porbeagles were brought onboard the vessel and a saltwater pump was inserted into the shark’s mouth to facilitate respiration. Total time on deck averaged less than 15 minutes. Sex, fork length (FL; cm), total length (TL; cm), and capture location were recorded. Females near or above the size at maturity (210-230 cm FL; Jensen et al., 2002) were examined for pregnancy via ultrasound following Sulikowski and Hammerschlag (2023). An Ibex EVO II portable ultrasound (E.I Medical Imaging) with a 60 mm curved linear array 5-2.5 MHz transducer (model 290470) capable of a 24 cm scan depth was used to obtain images of the reproductive tract. Scanning was performed on the ventral surface from the pectoral to the pelvic fin in both a transverse and longitudinal orientation to obtain cross-sectional and lengthwise images of the uterus, respectively.
Porbeagles were tagged with both a fin-mounted satellite linked transmitter (hereafter “finmount tag”) and a PSAT. The finmount tag attached to the predated porbeagle was a Sirtrack F6F FastGPS Argos transmitter (Lotek Wireless). The finmount tag was attached to the sharks’ first dorsal fin following established protocols (e.g., Gibson et al., 2021). Additionally, a 365-day PSATFLEX (Lotek Wireless) was looped through a hole in the first dorsal fin of each shark following Anderson et al. (2021). PSATs were programmed to collect pressure in dbar (i.e., depth in m, hereafter referred to as depth) and ambient water temperature (°C) at 20-s intervals for the pre-programmed 365-day deployment period, after which the tags were designed to float to the sea surface and transmit archived data (via Argos satellites) in compressed 2-hour bins (i.e., one depth and temperature record every 2 hours). The PSATs are designed to report daily light-based geolocation estimates that are determined onboard the tag by manufacturer proprietary software (Ekstrom, 2004) and thus raw light-level data are not reported. PSATs were programmed to release and transmit prematurely if pressure remained constant (± 5 m) for 3 days, indicating a mortality or a shed tag floating at the sea surface or washed ashore.
For geographic location data, PSAT light-based geolocation estimates were discarded due to a low number of reliable position estimates. In particular, light-based PSAT latitude estimates ranged from –90.1 to 78.1 and longitude estimates from –177.7 to 175.9. Latitudinal error estimates ranged from 0 to 67.0° (mean ± SD = 10.4° ± 15.0°) and longitudinal error estimates from 0.2 to 51.6° (mean ± SD = 3.4° ± 6.0°). The deep diving behavior of porbeagles while in mesopelagic habitats likely contributed to a proportion of the erroneous positions in each PSAT track (Campana et al., 2010; Heard et al., 2017). Position estimates from finmount tags were downloaded from Argos satellites (CLS America, Inc.). Argos provides location accuracy using the following location classes (LC): 3, 2, 1, 0, A, B, and Z (in decreasing accuracy), with accuracy ranging from < 250 m to > 5 km for LC 3 to B (Tougaard et al., 2008). There were no transmissions categorized as the poorest accuracy LC (Z).
It is standard practice for satellite telemetry data to be regularized and interpolated with a state-space model (e.g., Jonsen et al., 2023) when assessing the habitat use of marine species. Unfortunately, due to the limited number of finmount satellite transmissions before predation and necessary discarding of PSAT geolocation estimates, tag data could not be processed with a state-space model in this study. However, given satellite telemetry data were strictly used for helping to identify the general locations of the tagged porbeagle rather than assessing habitat use patterns, the highest-accuracy finmount tag transmission for a given day were plotted using the R package ‘ggmap’ (Kahle and Wickham, 2013), along with the capture location and the first satellite transmission received from the PSAT after pop-off. The minimum horizontal displacement of the shark was estimated by calculating the distance (km) between the capture location and the first satellite transmission received by the PSAT following pop-off. For depth and temperature data recorded by the PSAT, the temperature-integrated diving track was plotted across the time-series using the R package ‘ggplot’ (Wickham, 2016). Additionally, the average ambient temperatures at depths were calculated and compared between the week before apparent tag ingestion and during the tag ingestion period.
3 Results
A 223 cm FL pregnant porbeagle was captured and tagged southeast of Cape Cod, MA on October 28, 2020 (Figure 1). The shark’s PSAT transmitted its first location (LC 1) southwest of Bermuda 158 days after tagging, on April 3, 2021 (Figure 1). The minimum horizontal displacement of the shark was estimated to be 1,120 km. However, it’s important to note that there was a three-and-a-half-day gap between the last transmitted depth measurement and the first satellite transmission after pop-off, during which the PSAT was likely drifting at the sea surface (e.g., Peel et al., 2020). The shark’s finmount tag only transmitted one location on Georges Bank on November 12, 2020 (Figure 1). Based on PSAT depth data, the pregnant porbeagle was diving between the sea surface and approximately 100 m from the day of tagging until December 26, 2020 (Figure 2). During this time, ambient temperatures recorded by the tag ranged from 10.28 to 20.54°C (Figure 2). From late December to late March, the shark transitioned diving behavior, spending nighttime at approximately 100-200 m and daytime at approximately 600-800 m, suggesting that the shark had moved off of the continental shelf (Figure 2). During this period, the shark occupied water temperatures of 6.4 to 23.52°C (Figure 2). However, on March 24, 2021, ambient temperatures recorded by the PSAT increased, ranging between 16.4 and 24.72°C despite recording a similar depth range of approximately 150-600 m (Figure 2), indicating tag ingestion. In the week prior to the apparent predation, overall average temperature recorded by the tag was 17.4°C. During that time, average temperature was 19.1°C at 100-300 m depth and 16.1°C at 500-700 m depth. In comparison, during the apparent tag ingestion period, the overall average temperature recorded by the tag was 22.2°C. Average temperature was 21.8°C at 100-300 m depth and 22.2°C at 500-700 m depth during tag ingestion.
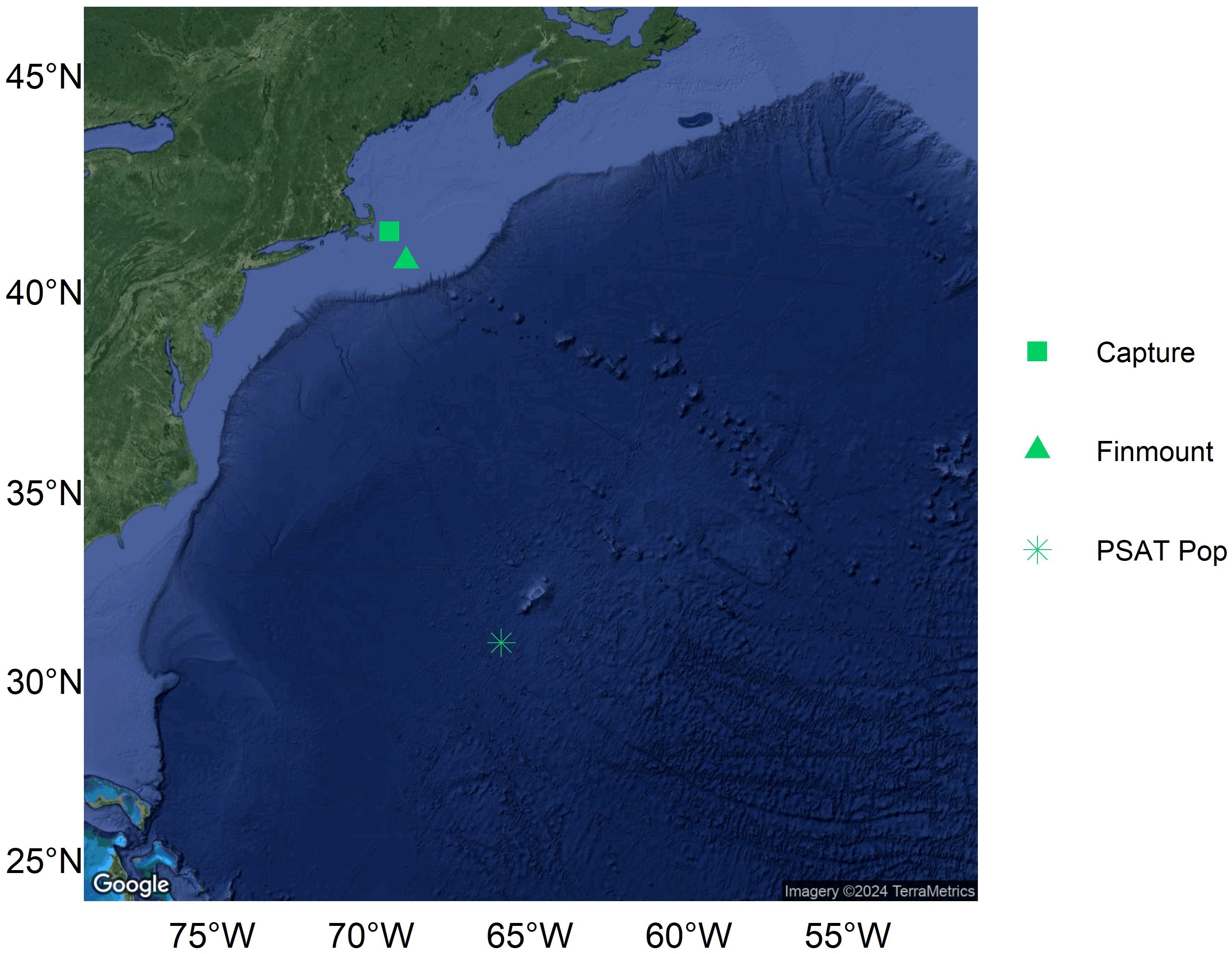
Figure 1 Map of the capture location, the single highest-accuracy daily finmount tag transmission, and the first transmission received from the PSAT after pop-off for a pregnant porbeagle tagged in the Northwest Atlantic. It is important to note that there is a time gap between the last depth measurement collected from the PSAT and the first transmission after pop-off.
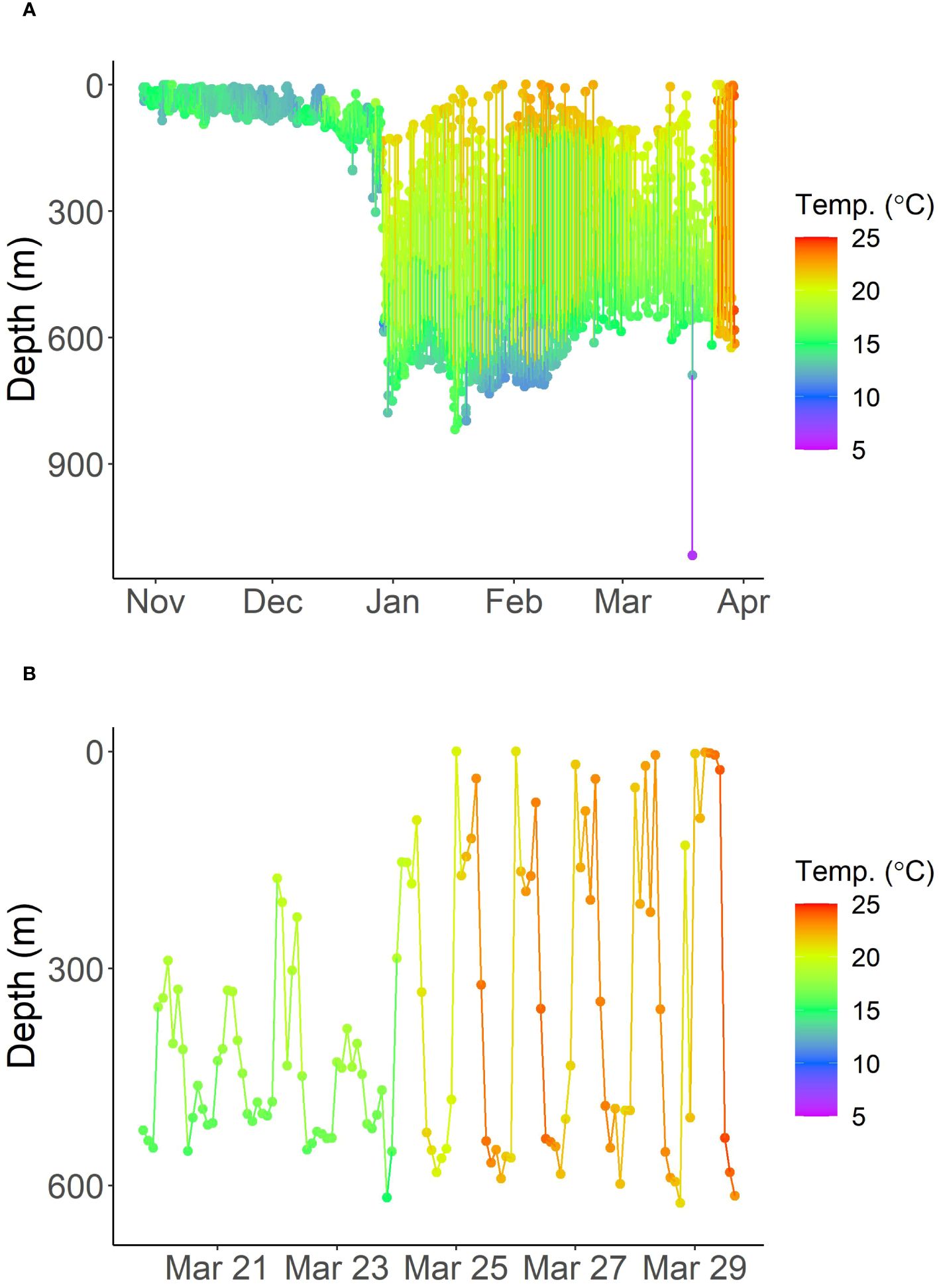
Figure 2 Depth and temperature time-series over the entire deployment (A) and the last week before pop-off (B) for the tagged porbeagle.
4 Discussion
Based on previous work (e.g., Anderson et al., 2021; Bowlby et al., 2021), the post-release diving behavior for the porbeagle tagged in this study appeared characteristic for the species and suggest that capture and handling effects during tagging were not influencing the observed mortality (Bowlby et al., 2021). In addition, PRM has been shown to largely occur immediately (or within 48 hours) of release in large pelagic sharks (e.g., Campana et al., 2016), and physiological recovery of lamnid sharks post-release suggest the group are generally resilient to capture and handling methods (Musyl and Gilman, 2019; Anderson et al., 2021; Bowlby et al., 2021). Based on the collective understanding of PRM and recovery in porbeagles and other lamnids, the PSAT data described herein are indicative of natural morality caused by predation rather than from stress from the capture and handling of the shark months prior to the observed mortality. In addition, given the slight change in diving pattern and premature release of the PSAT, it is unlikely that the observation reported here was related to movement into a warmer water body.
The depth and temperature data recorded by the sharks’ PSAT as well as the general location of predation can be used to identify potential predators. The sudden elevated temperatures (16.4-24.72°C) and stabilized thermal excess (5°C above ambient temperature on average) during ingestion were characteristic of gut temperatures reported for other large endothermic fish (Goldman, 1997; Sepulveda et al., 2004; Jorgensen et al., 2015). Large marine mammals, such as odontocete whales (e.g., Orcas Orcinus orca) maintain much higher internal temperatures (close to 40°C; Whittow et al., 1974; Strøm et al., 2019) and so were not considered as a potential predator in this instance. Two endothermic predator candidates large enough to predate upon mature porbeagles and located within the vicinity and at the time of year of the predation event include the white shark Carcharodon carcharias (Skomal et al., 2017) and shortfin mako Isurus oxyrhinchus (Santos et al., 2021). Following ontogenetic shifts in diet, prey species of adult white sharks and shortfin makos are known to include other elasmobranch species (and marine mammals) (e.g., Joyce et al., 2002). While fine-scale diving data for these potential predators are limited in the location that the predation event occurred, both white sharks (Skomal et al., 2017) and shortfin makos (Santos et al., 2021; Loefer et al., 2005) have been documented to occupy broad depth distributions from the sea surface to >300 m while in offshore waters near the Sargasso Sea. However, tagged shortfin makos mainly occupied depths less than 400 m and made rapid oscillatory dives between the sea surface and deeper depths during the day while in offshore waters of the North Atlantic (Santos et al., 2021; Loefer et al., 2005), which was not characteristic of the predator’s behavior in this study. In comparison to shortfin makos, large female white sharks spent more time at depths of 400-600 m while in mesopelagic waters of the North Atlantic (Skomal et al., 2017). Given this comparison, a white shark was likely the predator of our tagged porbeagle.
The data presented in this study is, to the best of our knowledge, the first evidence of predation on a porbeagle and provides novel insight into inter-specific interactions for this large, globally vulnerable (IUCN, 2023) shark species. Knowledge gaps persist regarding natural mortality rates for porbeagles, and this work suggests predation may be an underestimated source of natural mortality for the Northwest Atlantic population. For example, 11 porbeagles (excluding one immediate mortality) were tagged with year-long PSATs and transmitted data which could be used to opportunistically identify natural mortality events. Two (18.2%) natural mortality events were observed, including the predation documented in this study as well as the mortality (of unknown cause) of an adult female that also occurred near Bermuda approximately 50 days after tagging. However, it is important to consider that it has been suggested that marine animals tagged with tethered PSATs may be at increased risk of predation (e.g., tags act as lure or emits attractant signal) (Thorstad et al., 2013; Hall and James, 2021; Lennox et al., 2023). Nevertheless, continued research of mesopelagic predator-prey interactions using satellite tags is warranted to increase our knowledge of ecosystem dynamics in the open ocean and thus improve the efficacy of management strategies to maintain ecosystem health. For example, the observations presented here suggest mesopelagic waters near Bermuda are an area of important inter-species interaction and mortality for porbeagles and thus should be a location of future research efforts.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by Arizona State University Institutional Animal Care and Use Committee. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
BA: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Writing – original draft, Writing – review & editing, Project administration. LH: Writing – original draft, Writing – review & editing. KB: Writing – original draft, Writing – review & editing. JD: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Writing – review & editing. JS: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Fieldwork for this study was supported in part by the American Elasmobranch Society Donald R. Nelson award and the Arizona State University Graduate and Professional Student Association Graduate Research and Support Program. The Animal Telemetry Network provided satellite time for tags.
Acknowledgments
We would like to thank boat captains W. Hatch and S. Riddle as well as crew members for providing access to fishing vessels and assistance in fieldwork for this study. We also gratefully acknowledge B. Campbell for fieldwork assistance. Thank you to the Animal Telemetry Network for providing satellite time for tags. All capture, handling, and sampling protocols were approved by the Arizona State University Institutional Animal Care and Use Committee (20-1745R).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Anderson B. N., Bowlby H. D., Natanson L. J., Coelho R., Cortes E., Domingo A., et al. (2021). Preliminary estimate of post-release survival of immature porbeagles caught with rod-and-reel in the Northwest Atlantic Ocean. Mar. Ecol. Prog. Ser. 660, 153–159. doi: 10.3354/meps13603
Béguer-Pon M., Benchetrit J., Castonguay M., Aarestrup K., Campana S. E., Stokesbury M. J., et al. (2012). Shark predation on migrating adult American eels (Anguilla rostrata) in the Gulf of St. Lawrence. PLoS One 7, e46830. doi: 10.1371/journal.pone.0046830
Bowlby H. D., Benoît H. P., Joyce W., Sulikowski J., Coelho R., Domingo A., et al. (2021). Beyond post-release mortality: inferences on recovery periods and natural mortality from electronic tagging data for discarded lamnid sharks. Front. Mar. Sci. 8, 619190. doi: 10.3389/fmars.2021.619190
Campana S. E., Joyce W., Fowler M. (2010). Subtropical pupping ground for a cold-water shark. Can. J. Fish. Aquat. Sci. 67, 769–773. doi: 10.1139/F10-020
Campana S. E., Joyce W., Fowler M., Showell M. (2016). Discards, hooking, and post-release mortality of porbeagle (Lamna nasus), shortfin mako (Isurus oxyrinchus), and blue shark (Prionace glauca) in the Canadian pelagic longline fishery. ICES J. Mar. Sci. 73, 520–528. doi: 10.1093/icesjms/fsv234
Cortés E., Semba Y. (2020). Estimates of vital rates and population dynamics parameters of interest for porbeagle shark in the western North Atlantic and South Atlantic Ocean. Collect. Vol. Sci. Pap. ICCAT 77, 118–131.
Ekstrom P. A. (2004). An advance in geolocation by light Vol. Special Issue 58 (Memoirs of National Institute of Polar Research), 210–226.
Gibson K. J., Streich M. K., Topping T. S., Stunz G. W. (2021). New insights into the seasonal movement patterns of shortfin mako sharks in the Gulf of Mexico. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.623104
Goldman K. J. (1997). Regulation of body temperature in the white shark, Carcharodon carcharias. J. Comp. Physiol. B. 167, 423–429. doi: 10.1007/s003600050092
Goldsmith W. M., Scheld A. M., Graves J. E. (2017). Performance of a low-cost, solar-powered pop-up satellite archival tag for assessing post-release mortality of Atlantic bluefin tuna (Thunnus thynnus) caught in the US east coast light-tackle recreational fishery. Anim. Biotelemetry 5, 1–13. doi: 10.1186/s40317-017-0144-9
Hall K. E., James M. C. (2021). Predation of satellite-tagged juvenile loggerhead turtles Caretta caretta in the Northwest Atlantic Ocean. Endanger. Species Res. 46, 279–291. doi: 10.3354/esr01165
Heard M., Rogers P. J., Bruce B. D., Humphries N. E., Huveneers C. (2017). Plasticity in the diel vertical movement of two pelagic predators (Prionace glauca and Alopias vulpinas) in the southwestern Indian Ocean. Fish. Oceanog. 27, 199–211. doi: 10.1111/fog.12245
IUCN (2023). The IUCN Red List of Threatened Species. Version 2023-1. Available online at: https://www.iucnredlist.org (Accessed 19 March 2024).
Jensen C. F., Natanson L. J., Pratt H. L. Jr., Kohler N., Campana S. E. (2002). The reproductive biology of the porbeagle shark (Lamna nasus) in the western North Atlantic Ocean. Fish. Bull. 100, 727–738.
Jonsen I. D., Grecian W. J., Phillips L., Carroll G., McMahon C., Harcourt R. G., et al. (2023). aniMotum, an R package for animal movement data: rapid quality control, behavioural estimation and simulation. Methods Ecol. Evol. 14, 806–816. doi: 10.1111/2041-210X.14060
Jorgensen S. J., Gleiss A. C., Kanive P. E., Chapple T. K., Anderson S. D., Ezcurra J. M., et al. (2015). In the belly of the beast: resolving stomach tag data to link temperature, acceleration and feeding in white sharks (Carcharodon carcharias). Anim. Biotelem. 3, 1–10. doi: 10.1186/s40317-015-0071-6
Joyce W. N., Campana S. E., Natanson L. J., Kohler N. E., Pratt J. H.L., Jensen C. F. (2002). Analysis of stomach contents of the porbeagle shark (Lamna nasus Bonnaterre) in the northwest Atlantic. ICES J. Mar. Sci. 59, 1263–1269. doi: 10.1006/jmsc.2002.1286
Kahle D., Wickham H. (2013). ggmap: spatial visualization with ggplot2. R J. 5, 144–161. doi: 10.32614/RJ-2013-014
Kerstetter D. W., Polovina J., Graves J. E. (2004). Evidence of shark predation and scavenging on fishes equipped with pop-up satellite archival tags. Fish. Bull. 102, 750.
Lacroix G. L. (2014). Large pelagic predators could jeopardize the recovery of endangered Atlantic salmon. Can. J. Fish. Aquat. Sci. 71, 343–350. doi: 10.1139/cjfas-2013-0458
Lennox R. J., Dahlmo L. S., Ford A. T., Sortland L. K., Vogel E. F., Vollset K. W. (2023). Predation research with electronic tagging. Wildl. Biol. 2023, e01045. doi: 10.1002/wlb3.01045
Loefer J. K., Sedberry G. R., McGovern J. C. (2005). Vertical movements of a shortfin mako in the western North Atlantic as determined by pop-up satellite tagging. Southeastern Naturalist 4 (2), 237–246. doi: 10.1656/1528-7092(2005)004[0237:VMOASM]2.0.CO;2
Musyl M. K., Gilman E. L. (2019). Meta-analysis of post-release fishing mortality in apex predatory pelagic sharks and white marlin. J. Fish. 20, 466–500. doi: 10.1111/faf.12358
Natanson L. J., Deacy B. M., Joyce W., Sulikowski J. (2019). Presence of resting population of female porbeagles (Lamna nasus), indicating a biennial reproductive cycle, in the western North Atlantic Ocean. Fish. Bull. 117, 70–77. doi: 10.7755/FB
Natanson L. J., Mello J. J., Campana S. E. (2002). Validated age and growth of the porbeagle shark (Lamna nasus) in the western North Atlantic Ocean. Fish. Bull. 100, 266–278.
Peel L. R., Stevens G. M. W., Daly R., Keating Daly C. A., Collin S. P., Nogués J., et al. (2020). Regional movements of reef manta rays (Mobula alfredi) in Seychelles waters. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00558
Santos C. C., Domingo A., Carlson J., Natanson L. J., Travassos P., Macías D., et al. (2021). Movements, habitat use, and diving behavior of shortfin mako in the Atlantic Ocean. Front. Mar. Sci. 8, 686343. doi: 10.3389/fmars.2021.686343
Sepulveda C. A., Kohin S., Chan C., Vetter R., Graham J. B. (2004). Movement patterns, depth preferences, and stomach temperatures of free-swimming juvenile mako sharks, Isurus oxyrinchus, in the Southern California Bight. Mar. Biol. 145, 191–199. doi: 10.1007/s00227-004-1356-0
Skomal G. B., Braun C. D., Chisholm J. H., Thorrold S. R. (2017). Movements of the white shark Carcharodon carcharias in the North Atlantic Ocean. Mar. Ecol. Prog. Ser. 580, 1–16. doi: 10.3354/meps12306
Skomal G., Marshall H., Galuardi B., Natanson L., Braun C. D., Bernal D. (2021). Horizontal and vertical movement patterns and habitat use of juvenile porbeagles (Lamna nasus) in the western north Atlantic. Front. Mar. Sci. 8, 624158. doi: 10.3389/fmars.2021.624158
Strøm J. F., Rikardsen A. H., Campana S. E., Righton D., Carr J., Aarestrup K., et al. (2019). Ocean predation and mortality of adult Atlantic salmon. Sci. Rep. 9, 789. doi: 10.1038/s41598-019-44041-5
Sulikowski J. A., Hammerschlag N. (2023). A novel intrauterine satellite transmitter to identify parturition in large sharks. Sci. Adv. 9, eadd6340. doi: 10.1126/sciadv.add6340
Thorstad E. B., Rikardsen A. H., Alp A., Økland F. (2013). The use of electronic tags in fish research–an overview of fish telemetry methods. Turk. J. Fish. Aquat. Sci. 13, 881–896. doi: 10.4194/1303-2712-v13_5_13
Tougaard J., Teilmann J., Tougaard S. (2008). Harbour seal spatial distribution estimated from Argos satellite telemetry: overcoming positioning errors. Endanger. Species Res. 4, 113–122. doi: 10.3354/esr00068
Whittow G. C., Hampton I. F. G., Matsuura D. T., Ohata C. A., Smith R. M., Allen J. F. (1974). Body temperature of three species of whales. J. Mammal. 55, 653–656. doi: 10.2307/1379555
Wickham H. (2016). ggplot2: Elegant Graphics for Data Analysis (Springer-Verlag: New York). Available at: https://ggplot2.tidyverse.org, ISBN: ISBN 978-3-319-24277-4. doi: 10.1007/978-3-319-24277-4
Keywords: ecology, reproduction, inter-species interaction, Bermuda, Lamna nasus, white shark, shortfin mako
Citation: Anderson BN, Horstmyer L, Ballard KL, Dodd JF and Sulikowski JA (2024) First evidence of predation on an adult porbeagle equipped with a pop-off satellite archival tag in the Northwest Atlantic. Front. Mar. Sci. 11:1406973. doi: 10.3389/fmars.2024.1406973
Received: 25 March 2024; Accepted: 16 July 2024;
Published: 03 September 2024.
Edited by:
Emily Lester, Australian Institute of Marine Science (AIMS), AustraliaReviewed by:
Samantha Andrzejaczek, Stanford University, United StatesMark Andrew Royer, University of Hawaii at Manoa, United States
Copyright © 2024 Anderson, Horstmyer, Ballard, Dodd and Sulikowski. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Brooke N. Anderson, Ym5hbmRlcnNvQGdtYWlsLmNvbQ==