 Madeleine Hamamé
Madeleine Hamamé Gustavo Aedo
Gustavo Aedo Paula Ortiz
Paula Ortiz Andrés Olguín3
Andrés Olguín3 Luis Miguel Pardo
Luis Miguel Pardo
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 17 July 2024
Sec. Marine Biology
Volume 11 - 2024 | https://doi.org/10.3389/fmars.2024.1392758
This article is part of the Research TopicAdvances in crustacean research from the 10th International Crustacean CongressView all 12 articles
Small-scale fisheries have been typically data-limited despite their economic importance for local communities. This is especially true in zones where fishing operations occur in remote areas under harsh weather conditions. Crab fishery in Chile is exclusively artisanal, and marble crab (Metacarcinus edwardsii) has the highest landing records. This species is found in most parts of the coast of Chile, but it is mainly caught in the south of the country (including Patagonia). Fishery management is data-limited, and monitoring has not established the spatial and temporal variability baseline necessary to determine its exploitation status. This fishery is currently evaluated annually under a scheme based on reporting biological and fishery information, primarily from the landing ports and secondarily from fishing grounds. In the present study, we collected data from fishing grounds on board artisanal fishing vessels around 45°S during an annual cycle to establish indicators based on catch per unit effort (above and below minimum legal size), size, sex ratio, and the relationship between weight and size. Our results showed that fishery and biological indicators respond more to seasonal patterns than to expected spatial heterogeneity related to different fishing grounds: proportional stock density (lowest in winter), sex ratio (biased toward males in winter), retained catch (highest in autumn), and released catch (highest in winter), while the average size of the largest 10% of the sample only showed differences between males and females. Additionally, released catch varied according to soak time, which indicates the effectiveness of escape rings incorporated in the traps. These results lead to recommendations related to the current monitoring program, which should include the selection of a limited number of fishing grounds standardized by season and the incorporation of information related to soak time and the presence of escape rings. Finally, this study highlights the importance of on-board scientific monitoring for any fisheries, even those of small scale.
Since 2013, Chilean fisheries have been regulated by a legislative framework [General Law of Fisheries and Aquaculture (LGPA)] that establishes the maximum sustainable yield (MSY) as the main management objective (Tsikliras and Froese, 2018). This requires the implementation of stock assessment models, based on long-term abundance data and life-history parameters, which allow the estimation of biological reference points (BRPs) (Collie and Gislason, 2001; Morgan et al., 2014). In Chile, there are currently 44 stocks formally declared as fisheries, 27 of which have BRPs (Subpesca, 2022), a low percentage, considering that ca. 150 species of fish, crustaceans, mollusks, and other resources are reported in the national landing statistics. This implies that at least 70% of the species that landed in the country lack estimations of MSY, stock abundance, or fishery parameters.
Most of these data-limited fisheries are identified in the category of small-scale fisheries (SSFs) (Béné, 2006). In the Aysén Region, southern Chile (43°38′–49°16′S), only SSFs are allowed in the so-called “inner sea” since the marine ecosystem is restricted to narrow channels and fjords. Over the years, several fishing grounds have been established, maintaining mainly demersal and benthic fisheries that are the main sustenance for local economies. In this region, latitudinal and longitudinal productivity gradients are the result of a marked seasonal pattern associated with solar radiation, freshwater discharge, and precipitation (Aracena et al., 2011; Montero et al., 2011), creating different oceanographic conditions that influence species distribution and their behavioral patterns on a small scale. Therefore, any monitoring strategy should consider the possible effects of this spatial and temporal heterogeneity.
The marble crab, Metacarcinus edwardsii (Bell, 1835), is an SSF concentrated mainly in southern Chile (from 36°6′ to 45°3′S), with average annual landings of 4,300 t in the last 10 years (Sernapesca, 2022). The official records of marble crab landings in the Aysén Region date back to 1976 and have been increasing since 1991, being a persistent artisanal fishery for over 30 years. This region contributed on average 20% of this species’ landings (an average of 800 t in the last 10 years; Sernapesca, 2022), and 83% of local fishers have licenses to catch this species. This fishery is monitored annually under a scheme that records biological and fishery information, mainly from the landing ports (Olguín and Mora, 2022) since data collection from fishing grounds is difficult and costly. The remoteness of fishing grounds, typically requiring 8 to 48 hours of sailing from the main port, along with harsh weather conditions prevailing for most of the year, represent the primary obstacles to monitoring efforts. Under this context, the government monitoring program estimates total landings (in biomass), size structure, and sex ratio (Olguín and Mora, 2022). However, it lacks estimations for crucial fishery indicators such as effort, catch, and other operational variables like soak time, which necessitate direct collection from fishing operations. This data limitation does not allow the detection of spatial and temporal changes in biological and fishery indicators, and hence, the status of the fishery has not been established. As a result, the regulation of this fishery is restricted only to fishing licenses, gear requirements, minimum legal size, and the prohibition of harvesting of ovigerous females. So far, no regulations on fishing effort or total allowable catch have been implemented.
The main objective of this study was to determine the seasonal and spatial trends of the biological and fishery indicators of M. edwardsii in its southernmost fishing grounds, the Aysén Region. The results will allow for recommendations on improving the current monitoring program.
The study was carried out in northern Chilean Patagonia, in an area located around 45°20′S and 73°40′W (Figure 1). The marine ecosystem in this region is influenced by a main channel called Moraleda, which separates the archipelago to the west from the continental coast to the east. Fjords, channels, and numerous islands characterize this “inner sea”. Water exchange from the open ocean, glacial inputs, light regime, and freshwater discharge varied over latitudinal and longitudinal gradients, presenting a strong seasonal cycle that determines hydrographic conditions and hence productivity patterns (Aracena et al., 2011; Montero et al., 2011). Near the southern extreme of the Moraleda Channel, there is a shallow sill (60 m deep), called the “Meninea Constriction”, which partially isolates the southern part of the Moraleda Channel and Aysén Fjord from the open ocean. The sill constrains circulation, forming two basins (North and South); the southern basin is warmer, less saline, and more oxygenated than the northern one (Silva et al., 1995, Silva et al., 2000). The northern basin in its western part is influenced by water masses from the Moraleda Channel, i.e., estuarine waters on the surface, modified subantarctic waters as an intermediate water mass, and, in deeper zones, equatorial subsurface waters. In the east, the basin is influenced by waters from the Aysén Fjord. Two water masses are present during the year at the mouth of this fjord, varying their depth according to the season. At the mouth of the fjord, salty estuarine water (21–31 psu) dominates the upper 40 m of the water column, becoming deeper in winter (approximately 100 m). Below this water mass, modified subantarctic water mass dominates at different depths depending on the season (Guzman, 2004).
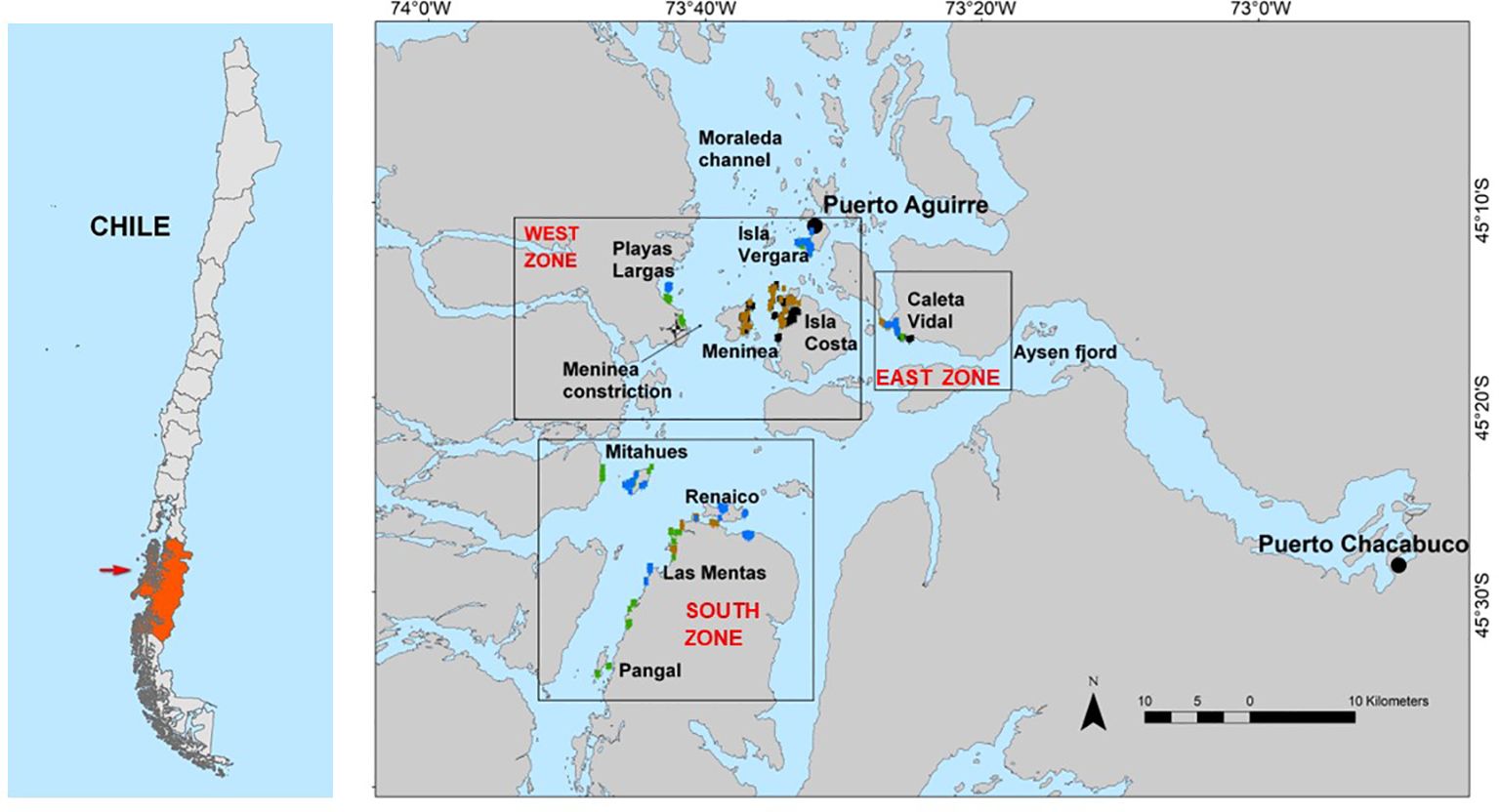
Figure 1 Study area; red squares show the main marble crab fishing grounds, south of Puerto Aguirre cove (45°10′S–45°30′W). Colors indicate the location of the fishing traps during autumn (brown), winter (black), spring (green), and summer (blue).
The study area was selected since it is the main regional fishing ground associated with marble crab (M. edwardsii). Fishers and their families reside near the fishing grounds, with travel times ranging from 10 min to several hours of navigation. Extraction occurs year-round, except for a summer break, typically from mid-December until early March, following commercial agreements with buyers.
In this context, 10 fishing grounds were sampled, with efforts to cover all seasons through an annual cycle. Zones were grouped considering oceanographic conditions associated with the Meninea Constriction and the influence of the Aysén Fjord; in this sense, the study areas were divided into the west (n = 4), east (n = 1), and south zones (n = 5, see polygons in Figure 1).
In the study area, the fishery was monitored in the three zones based on seasonal surveys (Table 1). Due to weather conditions, the southern zone could not be sampled during the winter season. Sampling was planned daily according to fishers’ activities since the assessment was made on board fishing boats using their traps as the sample unit. Traps are the only fishing gear legally allowed for crabs (Subpesca, 2013). Traps used by the fishers were truncated cone-shaped (Nasa type) and composed of three metal rings, one in the bottom, another in the middle, and a third upper ring to which a plastic entrance tunnel is attached; the diameter of the entrance varied between 25 and 35 cm (Figure 2). Most traps had escape rings with a diameter approximately of 8 cm. Traps were individually set on the seabed at a distance greater than 50 m between them. Therefore, it is assumed that there is no overlap between the effective fishing areas (Aedo and Arancibia, 2003). For each trap, the catch was recorded by the number of individuals, categorized into retained [carapace width (CW), ≥120 mm] and released (CW < 120 mm) [following the minimum legal size (MLS) of extraction, established in 120-mm CW; D. Ext. No. 9 of 1990 establishes MLS)]. This procedure allowed the estimation of catch per unit effort (CPUE) in a number of individuals. During each fishing trip, three or four traps were randomly selected to collect enough individuals (between 25 and 281 individuals per trap) to allow biological sampling, registering crab size, weight, and sex (Table 2).
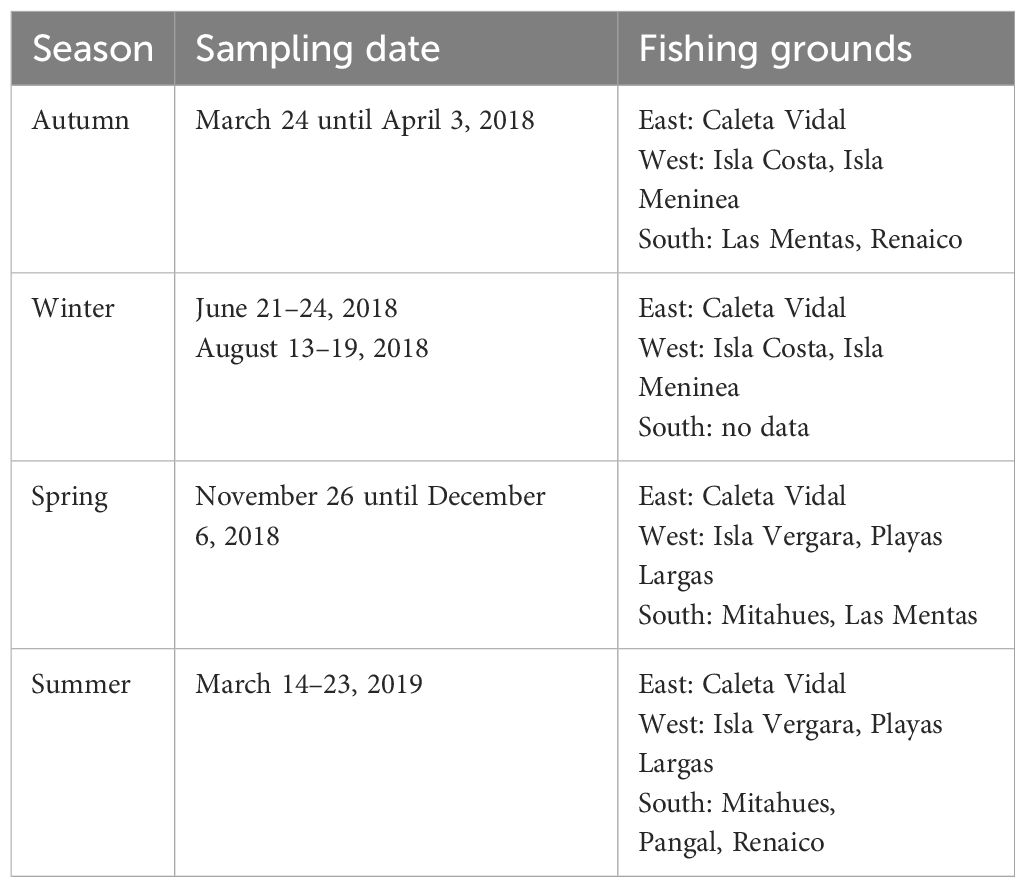
Table 1 Seasonal sampling in the different fishing zones, Aysén Region.

Figure 2 Boats and fishing gear commonly used by fishers to catch marble crabs in Aysén Region.
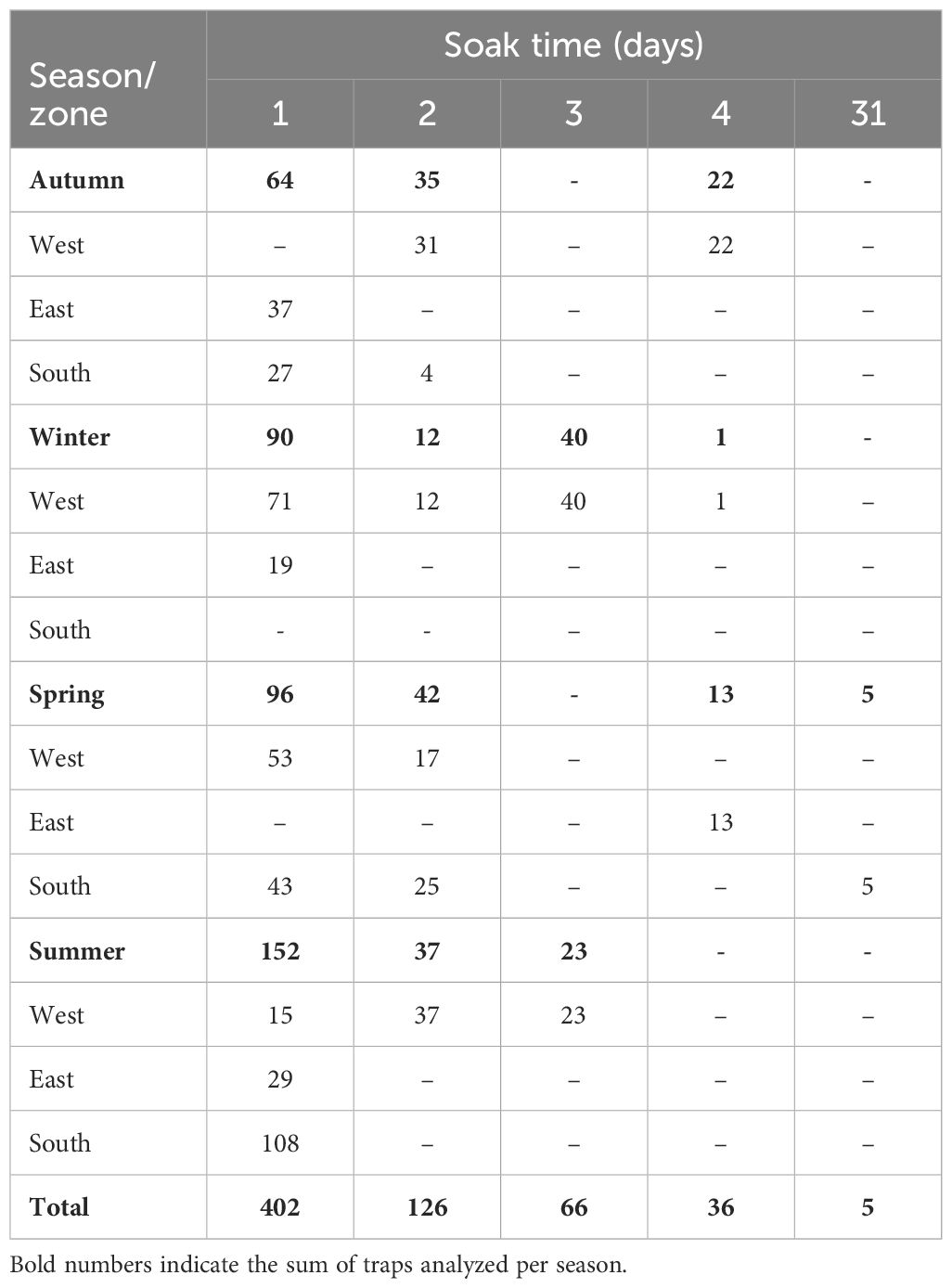
Table 2 Number of traps analyzed as a function of soak time.
Since 80% of the traps used in the extraction of marble crab have lateral escape rings, soak time could have influenced escape rates and hence CPUE. Consequently, the effect of soak time on released and retained CPUE was analyzed separately.
Seasonal and zone effects were analyzed based on five indicators: i) released and retained CPUE, ii) proportional stock density (PSD), iii) largest 10% of the sample (Lmax), iv) sex ratio, and v) length–weight relationship. PSD relates the number of individuals in a specific size class to the total number of individuals collected. The limit size adopted for the estimation of PSD was the MLS, and therefore, PSD estimates the percentage of individuals whose size exceeds this MLS. This indicator had been proposed by Froesse (2004), describing it as “let them spawn” since it is a measure of the percentage of mature specimens in the catch and hence can show trends of the status of a fishery. Data were also explored considering the average size of the largest 10% of the sample (Lmax) and selected as a measure that is less affected by environmental effects and recruitment variability (Miethe et al., 2016). Sex ratio was estimated as the proportion of the number of males in each trap to the total number of individuals (females + males) and was estimated using only individuals over MLS. Therefore, a sex ratio greater than 0.5 indicates an increased number of males compared to females. Length–weight relationship was calculated using the following formula:, where TW is body wet weight of crab in g, CW is carapace width in mm, and “a” and “b” are intercept and slope of equation, respectively. The length–weight relationship was transformed into a linear model by taking the natural logarithms of both sides of the equation (Patil and Patil, 2012), making the errors additive, and stabilizing the variances in the model. To test the isometric growth hypothesis (b = 3), a t-test was used:, where b^, and df (degrees of freedom, df = n − 2) are from the linear regression, and b0 is the specified value in the H0 (Maity, 2018). To compare the slopes of the regressions, first, the coefficients of the slope were estimated using the nlme package (Pinheiro et al., 2018), and then the slopes were compared using least square means (lsmeans; Lenth, 2016).
Non-parametric ANOVA with permutations (using the function aovp from lmPerm package; Wheeler and Torchiano, 2016) was used to search for differences in CPUE (released and retained) among fishing zones and seasons since the data were not normally distributed (Kolmogorov–Smirnov test, p < 0.05) and presented heterogeneity of variance (Levene’s test, p < 0.05). Post-hoc tests were performed using permutations pairwise comparisons (rcompanion package; Mangiafico, 2019). ANOVA test was used, with Tukey’s test as post-hoc analyses, for Lmax, PSD, and soak time since they presented a normal distribution and homogeneity of variance. A generalized linear model (GLM) test, assuming a quasi-binomial distribution of errors, was used to assess the sex ratio discrepancy between zones and seasons (Wilson and Hardy, 2002).
All calculations and analyses were conducted in the R programming environment (R Core Team, 2023).
The number of traps varied between boats from 26 and up to 37, depending on whether they had davits or not. A boat without a davit works with fewer traps compared to a larger boat with a davit that operates with two fishers and more traps. During the field trips, it was noticed that under good weather conditions and sufficient bait availability, fishers went daily through all their traps. In these cases, the soak time was typically 1 day. However, in instances of adverse weather conditions or limited bait availability, the soak time could extend beyond this period. The maximum soak time recorded was 31 days. Throughout the study period, all traps employed the same bait, consisting of 3 to 4 kg of Chilean mussels (Mytilus chilensis).
The seasonal surveys between autumn 2018 and summer 2019 resulted in 632 independent and validated records associated with catch per unit effort (CPUE). The soak time of these traps varied between 1 day (n = 402), 2 days (n = 126), 3 days (n = 63), 4 days (n = 36), and 31 days (n = 5).
Retained catch (CPUE ≥ 120 mm) did not show significant differences between 1 and 2 days, nor between 3 and 4 days of soak time (Figure 3A; Tukey’s test, p < 0.001). However, significant differences were detected at soak times of 1–2 days compared to 3–4 days (Tukey’s test; p < 0.001). The catch retained with 31 days of soak time was significantly different only from that recorded with 3–4 days (Tukey’s test; p < 0.05).
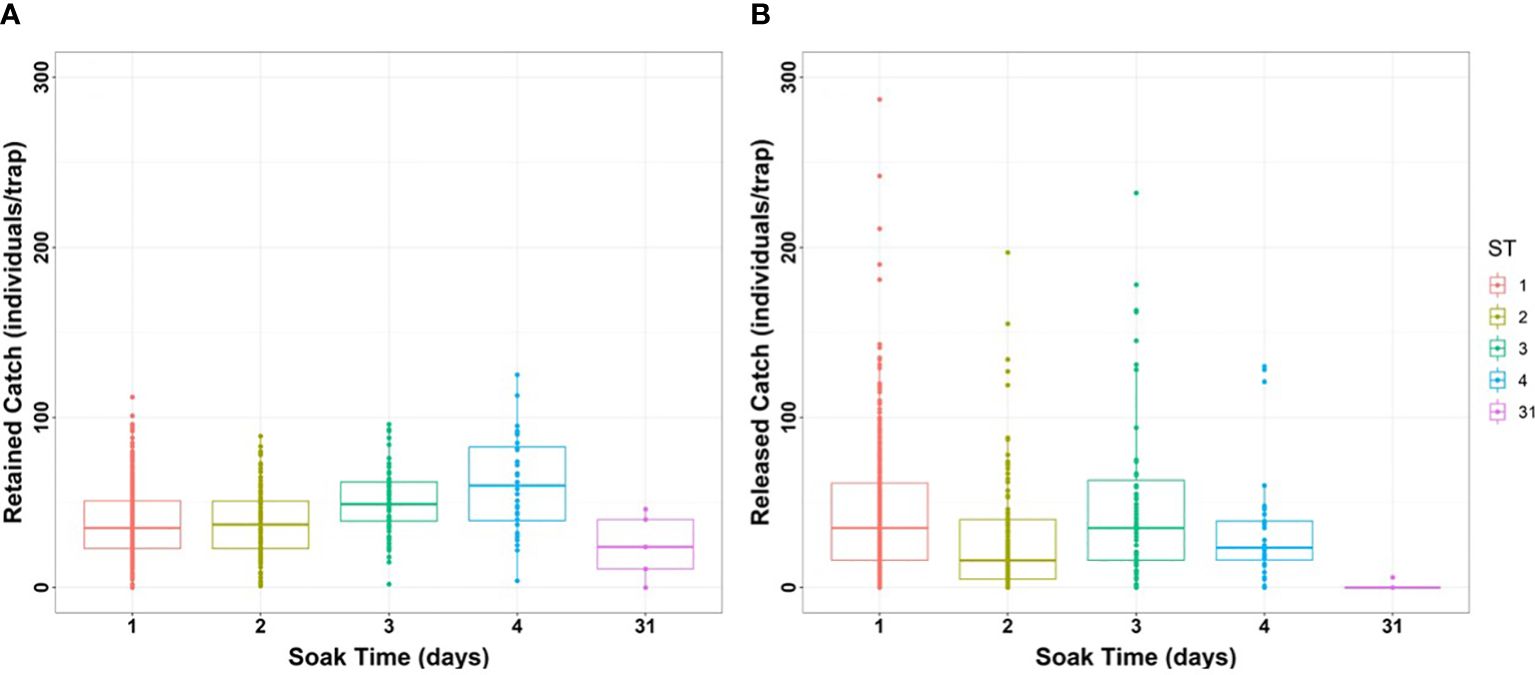
Figure 3 (A) Retained and (B) released catch (individuals/trap) of marble crab as a function of soak time (days).
Released catch revealed significant differences at soak times of 1 and 2 days and 2 and 3 days (Figure 3B; Tukey’s test; p < 0.001). For all other possible comparisons, no significant differences were found (Tukey’s test; p > 0.05).
The influence of soak time on retained and released catch enabled the selection of the most suitable datasets for analyzing spatial (fishing zones) and temporal (seasons) factors concerning various biological and fishery indicators. The analysis for retained CPUE and sex ratio considered catches with 1 and 2 days of soak time (n = 528). In contrast, released CPUE and most other biological indicators were analyzed only with 1 day of soak time (n = 402, Table 2). For the analysis of the length–weight relationship, the information corresponding to all soak times was considered (n = 632) since the relationship between both metric variables is not affected by the rates of entry and escape to the trap.
For retained catch (considering 1 and 2 days of soak time), the highest size frequencies were between 120- and 130-mm CW (51%) even though maximum sizes reached 190-mm CW.
Seasonal variability was evident across all zones (Figure 4A). In the eastern zone, the retained catch during autumn (average 53 ± SE 2) was higher than in winter (average 41 ± SE 4) and summer (average 43 ± SE 4) (pairwise permutation test; p < 0.05). Conversely, in the southern zone, there were no significant differences between autumn and summer (average 34 ± SE 2 and average 37 ± SE 2, respectively) (pairwise permutation test; p > 0.1). Additionally, in both seasons, the retained catch was higher than in spring (average 24 ± SE 2) (pairwise permutation test; p < 0.001). In the western zone, retained catch in autumn (average 47 ± SE 4) and winter (average 50 ± SE 2) was similar (pairwise permutation test; p > 0.05), being higher in both seasons compared to summer (average 38 ± SE 2) and spring (average 26 ± SE 1) (pairwise permutation test; p < 0.001). Retained catch in spring was the lowest of all seasons. Data for all zones were available only for summer and autumn; in summer, no differences were found between zones (aovp test; p > 0.1), but in autumn, the retained catch was lower in the south (average 34 ± SE 2) compared to the east (average 53 ± SE 2) and west zones (average 47 ± SE 4) (pairwise permutation test; p < 0.001). Retained catch in these latter zones was similar, presenting no significant differences (pairwise permutation test; p > 0.1).
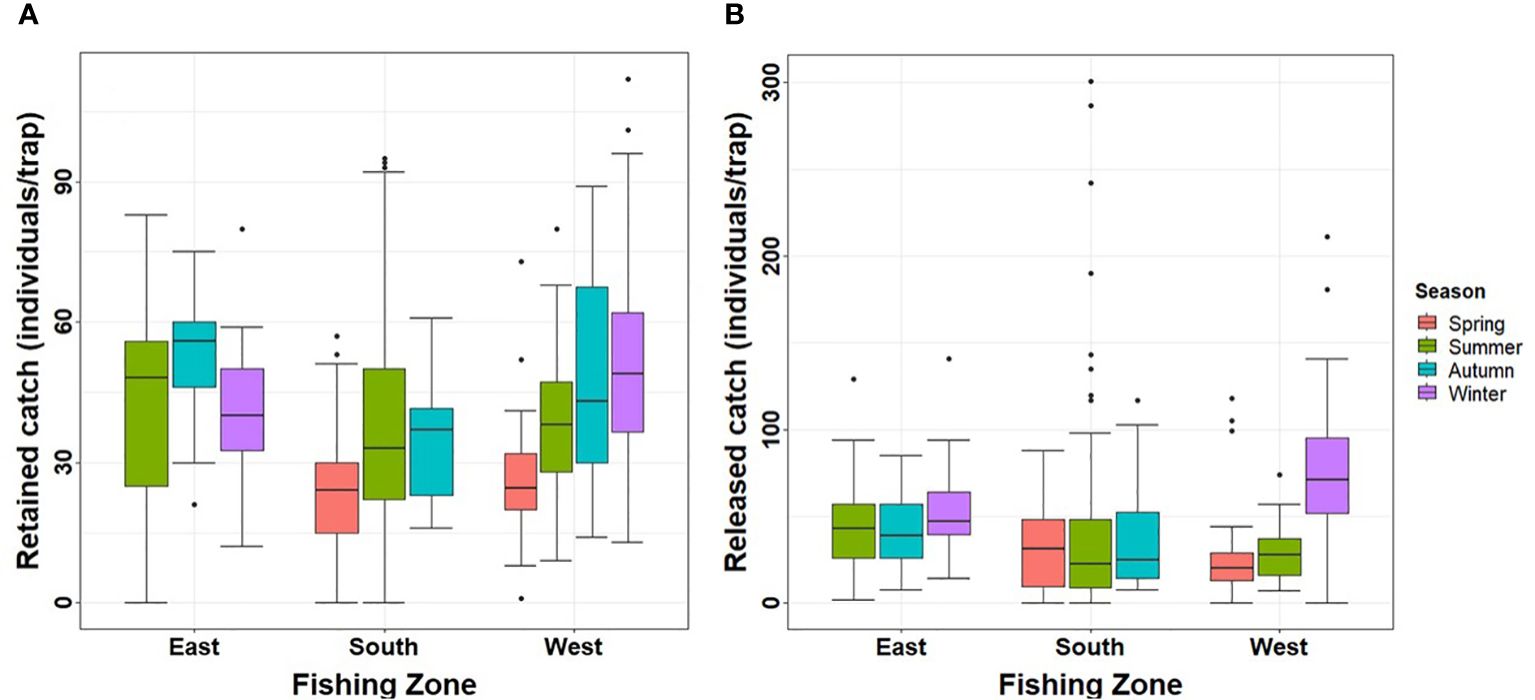
Figure 4 (A) Retained catch and (B) released catch (individuals/trap) estimated in three fishing zones during an annual cycle in Aysén Region.
For total released catch (considering 1 day of soak time), the highest size frequencies were found between 109- and 119-mm CW (72%), but individuals as small as 54-mm CW were retained in the traps. The only zone exhibiting seasonal variation in released catch was the west (Figure 4B), with winter showing higher values (average 77 ± SE 4) compared to the other seasons, namely, spring and summer (pairwise permutation test; p < 0.01). Data for all zones were available only for summer. When analyzing this season, no significant differences in the released catch were found between zones (pairwise permutation test; p > 0.05).
PSD (as a percentage of crabs ≥ MLS) could not be estimated for all seasons in the zones because the data considered only 1 day of soak time. However, in the east and west zones, PSD was minimal in winter, increasing toward spring and summer to a maximum in autumn (Figure 5). In the eastern zone, PSD in autumn was higher (average 60 ± SE 3) than in summer (average 52 ± SE 3; pairwise permutation test, p < 0.05) and winter (average 44 ± SE 3; pairwise permutation test, p < 0.001). In the western zone, PSD could not be estimated in autumn, but spring (average 54 ± SE 2) and summer (average 59 ± SE 4) presented a higher PSD compared to winter (average 40 ± SE 2; pairwise permutation test, p < 0.001). Conversely, in the south, no significant differences were found between seasons (ANOVA test; p > 0.1), although winter could not be sampled. In summer (where all zones were sampled), no significant differences were found (ANOVA test; p > 0.1).
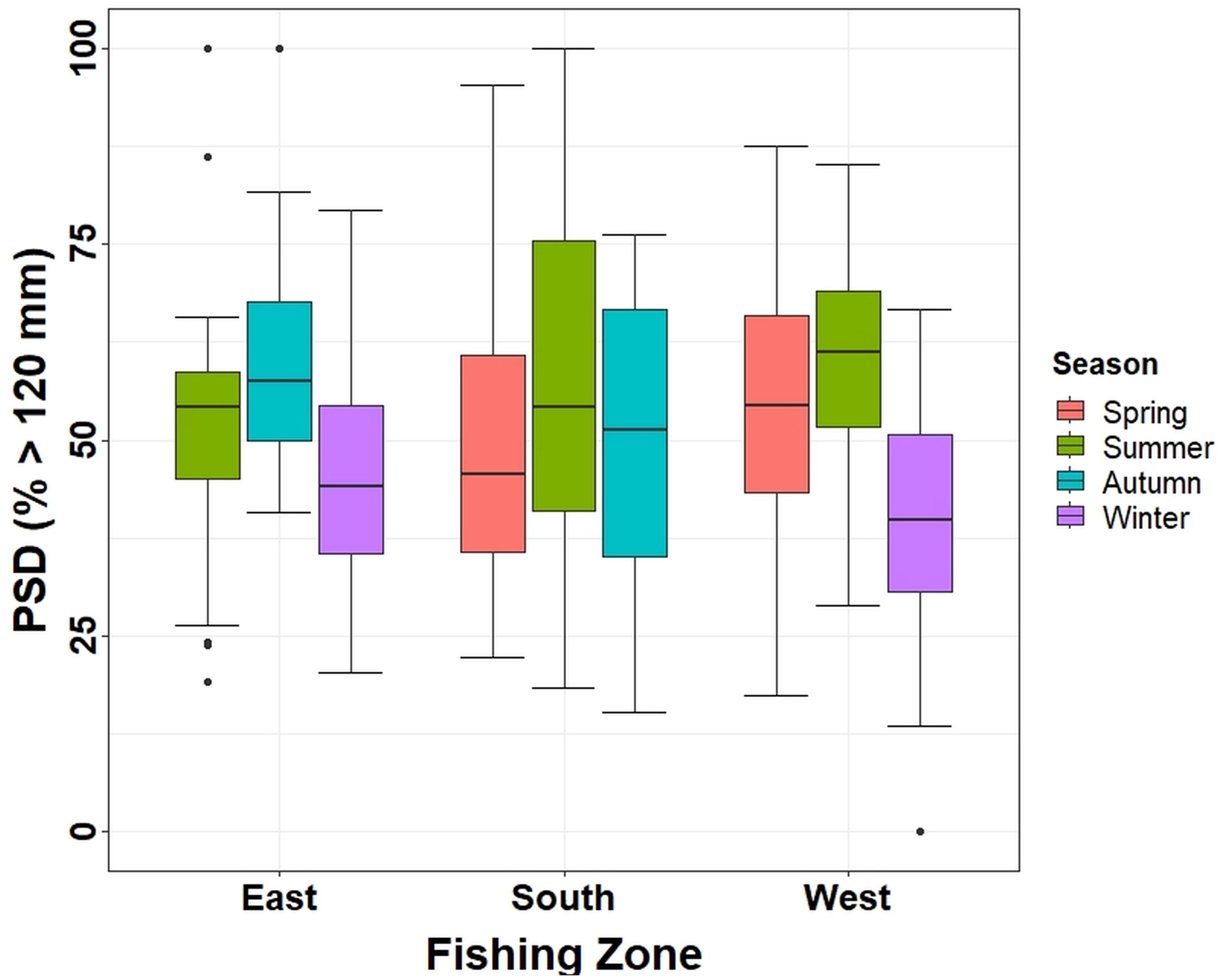
Figure 5 PSD (expressed as average percentage of individuals over 120 mm) estimated in the three fishing zones during an annual cycle in Aysén Region. PSD, proportional stock density.
Lmax (top 10%) were similar between seasons in the same zone (ANOVA test; p > 0.1) and were also similar in summer when all zones could be sampled (ANOVA test; p > 0.1). Lmax varied on average between 136- and 147-mm CW, having a minimum value of 110-mm CW and a maximum of 170-mm CW. Lmax were different between sexes (ANOVA test; p < 0.001), as males were larger (average 149 ± SE 1) compared to females (average 135 ± SE 1) (Figures 6A, B).
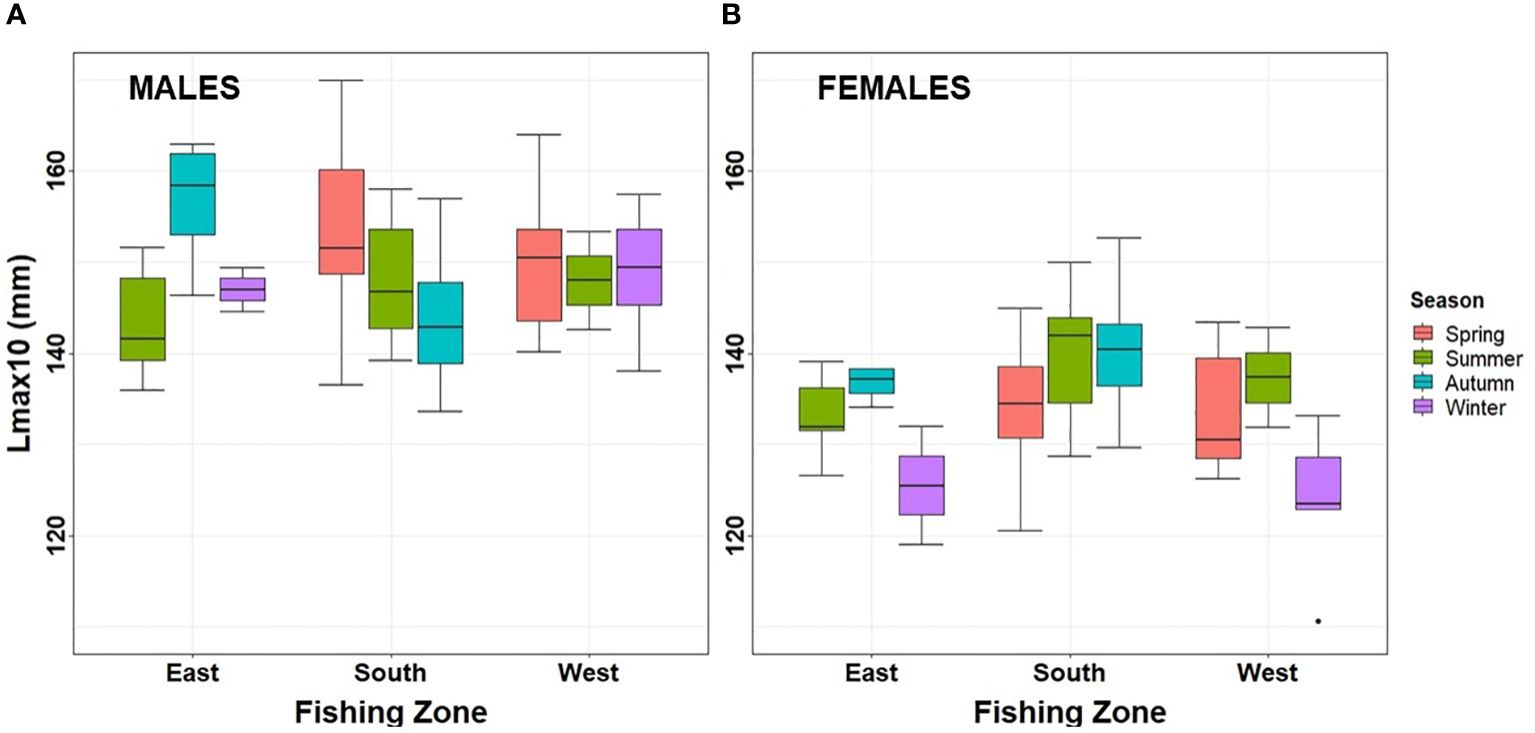
Figure 6 Lmax of (A) males and (B) females estimated in the three fishing zones during an annual cycle in Aysén Region. Lmax, largest 10% of the sample.
The sex ratio was analyzed based on individuals exceeding the MLS using both 1 and 2 days of soak time. For most seasons and zones, males dominated this fraction of the stock (Figure 7). Generally, males were predominant during winter (average 0.84 ± SE 0.05) compared to the other seasons. In contrast, in autumn in the west and south zones, females dominated the catch (average 0.34 ± SE 0.08), being statistically different from spring and summer in the south (GLM test; p < 0.05; Supplementary Material; Table S1) and spring and winter in the west (GLM test; p < 0.05). In summer and autumn when there were enough data to compare all zones, no significant differences were found during summer between zones (GLM test; p > 0.1), but in autumn, significant differences were found between the east zone (average 0.59 ± SE 0.03) and the south and west zones (GLM test; p < 0.05; Supplementary Material; Table S1).
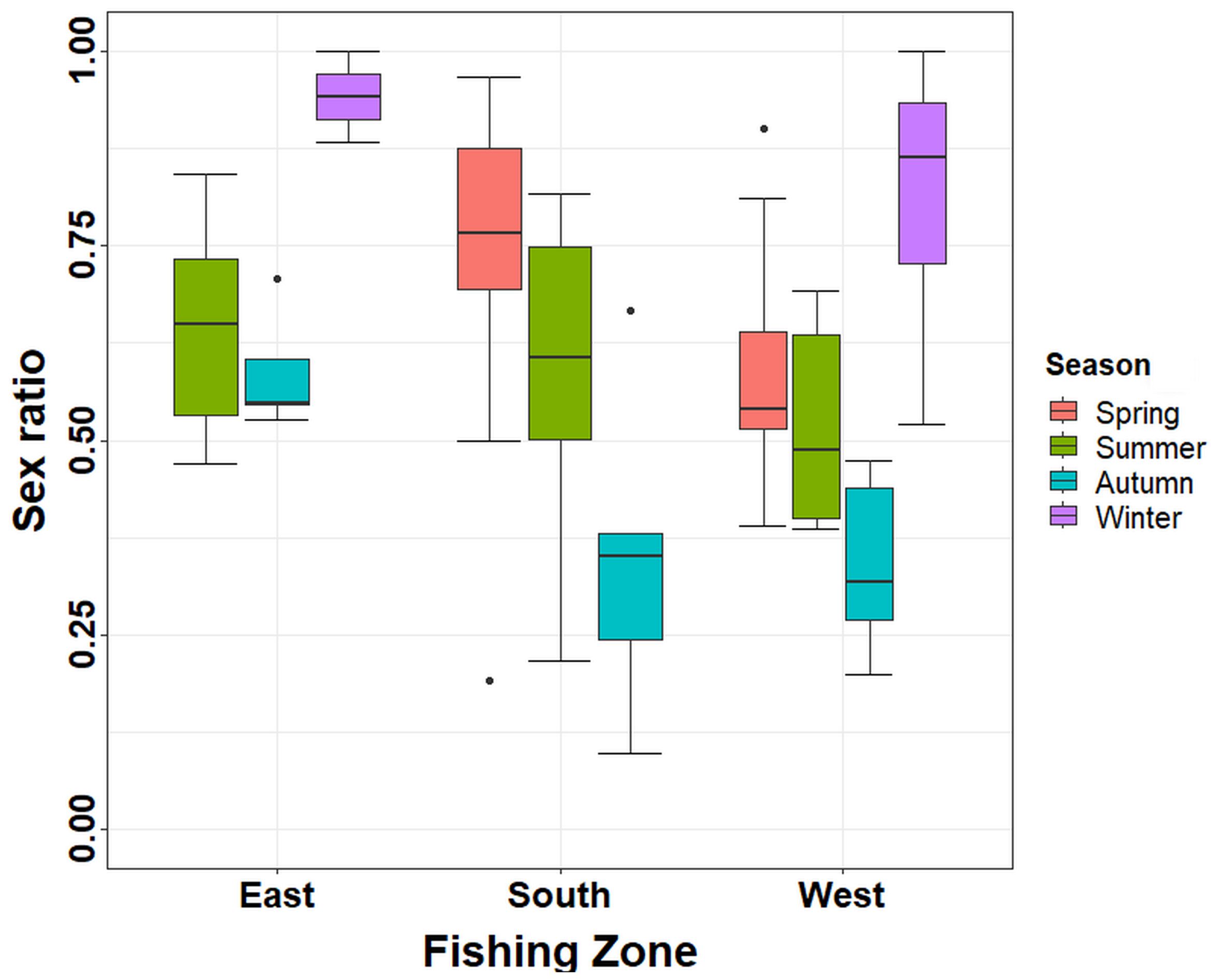
Figure 7 Sex ratio (expressed as average with standard error) of individuals over 120-mm CW estimated in the three fishing zones during an annual cycle in Aysén Region. CW, carapace width.
The sample sizes for the length–weight relationship varied between 591 and 2,033 individuals for the spatial analysis (zones) and between 515 and 1,361 individuals for the temporal analysis (seasons). For each sex in the different fishing zones and seasons, after the log transformation of data, the adjustment of the length–weight relationship was significant (Student’s t-test; p < 0.05) (Supplementary data, Tables S2 and S3). In terms of growth type, slope b was significantly higher than 3 in males and statistically less than 3 in females, indicating positive and negative allometric growth, respectively (Student’s t-test; p < 0.05) (Supplementary data, Tables S2 and S3). The comparison between the slopes of the regressions determined a significant effect of the zone (ANOVA test; p < 0.01) and season (ANOVA test; p < 0.001) only in the case of males, but not for females (in both factors: ANOVA test; p > 0.05). The slope b of males collected in the eastern zone was statistically lower than the estimated for the western (Tukey’s test; p < 0.05) and southern zones (Tukey’s test; p < 0.01). In addition, the slope b of males collected in spring was statistically higher than the estimated for autumn (Tukey’s test; p < 0.001), winter (Tukey’s test; p < 0.001), and summer (Tukey’s test; p < 0.01).
Our study is the first to collect data on the marble crab in the main fishing grounds of Northern Patagonia through a design implemented on board artisanal vessels during an annual cycle. Although we encountered sampling limitations since in some fishing zones data could not be obtained across seasons, most estimated indicators presented seasonal variability, and some patterns could be identified along the zones. For all zones, retained CPUE was higher during autumn compared to other seasons. In contrast, in spring, retained CPUE was the lowest. This seasonal variability could be associated with the reproductive cycle of the studied species. In autumn, trap efficiency could be enhanced by higher foraging activity related to increased energetic demand preceding spawning in females and gonadal recovery after mating in males (Pardo et al., 2013, 2015, 2020). In contrast, low retained CPUE in spring could be associated with an intensive mating period described for this species from the end of spring/beginning of summer (Pardo et al., 2013, 2016). It has been observed during this phase that male and female crabs reduce their feeding response (Skinner and Hill, 1987; Kennelly and Watkins, 1994). Other non-exclusive factors could be seasonal bathymetric migrations, described for this species in lower latitudes (Pardo et al., 2020), and stock depletion through the year (Bez et al., 2006) since the fishing period usually ends in spring.
Released CPUE was seasonal only in the western fishing zone, with the highest catch recorded during winter. Under the assumption that the catch per trap is proportional to the abundance and availability for both juveniles and adults, at least at short soak times (i.e., 1 day), an increase in the capture of individuals classified as pre-fishery recruits (dominated by individuals between sizes of 104 and 119 mm) could be associated with migratory movements (Pardo et al., 2020). In crabs, movement patterns have been mostly associated with changes in habitat use, search for refuge sites, and reproductive migration events (Hines et al., 1995; Muñoz et al., 2006; Curtis and McGaw, 2008). However, these hypotheses should be explored since in the eastern zone, no seasonal changes were observed in the released catch.
Other fishery indicators (PSD, Lmax, and sex ratio) showed seasonal variability. The main trend was associated with a lower proportion of stock catches in winter, principally by large males. This tendency was found in the east and west zones. The lack of seasonal variability in the southern zone could be explained by sampling difficulties in winter. The higher proportion of males found in winter could be explained by catch bias by sex, considering that during this period most females are carrying embryos. Since it had been demonstrated that their foraging activity is reduced during this stage (Howard, 1982), they are not easily drawn to the trap.
Our results showed that catches of crabs ≥120-mm CW were biased toward males in most of the seasons and zones. The sex ratio could get as high as 11 males per female, averaging between 2 and 3 males per female. Although the optimal sex ratio is unclear, the skewed ratio toward males and the presence of large males indicate that fishing is not adversely affecting these reproductive traits. In our study, males could be on average 14 mm larger than females, which is consistent with results found in areas of low fishing intensity (Pardo et al., 2017). Sexual dimorphism was evident not only in size but also in the relationship between size and weight. Females exhibited negative allometry, whereas males showed positive allometry, as previously described for this species (Pool et al., 1998; Olguín and Mora, 2022; Daza et al., 2023). Considering that females mate after molting, previous studies have found they preferentially mate with larger males due to the need for a pre- and post-copulatory embrace to ensure survival (Pardo et al., 2018). This mating process may also be favored by the greater weight growth in males compared to females (Waiho et al., 2015). In our study, higher weight in spring for males compared to other seasons agrees with this: during the mating period (i.e., spring), males could shift their energy investment toward growth (Sumer et al., 2013) to protect females.
Fishery stock monitoring based on the recording of variables on board artisanal vessels allows the establishment of indicators that compare the exploited (legal) and unexploited (non-legal) fractions, which in the case of the marble crab fishery corresponds to individuals below and above the minimum legal size of extraction (MLS ≥ 120 mm CW). For example, median PSD varied mostly between 40% and 60% for different fishing zones and seasons. This level of detailed fishing information provides higher resolution fishing information compared to solely monitoring retained catch from landing ports (Olguín and Mora, 2019).
Our results showed that soak time differences between traps are key to a correct estimation and, hence, comparison of biological and fishery indicators. Especially in zones where the presence of escape rings is a common practice among fishers (as in the Aysén Region), favoring after 1 day of soak time, the escape of small individuals through this mechanism. These results agreed with field observations since we could verify that bait decreases its effectiveness when soak time exceeds 1 day, causing small individuals to leave the traps through the escape rings. Literature also indicates that the capture of small individuals decreases with escape rings (Rosa-Pacheco and Ramírez-Rodríguez, 1996; Aguilar and Pizarro, 2006; Arana et al., 2011). From the operational point of view, this benefits not only the fishers hauling traps by hand (i.e., decreasing the weight of each trap) but also the juveniles since handling may affect survival. This ultimately highlights the importance of incorporating escape rings as a mandatory administrative measure.
Our results showed that the variability of fishery and biological indicators responds more to seasonal patterns than to expected heterogeneity across fishing grounds. These results allowed us to make monitoring recommendations for crab fisheries.
• Monitoring should be based on a representative and easily accessible number of fishing zones that could be monitored over time.
• Design should be oriented on annual seasonality that allows coupling between catch indicators (CPUE), size, and reproductive parameters.
• Biological and fishery indicators should be estimated with data from 1 or 2 days of soak time since the catch is affected by this variable.
• Trap design for capturing crabs must incorporate escape rings to reduce the catchability of small individuals. This not only has operational advantages but also improves the survival of non-commercial individuals.
• Monitoring on board artisanal vessels allows georeferencing with a high spatial resolution, giving an opportunity to evaluate the exploitation status of marble crabs at the scale of fishing zones (e.g., data-limited and index-based).
In conclusion, these results allow us to develop improved monitoring recommendations for this fishery in Northern Patagonia. Specifically, monitoring could be based on a more limited number of fishing grounds but should be standardized by season, soak time, and presence/absence of escape rings. Moreover, the correlation between indicator temporal variability with the reproductive cycle of this species highlights the necessity for monitoring its possible seasonal variations in parallel.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because during the study we did not sacrifice the animals, all individuals were return immediately after being weighed and measured to their marine environment trying to ensure their maximum survival.
MH: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. GA: Conceptualization, Data curation, Investigation, Methodology, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. PO: Funding acquisition, Investigation, Methodology, Project administration, Validation, Writing – review & editing. AO: Conceptualization, Validation, Writing – review & editing. LP: Conceptualization, Investigation, Methodology, Validation, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was funded by CORFO through grant BPCR 75810 “Línea base biológica-pesquera del recurso Jaiba marmola: insumos para su manejo sustentable en la Región de Aysén”. Partial funding was available through Programa Regional ANID R20F0002.
The authors would like to thank Brian Reid for the English revision of the manuscript and Nahuelquin and Legue families and Adan Mansilla for allowing us to go with them on their fishing trips and to collect samples. Luis Miguel Pardo thanks to FONDAP IDEAL 15150003.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1392758/full#supplementary-material
Aedo G., Arancibia H. (2003). Estimating the attraction area and the effective fishing area for Chilean lemon crab (Cancer porteri) using traps. Fisheries Res. 60, 267–272. doi: 10.1016/S0165-7836(02)00177-7
Aguilar M., Pizarro P. (2006). Empleo de ventanas de escape en trampas para la captura de jaiba peluda (Cancer setosus) en Iquique. Chile. Investigaciones Marinas Valparaíso 34, 63–70. doi: 10.4067/S0717-71782006000200007
Aracena C., Lange C. B., Iriarte J. L., Rebolledo L., Pantoja S. (2011). Latitudinal patterns of export production recorded in surface sediments of the Chilean Patagonian fjords (41–55°S) as a response to water column productivity. Continental Shelf Res. 31, 340–355. doi: 10.1016/j.csr.2010.08.008
Arana P., Orellana J. C., De Caso A. (2011). Escape vents and trap selectivity in the fishery for the Juan Fernández rock lobster (Jasus frontalis), Chile. Fisheries Res. 110, 1–9. doi: 10.1016/j.fishres.2011.03.021
Bell T. (1835). Observations on the genus Cancer of Dr. Leach (Platycarcinos Latr.), with descriptions of three new species. Proceedings of the Zoological Society of London 1835, 86–88.
Bez N., De Oliveira E., Duhamel G. (2006). Repetitive fishing, local depletion, and fishing efficiencies in the Kerguelen Islands fisheries. ICES J. Mar. Sci. 63, 532–542. doi: 10.1016/j.icesjms.2005.10.005
Béné C. (2006). Small-scale fisheries: assessing their contribution to rural livelihoods in developing countries. FAO Fisheries Circular No. 1008 (Rome: FAO), 46.
Collie J. S., Gislason H. (2001). Biological reference points for fish stocks in a multispecies context. Can. J. Fisheries Aquat. Sci. 58, 2167–2176. doi: 10.1139/cjfas-58-11-2167
Curtis D. L., McGaw I. J. (2008). A year in the life of a Dungeness crab: methodology for determining microhabitat conditions experienced by large decapod crustaceans in estuaries. J. Zoology 274, 375–385. doi: 10.1111/j.1469-7998.2007.00397.x
Daza E., Olguín A., Almonacid E., Mora P., Pacheco H. (2023). Programa de Seguimiento de las Principales Pesquerías Nacionales, año 2022: Pesquerías: Crustáceos Bentónicos. Informe Final IFOP-Ministerio Economía, 391.
Froesse R. (2004). Keep it simple: three indicators to deal with overfishing. Fish Fisheries 5, 86–91. doi: 10.1111/j.1467-2979.2004.00144.x
Guzman D. (2004). “Caracterización hidrográfica, oceanográfica y balance de nitrógeno y fosforo del fiordo Aysén. Tesis para obtener el grado de magister en oceanografía,” in Facultad de recursos naturales (Pontificia Universidad Católica de Valparaíso, Valparaíso. Chile).
Hines A. H., Wolcott T. G., González-Gurriarán E., González-Escalante J. L., Freire J. (1995). Movement patterns and migrations in crabs: telemetry of juvenile and adult behavior in Callinectes sapidus and Maja squinado. J. Mar. Biol. Assoc. United Kingdom 75, 27–42. doi: 10.1017/S0025315400015174
Howard A. E. (1982). The distribution and behaviour of ovigerous edible crabs (Cancer pagurus), and consequent sampling bias. J. duConseil Int. pour l’Exploration la Mer 40, 259–261. doi: 10.1093/icesjms/40.3.259
Kennelly S. J., Watkins D. (1994). Fecundity and reproductive period, and their relationship to catch rates of spanner crabs, Ranina ranina, off the east coast of Australia. J. Crustacean Biol. 14, 146–150. doi: 10.1163/193724094X00533
Lenth R. V. (2016). Least-squares means: the R package lsmeans. J. Stat. Software 69, 1–33. doi: 10.18637/jss.v069.i01
Maity R. (2018). Statistical methods in hydrology and hydroclimatology. (Singapure: Springer), 444. doi: 10.1007/978-981-10-8779-0
Mangiafico S. (2019). Functions to support extension education program evaluation (R Package ‘rcompanion’ Version 2.1.1). Available online at: https://rcompanion.org/handbook/G_14.html.
Miethe T., Dobby H., McLay A. (2016). The use of indicators for shellfish stocks and fisheries: A literature review. Scottish Mar. Freshw. Sci. 7, 78. doi: 10.7489/1764-1
Montero P., Daneri G., González H. E., Iriarte J. L., Tapia F. J., Lizárraga L., et al. (2011). Seasonal variability of primary production in a fjord ecosystem of the Chilean Patagonia: implications for the transfer of carbon within pelagic food webs. Continental Shelf Res. 31, 202–215. doi: 10.1016/j.csr.2010.09.003
Morgan M. J., Shelton P. A., Rideout R. M. (2014). An evaluation of fishing mortality reference points under varying levels of population productivity in three Atlantic cod (Gadus morhua) stocks. ICES J. Mar. Sci. 71, 1407–1416. doi: 10.1093/icesjms/fsu092
Muñoz C., Pardo L. M., Henriquez L., Palma A. (2006). Variaciones temporales en la composición y abundancia de cuatro especies de Cancer (Decapoda: Brachyura: Cancridae) capturadas con trampas en bahía San Vicente, Concepción (Chile central). Investigaciones Marinas Valparaíso 34, 9–21. doi: 10.4067/S0717-71782006000200002
Olguín A., Mora P. (2019). “Seguimiento General de Pesquerías: Crustáceos Bentónicos: Jaibas y Centolla X y XI Regione,” in IFOP, Ministerio de economía Fomento y Turismo, Valparaíso, Chile, vol. 149.
Olguín A., Mora P. (2022). “Seguimiento General de Pesquerías: Crustáceos Bentónicos: Jaibas y Centolla, Región de Los Lagos y Región de Aysén. 2021,” in IFOP, Ministerio de economía Fomento y Turismo, Valparaíso, Chile, vol. 171.
Pardo L. M., Riveros M., Chaparro O., Pretterebner K. (2018). . Ejaculate allocation in brachyura: what do males of Metacarcinusedwardsii respond to? Aquat. Biol. 27, 25–33. doi: 10.3354/ab00693
Pardo L. M., Riveros M., Fuentes J. P., Lopez-Greco L. (2013). Functional morphology of the seminal receptacle in the crab Metacarcinus edwardsii. Invertebrate Biol. 132, 386–393. doi: 10.1111/ivb.12038
Pardo L. M., Riveros M. P., Fuentes J., Pinochet R., Cárdenas C., Sainte-Marie B. (2017). High fishing intensity reduces females’ sperm reserve and brood fecundity in a eubrachyuran crab subject to sex- and size-biased harvest. ICES J. Mar. Sci. 74, 2459–2469. doi: 10.1093/icesjms/fsx077
Pardo L. M., Riveros M. P., Fuentes J. P., Rojas-Hernandez N., Veliz D. (2016). An effective sperm competition avoidance strategy in crabs drives genetic monogamy despite evidence of polyandry. Behav. Ecol. Sociobiology 70: 1, 73–81. doi: 10.1007/s00265-015-2026-6
Pardo L. M., Rosas Y., Fuentes J. P., Riveros M. P., Chaparro R. C. (2015). Fishery induces sperm depletion and reduction in male reproductive potential for crab species under male-biased harvest strategy. PloS One 10, e0115525. doi: 10.1371/journal.pone.0115525
Pardo L. M., Rubilar P. R., Fuentes J. P. (2020). North Patagonian estuaries appear to function as nursery habitats for marble crab (Metacarcinus edwardsii). Regional Stud. Mar. Science. 36, 101315. doi: 10.1016/j.rsma.2020.101315
Patil K. M., Patil M. U. (2012). Length-weight relationship and condition factor of freshwater crab Barytelphusagurini (Decapoda, Brachyura). J. Exp. Sci. 3, 13–15. doi: 10.3170/ajas.5.1.14902
Pinheiro J., Bates D., DebRoy S., Sarkar D. (2018). R Development Core Team; nlme: Linear and Nonlinear Mixed Effects Models. Rpackage version 3.1-137. Available online at: http://CRAN.R-project.org/package=nlme.
Pool H., Montenegro C., Canales C., Barahona N., Vicencio C. (1998). “Análisis de la Pesquería de jaibas en la X Región,” in proyecto FIP, vol. 76. , 96–35.
R Core Team (2023). R: A language and environment for statistical computing (Vienna: R Foundation for Statistical Computing). Available at: https://www.R-project.org.
Rosa-Pacheco R. D. L., Ramírez-Rodríguez M. (1996). Escape vents in traps for the fishery of the California spiny lobster, Panulirus interruptus, in Baja California Sur, Mexico. Cienc. Mar. Baja Calif. Mexico 22, 235–243. doi: 10.7773/cm.v22i2.849
Sernapesca (2022).Anuario Estadístico de pesca 2010-2022. Ministerio de Economía, Fomento y Reconstrucción. Sección Desembarques artesanales y Acuicultura. In: Servicio Nacional de Pesca y Acuicultura. Available online at: www.sernapesca.cl (Accessed September 10, 2023).
Silva N., Guzmán D., Valdenegro A. (2000). “Aysén sound, Chile,” in Estuarine system of the South American region: carbon, nitrogen and phosphorus fluxes. LOICZ Reports and Studies 15, Texel. Eds. Smith S. V., Dupra V., Crossland J. I. M., Crossland C. J. (The Netherlands), 55–64.
Silva N., Siervers H., Prado. R. (1995). Características oceanográficas y una proposición de circulación para algunos canales australes de Chile (41° 20’S, 46° 40’S). Rev. Biología Marina 30, 207–254.
Skinner D. G., Hill B. J. (1987). Feeding and reproductive behaviour and their effect on catchability of the spanner crab Ranina ranina. Mar. Biol. 94, 211–218. doi: 10.1007/BF00392933
Subpesca (2013). “Nómina nacional de pesquerías artesanales. Res. Ext. N° 3115, nov 2013,” in Ministerio de Economía, Fomento y Turismo, Valparaíso, Chile.
Subpesca (2022).Estado de situación de las principales pesquerías Chilenas. Año 2020. In: Departamento de Pesquerías. División de Administración Pesquera (Chile: Valparaíso). Available online at: https://www.subpesca.cl/portal/618/w3-article-117812.html (Accessed October 4, 2023).
Sumer C., Teksam I., Karatas H., Beyhan T., Aydin C. M. (2013). Growth and reproduction biology of the blue crab, callinectes sapidus rathbun 1896, in the beymelek lagoon (Southwestern coast of Turkey). Turkish J. Fisheries Aquat. Sci. 13, 675–684. doi: 10.4194/1303-2712-v13_4_13
Tsikliras A. C., Froese R. (2018). Maximum sustainable yield. 2nd ed. Ed. Fath B. (Amsterdam, Holanda: Elsevier), 1–20.
Waiho K., Mustaqim M., Fazhan H., Wan Norfaizza W. I., Megat F. H., Ikhwanuddin M. (2015). Mating behaviour of the orange mud crab, Scylla olivacea: The effect of sex ratio and stocking density on mating success. Aquaculture Rep. 2, 50–57. doi: 10.1016/j.aqrep.2015.08.004
Wilson K., Hardy I. C. W. (2002). Sex ratios concepts and research methods (Cambridge University Press Print), 48–92. doi: 10.1017/CBO9780511542053.004
Wheeler B., Torchiano M. (2016). lmPerm: permutation tests for linear models, version: 2.1.0. https://CRAN.R-project.org/package=lmPerm.
Keywords: M. edwardsii, monitoring, data-limited fishery, North Patagonia, small-scale fishery
Citation: Hamamé M, Aedo G, Ortiz P, Olguín A and Pardo LM (2024) Biological and fishery indicators for the small-scale marble crab fishery in Northern Patagonia: recommendations for improving a monitoring program and stock assessment of a data-limited fishery. Front. Mar. Sci. 11:1392758. doi: 10.3389/fmars.2024.1392758
Received: 28 February 2024; Accepted: 26 June 2024;
Published: 17 July 2024.
Edited by:
Kareen E. Schnabel, National Institute of Water and Atmospheric Research (NIWA), New ZealandReviewed by:
Ajit Kumar Mohanty, Indira Gandhi Centre for Atomic Research (IGCAR), IndiaCopyright © 2024 Hamamé, Aedo, Ortiz, Olguín and Pardo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Madeleine Hamamé, bWhhbWFtZUBjaWVwLmNs
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.