 Michael C. James
Michael C. James Kelly E. Hall
Kelly E. Hall Emily P. Bond
Emily P. Bond Scott Sherrill-Mix
Scott Sherrill-Mix Virginie Plot
Virginie Plot- 1Population Ecology Division, Fisheries and Oceans Canada, Bedford Institute of Oceanography, Dartmouth, NS, Canada
- 2Department of Microbiology, Genetics and Immunology, Michigan State University, East Lansing, MI, United States
- 3Groupe Local d'Observation et d'Identification des Cétacés Réunion, Saint-Paul, Réunion Island, France
Estimating mortality of sea turtles incidentally captured in fishing gear is essential to understanding fishery impacts on sea turtle populations. Loggerhead turtles (Caretta caretta) are a component of bycatch in pelagic longline fisheries, including those operating in Atlantic Canada. In this study, we used pop-up satellite archival tags (PSATs) attached to 62 loggerhead turtles incidentally hooked in Atlantic Canadian shallow-set pelagic longline gear (2012–2018) to estimate associated post-release survival. Consistent with conventional fishing practices, hooks were not removed from turtles prior to release. Analysis of diving behaviour, ocean temperature, and ambient light level was used to assign fates to turtles. Post-release survival was calculated using the Kaplan-Meier estimator with right censoring. Based on this method, the annual survival probability of an individual loggerhead post-release is 0.877 ± SE 0.052. While it is commonly assumed that hook location is an important variable in predicting fates of turtles after release, there was no difference in survival of deep-hooked (swallowed hook) versus shallow-hooked turtles. These results indicate higher survivorship among hooked loggerheads captured in shallow-set pelagic longline gear than prevailing wisdom and suggest that at least some of the population impacts on loggerhead turtles conventionally attributed to pelagic longline interactions may be linked to other threats.
1 Introduction
The ranges of many marine mammals, sharks, seabirds and sea turtles span broad oceanographic regions and overlap with areas of intense fishing activity (Johnson et al., 2005; Wallace et al., 2010; Bjørge et al., 2013; Sulikowski et al., 2020; Frankish et al., 2021). As a result, these species are vulnerable to incidental entanglement or hooking (collectively known as bycatch) in recreational, artisanal and commercial fishing gear. Such interactions represent a widespread threat and leading cause of population decline (Spotila et al., 2000; Lewison et al., 2004a, 2004b).
While much bycatch is discovered dead at the time of release, in some fisheries various species are regularly released alive (Carruthers et al., 2009; Gray and Kennelly, 2018). Given the scale of global bycatch, and the logistical challenges and cost associated with evaluating fates of incidentally caught and live released animals, post-release survival studies have focused on a relatively small number of species in only a fraction of fisheries. Much related research attention has been directed towards sharks (Eddy et al., 2016; Musyl and Gilman, 2018; Schaefer et al., 2019; Sulikowski et al., 2020) and other large commercially-valuable fish (Fenton et al., 2015; Tracey et al., 2016; Ferter et al., 2017; Jackson et al., 2018; Nielsen et al., 2018; Orbesen et al., 2018; Strøm et al., 2019). Although estimating post-release survival is also essential to understanding how bycatch may impact sea turtle populations, markedly few corresponding studies exist (Sasso and Epperly, 2007; Àlvarez de Quevedo et al., 2013; Swimmer et al., 2014).
Fishing activities overlap with key habitats used by sea turtles, including foraging areas, migration corridors and waters adjacent nesting beaches (Lewison et al., 2013). Around 85 000 sea turtles are reported as bycatch discard (live or dead at release) each year, but the true total could be two orders of magnitude greater than this (Wallace et al., 2010). For instance, it was estimated that in one year, 200 000 loggerhead turtles (Caretta caretta) were likely captured in pelagic longline fisheries alone (Lewison et al., 2004b).
To help inform bycatch management, studies estimating sea turtle bycatch rates have described the overlap between fishing areas and turtle distributions to better understand how and when turtles are most likely to interact with fisheries (Lewison et al., 2013). Differences in bycatch rates can reflect environmental factors, gear types, and fishing methods, including set depths, soak times, and hook and bait types (Watson et al., 2005; Gilman et al., 2007; Howell et al., 2008; Epperly et al., 2012). These same variables, which cannot always be controlled in post-release mortality studies, may affect survival outcomes of incidentally captured turtles. To assess population-level impacts on bycatch species, population demographics, as well as direct bycatch mortality and post-release survival must be considered (Lewison et al., 2013). In the absence of post-release survival data, true bycatch mortality may be greatly underestimated (Molina and Cooke, 2012).
Complex and confounding variables such as physiological stress, reduced feeding, or infections from retained gear (Innis et al., 2010; Cassoff et al., 2011; Phillips et al., 2015) can result in mortality events in the hours, days, or weeks following release. When turtle mortality is delayed following fishery interaction, it becomes difficult to attribute death to injury versus natural causes, and this uncertainty increases with time (Benoît et al., 2020). Additionally, confirmation of sea turtle fates through direct observation in the wild is not possible. Some studies have collected sea turtles following incidental capture in fisheries and have observed outcomes in aquaria (Rudloe and Rudloe, 2005; Casale et al., 2008). Although handling and captive husbandry introduce multiple confounding variables, Caracappa et al. (2018) reported that 22% of a sample of stranded loggerhead turtles presenting with ingested hooks in the mouth or proximal esophagus were successfully rehabilitated.
Biotelemetry allows for remote observation of turtles following fishery interactions. Satellite-linked platform transmitting terminals (PTTs) have been used to track turtles following capture in fishing gear (Bond and James, 2021), with mortality inferred from cessation of instrument transmission (Hays et al., 2003; Chaloupka et al., 2004; Snoddy and Williard, 2010; Mangel et al., 2011). However, transmission failure may be attributed to multiple causes, including electronic malfunction, battery depletion, biofouling and tag detachment (Hays et al., 2007, 2021), thus this approach is likely to overestimate absolute levels of mortality (Hays et al., 2003). Pop-up satellite archival tags (PSATs) archive depth, temperature and light-level data and then transmit summaries of these data through the Argos satellite system at the time of release from their hosts. PSATs were originally engineered to support research on large pelagic fish (e.g., Block et al., 1998), including assessment of movements, dive behaviour, and post-release survival (Block et al., 2001; Boustany et al., 2002; Graves et al., 2002; Domeier et al., 2003; Kerstetter et al., 2003; Horodysky and Graves, 2005; Tracey et al., 2016; Schaefer et al., 2019; Strøm et al., 2019; Schaefer et al., 2021). They have also been used in sea turtle research (Hall and James, 2021), including a small number of post-release mortality studies (Sasso and Epperly, 2007; Àlvarez de Quevedo et al., 2013; Swimmer et al., 2014; Maxwell et al., 2018).
Loggerhead sea turtles are listed as endangered under Canada’s Species at Risk Act (COSEWIC, 2014) and the Northwest Atlantic Distinct Population Segment, which includes loggerheads that forage in Atlantic Canada, is listed as threatened under the U.S. Endangered Species Act (76 FR 58868; October 24, 2011). The Northwest Atlantic Ocean supports high fishing effort (Lewison et al., 2004a; Wallace et al., 2010), including pelagic longline fisheries targeting swordfish, tuna and sharks (Stokes et al., 2012). Bycatch in these fisheries is considered to be a large source of injury and mortality for sea turtles (Stokes et al., 2012) and a primary threat to the recovery of loggerhead sea turtle populations (Lewison et al., 2013; Casale and Tucker, 2017; Ceriani and Meylan, 2017).
At high latitudes of the North Atlantic, large seasonal concentrations of loggerhead turtles can be associated with productive offshore waters, especially those subject to the warming influence of the Gulf Stream current (Gardner et al., 2008; Parra et al., 2023). In Atlantic Canada, loggerhead turtles have been documented from late spring through fall as regular bycatch in shallow-set large pelagic longline fisheries targeting yellowfin and bigeye tuna (Thunnus albacores, Thunnus obesus) and swordfish (Xiphias gladius) along the Scotian Shelf, Georges Bank and the Grand Banks (Brazner and McMillan, 2008; Gardner et al., 2008). To better understand the impact of temperate North Atlantic pelagic longline fisheries on loggerhead populations, accurate estimates of associated bycatch-related mortality are required. Robust sample sizes (tens of individuals) are conspicuously lacking in most wildlife telemetry studies, limiting confidence in results (Sequeira et al., 2019). To date, empirically-derived estimates of post-release mortality based on large sample sizes have not been available for loggerhead turtles interacting with temperate North Atlantic pelagic longline fisheries. To address this knowledge gap, we deployed PSATs on loggerhead turtles incidentally hooked in Atlantic Canadian shallow-set pelagic longline gear.
2 Materials and methods
2.1 Tagging
Over the course of five tagging seasons (2012–2018), PSATs (PAT-Mk10 (n = 38), Mini-PAT (n = 13), or SPLASH323-B (n = 11); Wildlife Computers, Redmond, WA) were deployed on 62 loggerhead turtles incidentally hooked in Atlantic Canadian shallow-set pelagic longline gear targeting tuna species and swordfish in the Canadian Exclusive Economic Zone. In contrast to deep-set fishing practices in other areas, when targeting swordfish, the Atlantic Canadian pelagic longline fishery typically sets gear in the upper 20 m with 4.5 m drop lines and 8 m gangions (Carruthers et al., 2011).
In the present study, hooked loggerheads were boarded with a dip net and hook location was photographed. PSATs were attached by fishers, fisheries observers or biologists using a ~20 cm tether anchored to the posterior margin of the carapace, as per Epperly et al. (2007). Curved carapace length (CCL) and curved carapace width (CCW) were measured using a flexible tape measure. Turtles hooked in the mouth or the jaw were classified as “shallow hooked” (Figures 1A–C), and turtles that were hooked in the glottis region or deeper (i.e., hook swallowed) were classified as “deep hooked” (Figure 1D). To best approximate conventional release practices, hooks (normally size 16 and mandated to be circle type and corrodible) were not removed. The length of monofilament left attached to the hook (if any) was inconsistently recorded, and could not be reliably discerned from photographs; therefore, this variable was not considered in the analysis.

Figure 1. Common hooking locations for loggerhead sea turtles incidentally hooked in Atlantic Canadian pelagic longline gear. "Shallow hooked" includes (A) mouth, (B) jaw, and (C) beak; and "deep hooked" includes (D) swallowed.
2.2 Tag programming
PSATs were programmed to release after a set interval (120–365 days after deployment; “scheduled” release type), or after a prolonged period (4 or 8 days) at a constant depth (± 1 m) (“premature” release type). PSATs were also programmed to release if the maximum depth threshold (1800 m) was reached (“too deep” release type). If the tag’s pressure sensor failed to initiate release at maximum depth, a mechanical release was triggered (“too deep” release type). Following release, PSATs are designed to float to the surface and transmit archived data until their batteries are fully depleted.
PSATs are geolocated by, and transmit data to, the Argos satellite network (http://www.argos-system.org/) and archive environmental data such as depth (m), temperature (°C), and light intensity. Depth and temperature data were collected and compiled by PSATs as six-hour histograms (time at depth and time at temperature) separated into either 12 or 14 bins ranging from 0 to >200 m and 8 to >32°C. The tags also recorded depth-temperature profiles from which daily minimum and maximum dive depths were extracted. Forty-nine tags (excluding Mini-PATs) were programmed to transmit opportunistically on the first of each month, which enabled confirmation that the tag remained attached and was operating as expected. Otherwise, data were archived and batch transmitted upon release. Argos data were decoded and exported using the Wildlife Computers Data Analysis Program (DAP; https://wildlifecomputers.com/).
2.3 Tag data
Dive and temperature data were truncated to include only observations when the tags were assumed to be attached to the animal (i.e., removing surface observations that occurred after premature tag detachments or observations associated with the prolonged depth threshold criteria). Average time spent at surface was calculated as the average daily percentage of time spent at 0–1 m, and the maximum dive depth corresponded to the deepest daily dive. These parameters have been widely used to characterize turtle diving behavior (e.g., Polovina et al. (2003); Swimmer et al. (2006), and Àlvarez de Quevedo et al. (2013)) and were examined throughout the tracking period to identify potential changes in dive patterns indicative of mortality events, such as a marked increase in time spent at surface and/or tags reaching depths beyond what is biologically probable for a living turtle (e.g., Figure 2). Recognizing that marked changes in dive patterns do not always reflect compromised health in turtles, but can also be associated with different habitats and changing environmental conditions (i.e., water temperature, bathymetry), we examined depth-temperature profiles and used Argos locations to determine whether turtles exploited neritic or oceanic habitat. Argos locations were also used to identify where tags released. For PSATs that did not transmit upon release, the last monthly Argos location was considered to be the end of the deployment.
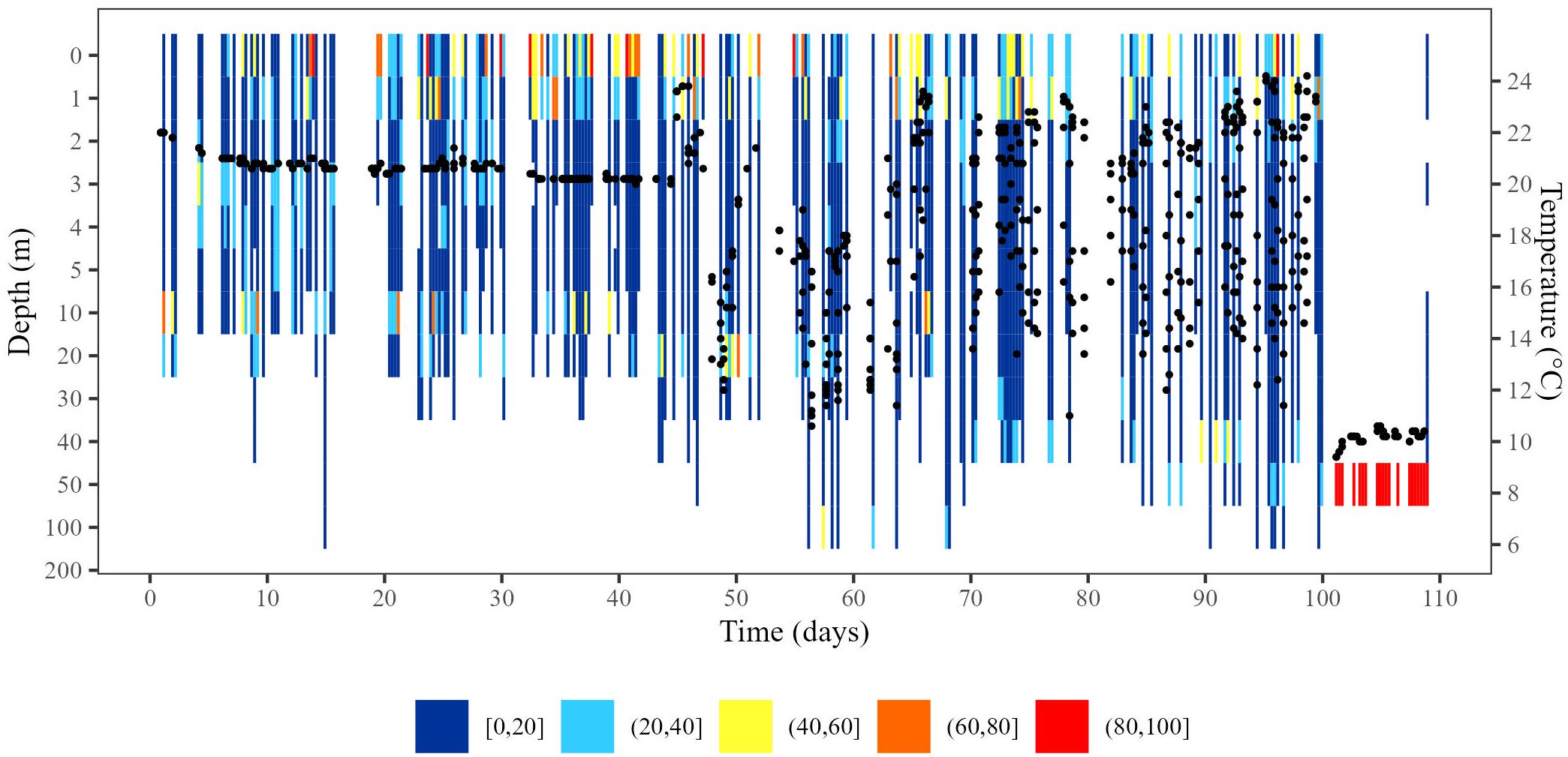
Figure 2. Behaviour of a “mortality assumed” turtle throughout PSAT deployment. X-axis indicates number of days tag was deployed; y-axis indicates depth (m), with black dots corresponding to temperature (secondary y-axis; °C). Colours indicate daily percentage of time at depth as indicated in legend. Tag 109809 was deployed on May 2, 2012, and released prematurely on August 18, 2012. On August 8, 2012, the turtle died and sank to the sea bottom, where the tag remained for 8 days before release (constant depth threshold was reached).
2.4 Turtle fate
As programmed, PSATs in this study allow turtle fate categorization as follows: (1) “survival assumed” (full programmed deployment was achieved and turtle appeared to be diving normally prior to tag release); (2) “survival suspected” (turtle appeared to be regularly diving, but tag detached for an unknown reason; tag remained at the surface for 4 or 8 days and released prematurely; tag did not report release type, but turtle appeared to be diving until last data transmission; or tag did not transmit upon release or insufficient dive data were transmitted, but monthly opportunistic transmissions were received); (3) “mortality assumed” (turtle exhibited a marked change in diving behavior until it sank and tag remained at constant depth for 4 or 8 days; or tag sank below 1800 m followed by tag releasing and remaining at the surface for 4 or 8 days).
2.5 Survival estimation
Based on the three possible fate assignments, annual survival probability of tagged turtles was estimated. Turtles with tags that did not transmit upon release were included as censored results (i.e., alive until last observation). Sixty-two individual turtle fates were used to calculate post-release survival.
All statistical analyses were conducted using R 3.6.1 (R Core Team, 2021). Survival was estimated using the Kaplan-Meier estimator with right censoring using the survival package in R (Kaplan and Meier, 1958; R Core Team, 2021; Therneau et al., 2021). This method makes no assumptions about the fate of turtles without inferred mortalities, which allows for estimation of survival even if an individual’s tracking duration is shorter than the PSAT's programmed deployment length and no time of mortality is determined. Turtles are considered alive at last observation when no further data are available and are subsequently censored. For example, “survival suspected” turtles that do not achieve full programmed deployments are censored at their last known observation. A Kaplan-Meier survival model was run and survival curves were tested using a log-rank test (Therneau et al., 2021) to determine if annual survival varied according to hooking location (i.e., hooking location was included in the model as an explicative variable). The model uses a status indicator of zero for survival (right censored) and one for mortality to estimate the survival probability. The resulting survival curve indicates the probability of survival over time, with each step in the curve indicating a mortality.
3 Results
Mean curved carapace length (CCL) across all tagged turtles (n = 62) was 65.45 ± SD 5.62 cm (range: 54.50–75.0 cm) and mean curved carapace width (CCW) was 63.67 ± SD 6.57 cm (range: 42.0–76.50 cm) (Supplementary Table 1). A total of 23 turtles were hooked in the mouth or the jaw and were classified as shallow hooked, and 39 turtles swallowed the hook and were classified as deep hooked (Supplementary Table 1).
3.1 Tag release and data transmission
Of the 62 tags deployed, 22 released as programmed (at scheduled time), 27 released prematurely, and 13 did not report release, but opportunistically transmitted on the first of the month (Supplementary Table 1). Hardware malfunction was attributed to two tags that released prematurely and for which the turtle was suspected to be alive at the end of tracking (Supplementary Table 1).
3.2 Turtle fates
Data from all 62 PSAT deployments were used to determine turtle fates. Twenty-two turtles were classified as “survival assumed”, 35 as “survival suspected”, and 5 as “mortality assumed” (Supplementary Table 1). Locations of turtles at the time of assumed fate were widely dispersed (Figure 3).
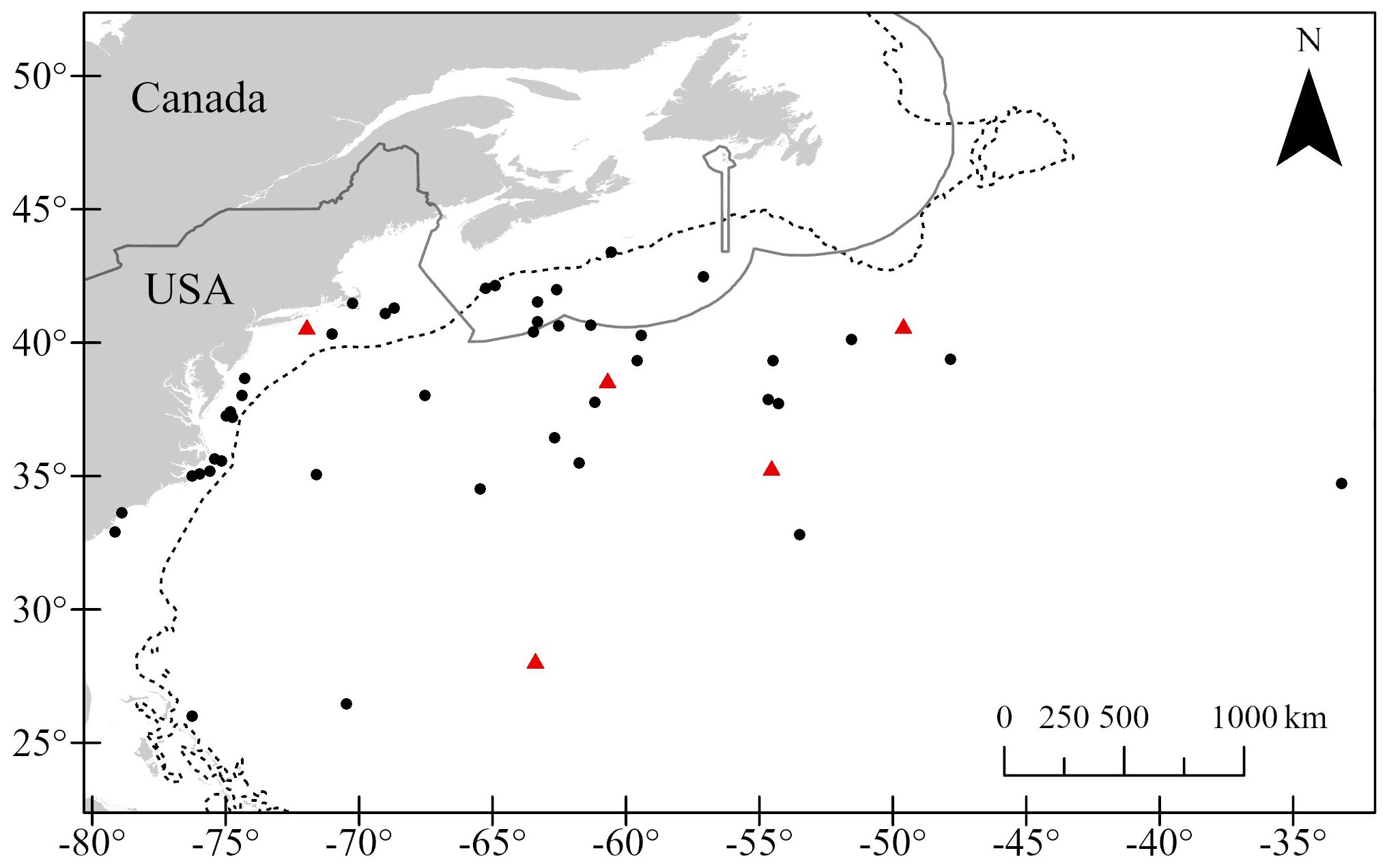
Figure 3. Post-release locations of 49 loggerhead turtles incidentally hooked in Atlantic Canadian pelagic longline gear, colored by fate: red triangles = "mortality assumed", black circles = "survival suspected" and "survival assumed". Dotted line indicates 1000 m isobath and the solid line is the Canadian Economic Exclusive Zone. Plot does not include tags that did not report a release location (n = 13).
For the “survival assumed” turtles (e.g., Figure 4), no marked change in daily proportion of time spent at surface was observed. Turtles spent 34.48 ± SD 1.75% of time at surface per day (range: 14.49–45.63%; Supplementary Table 1). Mean deepest daily dive depth was 49.31 ± SD 4.16 m (range: 28.0–77.20 m; Supplementary Table 1).
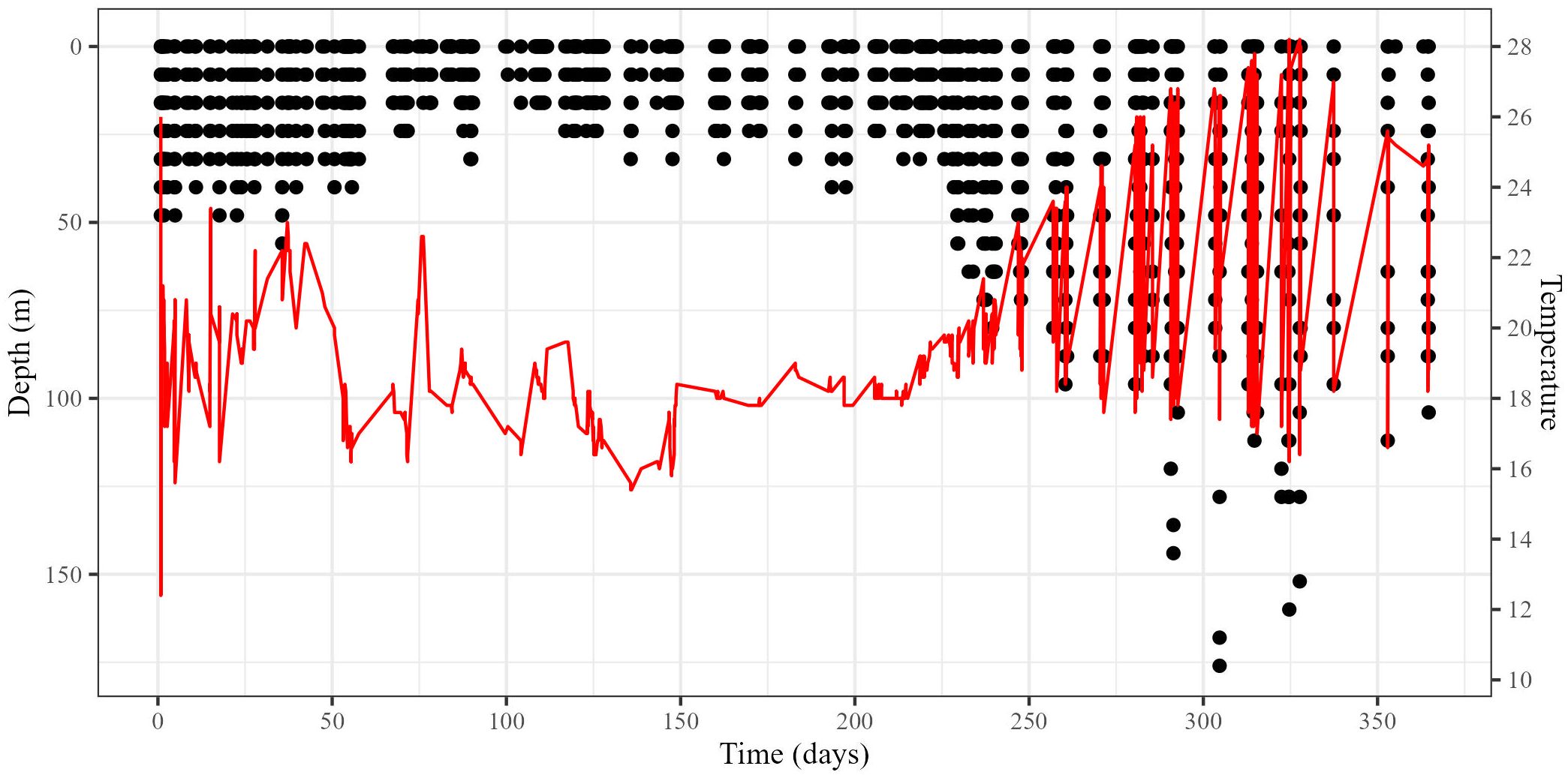
Figure 4. Depth (black points; primary y-axis) and temperature (red line; secondary y-axis) of a “survival assumed” turtle throughout PSAT deployment. Tag 121238 was deployed on October 6, 2012, and released as programmed after 365 days.
Among the “survival suspected” turtles, 22 PSATs prematurely released and 13 ceased transmissions without reporting release. Dive patterns did not change markedly throughout the tracking period for most PSATs that had released prematurely even though turtles occasionally exploited deeper depths (e.g., Figure 5). Overall, turtles spent 37.75 ± SD 2.52% of the day at the surface (range: 16.12–60.71%; Supplementary Table 1). The mean deepest daily dive was 55.24 ± SD 5.36 m (range: 22.22–97.82 m; Supplementary Table 1).
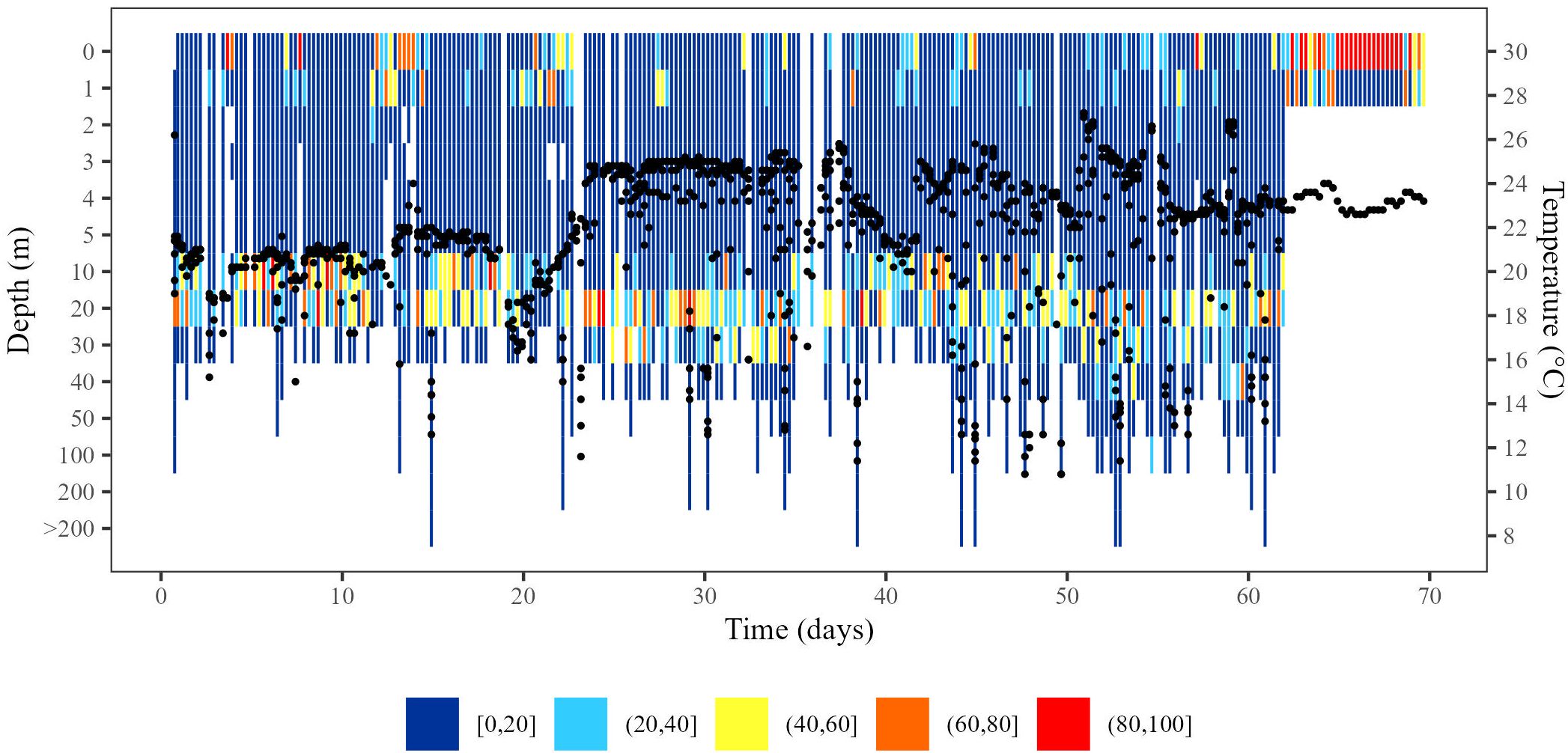
Figure 5. Behaviour of a “survival suspected” turtle throughout PSAT deployment. X-axis indicates number of days tag was deployed; y-axis indicates depth (m), with black dots corresponding to temperature (secondary y-axis; °C). Colours indicate daily percentage of time at depth as indicated in legend. Tag 117636 was deployed on July 9, 2012, and released prematurely on September 16, 2012, after remaining 8 days at the surface (70 day deployment). Turtle was assumed to be alive until September 8, 2012.
For the “mortality assumed” turtles (n = 4 deep hooked; n = 1 shallow hooked), one turtle died inshore and sank to the bottom (at 48 m) after 98.65 days of tracking (tag 109809; Figure 2) and two turtles died and sank offshore after 105.39 days and 21.30 days of tracking (at 1832 m and 1952 m; tags 117632 and 120016, respectively). Two turtles sank below the 1800 m tag release threshold depth (tag 148988 at 78.91 d and tag 148980 at 106.64 d). These depth patterns are clear indications of mortality.
3.3 Survival estimation
The annual probability of survival for an individual turtle post-release using the Kaplan-Meier estimator was 0.877 ± SE 0.052 (Figure 6; Table 1). A log-rank test indicated that there was no significant difference in survival between shallow-hooked and deep-hooked turtles (p = 0.20).
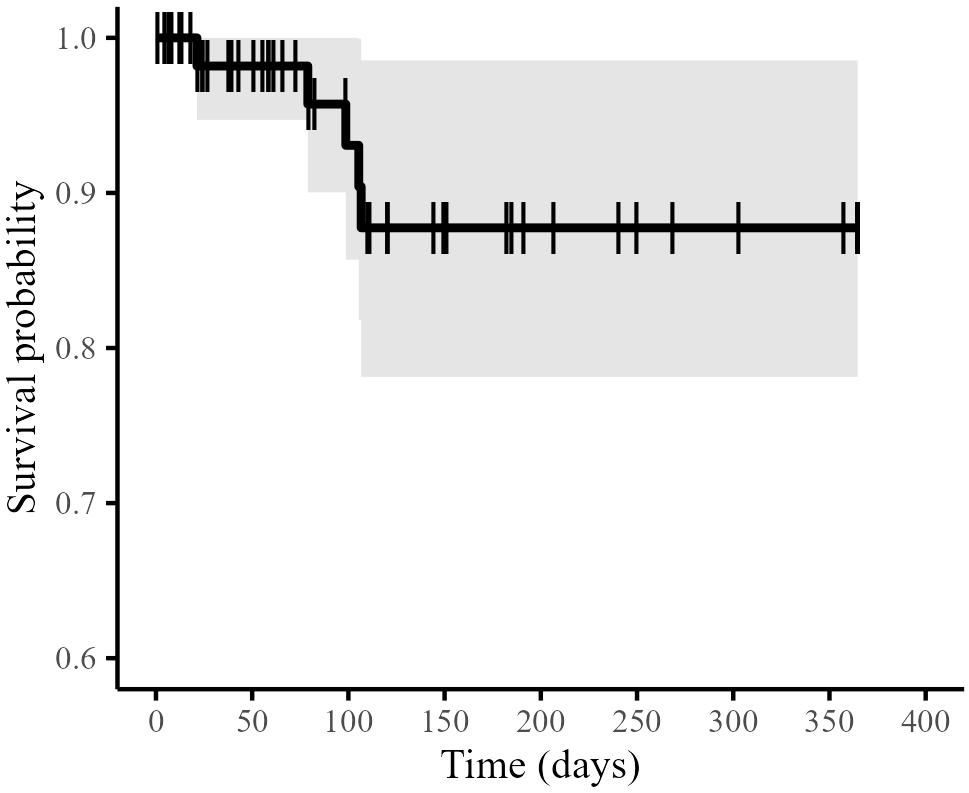
Figure 6. Kaplan-Meier estimates of overall annual survival of loggerhead turtles incidentally hooked in Atlantic Canadian pelagic longline gear. Solid line is the survivorship curve, with shaded 95% confidence intervals. Hashed lines are censored results. Each step is a mortality.
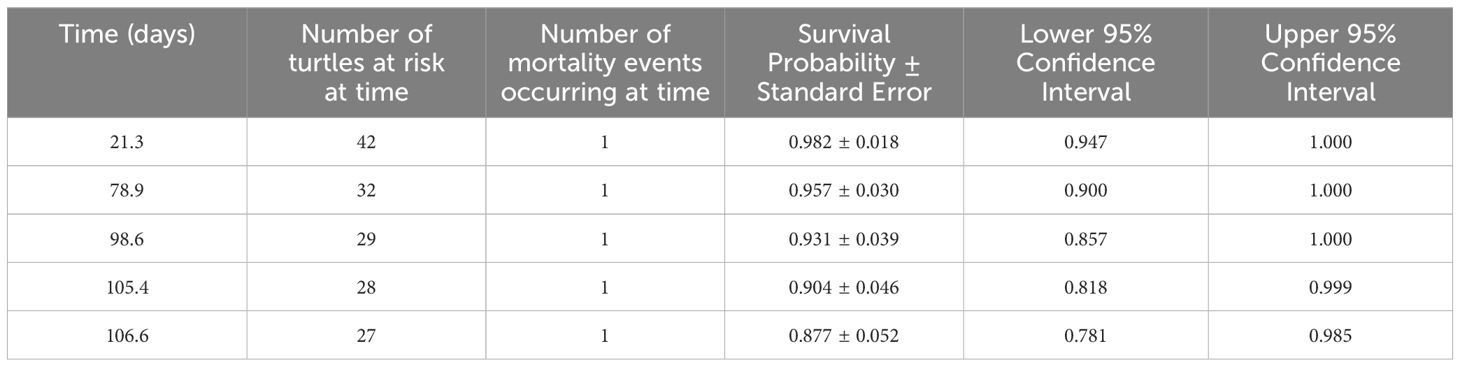
Table 1. Post-release survival of loggerhead turtles incidentally hooked in Atlantic Canadian pelagic longline gear: Kaplan-Meier estimator results.
4 Discussion
4.1 Survival estimation
Estimating post-release survival using PSATs relies on a suite of assumptions to assign an animal’s fate. First, normally only animals with tags that report release can be included in such analyses. However, unlike previous sea turtle post-release survival studies, we were able to include turtles with tags that did not report release as censored results because by enabling monthly opportunistic transmissions, we were able to identify dates when turtles were last known to be alive.
Second, prematurely-detached tags may not reflect mortality (Sasso et al., 2011). PSATs are programmed to release prematurely under conditions indicative of sea turtle mortality, including a prolonged period of submergence (4 or 8 days) at a constant depth (indicating the animal failed to surface to breathe) or descent to a depth > 1800 m (exceeding the expected depth maximum of loggerhead sea turtles). Premature release triggered by 4 or 8 days spent at a constant depth of 0 m (i.e., at the surface) may be indicative of hardware malfunction or tag attachment failure. For prematurely-released tags that were not definitive cases of mortality, if tag data indicated that turtles were diving normally up until tag release, we assumed they were alive at tag release.
Studies on other species, such as sharks, have assigned mortality on the basis of tags registering constant depth for 2 days (Sulikowski et al., 2020), but have also identified tags with attachment failures and deemed them survivors (Schaefer et al., 2019). In studies where tags released at the maximum threshold depth, sinking rate has been used to decipher whether the animal was sinking with the tag still attached or if the tag had released from the animal and was sinking on its own (Musyl and Gilman, 2018; Orbesen et al., 2018). In the present study, only positively buoyant tags were used, so instances of tags sinking to release threshold depth clearly indicated turtle mortalities as dead turtles initially sink (Schultz et al., 2022). Consistent with the approach of multiple fishery post-release studies on other species, such as sharks and tuna (Tracey et al., 2016; Musyl and Gilman, 2018; Schaefer et al., 2019, 2021), we used a Kaplan-Meier survival model with right censoring. With this method, the annual probability of survival for an individual loggerhead turtle post-release was 0.877 ± SE 0.052.
The only published estimate of post-release survival for sea turtles incidentally hooked in pelagic longline fisheries approximating our result is that of Sasso and Epperly (2007), who reported a survival rate of 0.81 (CI 95%: 0.55–0.94) from a known-fate model for juvenile loggerhead turtles captured by the US Atlantic fleet. Although we used instrumentation and attachment methods consistent with Sasso and Epperly (2007), their sample was smaller (n = 10 turtles corresponding to the hooking classifications used here versus n = 62); only lightly hooked turtles were tagged (including 4 hooked externally or in the beak versus all mouth hooked or deeper in the present study); and all fishing gear was removed from turtles before release (we left hooks in place).
Apart from Sasso and Epperly (2007), previously published studies of post-release fates of cheloniid sea turtles hooked in pelagic longline fisheries have occurred outside of the North Atlantic. All earlier studies have reported survival estimates lower than that of the present study: 0.81 (Sasso and Epperly, 2007), 0.635-0.692 (Àlvarez de Quevedo et al., 2013), and 0.72 (Swimmer et al., 2014). The disparity in results may be the result of smaller sample sizes; differences in environmental (oceanic) conditions; durations of hypoxia; fishing gear (e.g., hook size, gangion length); fishing practices (e.g., soak time and depth); and both reported and unreported turtle handling practices (e.g., time aboard, steps taken to prevent overheating). For example, we might expect higher mortality for hooked turtles subjected to longer periods of forced submergence prior to boarding (Snoddy et al., 2009; Franchini et al., 2021). Turtles may also be more likely to drown when hooked in deep-set gear; Fahlman et al. (2017) reported, for example, that the severity and fatality of gas embolism increases with depth. Further, it is possible that measures to remove hooks (as was standard practice in some previous studies) may inadvertently complicate post-release recovery. As concluded by both PSAT (Sasso and Epperly, 2007; Àlvarez de Quevedo et al., 2013; Swimmer et al., 2014) and rehabilitation studies of sea turtles (Casale et al., 2008), delayed mortality (i.e., weeks to months) is typical following incidental hooking. However, as the interval between initial interaction and mortality event increases, so does the uncertainty surrounding etiology.
Beyond these factors, methodological differences in tag attachment (e.g., tethered in this study and Sasso and Epperly (2007) versus epoxied as per Àlvarez de Quevedo et al. (2013) and Swimmer et al. (2014)); type of instrumentation used (make and models of tag and data streams collected); tag parameter settings (e.g., time to scheduled tag release); and, perhaps most importantly, how tagging data are interpreted, are expected to impact results. We suggest the higher survivorship result we found at least partially reflects our decision to take the precautionary step of enabling interim (one day per month) Argos transmissions by tags. This allowed us to determine when turtles were last alive, and thereby count them as “survival suspected” (again, turtles with tags that didn’t ultimately report at programmed release might otherwise be mistakenly classified as dead). We urge that this measure and others be taken in the future to help reduce potential bias towards diagnosing mortality in electronic tagging studies.
Swimmer et al. (2014) and Àlvarez de Quevedo et al. (2013) inferred post-release mortalities of loggerhead turtles hooked in pelagic longline fisheries occurring within 90 days of tagging. In the present study, among the five turtles that died, mortality occurred at 21.3 days (n = 1), 78.9 days (n = 1), and > 90 days (n = 3) after tagging. Sasso and Epperly (2007) reported a loggerhead mortality 177 days post-release. While post-release mortality can occur beyond the time frame for tag deployments investigated here (through infection, decreased mobility or feeding efficiency, chronic debilitation, etc.), with increased time post-release, individuals may experience additional threats (natural or anthropogenic), making it more difficult to attribute the cause of mortality to the initial fishery interaction. Fortunately, behavioural metrics such as dive patterns, both within and between individuals in a sample, can help clarify the condition of tagged animals throughout tag deployments and, as shown here, may be especially useful to consider in the time frame immediately before tags are prematurely released.
4.2 Hook removal
Many countries encourage or mandate their domestic pelagic longline fleets to adopt hard shell sea turtle de-hooking protocols developed with the intent of increasing post-release survival. Previous studies have indicated that hook removal from shallow-hooked turtles results in a low probability of mortality after release (Swimmer et al., 2006; Sasso and Epperly, 2007) provided that such removals are carefully executed (Parga, 2012). A higher probability of mortality has been attributed to leaving the hook in place in deep-hooked turtles equipped with PSATs (Àlvarez de Quevedo et al., 2013). However, it has been suggested by others that attempts to remove hooks that are lodged in the gastrointestinal tract beyond the proximal esophagus or that have perforated large vessels or organs may complicate injuries and potentially cause lethal damage (Parga, 2012), and as long as the associated monofilament is not pulled, turtles that swallow a hook often survive (Alegre et al., 2006; Valente et al., 2007).
In the present study, to emulate conventional fishing practices in Atlantic Canada and minimize potentially confounding variables, all turtles were released with hooks embedded. Nonetheless, the probability of survivorship was found to be higher than that reported in all previous studies, including those where hooks were removed. Moreover, we found that survival did not statistically vary between deep-hooked and shallow-hooked turtles, possibly in part due to the sample size of observed mortalities (n = 5). There was limited evidence that deep-hooked turtles experience higher mortality than shallow-hooked turtles based on the proportion estimated dead in each group: 10.3% (n = 39) versus 4.3% (n = 23), respectively. This result suggests that even if fishers carefully assess hooked turtles against eligibility criteria for de-hooking (which may be impractical), proceeding with hook removal may offer limited conservation benefit. This possibility is something that should be explored through further research.
It is important to note that while hooks were left embedded in the present study, by removing most or all of the attached monofilament more serious or lethal injuries caused by strangulation, tractions, lesions, or perforations in the gastrointestinal tract or adjacent organs (Work and Balazs, 2010) may have been prevented (Parga, 2012). Therefore, even when hooks are not removed, encouraging fishers to follow other recommendations for handling hooked turtles is important, as such practices may play a critical role in enhancing their post-release survival (Parga et al., 2015).
Apart from a potential effect of hook location, there are other variables associated with fishery interactions that may influence sea turtle post-release survival. In most cases these are difficult, if not impossible, to quantify. For example, hooked turtles may remain forcibly submerged for extended periods, experiencing a lack of oxygen that can result in a disruption of physiological parameters (Snoddy and Williard, 2010; Work and Balazs, 2010; Williard et al., 2015). To separate variables contributing to mortality associated with the fishery interaction from individual health status, improved indexes of body condition readily applied in the field at the time of turtle release are urgently required.
4.3 PSATs
In the present study, 21 tags (37%; excluding “mortality assumed” tags) released prematurely even though turtles appeared to be diving normally prior to release, and 13 tags (21%) did not report release (Supplementary Table 1). Sasso and Epperly (2007) used the same tag attachment protocol we followed and reported that 16% of PSATs released prematurely. Baseplates have been epoxied to the carapace as an anchor point for tethered PSATs in other sea turtle studies (e.g., Àlvarez de Quevedo et al., 2013). However, epoxy can detach from growing scutes after a short period of time (Mansfield et al., 2012), sink the towed tag, and potentially lead to a false diagnosis of turtle mortality. We suggest that direct attachment methods like the one we employed enhance PSAT retention and may be less likely to confound interpretation of results in sea turtle survival studies.
Incomplete receipt of archival datasets from PSATs by Argos following tag release was apparent in the current study; data gaps were associated with many tags. This phenomenon has been reported previously. Musyl et al. (2011) reviewed potential PSAT performance issues over 731 PSAT deployments on 19 marine species. Overall, the authors estimated that 21% of PSATs failed to transmit any data. Sasso and Epperly (2007) reported that 16% of PSATs attached to juvenile loggerheads turtles failed to transmit data. In the present study, the amount of recovered dive data varied widely among individuals. PSATs transmitted an average of 58.07 ± SD 4.95% deployment days; the maximum number of reporting days was 165.
5 Conclusions
Given the Northwest Atlantic loggerhead turtle population’s precarious conservation status and the regularity with which loggerhead turtles interact with pelagic longline fisheries in Atlantic Canadian waters (Brazner and McMillan, 2008) and elsewhere (Lewison et al., 2004b; Wallace et al., 2013), it is important to estimate the degree to which such interactions result in mortality using the best empirical methods available. We suggest that data collection and transmission by PSATs both during tag deployment and at pop-off provides the most complete record of turtle behaviour to support assessment of fates and survival modelling.
This study’s findings contribute to a better understanding of the impact of shallow-set pelagic longline fisheries on the recovery of the endangered Northwest Atlantic loggerhead turtle population and can help inform management measures. While pelagic longline fisheries have long been posited as a key anthropogenic threat to this species, our results suggest that, at least in the context of where and how the Atlantic Canadian fleet operates, post-release survival of incidentally hooked loggerheads is higher than what has been reported from previous research.
While we caution against extrapolating survival estimates derived from the present study to deep-set pelagic longline fisheries, our empirical findings stand in contrast to prevailing wisdom regarding loggerhead turtle mortality in pelagic longline gear. This is significant as fishery impacts on the loggerhead epipelagic life history stage in oceanic areas (again principally via interactions with pelagic longline gear) have traditionally been seen as especially limiting to population recovery (Lewison et al., 2004b). If, as we suggest, loggerhead mortality in the Northwest Atlantic shallow-set pelagic longline fishery is much lower than previously assumed, then population impacts in neritic areas may actually be an underestimated conservation concern and deserving of enhanced science and management attention.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by Fisheries and Oceans Canada Maritimes Region Animal Care Committee. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
MJ: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Writing – original draft, Writing – review & editing. KH: Data curation, Formal analysis, Methodology, Software, Visualization, Writing – review & editing. EB: Data curation, Methodology, Writing – review & editing. SS-M: Formal analysis, Methodology, Software, Writing – review & editing. VP: Formal analysis, Methodology, Software, Visualization, Writing – original draft.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Project funding was provided by Fisheries and Oceans Canada Internal Governance Strategy Science Program.
Acknowledgments
We thank L. Belskis, K. Clarke, S. Cole, S. Epperly, K. Martin, J. Porter, C. Sasso, and L. Stokes for thoughtful suggestions and guidance. E. Christiansen, M. Godfrey, C. Harms, C. Legner, and the North Carolina Aquarium Sea Turtle Assistance and Rehabilitation program kindly facilitated tag attachment trials and provided mentoring in loggerhead handling and sampling protocols. We are grateful to the Canadian Sea Turtle Network, Javitech Ltd., NOAA-National Marine Fisheries Service, and the Nova Scotia Swordfishermen’s Association for their support of this research. This work would not have been possible without the generous volunteer efforts of the Nova Scotia fishing captains and crews who assisted with instrument deployments.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1392582/full#supplementary-material
References
Alegre F., Parga M. L., del Castillo C., Pont S. (2006). “Study on the long-term effect of hooks in the mid-oesophagus of sea turtles,” in Book of abstracts. 26th annual symposium on sea turtle biology and conservation. 234 (Athens: International Sea Turtle Society), 234.
Àlvarez de Quevedo I., San Félix M., Cardona L. (2013). Mortality rates in by-caught loggerhead turtle Caretta caretta in the Mediterranean Sea and implications for the Atlantic populations. Mar. Ecol. Prog. Ser. 489, 225–234. doi: 10.3354/meps10411
Benoît H. P., Kneebone J., Tracey S. R., Bernal D., Hartmann K., Golet W. (2020). Distinguishing discard mortality from natural mortality in field experiments based on electronic tagging. Fish. Res. 230, 105642. doi: 10.1016/j.fishres.2020.105642
Bjørge A., Skern-Mauritzen M., Rossman M. C. (2013). Estimated bycatch of harbour porpoise (Phocoena phocoena) in two coastal gillnet fisheries in Norway 2006–2008. Mitigation and implications for conservation. Biol. Conserv. 161, 164–173. doi: 10.1016/j.biocon.2013.03.009
Block B. A., Dewar H., Blackwell S. B., Williams T. D., Prince E. D., Farwell C. J., et al. (2001). Migratory movements, depth preferences, and thermal biology of Atlantic bluefin tuna. Science 293, 1310–1314. doi: 10.1126/science.1061197
Block B. A., Dewar H., Farwell C., Prince E. D. (1998). A new satellite technology for tracking the movements of Atlantic bluefin tuna. Proc. Natl. Acad. Sci. 95, 9384–9389. doi: 10.1073/pnas.95.16.9384
Bond E. P., James M. C. (2021). Postrelease movements of leatherback turtles (Dermochelys coriacea) following incidental capture in fishing gear in the Atlantic Ocean. Fish. Bull. 119, 255–260. doi: 10.7755/FB.119.4.5
Boustany A. M., Davis S. F., Pyle P., Anderson S. D., Le Boeuf B. J., Block B. A. (2002). Expanded niche for white sharks. Nature 415, 35–36. doi: 10.1038/415035b
Brazner J. C., McMillan J. (2008). Loggerhead turtle (Caretta caretta) bycatch in Canadian pelagic longline fisheries: Relative importance in the western North Atlantic and opportunities for mitigation. Fish. Res. 91, 310–324. doi: 10.1016/j.fishres.2007.12.023
Caracappa S., Persichetti M. F., Piazza A., Caracappa G., Gentile A., Marineo S., et al. (2018). Incidental catch of loggerhead sea turtles (Caretta caretta) along the Sicilian coasts by longline fishery. PeerJ 6, e5392. doi: 10.7717/peerj.5392
Carruthers E. H., Neilson J. D., Smith S. C. (2011). Overlooked bycatch mitigation opportunities in pelagic longline fisheries: Soak time and temperature effects on swordfish (Xiphias gladius) and blue shark (Prionace glauca) catch. Fish. Res. 108, 112–120. doi: 10.1016/j.fishres.2010.12.008
Carruthers E. H., Schneider D. C., Neilson J. D. (2009). Estimating the odds of survival and identifying mitigation opportunities for common bycatch in pelagic longline fisheries. Biol. Conserv. 142, 2620–2630. doi: 10.1016/j.biocon.2009.06.010
Casale P., Freggi D., Rocco M. (2008). Mortality induced by drifting longline hooks and branchlines in loggerhead sea turtles, estimated through observation in captivity. Aquat. Conserv.: Mar. Freshw. Ecosyst. 18, 945–954. doi: 10.1002/aqc.894
Casale P., Tucker A. D. (2017). Caretta caretta (amended version of 2015 assessment). The IUCN Red List of Threatened Species. 2017, e.T3897A119333622. doi: 10.2305/IUCN.UK.2017-2.RLTS.T3897A119333622.en
Cassoff R. M., Moore K. M., McLellan W. A., Barco S. G., Rotstein D. S., Moore M. J. (2011). Lethal entanglement in baleen whales. Dis. Aquat. Org. 96, 175–185. doi: 10.3354/dao02385
Ceriani S. A., Meylan A. B. (2017). Caretta caretta (North West Atlantic subpopulation) (amended version of 2015 assessment). The IUCN Red List of Threatened Species, 2017: e.T84131194A119339029. doi: 10.2305/IUCN.UK.2017-2.RLTS.T84131194A119339029.en
Chaloupka M., Parker D., Balazs G. (2004). Tracking turtles to their death - reply to Hays et al. Mar. Ecol. Prog. Ser. 283, 301–302. doi: 10.3354/meps283301
COSEWIC (2014). COSEWIC assessment and status report on the Porbeagle Lamna nasus in Canada (Ottawa: Committee on the Status of Endangered Wildlife in Canada). xi + 40 pp.
Domeier M. L., Dewar H., Nasby-Lucas N. (2003). Mortality rate of striped marlin (Tetrapturus audax) caught with recreational tackle. Mar. Freshw. Res. 54, 435–445. doi: 10.1071/MF01270
Eddy C., Brill R., Bernal D. (2016). Rates of at-vessel mortality and post-release survival of pelagic sharks captured with tuna purse seines around drifting fish aggregating devices (FADs) in the equatorial eastern Pacific Ocean. Fish. Res. 174, 109–117. doi: 10.1016/j.fishres.2015.09.008
Epperly S. P., Watson J. W., Foster D. G., Shah A. K. (2012). Anatomical hooking location and condition of animals captured with pelagic longlines: The Grand Banks experiments 2002–2003. B. Mar. Sci. 88, 513–527. doi: 10.5343/bms.2011.1083
Epperly S. P., Wyneken J., Flanagan J. P., Harms C. A., Higgins B. (2007). Attachment of popup archival transmitting (PAT) tags to loggerhead sea turtles (Caretta caretta). Herpetol. Rev. 38, 419–425.
Fahlman A., Crespo-Picazo J. L., Sterba-Boatwright B., Stacy B. A., Garcia-Parraga D. (2017). Defining risk variables causing gas embolism in loggerhead sea turtles (Caretta caretta) caught in trawls and gillnets. Sci. Rep. 7, 2739. doi: 10.1038/s41598-017-02819-5
Fenton J., Ellis J. M., Falterman B., Kerstetter D. W. (2015). Habitat utilization of blackfin tuna, Thunnus atlanticus, in the north-central Gulf of Mexico. Environ. Biol. Fish 98, 1141–1150. doi: 10.1007/s10641-014-0347-3
Ferter K., Rikardsen A. H., Evensen T. H., Svenning M.-A., Tracey S. R. (2017). Survival of Atlantic halibut (Hippoglossus hippoglossus) following catch-and-release angling. Fish. Res. 186, 634–641. doi: 10.1016/j.fishres.2016.05.022
Franchini D., Valastro C., Ciccarelli S., Trerotoli P., Paci S., Caprio S., et al. (2021). Analysis of risk factors associated with gas embolism and evaluation of predictors of mortality in 482 loggerhead sea turtles. Sci. Rep. 11, 22693. doi: 10.1038/s41598-021-02017-4
Frankish C. K., Cunningham C., Manica A., Clay T. A., Prince S., Phillips R. A. (2021). Tracking juveniles confirms fisheries-bycatch hotspot for an endangered albatross. Biol. Conserv. 261, 109288. doi: 10.1016/j.biocon.2021.109288
Gardner B., Sullivan P. J., Morreale S. J., Epperly S. P. (2008). Spatial and temporal statistical analysis of bycatch data: Patterns of sea turtle bycatch in the North Atlantic. Can. J. Fish. Aquat. Sci. 65, 2461–2470. doi: 10.1139/F08-152
Gilman E., Kobayashi D., Swenarton T., Brothers N., Dalzell P., Kinan-Kelly I. (2007). Reducing sea turtle interactions in the Hawaii-based longline swordfish fishery. Biol. Conserv. 139, 19–28. doi: 10.1016/j.biocon.2007.06.002
Graves J. E., Luckhurst B. E., Prince E. D. (2002). An evaluation of pop-up satellite tags for estimating postrelease survival of blue marline (Makaira nigricans) from a recreational fishery. Fish. Bull. 100, 134–142.
Gray C. A., Kennelly S. J. (2018). Bycatches of endangered, threatened and protected species in marine fisheries. Rev. Fish. Biol. Fisher. 28, 521–541. doi: 10.1007/s11160-018-9520-7
Hall K. E., James M. C. (2021). Predation of satellite-tagged juvenile loggerhead turtles Caretta caretta in the Northwest Atlantic Ocean. Endang. Species. Res. 46, 279–291. doi: 10.3354/esr01165
Hays G. C., Bradshaw C. J. A., James M. C., Lovell P., Sims D. W. (2007). Why do Argos satellite tags deployed on marine animals stop transmitting? J. Exp. Mar. Biol. Ecol. 349, 52–60. doi: 10.1016/j.jembe.2007.04.016
Hays G. C., Broderick A. C., Godley B. J., Luschi P., Nichols W. J. (2003). Satellite telemetry suggests high levels of fishing-induced mortality in marine turtles. Mar. Ecol. Prog. Ser. 262, 305–309. doi: 10.3354/meps262305
Hays G. C., Laloë J.-O., Rattray A., Esteban N. (2021). Why do Argos satellite tags stop relaying data? Ecol. Evol. 11, 7093–7101. doi: 10.1002/ece3.7558
Horodysky A. Z., Graves J. E. (2005). Application of pop-up satellite archival tag technology to estimate postrelease survival of white marlin (Tetrapturus albidus) caught on circle and straight-shank (“J”) hooks in the western North Atlantic recreational fishery. Fish. Bull. 103, 84–96.
Howell E. A., Kobayashi D. R., Parker D. M., Balazs G. H., Polovina J. J. (2008). TurtleWatch: A tool to aid in the bycatch reduction of loggerhead turtles Caretta caretta in the Hawaii-based pelagic longline fishery. Endang. Species Res. 5, 267–278. doi: 10.3354/esr00096
Innis C., Merigo C., Dodge K., Tlusty M., Dodge M., Sharp B., et al. (2010). Health evaluation of leatherback turtles (Dermochelys coriacea) in the northwestern Atlantic during direct capture and fisheries gear disentanglement. Chelonian Conserv. Bi. 9, 205–222. doi: 10.2744/CCB-0838.1
Jackson L. S., Drymon J. M., Nelson T. R., Powers S. P. (2018). Biotelemetry based estimates of greater amberjack (Seriola dumerili) post-release mortality in the northern Gulf of Mexico. Fish. Res. 208, 239–246. doi: 10.1016/j.fishres.2018.07.017
Johnson A., Salvador G., Kenney J., Robbins J., Kraus S., Landry S., et al. (2005). Fishing gear involved in entanglements of right and humpback whales. Mar. Mamm. Sci. 21, 635–645. doi: 10.1111/j.1748-7692.2005.tb01256.x
Kaplan E. L., Meier P. (1958). Nonparametric estimation from incomplete observations. J. Am. Stat. Assoc. 53, 457–481. doi: 10.1080/01621459.1958.10501452
Kerstetter D. W., Luckhurst B. E., Prince E. D., Graves J. E. (2003). Use of pop-up satellite archival tags to demonstrate survival of blue marlin (Makaira nigricans) released from pelagic longline gear. Fish. Bull. 101, 939–948.
Lewison R. L., Crowder L. B., Read A. J., Freeman S. A. (2004a). Understanding impacts of fisheries bycatch on marine megafauna. Trends Ecol. Evol. 19, 598–604. doi: 10.1016/j.tree.2004.09.004
Lewison R. L., Freeman S. A., Crowder L. B. (2004b). Quantifying the effects of fisheries on threatened species: The impact of pelagic longlines on loggerhead and leatherback sea turtles. Ecol. Lett. 7, 221–231. doi: 10.1111/j.1461-0248.2004.00573.x
Lewison R. L., Wallace B. P., Alfaro-Shigueto J., Mangel J. C., Maxwell S. M., Hazen E. L. (2013). “Fisheries bycatch of marine turtles: Lessons learned from decades of research and conservation,” in The Biology of Sea Turtles. Eds. Wyneken J., Lohmann K. J., Musick J. A. (Boca Raton: CRC Press), 329–350.
Mangel J. C., Alfaro-Shigueto J., Witt M. J., Dutton P. H., Seminoff J. A., Godley B. J. (2011). Post-capture movements of loggerhead turtles in the southeastern Pacific Ocean assessed by satellite tracking. Mar. Ecol. Prog. Ser. 433, 261–272. doi: 10.3354/meps09152
Mansfield K. L., Wyneken J., Rittschof D., Walsh M., Lim C. W., Richards P. M. (2012). Satellite tag attachment methods for tracking neonate sea turtles. Mar. Ecol. Prog. Ser. 457, 181–192. doi: 10.3354/meps09485
Maxwell S. M., Witt M. J., Abitsi G., Aboro M. P., Agamboue P. D., Asseko G. M., et al. (2018). Sea turtles and survivability in demersal trawl fisheries: Do comatose olive ridley sea turtles survive post-release? Anim. Biotelem. 6, 11. doi: 10.1186/s40317-018-0155-1
Molina J. M., Cooke S. J. (2012). Trends in shark bycatch research: Current status and research needs. Rev. Fish. Biol. Fisher. 22, 719–737. doi: 10.1007/s11160-012-9269-3
Musyl M. K., Domeier M. L., Nasby-Lucas N., Brill R. W., McNaughton L. M., Swimmer J. Y., et al. (2011). Performance of pop-up satellite archival tags. Mar. Ecol. Prog. Ser. 433, 1–28. doi: 10.3354/meps09202
Musyl M. K., Gilman E. L. (2018). Post-release fishing mortality of blue (Prionace glauca) and silky shark (Carcharhinus falciformes) from a Palauan-based commercial longline fishery. Rev. Fish. Biol. Fisher. 28, 567–586. doi: 10.1007/s11160-018-9517-2
Nielsen J. K., Rose C. S., Loher T., Drobny P., Seitz A. C., Courtney M. B., et al. (2018). Characterizing activity and assessing bycatch survival of Pacific halibut with accelerometer Pop-up Satellite Archival Tags. Anim. Biotelem. 6, 10. doi: 10.1186/s40317-018-0154-2
Orbesen E. S., Brown C. A., Snodgrass D., Serafy J. E., Walter J. F. III (2018). At-vessel and postrelease mortality rates of bluefin tuna (Thunnus thynnus) associated with pelagic longline gear in the northern Gulf of Mexico. Fish. Bull. 117, 15–23. doi: 10.7755/FB.117.1.3
Parga M. L. (2012). Hooks and sea turtles: A veterinarian’s perspective. B. Mar. Sci. 88, 731–741. doi: 10.5343/bms.2011.1063
Parga M. L., Pons M., Andraka S., Rendón L., Mituhasi T., Hall M., et al. (2015). Hooking locations in sea turtles incidentally captured by artisanal longline fisheries in the Eastern Pacific Ocean. Fish. Res. 164, 231–237. doi: 10.1016/j.fishres.2014.11.012
Parra H., Machete M., Santos M., Bjorndal K. A., Vandeperre F. (2023). Incidental capture of sea turtles in the Northeast Atlantic Portuguese pelagic longline fishery. Fish. Res. 263, 106673. doi: 10.1016/j.fishres.2023.106673
Phillips B. E., Cannizzo S. A., Godfrey M. H., Stacy B. A., Harms C. A. (2015). Exertional myopathy in a juvenile green sea turtle (Chelonia mydas) entangled in a large mesh gillnet. Case Rep. Vet. Med. 2015, 604320. doi: 10.1155/2015/604320
Polovina J. J., Howell E., Parker D. M., Balazs G. H. (2003). Dive-depth distribution of loggerhead (Carretta carretta) and olive ridley (Lepidochelys olivacea) sea turtles in the central North Pacific: Might deep longline sets catch fewer turtles? Fish. Bull. 101, 189–193.
R Core Team (2021). R: A language and environment for statistical computing. Available online at: https://www.R-project.org/. [Accessed August 30, 2024].
Rudloe A., Rudloe J. (2005). Site specificity and the impact of recreational fishing activity on subadult endangered Kemp’s ridley sea turtles in estuarine foraging habitats in the northeastern Gulf of Mexico. Gulf Mex. Sci. 23, 2. doi: 10.18785/goms.2302.05
Sasso C. R., Epperly S. P. (2007). Survival of pelagic juvenile loggerhead turtles in the open ocean. J. Wildl. Manage. 71, 1830–1835. doi: 10.2193/2006-448
Sasso C. R., Epperly S. P., Johnson C. (2011). Annual survival of loggerhead sea turtles (Caretta caretta) nesting in peninsular Florida: A cause for concern. Herpetol. Conserv. Bio. 6, 443–448.
Schaefer K. M., Fuller D. W., Aires-da-Silva A., Carvajal J. M., Martínez-Ortiz J., Hutchinson M. R. (2019). Postrelease survival of silky sharks (Carcharhinus falciformis) following capture by longline fishing vessels in the equatorial eastern Pacific Ocean. B. Mar. Sci. 95, 355–369. doi: 10.5343/bms.2018.0052
Schaefer K., Fuller D., Castillo-Geniz J. L., Godinez-Padilla C. J., Dreyfus M., Aires-da-Silva A. (2021). Post-release survival of silky sharks (Carcharhinus falciformis) following capture by Mexican flag longline fishing vessels in the northeastern Pacific Ocean. Fish. Res. 234, 105779. doi: 10.1016/j.fishres.2020.105779
Schultz E. A., Cook M., Nero R. W., Caillouet R. J., Reneker J. L., Barbour J. E., et al. (2022). Point of no return: Determining depth at which sea turtle carcasses experience constant submergence. Chelonian Conserv. Bi. 21, 88–97. doi: 10.2744/CCB-1518.1
Sequeira A. M. M., Heupel M. R., Lea M.-A., Eguíluz V. M., Duarte C. M., Meekan M. G., et al. (2019). The importance of sample size in marine megafauna tagging studies. Ecol. Appl. 29, e01947. doi: 10.1002/eap.1947
Snoddy J. E., Landon M., Blanvillain G., Southwood A. (2009). Blood biochemistry of sea turtles captured in gillnets in the Lower Cape Fear River, North Carolina, USA. J. Wildl. Manage. 73, 1394–1401. doi: 10.2193/2008-472
Snoddy J. E., Williard A. S. (2010). Movements and post-release mortality of juvenile sea turtles released from gillnets in the lower Cape Fear River, North Carolina, USA. Endang. Species. Res. 12, 235–247. doi: 10.3354/esr00305
Spotila J. R., Reina R. D., Steyermark A. C., Plotkin P. T., Paladino F. V. (2000). Pacific leatherback turtles face extinction. Nature 405, 529–530. doi: 10.1038/35014729
Stokes L. W., Epperly S. P., McCarthy K. J. (2012). Relationship between hook type and hooking location in sea turtles incidentally captured in the United States Atlantic pelagic longline fishery. B. Mar. Sci. 88, 703–718. doi: 10.5343/bms.2011.1074
Strøm J. F., Rikardsen A. H., Campana S. E., Righton D., Carr J., Aarestrup K., et al. (2019). Ocean predation and mortality of adult Atlantic salmon. Sci. Rep. 9, 7890. doi: 10.1038/s41598-019-44041-5
Sulikowski J. A., Golet W., Hoffmayer E. R., Driggers W. B., Natanson L. J., Carlson A., et al. (2020). Observing post-release mortality for dusky sharks, Carcharhinus obscurus, captured in the U.S. pelagic longline fishery. Fish. Res. 221, 105341. doi: 10.1016/j.fishres.2019.105341
Swimmer Y., Arauz R., McCracken M., McNaughton L., Ballestero J., Musyl M., et al. (2006). Diving behavior and delayed mortality of olive ridley sea turtles Lepidochelys olivacea after their release from longline fishing gear. Mar. Ecol. Prog. Ser. 323, 253–261. doi: 10.3354/meps323253
Swimmer Y., Empey Campora C., McNaughton L., Musyl M., Parga M. (2014). Post-release mortality estimates of loggerhead sea turtles (Caretta caretta) caught in pelagic longline fisheries based on satellite data and hooking location. Aquat. Conserv.: Mar. Freshw. Ecosyst. 24, 498–510. doi: 10.1002/aqc.2396
Therneau T. M., Lumley T., Atkinson E., Crowson C. (2021). Survival Analysis. Available online at: https://github.com/therneau/survival. (accessed [August 30, 2024]).
Tracey S. R., Hartmann K., Leef M., McAllister J. (2016). Capture-induced physiological stress and postrelease mortality for Southern bluefin tuna (Thunnus maccoyii) from a recreational fishery. Can. J. Fish. Aquat. Sci. 73, 1547–1556. doi: 10.1139/cjfas-2015-0516
Valente A. L. S., Parga M. L., Velarde R., Marco I., Lavin S., Alegre F., et al. (2007). Fishhook lesions in loggerhead sea turtles. J. Wildl. Dis. 43, 737–741. doi: 10.7589/0090-3558-43.4.737
Wallace B. P., Kot C. Y., DiMatteo A. D., Lee T., Crowder L. B., Lewison R. L. (2013). Impacts of fisheries bycatch on marine turtle populations worldwide: Toward conservation and research priorities. Ecosphere 4, 40. doi: 10.1890/ES12-00388.1
Wallace B. P., Lewison R. L., McDonald S. L., McDonald R. K., Kot C. Y., Kelez S., et al. (2010). Global patterns of marine turtle bycatch. Conserv. Lett. 3, 131–142. doi: 10.1111/j.1755-263X.2010.00105.x
Watson J. W., Epperly S. P., Shah A. K., Foster D. G. (2005). Fishing methods to reduce sea turtle mortality associated with pelagic longlines. Can. J. Fish. Aquat. Sci. 62, 965–981. doi: 10.1139/f05-004
Williard A., Parga M., Sagarminaga R., Swimmer Y. (2015). Physiological ramifications for loggerhead turtles captured in pelagic longlines. Biol. Lett. 11, 20150607. doi: 10.1098/rsbl.2015.0607
Keywords: bycatch, loggerhead sea turtle, fisheries mortality, Canada, longline, satellite telemetry, archival tag, PSAT
Citation: James MC, Hall KE, Bond EP, Sherrill-Mix S and Plot V (2024) Post-release survival of loggerhead sea turtles (Caretta caretta) incidentally hooked in a North Atlantic pelagic longline fishery. Front. Mar. Sci. 11:1392582. doi: 10.3389/fmars.2024.1392582
Received: 27 February 2024; Accepted: 12 August 2024;
Published: 19 September 2024.
Edited by:
Mark E. Bond, Florida International University, United StatesReviewed by:
Ryan Knotek, New England Aquarium, United StatesYonat B. Swimmer, National Oceanic and Atmospheric Administration, United States
Copyright © 2024 James, Hall, Bond, Sherrill-Mix and Plot. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michael C. James, TWlrZS5KYW1lc0BkZm8tbXBvLmdjLmNh
†Present address: Emily P. Bond, Canadian Sea Turtle Network, Halifax, NS, Canada