
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 07 June 2024
Sec. Marine Fisheries, Aquaculture and Living Resources
Volume 11 - 2024 | https://doi.org/10.3389/fmars.2024.1375269
 Xosé A. Padin1*
Xosé A. Padin1* José M. F. Babarro1
José M. F. Babarro1 Pablo Otero2
Pablo Otero2 Miguel Gilcoto1Trinidad Rellán1Lino Suárez3
Miguel Gilcoto1Trinidad Rellán1Lino Suárez3 Anton Velo1
Anton Velo1 Laura G. Peteiro1*
Laura G. Peteiro1*A general decline in foundation species at the rocky intertidal has been observed during the last decades all around the world and primarily related to climate change. In agreement with that trend, the mussel aquaculture sector in Galicia (NW Spain), the main production area in Europe, has warned over the last years about a decline in the availability of wild mussel seed from the rocky coast. Here we compile for the first time, mussel seed collection reports by mussel farmers in Galicia for the period 2006–2021. We employed that dataset as a proxy of mussel recruitment evolution in the rocky shore for the last 16 years. Temporal analysis of our data confirmed the reported decline (-148 t yr-1), particularly pronounced from 2012 onwards. The data base also allowed us to analyze inter-annual variability according to both, climatic variations and management scenarios. Since cultivated mussels conform a meta-population with wild mussels from the rocky shore, alterations on the market preferences towards smaller individuals at harvest, could also contribute to a reduction in reproductive output. Our results show a decrease in life-time egg production under certain scenarios. Nonetheless, coastal upwelling seems to be the largest factor conditioning recruitment abundance, explaining as much as 60% of the variability observed. Decline on recruitment abundance was highly modulated by the observed increment in frequency of intense upwelling events, exceeding 500 m3 km-1 s-1 between July and November. Meridional winds also determined the spatial recruitment patterns, pointing to the large role of wind forcing on mussel larval dispersal. Our results highlights how alterations on upwelling regimes related to climate change can interact with mussel population dynamics and also condition aquaculture sustainability and food security.
Marine aquaculture is expected to provide a large part of the 60% increase in food production necessary to feed the world population by 2050 (Science Advice for Policy by European Academies, 2017). To ensure the growth of sustainable aquaculture, the increase should focus particularly on offshore aquaculture and the production of filter feeders (+100 Mt) (Science Advice for Policy by European Academies, 2017). Among these, the cultivation of bivalves has a minimal environmental footprint with lower CO2 emissions per gram of edible protein than any marine animal production (Froehlich et al., 2018; Kim and Kim, 2020; Gephart et al., 2021) and it provides important benthic and coastal ecosystem functions (Wilberg et al., 2011; Froján et al., 2016). Considering these factors, bivalve farming is an attractive alternative for the future expansion of sustainable seafood (Smaal et al., 2019; Costello et al., 2020; Gentry et al., 2020) and a substitute for the stagnant global fisheries production (Science Advice for Policy by European Academies, 2017).
The production of mussels which are the main aquaculture species in Europe, has reported a significant decreasing trend over the past two decades, mainly in northern countries (Avdelas et al., 2021). This decline has not yet been reported in Spanish mussel production, which is the first European producer with 90% of its production coming from Galicia (NW Spain; Labarta and Fernández-Reiriz, 2019). Nonetheless, forecasts from the Spanish Ministry of Agriculture, Fisheries and Food do point to a future stagnation by 2027 (SGP-MAPA, 2022).
The decline in European mussel cultivation has been attributed to several factors, including diseases, harmful algal blooms, and the impacts of climate change, as highlighted by previous studies (Álvarez-Salgado et al., 2009; Rodrigues et al., 2015; Fuentes-Santos et al. 2021; Outeiro et al., 2018). Another critical factor often cited for the decline in mussel production is the low and unpredictable availability of mussel seed (Filgueira et al., 2007; Avdelas et al., 2021). The dependence of current aquaculture on wild mussel seed stocks, which exhibit significant interannual fluctuation, compromises the sustainability of production in Northern European countries (Avdelas et al., 2021). Population dynamics shifts and mussel seed scarcity have been linked to global change (Handisyde et al., 2017; Petraitis and Dudgeon, 2020; Baden et al., 2021). Global change encompasses not only seawater warming but also a series of cascading effects across different temporal and spatial scales, potentially impacting the replenishment of natural populations (Scheffers et al., 2016).
The supply of mussel seed has not been considered a limiting factor for mussel (Mytilus galloprovincialis) cultivation on the Galician coast until recently. The main source of mussel seed comes from exposed intertidal rocky shores (Mariño et al., 1982; Pérez-Camacho et al., 1991), and it is tightly linked to the spawning of cultivated mussels, which concentrate the largest adult populations in Galicia (Fernández-Pulpeiro et al., 2002; Fernández-Pulpeiro and Aldariz, 2003). However, mussel farmers in the region have increasingly perceived a decrease in mussel seed availability from the rocky shore during the latter years of 2010s. This concern, communicated through the media, highlighted the lack of adequate spat stock monitoring programs to assess spatio-temporal variability in annual mussel seed availability and underscored the need for more flexible management strategies for its collection. Nonetheless, mussel farmers are required to officially report their annual mussel seed collections (tonnes and origin of the seed outplanted on their rafts), which they do with considerable variation in the frequency and detail on the information provided. However, these reports are the sole source of information on the mussel seed stock status in Galicia, and constitute a very valuable long-term time-series of mussel recruitment data. This data has not been systematically analyzed yet to link environmental factors with mussel seed spatio-temporal variability.
In addition to the possible impact of the changing environment, the recent scarcity of mussel seed in Galicia could also be related to the lower production of larger individuals, which may affect the reproductive efficiency of populations and lead to recruitment failures. The progressive preference for smaller individuals in the fresh mussel market has steadily increased over the last two decades, accounting for more than 60% of production in 2015 (Labarta and Fernández-Reiriz, 2019). This shift in production, driven by consumer preferences, also offers significant economic benefits for mussel growers by reducing culture cycles and losses, occasionally massive, attributed to dislodgement events during cultivation (Pérez-Camacho et al., 2013; Babarro et al., 2020). The shortening of cultivation timing could limit or reduce the number of spawning opportunities and optimal periods for reproduction before adult harvesting. In addition, adult size exponentially determines the number of gametes that can be produced which is proportional to spawner size (Oliveira et al., 2021) and could also determine its viability (Sukhotin and Flyachinskaya, 2009). Nonetheless, the analysis of the impact of harvesting progressively smaller mussels on the potential reproductive output of their populations and, consequently, the processes of fertilization and viability of larvae has not been investigated in detail for mussel culture in Galicia.
Therefore, our goal is to investigate the impact of both, environmental variability and shifts in the cultivation standards over the past two decades. We aim to understand their links to the interannual variability observed in mussel seed collection, using this as a proxy to assess the status and replenishment of natural mussel populations. With this purpose, we have analyzed, for the first time, the mussel seed collection reports from mussel farmers, compiling a 16-year time series of mussel seed recruitment. This information provides an updated assessment of the amount of mussel seed used in Galician mussel farming to sustain the current production levels and allows to identify the main environmental factors that explain its interannual variability. In addition, the spawning stock was also estimated to infer the impact of reducing the amount of large reproducers on the availability of mussel seed along the rocky shore. This approach deepens our understanding of how management practices and climatic alterations contribute to the scarcity of mussel seed. This will help to develop dynamic management measures that balance profitability with the long-term sustainability of mussel aquaculture, paying special attention to the interaction between climatic conditions and the maintenance of the natural stocks.
Mussel aquaculture in Spain is primarily concentrated in rias, coastal bays of tectonic origin, located on the Galician coast (NW of the Iberian Peninsula; Figure 1). Among these embayments, the four rias located in the southern part of the Galician coast, named the “Rías Baixas”, stand out for their high mussel production. The high productivity of these rias is largely influenced by upwelling regimes and river discharges (Álvarez-Salgado et al., 2000; Figueiras et al., 2002).
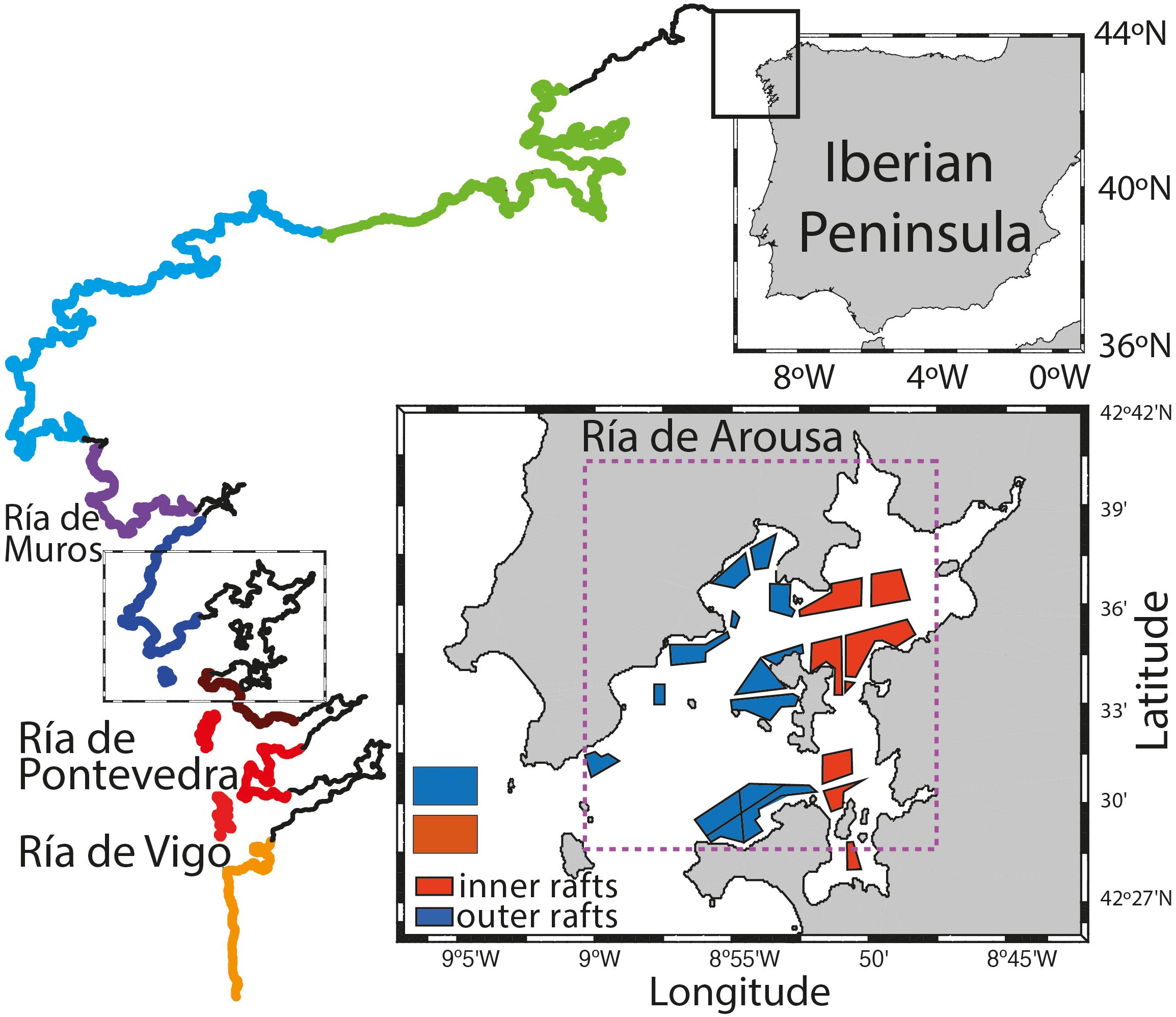
Figure 1 Map of the Galician coast with the coastal sectors for mussel seed collection (see Figure 8) and map of the Ría de Arousa, including the outer and inner mussel farming areas, and the window used for obtaining satellite measurements of temperature and chlorophyll.
The coast of Galicia represents the northern limit of the Canary Current Eastern Boundary Upwelling Ecosystems (Arístegui et al., 2009; Chavez and Messié, 2009). On the west coast of Galicia, due to coastal upwelling dynamics, the northerly wind component drives the surface waters offshore, inducing the rise of deeper, cooler, and nutrient-rich subsurface waters close to the coast. Southerly winds have the opposite effect, they induce coastal downwelling that piles up warmer, nutrient-poor surface waters near the coastline. Throughout the seasonal cycle, northerly winds and coastal upwelling dominate from April to September while southerly winds and downwelling prevail during fall and winter (Pardo et al., 2011). The primary production associated with the coastal upwelling of nutrient-rich waters (Pauly and Christensen, 1995) makes the Galician coast (Arístegui et al., 2009; Santos et al., 2011) one of the most productive regions in the oceans and ideal for the aquaculture of marine bivalves (Kämpf and Chapman, 2016; FAO, 2018; Varela et al., 2018).
Mussel aquaculture in the Galician coast is carried out in 3387 floating rafts, which are rectangular wooden grids with a maximum surface area of 550 m2 and a maximum lateral length of 27 meters. 500 ropes of 12 meters length are hung from each raft (Decree 174/2002 amending Decree 406/1996). The ria named “Ría de Arousa” is the largest of the Rías Baixas covering an area of 245 km2 and hosting 2319 mussel rafts, 68% of total rafts in Galicia, distributed in several polygons (Figure 1). Other rias such as the “Ría de Vigo” and “Ría de Pontevedra” are also important areas of mussel production accounting for 14% and 10% of total rafts, respectively.
Mussel cultivation begins with the outplanting of mussel seed obtained from the intertidal rocky shore or from subtidal suspended collectors (Figueira et al., 2007). At the end of the last century, approximately 65% of the producers used exclusively seed from the rocky shore, while 10% employed only seed from collector ropes and the rest combined the two seed sources (Mariño et al., 1982; Pérez-Camacho et al., 1991). Thus, the intertidal rocky shore is the main source of seed for mussel aquaculture in Galicia (Cáceres-Martínez et al., 1993; Molares and Fuentes, 1995; Brea Bermejo, 2009). The extraction of this wild seed is permitted from December to April, when mussel farmers are allowed to collect a maximum of 3500 kg of mussel seed per raft through manual scraping of wild seed on rocks, following the regulations set by the Regional Administration (DOG N° 228 Order of 26th October, 2000).
Although mussel larvae can be found throughout the year at these latitudes, the reproductive cycle of mussels is characterized by a main peak of spawning during spring followed by a minor peak during the autumn (Villalba, 1995; Philippart et al., 2012; Philippart et al., 2014). The pelagic life of mussel larvae has an average duration of 4 weeks. After that time, they would search for a settlement substrate to metamorphose on the rocky shore (Seed, 1969; Aguirre, 1979; Widdows, 1991; Cáceres-Martínez and Figueras, 1998). Therefore, mussel seed harvested from the rocky shore during winter and early spring corresponds mainly to the autumn spawning peak, while the offspring of the main spring spawning peak is collected in the collector ropes of the rafts.
Mussel seed collection reports were provided by mussel producers belonging to 6 associations of the Ría de Arousa, integrated into the OPMEGA organization (Organización de Productores de Mexillón de Galicia). These reports offer information on the amount of mussel seed harvested for each date, detailing the location of extraction and the raft of destination of the mussel seed. For this study, reports covering the period from the 2005–2006 to the 2021–2022 campaign (December to April) were used. Hereafter, these consecutive campaigns will be referenced by their closing year, thus referring to a study period from 2006 to 2022.
In total, 17363 records on mussel seed collections have been gathered over the entire study period. Inputs reporting the maximum authorized amount of mussel seed (3500 kg) on a single date have been excluded from the original database (totaling 261 entries) because of their lack of reliability. Accepted records were then grouped by destination raft for each campaign, resulting in a final database of 3720 annual reports of wild mussel seed harvested for individual rafts (Table 1). To account for the oceanographic heterogeneity of the Ría de Arousa (Alvarez-Salgado et al., 1996; Rosón, 1997), and differences in mussel productivity along the Ría (Pérez-Camacho et al., 2014; Babarro et al., 2018, Babarro et al., 2020; Padin et al., 2021), rafts were split into two distinct regions: an outer zone with 1262 rafts under greater oceanic influence and an inner, less productive, sheltered zone with 1057 rafts with higher river influence (Ulla and Umia rivers). This division ensured the availability of mussel seed collection reports from each raft group in every seed collection campaign (Table 1).
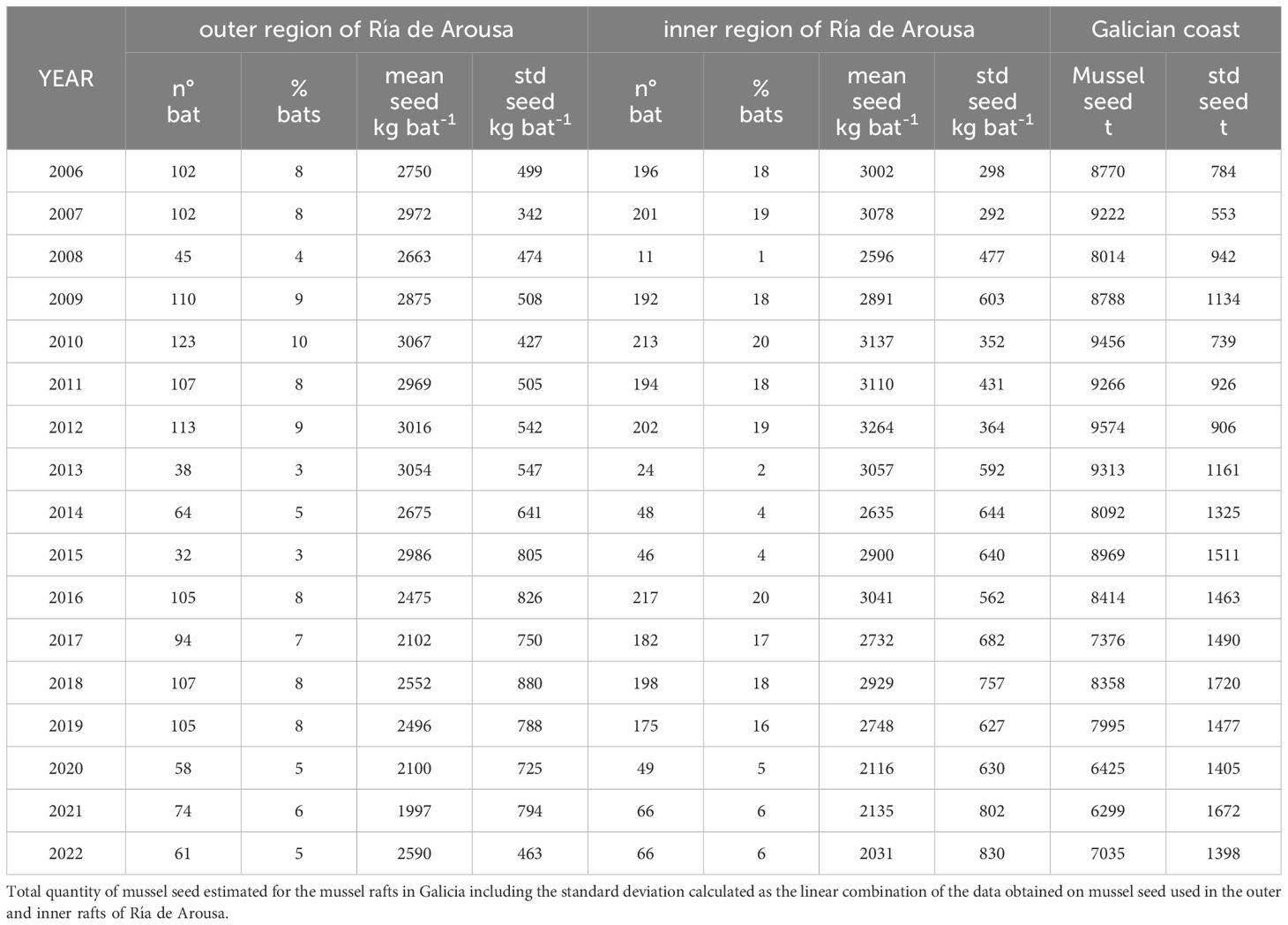
Table 1 Annual summary of the reports in the database including: number of mussel rafts, percentage of the total mussel rafts in the area, average quantity and standard deviation of mussel seed collected for each raft in the outer and inner areas of the Ría de Arousa.
The quantity of mussel seed collected annually from the reports of the Ría de Arousa was extrapolated to the total number of mussel rafts in Galicia, classified equally as outer and inner rafts, resulting in 1849 and 1538, respectively. A total of 339 rafts were excluded from the outer rafts group to account for the 10% of producers which exclusively employ seed from collector ropes, which are usually located in the outer areas of the embayments (Mariño et al., 1982; Pérez-Camacho et al., 1991). Therefore, the average values for the inner and outer rafts of the Ría de Arousa were extrapolated to a total of 1510 outer and 1538 inner rafts to estimate the total amount of wild mussel seed used by the aquaculture sector in Galicia.
The geographical location of the wild seed collection sites was mainly provided with colloquial names (headlands, nearby beaches, islands, coastal stones, close-by towns…) without including geographical coordinates. The spat collection area used by the mussel farmers of the Ría de Arousa extends along the entire Atlantic coast of Galicia, covering more than 300 km. This coast was divided into 7 continuous segments of outer rocky shore areas (Figure 1). The quantities of mussel seed declared in each report were allocated among the corresponding sectors.
The average distance (km) from the headquarters of each association, that provided their mussel collection reports, to the center of each of the mussel seed collection sector was calculated using Google Maps. Generalized additive models (GAM), as implemented in the mgcv library of R 3.6.2 (R Core Team, 2018), were used to investigate the relationship between the amount of mussel seed collected at each sector and the distance by road between the association and the center of each segment. Model validation included the verification of homogeneity, normality, and independence assumptions (Zuur et al., 2009).
Mussel production data are available at www.pescadegalicia.gal disaggregated by market destination in fresh and processed production. While mussels for the processing industry (canned, frozen, etc.) lack the information about specific size at harvest, mussels for fresh consumption are classified into three size categories according to the number of mussels per kg: small mussels (35 - 70 pieces kg-1), medium (24 - 35 pieces kg-1) and large (<24 pieces kg-1). Using the relationship between weight (W) and antero-posterior shell length (L) (L(mm)=23.398 W(g)0.3377; r2 = 0.9811), obtained from monitoring of mussel growth in the Ría de Arousa during several cultivation cycles (unpublished data), the mean extrapolated shell length for each size category was 62.83 mm, 76.14 mm and 85.27 mm for small, medium and large mussels, respectively. Subsequently, these lengths were used to estimate the potential egg production of each individual during the spring spawning according to the formula:log (N° Eggs mussel-1) = -1.43 + 2.99 logL (mm), R2 = 0.51 extracted from Oliveira et al. (2021) as the average of the northernmost populations evaluated, which are the most similar to the Galician coast. Autumn spawning capability was calculated with the same relationship, but dividing the result by 2 to account for the reduction of intensity of the autumn spawning peak with regard to the spring one (Philippart et al., 2012).
Following these assumptions, the potential egg production during each of the spawning peaks was estimated for an average production of 140K tonnes of mussels declared for fresh consumption during the 2006–2022 period. Since we assume mussel rafts are mainly seeded during winter, mussels are considered immature during their first spring under cultivation. Therefore, small mussels at harvest (8–10 months old) were considered to have only one spawning opportunity during the autumn peak (Aguirre, 1979), while medium (14–16 months old) and large sized mussels at harvest (17–19 months old) would contribute to both spawning peaks, first autumn and second spring under cultivation.
In order to estimate errors, the error propagation equation was applied (Bevington and Robinson, 2003) to each calculation step or transformation. The error for percentages of size category were estimated using the 25% of the standard deviation (6.25% of the variance) of the original time series and a 6.25% of the time covariance with the total fresh mussel production as initial errors. For the initial error of the number of mussel pieces per kilogram and size category a 25% of the size range for each category was used. Finally, for both sets of coefficients in the weight-length and length-egg potential equations a 1% of the absolute values of the coefficients was used to estimate their initial standard deviation.
The upwelling index at the geographical coordinates 42°N 10°W representing the region of interest, was obtained for the period between 2005 and 2021 from the Instituto Español de Oceanografías (http://www.indicedeafloramiento.ieo.es). This index is computed following Lavín et al. (1991) and based on 6-hourly geostrophic winds obtained from the Atmospheric Prediction System (NOGAPS) model, maintained by the Fleet Numerical Meteorology and Oceanography Center (FNMOC). This upwelling index has been previously identified as suitable for describing the oceanographic variability in the Rías Baixas (Gonzalez-Nuevo et al., 2014).
Chlorophyll-a concentration (Chl-a) and sea surface temperature (SST) were obtained from L4 satellite products with a spatial resolution of 0.05°. Whereas Chl-a was obtained from the Atlantic Ocean Color (GlobColour) Bio-Geo-Chemical product, SST was extracted from the European North West Shelf/Iberia Biscay Irish Seas product, both available at the Copernicus Marine Service (https://data.marine.copernicus.eu/). L4 products were selected because they are produced using a consolidated and consistent input dataset and have sufficient temporal coverage. The Chl-a and SST daily mean were estimated for the Ría de Arousa area, between 42.66°N - 42.46°N and 9°W - 8.78°W (Figure 1).
Daily discharge data for the main rivers flowing in the Ría de Arousa (Ulla and Umia) were obtained through the Spanish regional organization Augas de Galicia (https://augasdegalicia.xunta.gal/). Further information about gauging stations and the methodology can be obtained in Otero et al. (2010).
On one hand, temporal trends in environmental variables were explored with the Mann-Kendall test and Sen’s slope computed using the pyMannKendall library (Hussain and Mahmud, 2019). A variant of this test, the Correlated Seasonal Mann-Kendall test (Hipel and McLeod, 1994), was performed when seasonality and autocorrelation were present in the time series. On the other hand, Rodionov (2004) methodology was applied to detect level shift, based on a sequential t-test analysis on the time series. Statistically significant deviations from the mean value were calculated with a software provided by NOAA (http://www.beringclimate.noaa.gov/regimes/index.html). A 10-year cut-off length and a Huber parameter of 1 were set (deYoung et al., 2004). Pre-whitening was performed before shift analysis to prevent autocorrelation (Rodionov, 2004). It is important to note that shifts detected close to the tails of the time series must be interpreted with caution under this methodology. A 0.05 significance level was set in both analyses.
In addition, oceanographic conditions during the larval phase prior to recruitment on the rocky shore were summarized for each of the years where mussel seed collection reports were available. Taking into account the seasonal presence of mussel larvae (Philippart et al., 2012), an average pelagic larval duration of 30–40 days (Cáceres-Martínez and Figueras, 1998) and post-settlement growth rates of around 0.23 mm day-1 for early recruits (Peteiro et al., 2007), it can take 4 to 6 months from the time of spawning to reach an appropriate size for mussel seed collection (< 2 cm). Therefore, the seed collected between December and April was considered as planktonic between July and November. SST, Chla-a and river flow between July and November were characterized from their median values for each year. The upwelling index between July and November of each year was summarized as the sum of the daily values of the upwelling index greater than 500 m3 km-1 s-1. This value nearly coincides with the median of positive daily values of the upwelling index (493 m3 km-1 s-1) during the period 2005–2021 and has been previously described as a threshold for detrimental effects on mussel settlement at certain locations (Peteiro et al., 2011) and the value at which the upwelling index induces the greatest change in the water column temperature (Otero et al., 2023). Simple linear regressions were employed to test for relationships between the interannual variability of the total amount of coastal wild spat collected for seeding in rafts and the oceanographic conditions observed during the larval phase prior to recruitment on the rocky shore.
Finally, the sum of the meridional wind component from July to November each year was calculated to explore correlations (Pearson’s Correlation Coefficients) between the prevalent wind direction along-shore of each year and the amount of mussel spat collected at each of the 7 continuous segments of rocky coast designated.
The upwelling index computed in the outer waters of the Rías Baixas, SST and Chl-a estimations from satellite sensors and the fluvial discharge of the Umia and Ulla rivers were analyzed for the period 2005–2021. Monthly climatology values showed a marked seasonal pattern of these variables (Figure 2).

Figure 2 Boxplot of the seasonal variability of the (A) upwelling index, (B) sea surface temperature, (C) chlorophyll concentration, and (D) river discharge of Umia-Ulla rivers in the Ría de Arousa.
The upwelling index showed positive mean values between March and September with a maximum in August of 647 ± 761 m3 km-1 s-1 when northerly winds dominated (Figure 2A). In contrast, the autumn and winter seasons were characterized by negative values, especially in December with -454 ± 1027 m3 km-1 s-1 under the dominance of southerly winds. The variability of the upwelling index throughout the seasonal cycle was generally lower during the months with positive coastal upwelling (Figure 2A).
The daily SST of the Ría de Arousa was 15.05 ± 1.87°C with a total range between 10.67°C and 20.27°C (Figure 2B). The monthly mean temperature was highest in July with 17.2 ± 0.8°C while the coldest waters of 12.7 ± 0.5°C were observed in February. Chl-a concentration (Figure 2C), which had a mean value of 7.24 ± 6.90 mg m-3, showed the highest values during the upwelling season, with a peak of 9.6 ± 6.6 mg m-3 in August. In contrast, lower and less variable Chl-a was recorded during the autumn and winter, with a minimum mean value of 3.2 ± 1.9 mg m-3 in January. The joint discharge of the Umia and Ulla rivers (Figure 2D) showed a strong seasonal cycle with higher values from November to February, a period characterized by southerly winds and wet weather conditions, and a maximum of fluvial discharges in December.
Monthly mean variability 2005–2021 is shown (Figure 3) along with the 12-month moving average showing positive (red) and negative (blue) periods with respect to a zero baseline in the upwelling and to the mean in the Chl-a and SST series. The upwelling index alternated between positive and negative interannual anomalies shorter than 2 years between 2005 and 2015, with a clear predominance of positive anomalies in 2016–2021 (Figure 3A). In addition to a marked seasonal variability, the surface temperature in the Ría de Arousa also showed a succession of warm and cold interannual intervals throughout the study period (Figure 3B). These periods were mainly modulated by an inverse relationship with the interannual anomalies of the upwelling index. Periods of cool surface waters were coincident with positive upwelling index anomalies (2008–2009) while warm waters were observed during periods of particularly negative upwelling index (2006–2007). Nevertheless, the impact of upwelling did not seem to be as evident in the interannual temperature anomalies during other periods.
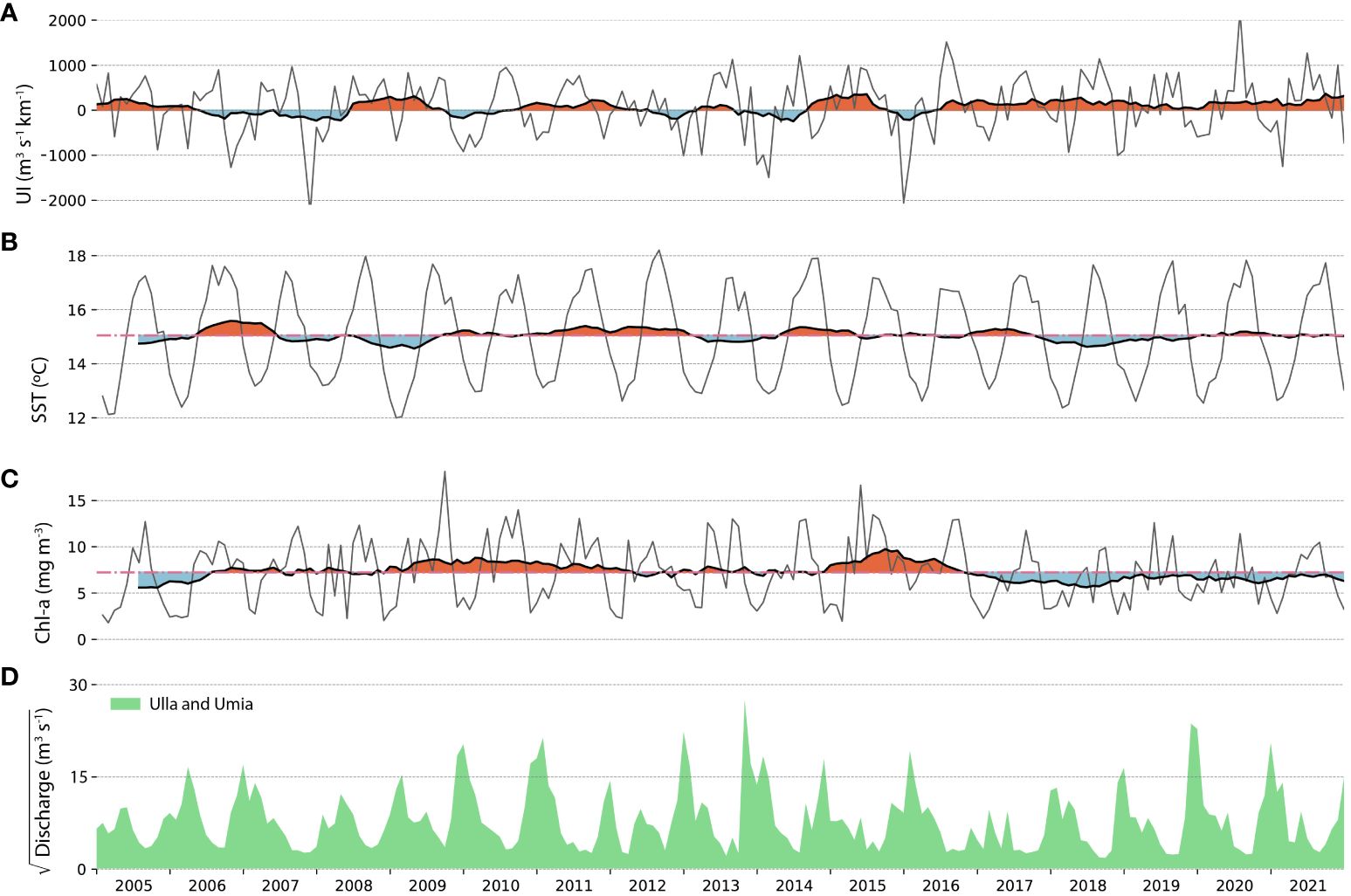
Figure 3 Monthly mean time series of (A) upwelling index, (B) chlorophyll concentration and (C) sea surface temperature in the Ría de Arousa and (D) the square root of the discharge of Umia-Ulla rivers instead of its mean monthly value in order to allow graphical visualization of maximum and minimum values. Red (blue) areas highlight positive (negative) anomalies from the mean in the Chl-a and SST, and from a zero-based line in the upwelling index. The black line is the 12-month average running mean of the time series.
Chl-a showed two clear phases throughout the study with a period between 2006 and 2015 dominated by positive anomalies and a final period with negative anomalies between 2016 and 2021 (Figure 3C).
None of these parameters showed a significant interannual trend in the period 2005–2021. Neither did the upwelling index and Chl-a, despite showing a significant shift in 2016.
A total of 3720 annual quantities of mussel seed collection from individual rafts were analyzed for the 17 years period (Table 1; Figure 4), with a large variability on the number of rafts recorded per year, which ranged from 56 to 336 for 2008 and 2018, respectively (Table 1). This information represented 7 ± 2% and 12 ± 7% (mean ± std) of the total number of 1262 outer and 1057 inner rafts in the Ría de Arousa, respectively. The average amount of mussel seed collected for the outer rafts (2679 ± 712 kg raft-1, n=1440) was significantly lower (two-sample z-test: p<0.05) than the average amount destined for the inner rafts (2917 ± 619 kg raft-1, n=2280) (Figure 4A). Despite this difference, the interannual variability is similar in the two groups with an initial increasing trend in mussel seed collection until 2012–2013 when maximum wild seed collection of 3264 kg raft-1 and 3054 kg raft-1 were observed for the inner and outer region, respectively (Figure 4B; Table 1). From these maximum values, mussel seed collection decreased until 2022 at a rate of -78 ± 31 kg raft-1 yr-1 in the outer rafts and -107 ± 22 kg raft-1 yr-1 in the inner rafts. This decreasing trend is also observed in a group of 12 singular rafts (10 outer and 2 inner) that provided data for the entire period 2006–2022. These rafts analyzed separately and standardized (Figure 4C), reinforce the decreasing trend of wild seed collected between 2013 and 2021 (Figure 4B).
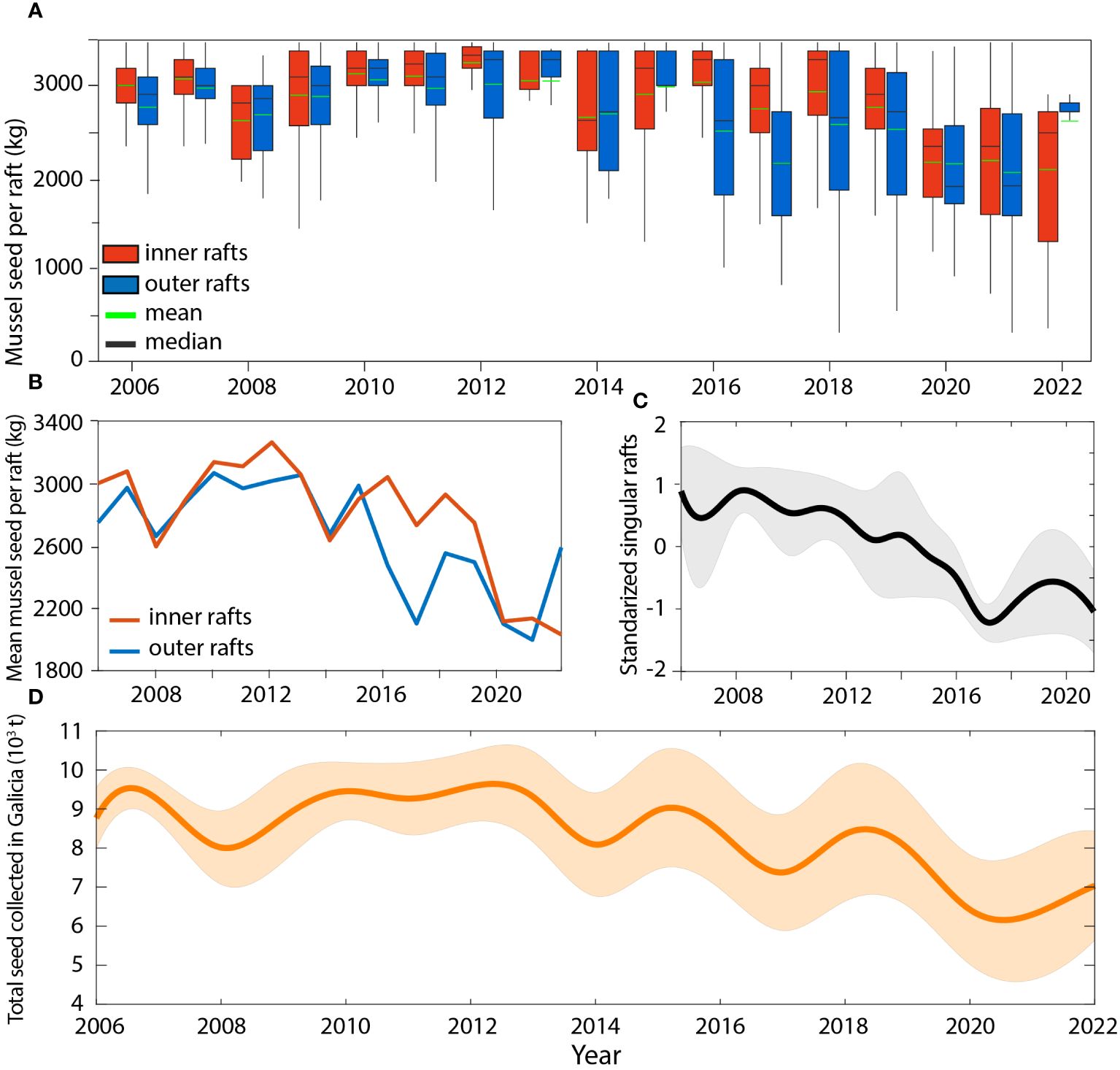
Figure 4 (A) Boxplot of the mussel seed quantity allocated to the inner and outer mussel farming areas of the Ría de Arousa. (B) Time series of the average mussel seed used in both outer and inner mussel rafts. (C) Time series of the mussel seed used in 12 rafts with data available for all years between 2006 and 2022. Values are normalized by dividing the difference between the reported quantities of wild seed and the mean value of mussel seed collected for each raft by the standard deviation. (D) Evolution of the total mussel seed quantity collected for all mussel rafts in Galicia based on Ría de Arousa data for all mussel rafts in Galicia.
The average amount of seed collected estimated for the total number of rafts on the Galician coast for the period 2006–2022 is 8316 ± 1027 tonnes (Figure 4D) with a maximum value of 9575 tonnes in 2012 and a minimum harvest of 6300 tonnes in 2021. A decreasing trend of -148 ± 36 t yr-1 (p-value<0.01, r2 = 0.53) characterized mussel seed availability on the Galician coast for the period 2006–2022 (Figure 4D; Table 1). This interannual decline was particularly pronounced from 2012 onwards with a rate of -288 ± 55 t yr-1 (p-value<0.01, r2 = 0.75).
With regard to the influence of oceanographic conditions during the larval phase on the interannual variability observed on the total amount of coastal wild spat collected for seeding in rafts only the sum of upwelling index values greater than 500 m3 km-1 s-1 and the median of the Chl-a concentration were significant (Figure 5).
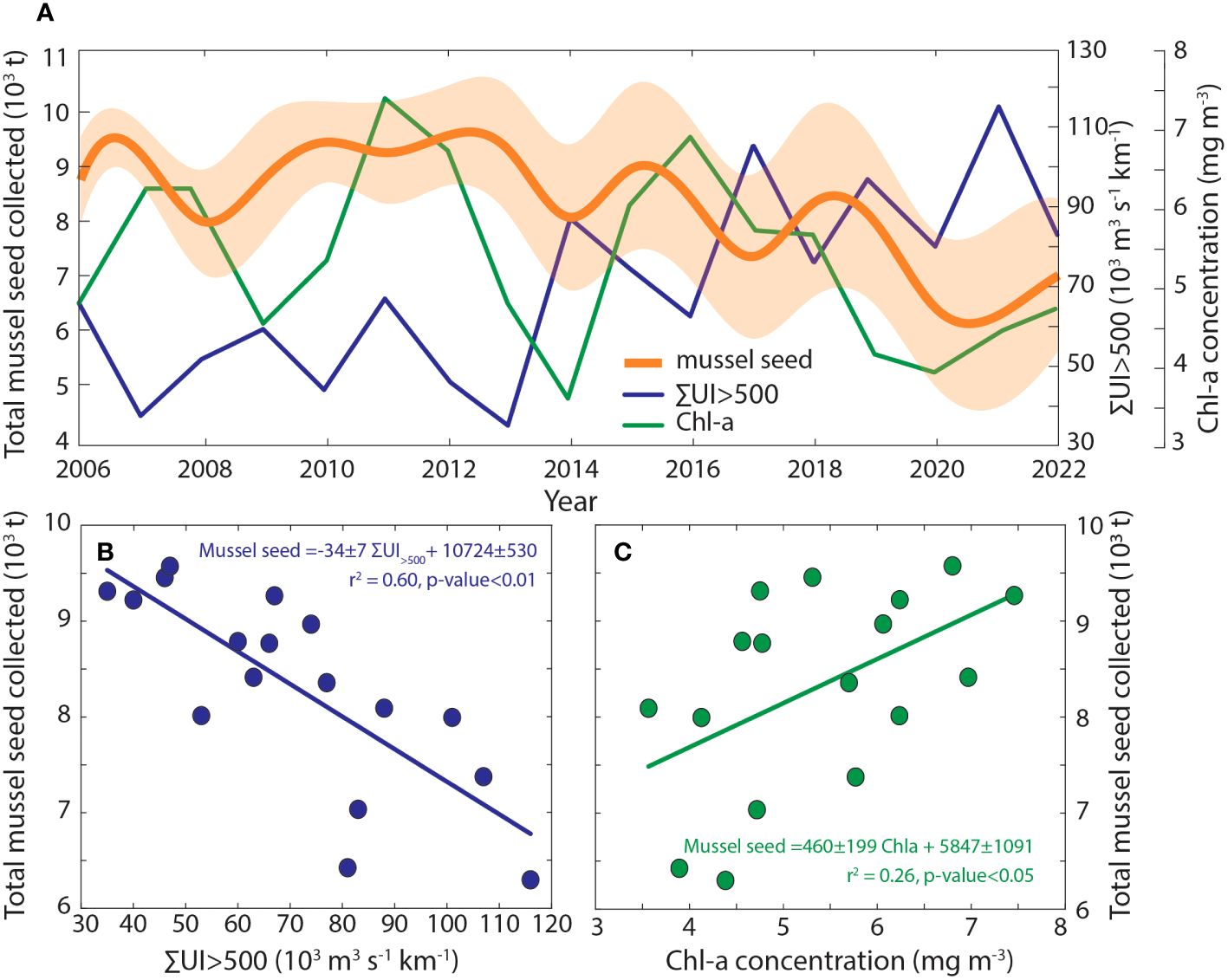
Figure 5 (A) Time series of the estimated mussel seed quantity for all mussel rafts in Galicia and the mean values of chlorophyll concentration and the sum of the upwelling index with values above 500 m3 km-1 s-1 between July and November. (B) Linear relationship between the total mussel seed quantity and the sum of the upwelling index with values above 500 m3 km-1 s-1 and (C) chlorophyll concentration.
The sum of upwelling index values greater than 500 m3 km-1 s-1 showed a significant negative correlation with the averaged amount of mussel seed collected (Figure 5A). The amount of intense events explained as much as 60% of the interannual variability observed on mussel seed collection along the Galician coast (r2 = 0.60; Figure 5B) In addition, the number of days with intense upwelling events showed an interannual increasing rate of 1.0 ± 0.4 days per year (r2 = 0.25; p<0.05) ranging from 38 to 70 days for the period between July and November between 2006 and 2022, and an average value of 51 days.
In contrast, the median of the Chl-a concentrations in the Ría de Arousa (Figure 5A) showed a positive relationship with the amount of mussel seed collected, explaining 26% of the interannual variability observed (r2 = 0.26; Figure 5C). On the other hand, SST and river discharge did not show significant correlations with the amount of mussel seed collected.
Mussel seed destined for the mussel raft cultivation in the Ría de Arousa was collected at more than 150 locations along ~300 km of the Atlantic coast of Galicia (Figure 1). Preferential sites for mussel seed collection of each association is mainly determined by distance (Figure 6A), with most of the seed collected within 50 km from the headquarters of the association and only 10% collected farther than 150 km. Accordingly, mussel farmers whose associations are located on the north shore of the Ría de Arousa were preferentially collecting seed from the northern sectors, while the opposite happened with the associations located on the south shore, which travelled preferentially to the southern locations. To analyze spatial variability of mussel seed availability during the study period without bias related to the distance to the association headquarters, only years with reports of associations from both shores of the Ría de Arousa were considered. The reported annual quantities of wild seed were weighted to equal the total amount of mussel collected from each shore before calculating the percentage of seed supplied from each segment of the Galician coast to the mussel rafts in the Ría de Arousa (Figure 6B).
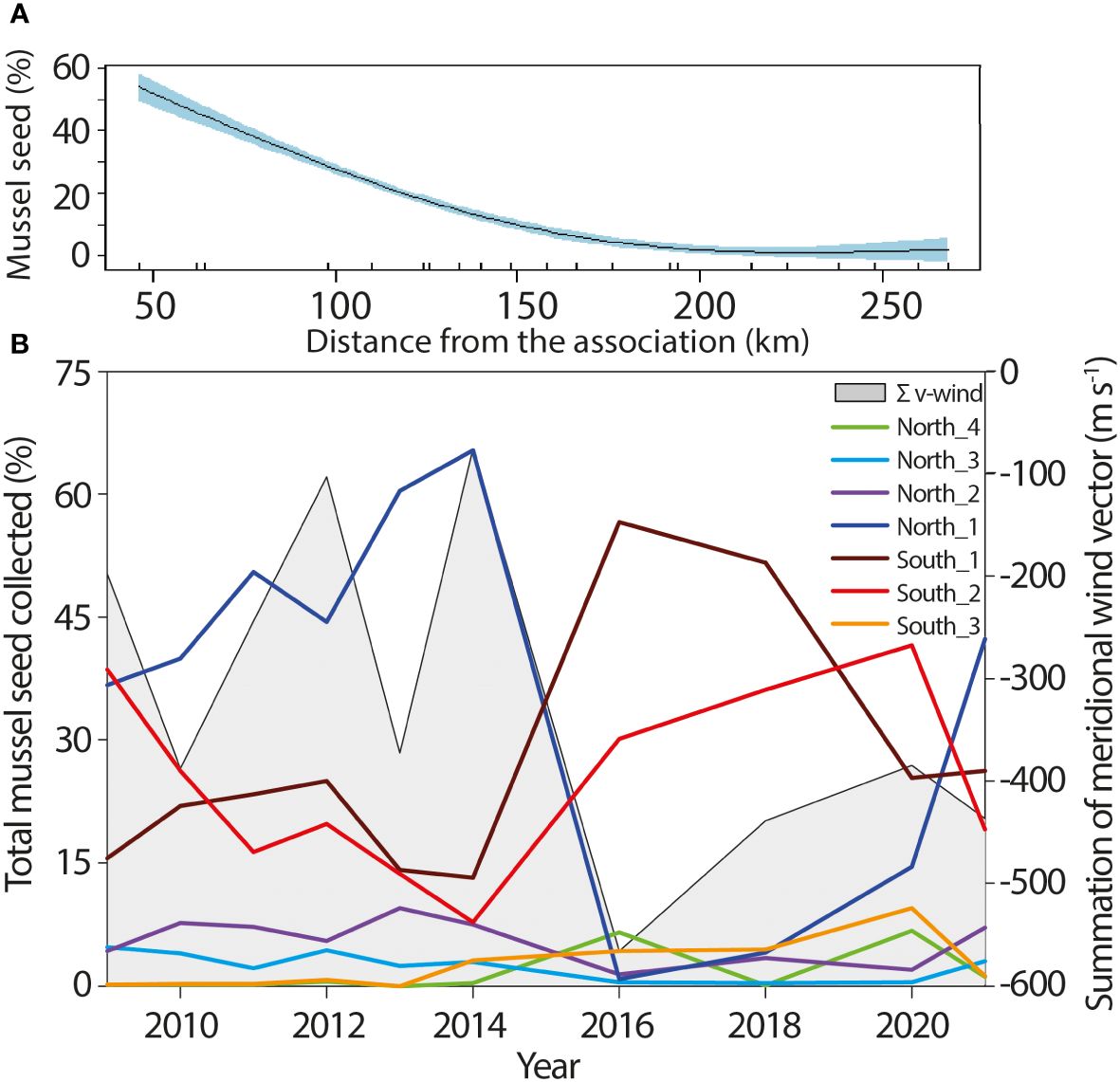
Figure 6 (A) Dependence of the percentage of mussel seed collected and the road distance from the mussel farmers’ association to the collection site. (B) Time series of the percentages of mussel seed collected in the coastal sectors described in Figure 1 and the sum of the meridional wind stress between July and November, where the lower negative values of Σv-winds indicate lower prevalence of northerly winds, since positive values of v indicate wind is coming from the South while negative values of v indicate wind is coming from the North.
The rocky shore immediately north (North_1) and south (South_1) of the Ría de Arousa were the preferred harvesting areas contributing 36 ± 22% and 27 ± 15% of the total mussel seeds respectively. The third most common origin was the southern shore of the Ría de Pontevedra (South_2) with 25 ± 11% while the southernmost area of the Galician coast (South_3) contributed only 2 ± 3% to the total seed collected. The areas to the north (North_2, North_3 and North_4) contributed 6 ± 3%, 2 ± 2% and 2 ± 3% of the total amount collected by mussel farmers of the Ría de Arousa, respectively. It is also worth noting that regular visits to the most distant locations (North 4 and South 3) only started to be visited regularly for mussel seed collection from 2013.
The amount of mussel seed collected within each coastal sector exhibited pronounced interannual variability. The collection of mussel seed between 2010 and 2014 was primarily conducted along the northern shoreline of Ría de Arousa (North_1), surpassing 60% in 2013 and 2014. Although with a lower contribution to the total amount of mussel seed collected, North_2 and North_3 sectors showed high positive correlation coefficients with the North_1 segment (r= 0.89 and r=0.65, respectively; Table 2). In contrast, the northernmost and more distant sector (North_4) exhibited a significant inverse correlation with regard to the rest of sectors to the north of Ría de Arousa (Table 2).
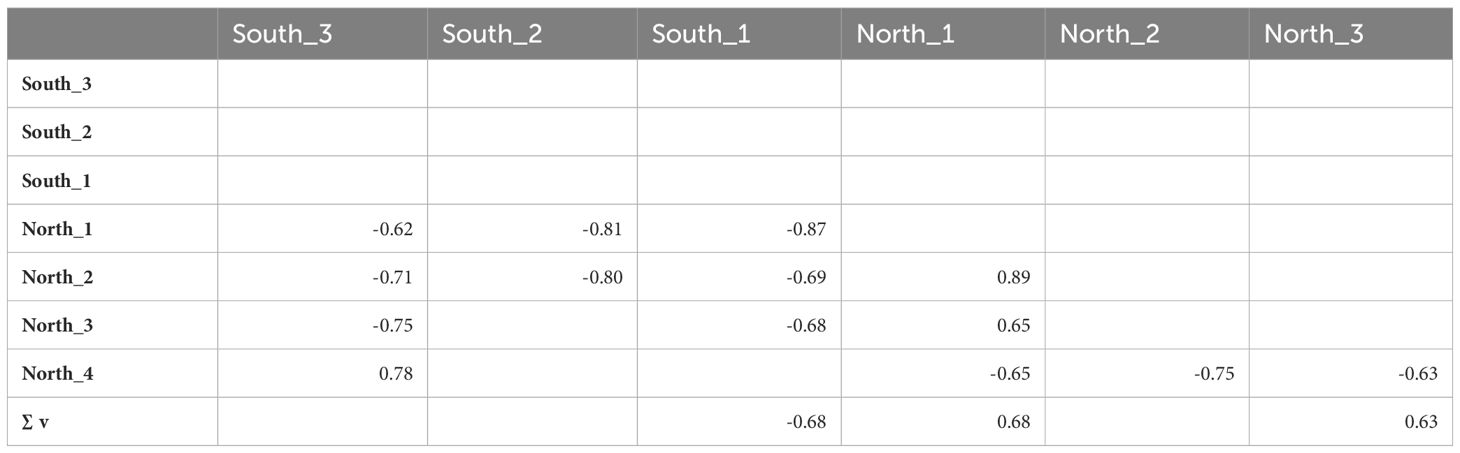
Table 2 Correlation matrix with the significant coefficients between the percentages of wild mussel seed collected in each of the coastal sectors between 2006–2022 and the sum of the meridional wind component (∑V) between July and November.
In the period 2016–2020, the main sector for harvesting wild seed shifted to the southern segments of Ría de Arousa. This transition was notably pronounced in 2016 when the southern shore of the Ría de Arousa (South_1) supplied a maximum value of 55% of the total mussel seed, while the northern shore (North_1) contributed only by 1% (Figure 6B). Even the sector encompassing the southern coast of Ría de Pontevedra and the northern region of Ría de Vigo (South_2) emerged as the preferred region for seed collection in the year 2020. During the latest campaign, 2022, the coastal segment that contributed the highest percentage was once again North_1 sector, although the quantities of wild seed harvested from the north and south of Ría de Arousa were similar. In general, there is an inverse relationship between the amount of mussel seed collected from the north and south sectors (Table 2). However, while the northern areas demonstrated some consistency in their interannual variability, the three southern segments of the Ría de Arousa, which represent the margins of important mussel production areas such as the Ría de Arousa, Ría de Pontevedra and the Ría de Vigo did not exhibit statistically significant correlations between them. An unexpected finding was the significant positive correlation (r=0.78) between the northern (North_4) and southern (South_3) sectors, located at the furthest points from Ría de Arousa.
The two distinct periods evidenced by contrasting mussel preference collecting zones also exhibited a clear disparity in the prevailing meridional wind patterns. These patterns were determined by calculating the sum of the meridional wind component from July to November each year. The preference of areas north of the Ría de Arousa as the main seed source during the period 2010–2014 was observed under a greater dominance of southerly winds (Figure 6). Conversely, a shift towards a dominance of northerly winds in the period from 2016 to 2020 placed the harvesting areas to the south of Ría de Arousa. The correlation coefficient between the southern wind component and the main mussel harvesting areas on both, the southern (South_1) and northern (North_1) shores of Ría de Arousa exhibited identical magnitudes (|r|=0.68) but opposite signs, with a negative relationship for the southern shore and a positive one for the northern shore (Table 2). In addition to these areas, a statistically significant correlation with wind patterns was only observed in the sector North_3. This segment, which does not have any major estuaries, is located in the westernmost corner of the Galician coast just north of the mussel production areas. The positive correlation between southerly wind and wild seed collected in this area was 0.63 despite having an exclusively natural mussel population.
The mussel aquaculture production in Galicia for the period 2006–2022 (Figure 7A; Table 3) was mostly dedicated to fresh consumption (53%), while 47% went to the processed industry for canned, frozen or other derivative products.
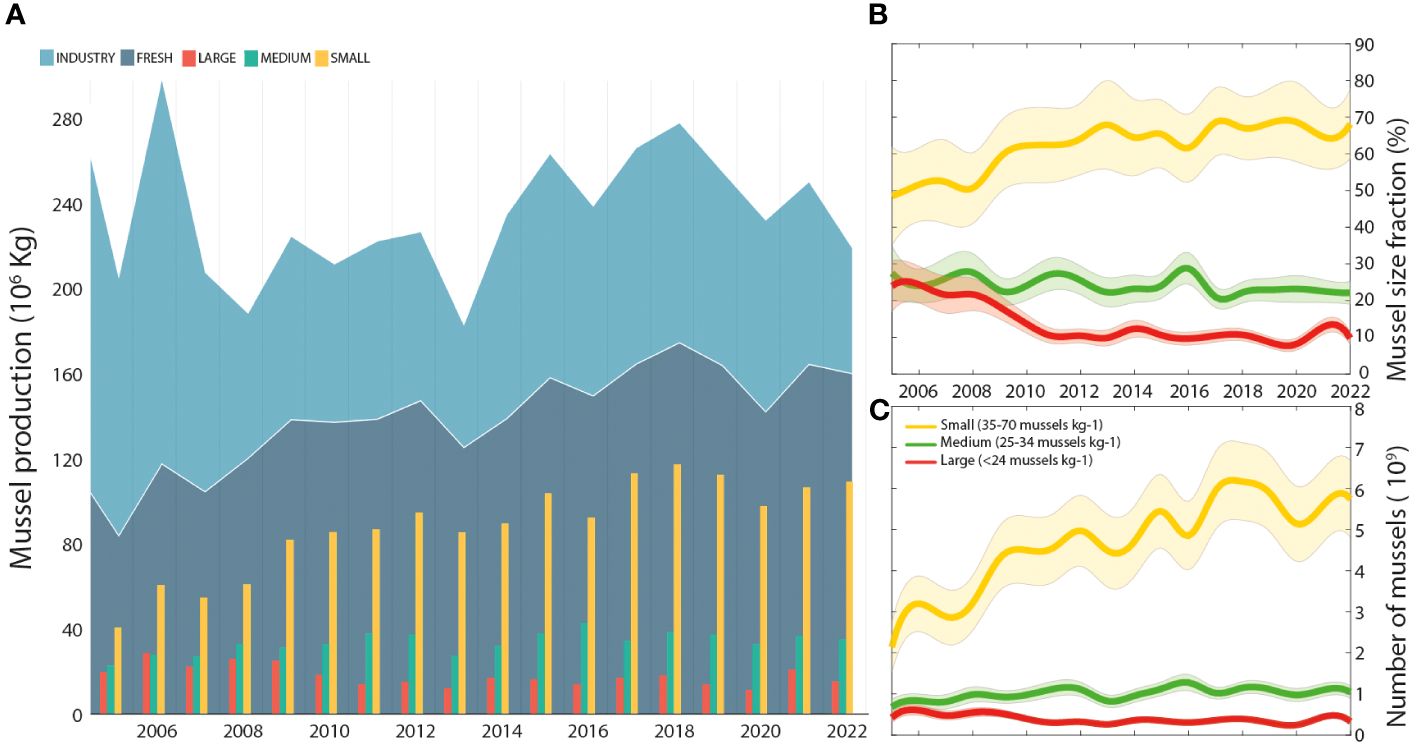
Figure 7 (A) Mussel production destined for the industry and for fresh consumption from 2006 to 2022, indicating the proportion of mussel sizes. (B) Evolution of the number of individuals produced for fresh consumption in each size class of mussels: small, medium, and large. (C) Number of individuals produced for fresh consumption in each mussel size class: small, medium and large.
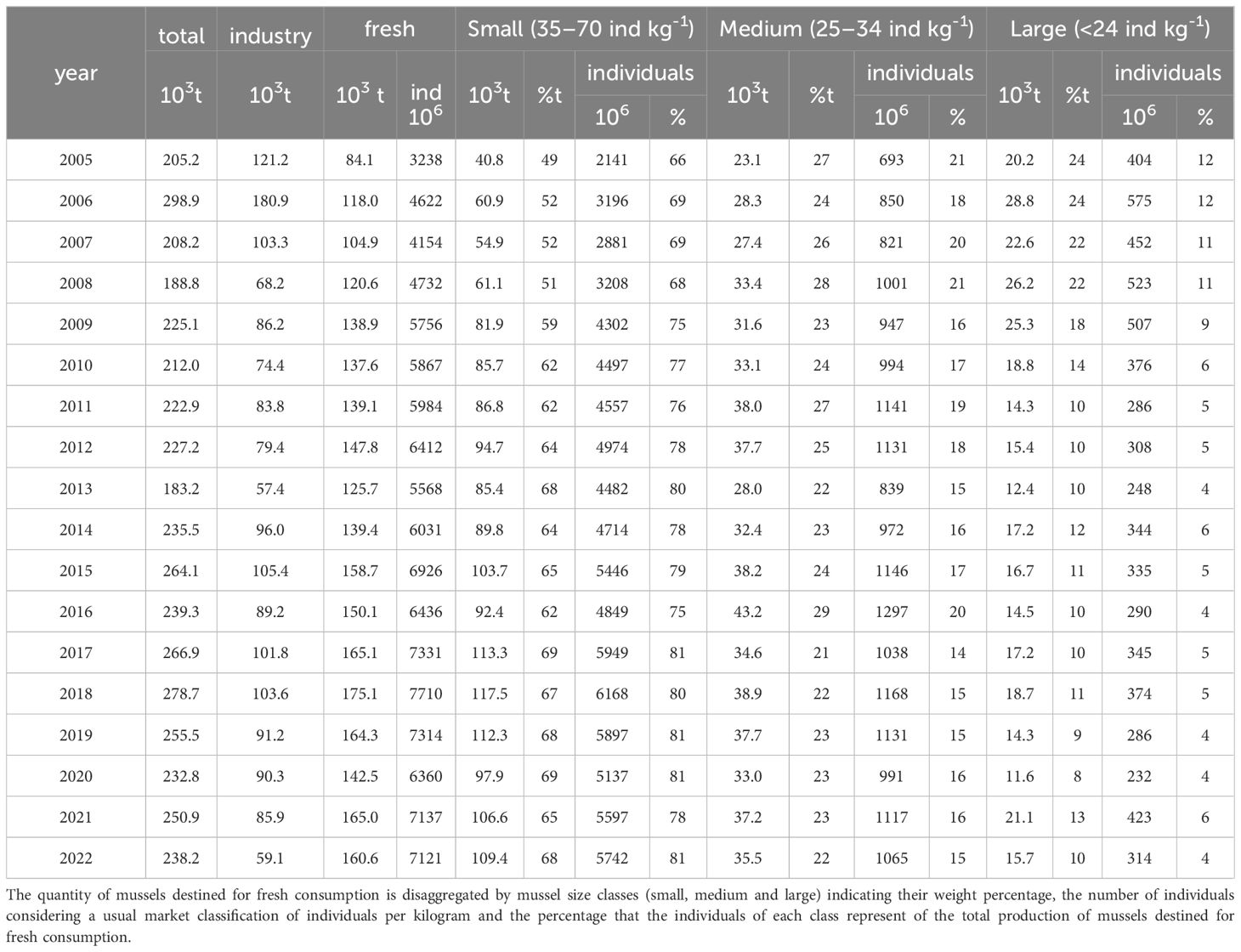
Table 3 Annual summary of mussel production collected by www.pescadegalicia.com for industry and for fresh consumption.
The total mussel aquaculture production exhibited in general terms a significantly positive trend (3703 ± 1135 t yr-1; r2 = 0.40) for the studied period when the peak of production of 2006 is excluded (an outstanding production year of 290K tonnes dedicated towards industry (65.5%).
In addition to the increase in total mussel production, a progressive shift towards small commercial sizes was also observed (Figure 7B). The fraction of production dedicated to small mussels (45–70 pieces kg-1) increased at a rate of 3660 t yr-1, going from 50% to almost 70% of the fresh production during the study period. Accordingly, large mussels (<24 pieces kg-1) decreased their contribution to the fresh production at a rate of -543 t yr-1, dropping from 25% to 10% of the production during the study period. Medium-sized mussels kept more or less the same percentage of the fresh production (24%), with a slight decreasing rate of -0.2% yr-1. This shift in market preferences towards smaller individuals is even more evident when the percentage of mussel production in tonnes is transformed to the number of mussels of each size (Figure 7C).
The impact of this change in the number of individuals of each size was analyzed in terms of their potential egg production (Figure 8). For this purpose, egg production was estimated for a stable production of 140K tonnes with the variability of the observed mussel size fraction. The mean egg production in autumn in both situations was 36 1012 eggs with a significant increasing interannual trend of 0.07 ± 0.01 1012 eggs yr-1. When including spring in this analysis to calculate lifetime egg production of the cultivated population, the mean potential production would almost doubled (61 1012 eggs), but a decreasing trend of -0.60 ± 0.09 1012 eggs yr-1 arise, since the fraction of small mussels would be harvested before their reach their second spring and the contribution of big mussels have continuously decreased over the last decade (Figure 8).
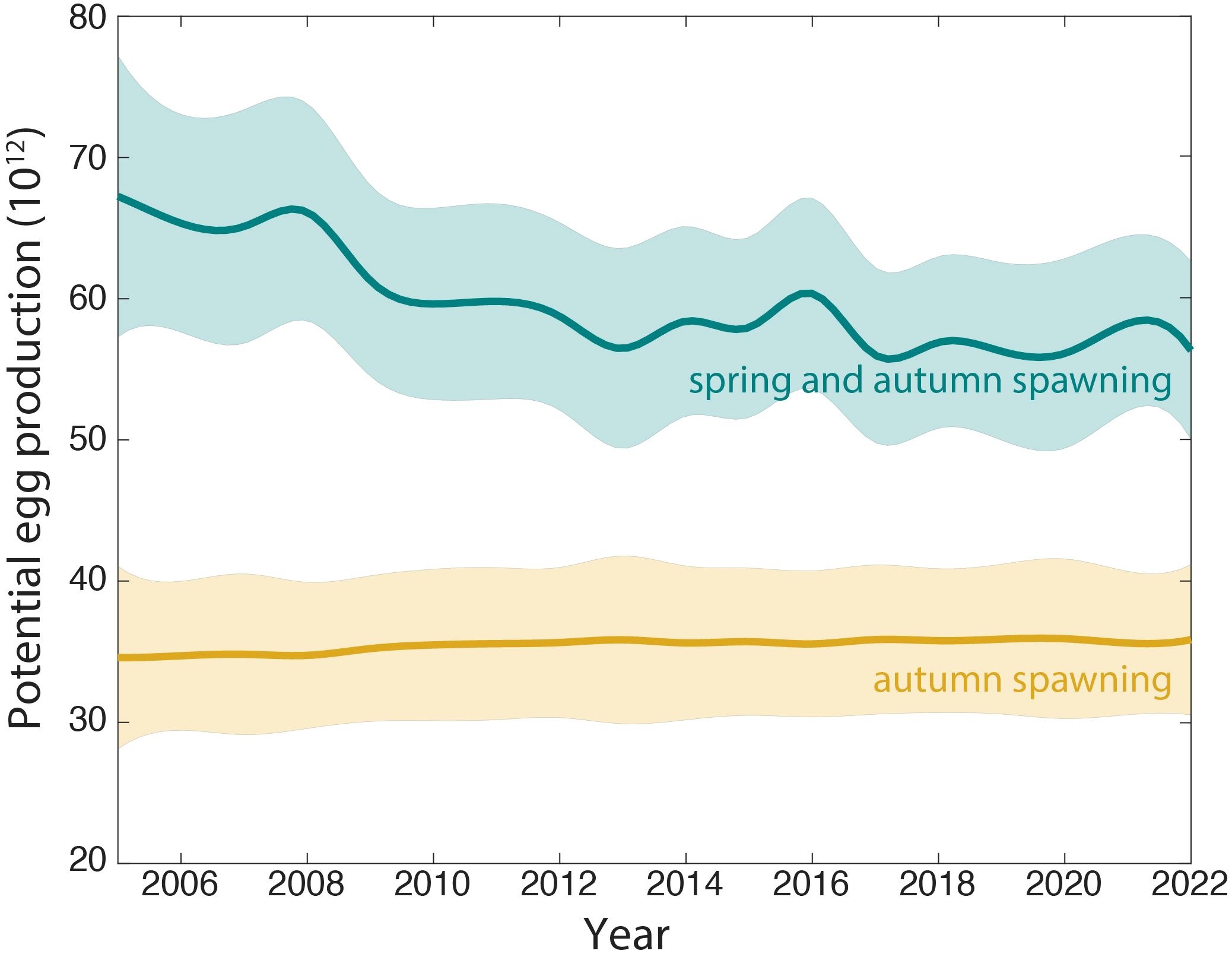
Figure 8 Time series of the estimated potential egg production of an average production of 140K tonnes but with the observed interannual variability of each size category in autumn (yellow lines) and in spring and autumn (blue lines) using the relationship between mussel size and number of eggs described by Oliveira et al. (2021).
This analysis is based on the first compilation of reports on the quantities of mussel seed collected and declared by mussel farmers in the Ría de Arousa for the period 2006–2022. This large amount of unpublished information constitutes the only time series record of mussel seed availability along the Galician coast. Despite certain methodological limitations, this information allows for the first time to study the effect of changes in aquaculture management and environmental variability on the regulation of mussel settlement.
This 17-year time series has revealed the progressive decline in the amount of wild mussel seed collected (Figure 4) on the Galician coast since 2012. This decline is more pronounced in the inner rafts (-107 ± 22 kg year-1 raft-1) than in the outer ones (-78 ± 31 kg year-1 raft-1) which on average use a 9% more mussel spat from the rocky shore for seeding their ropes (2917 ± 619 kg raft-1 and 2679 ± 712 kg raft-1 for the inner and outer rafts respectively). The lower dependency on wild seed in the rafts located in the outer estuarine zones would be associated with the higher efficiency of suspended collector ropes for seed capture at the most exposed locations (Pérez-Camacho et al., 1991; Peteiro et al., 2007; Peteiro et al., 2011).
The extrapolation of these data to the entire Galician production, resulted in an annual quantity of wild seeds collected of 8316 tonnes on average, with a range that oscillates between 9575 and 6300 tonnes estimated for 2012 and 2021, respectively. These quantities of wild mussel seed are higher than the 7500 tonnes (Camacho et al., 1995) and 6540 tonnes (Cáceres-Martínez and Figueras, 2007) estimated at the end of the 20th century as the amount of mussel seed necessary to maintain mussel cultivation. Considering that 10% of the rafts use exclusively collector seed and 25% combine both sources (Mariño et al., 1982; Pérez-Camacho et al., 1991), the contribution of collector ropes to the total seed would be approximately 22.5%. Assuming these percentages, the wild seed contribution from the rocky shore would have been around 5440 tonnes according to these previous studies (Camacho et al., 1995; Cáceres-Martínez and Figueras, 2007) and would have increased by 53% to our average estimate of 8316 tonnes. Furthermore, our calculations, which would represent 77.5% of the mussel seed destined for cultivation, would also raise the total seed required to an average of 10730 tonnes.
The entire time series of wild seed destined for the total number of mussel rafts in Galicia estimated from mussel farmers’ reports exhibited an annual reduction of 148 ± 36 t yr-1 which reached 288 ± 55 t yr-1 from 2012 onwards. This decline follows the reduction in recruitment and stability observed in natural mussel populations across different coasts of the Atlantic and Pacific Oceans which have been mainly attributed to environmental changes and overexploitation (Sorte et al., 2017; Commito et al., 2019; Petraitis and Dudgeon, 2020; Avdelas et al., 2021). The variable explaining the larger amount of this variability (60%) was the intensity of upwelling between July and November, specifically the recurrence of intense upwelling events (>500 m3 km-1 s-1), which have a detrimental effect on the abundance of mussel seed. Similar results have been reported before in the NW Pacific Coast (Menge et al., 2022), and also at the Galician rías, where the adaptation of mussel larvae to the characteristic intermittence of upwelling has been highlighted (Peteiro et al., 2011). Prevailing northerly winds, favorable to nearshore upwelling, may be behind the offshore transport of larvae during the pelagic phase of the mussel and can also prevent the accumulation of phytoplankton. Recently, a study by Otero et al. (2023) has determined that upwelling index values around 500 m3 km-1 s-1 would be a threshold for the activation of temperature changes in the water column of rías, and presumably, the limit for most noticeable effects on actual upwelling conditions. Our data also showed an increase in the number of days with an upwelling index above this threshold value over the period 2006–2022. Although many larval behaviors have evolved to avoid offshore wastage in upwelling systems (Shanks and Brink, 2005; Queiroga et al., 2007), several studies agree on reporting lower recruitment rates in areas with persistent downwelling or upwelling conditions (Navarrete et al., 2005; Broitman et al., 2008; Dudas et al., 2009; Menge et al., 2003; Smith et al., 2009). Intermittent upwelling, and the reversal currents associated with that process, might play a major role in regulating larval behavior and transport, as has been described before (Guisande et al., 2001; Otero et al., 2008, Otero et al., 2009; Peteiro et al., 2011; Pfaff et al., 2011; Bashevkin et al., 2020). Our results would confirm the importance of the frequency and variability of wind-driven currents as a key factor in recruitment success for autumn mussel spawning on the Galician coast (Pérez-Camacho and Román-Cabello, 1979; Mariño et al., 1982; Cáceres-Martínez et al., 1993; Villalba, 1995).
As stated above, the system has an evident response to regional winds, modulated in their persistence and duration by the atmospheric variability of the North Atlantic (Fraga, 1981; Blanton et al., 1984). Climate indices such as the North Atlantic Oscillation (NAO; Hurrell, 1995) and the East Atlantic (EA; Barnston and Livezey, 1987) capture the variability of recurrent spatial patterns at the basin level. The coupled patterns of the NAO and EA enhance upwelling, especially in winter (Dickson et al., 1988; deCastro et al., 2008). The decrease in seed availability began in 2012, coinciding precisely with significant changes in NAO and EA, which experienced a positive shift in their mean values (see Figure 5 in Otero et al., 2023) that contributed to the expansion of the subpolar gyre (de Jong and de Steur, 2016; Josey et al., 2018) and consequently, a freshening of the surface waters in the eastern subpolar North Atlantic basin (Holliday et al., 2020). Although it is difficult to establish a cause-effect relationship due to the different scales involved, it is not negligible that abrupt atmospheric, thermohaline, and seed availability changes coincide in time. In contrast to NAO, EA and seed variables showed a significant correlation between winter EA values and subsequent seed reports (r=0.44; p=0.01). Understanding system variability through these indices could potentially contribute to predicting the recruitment success of wild seeds on the coast.
In addition to the influence on larval transport processes, the wind pattern also affects temperature and food availability in the area. In this respect, increasing temperature has a complex effect on the mussel population, since it increases growth but could also have a detrimental effect on condition index and reproductive output when certain thresholds are surpassed (Franz et al., 2019; Menge et al., 2022), although its effect is largely modulated by food availability. Our results did not show a significant effect of temperature on mussel seed abundance, but we do detect a positive effect of Chl-a, which explains 25% of the variability observed. Chlorophyll concentration has a well-established effect on larval production and settlement of mussel recruits (Suárez et al., 2005; Broitman et al., 2008; Peteiro et al., 2010, Peteiro et al., 2011; Fuentes-Santos et al., 2014; Fuentes-Santos and Labarta, 2015; Fuentes-Santos et al., 2016), and is a proxy for food availability that would affect both, the physiological condition of the spawning mussels and larval development.
The inverse correlation between Chl-a and the annual cumulative sum of the upwelling index exceeding 500 m3 km-1 s-1 (p-value=0.11) seems contradictory to the expected direct response of phytoplankton biomass to increased nutrient inputs resulting from enhanced upwelling (Gregg et al., 2005). However, the growth of phytoplankton is favored by the succession of upwelling and downwelling events (Figueiras et al., 2002). The intense and/or persistent upwelling events could hinder from assimilating the nutrient injection due to the reduction of the retention time of the water within the estuary (Gilcoto et al., 2007; Largier, 2020) and the flushing of phytoplankton biomass offshore (Álvarez-Salgado et al., 2008). In this sense, increasing intensity of upwelling has also been related to a reduction of the impact of harmful algal blooms in the Rías Baixas (Álvarez-Salgado et al., 2008). Although coastal upwelling has a direct positive effect on productivity and consequently on mussel growth (Pérez-Camacho et al., 1991; Figueiras et al., 2002) and condition index (Blanton et al., 1987; Figueiras et al., 2002), maintaining its characteristic intermittent nature is key to ensure population equilibrium, particularly to ensure recruitment and replenishment of natural populations (Peteiro et al., 2011).
Our results suggest an important role of upwelling intensity in the offshore advection of larvae and its distribution along the coast as has been reported before (McQuaid and Phillips, 2000; Ruiz-Villarreal et al., 2008; Pineda et al., 2010; Peteiro et al., 2011; Reis, 2015; Nolasco et al., 2018; Demmer et al., 2022). The selection of wild seed collection sites has a strong dependence on the meridional component of the wind between July and November. According to the correlations observed between the preferential areas for mussel collection and the meridional wind component, the prevalence of upwelling favorable winds observed between 2016 and 2020 favored the settlement on the southern areas of the Galician coast, while downwelling favorable winds registered from 2010 to 2014 favored larval transport towards the north. The role of wind in controlling the connectivity of mussel populations has been reported before as a key element regulating meta-population persistence (Peteiro et al., 2011; Gomes et al., 2016; Franz et al., 2019; Bashevkin et al., 2020; Demmer et al., 2022). Since most of the mussel rafts are concentrated in Ría de Arousa, this is a key “source” sub-population for the Galician coast (Reis, 2015), and predominant winds will highly determine the role and relevance of different “sink” sub-populations and regions along the coast. Some exercises on meta-population modelling to understand key processes for mussels populations persistence (Carson et al., 2011) have highlighted the relevance of self-recruitment at the main source subpopulation as a key factor to ensure meta-population growth.
The distribution of wild seed collection sites also shows how mussel farmers started to travel longer distances since 2014 (North 4 and South 3; Figure 6), indicating an increase in the effort needed to obtain the required mussel spat for seeding their rafts. The observed decline in mussel seed availability also encompasses the general trend of declining natural mussel populations observed along the Atlantic and Pacific coasts (Sorte et al., 2017; Commito et al., 2019; Franz et al., 2019; Petraitis and Dudgeon, 2020; Avdelas et al., 2021; Baden et al., 2021; Menge et al., 2022). Certain studies document as much as a 60% reduction in mussel populations during the last 40 years (Sorte et al., 2017), motivated by mass mortality events but also by continuous declines in mussel recruitment (M. edulis) estimated at 15.7% less recruitment per year by Petraitis and Dudgeon (2020) in the western North Atlantic. Over-exploitation and climate change are the most often reported causes of this decline (Baden et al., 2021; Menge et al., 2022).
The management shifts of the aquaculture sector towards the harvest of smaller individuals due to market preferences and larger proficiency, could partly motivate the high demand for wild mussel seed and even be responsible for the reduction of this availability. Intuitively, a shorter cultivation period would require a higher supply of seed to maintain mussel production as observed when comparing the current wild seed supply estimated in this study (8316 tonnes) with that reported at the end of the 20th century (Pérez-Camacho et al., 1995; Cáceres-Martínez and Figueras, 2007). However, this increased use of seed did not translate into increased production. Our results show a lack of correlation between wild seed availability and total mussel production in the same year and a negative relationship with the following year’s production (p-value<0.05, r2 = 0.30).
This unexpected no-correlation between wild seed availability and mussel production could be related to numerous and complex factors. Among them, coastal upwelling emerges as a significant and oppositely controlling forcing between seed availability and mussel production. As described in this research, intense coastal upwelling events increase the probability of offspring loss due to offshore transport of mussel larval stages. Conversely, coastal upwelling was directly related to mussel growth (Pérez-Camacho et al., 1991; Figueiras et al., 2002), condition index (Blanton et al., 1987; Figueiras et al., 2002) and even the reduction of production closures due to harmful algal blooms (Álvarez-Salgado et al., 2008). Upwelling has been even suggested as a resilience mechanism against stressful conditions because of the supply of food, which might help to maintain the known efficiency of mussel farming in Galicia (Pérez-Camacho and Labarta, 2004; Kamermans and Capelle, 2019) even in a context of generalized production recession in Europe (Avdelas et al., 2021). The ability of coastal upwelling to act as a counterbalance between larval recruitment success and adult mussel physiological conditions may be related to its intensity and seasonality, which may also determine the net impact on primary production and transport processes.
Other factors that could explain the null impact of the quantity of seed collected in the coast and mussel production would be specific aspects of the mussel culture process in rafts. The most obvious explanation would be a much larger contribution of seeds from the collector ropes than suggested previously, which would compensate for the observed decline in wild seeds. However, no information is available on this seed contribution to the rafts. Another explanation could be that the use of the wild seed in the rafts above the threshold of seeded individuals would have a negative effect on production. In fact, the number of mussel seeds is indeterminate as the amount of wild seed collected is reported in kilograms. The high size heterogeneity of this resource, which also incorporates small mussel seeds (<1 mm), could account for a considerable number of individuals. In addition, the need for completely covering the ropes with mussel seed to avoid epifauna fixation makes it infeasible to manage the density or number of wild seed sown during the onset of cultivation. In this situation, shelf-thinning due to interspecific competition for food and space determines the number of mussels that will initiate the subsequent fattening process (Camacho et al., 1995).
The reduction of the traditional 18-month cultivation cycle to less than 12 months can also have a significant effect on the availability of mussel seeds in the rocky shore by reducing the spawning opportunities for adult mussels before harvest. In particular, the production of smaller mussels, mainly seeded in rafts during the winter-spring and harvested in late winter-early spring, would only participate in the fall spawning, as they would be immature during their first spring (Villalba, 1995) and harvested before the next one. When considering a stable mussel production dedicated to the fresh market and only altering the proportion of individuals of each size according to the progression over the last 16 years, a detrimental effect on the total reproductive output is observed only when both reproductive peaks are taken into account. The greater number of smaller individuals would compensate for the loss of spawning potential of larger individuals during the fall peak, but during spring, this potential is reduced. It should be noted that these estimates do not take into account the possible loss of fecundity due to the reduction in egg diameter, which depends on the size of the broodstock (Dare et al., 2004). The large sensitivity of population dynamics to changes in adult fecundity has been previously pointed out, signaling reproductive output as the most relevant factor controlling meta-population persistence, above survival and growth of juvenile stages or even population connectivity (Carson et al., 2011). Specific studies evaluating the reproductive performance of the adult population and its viability under different production scenarios (% of production of different sizes) are lacking to determine the actual impact of changes in the mussel farming strategy in Galicia on the availability of mussel seed.
In this scenario of climate change, the intensification of coastal upwelling is the most accepted hypothesis (Bakun, 1990) and forecasted by future climate projections (Rykaczewski et al., 2015; Cordeiro Pires et al., 2016; Álvarez et al., 2017; Soares et al., 2017; Sousa et al., 2017; Sousa et al., 2020). These climate models indicate that the increase in coastal upwelling will be particularly significant between May and October (Rykaczewski et al., 2015; Álvarez et al., 2017; Sousa et al., 2017; Sousa et al., 2020), which practically coincides with the pelagic phase considered in this study for the mussel seed collected from the rocky shore. This seasonal intensification would project more adverse conditions in the future for mussel larval recruitment success (Bashevkin et al., 2020), exacerbating wild mussel seed supply problems. However, these projections are still not confirmed by trends observed along the Galician coast to date (Álvarez-Salgado et al., 2008; Pérez et al., 2010; Pardo et al., 2011; Barton et al., 2013; Sydeman et al., 2014), where some data indicates even a decrease on the average intensity and duration of northerly winds during the upwelling season (Álvarez-Salgado et al., 2008).
In parallel to the possible increase in coastal upwelling and nutrient input into surface waters, global warming would promote stratification of the water column limiting fertilization (Sousa et al., 2020). The reduction in nutrient injection and possible decrease in phytoplankton abundance (Behrenfeld et al., 2006; Li and Harrison, 2008; Richardson, 2008; Sommer and Lengfellner, 2008; Morán et al., 2010) would have a direct negative impact on mussel production (Des et al., 2020). In addition to affecting mussel growth, suboptimal dietary conditions would also compromise the physiological condition of mussels, influencing their response to other environmental stressors, such as ocean acidification (Lassoued et al., 2019; Bashevkin et al., 2020; Lassoued et al., 2021), which is particularly pronounced in coastal waters off the Galician coast (Padin et al., 2020).
These environmental perturbations related to global change would also have an impact on reproductive seasonality (Edwards and Richardson, 2004; Philippart et al., 2012, Philippart et al., 2014) and larval survival (Poloczanska et al., 2013). However, it is important to note that climate projections are particularly inaccurate in coastal areas (Sarà et al., 2018; Stavrakidis-Zachou et al., 2018; Falconer et al., 2020). Therefore, incorporating these climate models as tools to improve the management and sustainability of mussel production requires better climate projections (Ahmed et al., 2019).
The sustainability and resilience of mussel aquaculture must be based on a thorough understanding of the environmental and management factors that regulate natural and cultured populations and their complex interactions. Understanding the processes underlying the variability of mussel seed availability is crucial not only to secure the future of the mussel aquaculture sector, but also to ensure the stability of wild populations. This is particularly significant in the context of global change, where cascading effects on marine ecosystems significantly impact mollusk aquaculture (Handisyde et al., 2017) due to their high sensitivity to environmental conditions (FAO, 2018). Our results highlight how such knowledge needs to be supported by long-term monitoring systems, ensuring the availability of time series long enough to detect the effect of environmental and management changes on aquaculture systems and their impact on natural populations.
Mussel seed collection reports from mussel producers in the Ría de Arousa are a useful source of information to know the availability of mussel seed, inter-annual variability and identification of the forcings that determine its highly variable stock on the rocky coast. The present analysis showed a decline in the availability of wild seed on the coast over the period 2006–2021. Extrapolation of these results to the total number of mussel rafts in Galicia would estimate the average annual quantity of seed collected to be 8316 tonnes within a range of 9575 tonnes in 2012 and 6300 tonnes in 2021. The entire time series of mussel seeds collected for initiating cultivation across all Galician rafts exhibited a reduction of 148 ± 36 t yr-1 which was particularly marked from 2012 onwards (288 ± 55 t yr-1). These values update the estimates made in the 20th century for the supply of wild seed from the coast at 5440 tonnes (Camacho et al., 1995; Cáceres-Martínez and Figueras, 2007).
The decreasing availability of mussel seed was related to wind and chlorophyll concentration during the larval stage of recruits collected between December and April. The sum of coastal upwelling episodes greater than 500 m3 km-1 s-1 explained 60% of the observed interannual variability of collected wild seed. The meridional component of the wind also determined the settlement pattern and dispersal of mussel seed along the coastline on both sides of the Ría de Arousa, the main source of mussel larvae as it accumulates 68% of the mussel rafts on the Galician coast. Chlorophyll also showed a significant correlation with seed availability explaining a 25%. These changes appear to be associated with larger-scale alterations observed in the Atlantic Ocean that could significantly link wild seed recruitment success to basin-scale climate indices.
The results reported here corresponded to a period with a marked shift in the farming strategy of the sector with regard to the increased production of small-sized mussels to meet market demand and improve profitability. As a consequence, the number of individuals farmed increased by 75% for the period 2006–2021 at the same time as the growing period was reduced. These changes in culture practices may also affect the reproductive potential of mussels as it limits spawning opportunities and the number of gametes which is proportional to the size of broodstock. This potential impact was analyzed based on a mean production value including the inter-annual variability of sizes observed over the period. Such simplistic approximation showed that the reproductive potential would remain stable in the autumn spawning due to the increase in the number of spawners while the sum of spring and autumn spawning would show a significant decrease in reproductive potential linked to reduced optimal reproductive time opportunities.
This first analysis of mussel seed harvesting reports highlights the importance of large monitoring time series for understanding the processes that determine the prosperity or decline of exploited wild seed stocks. Improving the quality and reliability of this information is therefore a necessity for the optimal management of this resource in the near future. The participation of observers to control the quantity of seed collected, the identification of collection areas by geographical coordinates and the digitalization of this information are aspects that need to be implemented in the compilation of these reports. In addition, in situ monitoring of environmental conditions jointly with the wild seed stock and size distribution during collection could allow a more rational establishment of both the collection period and areas, as well as a more precise estimation of the number of individuals used during the seeding process in the rafts. The contribution of collectors should also be included in such reports. Environmental conditions and changes in the availability of wild seed reported here may suggest that the contribution of mussel from the latter collector source is higher than previously considered (22.5%) and most likely would have considerable temporal and spatial variability to be determined in detail. Our data would indicate that throughout the study period the contribution of the collector ropes together with other factors of the cultivation process would contribute to compensate for the lack of seed from the rocky shore.
This knowledge would also represent an advance in addressing the impact of climatic changes on mussel seed availability, as well as how changes in mussel aquaculture management may interact with those that directly or indirectly drive changes in the replenishment and growth of natural mussel populations.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The manuscript presents research on animals that do not require ethical approval for their study.
XP: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Validation, Visualization, Writing – original draft, Writing – review & editing. JB: Conceptualization, Formal analysis, Funding acquisition, Investigation, Supervision, Validation, Writing – original draft, Writing – review & editing. PO: Conceptualization, Data curation, Formal analysis, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. MG: Funding acquisition, Supervision, Validation, Writing – original draft, Writing – review & editing. TR: Data curation, Writing – review & editing. LS: Data curation, Writing – review & editing. AV: Data curation, Funding acquisition, Writing – original draft, Writing – review & editing. LP: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was funded by MCIN/AEI/10.13039/501100011033 through REDEIRA (Ref: TED2021-132188B-I00/AEI/10.13039/501100011033/Unión Europea NextGenerationEU/PRTR) and STRAUSS (Ref: PID2019-106008RB-C21) projects, as well as by The Marine Science Program, ThinkInAzul, in Galicia supported by Ministerio de Ciencia e Innovación and Xunta de Galicia (Working Package: PT7) with funding from European Union NextGenerationEU (PRTR-C17.I1) and European Maritime and Fisheries Fund.
We thank the organization OPMEGA (Organización de Productores de Mexillón de Galicia) for their help in obtaining the necessary information from the mussel farmers of the different associations and “Plataforma Tecnolóxica da Pesca’’ from the Galician Government (Consellería do Mar - Xunta de Galicia) for providing data on the mussel total production by market destinations. Thanks to the Copernicus programme of the European Commission for the distribution of the chlorophyll remote sensing product.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aguirre M. (1979). Biología del mejillón (“M. Edulis”) de cultivo de la Ría de Vigo: N.° 276. Bol. Inst. Esp. Oceanogr. 5, 107–160.
Ahmed N., Thompson S., Glaser M. (2019). Global aquaculture productivity, environmental sustainability, and climate change adaptability. Environ. Manage. 63, 159–172. doi: 10.1007/s00267-018-1117-3
Álvarez I., Lorenzo M. N., deCastro M., Gomez-Gesteira M. (2017). Coastal upwelling trends under future warming scenarios from the CORDEX project along the Galician coast (NW Iberian Peninsula). Int. J. Climatol. 37, 3427–3438. doi: 10.1002/joc.4927
Álvarez-Salgado X. A., Gago J., Míguez B. M., Gilcoto M., Pérez F. F. (2000). Surface waters of the NW Iberian margin: Upwelling on the shelf versus out welling of upwelled waters from the Rías Baixas. Estuar. Coast. Shelf Sci. 51, 821–837. doi: 10.1006/ecss.2000.0714
Álvarez-Salgado X. A., Labarta U., Fernández-Reiriz M. J., Figueiras F. G., Rosón G., Piedracoba S., et al. (2008). Renewal time and the impact of harmful algal blooms on the extensive mussel raft culture of the Iberian coastal upwelling system (SW Europe). Harmful Algae 7, 849–855. doi: 10.1016/j.hal.2008.04.007
Alvarez-Salgado X., Rosón G., Pérez F., Figueiras F., Pazos Y. (1996). Nitrogen cycling in an estuarine upwelling system, the Ría de Arousa (NW Spain). I. Short-time-scale patterns of hydrodynamic and biogeochemical circulation. Mar. Ecol. Prog. Ser. 135, 259–273. doi: 10.3354/meps135259
Álvarez-Salgado X. A., Fernández-Reiriz M.-J., Labarta U., Filgueira R., Peteiro L., Figueiras F. G., et al. (2009). Influencia Do Cambio Climático No Cultivo Do Mexillón Das Rías Galegas. Evidencias e impactos do cambio climático en Galicia. Ed. Galicia X. 373–389.
Arístegui J., Barton E. D., Álvarez-Salgado X. A., Santos A. M. P., Figueiras F. G., Kifani S., et al. (2009). Sub-regional ecosystem variability in the Canary Current upwelling. Prog. Oceanogr. Eastern Boundary Upwelling Ecosystems: Integr. Comp. Approaches 83, 33–48. doi: 10.1016/j.pocean.2009.07.031
Avdelas L., Avdic-Mravlje E., Borges Marques A. C., Cano S., Capelle J. J., Carvalho N., et al. (2021). The decline of mussel aquaculture in the European Union: causes, economic impacts and opportunities. Rev. Aquac. 13, 91–118. doi: 10.1111/raq.12465
Babarro J. M. F., Abad M. J., Gestoso I., Silva E., Olabarria C. (2018). Susceptibility of two co-existing mytilid species to simulated predation under projected climate change conditions. Hydrobiologia 807, 247–261. doi: 10.1007/s10750-017-3397-7
Babarro J. M. F., Filgueira R., Padín X. A., Longa Portabales M. A. (2020). A novel index of the performance of Mytilus galloprovincialis to improve commercial exploitation in aquaculture. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00719
Baden S., Hernroth B., Lindahl O. (2021). Declining populations of mytilus spp. in north atlantic coastal waters—A swedish perspective. J. Shellfish Res. 40, 269–296. doi: 10.2983/035.040.0207
Bakun A. (1990). Global climate change and intensification of coastal ocean upwelling. Science 247, 198–201. doi: 10.1126/science.247.4939.198
Barnston A. G., Livezey R. E. (1987). Classification of seasonality and persistence of low-frequency atmospheric circulation patterns. Mon Wea Rev. 115, 1083–1126. doi: 10.1175/1520-0493(1987)115<1083:CSAPOL>2.0.CO;2
Barton E. D., Field D., Roy C. (2013). Canary Current upwelling: More or less? Prog. Oceanogr. 116, 167–178. doi: 10.1016/j.pocean.2013.07.007
Bashevkin S. M., Dibble C. D., Dunn R. P., Hollarsmith J. A., Ng G., Satterthwaite E. V., et al. (2020). Larval dispersal in a changing ocean with an emphasis on upwelling regions. Ecosphere 11, e03015. doi: 10.1002/ecs2.3015
Behrenfeld M. J., O’Malley R. T., Siegel D. A., McClain C. R., Sarmiento J. L., Feldman G. C., et al. (2006). Climate-driven trends in contemporary ocean productivity. Nature 444, 752–755. doi: 10.1038/nature05317
Bevington P. R., Robinson D. K. (2003). “Data reduction and error analysis for the physical sciences,”, 3 ed (McGraw-Hill, New York), 320 pp.
Blanton J., Atkinson L., Castillejo F. F., Montero A. L. (1984). Coastal upwelling off the Rias Bajas, Galicia, Northwest Spain I: hydrographic studies. Rapp. Procès-Verbaux La Réun. Biol. Product. N. Atl. Shelf Areas 183, 79–90. Available at: https://digitalcommons.odu.edu/ccpo_pubs/341/.
Blanton J. O., Tenore K. R., Castillejo F., Atkinson L. P., Schwing F. B., Lavin A. (1987). The relationship of upwelling to mussel production in the rias on the western coast of Spain. J. Mar. Res. 45, 497–511. doi: 10.1357/002224087788401115
Brea Bermejo E. (2009). Cartografiado y dinámica de poblaciones de los bancos naturales de semilla de mejillón en las costas atlánticas gallegas. PhD Thesis (Santiago de Compostela, Spain: Univ Santiago de Compostela).
Broitman B. R., Blanchette C. A., Menge A., Lubchenco J., Krenz C., Foley M., et al. (2008). Spatial and temporal patterns of invertebrate recruitment along the west coast of the United States. Ecol. Monogr. 78, 403–421. doi: 10.1890/06-1805.1
Cáceres-Martínez J., Figueras A. (1998). Mussel (Mytilus galloprovincialis Lamarck) colonization on artificial substrates in the Ria de Vigo of NW Spain. J. Shellfish Res. 17, 153–157.
Cáceres-Martínez J., Figueras A. (2007). Vida larvaria del mejillón. In El desarrollo larvario del mejillón (Mytilus galloprovincialis) en Galicia (Madrid: Consejo Superior de Investigaciones Científicas (Spain)), 289.
Cáceres-Martínez J., Robledo J. A., Figueras Huerta A. (1993). Settlement of mussels Mytilus galloprovincialis on an exposed rocky shore in Ria de Vigo, NW Spain. Mar. Ecol. Prog. Ser. 93, 195–198. doi: 10.3354/meps093195
Camacho A. P., Labarta U., Beiras R. (1995). Growth of mussels (Mytilus edulis galloprovincialis) on cultivation rafts: influence of seed source, cultivation site and phytoplankton availability. Aquaculture 138, 349–362. doi: 10.1016/0044-8486(95)01139-0
Carson H. S., Cook G. S., López-Duarte P. C., Levin L. A. (2011). Evaluating the importance of demographic connectivity in a marine metapopulation. Ecology 92, 1972–1984. doi: 10.1890/11-0488.1
Chavez F. P., Messié M. (2009). A comparison of eastern boundary upwelling ecosystems. Prog. Oceanogr. 83, 80–96. doi: 10.1016/j.pocean.2009.07.032
Commito J. A., Jones B. R., Jones M. A., Winders S. E., Como S. (2019). After the fall: Legacy effects of biogenic structure on wind-generated ecosystem processes following mussel bed collapse. Diversity 11, 11. doi: 10.3390/d11010011
Cordeiro Pires A., Nolasco R., Rocha A., Ramos A. M., Dubert J. (2016). Climate change in the Iberian Upwelling System: a numerical study using GCM downscaling. Clim. Dyn. 47, 451–464. doi: 10.1007/s00382-015-2848-y
Costello C., Cao L., Gelcich S., Cisneros-Mata M.Á., Free C. M., Froehlich H. E., et al. (2020). The future of food from the sea. Nature 588, 95–100. doi: 10.1038/s41586-020-2616-y
Dare P. J., Bell M. C., Walker P., Bannister R. C. A. (2004). Historical and current status of cockle and mussel stocks in The Wash. CEFAS Lowestoft, 85pp.
deCastro M., Gómez-Gesteira M., Alvarez I., Lorenzo M., Cabanas J. M., Prego R., et al. (2008). Characterization of fall–winter upwelling recurrence along the Galician western coast (NW Spain) from 2000 to 2005: Dependence on atmospheric forcing. J. Mar. Syst. Oceanography Bay Biscay 72, 145–158. doi: 10.1016/j.jmarsys.2007.04.005
de Jong M. F., de Steur L. (2016). Strong winter cooling over the Irminger Sea in winter 2014–2015, exceptional deep convection, and the emergence of anomalously low SST. Geophys. Res. Lett. 43, 7106–7113. doi: 10.1002/2016GL069596
Demmer J., Robins P., Malham S., Lewis M., Owen A., Jones T., et al. (2022). The role of wind in controlling the connectivity of blue mussels (Mytilus edulis L.) populations. Mov. Ecol. 10, 3. doi: 10.1186/s40462-022-00301-0
Des M., Gómez-Gesteira M., deCastro M., Gómez-Gesteira L., Sousa M. C. (2020). How can ocean warming at the NW Iberian Peninsula affect mussel aquaculture? Sci. Total Environ. 709, 136117. doi: 10.1016/j.scitotenv.2019.136117
deYoung B., Harris R., Alheit J., Beaugrand G., Mantua N., Shannon L. (2004). Detecting regime shifts in the ocean: data considerations. Prog. Oceanogr. 60, 143–164. doi: 10.1016/j.pocean.2004.02.017
Dickson R. R., Meincke J., Malmberg S.-A., Lee A. J. (1988). The “great salinity anomaly” in the Northern North Atlantic 1968–1982. Prog. Oceanogr. 20, 103–151. doi: 10.1016/0079-6611(88)90049-3
Dudas S. E., Rilov G., Tyburczy J., Menge B. A. (2009). Linking larval abundance, onshore supply and settlement using instantaneous versus integrated methods. Mar. Ecol. Prog. Ser. 387, 81–95. doi: 10.3354/meps08095
Edwards M., Richardson A. J. (2004). Impact of climate change on marine pelagic phenology and trophic mismatch. Nature 430, 881–884. doi: 10.1038/nature02808
Falconer L., Hjøllo S. S., Telfer T. C., McAdam B. J., Hermansen Ø., Ytteborg E. (2020). The importance of calibrating climate change projections to local conditions at aquaculture sites. Aquaculture 514, 734487. doi: 10.1016/j.aquaculture.2019.734487
FAO (2018). World food and agriculture: statistical pocketbook 2018 (Rome: Food and Agriculture Organization of the United Nations (FAO)).
Fernández-Pulpeiro E., Aldariz J. C. (2003). “Fauna asociada a sustratos rocosos: intermareales y sublitorales de las costas gallegas,” in El impacto del Prestige: análisis y evaluación de los daños causados por el accidente del Prestige y dispositivos para la regeneración medioambiental y recuperación económica de Galicia (A Coruña (Spain): Fundación Pedro Barrié de la Maza : Instituto de Estudios Económicos), 169–188.
Fernández-Pulpeiro E., Aldariz J. C., Lustres-Perez V., Brea Bermejo E. (2002). Prospección, cartografiado y gestión de los bancos naturales de semilla de mejillón (Final report for the fisheries and marine departments of Xunta de Galicia) (Santiago de Compostela: University of Santiago de Compostela).
Figueiras F. G., Labarta U., Fernández Reiriz M. J. (2002). Coastal upwelling, primary production and mussel growth in the Rías Baixas of Galicia. Hydrobiologia 484, 121–131. doi: 10.1023/A:1021309222459
Fraga F. (1981). “Upwelling off the galician coast, northwest Spain,” in Coastal Upwelling. Ed. Richards F. A. (Whasington DC (USA): American Geophysical Union), 176–182.
Franz M., Barboza F. R., Hinrichsen H.-H., Lehmann A., Scotti M., Hiebenthal C., et al. (2019). Long-term records of hard-bottom communities in the southwestern Baltic Sea reveal the decline of a foundation species. Estuar. Coast. Shelf Sci. 219, 242–251. doi: 10.1016/j.ecss.2019.02.029
Froehlich H. E., Gentry R. R., Halpern B. S. (2018). Global change in marine aquaculture production potential under climate change. Nat. Ecol. Evol. 2, 1745–1750. doi: 10.1038/s41559-018-0669-1
Froján C. R. S. B., Cooper K. M., Bolam S. G. (2016). Towards an integrated approach to marine benthic monitoring. Mar. pollut. Bull. 104, 20–28. doi: 10.1016/j.marpolbul.2016.01.054
Fuentes-Santos I., Cubillo A. M., Fernández-Reiriz M. J., Labarta U. (2014). Dynamic self-thinning model for sessile animal populations with multilayered distribution. Rev. Aquac. 6, 115–127. doi: 10.1111/raq.12032
Fuentes-Santos I., Labarta U. (2015). Spatial patterns of larval settlement and early post-settlement survivorship of Mytilus galloprovincialis in a Galician Ría (NW Spain). Effect on recruitment success. Reg. Stud. Mar. Sci. 2, 1–10. doi: 10.1016/j.rsma.2015.08.006
Fuentes-Santos I., Labarta U., Álvarez-Salgado X. A., Fernández-Reiriz M. J. (2016). Solar irradiance dictates settlement timing and intensity of marine mussels. Sci. Rep. 6, 29405. doi: 10.1038/srep29405
Fuentes-Santos I., Labarta U., Fernández-Reiriz M. J., Kay S., Hjøllo S. S., Alvarez-Salgado X. A. (2021). Modeling the impact of climate change on mussel aquaculture in a coastal upwelling system: a critical assessment. Sci. Total Environ. 775, 145020. doi: 10.1016/j.scitotenv.2021.145020
Gentry R. R., Alleway H. K., Bishop M. J., Gillies C. L., Waters T., Jones R. (2020). Exploring the potential for marine aquaculture to contribute to ecosystem services. Rev. Aquac. 12, 499–512. doi: 10.1111/raq.12328
Gephart J. A., Henriksson P. J. G., Parker R. W. R., Shepon A., Gorospe K. D., Bergman K., et al. (2021). Environmental performance of blue foods. Nature 597, 360–365. doi: 10.1038/s41586-021-03889-2
Gilcoto M., Pardo P. C., Álvarez-Salgado X. A., Pérez F. F. (2007). Exchange fluxes between the Ría de Vigo and the shelf: A bidirectional flow forced by remote wind. J. Geophys. Res. 112, C06001. doi: 10.1029/2005JC003140
Gomes I., Peteiro L., Albuquerque R., Nolasco R., Dubert J., Swearer S., et al. (2016). Wandering mussels: using natural tags to identify connectivity patterns among Marine Protected Areas. Mar. Ecol. Prog. Ser. 552, 159–176. doi: 10.3354/meps11753
Gonzalez-Nuevo G., Gago J., Cabanas J. M. (2014). Upwelling index: a powerful tool for marine research in the NW Iberian upwelling system. J. Oper. Oceanogr. 7, 47–57. doi: 10.1080/1755876X.2014.11020152
Gregg W. W., Casey N. W., McClain C. R. (2005). Recent trends in global ocean chlorophyll. Geophys. Res. Lett. 32, L03606. doi: 10.1029/2004GL021808
Guisande C., Cabanas J. M., Vergara A. R., Riveiro I. (2001). Effect of climate on recruitment success of Atlantic Iberian sardine Sardina pilchardus. Mar. Ecol. Prog. Ser. 223, 243–250. doi: 10.3354/meps223243
Handisyde N., Telfer T. C., Ross L. G. (2017). Vulnerability of aquaculture-related livelihoods to changing climate at the global scale. Fish Fish. 18, 466–488. doi: 10.1111/faf.12186
Hipel K. W., McLeod A. I. (1994). “Time series modelling of water resources and environmental systems,” in Developments in Water Science, 1st edition (Amsterdam: Elsevier Science).
Holliday N. P., Bersch M., Berx B., Chafik L., Cunningham S., Florindo-López C., et al. (2020). Ocean circulation causes the largest freshening event for 120 years in eastern subpolar North Atlantic. Nat. Commun. 11, 585. doi: 10.1038/s41467-020-14474-y
Hurrell J. W. (1995). Decadal trends in the north Atlantic oscillation: regional temperatures and precipitation. Science 269, 676–679. doi: 10.1126/science.269.5224.676
Hussain M. M., Mahmud I. (2019). pyMannKendall: a python package for non-parametric Mann Kendall family of trend tests. J. Open Source Softw 4, 1556. doi: 10.21105/joss.01556
Josey S. A., Hirschi J. J.-M., Sinha B., Duchez A., Grist J. P., Marsh R. (2018). The recent Atlantic cold anomaly: causes, consequences, and related phenomena. Annu. Rev. Mar. Sci. 10, 475–501. doi: 10.1146/annurev-marine-121916-063102
Kamermans P., Capelle J. (2019). Provisioning of mussel seed and its efficient use in culture, in: Goods and Services of Marine Bivalves. Springer pp, 27–49. doi: 10.1007/978–3-319–96776-9_3
Kämpf J., Chapman P. (2016). Upwelling Systems of the World (Cham: Springer International Publishing). doi: 10.1007/978–3-319–42524-5
Kim S.-U., Kim K.-Y. (2020). Impact of climate change on the primary production and related biogeochemical cycles in the coastal and sea ice zone of the Southern Ocean. Sci. Total Environ. 751, 141678. doi: 10.1016/j.scitotenv.2020.141678
Labarta U., Fernández-Reiriz M. J. (2019). The Galician mussel industry: Innovation and changes in the last forty years. Ocean Coast. Manage. 167, 208–218. doi: 10.1016/j.ocecoaman.2018.10.012
Largier J. L. (2020). Upwelling bays: how coastal upwelling controls circulation, habitat, and productivity in bays. Annu. Rev. Mar. Sci. 12, 415–447. doi: 10.1146/annurev-marine-010419-011020
Lassoued J., Babarro J. M. F., Padín X. A., Comeau L. A., Bejaoui N., Pérez F. F. (2019). Behavioral and eco-physiological responses of the mussel Mytilus galloprovincialis to acidification and distinct feeding regimes. Mar. Ecol. Prog. Ser. 626, 97–108. doi: 10.3354/meps13075
Lassoued J., Padín X. A., Comeau L. A., Bejaoui N., Pérez F. F., Babarro J. M. F. (2021). The Mediterranean mussel Mytilus galloprovincialis: responses to climate change scenarios as a function of the original habitat. Conserv. Physiol. 9, 10.1093. doi: 10.1093/conphys/coaa114
Lavín A., Díaz del Río G., Cabanas J. M., Casas G. (1991). Afloramiento en el noroeste de la Peninsula Iberica. Indices de afloramiento para el punto 43°N 11°W periodo 1966–1989. Inf. Téc. Inst. Esp. Oceanogr. 91, 1–40.
Li W. K. W., Harrison W. (2008). Propagation of an atmospheric climate signal to phytoplankton in a small marine basin. Limnol. Oceanogr. 53, 1734–1745. doi: 10.4319/lo.2008.53.5.1734
Mariño J., Pérez-Camacho A., Román-Cabello G. (1982). El cultivo del mejillón en la Ría de Arosa. Bol. Inst. Esp. Oceanog. 7 (2), 297–308.
McQuaid C. D., Phillips T. E. (2000). Limited wind-driven dispersal of intertidal mussel larvae: In situ evidence from the plankton and the spread of the invasive species Mytilus galloprovincialis in South Africa. Mar. Ecol. Prog. Ser. 201, 211–220. doi: 10.3354/meps201211
Menge B. A., Gravem S. A., Johnson A., Robinson J. W., Poirson B. N. (2022). Increasing instability of a rocky intertidal meta-ecosystem. Proc. Natl. Acad. Sci. 119, e2114257119. doi: 10.1073/pnas.2114257119
Menge B. A., Lubchenco J., Bracken M. E. S., Chan F., Foley M. M., Freidenburg T. L., et al. (2003). Coastal oceanography sets the pace of rocky intertidal community dynamics. Proc. Natl. Acad. Sci. U. S. A. 100, 12229–12234. doi: 10.1073/pnas.1534875100
Molares J., Fuentes J. (1995). Recruitment of the mussel Mytilus galloprovincialis on collectors situated on the intertidal zone in the Ria de Arousa (NW Spain). Aquaculture 138, 131–137. doi: 10.1016/0044-8486(95)01085-8
Morán X. A. G., López-Urrutia Á., Calvo-Díaz A., Li W. K. W. (2010). Increasing importance of small phytoplankton in a warmer ocean. Glob. Change Biol. 16, 1137–1144. doi: 10.1111/j.1365-2486.2009.01960.x
Navarrete S. A., Wieters E. A., Broitman B. R., Castilla J. C. (2005). Scales of benthic-pelagic coupling and the intensity of species interactions: From recruitment limitation to top-down control. Proc. Natl. Acad. Sci. U. S. A. 102, 18046–18051. doi: 10.1073/pnas.0509119102
Nolasco R., Gomes I., Peteiro L., Albuquerque R., Luna T., Dubert J., et al. (2018). Independent estimates of marine population connectivity are more concordant when accounting for uncertainties in larval origins. Sci. Rep. 8, 2641. doi: 10.1038/s41598–018-19833-w
Oliveira G. F., Siregar H., Queiroga H., Peteiro L. G. (2021). Main drivers of fecundity variability of mussels along a latitudinal gradient: lessons to apply for future climate change scenarios. J. Mar. Sci. Eng. 9, 759. doi: 10.3390/jmse9070759
Otero J., Álvarez-Salgado X. A., González A. F., Gilcoto M., Guerra A. (2009). High-frequency coastal upwelling events influence octopus vulgaris larval dynamics on the NW Iberian shelf. Mar. Ecol. Prog. Ser. 386, 123–132. doi: 10.3354/meps08041
Otero J., Álvarez-Salgado X. A., González A. F., Miranda A., Groom S. B., Cabanas J. M., et al. (2008). Bottom-up control of common octopus Octopus vulgaris in the Galician upwelling system, northeast Atlantic Ocean. Mar. Ecol. Prog. Ser. 362, 181–192. doi: 10.3354/meps07437
Otero P., Cabrero Á., Alonso-Pérez F., Gago J., Nogueira E. (2023). Temperature and salinity trends in the northern limit of the Canary Current Upwelling System. Sci. Total Environ. 901, 165791. doi: 10.1016/j.scitotenv.2023.165791
Otero P., Ruiz-Villarreal M., Peliz Á., Cabanas J. M. (2010). Climatology and reconstruction of runoff time series in northwest Iberia: influence in the shelf buoyancy budget off Ría de Vigo. Sci. Mar. 74, 247–266. doi: 10.3989/scimar.2010.74n2247
Outeiro L., Byron C., Angelini R. (2018). Ecosystem maturity as a proxy of mussel aquaculture carrying capacity in Ria de Arousa (NW Spain): A food web modeling perspective. Aquaculture 496, 270–284. doi: 10.1016/j.aquaculture.2018.0
Padin X. A., Babarro J. M. F., Silva E., Longa Portabales M.Á., Calvo S., Nolasco R. (2021). Variability in strength of byssus attachment and index condition of subtidal mussels during the maximum growth stage. Aquac. Res. 52, 3485–3497. doi: 10.1111/are.15209
Padin X. A., Velo A., Pérez F. F. (2020). ARIOS: a database for ocean acidification assessment in the Iberian upwelling system, (1976–2018). Earth Syst. Sci. Data 12, 2647–2663. doi: 10.5194/essd-12-2647-2020
Pardo P. C., Padín X. A., Gilcoto M., Farina-Busto L., Pérez F. F. (2011). Evolution of upwelling systems coupled to the long-term variability in sea surface temperature and Ekman transport. Clim. Res. 48, 231–246. doi: 10.3354/cr00989
Pauly D., Christensen V. (1995). Primary production required to sustain global fisheries. Nature 374, 255–257. doi: 10.1038/374255a0
Pérez F. F., Padín X. A., Pazos Y., Gilcoto M., Cabanas M., Pardo P. C., et al. (2010). Plankton response to weakening of the Iberian coastal upwelling. Glob. Change Biol. 16, 1258–1267. doi: 10.1111/j.1365-2486.2009.02125.x
Pérez-Camacho A., Aguiar E., Labarta U., Vinseiro V., Fernández-Reiriz M. J., Álvarez-Salgado X. A. (2014). Ecosystem-based indicators as a tool for mussel culture management strategies. Ecol. Indic. 45, 538–548. doi: 10.1016/j.ecolind.2014.05.015
Pérez-Camacho A., González R., Fuentes J. (1991). Mussel culture in Galicia (N.W. Spain). Aquaculture Biol. Cultivation Mussels 94, 263–278. doi: 10.1016/0044–8486(91)90122-N
Pérez-Camacho A., Labarta U. (2004). Rendimientos y producción del mejillón: bases tecnológicaspara la innovación. In: Bateeiros, mar, mejillión. Una perspectivabioeconómica. Fundación CaixaGalicia. (ed.). Spain, 262.
Pérez-Camacho A., Labarta U., Vinseiro V., Fernández-Reiriz M. J. (2013). Mussel production management: raft culture without thinning-out. Aquaculture 406, 172–179. doi: 10.1016/j.aquaculture.2013.05.019
Pérez-Camacho A., Román-Cabello G. (1979). Estudio del mejillón y de su epifauna asociada en los cultivos flotantes de la Ría de Arosa. II. Crecimiento, mortalidad y producción del mejillón. Bol. Inst. Esp. Oceanog. 267, 22–41. Available at: https://digital.csic.es/handle/10261/312622.
Peteiro L., Filgueira R., Labarta U., Fernández-Reiriz M. J. (2007). Settlement and recruitment patterns of Mytilus galloprovincialis L. in the Ría de Ares-Betanzos (NW Spain) in the years 2004/2005. Aquac. Res. 38, 957–964. doi: 10.1111/are.2007.38.issue-9
Peteiro L. G., Filgueira R., Labarta U., Fernández-Reiriz M. J. (2010). The role of fish predation on recruitment of Mytilus galloprovincialis on different artificial mussel collectors. Aquac. Eng. 42, 25–30. doi: 10.1016/j.aquaeng.2009.09.003
Peteiro L. G., Labarta U., Fernández-Reiriz M. J., Antón Álvarez-Salgado X., Filgueira R., Piedracoba S. (2011). Influence of intermittent-upwelling on Mytilus galloprovincialis settlement patterns in the Ría de Ares-Betanzos. Mar. Ecol. Prog. Ser. 443, 111–127. doi: 10.3354/meps09433
Petraitis P. S., Dudgeon S. R. (2020). Declines over the last two decades of five intertidal invertebrate species in the western North Atlantic. Commun. Biol. 3, 1–7. doi: 10.1038/s42003-020-01326-0
Pfaff M. C., Branch G. M., Wieters E. A., Branch R. A., Broitman B. R. (2011). Upwelling intensity and wave exposure determine recruitment of intertidal mussels and barnacles in the southern Benguela upwelling region. Mar. Ecol. Prog. Ser. 425, 141–152. doi: 10.3354/meps09003
Philippart C. J. M., Amaral A., Asmus R., van Bleijswijk J., Bremner J., Buchholz F., et al. (2012). Spatial synchronies in the seasonal occurrence of larvae of oysters (Crassostrea gigas) and mussels (Mytilus edulis/galloprovincialis) in European coastal waters. Estuar. Coast. Shelf Sci. ECSA 46 Conf. Proc. 108, 52–63. doi: 10.1016/j.ecss.2012.05.014
Philippart C. J. M., Bleijswijk J. D. L., Kromkamp J. C., Zuur A. F., Herman P. M. J. (2014). Reproductive phenology of coastal marine bivalves in a seasonal environment. J. Plankton Res. 36, 1512. doi: 10.1093/plankt/fbu073
Pineda J., Porri F., Starczak V., Blythe J. (2010). Causes of decoupling between larval supply and settlement and consequences for understanding recruitment and population connectivity. J. Exp. Mar. Biol. Ecol. 392, 9–21. doi: 10.1016/j.jembe.2010.04.008
Poloczanska E. S., Brown C. J., Sydeman W. J., Kiessling W., Schoeman D. S., Moore P. J., et al. (2013). Global imprint of climate change on marine life. Nat. Clim. Change 3, 919–925. doi: 10.1038/nclimate1958
Queiroga H., Cruz T., dos Santos A., Dubert J., González-Gordillo J. I., Paula J., et al. (2007). Oceanographic and behavioral processes affecting invertebrate larval dispersal and supply in the western Iberia upwelling ecosystem. Prog. Oceanogr. Ecol. Functioning Iberian Seas: A synthesis GLOBEC Res. Spain Portugal 74, 174–191. doi: 10.1016/j.pocean.2007.04.007
R Core Team (2018). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing).
Reis J. L. C. (2015). Modelação da dispersão e conectividade larvar a NW da Península Ibérica. Aveiro, Portugal: University of Aveiro. Available at: http://hdl.handle.net/10773/16395.
Richardson A. J. (2008). In hot water: zooplankton and climate change. ICES J. Mar. Sci. 65, 279–295. doi: 10.1093/icesjms/fsn028
Rodionov S. N. (2004). A sequential algorithm for testing climate regime shifts. Geophys. Res. Lett. 31, L09204. doi: 10.1029/2004GL019448
Rodrigues L. C., Bergh J. C. J. M. V. D., Massa F., Theodorou J. A., Ziveri P., Gazeau F. (2015). Sensitivity of mediterranean bivalve mollusc aquaculture to climate change, ocean acidification, and other environmental pressures: findings from a producer survey. J. Shellfish Res. 34, 1161–1176. doi: 10.2983/035.034.0341
Rosón G. (1997). A non-stationary box model to determine residual fluxes in a partially mixed estuary, based on both thermohaline properties: application to the Ria de Arousa (NW Spain). Estuar. Coast. Shelf Sci. 44, 249–262. doi: 10.1006/ecss.1996.0127
Ruiz-Villarreal M., Otero P., Cobas-García M., Díaz-Conde M. P., Almeida M. M. (2008). Variability of circulation off N and NW Iberia: insights from a forecast model.
Rykaczewski R. R., Dunne J. P., Sydeman W. J., García-Reyes M., Black B. A., Bograd S. J. (2015). Poleward displacement of coastal upwelling-favorable winds in the ocean’s eastern boundary currents through the 21st century. Geophys. Res. Lett. 42, 6424–6431. doi: 10.1002/2015GL064694
Santos F., Gomez Gesteira M., deCastro M. (2011). Coastal and oceanic SST variability along the western Iberian Peninsula. Cont. Shelf Res. 31, 2012–2017. doi: 10.1016/j.csr.2011.10.005
Sarà G., Gouhier T. C., Brigolin D., Porporato E. M. D., Mangano M. C., Mirto S., et al. (2018). Predicting shifting sustainability trade-offs in marine finfish aquaculture under climate change. Glob. Change Biol. 24, 3654–3665. doi: 10.1111/gcb.14296
Science Advice for Policy by European Academies (SAPEA) (2017). Food from the oceans: Evidence review report (Berlin: Science Advice for Policy by European Academy (SAPEA)). doi: 10.26356/foodfromtheoceans
Scheffers B. R., Meester L. D., Bridge T. C. L., Hoffmann A. A., Pandolfi J. M., Corlett R. T., et al. (2016). The broad footprint of climate change from genes to biomes to people. Science, 354. doi: 10.1126/science.aaf7671
Seed R. (1969). The ecology of Mytilus edulis L. (Lamellibranchiata) on exposed rocky shores : I. Breeding and Settlement. Oecologia 3, 277–316. doi: 10.1007/BF00390380
SGP-MAPA (2022). Contribución de España a las Directrices estratégicas para una acuicultura de la UE más sostenible y competitiva 2021-2030. Secretaría General de pesca. (Madrid, España: Ministerio de Agricultura, Pesca y Alimentación), 170. Available at: https://www.mapa.gob.es/es/pesca/temas/acuicultura/esacui_21_30_sgp_v2_feb_2024_tcm30-636188.pdf.
Shanks A. L., Brink L. (2005). Upwelling, downwelling, and cross-shelf transport of bivalve larvae: test of a hypothesis. Mar. Ecol. Prog. Ser. 302, 1–12. doi: 10.3354/meps302001
Smaal A. C., Ferreira J. G., Grant J., Petersen J. K., Strand Ø. (Eds.) (2019). Goods and Services of Marine Bivalves (Switzerland: Springer Nature). doi: 10.1007/978–3-319–96776-9
Smith J. R., Fong P., Ambrose R. F. (2009). Spatial patterns in recruitment and growth of the mussel Mytilus californianus (Conrad) in southern and northern California, USA, two regions with differing oceanographic conditions. J. Sea Res. 61, 165–173. doi: 10.1016/j.seares.2008.10.009
Soares P. M. M., Lima D. C. A., Cardoso R. M., Semedo A. (2017). High resolution projections for the western Iberian coastal low level jet in a changing climate. Clim. Dyn. 49, 1547–1566. doi: 10.1007/s00382-016-3397-8
Sommer U., Lengfellner K. (2008). Climate change and the timing, magnitude, and composition of the phytoplankton spring bloom. Glob. Change Biol. 14, 1199–1208. doi: 10.1111/j.1365-2486.2008.01571.x
Sorte C. J. B., Davidson V. E., Franklin M. C., Benes K. M., Doellman M. M., Etter R. J., et al. (2017). Long-term declines in an intertidal foundation species parallel shifts in community composition. Glob. Change Biol. 23, 341–352. doi: 10.1111/gcb.13425
Sousa M. C., deCastro M., Alvarez I., Gomez-Gesteira M., Dias J. M. (2017). Why coastal upwelling is expected to increase along the western Iberian Peninsula over the next century? Sci. Total Environ. 592, 243–251. doi: 10.1016/j.scitotenv.2017.03.046
Sousa M. C., Ribeiro A., Des M., Gomez-Gesteira M., deCastro M., Dias J. M. (2020). NW Iberian Peninsula coastal upwelling future weakening: Competition between wind intensification and surface heating. Sci. Total Environ. 703, 134808. doi: 10.1016/j.scitotenv.2019.134808
Stavrakidis-Zachou O., Papandroulakis N., Sturm A., Anastasiadis P., Wätzold F., Lika K. (2018). Towards a computer-based decision support system for aquaculture stakeholders in Greece in the context of climate change. Int. J. Sustain. Agric. Manage. Inform. 4, 219–234. doi: 10.1504/IJSAMI.2018.099235
Suárez M. P., Alvarez C., Molist P., San Juan F. (2005). Particular aspects of gonadal cycle and seasonal distribution of gametogenic stages of Mytilus galloprovincialis cultured in the estuary of Vigo. J. Shellfish Res. 24, 531–540. doi: 10.2983/0730–8000(2005)24[531:PAOGCA]2.0.CO;2
Sukhotin A. A., Flyachinskaya L. P. (2009). Aging reduces reproductive success in mussels Mytilus edulis. Mech. Ageing Dev. 130, 754–761. doi: 10.1016/j.mad.2009.09.005
Sydeman W. J., García-Reyes M., Schoeman D. S., Rykaczewski R. R., Thompson S. A., Black B. A., et al. (2014). Climate change. Climate change and wind intensification in coastal upwelling ecosystems. Science 345, 77–80. doi: 10.1126/science.1251635
Varela R., Lima F. P., Seabra R., Meneghesso C., Gómez-Gesteira M. (2018). Coastal warming and wind-driven upwelling: A global analysis. Sci. Total Environ. 639, 1501–1511. doi: 10.1016/j.scitotenv.2018.05.273
Villalba A. (1995). Gametogenic cycle of cultured mussel, Mytilus galloprovincialis, in the bays of Galicia (N.W. Spain). Aquaculture 130, 269–277. doi: 10.1016/0044-8486(94)00213-8
Widdows J. (1991). Physiological ecology of mussel larvae. Aquaculture Biol. Cultivation Mussels 94, 147–163. doi: 10.1016/0044–8486(91)90115-N
Wilberg M., Livings M., Barkman J., Morris B., Robinson J. (2011). Overfishing, disease, habitat loss, and potential extirpation of oysters in upper Chesapeake Bay. Mar. Ecol. Prog. Ser. 436, 131–144. doi: 10.3354/meps09161
Keywords: mussel aquaculture, mussel seed availability, recruitment time series, upwelling, reproductive output, management, climate change
Citation: Padin XA, Babarro JMF, Otero P, Gilcoto M, Rellán T, Suárez L, Velo A and Peteiro LG (2024) The declining availability of wild mussel seed for aquaculture in a coastal upwelling system. Front. Mar. Sci. 11:1375269. doi: 10.3389/fmars.2024.1375269
Received: 23 January 2024; Accepted: 21 May 2024;
Published: 07 June 2024.
Edited by:
Jorge Paramo, University of Magdalena, ColombiaReviewed by:
Fabian J. Tapia, University of Concepcion, ChileCopyright © 2024 Padin, Babarro, Otero, Gilcoto, Rellán, Suárez, Velo and Peteiro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xosé A. Padin, cGFkaW5AaWltLmNzaWMuZXM=; Laura G. Peteiro, bHBldGVpcm9AaWltLmNzaWMuZXM=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.