
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci. , 03 May 2024
Sec. Marine Megafauna
Volume 11 - 2024 | https://doi.org/10.3389/fmars.2024.1362169
 Danilo Malara1*
Danilo Malara1* Pietro Battaglia2
Pietro Battaglia2 Martin C. Arostegui3
Martin C. Arostegui3 Jonathan J. Dale4
Jonathan J. Dale4 Barbara A. Block4
Barbara A. Block4 Camrin D. Braun3
Camrin D. Braun3 Massimo Brogna5
Massimo Brogna5 Silvestro Greco6
Silvestro Greco6 Teresa Romeo7,8
Teresa Romeo7,8The Mediterranean spearfish (Tetrapturus belone) is a highly migratory predator, also exploited by commercial and recreational fisheries although its catches are poorly reported. The fishing pressure on this species is currently unknown because catch and landing data are fragmentary. Furthermore, more ecological information (e.g. migratory movements) is needed to establish eventual management measures, and ICCAT has not examined the population status yet. Understanding species migratory movements is crucial for addressing conservation challenges and supporting management decisions. In this study we investigated the migratory movements, seasonal patterns, vertical behavior, and thermal preference of T. belone, with the aim to improve information on its ecology and behavior. Overall, six individuals were tagged in the Strait of Messina and Tyrrhenian Sea with pop-up satellite tags and their movements were mainly restricted to the central Mediterranean. Utilization distributions derived from geolocation revealed an overall and seasonal importance of the Tyrrhenian Sea and Strait of Sicily, as well as variability in winter habitat use. While the fish exhibited similar depth preference, spending the majority of time at depths ≤ 10 m, we observed that the Mediterranean spearfish is also able to perform sporadic deep dives below 200 m. Vertical movement showed a bimodal pattern common in other billfish species, characterized by occupation of shallower, warmer waters during the night and deeper, cooler waters during the day. This research increases knowledge on the migration ecology and habitat preference of T. belone which is important for management of this understudied species.
Billfishes (swordfish, sailfish, spearfishes and marlins) are large highly migratory predators including 12 species and two families, i.e., Xiphiidae and Istiophoridae (Nakamura, 1985). They are considered important fishery resources in many countries (Collette et al., 2011; Pons et al., 2017), with populations impacted by both commercial and recreational fishing (Holland et al., 1998; Ditton and Stoll, 2003). Globally, less than half of billfish stocks exhibit healthy biomass levels and nearly a quarter are still experiencing overfishing (Pons et al., 2017). However there are cases (i.e. most spearfishes) where catch data are discontinuously reported and stocks have not yet been assessed (Punt et al., 2015; Pons et al., 2017). Technological advances have significantly increased our understanding of billfish biology and ecology, including through the use of electronic satellite tags to reveal horizontal and vertical movement behaviors (Braun et al., 2015). Previous satellite tagging studies have primarily focused on swordfish, sailfish, and marlins, while only a limited amount of research has been devoted to spearfishes (Tetrapturus spp.). Indeed, over 1000 satellite tags were deployed on billfish globally by 2014, however only 0.2% were on spearfish species (Braun et al., 2015), highlighting the need for more research on spearfishes.
Among spearfishes, Tetrapturus belone is endemic from the Mediterranean Sea (MED), where it is almost exclusively distributed (Nakamura, 1985; Collette and Graves, 2019). The status of this stock has not yet been assessed by ICCAT, due to the lack of sufficient data, although the species is assessed as Least Concern in the IUCN Red List of Threatened Species (Collette et al., 2023). Information on the distribution patterns and seasonal movements of T. belone in the MED is still incomplete and data on the occurrence of this species in the Levantine Basin (Bariche and Fricke, 2020; Gerovasileiou et al., 2020; Saad et al., 2024) and western MED (Robins and de Sylva, 1963) are often fragmentary. However, T. belone is distributed around the Italian peninsula, primarily found in the Strait of Messina, Tyrrhenian Sea and Strait of Sicily (Di Natale et al., 2005, 2011; Di Natale, 2012). Furthermore, many of the fisheries taking T. belone occurred (i.e. driftnets) or are occurring (i.e. longlines, harpoon, recreational fishing) mostly in spring, summer or early autumn. This temporal bias may limit our understanding of the species’ distribution during late autumn and winter. Additionally, there is a lack of information regarding the distribution of juvenile individuals.
T. belone is an important fish resource for some Mediterranean local markets, in particular in the area of the Strait of Messina where this species is caught and commercialized by local professional harpoon fishers (Romeo et al., 2009a, 2015; Malara et al., 2020). However, catch and landings data are often under-reported (Collette et al., 2023), thus contributing to the lack of information on this species. Most data available in the literature comes from catch series obtained from harpoon (e.g., Di Natale et al., 2005; Romeo et al., 2015), driftnets (currently banned) and longline fisheries (Di Natale et al., 2003). According to Collette et al. (2023), landings are largely unknown, but they have recently increased (above 100 t/year), despite only Italy, Spain and Portugal occasionally and partly providing catch data to ICCAT. Recently, some fragmentary additional data are also available from recreational fishing (Collette et al., 2023) and the European Union Marine Strategy Framework Directive.
According to the available knowledge, the Strait of Messina and surrounding areas play an important role in different life stages of the Mediterranean spearfish providing nursery, reproductive and foraging habitats (Spartà, 1960). Adult T. belone feed primarily on epipelagic fish and cephalopods (Romeo et al., 2009a, 2012), but can also act as a gelatinous plankton opportunistic consumer (Cardona et al., 2012). The regular presence of T. belone in the Strait of Messina during late spring and summer (mainly from May to July/August) is thought to be related to reproduction (Spartà, 1960; Potoschi, 2000) but may also be associated with food availability (Robins and de Sylva, 1963).
To date, there is minimal information about the movements, behavior, ethology, and ecology of T. belone (Robins and de Sylva, 1963; Braun et al., 2015). Only recently, one study investigated the horizontal and vertical movement behavior of T. belone in the MED collecting data on one tagged individual (Arostegui et al., 2019a) while another tagging study failed due to premature death and a subsequent scavenging event (Malara et al., 2020). Arostegui et al. (2019a) discovered that, on average, T. belone spent 93% of each 24-hr period above 30 m and preferred water layers between 26–28°C but occasionally descended into deeper, colder waters. In addition, the tagged fish exhibited a diel vertical migration spending more time in the near-surface at night and in deeper water layers during the day.
The aim of this paper is to help fill the knowledge gap on T. belone by investigating the movements and behavior of multiple individuals of this species in Mediterranean waters via satellite tagging. Specifically, we document the seasonal distribution and assess the depth and thermal preference of T. belone, relating the behavior to the available environmental data from tag-recorded parameters and to the diel cycle. These data will be useful as they increase knowledge on the species’ behavior and can help guide management decisions.
Pop-up satellite archival transmitting (PSAT) X-tags (Microwave Telemetry, Inc.) were used to perform satellite tagging on T. belone and deployed in the Strait of Messina, off north-eastern Sardinia, and off the north-western coast of Sicily (Figure 1). The weight of each tagged individual was visually estimated by fishers and researchers.
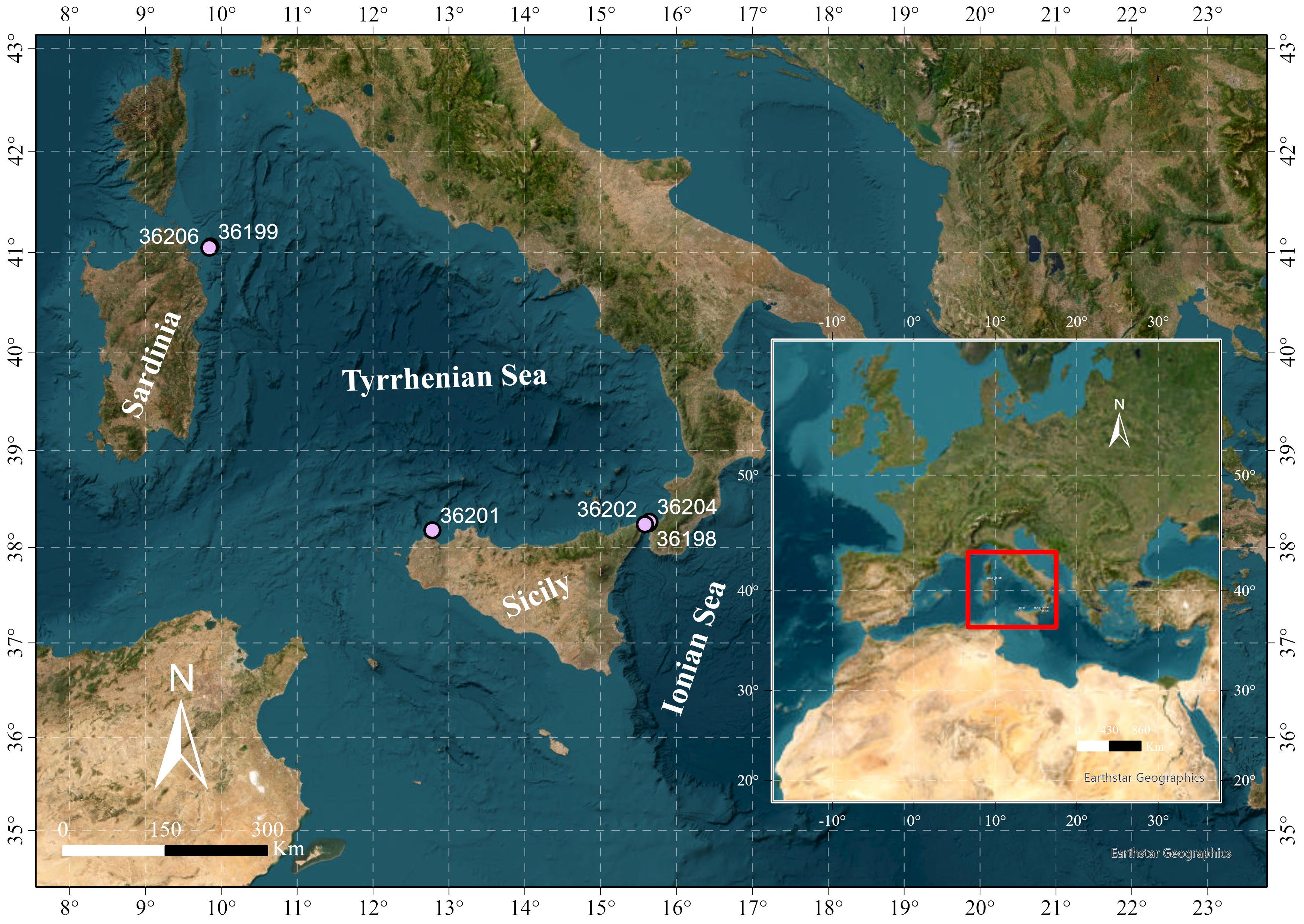
Figure 1 Sampling area. Points indicate the position of individual tagged animals during tag deployment.
The Strait of Messina is located in the central Mediterranean Sea and is the connection point between the Ionian and Tyrrhenian basins. This area is important because of the strong and turbulent tidal currents which allow colder, saltier and nutrient-rich deep Ionian waters to mix with shallower and warmer Tyrrhenian waters having a different density (Mosetti, 1991). In addition, the Ionian bottom morphology drives deep water rich in nutrients to the surface, thanks to upwelling currents, sustaining high levels of primary production (Spanò and De Domenico, 2017).
Cape San Vito is the western geographic limit of the Gulf of Castellammare (the widest bay in the northern coast of Sicily with >70 km of coastline). Here, easterly anticyclonic currents are responsible for the water circulation inside the Gulf (Istituto Idrografico della Marina, 1982). The seafloor morphology consists of a narrow or absent continental shelf above a very steep continental slope that steps down up to 720 m in the eastern part and up to 1100 m in the northern (Sulli et al., 2021). In addition, this area is characterized by submarine canyons (Sulli et al., 2021).
The Sardinia Island is situated between the Sardinia Sea on the west and Tyrrhenian Sea on the east. The Sardinia Sea is characterized by the presence of anticyclonic eddies and upwelling events (Ribotti et al., 2004), whereas, currents are generated by the action of water masses from the Western Tyrrhenian sea to the east of the island. The area around Sardinia Island is highly productive due to wind action from the Strait of Bonifacio that creates eddies and also mixing of different water masses from other parts of the MED (i.e. Modified Atlantic Waters, Levantine Intermediate Waters, etc.; Olita et al., 2013).
Based on the geographical location, two different fishing and tagging methods were used:
1. Strait of Messina: tagging operations were conducted by researchers on board two professional fishing vessels, which usually practice traditional swordfish harpoon fishing in this study area (Romeo et al., 2015; Battaglia et al., 2018). Individuals were sighted by experienced professional fishers located on a pulpit at the top of a mast (for a description and images of these fishing activities see Battaglia et al., 2018). The tagging operation was performed using a modified 3.5 m harpoon pole without taking the fish on board. This method was similar to the one used by Malara et al. (2020), but a compressed rubber stopper was attached to the application pin to prevent the pin from penetrating the flesh more than 4 cm. The pole was also equipped with an action camera (GoPro Hero7) to record the tagging activity. The tags were applied into the dorsal musculature near the base of the dorsal fin, behind the head, using a Domeier anchor dart attachment (Table 1).
2. North-eastern Sardinia and north-western coast of Sicily: the other tags were deployed using sportfishing vessels. The fish were caught on rod-and-reel with trolled lures and tagged by recreational anglers. The maximum duration of fighting was 7 minutes and the handling time was about 3 min. The tags were applied using a 1.5 m pole equipped with the previously described compressed rubber stopper. Tags were inserted into the dorsal musculature near the dorsal fin using a Domeier anchor dart attachment (Table 1).
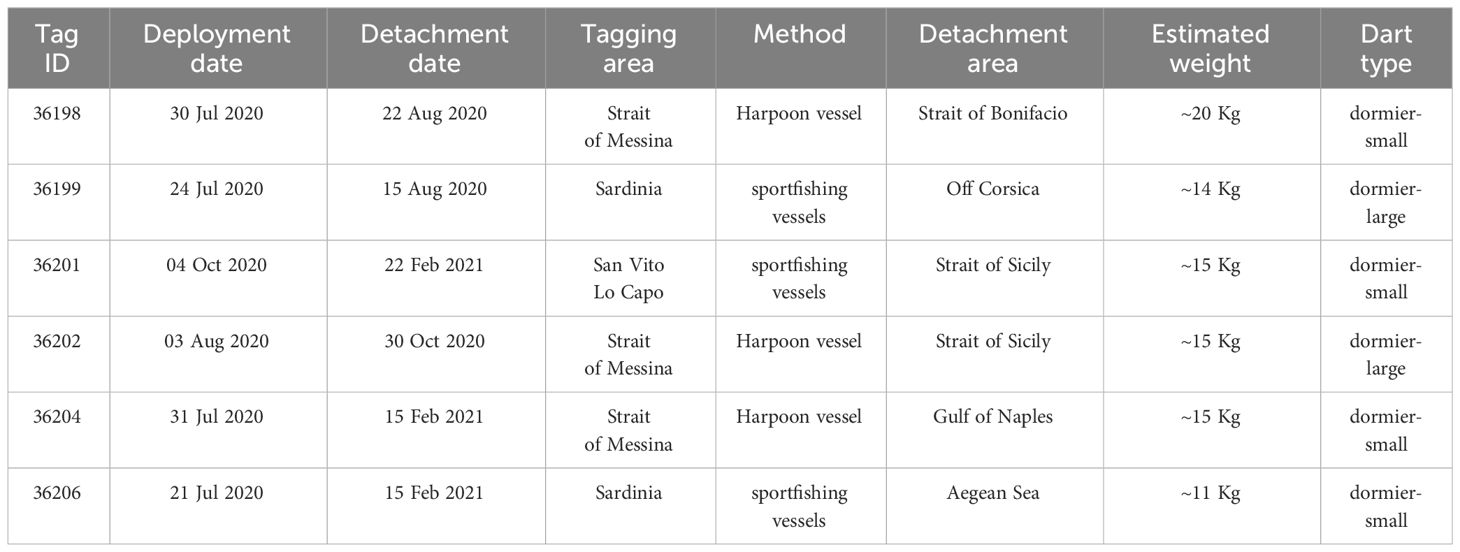
Table 1 Summary of tag IDs, deployment and detachment dates, area of tagging, estimated tagged animal weight (Kg) and dart type.
Tags were programmed for 240-d deployments in which they recorded temperature, pressure (depth) and light measurements at 2-min resolution when the tag was in data collection mode. After pop-off, the tags transmitted summarized versions of stored data via Argos satellites. Transmitted data resolution was dependent on deployment duration: resolution of 15 min when deployment duration was <120 days or resolution of 15–30 min when deployment duration was 120–240 days. Data were logged to onboard memory and summarized into a time series of depth and temperature.
Geolocation of the tagged Mediterranean spearfish was conducted using the HMMoce package for R (Braun et al., 2018). This gridded hidden Markov model approach compares diverse tag-based observations against remote sensing and data-assimilating oceanographic model outputs to generate likelihoods of a tagged individual’s location and behavior at each time step of its deployment. At 24-hr intervals, we calculated six separate likelihoods: (a) bathymetry calculated by excluding areas shallower than the tag-recorded maximum depth in the SRTM30_PLUS dataset (Becker et al., 2009), light-based (b) latitude and (c) longitude determined by manufacturer post-processing, (d) sea surface temperature (SST) generated from comparing tag-based SST values (< 10 m depth) against the Multi-scale Ultra-high Resolution (MUR) SST dataset (NASA/JPL, 2015), and a time series of tag-based depth-temperature profile data (15-s resolution) against the global 1/12o Hybrid Coordinate Ocean Model (HYCOM; Bleck, 2002; Chassignet et al., 2007) in terms of e) integrated ocean heat content (Luo et al., 2015) and (f) the daily depth-temperature products at the standard depth levels. The data-assimilating HYCOM reanalysis product adequately represents real oceanographic processes and variability in the MED (Wang et al., 2023), justifying the potential inclusion of depth-temperature likelihoods derived from comparison with this ocean model. All likelihood grids were resampled to 0.08o spatial resolution to match the native HYCOM grid. Furthermore, all likelihood calculations excluded any delta-limited temperature and depth measurements, which are potentially over- or underestimated measurements flagged by the tag manufacturer’s software because of data compression limitations onboard the tags. Similarly, the light-based latitude and longitude likelihood calculations excluded raw light-based estimates when the raw estimates exceeded the five-day rolling mean by more than one degree.
For each fish, we tested three alternative likelihood combinations as inputs to the hidden Markov model to determine which yielded the best geolocation: (1) bathymetry + latitude + longitude + SST, (2) bathymetry + latitude + longitude + SST + HYCOM ocean heat content, and (3) bathymetry + latitude + longitude + SST + HYCOM depth-temperature. The resulting observation likelihoods were convolved with a diffusive movement kernel for a single behavior state. Parameter estimation of behavior state movement used bound-constrained optimization (Byrd et al., 1995). Parameter bounds and the initial value for the movement kernel were informed by the daily displacement rates (km/d) estimated from previous telemetry studies of Mediterranean spearfish (Arostegui et al., 2019a) and white marlin (Kajikia albida; Hoolihan et al., 2015). We used Akaike information criterion (AIC) weight (Wagenmakers and Farrell, 2004) to identify the best-fit model resulting from the three different input likelihood combinations we tested. The daily posterior likelihood surfaces generated by the best-fit models were summed for each fish to yield their time-integrated, spatial utilization distributions (UDs) throughout the overall deployment and seasonally (Winter: Dec-Feb | Spring: Mar-May | Summer: Jun-Aug | Fall: Sep-Nov). The most probable track for each deployment was generated with the Viterbi method, a global decoding solution that controls path admissibility with the daily posterior likelihoods to generate realistic movement tracks in areas of complex topography that is applicable to both demersal (Nielsen et al., 2023) and pelagic species (Arostegui et al., 2024).
Seasonal time at depth (TAD) and time at temperature (TAT) histograms were obtained by the analysis of raw depth and temperature time series across fish. using the RchivalTag package (v. 0.1.2; Bauer, 2018) in Rstudio (Posit team, 2023). While our analyses focused primarily on the movements of T. belone, we acknowledge the importance of the well-marked thermocline in the Mediterranean Sea, which varies in depth and consistency from mid-spring to mid-autumn.
To address this concern, we calculated the daily thermocline depth by interpolating temperature and depth at a resolution of 10 meters and then averaging per location (Sicilian Channel, Tyrrhenian Sea, Ionian Sea, and Aegean Sea) and seasons. Additionally, for each location, we obtained the tag-based sea surface temperature (SST) by averaging the seasonal temperature in the first 5 meters of the water column.
In order to assess the preferential depth of T. belone and to relate its movements to water temperature, we calculated the proportion of time spent by the tagged individual in each depth (0–10, 10-20, 20-30, 30-40, 40-50, 50-60, 60-70, 70-80, 80-90, 90-100, >100 m) and temperature (< 16, 16-18, 18-20, 20–22, 22–24, 24–26, 26–28, 28-30, >30°C) bin. These proportions were estimated for each 24-hour period and then averaged to determine the mean TAD and TAT profiles. Additionally, data were further classified into nautical daytime (dawn to dusk) and nautical nighttime (dusk to dawn) categories to evaluate circadian changes in depth and temperature distribution.
To calculate nautical dawn and dusk data for each geolocation point, geographical coordinates estimated by the tracking model were utilized, along with the ‘suncalc’ package (v. 0.5.0; Thieurmel and Elmarhraoui, 2019). To mitigate potential biases related to anomalous fish behavior due to the tagging operation or possible mortality events, data from the deployment and detachment days were excluded from the analysis.
The distance between deployment and detachment points of the calculated tracks position, the total distance travelled, and the daily distance travelled were calculated using the R package “geosphere” (V. 1.5-14). Kruskal-Wallis rank sum test (Kruskal and Wallis, 1952) followed by Conover-Iman post-hoc test and Bonferroni p-value adjust (Conover and Iman, 1979; Conover, 1999) were used to test potential differences in depth or thermal preference, while Two-sample Wilcoxon test (Bauer, 1972; Hollander et al., 2014) was used to investigate if there were significant diel differences in depth distribution or thermal preference.
Six individuals of T. belone were tagged ranging between 11 and 20 kg (mean ± SD = 15 ± 2.64 kg; Table 1). All of the deployed tags transmitted data, covering different periods of the year (summer, fall and winter), but no fish had deployment coverage in spring. Deployment durations ranged from 23 to 210 days (mean ± SD: 115 ± 76), for a total of 688 days of observation data from tags. Overall, 150 days (30 ± 7) were recorded in summer, 300 days (75 ± 16) in fall, and 238 days (79 ± 3) in winter (Table 2). Premature detachments of two tags (tag n. 36199 and 36202) were potentially due to mortality events, whereas in the other cases the tag and leader apparently detached from the fish which could be related to a pull out of the dart or failure at the leader.
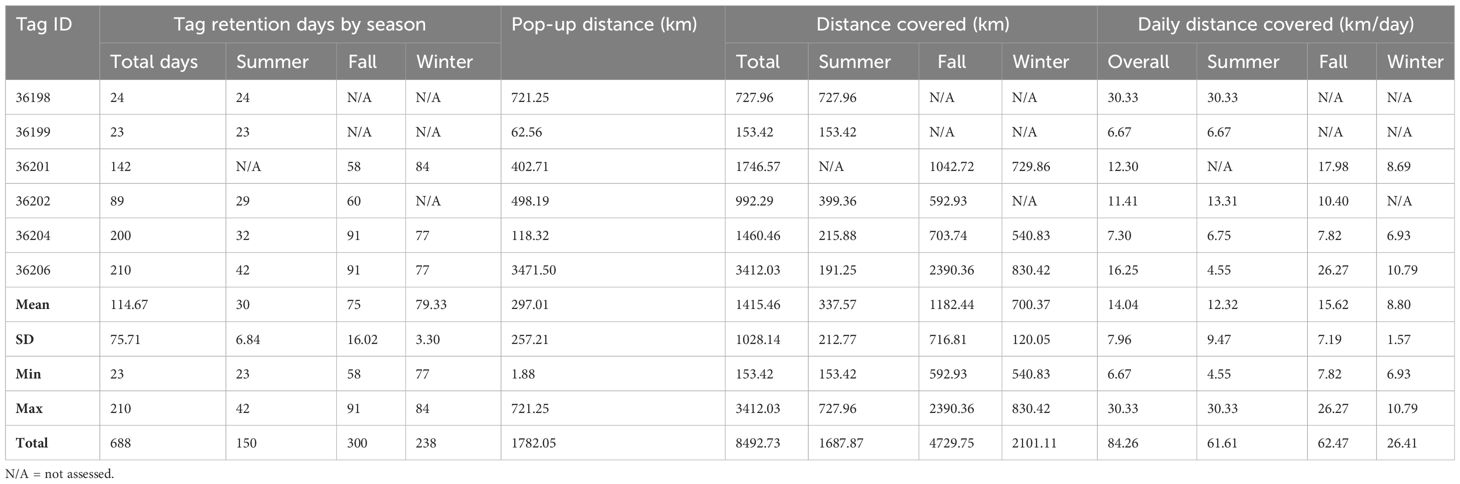
Table 2 Summary of the total tagging period and distance travelled by each tagged T. belone.
Tagged individuals remained within the MED and none of the fish went west of Corsica s Sardinia and/or into Adriatic waters (Figure 2). Generally, fish movements were restricted to the Tyrrhenian Sea and central Mediterranean, but in one case (tag n. 36206) the fish reached the Aegean Sea before the tag detached. The fish covered a total distance of 8492.73 km (1415.46 ± 1028.14 km) with an average speed of 14.04 ± 7.96 km/day.
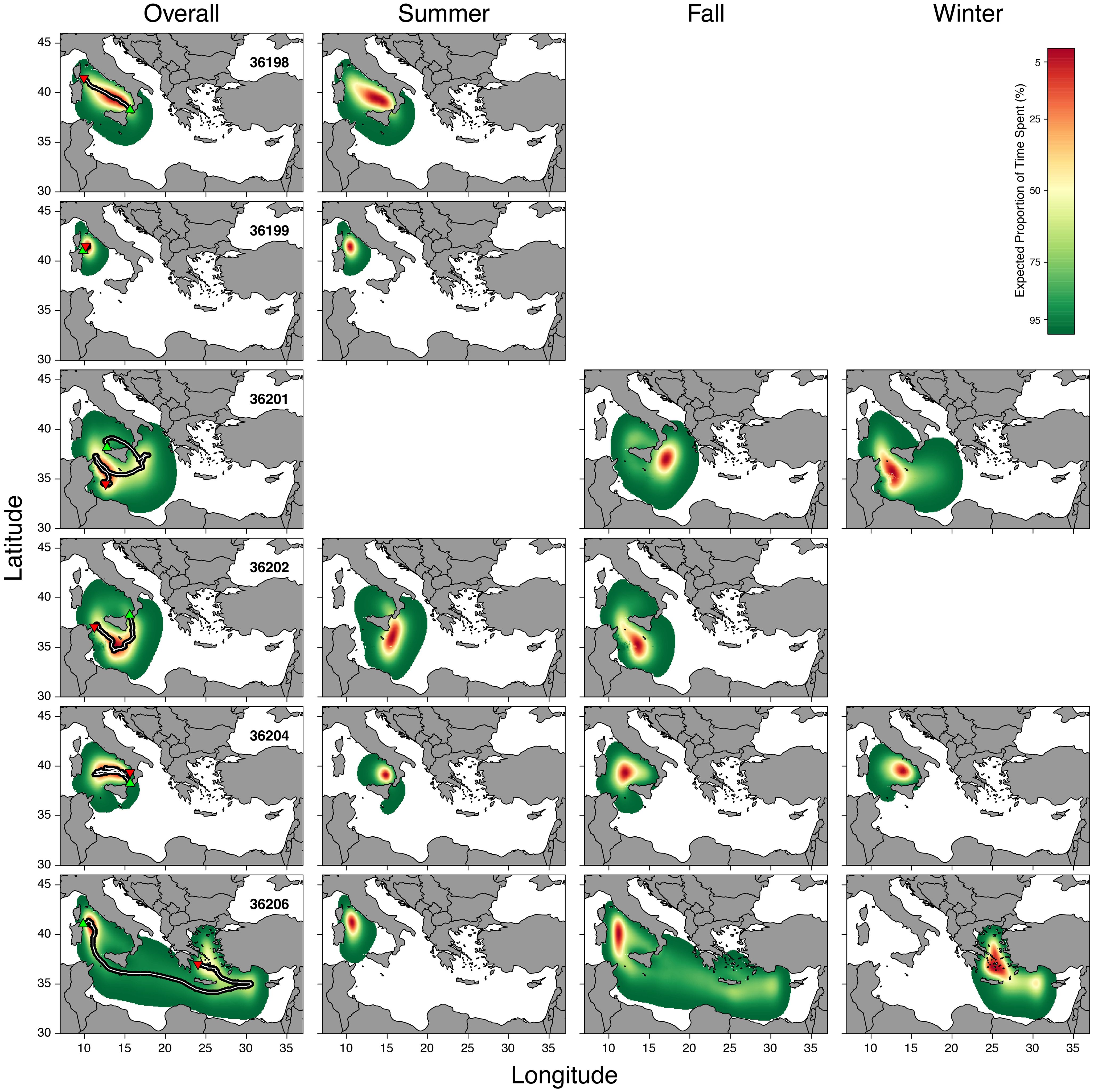
Figure 2 Graphic representation of the utilization distribution (UD) for all tagged fish, deployment and detachment locations (upward green and downward red triangles, respectively), overall utilization distributions with daily positions (black underlying dots) and resulting track lines (white lines). Blank panels represent seasons not included in the tag deployment. Color bar refers to the expected proportion of time spent.
Analysis of utilization distribution patterns (Figure 3) revealed a high importance of the Tyrrhenian basin and Strait of Sicily for Mediterranean spearfish movements and habitat use. The examined data showed that the core habitat (5% UD) for Mediterranean spearfish was mainly concentrated in the Tyrrhenian Sea, where tagged individuals remained for long periods during their horizontal movements across all seasons.
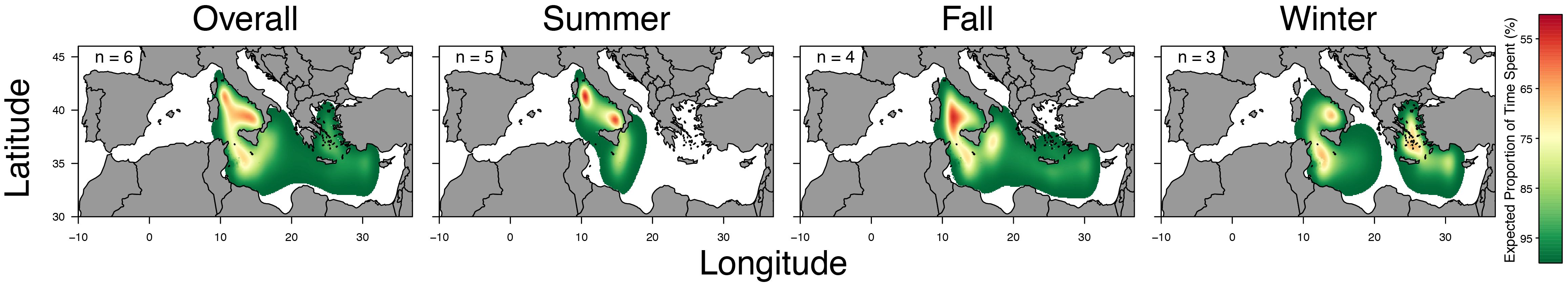
Figure 3 Graphic representation of the overall and seasonal utilization distribution (UD) across fish. Color bar refers to the expected proportion of time spent.
In summer, the core habitat occurred in two areas of the Tyrrhenian Sea: the northern Tyrrhenian as well as between the Aeolian archipelago and the southern Italian peninsula. The estimate of core habitat was wider in fall and mainly centered between Sardinia and Sicily, although the estimate records a small percentage of core habitat in the Ionian Sea and Strait of Sicily. In winter, the core habitat was mainly located in three different Mediterranean areas, i.e. waters off the Aeolian archipelago, Strait of Sicily off Tunisian coasts and Greek waters in the Aegean Sea.
Home range (<50% UD) habitat of T. belone overlapped with Tyrrhenian Sea, central Mediterranean basin and the eastern Mediterranean up to Cyprus. This pattern is also observed during fall, but considerable seasonal and individual variation in home range size were observed. For instance, individuals displayed a reduced home range in summer, expanding only between northern Tyrrhenian and Libyan coasts. However, the easterly expansion during the fall and winter was driven by the movement of a single fish.
The six tagged individuals largely remained in the upper epipelagic layer above 50 m across all seasons (Figure 4), spending approximately 99% of time at or above this depth, with a preference for the first 10 m (74.64 ± 14.88%; Figure 5A). Significant differences were found in depth use within all seasons (KWsummer: df = 10, χ2 = 1038.09, P < 2.2e-16; KWfall: df = 10, χ2 = 1422.88, P < 2.2e-16; KWwinter: df = 10, χ2 = 914.67, P < 2.2e-16), with T. belone spending a significantly greater amount of time in the first 10 m of water (p < 2.2e-16; Figure 5A; Supplementary Table S1). There were also significant differences in diel depth use, mainly due to higher time spent at 0 – 10 m during the night and 20 – 50 m during the day (Figure 5A; Supplementary Figure S1A). Below 50 m there was no diel difference in the time spent at 60 – 70 m and > 90 m (Supplementary Figure S1A). The maximum recorded seasonal depths were 167.0, 322.8, and 172.1 m in summer, fall and winter, respectively (Table 3; Figure 6A). Only one individual (n. 36206) reached depths below 200 m, performing a total of 4 dives into the mesopelagic zone during fall (Figure 6A; Supplementary Figure S2).
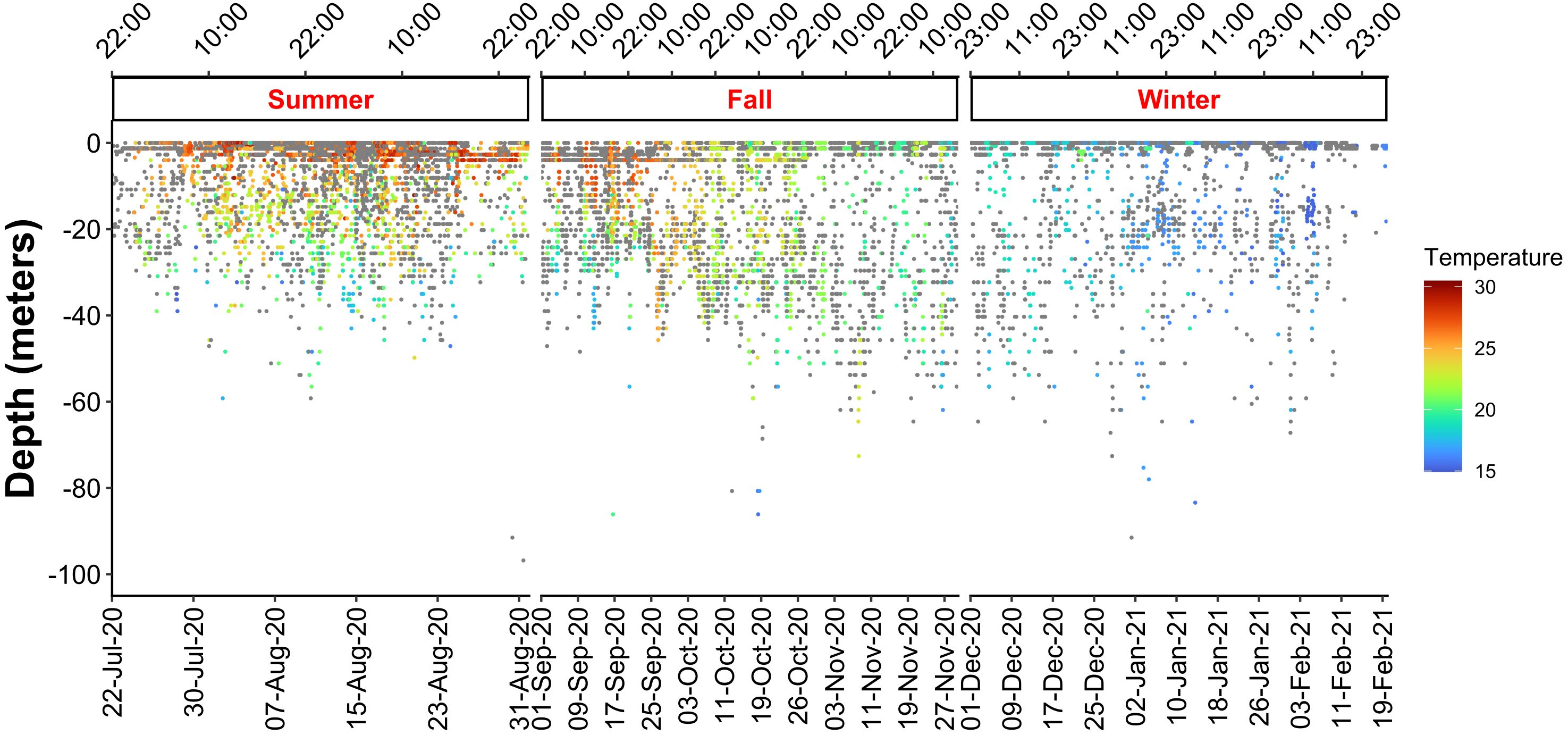
Figure 4 Seasonal representation of the depth-temperature experienced by T. belone. Color dots indicate the temperature achieved by the fish during the various seasons at each depth. Grey dots indicate depth recorded by the tag without associated temperature.
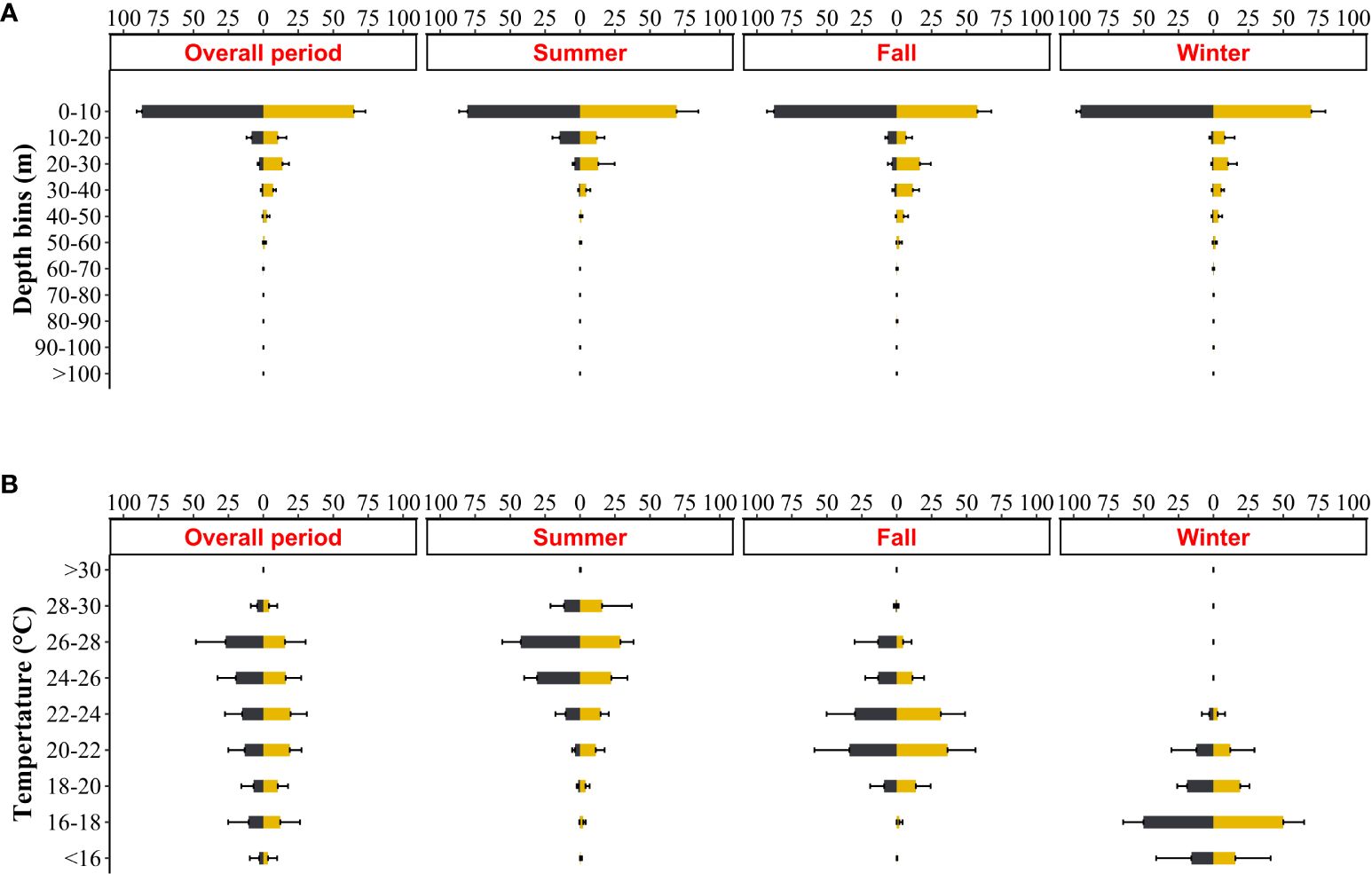
Figure 5 Diel percentage of time spent at depth (A) and temperature (B) across tagged fish during the overall period and different seasons (Summer, Fall and Winter). Yellow bars indicate daytime while black bars indicate night-time. Depth bins were: 0-10, 10-20, 20-30, 30-40, 40-50, 50-60, 60-70, 70-80, 80-90, 90-100 and >100 m. Temperature bins: <16, 16-18, 18-20, 20-22, 22-24, 24-26, 26-28, 28-30, >30.
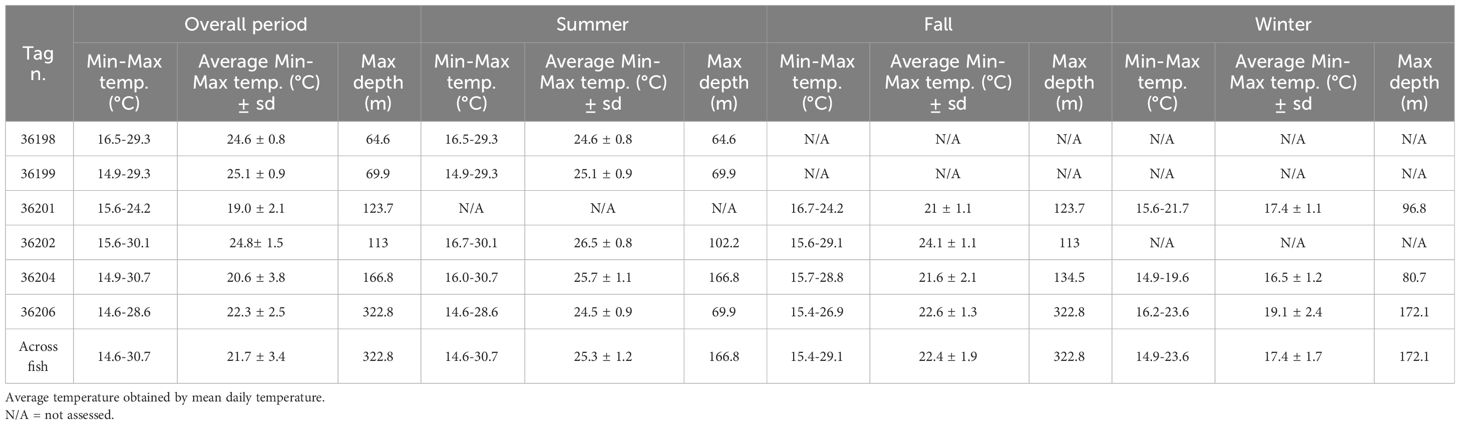
Table 3 Summary of the seasonal maximum and minimum temperature, the seasonal average ± sd temperature and the seasonal maximum depth recorded by the tags.
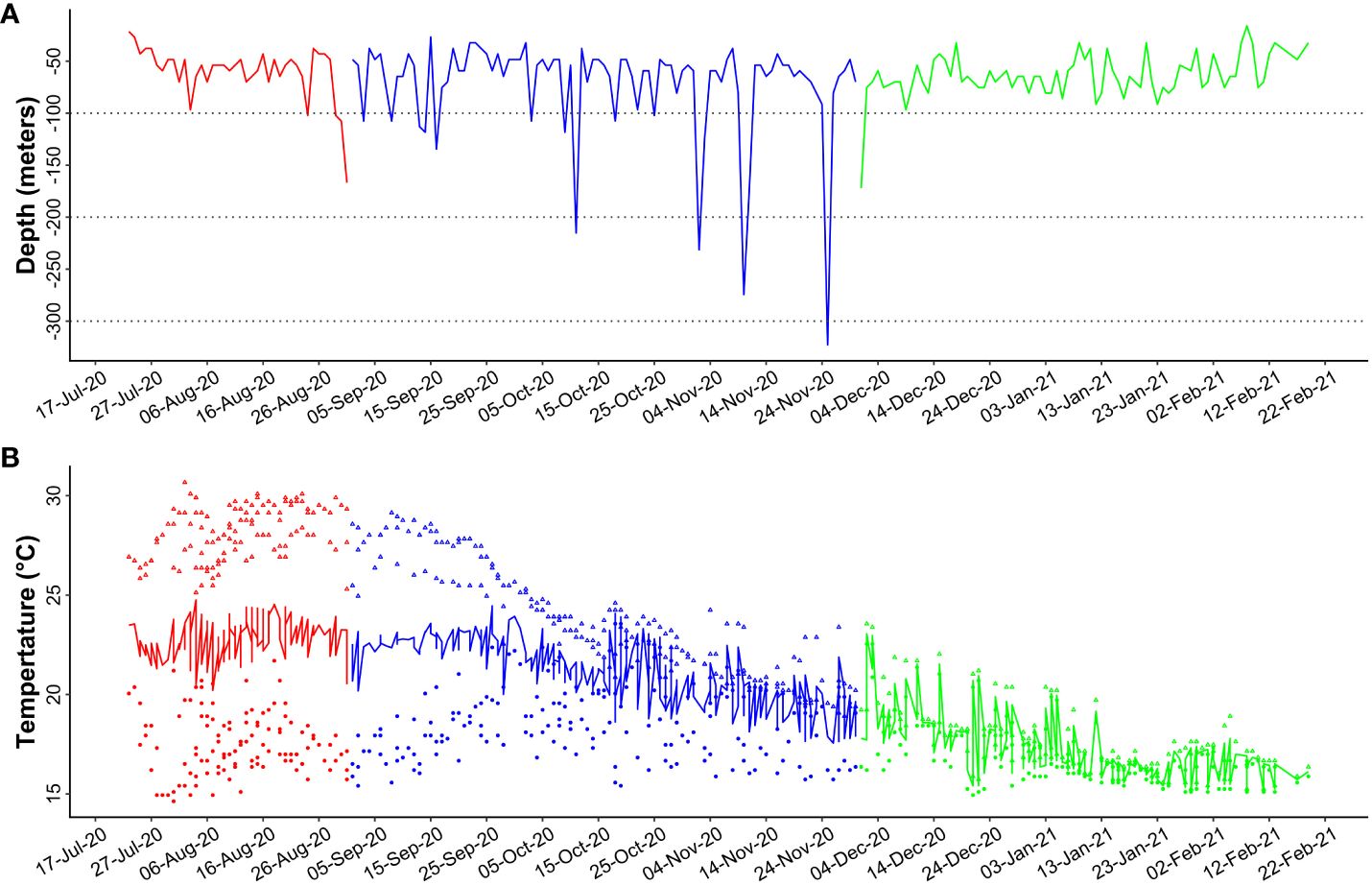
Figure 6 Mean maximum depth (A) and mean maximum, minimum and average temperature (B) recorded by all tags during the tracking period. Red line indicates summer period, blue line the fall, green line winter period. Black dot lines in A indicate 100, 200 and 300 depth limits.
Tagged individuals encountered environmental temperatures ranging from 14.6 and 30.7°C in summer (mean T = 25.3 ± 1.2°C), between 15.4 and 29.1°C in fall (mean T = 22.4 ± 1.9°C) and between 14.9 and 23.6°C in winter (mean T = 17.4 ± 1.7°C) (Table 3; Figure 6B). More than 85% of time was spent between 24 and 30°C in summer (Figure 5B), and time spent between 26 and 28°C was significantly higher (34.51% ± 5.99) than other temperature bins (Supplementary Table S2). Tetrapturus belone spend 78% of the time between 20 and 26°C in fall. During this season, the time spent between 20 and 22°C (35.29% ± 22.17) and between 22 and 24°C (30.96% ± 18.51) were significantly higher than the period spent at other temperatures. In winter, T. belone encountered temperatures between 16 and 20°C for about 85% of time and the highest percentage of time (50.06% ± 14.71) was spent between 16 and 18°C.
Differences in the diel pattern of thermal habitat use were also observed (Figure 5B, Supplementary Figure S1B). During summer, T. belone spent significantly more time between 24 and 28°C at night and cooler temperature during the day. No statistical differences were present at temperatures > 28°C (Supplementary Figure S1B). In fall, temperature 20 – 24°C is occupied mainly during the night while daytime is restricted between 16 and 20°C (Supplementary Figure S1B). There were no differences in time spent during the day/night period at temperature cooler than 16°C and hotter than 24°C, and no differences in diel temperature use was found in the winter period (Supplementary Figure S1B).
Our analysis of tagged fish revealed seasonal variations in Sea Surface Temperature (SST) across the Mediterranean Sea, as summarized in Table 4. However, no data were available for the Aegean Sea and Sicilian Channel in summer, nor during the winter in the Ionian Sea. Through summer, temperatures ranged between 23.6°C and 28.9°C, with an average SST of 26.4°C ± 1.2°C. The Ionian Sea exhibited the highest average temperature (27.7°C) among all regions. In fall, SST ranged from 18.7°C to 27.1°C, with an average of 23.1°C ± 2.4°C, observed across the Ionian Sea, Sicilian Channel, and Tyrrhenian Sea. The warmest temperatures were recorded in the Sicilian Channel (24.4°C), while the lowest were in the Ionian Sea (21.1°C). In winter, SST decreased across all seas, ranging from 15.2°C to 21.0°C, with an average of 17.2°C ± 1.3°C, and the Tyrrhenian Sea experiencing the coolest temperatures.
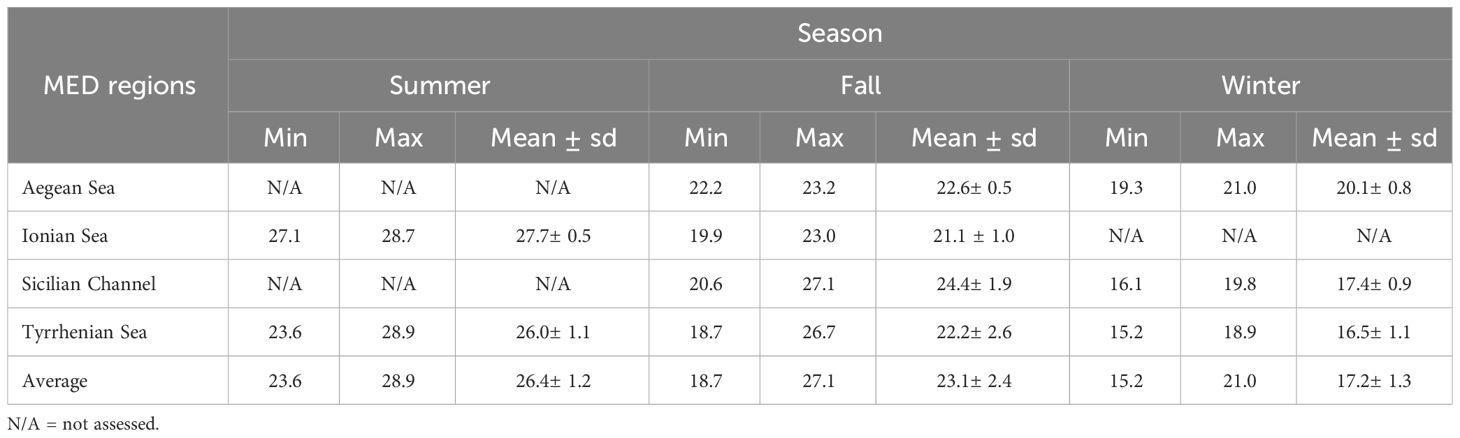
Table 4 Seasonal variation in tag-based sea surface temperature in different MED regions.
Due to the resolution of the tag data, coupled with the vertical behavior of the tagged individuals, we were not able to calculate the thermocline and mixed layer depth reliably, and therefore we did not include this analysis.
This research documented the seasonal variability in horizontal and vertical movements, as well as the thermal and depth occupation, of six T. belone in the MED using PSAT tags. Our data supplement and permit inference beyond the information from the first tagging experiment on this species (Arostegui et al., 2019a) which was restricted to a single individual.
In general, tagged spearfish dispersed from the release locations, moving to other areas of the Tyrrhenian Sea, Strait of Sicily, Ionian Sea and, in one case, towards Cypriot and Greek waters, where this species is considered less common (Akyol, 2020; Gerovasileiou et al., 2020). None of the tagged fish visited the western MED beyond Sardinia or to the Adriatic Sea, although, no data are available in spring, due to the premature detachment of the tags. T. belone showed significant mobility within the Mediterranean basin, with a mean straight-line distance from the tag area of 297.01 km, a mean total track distance of 1415.46 km, and mean daily displacement rate of 14.04 km/day. The only direct comparison could be done with the T. belone tagged in summer 2015 in the MED (Arostegui et al., 2019a), which travelled at a matching mean speed of 14.51 km/day. Similarly high extents of dispersal and degrees of mobility are known from other billfishes such as Kajikia audax, Istiompax indica and Makaira nigricans in other areas (Domeier, 2006; Rohner et al., 2020, 2021; Andrzejaczek et al., 2023).
Despite the variation in movement behavior among individuals, in most cases, fish remained within the Tyrrhenian Sea or around Sicily. Previous research suggests these areas are regularly used by T. belone as feeding (Romeo et al., 2009a, 2012) and reproductive grounds (Spartà, 1960; De Sylva, 1975). The spawning period of T. belone occurs between May and September, with eggs and larvae found from late spring to summer and juveniles in the fall in the Strait of Messina (Spartà, 1960). Therefore, our results support the hypothesis that the Strait of Messina and the surrounding seas are important areas for the ecology and life-history of T. belone. Previous studies have also identified the Strait of Messina and southern Tyrrhenian Sea as important spawning and/or nursery grounds for other large pelagic fishes such as swordfish and bluefin tuna (Nishida et al., 1998; Romeo et al., 2009b; Di Natale et al., 2018). The occurrence of juvenile spearfish as far east in the MED as the waters of Israel and Lebanon (Nakamura, 1985) and seasonal influxes of adult spearfish (some observed in pairs or aggregations seemingly engaged in courtship and/or spawning) as far west as Mallorca (Vince Riera, pers. comm.) and the southern coast of France (Bill François, pers. comm.) suggest the presence of spawning grounds throughout the basin. Yet, none of the tagged individuals in this study migrated to these longitudinally distant locations. Further long-term tagging to be carried out in more areas of the MED, is needed to elucidate the stock structure and potential connectivity or regional fidelity of Mediterranean spearfish. Furthermore, while our study provided evidence of the movements of T. belone within the Tyrrhenian Sea, around Sicily and in one case also in eastern MED, it is important to acknowledge that the species’ distribution extends beyond these regions. Additional literature suggests that T. belone is also present in various parts of the Mediterranean, including the Adriatic Sea and the Ligurian Sea (Dulcic and Soldo, 2004; Psomadakis et al., 2012). Moreover, the species is expected to inhabit the entire Mediterranean Sea, encompassing areas such as the Strait of Gibraltar and the near Atlantic region adjacent to the Strait (Nakamura, 1985; Collette and Graves, 2019). Therefore, future research efforts should aim to explore the movement patterns and distribution of T. belone across the broader Mediterranean basin to obtain a comprehensive understanding of its ecology and population dynamics.
Mediterranean spearfish in this study preferred the epipelagic environment as suggested in a previous study (Arostegui et al., 2019a), with more than 99% of time spent above 50 m and more than 70% at 0-10 m. Despite the MED being a temperate basin with unique oceanography, the epipelagic behavior was also observed in billfishes living in other oceanic areas (mostly tropical and equatorial) characterized by different physical-chemical features: T. angustirostris (Arostegui et al., 2019b, 2024), M. nigricans (Andrzejaczek et al., 2023), K. albida (Vaudo et al., 2017), T. pfluegeri (Kerstetter et al., 2009), I. indica and K. audax (Rohner et al., 2022). In addition, tagged individuals were observed to spend nearly 99% of their time above the 50-meter mark, suggesting a foraging strategy that focuses on prey within the epipelagic zone (Romeo et al., 2009a, 2012). However, some differences between night and daylight were observed, with the tendency to occupy deeper depths during the day than night. This was also observed in other billfishes such as T. pfluegeri (Kerstetter et al., 2009), M. nigricans (Andrzejaczek et al., 2023), Kajikia albida (Vaudo et al., 2017), and Istiophorus platypterus (Pohlot and Ehrhardt, 2017) but the reverse of the pattern characteristic of T. angustirostris (Arostegui et al., 2019b, 2024). Furthermore, recent study by Arostegui et al. (2019a) reported a maximum depth of 84 m reached by an individual equipped with a PSAT tag, while Nakamura (1985) supposed a vertical distribution not exceeding 200 m. However, we observed for the first time that this species is also able to perform deep excursions into mesopelagic waters, with recorded dives below 200 m and a maximum depth of 322.8 m.
Diving into the mesopelagic zone below 200 m may serve various ecological functions (e.g. predator avoidance, feeding activity; Braun et al., 2022) and has been observed in other billfish species (i.e., M. nigricans, I. indica, K. audax; Rohner et al., 2022; Andrzejaczek et al., 2023). We found that the depth range of T. belone extended to greater depths during fall and winter, reaching depths > 200 m. This behavior is likely due to the changes in the seasonal thermocline during fall and winter (Houpert et al., 2015). The seasonal variations in the depth of the mixed layer within the Mediterranean Sea significantly influence the vertical distribution of thermoclines, subsequently affecting the spatial distribution of prey species (Sabatés et al., 2009). Mediterranean spearfish, similar to other billfishes, are likely to modulate their vertical movements in response to these fluctuations (Braun et al., 2015), aiming to optimize feeding opportunities within the portion of the water column that is accessible to this species based on its thermal physiology (Arostegui et al., 2019a). These fluctuations in the MED mixed layer depth play a pivotal role in shaping the thermal structure of the water column, thereby potentially influencing the behavior and depth preferences of apex predators as documented elsewhere (Block et al., 2001). While we could not retrieve the thermocline depth and or mixed layers information from our tagging data, we were able to obtain sea surface temperature of the water column < 5m. Sea surface temperature is vital for influencing the mixing layer and thermocline depth (Houpert et al., 2015), and it is crucial during spawning seasons, enabling animals to spawn when the water temperature is optimal (Boyce et al., 2008). Indeed, environmental temperature affects metabolic rate (Fry, 1947; Clarke and Johnston, 1999) which, among other parameters, influences billfish distribution and behavior (e.g., Dale et al., 2022).
During winter months in the MED, heightened wind activity and cooler temperatures lead to increased mixing of the water column and the formation of deeper mixed layers (Heslop et al., 2012). Similarly, our tags recorded SST in winter exhibited a significant region-specific temperature decrease (Table 4). The coolest temperatures were recorded in the Tyrrhenian Sea, followed by the Sicilian Channel and the Aegean Sea. These cooler temperatures are characteristic of winter conditions and are potentially influenced by seasonal currents or regional weather patterns (Zveryaev, 2015). Conversely, in the summer months, the MED experiences shallower mixed layers due to warmer temperatures and reduced wind activity (Heslop et al., 2012). Tag-based SST reached its peak, with the Ionian Sea recording the highest average temperature, likely due to exposure to solar radiation contributing to its warmer temperatures. The Tyrrhenian Sea, although slightly cooler than the Ionian Sea, still maintained warm temperatures conducive to fish activity (Table 4). In response, large pelagic organisms may seasonally shift the distribution of their vertical movements to continually exploit prey associated with this thermal gradient, promoting increased metabolic rates and feeding activity among fish populations. In the fall, the Mediterranean Sea experiences changes in water thermocline and mixing, affecting large pelagic animals. Cooling temperatures and shifting wind patterns lead to a shallowing of the thermocline and increased water column mixing (Houpert et al., 2015). SST exhibited notable variation across different regions. The Aegean Sea and Tyrrhenian Sea recorded moderate temperatures, while the Ionian Sea experienced slightly cooler conditions. On the contrary, the Sicilian Channel exhibited significantly warmer temperatures, likely influenced by local environmental factors. These changes can influence spawning behaviors, migration patterns, and foraging strategies of large pelagic animals as they adapt to shifting environmental conditions in pursuit of prey and suitable spawning grounds.
The sea surface temperatures discussed here closely align with the mean seasonal SST observed in the MED across all regions (Skliris et al., 2011; Shaltout and Omstedt, 2014; Zveryaev, 2015; García-Monteiro et al., 2022). The rising sea surface temperatures in the Mediterranean Sea significantly impact marine ecosystems (Calvo et al., 2011; Lange, 2020), affecting billfish populations (Dell’Apa et al., 2018). Satellite data confirms this rise (Calvo et al., 2011; Lange, 2020), potentially altering billfish distribution, behavior, and health (Alix et al., 2020). This could lead to shifts in habitat range and migratory patterns (Alix et al., 2020). Additionally, higher sea temperatures may decrease billfish growth rates, reproductive success, and increase susceptibility to diseases (Alix et al., 2020). Understanding these impacts is crucial for conserving marine ecosystems, especially for apex predators.
The results of this paper improve the current understanding of the ecology, migratory behavior, and habitat preference of T. belone, as well as help in identifying crucial knowledge gaps that might help understand the distribution of the species within its range of occurrence. In particular, to better understand the migratory patterns of this species it is necessary to increase the tagging efforts, expanding to a broader range of the Mediterranean. In concert with approaches from other disciplines, such as genetics, additional tagging data would help resolve whether there is a single panmictic stock or multiple differentiated stocks within the basin, which is paramount to ensuring appropriate management strategies for this fish resource. Furthermore, additional research must also be conducted to collect yearly/seasonal catch data (landings by country, season, size composition) as well as life history information that are key factors for future stock assessments of this fishery resource.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was approved by SZN animal welfare body “case 04/2020/ec AWB-SZN -27 June 2020 – ricerca in natura fuori applicazione DLgs 26/2014”. The study was conducted in accordance with the local legislation and institutional requirements.
DM: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Validation, Visualization, Writing – original draft, Writing – review & editing. PB: Conceptualization, Investigation, Methodology, Supervision, Writing – original draft, Writing – review & editing. MA: Formal Analysis, Writing – original draft, Writing – review & editing, Data curation. JD: Writing – original draft, Writing – review & editing. BB: Writing – original draft, Writing – review & editing. CB: Formal Analysis, Writing – original draft, Writing – review & editing. MB: Writing – original draft, Writing – review & editing, Investigation. SG: Funding acquisition, Writing – original draft, Writing – review & editing, Supervision. TR: Conceptualization, Funding acquisition, Supervision, Writing – original draft, Writing – review & editing, Project administration.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. Funding has been provided by the Sicilian Department of Mediterranean Fisheries (PO FEAMP 2014-2020 funds; Measure 1.40, letter c; CUP G65C18000020009, project code 02/RBC/18). MA was supported by the Postdoctoral Scholar Program at Woods Hole Oceanographic Institution with funding provided by the Dr. George D. Grice Postdoctoral Scholarship Fund, as well as the WHOI President’s Innovation Fund.
The scientific collaboration for the Mediterranean Spearfish Program was developed under the umbrella of the Memorandum of Understanding between Stazione Zoologica Anton Dohrn (SZN), The International Game Fish Association (IGFA), Sea Life Care International (SLCI) and Stanford University (SUN). The SZN tagging operation were conducted according to the SZN animal welfare body “case 04/2020/ec AWB-SZN -27 June 2020 – ricerca in natura fuori applicazione DLgs 26/2014”, as well as by the International Game Fish Association (IGFA) and Sea Life Care International (SLCI), off Sardinia as part of the collaboration and the IGFA Great Marlin Race (IGMR) citizen science program (https://www.igfa.org) aiming to aid billfish conservation. We thank Bill François (École Normale Supérieure) and Vince Riera (Fishing in Mallorca, https://www.fishing-mallorca.net/en/) for sharing sightings information on T. belone in the western MED. We thank Jason Schratwieser and Bruce Pohlot from IGFA for reviewing the manuscript prior to submission as well as facilitating the tag orders and delivery. We also thank the excellent crew of “Aquila di mare” and “Antonio Padre” for assistance with the field work. A special thanks goes to Sandro Onofaro, President of Yacht Club Porto Rotondo’s, YCPR Fishing Division’s members in Sardinia, Carlo Sabatini (MY Briciola), Cristiano Berera (MY Nibbio), Stephen Palmer and Capt. Sergio Asara (MY Spicy Tuna), for their assistance off Sardinian coast, Michel Marchandise and Capt. Pascal Louis (MY Moana) for their assistance in San Vito lo Capo (TP).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1362169/full#supplementary-material
Akyol O. (2020). Additional record of Mediterranean spearfish, Tetrapturus belone Rafinesque 1810 (Istiophoridae) from Izmir Bay (northern Aegean Sea). COMU. J. Mar. Sci. Fish. 3, 146–148. doi: 10.46384/jmsf.814214
Alix M., Kjesbu O. S., Anderson K. C. (2020). From gametogenesis to spawning: How climate-driven warming affects teleost reproductive biology. J. Fish. Biol. 97, 607–632. doi: 10.1111/jfb.14439
Andrzejaczek S., Mikles C. S., Dale J. J., Castleton M., Block B. A. (2023). Seasonal and diel habitat use of blue marlin Makaira nigricans in the North Atlantic Ocean. ICES. J. Appl. Ichthyol. 80, (4), 1002–1015 doi: 10.1093/icesjms/fsad020
Arostegui M. C., Braun C. D., Gaube P. (2019a). Movement and thermal niche of the first satellite-tagged Mediterranean spearfish (Tetrapturus belone). Fish. Oceanogr. 28, 327–333. doi: 10.1111/fog.12413
Arostegui M. C., Gaube P., Bowman M., Nakamaru K., Braun C. D. (2024). Fishery-independent and -dependent movement data aid in defining the stock structure of a data-deficient billfish. Fish. Res. 271, 106923. doi: 10.1016/j.fishres.2023.106923
Arostegui M. C., Gaube P., Braun C. D. (2019b). Movement ecology and stenothermy of satellite-tagged shortbill spearfish (Tetrapturus angustirostris). Fish. Res. 215, 21–26. doi: 10.1016/j.fishres.2019.03.005
Bariche M., Fricke R. (2020). The marine ichthyofauna of Lebanon: an annotated checklist, history, biogeography, and conservation status. Zootaxa 4775, 1–157. doi: 10.11646/zootaxa.4775.1.1
Battaglia P., Perzia P., Pedà C., Esposito V., Consoli P., Andaloro F., et al. (2018). Evolution, crisis and new scenarios of the Italian swordfish harpoon fishery. Reg. Stud. Mar. Sci. 21, 94–101. doi: 10.1016/j.rsma.2017.09.006
Bauer D. F. (1972). Constructing confidence sets using rank statistics. J. Am. Stat. Assoc. 67, 687–690. doi: 10.1080/01621459.1972.10481279
Bauer R. (2018) Rchivaltag: Analyzing archival tagging data. R package. Available online at: https://CRAN.R-project.org/package=RchivalTag.
Becker J. J., Sandwell D. T., Smith W. H. F., Braud J., Binder B., Depner J., et al. (2009). Global bathymetry and elevation data at 30 arc seconds resolution: SRTM30_PLUS. Mar. Geod. 32, 355–371. doi: 10.1080/01490410903297766
Bleck R. (2002). An oceanic general circulation model framed in hybrid isopycnic-Cartesian coordinates. Ocean. Model. 4, 55–88. doi: 10.1016/S1463-5003(01)00012-9
Block B. A., Dewar H., Blackwell S. B., Williams T. D., Prince E. D., Farwell C. J., et al. (2001). Migratory movements, depth preferences, and thermal biology of Atlantic bluefin tuna. Science 293, 1310–1314. doi: 10.1126/science.1061197
Boyce D. G., Tittensor D. P., Worm B. (2008). Effects of temperature on global patterns of tuna and billfish richness. Mar. Ecol. Prog. Ser. 355, 267–276. doi: 10.3354/meps07237
Braun C. D., Arostegui M. C., Thorrold S. R., Papastamatiou Y. P., Gaube P., Fontes J., et al. (2022). The functional and ecological significance of deep diving by large marine predators. Ann. Rev. Mar. Sci. 14, 129–159. doi: 10.1146/annurev-marine-032521-103517
Braun C. D., Galuardi B., Thorrold S. R. (2018). HMMoce: An R package for improved geolocation of archival-tagged fishes using a hidden Markov method. Methods Ecol. Evol. 9, 1212–1220. doi: 10.1111/2041-210X.12959
Braun C. D., Kaplan M. B., Horodysky A. Z., Llopiz J. K. (2015). Satellite telemetry reveals physical processes driving billfish behavior. Anim. Biotelemetry. 3, 2. doi: 10.1186/s40317-014-0020-9
Byrd R. H., Lu P., Nocedal J., Zhu C. (1995). A limited memory algorithm for bound constrained optimization. SIAM. J. Sci. Comput. 16, 1190–1208. doi: 10.1137/0916069
Calvo E., Simó R., Coma R., Ribes M., Pascual J., Sabatés A., et al. (2011). Effects of climate change on Mediterranean marine ecosystems: the case of the Catalan Sea. Clim. Res. 50, 1–29. doi: 10.3354/cr01040
Cardona L., Álvarez de Quevedo I., Borrell A., Aguilar A. (2012). Massive consumption of gelatinous plankton by mediterranean apex predators. PloS One 7, e31329. doi: 10.1371/journal.pone.0031329
Chassignet E. P., Hurlburt H. E., Smedstad O. M., Halliwell G. R., Hogan P. J., Wallcraft A. J., et al. (2007). The HYCOM (HYbrid Coordinate Ocean Model) data assimilative system. J. Mar. Syst. 65, 60–83. doi: 10.1016/j.jmarsys.2005.09.016
Clarke A., Johnston N. M. (1999). Scaling of metabolic rate with body mass and temperature in teleost fish. J. Anim. Ecol. 68, 893–905. doi: 10.1046/j.1365-2656.1999.00337.x
Collette B. B., Carpenter K., Polidoro B., Juan-Jordá M., Boustany A., Die D., et al. (2011). High value and long life—double jeopardy for tunas and billfishes. Science 333, 291–292. doi: 10.1126/science.1208730
Collette B. B., Di Natale A., Graves J., Pohlot B., Schratwieser J. (2023). Tetrapturus belone. The IUCN red List of Threatened Species 2023. e.T170334A204737590. doi: 10.2305/IUCN.UK.2023-1.RLTS.T170334A204737590.en
Collette B. B., Graves J. (2019). Tunas and Billfishes of the World (Baltimore, Maryland: Johns Hopkins University Press).
Conover W., Iman R. (1979). “On multiple-comparisons procedures: Oak Ridge, TN, USA: Informal Report; Office of Scientific and Technical Information (OSTI)
Dale J. J., Brodie S., Carlisle A. B., Castleton M., Hazen E. L., Bograd S. J., et al. (2022). Global habitat loss of a highly migratory predator, the blue marlin (Makaira nigricans). Divers. Distrib. 28, 2020–2034. doi: 10.1111/ddi.13606
Dell’Apa A., Carney K., Davenport T. M., Carle M. V. (2018). Potential medium-term impacts of climate change on tuna and billfish in the Gulf of Mexico: A qualitative framework for management and conservation. Mar. Environ. Res. 141, 1–11. doi: 10.1016/j.marenvres.2018.07.017
De Sylva D. (1975). “Synopsis of biological data on the Mediterranean Spearfish Tetrapturus belone, Rafinesque,” in Proceedings of the International Billfish Symposium Part 3. Species Synopses. Eds. Shomura R. S., Williams F. (NOAA Technical Report NMFS SSRF-675, Kailua-Kona, Hawaii), 121–131.
Di Natale A. (2012). I grandi pelagici: stato delle risorse. In Rapporto annuale sullo stato delle risorse biologiche nei mari circostanti l’Italia: ed. A. Mannini and G. Relini, SIBM, MIPAAF. Biol. Mar. Medit. 19, 207–227.
Di Natale A., Addis P., Cau A., Garibaldi F., Piccinetti C., Orsi Relini L. (2011). “The state of large pelagics,” in The state of Italian marine fisheries and aquaculture. Eds. Cataudella S., Spagnolo M. (MIPAAF, Roma), 137–147.
Di Natale A., Cañadas A., Vázquez-Bonales J. A., Tensek S., Pagá-García A. (2018). Report of the ICCAT GBYP aerial survey for Bluefin spawning aggregation in 2017. ICCAT. ICCAT. Coll.Vol.Sci.Pap. 74, 3172–3204.
Di Natale A., Celona A., Mangano A. (2005). A series of catch records by the harpoon fishery in the Strait of Messina from 1976 to 2003. ICCAT. Coll.Vol.Sci.Pap. 58, 1348–1359.
Ditton R., Stoll J. (2003). Social and economic perspective on recreational billfish fisheries. Mar. Freshw. Res. 54, 545–554. doi: 10.1071/MF01279
Domeier M. L. (2006). An analysis of Pacific striped marlin (Tetrapturus audax) horizontal movement patterns using pop-up satellite archival tags. Bull. Mar. Sci. 79, 811–825.
Dulcic J., Soldo A. (2004). The Mediterranean spearfish, Tetrapturus belone Rafinesque 1810, in the Adriatic waters: new records and a review of Adriatic records. Ann. Ser. Hist. Nat. 4, 45–48.
García-Monteiro S., Sobrino J. A., Julien Y., Sòria G., Skokovic D. (2022). Surface Temperature trends in the Mediterranean Sea from MODIS data during years 2003–2019. Reg. Stud. Mar. Sci. 49, 102086. doi: 10.1016/j.rsma.2021.102086
Gerovasileiou V., Akyol O., Al-Hosne Z., Alshikh Rasheed R., AtaÇ E., Bello G., et al. (2020). New records of rare species in the Mediterranean Sea (May 2020). Mediterr. Mar. Sci. 21, 340–359. doi: 10.12681/mms.22148
Heslop E. E., Ruiz S., Allen J., López-Jurado J. L., Renault L., Tintoré J. (2012). Autonomous underwater gliders monitoring variability at “choke points” in our ocean system: A case study in the Western Mediterranean Sea. Geophys. Res. Lett. 39, L20604. doi: 10.1029/2012GL053717
Holland S. M., Ditton R. B., Graefe A. R. (1998). An ecotourism perspective on billfish fisheries. J. Sustain. Tour. 6, 97–116. doi: 10.1080/09669589808667305
Hollander M., Wolfe D. A., Chicken E. (2014). Nonparametric Statistical Methods. 3rd Edition (New York: John Wiley & Sons Inc).
Hoolihan J. P., Luo J., Snodgrass D., Orbesen E. S., Barse A. M., Prince E. D. (2015). Vertical and horizontal habitat use by white marlin Kajikia albida (Poey 1860) in the western North Atlantic Ocean. ICES. J. Mar. Sci. 72, 2364–2373. doi: 10.1093/icesjms/fsv082
Houpert L., Testor P., Durrieu de Madron X., Somot S., D’Ortenzio F., Estournel C., et al. (2015). Seasonal cycle of the mixed layer, the seasonal thermocline and the upper-ocean heat storage rate in the Mediterranean Sea derived from observations. Prog. Oceanogr. 132, 333–352. doi: 10.1016/j.pocean.2014.11.004
Istituto Idrografico della Marina (1982). Atlante delle correnti superficiali dei mari italiani (Genova: Istituto Idrografico della Marina).
Kerstetter D. W., Orbesen E. S., Snodgrass D., Prince E. D. (2009). Movements and habitat utilization of two longbill spearfish Tetrapturus pfluegeri in the eastern tropical South Atlantic Ocean. Bull. Mar. Sci. 85, 173–182.
Kruskal W. H., Wallis W. A. (1952). Use of ranks in one-criterion variance analysis. J. Am. Stat. Assoc. 47, 583–621. doi: 10.2307/2280779
Lange M. (2020). “Climate change in the mediterranean: environmental impacts and extreme events,” in European Institute of the Mediterranean Yearbook 2020 (IEMed, Barcelona), 30–45.
Luo J., Ault J. S., Shay L. K., Hoolihan J. P., Prince E. D., Brown C. A., et al. (2015). Ocean heat content reveals secrets of fish migrations. PloS One 10, e0141101. doi: 10.1371/journal.pone.0141101
Malara D., Battaglia P., Consoli P., Arcadi E., Canese S., Greco S., et al. (2020). Evidence of a predation event on a tagged Mediterranean spearfish (Tetrapturus belone; Pisces, Istiophoridae), inferred from pop-up satellite tagging data. Aquat. Living. Resour. 33, 23. doi: 10.1051/alr/2020024
Mosetti F. (1991). “Tidal and other currents in the Straits of Messina, The Straits of Messina Ecosystem,” in Proceedings of the Symposium held in Messina 4–6 April (Università degli Studi di Messina, Messina).
Nakamura I. (1985). FAO species catalogue. Vol. 5 Billfishes of the World. An annotated and illustrated catalogue of marlins, sailfishes, spearfishes and swordfishes known to date. FAO Fisheries. Synopsis. 5, 1–65.
NASA/JPL (2015) GHRSST Level 4 MUR Global Foundation Sea Surface Temperature Analysis (v4.1). Available online at: http://podaac.jpl.nasa.gov/dataset/MUR-JPL-L4-GLOB-v4.1.
Nielsen J. K., Bryan D. R., Rand K. M., Arostegui M. C., Braun C. D., Galuardi B., et al. (2023). Geolocation of a demersal fish (Pacific cod) in a high-latitude island chain (Aleutian Islands, Alaska). Anim. Biotelemetry. 11, 29. doi: 10.1186/s40317-023-00340-3
Nishida T., Tsuji S., Segawa K. (1998). Spatial data analyses of Atlantic bluefin tuna larval surveys in the 1994 ICCAT BYP. ICCAT. Coll.Vol.Sci.Pap. 48, 107–110.
Olita A., Ribotti A., Fazioli L., Perilli A., Sorgente R. (2013). Surface circulation and upwelling in the Sardinia Sea: A numerical study. Cont. Shelf. Res. 71, 95–108. doi: 10.1016/j.csr.2013.10.011
Pohlot B. G., Ehrhardt N. (2017). An analysis of sailfish daily activity in the Eastern Pacific Ocean using satellite tagging and recreational fisheries data. ICES. J. Mar. Sci. 75, 871–879. doi: 10.1093/icesjms/fsx082
Pons M., Branch T. A., Melnychuk M. C., Jensen O. P., Brodziak J., Fromentin J. M., et al. (2017). Effects of biological, economic and management factors on tuna and billfish stock status. Fish. Fish. 18, 1–21. doi: 10.1111/faf.12163
Posit team (2023). RStudio: integrated development for R (Boston, MA: Posit Software, PBC). Available at: http://www.posit.co.
Potoschi A. (2000). Biological aspects of Tetrapturus belone (Raf. 1810) in the straits of messina. Biol. Mar. Mediterr. 7, 819–824.
Psomadakis P. N., Giustino S., Vacchi M. (2012). Mediterranean fish biodiversity: an updated inventory with focus on the Ligurian and Tyrrhenian seas. Zootaxa 326, 1–46. doi: 10.11646/zootaxa.3263.1.1
Punt A. E., Su N.-J., Sun C.-L. (2015). Assessing billfish stocks: A review of current methods and some future directions. Fish. Res. 166, 103–118. doi: 10.1016/j.fishres.2014.07.016
Ribotti A., Puillat I., Sorgente R., Natale S. (2004). Mesoscale circulation in the surface layer off the southern and western Sardinia Island in 2000–2002. Chem. Ecol. 20, 345–363. doi: 10.1080/02757540410001727963
Robins C. R., de Sylva D. P. (1963). A new western atlantic spearfish, tetrapturus pfluegeri, with a redescription of the mediterranean spearfish tetrapturus belone. Bull. Mar. Sci. 13, 84–122.
Rohner C. A., Bealey R., Fulanda B. M., Everett J. D., Richardson A. J., Pierce S. J. (2021). Movement ecology of black marlin Istiompax indica in the Western Indian Ocean. J. Fish. Biol., 9(3), 1044–1059. doi: 10.1111/jfb.14809
Rohner C. A., Bealey R., Fulanda B. M., Pierce S. J. (2020). Movement and habitat use of striped marlin Kajikia audax in the Western Indian Ocean. J. Fish. Biol. 97, 1415–1427. doi: 10.1111/jfb.14508
Rohner C. A., Bealey R., Fulanda B. M., Prebble C. E. M., Williams S. M., Pierce S. J. (2022). Vertical habitat use by black and striped marlin in the Western Indian Ocean. Mar. Ecol. Prog. Ser. 690, 165–183. doi: 10.3354/meps14043
Romeo T., Battaglia P., Pedà C., Perzia P., Consoli P., Esposito V., et al. (2012). Pelagic cephalopods of the central Mediterranean Sea determined by the analysis of the stomach content of large fish predators. Helgol. Mar. Res. 66, 295–306. doi: 10.1007/s10152-011-0270-3
Romeo T., Battaglia P., Raicevich S., Perzia P., Andaloro F. (2015). Swordfish harpoon fishery in the mediterranean sea: Recent data to implement the marine strategy framework directive and the ecap (ecosystem approach) process. Fish. Res. 161, 191–199. doi: 10.1016/j.fishres.2014.07.013
Romeo T., Consoli P., Castriota L., Andaloro F. (2009a). An evaluation of resource partitioning between two billfish, Tetrapturus belone and Xiphias gladius, in the central Mediterranean Sea. J. Mar. Biol. Assoc. U. K. 89, 849–857. doi: 10.1017/S0025315408002087
Romeo T., Consoli P., Greco S., Canese S., Andaloro F. (2009b). Swordfish (Xiphias gladius, Teleostea: Xiphiidae) surface behavior during reproductive period in the central Mediterranean Sea (southern Tyrrhenian Sea). Mar. Biodivers. Rec. 2, e45. doi: 10.1017/s1755267209000578
Saad A., Khrema L., Alnesser A., Barakat I., Capapé C. (2024). First substantiated record of Mediterranean spearfish Tetrapturus belone (istiophoridae) from the Syrian coast. Thalass. Salentina. 45, 11–16. doi: 10.1285/i15910725v45p11
Sabatés A., Salat J., Palomera I., Raya V. (2009). Physical and biological processes controlling the distribution of fish larvae in the NW Mediterranean. Prog. Oceanogr. 74, 355–376. doi: 10.1016/j.pocean.2007.04.017
Shaltout M., Omstedt A. (2014). Recent sea surface temperature trends and future scenarios for the Mediterranean Sea. Oceanologia 56, 411–443. doi: 10.5697/oc.56-3.411
Skliris N., Sofianos S. S., Gkanasos A., Axaopoulos P., Mantziafou A., Vervatis V. (2011). Long-term sea surface temperature variability in the Aegean Sea. Adv. Oceanogr. Limnol. 2, 125–139. doi: 10.1080/19475721.2011.601325
Spanò N., De Domenico E. (2017). “Biodiversity in Central Mediterranean Sea,” in Mediterranean Identities - Environment, Society, Culture. Ed. Fuerst-Bjelis B. (IntechOpen, Rijeka, Crozia), 129–148.
Spartà A. (1960). Biologia e pesca di Tetrapterus belone e sue forme postlarvali. Boll. Pesca. Pisc. Idrobiol. 15, 20–24.
Sulli A., Agate M., Zizzo E., Gasparo Morticelli M., Lo Iacono C. (2021). Geo-hazards of the San Vito peninsula offshore (southwestern Tyrrhenian Sea). J. Maps. 17, 185–196. doi: 10.1080/17445647.2020.1866703
Thieurmel B., Elmarhraoui A. (2019) Suncalc: Compute sun position, sunlight phases, moon position and lunar phase. Available online at: https://CRAN.R-project.org/package=suncalc.
Vaudo J. J., Byrne M. E., Wetherbee B. M., Harvey G. M., Mendillo A. Jr, Shivji M. S. (2017). Horizontal and vertical movements of white marlin, Kajikia albida, tagged off the Yucatán Peninsula. ICES. J. Mar. Sci. 75, 844–857. doi: 10.1093/icesjms/fsx176
Wagenmakers E.-J., Farrell S. (2004). AIC model selection using Akaike weights. Psychon. Bull. Rev. 11, 192–196. doi: 10.3758/BF03206482
Wang H., You Z., Guo H., Zhang W., Xu P., Ren K. (2023). Quality assessment of sea surface salinity from multiple ocean reanalysis products. J. Mar. Sci. Eng. 11, 54. doi: 10.3390/jmse11010054
Keywords: Pelagic predator, PSAT, Temperature niche, Mediterranean Sea, Billfish
Citation: Malara D, Battaglia P, Arostegui MC, Dale JJ, Block BA, Braun CD, Brogna M, Greco S and Romeo T (2024) Satellite tagging insights into the seasonal movements and behavior of Mediterranean spearfish (Tetrapturus belone, Istiophoridae). Front. Mar. Sci. 11:1362169. doi: 10.3389/fmars.2024.1362169
Received: 27 December 2023; Accepted: 19 April 2024;
Published: 03 May 2024.
Edited by:
Fausto Tinti, University of Bologna, ItalyReviewed by:
Antonio Di Natale, Fondazione Acquario di Genova Onlus, ItalyCopyright © 2024 Malara, Battaglia, Arostegui, Dale, Block, Braun, Brogna, Greco and Romeo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Danilo Malara, ZGFuaWxvLm1hbGFyYUBzem4uaXQ=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.