 Diego Cepeda
Diego Cepeda Nuria Sánchez
Nuria Sánchez Adriana Spedicato4
Adriana Spedicato4 Emma Michaud
Emma Michaud Daniela Zeppilli
Daniela Zeppilli- 1Univ. Brest, CNRS, Ifremer, UMR6197 Biologie et Ecologie des Ecosystèmes marins Profonds, Plouzané, France
- 2Autonomous University of Madrid (UAM), Faculty of Sciences, Department of Biology, Centre for Research on Biodiversity and Global Change (CIBC-UAM), Madrid, Spain
- 3Department of Biodiversity, Ecology and Evolution (BEE), Faculty of Biological Sciences, Complutense University of Madrid (UCM), Madrid, Spain
- 4Univ. Brest, CNRS, IRD, IFREMER, LEMAR, Plouzané, France
Meiofaunal communities are dominated by free-living nematodes and harpacticoid copepods. Most of the studies defining the environmental drivers that influence meiofauna focus on these profuse groups. The ‘rare’ meiofaunal organisms (densities <5%) are frequently overlooked, making it difficult to know which abiotic variables influence their populations, or their ecological role in the benthic ecosystems. In this study, we explore the ecological factors that characterize a low human-polluted mangrove in French Guiana (western Atlantic Ocean) along an urban-to-natural gradient, focusing on the ‘rare’ phylum Kinorhyncha. This taxon seems to be mainly influenced by sediment size, being more abundant in silty sediments. The presence of microorganisms also enhances the community, likely due to a higher food availability. Oppositely, some trace metals of both lithogenic and anthropogenic origin, as well as predominance of coarser sediment particles, seem to negatively affect their populations. The importance of including the lesser abundant groups of meiofauna in ecological studies is evident, since it allows to have a better idea of the ecological state of marine benthic environments.
1 Introduction
Meiofauna includes a diverse assemblage of microscopic benthic invertebrates usually smaller than 1 mm and larger than 0.06 mm (Giere, 2009). Free-living nematodes and harpacticoid copepods frequently dominate the meiofaunal communities (James and Mark, 2004; Giere, 2009; Punniyamoorthy et al., 2021), consequently drawing the attention of the scientific community. Several other meiofaunal taxa are frequently overlooked, mainly due to a lack of experts (Balsamo et al., 2020). This is the case of Kinorhyncha, a commonly neglected phylum in ecological studies of marine meiofauna due to its low abundance (Hodda and Nicholas, 1986; Schrijvers et al., 1995; Bianchelli et al., 2010; Della Patrona et al., 2016). However, some studies showed these animals can become dominant in specific habitats, including some riverine and fringe mangroves (Sarma and Wilsanand, 1994; Gomes et al., 2002; Santos et al., 2009; Ostmann et al., 2012; Annapurna et al., 2015; Cepeda et al., 2022b). In addition, low abundant taxa have shown to be useful when characterizing meiofaunal assemblages in certain areas or when assessing meiofauna sensitivity to pollutants (Vanreusel, 1990; Giere, 2009; Sevastou et al., 2013; Sánchez and Gómez-León, 2024). Currently, 42 species of Kinorhyncha are known to inhabit mangroves (Cepeda et al., 2022a, b; Sørensen, 2023). Some of these species belong to the Echinoderes coulli-group, characterized by an enlarged nephridial sieve plate likely related to cope with fluctuating salinities, tolerating both brackish and hypersaline conditions (Ostmann et al., 2012). Biological data on kinorhynchs is rather scarce, but it is hypothesized that species living in shallow waters exhibit patched, reduced, natural distributions due to their limited mobility through the sediment particles and the lack of dispersal larva, together with their preference for specific abiotic conditions of the sediment. Furthermore, kinorhynchs are preys for macrofauna, including some polychaetes, bivalves, decapods and mysidaceans (Neuhaus, 2013). What the kinorhynchs themselves feed on, however, is still a mystery.
Still, little is known about the main abiotic factors that model Kinorhyncha communities. Some studies proved that both kinorhynch species diversity and community composition are correlated to sediment particle features (Landers et al., 2018; Grzelak and Sørensen, 2019; Landers et al., 2019, 2020; Cepeda et al., 2020b; Hoffman et al., 2021). In addition, the organic enrichment may have a variable effect on kinorhynchs by increasing or decreasing density and richness (Ostmann et al., 2012; Dal Zotto et al., 2016; Landers et al., 2020; Cepeda et al., 2020b; Hoffman et al., 2021). Trace metals are not thought to directly influence kinorhynch communities, but positive correlations with density have been found in the Gulf of Mexico (Landers et al., 2018, 2019, 2020; Hoffman et al., 2021). In CO2 vents and deep-sea pockmarks, Kinorhyncha community composition is mainly driven by the accumulation of reduced compounds and pH (Álvarez-Castillo et al., 2015; Sánchez et al., 2020; Cepeda et al., 2020a).
Michelet et al. (2021) conducted a study on meiofauna (focused on nematodes) sensitivity to low human impact in a French Guiana mangrove (western Atlantic Ocean), showing that nematodes might be affected by low contaminant concentrations. These preliminary results also shown that Kinorhyncha reached ca. 5% of the overall meiofauna abundance in some stations, which is in the range of values reported from extreme environments (Zeppilli et al., 2018). In the present study, we analyze the relationships between the Kinorhyncha community and different abiotic factors in the same mangrove. Our main goals are: i) to identify the most significant abiotic parameters influencing the Kinorhyncha community, and ii) to define the role that this ‘rare’ group can have in the ecological characterization of a meiofaunal community in mangrove ecosystems.
2 Materials and methods
2.1 Sampling site, extraction of meiofauna and environment characterization
The present work is based on sediment samples previously analyzed by Michelet et al. (2021); Cepeda et al. (2022a) and Fiard et al. (2022). The mangrove study area is located at the Cayenne Estuary (French Guiana, western Atlantic Ocean), near to the city of Cayenne (ca. 66,000 inhabitants) (INSEE, 2023). Sampling was conducted from November 19th to 21st, 2017, in three sampling stations (from now on: S1, S2 and S3) following an urban-to-natural gradient. The sampling points were selected in the polyhaline area of the estuary to exclude salinity differences (Michelet et al., 2021) at 0 m depth during low tide. S1 is situated at the Crique Fouillé Tributary (04°54′53.208″ N, 52°20′15.9324″ W) and it is the most impacted by sewage discharges, having a higher proportion of sand, pollutants (both metals and organic contaminants) and total organic carbon. S2, located near the confluence of the Cayenne and Montsinéry Rivers (04°53′49.2288″ N, 52°22′27.714″ W), is considered as moderate to lowly influenced by pollution as it receives some water from the suburban Cayenne drainage basin. This station has a higher content in silt and clay, pigments and organic pollutants, as well as reducing conditions. S3 (reference station), located ca. 14 km upstream of the Cayenne River (04°51′31.9716″ N, 52°23′59.5248″ W) constitutes a natural area far from human influence. This station is characterized by higher proportions of silt and clay and oxidizing conditions, although Fiard et al. (2022) found relatively high concentrations of dieldrin and naphthalene in the deepest layers of the sediment. Detailed maps of the location of the sampling stations can be found in Michelet et al. (2021) and Cepeda et al. (2022a).
Three sediment cores (from now on: A, B and C) spaced by 2 m, made of Plexiglas® tubes of 10.4 cm internal diameter and 20 cm height, were collected from each station. The sediment column of each core was sliced horizontally in three depth layers (from now on: L1, L2 and L3) as follows: L1, 0–2 cm; L2, 2–10 cm; L3, 10–16 cm. Two subsamples of 1.77 cm2 (total of 3.54 cm2) were taken in each sediment layer for meiofauna study, being fixed in 4% formalin buffered with sodium bicarbonate. Subsamples for particle size analyses were kept at 4°C, whereas subsamples for biogeochemistry analyses were frozen at -80°C.
Detailed information about extraction of meiofauna and study of kinorhynchs may be found in Michelet et al. (2021) and Cepeda et al. (2022a), respectively. Detailed information on methodology concerning environment characterization and biogeochemical analyses is in Michelet et al. (2021) and Fiard et al. (2022). The abiotic variables that we used in this study were: physicochemical parameters, organic matter, prokaryotic community, pigments, metals and metalloids, and organic contaminants. Information about these variables may be found in the initial dataset (Michelet et al., 2021, doi: 10.5281/zenodo.4592299). Ordination analyses grouped cores and sediment layers of the same station together, which determines there is a noticeable change in the environment between the sampling stations. These differences were mainly determined by sediment granulometry, organic matter, accumulation of pigments, redox potential and presence of certain metals and organic pollutants (Michelet et al., 2021).
2.2 Statistics
The dataset of the present study was built following the sampling strategy, which included three sampling stations (S1, S2 and S3), three cores per station (A, B and C) and three sediment layers per core (L1, L2 and L3). For each sediment layer, the Kinorhyncha density (ind/10 cm2) was used as the response variable. All the statistical analyses were run on R v.4.1.2 environment (R Core Team, 2022).
Kruskal-Wallis (KW) analyses were performed to assess differences in Kinorhyncha density. KWs were used because normality and homoscedasticity of the variables were violated. Two different approaches were considered: vertical profile analysis (throughout the sediment depth profile, split in three layers) and interstation analysis (among the three sampling stations following an urban-to-natural gradient). Wilcoxon tests for pairwise comparisons were carried out after KWs.
To test the relationship between these variables over kinorhynch density, linear correlation coefficients using the non-parametric Spearman coefficient ρ (as data normality was not verified) were calculated. Moreover, a redundancy analysis (RDA) was performed with the package ‘vegan’ (Oksanen et al., 2022) to extract and summarize the variation of the Kinorhyncha density due to the environmental variables between the sampled stations, considering that complex and non-linear interactions between multiple environmental variables may act over the Kinorhyncha community. Explanatory variables were centered, standardized, and normalized, whereas community data was Hellinger-transformed; multicollinearity of explanatory variables was tested calculating the variance inflation factor, which reduced the included variables in the RDA to percentage of sand, redox potential, total organic carbon, total prokaryotic DNA, chlorophyll a, bismuth, cadmium, lead, manganese, mercury, polychlorinated biphenyls, and pesticides. A permutational multivariate analysis of variance (PERMANOVA) was subsequently conducted to determine significant differences in the Kinorhyncha density according to the analyzed variables.
3 Results
3.1 Characterization of the Kinorhyncha community
A total of 379 kinorhynch specimens belonging to two species, Echinoderes angelae Cepeda et al., 2022a and E. guianensis Cepeda et al., 2022b, were collected. Most of the specimens corresponded to E. angelae, whereas only four representatives of E. guianensis were found at S2. Animal densities ranged 0–353 ind/10cm2, with mean of 44 ind/10cm2. S2 showed the highest values, closely followed by S3. The three studied stations potentially differ in density of Kinorhyncha, as marginally significant differences were found (p=0.05). The Wilcoxon test found differences in density between S1 and the remaining stations (Table 1, Figure 1B).
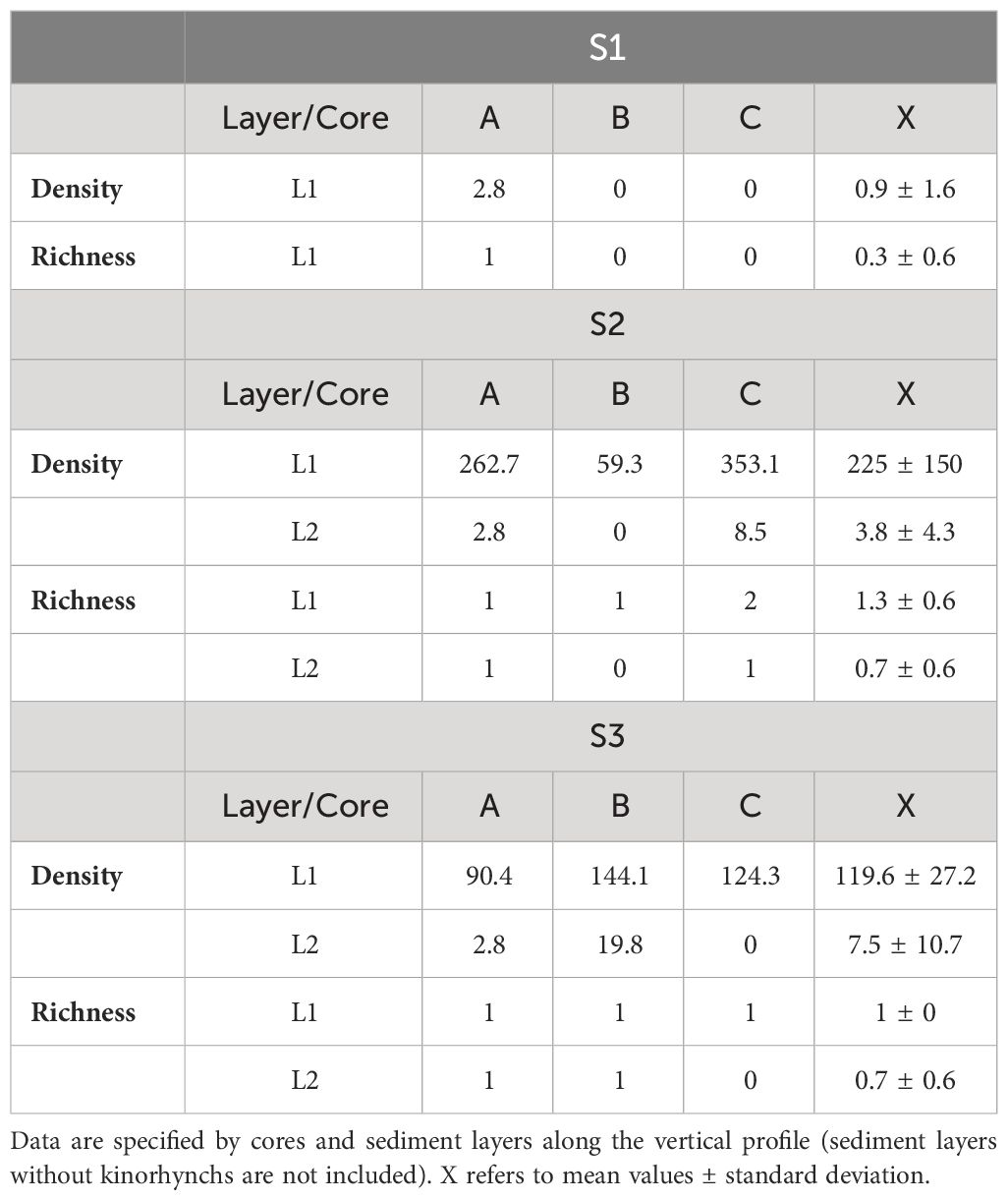
Table 1 Raw, mean values and standard deviation of density (ind/10cm2) and richness (number of species) of Kinorhyncha in the sampling sites.
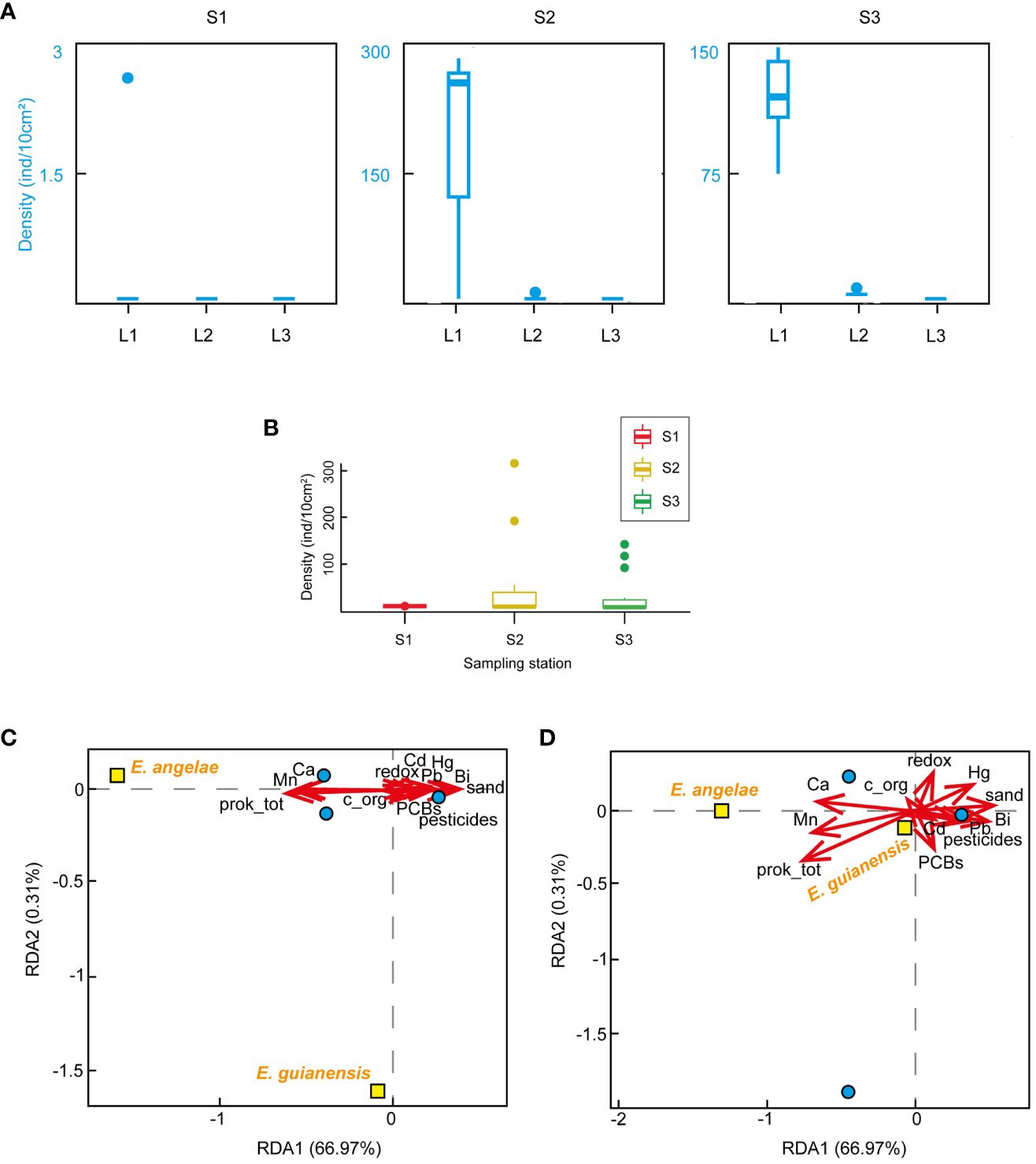
Figure 1 (A) Kinorhyncha density (ind/10cm2) along the vertical profile (i.e., sediment depth) of the three sampling stations. (B) Interstation analysis of Kinorhyncha density (ind/10cm2) including values of the three sampling stations. (C) RDA triplot scaling 1, showing similarities between objects in the response matrix. (D) RDA triplot scaling 2, showing the effects of environmental variables. Boxplots represent the median value (horizontal, thicker line within the box), the distributions of 50% of the data (the box), and the highest and lowest values within 95% of the distribution (the whisker). In the RDA graphics, blue circles represent sites and yellow squares represent species.
Concerning the vertical profile analysis, maximum values of density were found in L1, subsequently fading towards L3, where kinorhynchs were completely absent (Table 1, Figure 1A). Statistically significant differences in Kinorhyncha density were found along the vertical sediment profile in S2 and S3 (p=0.03), indicating an effect of sediment depth in the number of specimens (Table 1, Figure 1A).
3.2 Effect of environment over Kinorhyncha
Kinorhyncha density correlated positively with silt, clay, total prokaryotic DNA, some pigments (chlorophyll a, β-carotene, pheophytin a, unknown pigments 3 and 4) and manganese, and negatively with sand, bismuth, mercury, and molybdenum (Table 2).
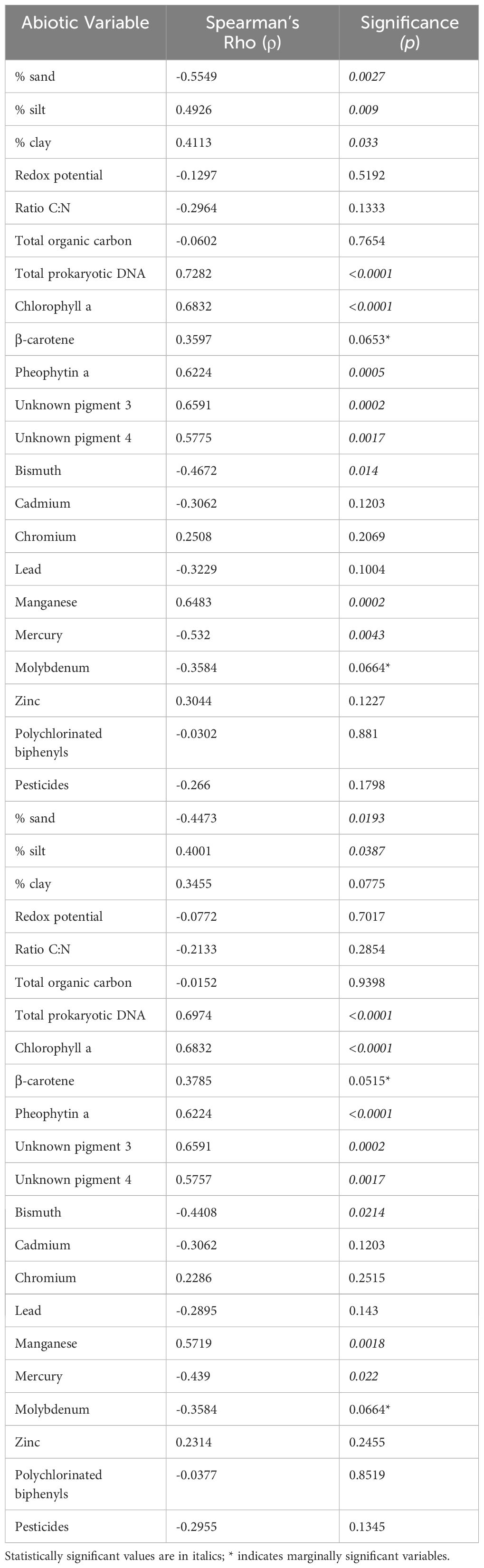
Table 2 Spearman rank correlations between Kinorhyncha density and most relevant abiotic variables.
The environmental variables included in the RDA were significant (F=2.3995, p=0.036) in explaining 67.3% of the variation in Kinorhyncha density across sites (Table 3, Figures 1C, D). Particularly meaningful predictors were the percentage of sand (F=7.0123, p=0.013), total organic carbon (F=7.6543, p=0.005), total prokaryotic DNA (F=7.674, p=0.015), and bismuth (F=3.2599, p=0.07) (Table 3, Figures 1C, D).
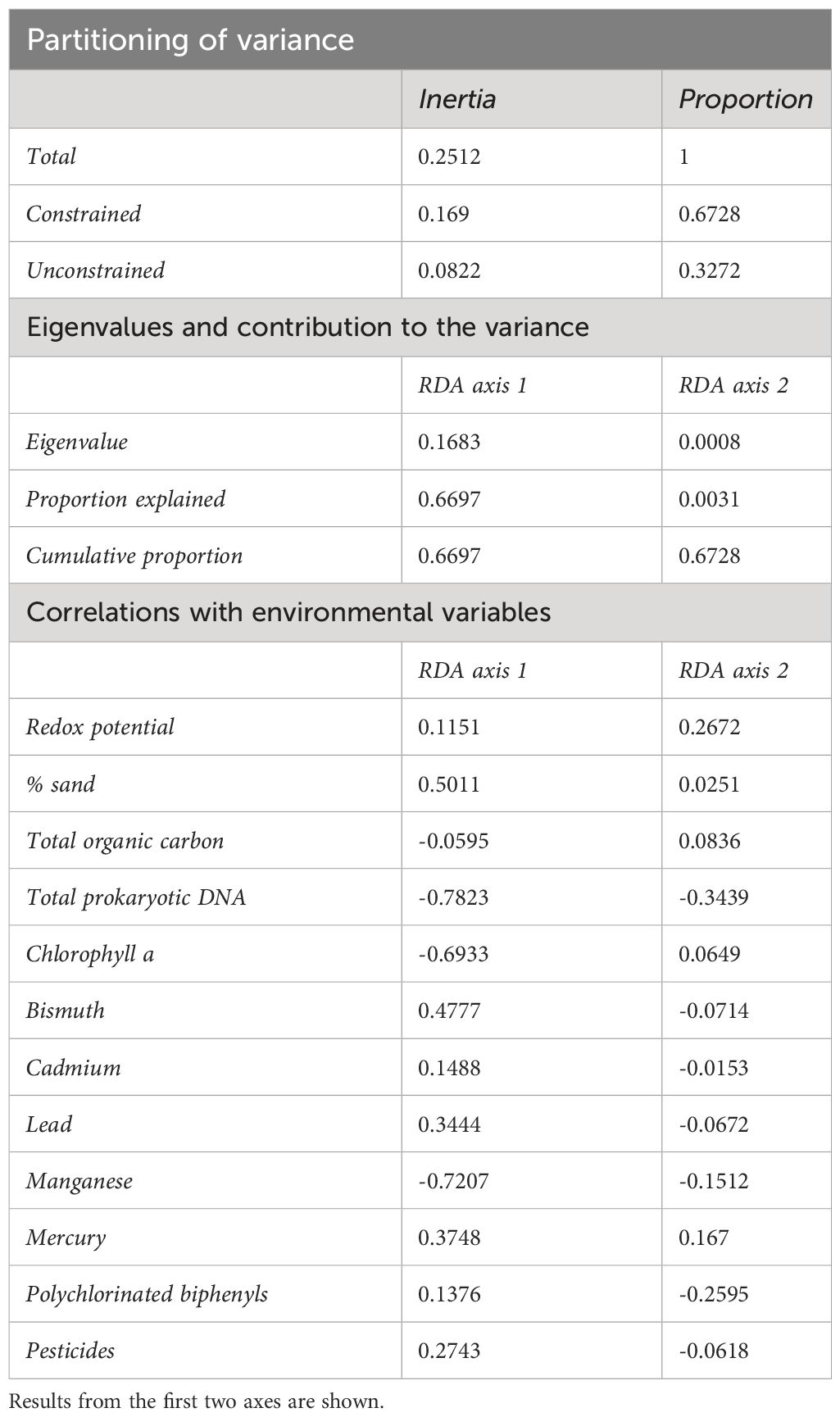
Table 3 Redundancy analysis (RDA) results on relationships between Kinoryncha species density and environmental variables.
4 Discussion
4.1 Kinorhyncha community along the vertical profile
Sediment depth plays a key role when explaining changes in Kinorhyncha community in the studied area. In S2 and S3, density of kinorhynchs significantly decreases along the vertical profile, from values of ca. 60–350ind/10cm2 at the upper layers to the complete absence of kinorhynchs at the deeper layers. Although similar results were not found in S1, the general low number of recovered specimens likely prevented us from finding the same decreasing trend along the vertical profile.
In our study, the majority of the Kinorhyncha community was concentrated in the upper sediment layer (0–2 cm). However, both oxidizing (S1 and S3) and reducing (S2) conditions were found at the upper layer, and this does not seem to affect Kinorhyncha density. We can assume that the species present here have some kind of physiological adaptation that allows them to tolerate variable oxygenation conditions in the marine sediment. However, when the reducing conditions significantly increase from the first two centimeters, Kinorhyncha density abruptly decreases. This result agrees with almost all previous meiobenthic studies (Kotwicki et al., 2005), since vertical distribution of meiofauna is mainly defined by the redox potential discontinuity layer, which marks the boundary between predominantly oxidizing and reducing conditions (Coull, 1988; Gerwing et al., 2013). Even in riverine mangroves, where water level strongly varies daily (in French Guiana, up to 1.5 m) (Service Hydrographique et Océanographique de la Marine, 1975), vertical migration of Kinorhyncha caused by water shortage seems not to be influential, according to this study and previous ones.
4.2 Kinorhyncha community along the urban-to-natural gradient
The application of RDA provides valuable information on how environmental factors affect the distribution and abundance of kinorhynchs in the studied mangrove, helping to identify which environmental variables are most important for community structure. This analysis also considers all environmental variables and the complex (not necessarily linear) relationships they have with our response variable (Kinorhyncha density) simultaneously (Zuur et al., 2007). Thus, we can consider it a powerful tool that allows us to explore and better understand the ecological role of kinorhynchs, in this case in a mangrove swamp.
The biggest differences in Kinorhyncha density were found between S1, which represents the most influenced area of the mangrove by wastewater emissions, and the remaining sampling stations. At S1, a single kinorhynch specimen of Echinoderes angelae was found, whereas the remaining specimens of both E. angelae and E. guianensis were recovered at the other stations, both showing high densities. Thus, the studied urban-to-natural gradient also defines important changes in the Kinorhyncha community, at least between the most polluted area (S1) and the remaining, lowly affected sampling areas (S2 and S3). Kinorhynchs seem to be negatively influenced by the combination of high percentages of sand and accumulation of certain metals (bismuth, mercury, and molybdenum) that characterized S1. Oppositely, silty sediments with a rich prokaryotic community, high amount of organic carbon, prevalence of certain pigments and manganese, promoted higher density of kinorhynchs at S2 and S3.
Kinorhynchs are usually more abundant in silty sediments. This is likely related to their body morphology, dimensions, external morphological features, and locomotion way (Grzelak and Sørensen, 2019). Most kinorhynchs are part of the burrowing meiofauna that move actively through the sediment. In finer sediments, the possession of a fusiform body shape (like that of the two reported species) facilitates the burrowing through the smallest interstices by simple movements of the body (Cepeda et al., 2020b). The results herein obtained coincide with those of other areas (Dal Zotto et al., 2016; Landers et al., 2018; Grzelak and Sørensen, 2019; Landers et al., 2019; Hoffman et al., 2021) reinforcing the fact that fusiform kinorhynchs seem to be mainly present in sediments rich in silt and clay.
The presence of a rich prokaryotic community, together with high concentrations of pigments (which reflects the presence of microphytes and potential food sources) may also enhance Kinorhyncha communities. Kinorhynch density was positively correlated to both chlorophyll a and pheophytin a, a degradation product of chlorophyll a (Michelet et al., 2021), which could indicate a degradation process beneficious for kinorhynchs in terms of source of organic matter. Usually, coarser sediments with larger interstitial spaces host a wider variety of food sources (Vanaverbeke et al., 2011). However, fine sediments may contain enough food sources and oxygen in the first centimeters to allow kinorhynchs flourishing (Nomaki et al., 2008; Dal Zotto et al., 2016). Thus, the specific variety of the nutrients present in fine sediments may benefit somehow the proliferation of, at least, small and fusiform kinorhynchs, such as E. angelae and E. guianensis.
Trace metal contamination has shown to negatively influence Kinorhyncha communities along the Louisiana continental shelf (Landers et al., 2019). However, recent investigations have shown that kinorhynch density increased in sediments where high percentages of silt and clay were associated to trace metals (Landers et al., 2018; Landers et al., 2020; Hoffman et al., 2021). Indeed, sediment grain size and the lithogenic origin of such metals seem to be the most determinant abiotic factors shaping Kinorhyncha communities (Grzelak and Sørensen, 2019; Landers et al., 2020; Cepeda et al., 2020b). Sediment grain size seems to be a much more powerful explanatory variable than trace metals content for kinorhynch communities. Moreover, metals do not often exceed the toxicity threshold of the official guidelines (Michelet et al., 2021), thus their effect may be no detectable. In the studied mangrove, metals and metalloids also have a lithogenic origin with succession of co-precipitation and co-complexification geochemical processes (Marchand et al., 2006), except for bismuth, mercury and molybdenum that had a wastewater origin (Michelet et al., 2021). Indeed, such metals negatively correlated with Kinorhyncha density. Notably, mercury content (0.64 mg/kg) was above the established hazardous concentration for benthic invertebrates (Conder et al., 2015; Michelet et al., 2021), which would explain the negative effect observed in the present study.
5 Conclusion
In the present study, Kinorhyncha from Cayenne Estuary’s mangrove seems to be mainly influenced by sediment grain size, as silt and clay enhance density of these animals. The presence of potential food sources along the first sediment centimeters, where kinorhynchs are most abundant, also would make their population flourish. Even at low concentration, certain trace metals and metalloids may negatively affect the Kinorhyncha community. Eventually, the use of ‘rare’ meiofaunal groups (often present in low abundances), identified at species level, must be coupled with that of other dominant taxa (such as nematodes and copepods). Such combination provides more accurate information to characterize the quality of benthic habitats.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
DC: Formal analysis, Investigation, Methodology, Software, Writing – original draft, Writing – review & editing. NS: Investigation, Writing – review & editing. AS: Investigation, Writing – review & editing. EM: Conceptualization, Funding acquisition, Methodology, Resources, Supervision, Writing – review & editing. DZ: Conceptualization, Funding acquisition, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by Office de l’Eau de la Guyane (OEG) and Office Français de la Biodiversité (OFB) under the framework of the European Water Framework Directive (French working group ‘Mangroves DCE’, composed of G. Dirberg, C. Hubas, D. Lamy, P. Cuny, C. Militon, R. Walcker, F. Fromard, I. Bihannic and E. Michaud.
Acknowledgments
The authors thank the LEEISA laboratory (A. Gardel, T. Maury) and IRD for providing laboratory facilities, C. Michelet for the meiofauna sorting during her Master’s Internship, and the support from the project “Biodiversity underestimation in our blue planet: artificial intelligence revolution in benthic taxonomy (BlueRev)” funded by Ifremer, ISblue and Equinor.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Álvarez-Castillo L., Hermoso-Salazar M., Estradas-Romero A., Prol-Ledesma R. M., Pardos F. (2015). First records of Kinorhyncha from the Gulf of California: horizontal and vertical distribution of four genera in shallow basins with CO2 venting activity. Cahiers Biol. Mar. 56, 271–281. doi: 10.21411/CBM.A.65052383
Annapurna C., Rao M. S., Bhanu C. H. V. (2015). Distribution of meiobenthos off Kakinada Bay, Gaderu and Coringa estuarine complex. J. Mar. Biol. Assoc. India 57, 17–26.
Balsamo M., Artois T., Smith J. P. S. III, Todaro M. A., Guidi L., Leander B. S., et al. (2020). The curious and neglected soft-bodied meiofauna: Rouphozoa (Gastrotricha and Platyhelminthes). Hydrobiologia 847, 2613–2644. doi: 10.1007/s10750-020-04287-x
Bianchelli S., Gambi C., Zeppilli D., Danovaro R. (2010). Metazoan meiofauna in deep-sea canyons and adjacent open slopes: a large-scale comparison with focus on the rare taxa. Deep-Sea Res. Part I: Oceanogr. Res. Pap. 57, 420–433. doi: 10.1016/j.dsr.2009.12.001
Cepeda D., Gayet N., Spedicato A., Michaud E., Zeppilli D. (2022a). Two new species of the Echinoderes coulli-group (Kinorhyncha: Cyclorhagida: Echinoderidae) from a low human-impacted mangrove swamp in French Guiana (western Atlantic Ocean). Zoologischer Anzeiger 301, 179–195. doi: 10.1016/j.jcz.2022.10.008
Cepeda D., González-Casarrubios A., Sánchez N., Spedicato A., Michaud E., Zeppilli D. (2022b). Two new species of mud dragons (Scalidophora: Kinorhyncha) inhabiting a human-impacted mangrove from Mayotte (Southwestern Indian Ocean). Zoologischer Anzeiger 301, 23–41. doi: 10.1016/j.jcz.2022.09.001
Cepeda D., Pardos F., Zeppilli D., Sánchez N. (2020a). Dragons of the deep sea: Kinorhyncha communities in a pockmark field at Mozambique Channel, with the description of three new species. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00665
Cepeda D., Trigo D., Pardos F., Sánchez N. (2020b). Does sediment composition sort kinorhynch communities? An ecomorphological approach through geometric morphometrics. Sci. Rep. 10, 2603. doi: 10.1038/s41598-020-59511-4
Conder J. M., Fuchsman P. C., Grover M. M., Magar V. S., Henning M. H. (2015). Critial review of mercury sediment quality values for the protection of benthic invertebrates. Environ. Toxicol. Chem. 34, 6–21. doi: 10.1002/etc.2769
Coull B. C. (1988). “Ecology of the marine meiofauna,” in Introduction to the Study of Meiofauna. Eds. Higgins R. P., Thiel H. (Smithsonian Institution Press, Washington D.C.), 18–38.
Dal Zotto M., Santulli A., Simonini R., Todaro M. A. (2016). Organic enrichment effects on a marine meiofauna community, with focus on Kinorhyncha. Zoologischer Anzeiger 265, 127–140. doi: 10.1016/j.jcz.2016.03.013
Della Patrona L., Marchand C., Hubas C., Molnar N., Deborde J., Meziane T. (2016). Meiofauna distribution in a mangrove forest exposed to shrimp farm effluents (New Caledonia). Mar. Environ. Res. 119, 100–113. doi: 10.1016/j.marenvres.2016.05.028
Fiard M., Cuny P., Sylvi L., Hubas C., Jézéquel R., Lamy D., et al. (2022). Mangrove microbiota along the urban-to-rural gradient of the Cayenne estuary (French Guiana, South America): Drivers and potential bioindicators. Sci. Total Environ. 807, 150667. doi: 10.1016/j.scitotenv.2021.150667
Gerwing T. G., Gerwing A. M. A., Drolet D., Hamilton D. J., Barbeau M. A. (2013). Comparison of two methods of measuring the depth of the redox potential discontinuity in intertidal mudflat sediments. Mar. Ecol. Prog. Ser. 487, 7–13. doi: 10.3354/meps10407
Giere O. (2009). “Meiobenthology,” in The Microscopic Motile Fauna of Aquatic Sediments, 2nd ed (Universität Hamburg, Hamburg).
Gomes C. A. A., Dos Santos P. J. P., Alves T. N. C., Rosa-Filho J. S., Souza-Santos L. P. (2002). Variaçao temporal da meiofauna em Area de Manguezal em Itamaraca – Pernambuco Vol. 24 (Atlântica, Rio Grande), 89–96.
Grzelak K., Sørensen M. V. (2019). Diversity and community structure of kinorhynchs around Svalbard: first insights into spatial patterns and environmental drivers. Zoologischer Anzeiger 282, 31–43. doi: 10.1016/j.jcz.2019.05.009
Hodda M., Nicholas W. L. (1986). Temporal changes in littoral meiofauna from the Hunter River estuary. Aust. J. Mar. Freshw. Res. 37, 729–741. doi: 10.1071/MF9860729
Hoffman K. P., Sánchez N., Sørensen M. V., Ingels J., Landers S. C. (2021). Kinorhynch communities of Mobile Bay and the Alabama continental shelf. Cahiers Biol. Mar. 62, 371–380. doi: 10.21411/CBM.A.B0EA3C57
INSEE, Institut National de la Statistique et des Études Économiques (2023) Commune de Cayenne, (97302), Dossier complet. Available at: https://www.insee.fr/fr/statistiques/2011101?geo=COM-97302 (Accessed March 06, 2023).
James W. N., Mark D. B. (2004). Marine biology: an ecological approach (San Francisco, USA: Person Education Inc).
Kotwicki L., De Troch M., Urban-Malinga B., Gheskiere T., Węslawski J. M. (2005). Horizontal and vertical distribution of meiofauna on sandy beaches of the North Sea (The Netherlands, Belgium, France). Helgoland Mar. Res. 59, 255–264. doi: 10.1007/s10152-005-0001-8
Landers S. C., Bassham R. D., Miller J. M., Ingels J., Sánchez N., Sørensen M. V. (2020). Kinorhynch communities from Alabama coastal waters. Mar. Biol. Res. 16, 494–504. doi: 10.1080/17451000.2020.1789660
Landers S. C., Sørensen M. V., Beaton K. R., Jones C. M., Miller J. M., Stewart P. M. (2018). Kinorhynch assemblages in the Gulf of Mexico continental shelf collected during a two-year survey. J. Exp. Mar. Biol. Ecol. 502, 81–90. doi: 10.1016/j.jembe.2017.05.013
Landers S. C., Sørensen M. V., Sánchez N., Beaton K. R., Miller J. M., Ingels J. (2019). Kinorhynch communities on the Louisiana continental shelf. Proc. Biol. Soc. Washington 132, 1–14. doi: 10.2988/18-00008
Marchand C., Lallier-Vergès E., Baltzer F., Albéric P., Cossa D., Baillif P. (2006). Heavy metals distribution in mangrove sediments along the Mobile Coastline of French Guiana. Mar. Chem. 98, 1–17. doi: 10.1016/j.marchem.2005.06.001
Michelet C., Zeppilli D., Hubas C., Baldrighi E., Cuny P., Dirberg G., et al. (2021). First assessment of the benthic meiofauna sensitivity to low human-impacted mangroves in French Guiana. Forests 12, 338. doi: 10.3390/f12030338
Neuhaus B. (2013). “Kinorhyncha (=Echinodera),” in Handbook of Zoology, Gastrotricha, Cycloneuralia and Gnathifera, Volume 1: Nematomorpha, Priapulida, Kinorhyncha, Loricifera. Ed. Schmidt-Rhaesa A. (Berlin, Germany), 181–348. doi: 10.1515/9783110272536.181
Nomaki H., Ogawa N. O., Ohkouchi N., Suga H., Toyofuku T., Shimanaga M., et al. (2008). Benthic foraminifera as trophic links between phytodetritus and benthic metazoans: carbon and nitrogen isotopic evidence. Mar. Ecol. Prog. Ser. 357, 153–164. doi: 10.3354/meps07309
Oksanen J., Simpson G., Blanchet F., Kindt R., Legendre P., Minchin P., et al. (2022) vegan: Community Ecology Package. R package version 2.6-4. Available at: https://CRAN.R-project.org/package=vegan.
Ostmann O., Nordhaus I., Sørensen M. V. (2012). First recording of kinorhynchs from Java, with the description of a new brackish water species from a mangrove-fringed lagoon. Mar. Biodivers. 42, 79–91. doi: 10.1007/s12526-011-0094-z
Punniyamoorthy R., Murugesan P., Mahadevan G., Sánchez A. (2021). Benthic meiofaunal diversity in four zones of Pichavaram mangrove forest, India. J. Foraminiferal Res. 51, 294–307. doi: 10.2113/gsjfr.51.4.294
R Core Team (2022). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/.
Sánchez A., Gómez-León A. (2024). Azoic sediments and benthic foraminifera: environmental quality in a subtropical coastal lagoon in the Gulf of California. Environ. Res. 244, 117924. doi: 10.1016/j.envres.2023.117924
Sánchez N., Zeppilli D., Baldrighi E., Vanreusel A., Lahitsiresy M. G., Brandily C., et al. (2020). A threefold perspective on the role of a pockmark in benthic faunal communities and biodiversity patterns. Deep Sea Res. Part I: Oceanogr. Res. Pap. 167, 103425. doi: 10.1016/j.dsr.2020.103425
Santos P. J. P., Botter-Carvalho M., Do Nascimento-Junior A. B., Marinho R. G. C., Carvalho P. V. V. C., Valença A. P. M. C. (2009). Response of estuarine meiofauna assemblage to effects of fertilizer enrichment used in the sugar cane monoculture. Pernambuco, Brazil. Braz. J. Oceanogr. 57, 43–55. doi: 10.1590/S1679-87592009000100005
Sarma A., Wilsanand V. (1994). Littoral meiofauna of Bhitarkanika mangroves of river Mahanadi system, east coast of India. Indian J. Mar. Sci. 25, 302–306.
Schrijvers J., Okondo J., Steyaert M., Vincx M. (1995). Influence of epibenthos on meiobenthos of the Ceriops tagal mangrove sediment at Gazi Bay, Kenya. Mar. Ecol. Prog. Ser. 128, 247–259.
Service Hydrographique et Océanographique de la Marine (1975). Service hydrographique et océanographique de la marine instructions nautiques: amérique du sud (Côte Est). 4, 325.
Sevastou K., Lampadariou N., Polymenakou P. N., Tselepides A. (2013). Benthic communities in the deep Mediterranean Sea: exploring microbial and meiofaunal patterns in slope and basin ecosystems. Biogeosci. Discuss. 9, 17539–17581. doi: 10.5194/bg-10-4861-2013
Sørensen M. V. (2023). New data on the Echinoderes coulli-group (Kinorhyncha: Cyclorhagida: Echinoderidae): a new species from New Caledonia mangroves, and a redescription of E. bengalensis (Timm 1958). Zoologischer Anzeiger 302, 90–101. doi: 10.1016/j.jcz.2022.11.009
Vanaverbeke J., Merckx B., Dgraer S., Vincx M. (2011). Sediment-related distribution patterns of nematodes and macrofauna: two sides of the benthic coin? Mar. Environ. Res. 71, 31–40. doi: 10.1016/j.marenvres.2010.09.006
Vanreusel A. (1990). Ecology of the free-living marine nematodes from the Voordelta (Southern Bight of the North Sea). I, Species composition and structure of the nematode communities. Cahiers Biol. Mar. 31, 439–462. doi: 10.21411/CBM.A.3D2ACB7F
Zeppilli D., Leduc D., Fontanier C., Fontaneto D., Fuchs S., Gooday A. J., et al. (2018). Characteristics of meiofauna in extreme marine ecosystems: a review. Mar. Biodivers. 48, 35–71. doi: 10.1007/s12526-017-0815-z
Keywords: mud dragons, meiofauna, pollution, trace metals, sediment grain size, mangrove forests, benthic habitat, anthropogenic pressure
Citation: Cepeda D, Sánchez N, Spedicato A, Michaud E and Zeppilli D (2024) Environmental drivers modelling the mangrove Kinorhyncha community along an urban-to-natural gradient in French Guiana (western Atlantic Ocean). Front. Mar. Sci. 11:1342763. doi: 10.3389/fmars.2024.1342763
Received: 22 November 2023; Accepted: 04 March 2024;
Published: 15 March 2024.
Edited by:
Alexander Ereskovsky, UMR7263 Institut méditerranéen de biodiversité et d’écologie marine et continentale (IMBE), FranceReviewed by:
Ajit Kumar Mohanty, Indira Gandhi Centre for Atomic Research (IGCAR), IndiaAlberto Sánchez-González, National Polytechnic Institute (IPN), Mexico
Copyright © 2024 Cepeda, Sánchez, Spedicato, Michaud and Zeppilli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Diego Cepeda, ZGllZ28uY2VwZWRhQHVhbS5lcw==