
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 19 March 2025
Sec. Marine Ecosystem Ecology
Volume 11 - 2024 | https://doi.org/10.3389/fmars.2024.1320194
This article is part of the Research TopicTropical Blue Carbon: Challenges and OpportunitiesView all 8 articles
 Jorge Alfredo Herrera-Silveira1
Jorge Alfredo Herrera-Silveira1 Erika Betzabeth Palafox-Juárez2,3
Erika Betzabeth Palafox-Juárez2,3 Juan Enrique Mendoza-Martínez1*
Juan Enrique Mendoza-Martínez1* Jorge Luis Montero Muñoz1
Jorge Luis Montero Muñoz1 Claudia Teutli Hernández4
Claudia Teutli Hernández4Seagrass meadows play a crucial role due to their ability to store carbon, mitigate the effects of climate change, and provide important ecosystem services such as coastal protection and water quality improvement. It has been observed that their carbon storage capacity varies depending on factors such as leaf structure, substrate type, depth, hydrodynamics, and spatial configuration. To assess the relationship between the landscape configuration of seagrass meadows and carbon storage, ten reef lagoons from the northern and southern regions of the Veracruz Reef System National Park were analyzed. Vegetation and soil data were collected from 513 sampling sites, and biomass organic carbon (BOC) and soil organic carbon (SOC) samples were taken at 153 of these sites, down to a depth of 25 cm. Through a supervised classification of WorldView2 satellite imagery, landscape maps were generated, and their composition and configuration were analyzed using 13 landscape metrics. A Generalized Additive Model fitted to distance-based redundancy analysis, was then applied to identify the relationship between landscape configuration and carbon storage (SOC5cm). Three types of seagrass meadows were identified: low density on rock, medium density on sand and rock, and high density on sand. Significant differences were observed in both carbon concentration and landscape configuration. High-density meadows exhibited the highest carbon concentrations in both biomass and soil (SOC25cm), while low-density meadows on rock recorded the lowest concentrations. The results showed that in the northern region, landscapes were more fragmented and had lower carbon storage capacity, whereas in the southern region, more cohesive and connected seagrass meadows stored greater amounts of carbon. Landscape metrics such as mean patch size and aggregation index were correlated with SOC5cm variations in southern landscapes, while edge density was the most influential metric in the north. The findings indicated that more fragmented landscapes with higher edge density exhibited lower carbon storage capacity, likely due to greater exposure to erosive processes. In contrast, more connected and cohesive landscapes, particularly in regions less affected by human activities, retained larger amounts of carbon in both biomass and soil. These findings highlight the importance of landscape configuration in the carbon storage capacity of seagrass meadows and emphasize the need to conserve their structural integrity to maximize their potential as carbon sinks and other ecosystem services.
Seagrasses are widespread around tropical and temperate shores. They develop in shallow areas where light, hydrodynamics, and substrate conditions are adequate for settlement, such as estuaries, marshes, swamps, lagoons, and reef lagoons (Short et al., 2007; Duarte et al., 2010, 2013; Davidson and Finlayson, 2019). Seagrass meadows are among the most important and productive habitats in coastal zones worldwide (Nordlund et al., 2016). They constitute the basis of many ecologically important marine habitats and provide several environmental services, such as protection of the coast, improved water quality (Heck et al., 2008; Kennedy et al., 2010; Mellbrand et al., 2011), feeding, shelter, and breeding areas for numerous faunal species (Nagelkerken, 2009; Boström et al., 2011; Nordlund et al., 2018). Costanza et al. (2014) estimated the global annual value of seagrass/macroalgae ecosystem services at US$ 28,916 ha year-1, while Short et al. (2011) estimated a value of US $34,000 ha year-1 based only on the nutrient cycle function.
Seagrasses, mangroves, and salt marshes are part of the blue carbon ecosystem. Owing to their important contributions in sequestering and storing carbon from the atmosphere and oceans, they are key elements in mitigating the effects of global climate change (Howard et al., 2014). The importance of coastal marine vegetation as a carbon stock is evident. Seagrasses make up approximately 0.1% of the total ocean floor and contribute 10–18% of the total oceanic carbon burial (Mcleod et al., 2011). Their reserves can originate from allochthonous or autochthonous sources and are fixed as living biomass or buried in the soil (Kennedy et al., 2010; Howard et al., 2018; Davidson and Finlayson, 2019). Estimates indicate that up to 19.9 Pg C is stored in the top meter of seagrass soil globally (Fourqurean et al., 2012), while their carbon accumulation rates range from 1.6 to 9.4 Tg C yr-1 (Miyajima and Hamaguchi, 2019) due to their wide distribution and constant exchange of matter and energy with other ecosystems.
Seagrasses form natural mosaics (landscapes) of homogeneous units (patches) distributed as islands within areas of sand, rock, or coral (matrix), adjusting to the patch matrix model, which is derived from the island biogeography theory (MacArthur and Wilson, 2001; Boström et al., 2011; Pittman et al., 2011). Landscape composition (size, number, and patch shape) and its configuration or spatial arrangement (distribution, density, and isolation) result from the interaction between abiotic and biotic factors and natural and anthropogenic disturbances (Boström et al., 2011; Pittman et al., 2011; Kupfer, 2012; Zarnetske et al., 2017). Factors such as water quality, hydrodynamics, substrate type, sediment transport, and rhizome growth rate define the conditions, density, and distribution of seagrass beds (Yeager et al., 2016; Uhrin and Turner, 2018; Pierrejean et al., 2023). Hurricanes, storms, and anthropogenic factors, such as eutrophication, dredging, and removal by direct impact of vessels, which can cause grounding, propeller scars, and damage from anchors, result in changes in the structure, connectivity, and fragmentation of seagrass (Macreadie et al., 2009; Montefalcone et al., 2010; Ricart et al., 2017; Holmquist et al., 2018; Spivak et al., 2019). Recent studies have suggested that the spatial configuration of seagrass influences the carbon fluxes between meadows and neighboring habitats (Grober-Dunsmore et al., 2009; Hyndes et al., 2014; Ricart et al., 2017). Therefore, analyzing the influence of seagrass meadow arrangements on their capacity to bury and store carbon provides substantial elements for designing conservation strategies for this important blue carbon ecosystem that contributes to mitigating the effects of global climate change, among many other environmental services.
The Veracruz Reef Systems National Park (VRSNP) has 50 reef systems, making it the largest reef system in the Central Gulf of Mexico (GM). It hosts the highest biodiversity of reef species in western GM and is at the midpoint of the southwestern GM reef corridor (Ortiz-Lozano et al., 2013, 2021; Horta-Puga et al., 2022). Veracruz reefs are exposed to natural and anthropogenic variability that influences the configuration of their seascape. This study aimed to estimate carbon stocks in seagrass meadows across 10 reef systems and assess their variation in relation to meadow configuration and spatial arrangement.
The VRSNP is located on the continental shelf of Veracruz state, Mexico, with extreme coordinates: 19°0’–19°16’ N; 95°45’–96°12’ W (Figure 1). It includes 50 coastal and platform coral reefs (DOF (Diario Oficial de la Federación), 2017; Liaño-Carrera et al., 2019) distributed in two clusters separated by the Jamapa River flow. The reefs of the northern region are characterized by being the smallest and shallowest and being exposed to greater anthropic influence as they are located in front of the City of Veracruz, the most populated in the state, and in the vicinity of Veracruz Harbor, the largest and most important commercial seaport in Mexico, built in the 18th century on land reclaimed from the sea; therefore, there is continuous and intense maritime, tourist, and fishing traffic. In contrast, the southern reefs are the largest and deepest in the park and receive discharge from the Jamapa River during the rainy season (Horta-Puga et al., 2016; Avendaño-Alvarez et al., 2017; Castañeda-Chávez and Lango-Reynoso, 2021; Salas-Monreal et al., 2022). They are in front of the fishing town of Antón Lizardo, approximately 20 km southeast of Veracruz City. In this region, naval traffic is mainly used for fishing activities; therefore, there is less anthropogenic influence than in the northern region (DOF (Diario Oficial de la Federación), 2017; Mapel-Hernández et al., 2021).
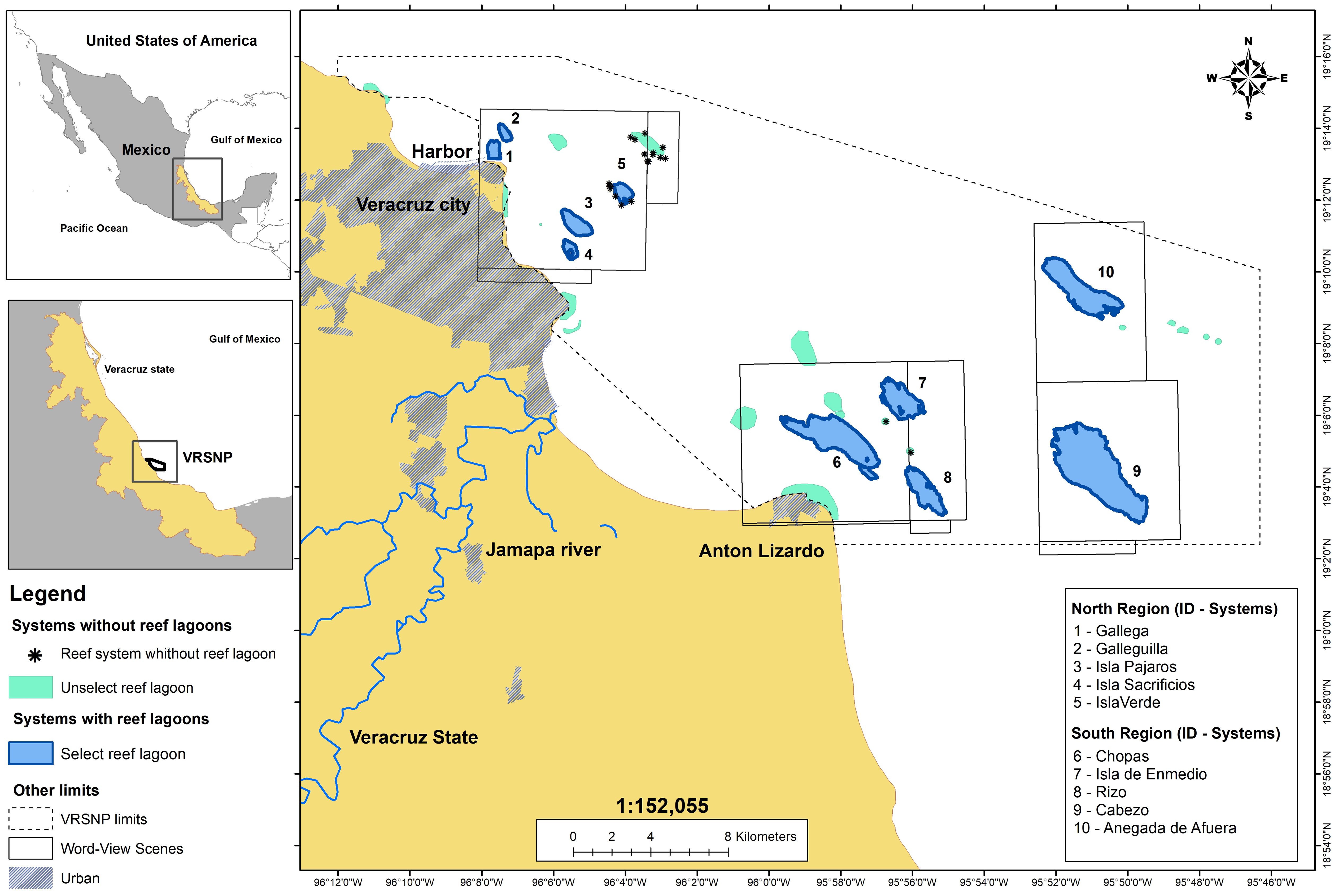
Figure 1. Location and distribution of the reef lagoons considered in the study.
Of the 50 reefs in the VRSNP, only ten contain seagrass meadows (Terrados et al., 2008; Ramírez-García and Pedraza-Venegas, 2019): Gallega, Galleguilla, Isla Pajaros, Isla Sacrificios, and Isla Verde in the north and Chopas, Isla de En Medio, Rizo, Cabezo, and Anegada de Afuera in the south (Figure 1). These reefs were selected for this study.
The process of evaluating the influence of landscape configuration on seagrass carbon storage capacity is presented in Figure 2 and is detailed in the following sections.
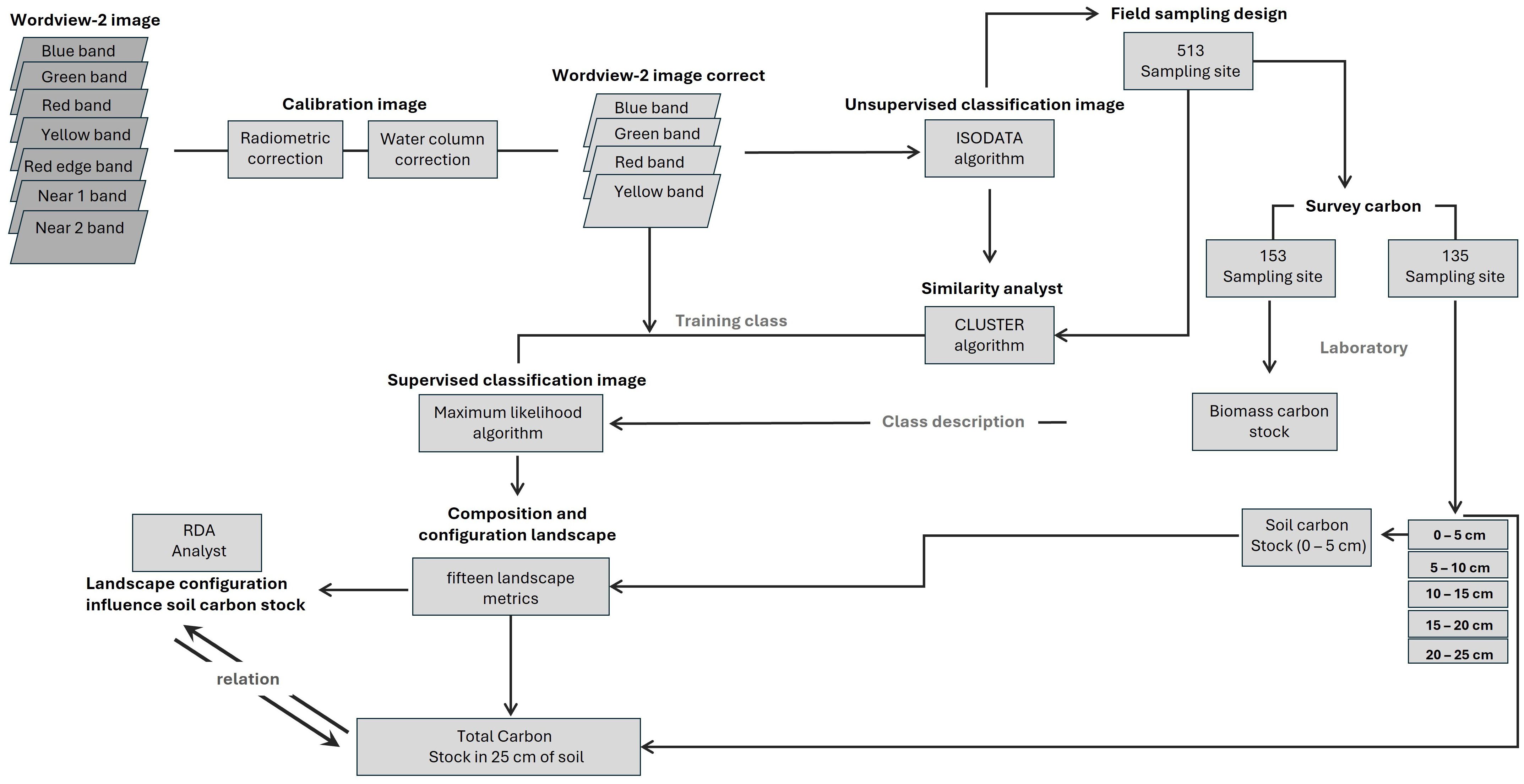
Figure 2. Schematic workflow to seagrass mapping and carbon stock relationship.
A Word-View-2 multispectral satellite imagery mosaic was constructed to cover the study area (Digital-Globe, 2017; Table 1). These images are widely recommended for seagrass studies because of their high precision and resolution (Updike and Comp, 2010; Coffer et al., 2020). To enhance the accuracy of the image data, an atmospheric correction was applied to remove atmospheric effects (Berk et al., 2003) using ERDAS Imagine software version 2014. A radiometric correction was performed in ENVI 5.3 software to adjust radiance or reflectance values, compensating for variations in illumination conditions and sensor response. Additionally, a water column correction was applied to mitigate the distortion caused by water on the signal received by the sensor (Maritorena et al., 1994; Stumpf et al., 2003).
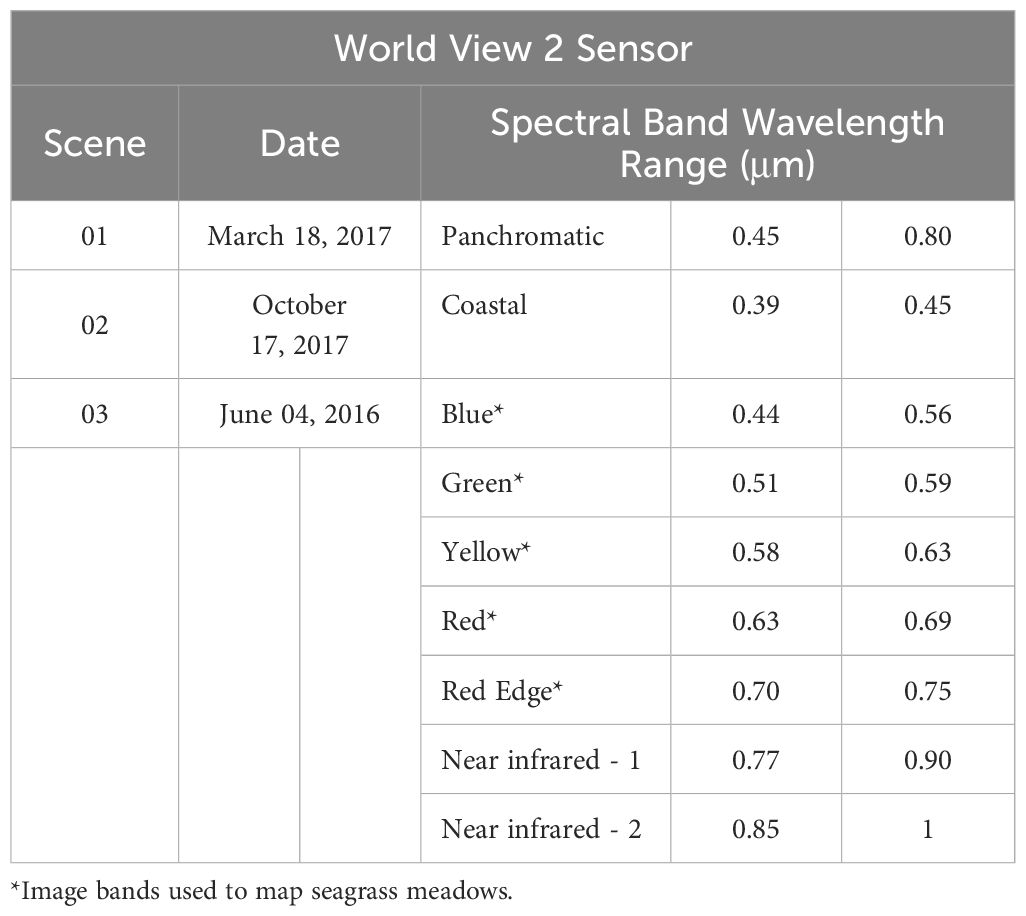
Table 1. Characteristics of Images used.
WorldView-2 imagery has eight bands in the visible and near-infrared (NIR) spectrum. Except for the two NIR bands, the other six bands (Coastal, Blue, Green, Yellow, Red, and Red Edge) could be used to map seagrass. For this study, the coastal blue and red edge bands were not considered, as they provide little information for classification (Su and Huang, 2019) compared to the other bands (blue, green, yellow, and red). Blue, green, yellow, and red bands were calculated using the Isodata method to distinguish spectrally different areas at the bottom of the lagoons; this variability was used as a base to design stratified sampling, obtaining 513 sites, with 162 in the north and 351 in the south (Figure 3). The number of sampling sites in each lagoon system varied according to its extent. This strategy improved the selection of sampling sites and captured spatial variability in seagrass meadows in past studies (Arellano-Méndez et al., 2016; Palafox-Juárez and de los Ángeles Liceaga-Correa, 2017; Cuevas et al., 2021).
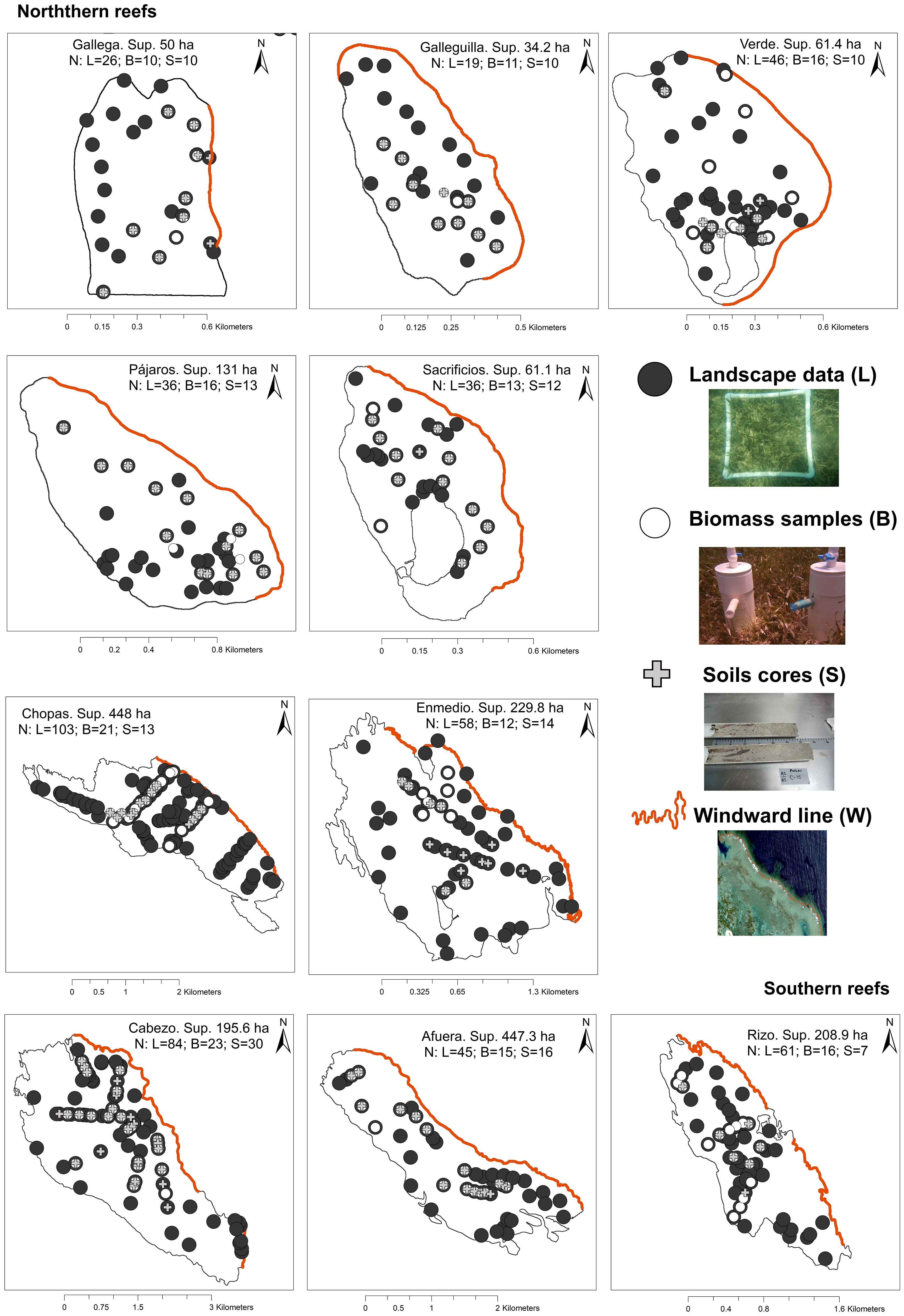
Figure 3. Location of sampling sites to identify seagrass landscape, biomass and soil carbon stock.
The survey was conducted between February and July 2018 using free diving or SCUBA methods, depending on the depth of each site. To characterize seagrass landscapes, a radius of 5 meters was established around each site coordinate (n=513; Figure 3). From the central point, a 0.25 m² quadrant was randomly distributed within each region defined by the cardinal directions (NE, SE, SW, and NW). In each quadrant, the dominant substrate type, abundance, coverage, and seagrass species were recorded in situ. Additionally, photographic evidence was collected for subsequent background analysis following the methodology of Hill and Wilkinson (2004).
Regarding environmental variables, depth was recorded at the centroid of each site using a HONDEX PS-7 sonde. Additionally, the percentage of incident light on the water surface, within the water column, and on the bottom was measured using a Li-Cor LI-250A console and an LI-193 submarine sensor. Finally, for each reef lagoon, a survey was conducted to delineate the windward line and estimate the length of the reef crest.
To evaluate carbon stocks, the standing crop method proposed by Milner and Hughes (1968) and the CARICOMP technique (2001) were used. Biomass (N=153) and soil samples (N=135) were taken at 153 sites (Figure 3). Encrusting coral, rock, or dead coral debris prevented the collection of soil samples at 18 of the 513 selected sites. Each sample was taken in duplicate (306 total samples for biomass and 270 total samples for soil) using PVC core samplers with diameters of 15 cm and depths of 10 cm for biomass and diameters of 6 cm and depths of 60 cm for soil. Biomass samples were stored in Ziploc bags and preserved at 5°C for subsequent laboratory analysis.
For the soil samples, core samplers with sharp edges at the bottom of the PVC corers were used and inserted vertically using a combination of manual and mechanical percussion (Serrano et al., 2012). Once inserted into the target depth, a PVC cap was placed on the top of the corer to create a vacuum and obtain the soil profile sample. Not all cores reached the target depth of 60 cm, so only the first 25 cm of depth were used. The extracted soil cores were then capped on both ends and stored vertically at -5°C for subsequent sectioning into five 5 cm subsamples in the laboratory. To prevent soil compression from the collection method, a correction factor was calculated based on the ratio of the sample length to the inserted PVC corer length. The corrected sample length was determined by multiplying the desired depth interval by the compaction correction factor (Howard et al., 2018). On average, the compression factor was 0.89.
A database was constructed using data from 513 sites, resulting in 2,052 data points (four quadrants per site). A fourth root transformation was applied to the data collected at the quadrat level, including seagrass cover and substrate characteristics, to normalize the frequency distribution. To explore the similarity of seagrass cover and substrate among sites, a cluster analysis was performed using the Bray-Curtis similarity index, following the recommendations of Clarke et al. (2014) and utilizing PRIMER v6 software (Gorley and Clarke, 2006). To define seagrass landscape types, the grouped data were used as training sets to classify WorldView-2 images using the blue, green, red, and yellow bands (Su and Huang, 2019) with the maximum likelihood method in Envi 5 (Bello-Pineda et al., 2015; Palafox-Juárez and de los Ángeles Liceaga-Correa, 2017). Finally, the kappa index (Lillesand et al., 2015) was calculated to evaluate the accuracy of the seagrass landscape maps using data from 200 randomly selected sampling sites that were not used in the classification.
According to the patch matrix model, seagrass meadows are represented as binary landscapes, where seagrasses are the elements of interest, and unvegetated areas constitute the matrix (Collinge, 2009; MacArthur and Wilson, 2001; Boström et al., 2011; Wedding et al., 2011; Singh et al., 2014; (McGarigal, 2015; Ricart et al., 2017). Fifteen landscape metrics (Table 2) were calculated using the LecoS plugin (Jung, 2016) in QGIS 3.14 software to analyze the spatial composition and configuration of the seagrass landscape.
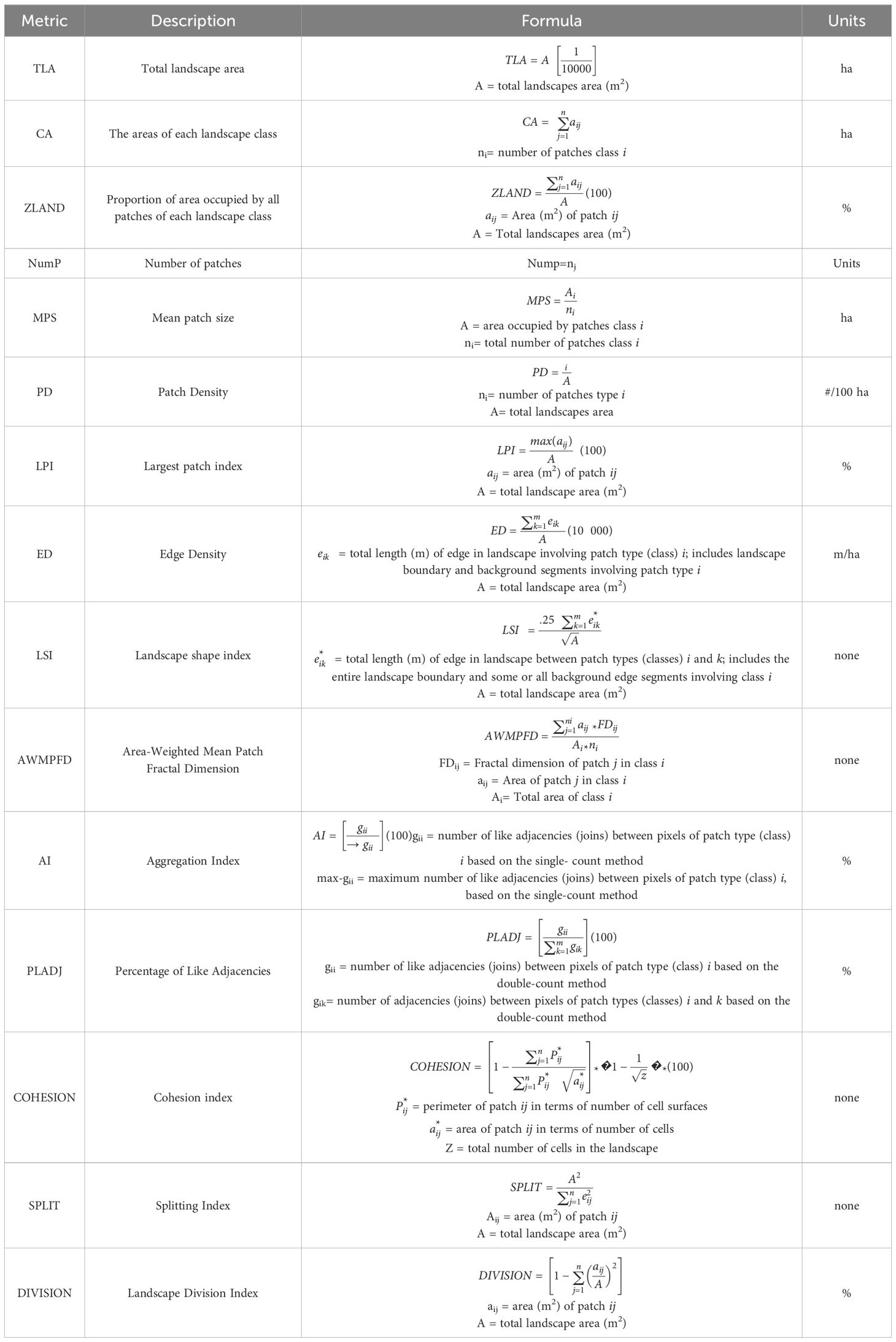
Table 2. Estimated landscape metric details (McGarigal, 2015).
The seagrass samples were rinsed with fresh water to remove sediments, carbonates, seaweed, detritus, and other organisms. Epiphytes were manually removed from the leaves using a blade. To obtain the biomass organic carbon stock (BOC), both the aboveground (leaves and stems, living or dead) and belowground (roots and rhizomes) biomass were considered. The components were oven-dried at 60°C for 72 hours or until a constant weight was achieved. This dry weight was converted to carbon equivalents, assuming a carbon content of 35% of the seagrass biomass dry weight (Fourqurean et al., 2012). Finally, the results were extrapolated to megagrams of carbon per hectare (Mg C ha-1-1) using the area sampled by the core sampler.
The soil cores were cut crosswise and sectioned every 5 cm according to the corrected length. A sample from each core section was dried in an oven at 70°C for 72 hours (Howard et al., 2014). The apparent density (Pb) was estimated from the dry weight. The total carbon content (% TC) in each soil section, up to a depth of 25 cm, was determined using a Flash EA 1112 elemental analyzer (CHN). The inorganic carbon content (% IC) was estimated by acidifying a 30 mg subsample with 100 ml of HCl for 4 hours at a concentration of 12 mol L-1 (Harris et al., 2001). The organic carbon content (% OC) was calculated by subtracting the inorganic carbon content from the total carbon content of each core section. The soil organic carbon stock (SOC) was then calculated to a depth of 25 cm and extrapolated to megagrams of carbon per hectare (Mg C ha-1) using Equation 1.
where Pb is Dry bulk density (g cm3), Sn is Soil core section (cm), and OC is Organic carbon (%).
To evaluate the effect of seagrass meadow structure on soil organic carbon accumulation, we compared the average organic carbon concentrations (% OC) along the soil profile across different seagrass landscapes within each region. Additionally, to determine the magnitude of organic carbon stored in biomass and soil for each landscape across the ten reef systems, the BOC and SOC values were analyzed using nonparametric confidence intervals with the adjusted bootstrap percentile (BCa) method (Davison and Hinkley, 1997), with B=1,000 replications. The BCa interval is a second-order correction that addresses bias and skewness in the distribution of bootstrap estimates through an accelerated bias-correction method (Chernick and LaBudde, 2014). BCa intervals were calculated using the boot. ci function in R software (R Core Team, 2018) with the boot library (Canty and Ripley, 2019).
Given the importance of the top few centimeters of soil about the presence or absence of the canopy (Tanaya et al., 2018; Santos et al., 2019), we evaluated the relationship between different spatial configurations of seagrass landscapes and soil carbon stock by using the average value of the top 5 centimeters of soil (SOC5cm). The statistical analysis was conducted in two phases to assess the impact of landscape configuration on the SOC5cm of seagrass meadows. First, principal component analysis (PCA) was performed to select the relevant variables (landscape metrics), using loading scores as the statistical criterion. Metrics with high loadings were considered to have a greater contribution to the variance explained by PCA1.
The selected landscape metrics were then used in canonical ordination analysis as predictor variables for carbon stock. Second, a dataset comprising SOC5cm and landscape metrics was analyzed using distance-based redundancy analysis (db-RDA) (Legendre and Anderson, 1999). The db-RDA is a canonical ordination method that relates a triangular matrix to response variables, such as landscape metrics (Legendre and De Caceres, 2013). This analysis used soil carbon stock data, and a triangular matrix was computed using Euclidean distance. Variance Inflation Factors (VIF) were employed to identify and select variables to mitigate multicollinearity among landscape metrics. When collinearity exceeded 0.8, only one variable was retained to ensure that the VIF consistently remained below five within the metric set (Quinn and Keough, 2002).
The db-RDA statistical approach effectively identifies landscape metrics that best explain variations in soil carbon storage. To quantify the influence of these metrics on soil carbon storage, linear models or Generalized Additive Models (GAM) were fitted to the RDA results. The db-RDA analysis was conducted using the capscale function from the vegan package (Oksanen et al., 2020) version 2.6-4 within the R environment (R Core Team, 2023).
Finally, to evaluate the correlations between independent variables (landscape metrics) and changes in soil carbon stock across different landscape types, we employed the envfit and ordisurf functions from the vegan package (Oksanen et al., 2020) in the R environment (R Core Team, 2023). These functions provide squared correlation coefficients based on a permutation test with 9,999 random permutations.
Seagrass landscapes in the VRSNP are characterized by the presence of seagrass beds composed mainly of Thalassia testudinum (Tt) and, to a lesser extent, mixed beds of Tt and Syringodium filiforme (Sf), growing on a combination of sandy and rocky substrates. In areas described as having rocky substrates, the seagrass does not grow directly on solid rock but rather in an unconsolidated layer of sand and sediment that overlays bedrock or coral rubble. The thickness of this sediment layer varies across the lagoon systems, but it is generally sufficient to allow for the penetration of seagrass rhizomes. In some areas, coral rubble also contributes to the substrate, providing additional structure for seagrass colonization. This stratified nature of the substrate allows for the coexistence of both sand and rock components in the seagrass beds (Figure 4).

Figure 4. Bottom characteristics in each seagrass landscape. (A) LD-R: Low density over rocks; (B) MD-SR: Medium density over sand-rock; (C) HD-S: High density over sand.
The species composition, seagrass cover, and associated substrate defined the distribution of three seagrass landscapes embedded within an unvegetated matrix of coral, rock, or sand (unvegetated area) (Figure 5). Low density over rocks (LD-R) was characterized by a seagrass bed mix of Tt and Sf growing on rocky substrates, with the lowest seagrass cover (aboveground) and rhizoidal structures (belowground) of all lagoon systems. The average shoot density for this landscape was estimated at 264.6 to 848.8 shoots/m2 for the north and 346.6 to 905.4 shoots/m2 for the south (Supplementary Table 1; Figure 4A).
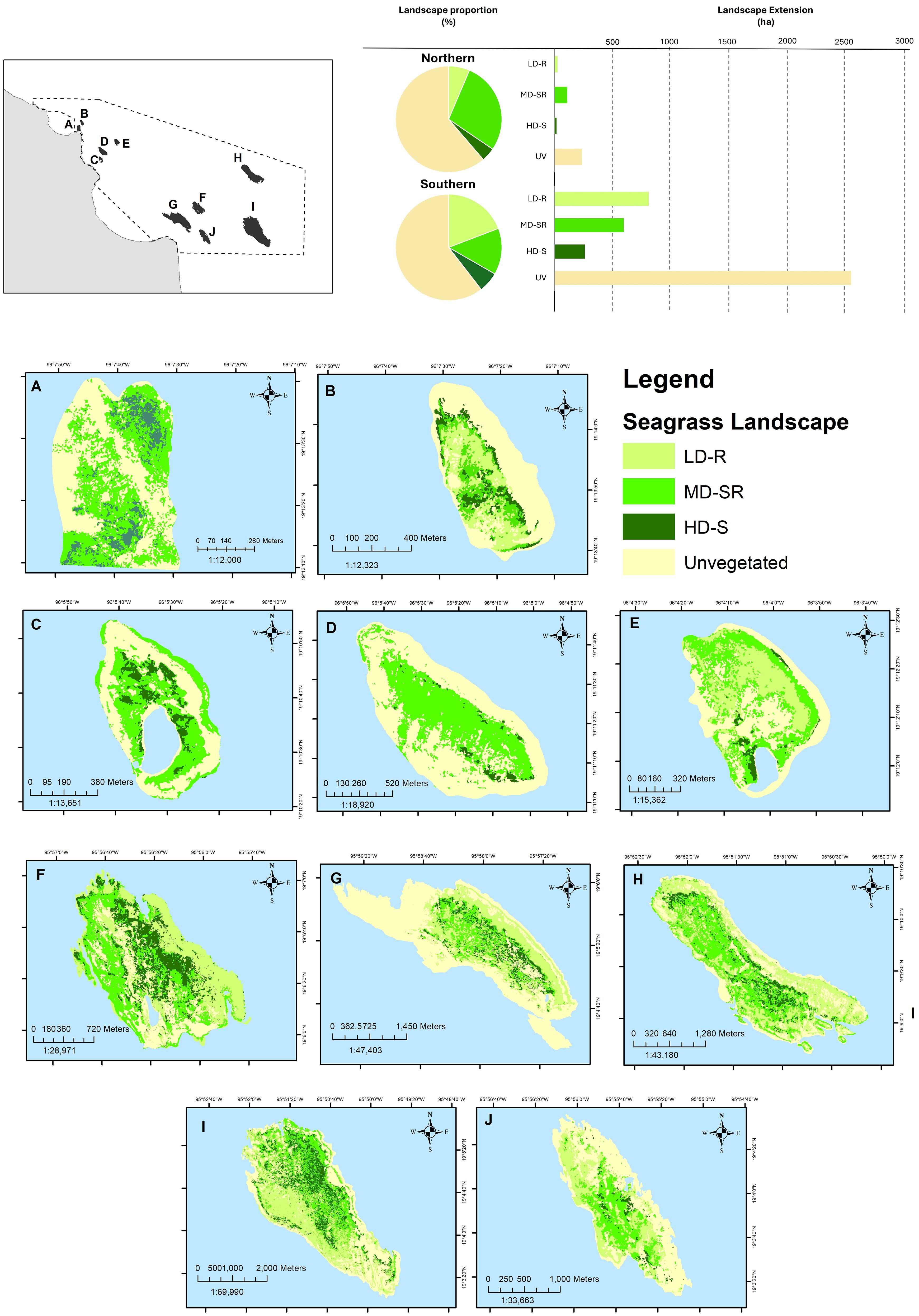
Figure 5. Distribution of seagrass landscapes in the ten lagoon systems. LD-R: Low density over rocks; MD-SR: Medium density over sand-rock; HD-S: High density over sand. (A) Gallega; (B) Galleguilla; (C) Isla Verde; (D) Isla Pájaros; (E) Isla Sacrificios; (F) Chopas; (G) Isla de en medio; (H) Isla Cabezo; (I) Anegada de Afuera; (J) Rizo.
Medium density over sand-rock (MD-SR) was defined as seagrass beds mainly composed of Tt and Sf in a lesser proportion over a sandy, rocky substrate. The seagrass structural complexity was larger than LD-R, and the average shoot density was 452.7 to 1,991.9 shoots/m2 in the north and 282.9 to 1,499.6 shoots/m2 in the south (Figure 4B). High density over sand (HD-S) was characterized by seagrass meadows with the highest shoot density (719.2 to 1,754.2 shoots/m2 in the north and 169.8 to 1,612.8 shoots/m2 in the south). Tt dominated the predominantly sandy bottom (Supplementary Table 1; Figure 4C).
The distribution and extent of seagrasses in the VRSNP are characteristics of each reef system, including the linear length of the reef crest, depth, light incidence, and distance to the windward side. In the northern systems, reef lagoons are bordered by an average windward length of 1.9 km (Supplementary Table 2), with Gallega having the shortest length (0.7 km) and I. Pajaros the longest (3 km). These northern systems are generally small and shallow (0.4 m to 1.6 m deep), with minimal light incidence in Gallega (17%) and a range between 40% and 53% for the other systems in this region. In contrast, southern systems are larger, with a more extensive linear area protected by the windward side (6 km), deeper waters ranging from 0.5 m to 2.5 m, and light incidence between 18% and 54% (Supplementary Table 2).
Reef lagoons occupied a total area of 4,592 ha; 91.6% (4,208 ha) corresponded to the southern systems and 8.4% (383 ha) to the northern systems. This extent is considered to be vegetated and unvegetated areas, particularly those with seagrass meadows, covering 1,810 ha, with 1,662 ha in the south and 148.6 ha in the north (Figure 5; Supplementary Table 3). This seagrass distribution presented low error percentages (89% - 94%) according to the overall kappa coefficient of the classification (Supplementary Table 4).
Concerning the extent, LD-R was the most extensive landscape in the southern region (809 ha), and MD-SR (107.8 ha) was the most extensive in the north. In the south, LD-R landscapes were distributed at an average linear distance of 712.2 ± 139.8 m from the reef crest relative to other landscapes in that region. In contrast, in the north, MD-SR landscapes were distributed at an intermediate distance from the reef crest (203.5 ± 35.5 m) relative to other landscapes (Supplementary Table 3). High density over the sand landscape was the least extensive in both regions: 16.1 ha in the northern with a greater distance from the reef crest (239.6 ± 58.3 m) and 259 ha in the southern systems with a shorter distance (572.1 ± 84.6 m) from the reef crest. HD-S and MD-SR were present in all systems, whereas LD-R was absent in all three systems in the north (Supplementary Table 3).
Regarding patch size (MPS), in the northern region, the largest patches were present in the LD-R of Galleguilla and Verde, with an average patch size of 1,400 m2. In contrast, the smallest fragments were found at HD-S landscape of Gallega (MPS=230 m2). The largest patches were observed in the south region in LD-R and MD-SR at Chopas (MPS = 1,940 and 1,650 m2, respectively). Smaller fragments were observed in the HD-S of A. Afuera (MPS =350 m2).
Edge Density (ED) provides information about the connectivity of patches of the same type with neighboring habitats; a high edge density reduces connectivity and vice versa. In the case of northern landscapes, the ED varies between 50.4 m/ha for HD-S in I—pajaros to 1,077.8 m/ha for the MD-SR landscape of the Gallega system. For the southern landscapes, the range of variation is narrowest, with an ED = 70.8 for HD-S in Rizo and 721.5 m/ha for LD-R in the Cabezo system.
The Area-Weighted Mean Patch Fractal Dimension (AWMPFD) provides information about patch shape complexity based on the area-perimeter relationship. For the landscapes in the northern region, the highest AWMPFD was observed in the MD-SR landscape in Gallega and Isla Verde, with 1.5 and 1.4, respectively, while the lowest values occurred in HD-S of I—Pajaros, with 1.2. Regarding the southern landscapes, the results showed that the MD-SR and LD-R landscapes presented high sinuosity in the patches of all systems, with an AWMPFD of 1.5. The lowest values (AWMPFD =1.2) were observed in the HD-S landscape for Rizo.
Regarding the spatial distribution of the fragment landscape, patch density (PD) values showed that, in general, northern landscapes were more dispersed than southern ones, highlighting HD-S in Gallega (PD = 0.44 patches/100 ha) with the greatest dispersion in the study area, while for the southern region, the most dispersed landscape was HD-S in A. Afuera (PD = 0.28/100 ha). Regarding low dispersion, the results showed that, in the north, the lowest dispersion was observed in LD-R at Galleguilla and Verde and MD-SR at Isla Sacrificios, all with PD = 0.072/100 ha. In contrast, LD-R at Chopas was the least dispersed in the southern region in the study area (PD = 0.05 patches/100 ha).
Regarding the Agrupation Index (AI), this index describes the aggregation degree of patches of the same type; for the northern systems, it was observed that the HD-S landscape in Gallega presents the lowest aggregation with an AI = 81.8%, while the greatest aggregations occur in the MD-SR landscape of Isla Pajaros, with an AI = 95.3%. Regarding the southern landscapes, it was observed that the lowest aggregation is in MD-S landscapes in Cabezo and HD-S in Chopas, with an AI of 86.5%. In comparison, the highest aggregation occurred in the LD-R landscape of the Chopas system with AI = 93.4%.
Finally, concerning Percentage of Like Adjacencies (PLADJ), which explains the degree of aggregation between patches in different habitats, the results showed the same trend observed for IA. The PLADJ varied from 81.1% in HD-S in Gallega to 95% in MD-SR in Isla Pajaros for the northern systems. At the same time, for the southern landscapes, it observed a lower variation of aggregation degree, with the highest (93.2%) and lowest (PLADJ = 86.2%) patch aggregation in the same system, LD-R and HD-S in Cabezo, respectively.
The organic carbon concentrations (% OC) along the soil profile showed differences between landscapes with different structures and regions (Figure 6). Seagrass landscapes with lower foliar structure (LD-R) exhibited the lowest OC concentrations, with small variations between 4% and 5% along the soil profile. On the other hand, in MD-SR and HD-S landscapes, where the seagrass foliar structure was greater, the organic carbon concentrations in the soil were also higher, ranging from 4.5% to 6%, with a slight decrease in deeper soil horizons (> 10 cm), except for HD-S landscapes in the northern region, where the upper soil horizons had the lowest OC concentrations (4.7%).
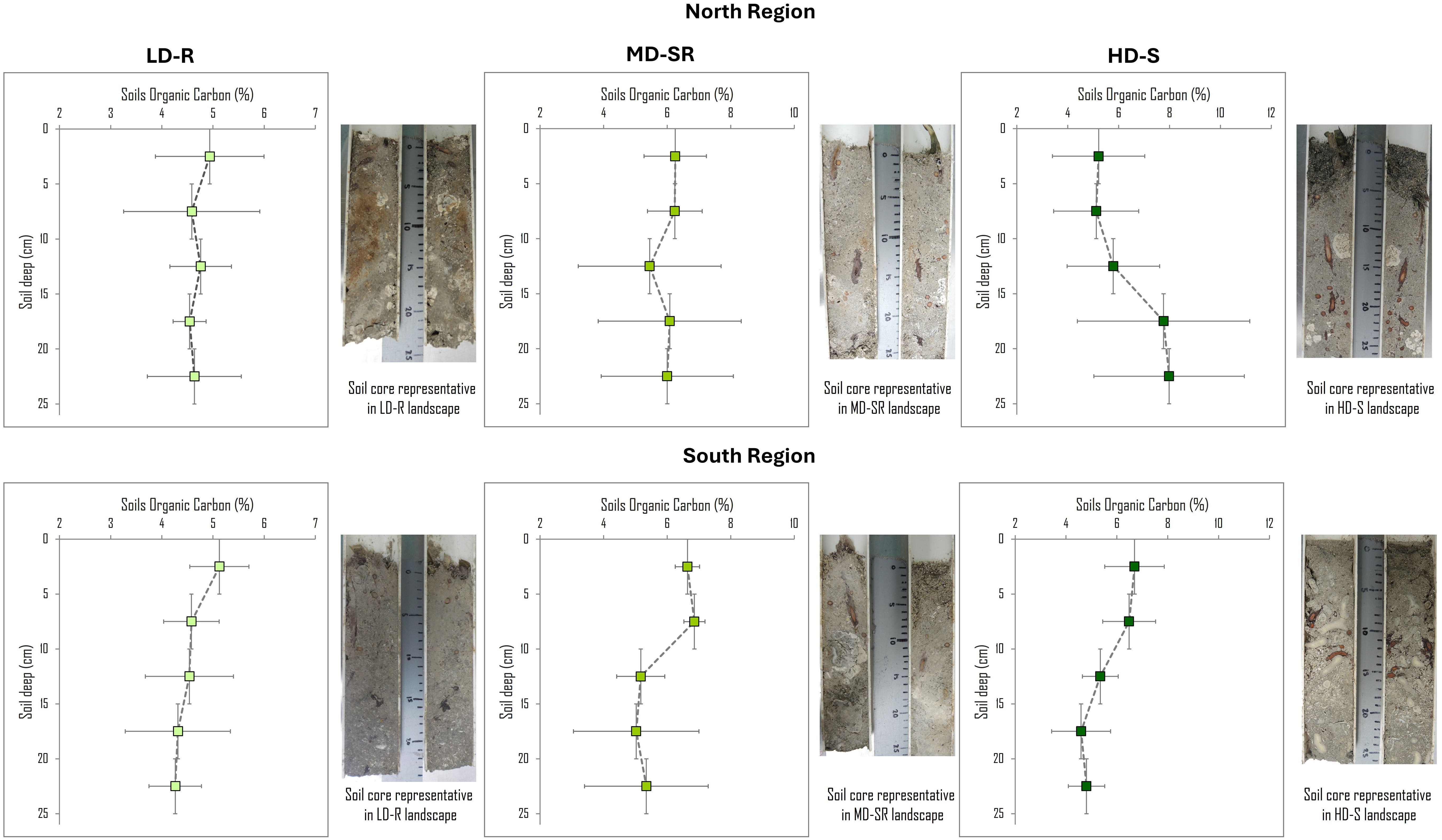
Figure 6. Organic carbon concentration (% OC) and depth profiles of seagrass landscape by region in VRSPN. LD-R: Low density over rocks; MD-SR: Medium density over sand-rock; HD-S: High density over sand. Data are mean values ± standard error (SE) of Organic Carbon; for the Y axis, the depth of each profile is the average value of each section in the profile; 2.5 cm = 0–5 cm; 7.5 cm = 5-10 cm; 12.5 cm = 10-15 cm; 17.5 cm = 15-20 cm; 22.5 cm = 20-25 cm).
The 95% confidence interval Bootstrap BCa analysis results demonstrated significant differences in carbon storage of BOC among the seagrass landscapes from the north compared to those from the south in the VRSNP. In the southern systems, the three seagrass landscapes store a lower concentration of carbon in biomass (BOC), with confidence intervals ranging from 0.5 - 0.9 Mg C ha-1 for LD-R in Rizo and 1.6 - 5.3 Mg C ha-1 for HD-S in A. Afuera (Supplementary Table 5). In contrast, the HD-S landscapes of I. Sacrificio (3.2 - 8.2 Mg C ha-1) and Gallega (3.7 - 6.9 Mg C ha-1) in the north showed higher BOC values with significant differences compared to the landscapes from the south. However, high variation ranges were observed within each seagrass landscape of the ten reefs studied, indicating significant site variability (Figure 7).
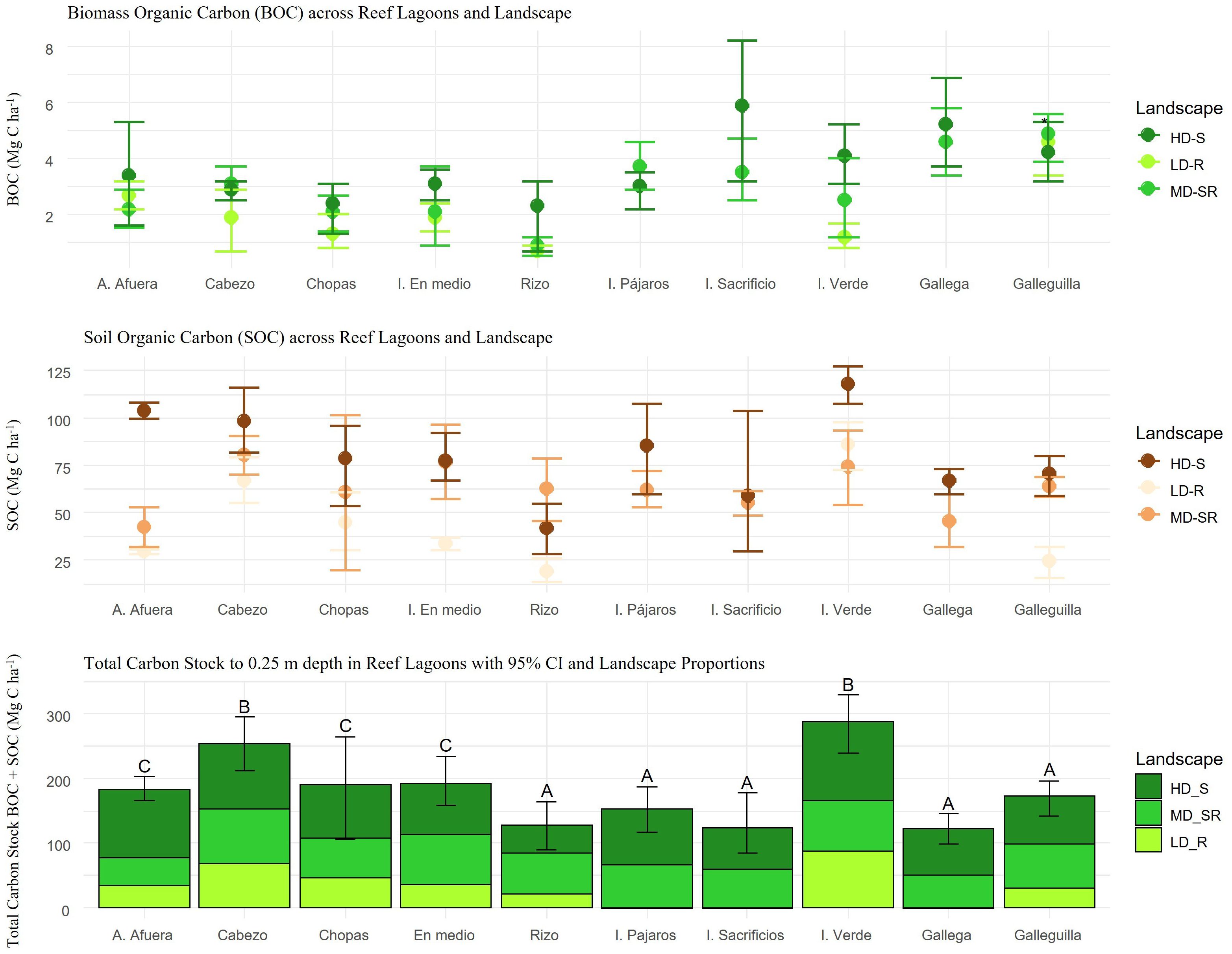
Figure 7. Bootstrap Bca confidence interval 95% (CI) of A) biomass organic carbon stock (BOC) and B) soils organic carbon stock (SOC) of the landscape class. C) Variation of total carbon stock (BOC + SOC) of the landscape class across reef lagoons (mean ± 95% CI). Letters indicate significant differences in biomass (p < 0.01). LD-R: Low density over rocks; MD-SR: Medium density over sand-rock; HD-S: High density over sand.
Regarding SOC, the 95% confidence interval Bootstrap BCa analysis results demonstrated significant differences between HD-S landscapes in the north and the south (Figure 7). The SOC was standardized and calculated to a depth of 25 cm for all sites. However, the total soil depth varied among landscapes. In the northern systems, SOC varied from 107.4 - 126.9 Mg C ha-1 in HD-S of Isla Verde with an average total soil profile depth of 27.6 ± 6.1 cm and from 59.5 - 107.4 Mg C ha-1 in HD-S of the I. Pajaros system with an average depth of 22.2 ± 1.7 cm. For the southern landscapes, SOC varied from 99.6 - 107.8 Mg C ha-1 in HD-S of A. Afuera over a total soil profile depth of 36.9 ± 3.1 cm and from 81.4 - 115.6 Mg C ha-1 in HD-S of the Cabezo system with an average depth of 25.7 ± 1.6 cm (Supplementary Table 5). The landscapes with the lowest SOC were Galleguilla in the north and I. Rizo in the south, with intervals ranging from 32.2 - 59.4 Mg C ha-1 in 22.7 ± 1.7 cm of soil and from 13.5 - 26 Mg C ha-1 in 17.6 ± 1.2 cm of soil, respectively.
Finally, the bottom panel of Figure 7 shows the total carbon stock (BOC + SOC) in the landscape system within the VRSNP and the differences in carbon storage capacity. The systems with the highest total carbon stock were Cabezo (95% CI = 212.4 - 294.9 Mg C ha-1) in the south and the I. Verde reef (95% CI = 238.9 - 392 Mg C ha-1) in the north. In contrast, the systems with the lowest total carbon stock were Gallega (95% CI = 98.7 - 145.2 Mg C ha-1) and the I. Sacrificios reef (95% CI = 83.8 - 177.6 Mg C ha-1) in the north and Rizo (95% CI = 89.1 - 164.3 Mg C ha-1) in the south.
The distribution of carbon among the different landscapes is also evident. The HD-S and MD-SR landscapes generally contribute to a higher carbon storage capacity across all reefs, both in BOC (2%) and SOC (40.4%). Notably, although reefs in the north exhibit higher BOC accumulation (Figure 7 top and middle panels), the proportions of carbon stored in biomass (BOC) are comparatively lower. For instance, in the I. Verde reef, despite having one of the highest total carbon stocks (286.3 ± 24.1 Mg C ha-1), the relative contribution of BOC is lower (2.8%) compared to SOC (97.2%). Conversely, in Gallega, BOC represents a larger proportion of the total carbon storage (8.1%) despite having a lower capacity to store SOC (91.9%).
In the south, reefs like Cabezo stand out for their high total carbon stock (253.6 ± 21.1 Mg C ha-1), with a more balanced distribution between BOC and SOC than other reefs in the region. This contrasts with Rizo, which shows a lower carbon storage capacity (127.4 ± 27.6 Mg C ha-1), particularly in its biomass (3.1%).
The distance-based redundancy analysis (db-RDA) shows the influence of different metrics on the variation of the top 5 centimeters of soil organic carbon (SOC5cm) across different landscapes present in the VRSNP. For the northern systems, no significant linear relationships were found between landscape configuration and variations in SOC5cm (Figure 8; Table 3). However, the adjusted generalized additive model (GAM) showed a significant non-linear relationship between edge density (ED) and carbon accumulation, with an F-value of 0.77 and p = 0.08, explaining 49.4% of the total deviation.
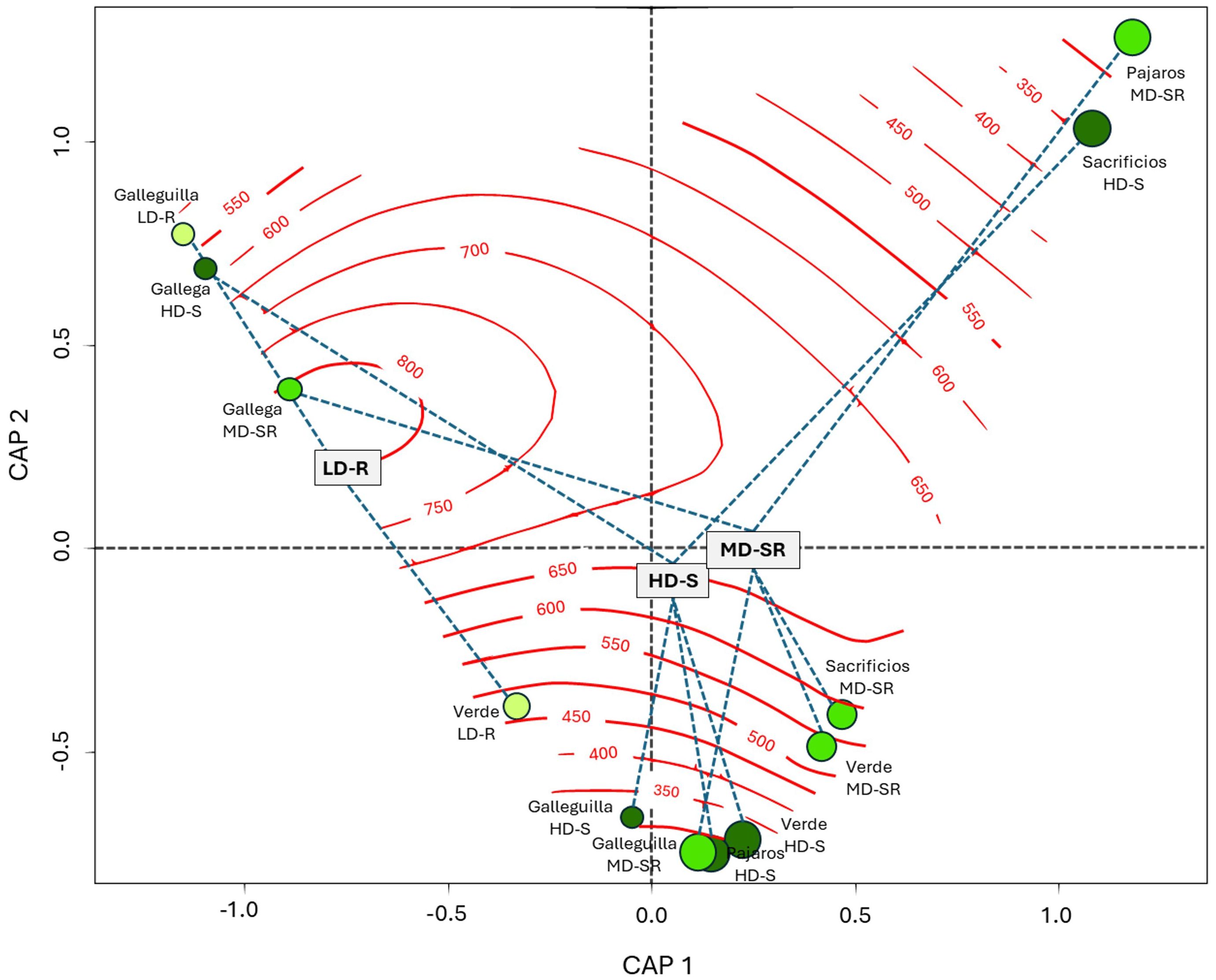
Figure 8. Redundancy Analysis (RDA) between top 5 centimeters of soil organic carbon (SOC5cm) stock (Mg C ha-1) and Landscape metrics in seagrass meadows of the North. LD-R: Low density over rocks; MD-SR: Medium density over sand-rock; HD-S: High density over sand.
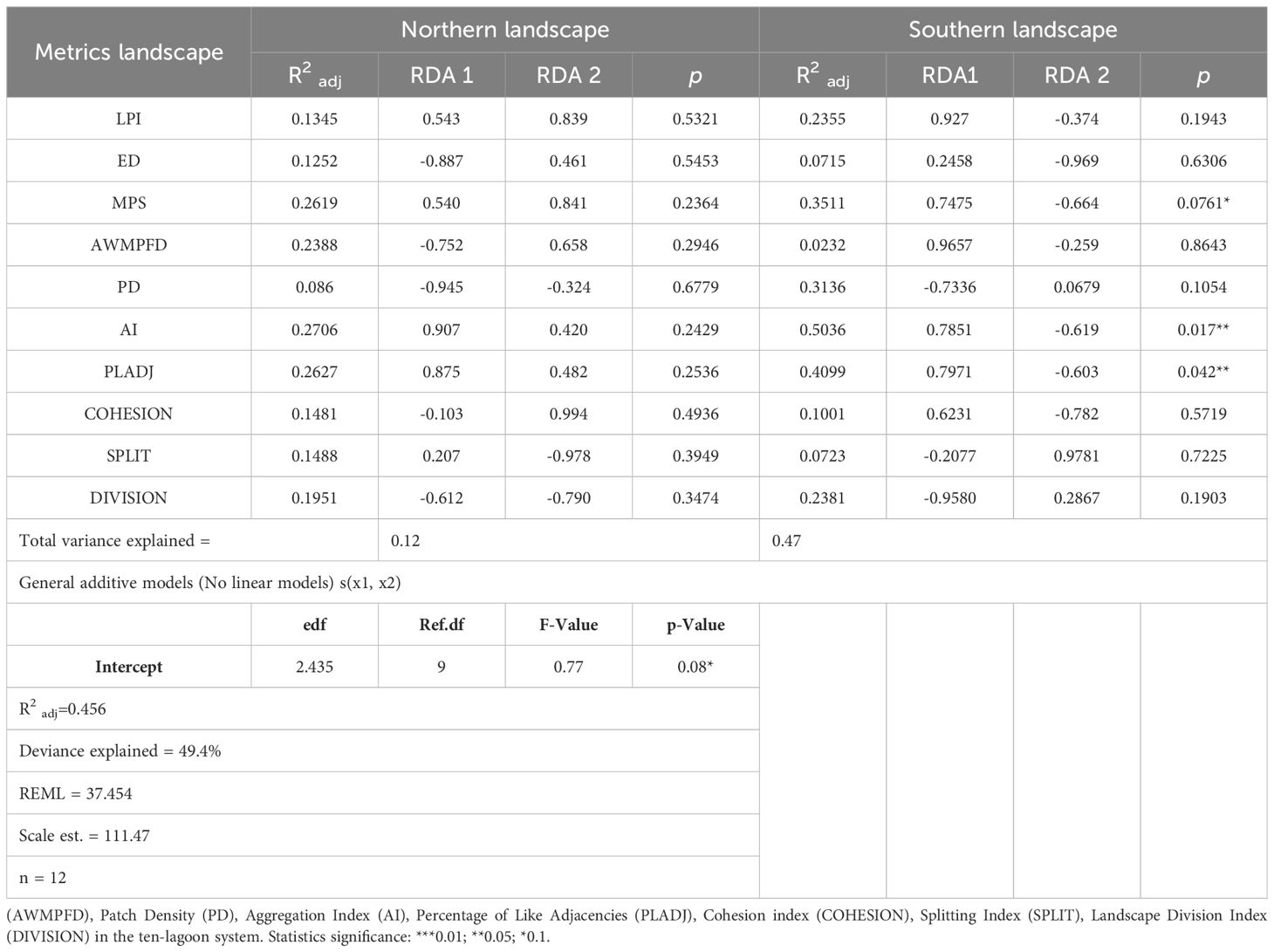
Table 3. Statistical summary of Redundancy Analysis (RDA) between Soil Organic Carbon Stock (Mg C ha-1) of top 5 cm (SOC5cm) seagrass meadows and different landscape metrics of class area (CA), Largest patch index (LPI), Edge Density (ED), Mean patch size (MPS), Area-Weighted Mean Patch Fractal Dimension.
The ordination plot displays isolines representing the increase and decrease of Edge Density (ED) about SOC5cm values. In this study, edge density explained 45.6% (adjusted R2) of the variation in SOC5cm. It was observed that landscapes with higher edge densities, such as LD-R in Galleguilla and Verde and MD-SR and HD-S in Gallega, tended to store less carbon in the soil.
On the other hand, the landscapes that stored more carbon in the soil were the HD-S in Sacrificios and Verde and the MD-SR in Pajaros and Verde. Although variable in structure, these landscapes exhibited less fragmentation or a higher degree of internal connectivity, which could have allowed for greater carbon retention despite possible variations in ED.
In the southern landscapes, a significant linear relationship was observed between landscape configuration and SOC (Figure 9). On Axis 1 (CAP1), positive correlations were found for the Largest Patch Index (LPI) (r2 = 0.2355, p = 0.1943) and the Cohesion Index (COHESION) (r2 = 0.1001, p = 0.5719); significant correlations were also found for Mean Patch Size (MPS) (r2 = 0.3511, p = 0.0761), Aggregation Index (AI) (r2 = 0.5036, p = 0.0177), and Percentage of Like Adjacencies (PLADJ) (r2 = 0.4099, p = 0.0429).
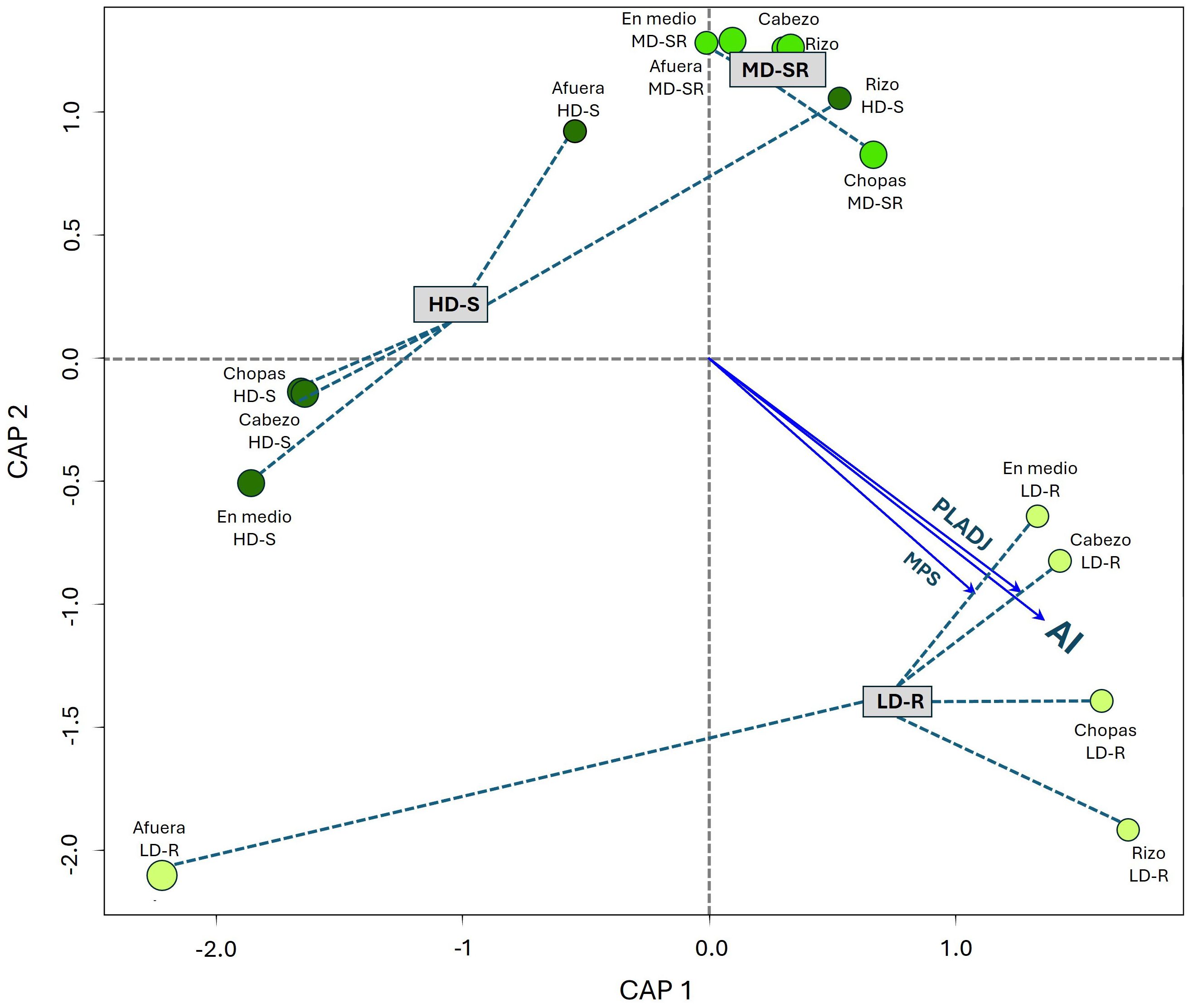
Figure 9. Redundancy Analysis (RDA) between top 5 centimeters of soil organic carbon (SOC5cm) stock (Mg C ha-1) and Landscape metrics in seagrass meadows of the south. LD-R: Low density over rocks; MD-SR: Medium density over sand-rock; HD-S: High density over sand.
Meanwhile, on Axis 2 (CAP2), the observed correlations were negative and insignificant concerning SOC5cm variation (Table 3). This indicates that variations in SOC5cm along this axis are less influenced by landscape configuration and more likely influenced by other factors not considered in this analysis.
The results reveal distinct seagrass landscape configurations in the VRSNP, with significant variations between the northern and southern systems. In the southern regions, larger windward and reef areas (5.8 ± 1.6 km; 841.7 ± 887.4 ha) and greater depths (1.3 ± 0.2 m) were observed, potentially creating more favorable conditions for the development and persistence of seagrass beds (Schultz et al., 2011; East et al., 2020); this may explain the presence of larger, more interconnected patches with a higher degree of aggregation (MPS: 0.19 ± 0.04 ha; AI: 89.6 ± 3.7%; PD: 0.16 ± 0.10 patches/100 ha) (Supplementary Tables 2, 3). Conversely, in the northern landscape’s environmental conditions, such as reef lagoons with reduced light availability (38.9 ± 12.3%), shallower depths (0.7 ± 0.1 cm) and a short windward (1.8 ± 0.8 km) appear to limit the development and persistence of seagrass beds (Patriquin et al., 2024), restricting to smaller lagoons (76 ± 30.7 ha). These factors seem to contribute to the lower extension and complexity of seagrass landscapes in this region (MPS: 805 ± 410 m²; AI: 88.8 ± 3.7%; PD: 0.16 ± 0.10 patches/100 ha).
According to van Der Heide et al. (2010), the structural complexity of seagrass landscapes is closely related to areas with low hydrodynamics, which influences the spatial distribution patterns of seagrasses in various ways. We observed landscapes with medium-density seagrass and sparse reef (MD-SR) at the northern systems; high-density seagrass (HD-S) are generally situated at greater average distances from the reef crest compared to low-density seagrass and reef landscapes (LD-R), which are located closer to the crest (average distance to reef crest for MD-SR: 212.5 ± 39 m; HD-S: 239.6 ± 58.3 m; LD-R: 203.5 ± 35.5 m). This increased distance from the reef crest may facilitate the development of more complex foliar structures in MD-SR (452.7 – 1,191.9 shoot m2) and HD-S (719.2 – 1,754.2 shoot m2) landscapes, which also exhibit higher levels of aggregation and patch density.
With regard to southern systems, the pattern is inverted; landscapes with higher foliar structures, such as HD-S (169.8 – 1,612.8 shoot m2) and MD-SR (282.9 – 1,499.6 shoot m2), are located closer to the reef crest (HD-S: 572.1 ± 84.6 m; MD-SR: 476.1 ± 67.5 m).This proximity may account for their more aggregated and interconnected configurations compared to LD-R landscapes, which are distributed farther from the reef crest (712.3 ± 139.8 m) and exhibit lower structural foliar density (346.6 - 905.4 shoot m2). The closer proximity of HD-S and MD-SR landscapes to the reef crest likely contributes to the higher structural complexity and greater heterogeneity observed in the southern seagrass landscapes (Supplementary Table 1), because a high shoot density enhances anchorage, thereby contributing to sediment stabilization and reducing the risk of meadow uprooting (Larkum et al., 2006; Badalamenti et al., 2015). Although the dominant hydrodynamic processes were not examined in this study, similar studies (Oreska et al., 2017; Ricart et al., 2017; Alemu et al., 2022) have reported that distance to the reef crest and soil composition influence the complexity of spatial patterns, as observed in the HD-S and MD-SR landscapes in both regions.
The distribution and extent of seagrasses in the VRSNP seems to be influenced by the characteristics of each reef system, including the linear length of the reef crest, depth, light incidence, and distance to the windward side (Supplementary Table 2). According to Lara et al. (1992) and Jones et al. (2008), southern reef systems are characterized by extensive, diverse, and high coral cover, indicating greater structural complexity. In contrast, northern systems comprise smaller, simpler, and less diverse reefs. These natural structural differences reflect the spatial configuration of seagrass landscapes identified in this study, with northern landscapes being more homogeneous and southern landscapes exhibiting greater complexity and diversity.
The configuration of seagrass landscapes, including the spatial arrangement and distribution of patches, plays a crucial role in regulating the flow of matter and energy between similar habitats. In the VRSNP, the larger and more complex rocky and coral areas in the southern region contribute to a more heterogeneous landscape compared to the northern region, where the reefs are smaller, simpler, and less diverse (Lara et al., 1992; Jones et al., 2008; Allende-Arandía et al., 2016). The increased complexity of southern reef systems provides enhanced protection for seagrass meadows, as evidenced by the larger and more expansive reef lagoons (Kirkman and Kirkman, 2000; Robbins and Bell, 2000; Arellano-Méndez et al., 2016).
In addition, there are natural stressors such as tropical storms, hurricanes, cold fronts, and red tides, as well as discharges from Jamapa, Papaloapan, and Antigua Rivers, that influence the seagrass landscape configuration in each region (Ortiz-Lozano et al., 2013); specifically in the north, anthropogenic activities, such as the operation of Veracruz Harbor, the presence of densely populated areas, and intense tourist activities (Vargas-Hernández et al., 1993; Horta-Puga et al., 2007; Valadez-Rocha and Ortiz-Lozano, 2013) could represent additional factors influencing the configuration of seagrass landscapes in this region. These combined natural and human factors may explain some of the observed differences between northern and southern systems, although further investigation would be required to determine the relative impact of each.
However, although anthropogenic factors were not directly measured in this study, it is possible that the northern systems are also exposed to such influences, which may contribute to the observed differences in landscape configuration. These factors could increase pressure on seagrass meadows and alter water quality and sedimentation patterns (Dahl et al., 2022). In the northern systems, reef lagoons are small and shallow (0.4 m to 1.6 m deep), with minimal light incidence in Gallega (17%); the southern systems, by contrast, are larger, with deeper waters (0.5 m to 2.5 m) and greater light incidence (38%). These physical and environmental differences may influence the carbon retention capacity of the different seagrass landscapes. The combination of natural and human-induced stressors in the northern systems may contribute to the reduced extent and complexity of seagrass landscapes compared to the more protected and expansive systems in the south, reinforcing the importance of considering both foliar structure and environmental context in evaluating SOC5cm stores.
The results of this study confirm that the type of seagrass landscape, in terms of foliar composition, significantly influences soil (SOC) and biomass (BOC) organic carbon reserves (Supplementary Table 5; Figure 5) despite the high variability observed across the ten lagoon reefs evaluated. HD-S landscapes exhibited the highest SOC (41.8–117.6 Mg C ha-1) and BOC (2.3–5.6 Mg C ha-1) concentration, while LD-R landscapes had the lowest (SOC= 19.2–86.2 Mg C ha-1 and BOC= 0.8–4.7 Mg C ha-1). However, the content and reserves of organic carbon associated with seagrasses are known to be highly variable (Fourqurean et al., 2012). Numerous studies have demonstrated that multiple factors, operating at different scales, influence this variability. These factors include substrate type, foliar structure of the meadow (Alongi et al., 2016), depth, hydrodynamics, landscape configuration, and geographic location (Sleeman et al., 2005; Watanabe and Kuwae, 2015; Samper-Villarreal et al., 2016; Gullström et al., 2018; Jiménez-Ramos et al., 2019; Mazarrasa et al., 2021). These studies, along with recent large-scale investigations (Serrano et al., 2016; Mazarrasa et al., 2018, 2021; Kennedy et al., 2022), highlight that variability in carbon accumulation in seagrass meadows is influenced by processes occurring across both global and landscape scales, including the specific environmental and geomorphological conditions observed in the VRSNP (Oreska et al., 2017; Kindeberg et al., 2018; Lima et al., 2020; Alemu et al., 2022).
Specifically, at a fine scale (each seagrass landscape type), substrate type, depth, and light incidence might limit the development of denser seagrass beds and reduce their capacity to retain carbon in the soil through canopy action (Waycott et al., 2009; Van Tussenbroek, 2011; Miyajima and Hamaguchi, 2019; Lewis et al., 2020). Additionally, soils composed of finer particles, such as sandy soils, generally have a greater ability to retain organic carbon compared to rocky soils, regardless of seagrass growth (Serrano et al., 2016; Mazarrasa et al., 2018). Similarly, in areas with reduced hydrodynamic activity and proximity to the reef crest, we observed that increased shoot density and foliar complexity enhance biomass carbon accumulation (BOC), suggesting that the biotic components of seagrass meadows play a key role in driving carbon storage, especially in shallow, sandy substrates. This is attributed to lower organic matter decomposition rates and higher sediment burial rates (Gacia and Duarte, 2001; Hendriks et al., 2008; Kennedy et al., 2010; Uhrin and Turner, 2018). Our results describe BOC values (HD-S of I. Sacrificio= 5.9 ± 1.2 Mg C ha-1) in seagrass meadows with higher structural complexity (range= 719.2 - 1754.2 shoot m2), predominantly occurring on sandy substrates and situated in areas shielded from wave-induced hydrodynamic stress (Figure 4; Supplementary Table 3).
This complex interaction of factors is further evidenced by the variation of soil organic carbon content (OC%) along a 25 cm profile in three distinct seagrass landscapes of the VRSNP. The HD-S landscape of the southern region and the MD-SR landscape of the northern region showed the highest carbon concentrations (6.3% and 6.1%, respectively) in the top 5-10 cm of soil, gradually declining to 4% at depths >15 cm. This pattern aligns with findings from reef-associated meadows where organic carbon accumulation is higher in the upper soil layers and decreases with depth (Stankovic et al., 2023). Such vertical profiles are characteristic of seagrass meadows where the carbon sequestration is most active near the surface, supported by the dense foliar canopy and low hydrodynamic conditions (Mazarrasa et al., 2018).
In contrast, LD-R landscapes in the northern and southern regions displayed a more uniform distribution of organic carbon content along the soil profile (4.6% to 5.1%), suggesting a stable environment with continuous organic matter input and preservation throughout the 25 cm profile. This linear pattern might indicate less disturbance and a more balanced carbon sequestration process (Kennedy et al., 2010). However, the HD-S landscape in the northern region exhibits an atypical pattern, with lower organic carbon content (4.7%) in the upper soil layers that increases with depth. This inverse distribution suggests that processes other than typical carbon sequestration dynamics, possibly including recent disturbances, have affected the upper layers, leading to a reorganization of carbon stocks or differential preservation processes in deeper soil layers (Gillis et al., 2014). This unusual pattern might indicate a history of natural or anthropogenic impacts that have disrupted the surface soil layers, causing a shift in where carbon is stored within the profile (Dahl et al., 2023).
At broader spatial scales, factors such as available colonization area, external carbon inputs, and wind- or storm-driven wave action have been reported to influence the morphology, extent, and dispersal patterns of seagrass meadow fragments (Vacchi et al., 2012; Oprandi et al., 2020), which in turn influenced the potential total carbon reserves in the reef system. This suggests that carbon variability across the 27 seagrass landscapes within the VRSNP is driven by both fine- and coarse-scale factors. The large size of the southern systems (Lara et al., 1992; Jones et al., 2008; DOF (Diario Oficial de la Federación), 2017) provides greater space and connectivity for seagrass growth, thereby enhancing carbon capacity in HD-S of Cabezo in the south and HD-S of I. Verde (Ca = 143.8 ha and BOC+SOC= 83.9 – 118.8 Mg C ha-1 and Ca = 1.7 ha and BOC+SOC= 110.5 – 132.1 Mg C ha-1, respectively), while the smaller extent of these landscapes in the northern systems reduces their capacity to retain carbon, as observed in HD-S of Gallega in the north region and HD-S of Rizo in the south (Ca = 5.2 ha and BOC+SOC= 62.9 – 79.8 Mg C ha-1 and Ca = 5.4 ha and BOC+SOC= 29.1 -57.6 Mg C ha-1, respectively). This suggests that, if the effects of coarse-scale factors are intensified, smaller and deeper seagrass landscapes may be more vulnerable to loss of seagrass cover and seagrass fragmentation. These findings suggest that if the influence of coarse-scale factors intensifies, smaller and deeper seagrass landscapes may become more susceptible to seagrass cover loss and fragmentation. However, the observed variation in carbon storage across seagrass landscapes was slightly greater than the range reported for Thalassia testudinum-dominated meadows in shallow reef lagoons (Van Tussenbroek, 2011; Guerra-Vargas et al., 2020).
Research has identified various factors operating at both fine and coarse scales that influence landscape composition and configuration, ultimately shaping seagrass meadow structure and, consequently, carbon storage within biomass (Waycott et al., 2009; Van Tussenbroek, 2011; Vacchi et al., 2012; Miyajima and Hamaguchi, 2019; Oprandi et al., 2020) In northern landscapes, our results reveal a limited direct correlation between variations in surface soil carbon (SOC5cm) and configuration-defining metrics. This indicates that the impact of edge density on soil carbon accumulation is not uniform but rather fluctuates based on the internal structure of the landscapes within each reef, with factors such as meadow size and spatial location playing a crucial role.
Landscape metrics related to perimeter and area have frequently been used to quantify different degrees of fragmentation in seagrass landscapes (Serrano et al., 2014; Wang et al., 2014). The loss of cover and habitat can lead to landscape transformation, influencing patterns and processes, including edge effects (Yarnall et al., 2022), which in turn may affect functions such as carbon storage capacity (Trevathan-Tackett et al., 2018). The results showed the landscapes HD-S, MD-SR in Gallega, and LD-R in Galleguilla have a lower storage of SOC5cm when edge density increases (550 - 800 m/ha). In contrast, landscapes with lower edge density (200 - 400 m/ha) such as HD-S in I. Sacrificios and MD-SR in I. Pajaros showed a greater storage of SOC5cm.
Ricart et al. (2015) observed reduced carbon accumulation at the edges of seagrass patches compared to their interior zones, which may explain the strong non-linear relationship between edge density and SOC5cm variation identified in this study; according to Granata et al. (2001) and Folkard (2005), this may be attributed to turbulence processes at the sand-seagrass interface, which increase sediment resuspension and transport of seagrass-derived materials from the meadow edges, thereby reducing the deposition of suspended particles.
Therefore, according to Fahrig (2017), the effects of fragmentation, as measured by edge density (ED), can be more pronounced in smaller landscapes due to the higher proportion of edge relative to the total area. This is consistent with our findings in northern landscapes, where the reduced size of the landscapes seems to exacerbate the negative effects of fragmentation on soil carbon accumulation (Bustamante et al., 2016). Edge effects are commonly addressed in monospecific meadows and on a small scale, where the patch area is smaller, and the effects of a gradient of ecological conditions spanning from the habitat-matrix interface to the center of the habitat patch are more evident (Ries et al., 2004; Ricart et al., 2015).
Numerous studies (Ries et al., 2004; Laurance, 2008; Haddad et al., 2015) have suggested that edge effects at the patch scale drive fragmentation dynamics at the landscape scale by increasing the number of patches with constant extension. Habitat loss and configuration change occur simultaneously, making it difficult to isolate the impact of each component (Long et al., 2010). This suggests that the high fragmentation (e.g., ED) could reduce carbon retention efficiency (e.g., SOC5cm) in these systems due to increased exposure of edges to erosive processes and reduced internal cohesion. Our results provide an initial approach to relating ecological processes, such as carbon storage in seagrasses, to landscape metrics.
On the other hand, we observed a large number of patches (227 fragments), predominantly small (226.9 m2) and with a high patch density (PD = 0.44), in the Gallega HD-S landscape, the most fragmented in the VRSNP. This is probably due to its proximity to the shipping channel of the most important commercial port, the Veracruz City, and tourist activities (Valadez-Rocha and Ortiz-Lozano, 2013; DOF (Diario Oficial de la Federación), 2017; Gil-Agudelo et al., 2020). Furthermore, Terrados and Ramírez-García (2011) observed low seagrass cover and high fragmentation in other lagoons of the VRSNP, attributed to increased exposure of the meadows to wave action and reef flattening due to human activities. Our results indicated that the northern HD-S landscapes retain less organic carbon in the surface strata (OC= 4.7% in <10 cm) compared to deeper layers (OC= 6% to 8% in 10 – 25 cm of depth soil). Additionally, the Gallega HD-S landscape exhibited the most fragmented seagrass meadows, with the lowest soil carbon storage (64 ± 3.3 Mg C ha-1). This suggests carbon loss and fragmentation, potentially linked to nearby anthropogenic disturbances, as reported by Valadez-Rocha and Ortiz-Lozano (2013).
On the other hand, landscapes metrics such as MPS, AI, and PLADJ were more significant in the south, where systems present a more heterogeneous configuration, reflecting that cohesion and internal connectivity favors SOC5cm accumulation. This suggests that connectivity and landscape configuration are key determinants of ecosystem functionality (Sweatman et al., 2017), which is reflected in soil carbon retention (Fahrig, 2003), as observed in the results obtained from southern landscapes. Understanding how the composition and configuration of seagrass landscapes affect variations in SOC5cm can provide valuable information for managing blue carbon ecosystems (Fahrig, 2017), particularly in scenarios with projected increases in the frequency, duration, and intensity of extreme events (Carlson et al., 2018; Correia and Smee, 2022) or in the context of ecosystem restoration (Ziegler et al., 2018; Olson et al., 2019).
This study highlights the influence of natural factors such as geomorphology, substrate type, depth, and hydrodynamics in shaping the configuration and composition of seagrass landscapes within the Veracruz Reef System National Park. These factors contribute to the variation in seagrass structure and distribution between the northern and southern regions, where three distinct landscape types (LD-R, MD-SR, and HD-S) were identified. Among these, the HD-S landscape, characterized by its higher shoot density and structural complexity, exhibited the greatest potential for carbon storage.
Although anthropogenic impacts were not directly measured in this study, previous research suggests fragmentation in northern landscapes were in part driven by increased human activity. Our results demonstrate that smaller northern landscapes with higher edge density stored less surface carbon. A greater proportion of edges exposed to erosive processes, such as sediment resuspension and the loss of organic matter, likely decreases the capacity of these ecosystems to retain surface carbon. In contrast, the more cohesive and interconnected seagrass meadows in the southern region, which experience fewer human pressures, showed greater efficiency in carbon storage. These findings underscore the critical need to conserve seagrass landscapes by preventing fragmentation and preserving their structural integrity to maximize their carbon sequestration potential.
The importance of protecting seagrasses extends beyond their role in carbon storage. As vital coastal ecosystems, seagrasses provide essential services such as supporting biodiversity, stabilizing shorelines, and improving water quality. Their conservation is crucial for maintaining these benefits and addressing climate change. Efforts to protect and restore seagrass ecosystems, especially in areas vulnerable to human impact, are essential to ensure they continue to play a key role in global climate adaptation and mitigation.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
JH-S: Writing – review & editing, Supervision, Resources, Project administration, Investigation, Funding acquisition, Conceptualization. EP-J: Writing – review & editing, Writing – original draft, Supervision, Formal Analysis, Data curation. JuM-M: Writing – review & editing, Writing – original draft, Software, Methodology, Formal analysis, Data curation. JoMM: Writing – review & editing, Supervision, Formal analysis, Data curation, Conceptualization. CTH: Writing – review & editing, Supervision, Formal Analysis, Data curation, Conceptualization.
The author(s) declare that financial support was received for the research and/or publication of this article. This research was conducted as part of the project titled 'Evaluation and Monitoring of Seagrasses in the PNSAV in the Context of the Expansion Project of the Port of Veracruz in the Northern Zone' (Contract No. API-GI-CS-62601-066-17).
We want to acknowledge and thank the technicians and researchers who participated in the planning and execution of this report. Special recognition goes to Isaac Ramírez and his group of collaborators at ARGOS Consultants for their administrative efforts and logistical support; Dr. Martín Bolaños and his group from Programa Mexicano del Carbono for the administration of the project; Comisión Nacional de Áreas Naturales Protegidas for the facilities granted for surveying the Parque Nacional Sistema Arrecifal Veracruzano; And collaborators of Cinvestav- for their support in interpreting the results. Special thanks go to Mr. José Luis Zapién, captain of a boat with register # 3002189813-8 from Antón Lizardo municipality, and his ship’s company. We are also deeply grateful to the reviewers for their insightful comments and contributions, which significantly enriched and improved this study.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1320194/full#supplementary-material
Alemu J. B., Yaakub S. M., Yando E. S., San Lau R. Y., Lim C. C., Puah J. Y., et al. (2022). Geomorphic gradients in shallow seagrass carbon stocks. Estuarine Coast. Shelf Sci. 265, 107681.
Allende-Arandía M. E., Zavala-Hidalgo J., Romero-Centeno R., Mateos-Jasso A., Vargas-Hernández J. M., Zamudio L. (2016). Analysis of ocean current observations in the northern veracruz coral reef system, Mexico: 2007–12. J. Coast. Res. 317, 46–55. doi: 10.2112/jcoastres-d-14-00148.1
Alongi D. M., Murdiyarso D., Fourqurean J. W., Kauffman J. B., Hutahaean A., Crooks S., et al. (2016). Indonesia’s blue carbon: a globally significant and vulnerable sink for seagrass and mangrove carbon. Wetlands Ecol. Manage. 24, 3–13. doi: 10.1007/s11273-015-9446-y
Arellano-Méndez L., Bello-Pineda J., Aké-Castillo J. A., Pérez-España H., Martinez-Cardenas L. (2016). Spatial distribution and ecological structure of Thalassia testudinum (Hydrocharitaceae) in two reefs of Veracruz Reef System National Park, Mexico. Rev. Biol. Trop. 64, 427–448. doi: 10.15517/rbt.v64i2.19810
Avendaño-Alvarez O., Salas-Monreal D., Marin-Hernandez M., Salas-de-Leon D. A., Monreal-Gomez M. A. (2017). Annual hydrological variation and hypoxic zone in a tropical coral reef system. Region. Stud. Mar. Sci. 9, 145–155. doi: 10.1016/j.rsma.2016.12.007
Badalamenti F., Alagna A., Fici S. (2015). Evidences of adaptive traits to rocky substrates undermine paradigm of habitat preference of the Mediterranean seagrass Posidonia oceanica. Sci. Rep. 5, 8804. doi: 10.1038/srep08804
Bello-Pineda J., Reyna-González P., Aguilera-Arias J., Arellano-Méndez y H. Pérez-España L. (2015). “Incorporando análisis espacial para la toma de decisiones en el Parque Nacional Sistema Arrecifal Veracruzano,” in Aportes al conocimiento del Sistema Arrecifal Veracruzano: hacia el Corredor Arrecifal del Suroeste del Golfo de México. Eds. Granados-Barba A., Ortiz-Lozano L. D., Salas-Monreal D., González-Gándara C. (Universidad Autónoma de Campeche, Mexico), 281–300, ISBN: ISBN 978-607-8444-11-3.
Berk A., Anderson G. P., Acharaya P. K., Hoke M. L., Chetwynd J. H., Bernstein L. S., et al. (2003). MODTRAN 4 Version 3 Revision 1 User’s Manual. 2003 (Washington, DC, USA: Air Force Research Laboratory Naval Research Laboratory Remote Sensing Division).
Boström C., Pittman S. J., Simenstad C., Kneib R. T. (2011). Seascape ecology of coastal biogenic habitats: advances, gaps, and challenges. Mar. Ecol. Prog. Ser. 427, 191–217. doi: 10.3354/meps09051
Bustamante M. M., Roitman I., Aide T. M., Alencar A., Anderson L. O., Aragão L., et al. (2016). Toward an integrated monitoring framework to assess the effects of tropical forest degradation and recovery on carbon stocks and biodiversity. Global Change Biol. 22, 92–109. doi: 10.1111/gcb.13087
Canty B., Ripley B. D. (2019). Boot: bootstrap R (S-Plus) functions. R package version. 1.3–26. Available online at: https://www.r-project.org/ (Accessed January 28, 2023).
CARICOMP, C. C. M. P (2001). CARICOMP Methods Manual Levels 1 and 2: Methods for Mapping and Monitoring of Physical and Biological Parameters in the Coastal Zone of the Caribbean (Kingston, Jamaica: Centre for Marine Sciences, University of the West Indies).
Carlson D. F., Yarbro L. A., Scolaro S., Poniatowski M., McGee-Absten V., Carlson P. R. Jr. (2018). Sea surface temperatures and seagrass mortality in Florida Bay: spatial and temporal patterns discerned from MODIS and AVHRR data. Remote Sens. Environ. 208, 171–188. doi: 10.1016/j.rse.2018.02.014
Castañeda-Chávez M., Lango-Reynoso F. (2021). “Impact of the jamapa river basin on the gulf of Mexico,” in River deltas research. Recent advances. Ed. Manning A. J. (IntechOpen, London), 247–263.
Chernick M. R., LaBudde R. A. (2014). An introduction to bootstrap methods with applications to R (New Jersey, USA: John Wiley & Sons).
Clarke K. R., Tweedley J. R., Valesini F. J. (2014). Simple shade plots aid better long-term choices of data pre-treatment in multivariate assemblage studies. J. Mar. Biol. Assoc. United Kingdom 94, 1–16. doi: 10.1017/S0025315413001227
Coffer M. M., Schaeffer B. A., Zimmerman R. C., Hill V., Li J., Islam K. A., et al. (2020). Performance across WorldView-2 and RapidEye for reproducible seagrass mapping. Remote Sens. Environ. 250, 112036. doi: 10.1016/j.rse.2020.112036
Correia K. M., Smee D. L. (2022). A meta-analysis of tropical cyclone effects on seagrass meadows. Wetlands 42, 108. doi: 10.1007/s13157-022-01611-0
Costanza R., De Groot R., Sutton P., van der Ploeg S., Anderson S. J., Kubiszewski I., et al. (2014). Changes in the global value of ecosystem services. Global Environ. Change 26, 152–158. doi: 10.1016/j.gloenvcha.2014.04.002
Cuevas E., Uribe-Martínez A., Morales-Ojeda S. M., Gómez-Ruíz P. A., Núñez-Lara E., Teutli-Hernández C., et al. (2021). Spatial configuration of seagrass community attributes in a stressed coastal lagoon, southeastern Gulf of Mexico. Region. Stud. Mar. Sci. 48, 102049. doi: 10.1016/j.rsma.2021.102049
Dahl M., Ismail R., Braun S., Masqué P., Lavery P. S., Gullström M., et al. (2022). Impacts of land-use change and urban development on carbon sequestration in tropical seagrass meadow sediments. Mar. Environ. Res. 176, 105608. doi: 10.1016/j.marenvres.2022.105608
Dahl M., McMahon K., Lavery P. S., Hamilton S. H., Lovelock C. E., Serrano O. (2023). Ranking the risk of CO2 emissions from seagrass soil carbon stocks under global change threats. Global Environ. Change 78, 102632. doi: 10.1016/j.gloenvcha.2022.102632
Davidson C., Finlayson M. (2019). Updating global coastal wetland areas presented in Davidson and Finlayson, (2018). Mar. Freshw. Res. 70, 1195–1200. doi: 10.1071/MF19010
Davison A. C., Hinkley D. V. (1997). Bootstrap methods and their application (No. 1). Cambridge university press. Diario Oficial de la Federación DOF., (2017). Acuerdo por el que se da a conocer el resumen del Programa de Manejo del Área Natural Protegida con categoría de Parque Nacional la zona conocida como Sistema Arrecifal Veracruzano (Mexico City, Mexico: CONANP).
Digital-Globe, I. (2017). Digital globe core imagery products guide. 2 p. Available online at: https://resources.maxar.com/data-sheets/worldview-2 (Accessed October 28, 2024).
DOF (Diario Oficial de la Federación). (2017). Acuerdo por el que se da a conocer el resumen del Programa de Manejo del Área Natural Protegida con categoría de Parque Nacional la zona conocida como Sistema Arrecifal Veracruzano. Mexico City, Mexico: CONANP. Available online at: https://dof.gob.mx/nota_detalle.php?codigo=5483733&fecha=22/05/2017#gsc.tab=0 (Accessed October 28, 2024).
Duarte C. M., Losada I. J., Hendriks I. E., Mazarrasa I., Marbà N. (2013). The role of coastal plant communities for climate change mitigation and adaptation. Nat. Climate Change 3, 961–968. doi: 10.1038/nclimate1970
Duarte C. M., Marbà N., Gacia E., Fourqurean J. W., Beggins J., Barrón C., et al. (2010). Seagrass community metabolism: Assessing the carbon sink capacity of seagrass meadows. Global biogeochem. cycles 24. doi: 10.1029/2010GB003793
East H. K., Perry C. T., Beetham E. P., Kench P. S., Liang Y. (2020). Modelling reef hydrodynamics and sediment mobility under sea level rise in atoll reef island systems. Global Planet. Change 192, 103196. doi: 10.1016/j.gloplacha.2020.103196
Fahrig L. (2003). Effects of habitat fragmentation on biodiversity. Annu. Rev. ecology evolution systematics 34 (1), 487–515. doi: 10.1146/annurev.ecolsys.34.011802.132419
Fahrig L. (2017). Ecological responses to habitat fragmentation per se. Annu. Rev. ecol. evol. system. 48, 1–23. doi: 10.1146/annurev-ecolsys-110316-022612
Folkard A. M. (2005). Hydrodynamics of model Posidonia oceanica patches in shallow water. Limnol. oceanogr. 50, 1592–1600. doi: 10.4319/lo.2005.50.5.1592
Fourqurean J. W., Duarte C. M., Kennedy H., Marbà N., Holmer M., Mateo M. A., et al. (2012). Seagrass ecosystems as a globally significant carbon stock. Nat. Geosci. 5, 505–509. doi: 10.1038/NGEO1477
Gacia E., Duarte C. M. (2001). Sediment retention by a mediterranean posidonia oceanica meadow: the balance between deposition and resuspension. Estuarine Coast. shelf Sci. 52, 505–514. doi: 10.1006/ecss.2000.0753
Gil-Agudelo D. L., Cintra-Buenrostro C. E., Brenner J., González-Díaz P., Kiene W., Lustic C., et al. (2020). Coral reefs in the Gulf of Mexico large marine ecosystem: conservation status, challenges, and opportunities. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00807
Gillis L. G., Bouma T. J., Jones C. G., Van Katwijk M. M., Nagelkerken I., Jeuken C. J. L., et al. (2014). Potential for landscape-scale positive interactions among tropical marine ecosystems. Mar. Ecol. Prog. Ser. 503, 289–303. doi: 10.3354/meps10716
Granata T. C., Serra T., Colomer J., Casamitjana X., Duarte C. M., Gacia E. (2001). Flow and particle distributions in a nearshore seagrass meadow before and after a storm. Mar. Ecol. Prog. Ser. 218, 95–106. doi: 10.3354/meps218095
Grober-Dunsmore R., Pittman S. J., Caldow C., Kendall M. S., Frazer T. K. (2009). “A Landscape Ecology Approach for the Study of Ecological Connectivity Across Tropical Marine Seascapes,” in Ecological Connectivity among Tropical Coastal Ecosystems. Ed. Nagelkerken I. (Springer Netherlands, Dordrecht), 493–530. doi: 10.1007/978-90-481-2406-0_14
Guerra-Vargas L. A., Gillis L. G., Mancera-Pineda J. E. (2020). Stronger together: do coral reefs enhance seagrass meadows “blue carbon” potential? Front. Mar. Sci. 628. doi: 10.3389/fmars.2020.00628
Gullström M., Lyimo L. D., Dahl M., Samuelsson G. S., Eggertsen M., Anderberg E., et al. (2018). Blue carbon storage in tropical seagrass meadows relates to carbonate stock dynamics, plant–sediment processes, and landscape context: insights from the western Indian Ocean. Ecosystems 21, 551–566. doi: 10.1007/s10021-017-0170-8
Haddad N. M., Brudvig L. A., Clobert J., Davies K. F., Gonzalez A., Holt R. D., et al. (2015). Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 1, e1500052. doi: 10.1126/sciadv.1500052
Harris D., Horwáth W. R., Van Kessel C. (2001). Acid fumigation of soils to remove carbonates prior to total organic carbon or carbon-13 isotopic analysis. Soil Sci. Soc. America J. 65, 1853–1856. doi: 10.2136/sssaj2001.1853
Heck K. L., Carruthers T. J., Duarte C. M., Hughes A. R., Kendrick G., Orth R. J., et al. (2008). Trophic transfers from seagrass meadows subsidize diverse marine and terrestrial consumers. Ecosystems 11, 1198–1210. doi: 10.1007/s10021-008-9155-y
Hendriks I. E., Sintes T., Bouma T. J., Duarte C. M. (2008). Experimental assessment and modeling evaluation of the effects of the seagrass Posidonia oceanica on flow and particle trapping. Mar. Ecol. Prog. Ser. 356, 163–173. doi: 10.3354/meps07316
Hill J., Wilkinson C. L. I. V. E. (2004). Methods for ecological monitoring of coral reefs (Townsville, Australia: Australian Institute of Marine Science), 117.
Holmquist J. R., Windham-Myers L., Bliss N., Crooks S., Morris J. T., Megonigal J. P., et al. (2018). Accuracy and precision of tidal wetland soil carbon mapping in the conterminous United States. Sci. Rep. 8, 9478. doi: 10.1038/s41598-018-26948-7
Horta-Puga G., Vargas-Hernández J. M., Carricart-Ganivet J. P. (2007). “Reef corals,” in Coral reefs of the southern gulf of mexico. Eds. Tunnell J. W. Jr., Chávez E. A., Withers K. (College Station, TX: Texas A&M University Press), 95–101.
Horta-Puga G., Cházaro-Olvera S., Winfield I., Lozano-Aburto M. A., Arenas-Fuentes V. (2016). Heavy metals in macroalgae from the Veracruz Reef System, Southern Gulf of Mexico Metales pesados en macroalgas del Sistema Arrecifal Veracruzano, sur del Golfo de México. Rev. Bio Cienc. 3, 326–339. doi: 10.15741/revbio.03.01.07
Horta-Puga G., Morales-Aranda A. A., Tello-Musi J. L. (2022). The reef fish assemblage of a coral reef system in the southwestern Gulf of Mexico. Rev. Mexicana Biodiversidad 93, 11. doi: 10.22201/ib.20078706e.2022.93.4156
Howard J. L., Creed J. C., Aguiar M. V., Fourqurean J. W. (2018). CO2 released by carbonate sediment production in some coastal areas may offset the benefits of seagrass “Blue Carbon” storage. Limnol. Oceanogr. 63, 160–172. doi: 10.1002/lno.10621
Howard J., Hoyt S., Isensee K., Telszewski M., Pidgeon E. (2014). Coastal blue carbon: methods for assessing carbon stocks and emissions factors in mangroves, tidal salt marshes, and seagrasses (Arlington, Virginia, USA: Conservation International, Intergovernmental Oceanographic Commission of UNESCO, International Union for Conservation of Nature).
Hyndes G. A., Nagelkerken I., McLeod R. J., Connolly R. M., Lavery P. S., Vanderklift M. A. (2014). Mechanisms and ecological role of carbon transfer within coastal seascapes. Biol. Rev. 89, 232–254. doi: 10.1111/brv.12055
Jiménez-Ramos R., Egea L. G., Vergara J. J., Bouma T. J., Brun F. G. (2019). The role of flow velocity combined with habitat complexity as a top–down regulator in seagrass meadows. Oikos 128, 64–76. doi: 10.1111/oik.05452
Jones J., Withers K., Tunnell J. W. (2008). “Comparison of benthic communities on six coral reefs in the Veracruz Reef System (Mexico),” in Proceedings of the 11th International Coral Reef Symposium. 757–760.
Jung M. L. (2016). A python plugin for automated landscape ecology analysis. Ecol. Inform 31, 18–21. doi: 10.1016/j.ecoinf.2015.11.006
Kennedy H., Beggins J., Duarte C. M., Fourqurean J. W., Holmer M., Marbà N., et al. (2010). Seagrass sediments as a global carbon sink: Isotopic constraints. Global biogeochem. cycles 24. doi: 10.1029/2010GB003848
Kennedy H., Pagès J. F., Lagomasino D., Arias-Ortiz A., Colarusso P., Fourqurean J. W., et al. (2022). Species traits and geomorphic setting as drivers of global soil carbon stocks in seagrass meadows. Global Biogeochem. Cycles 36, e2022GB007481. doi: 10.1029/2022gb007481
Kindeberg T., Ørberg S. B., Röhr M. E., Holmer M., Krause-Jensen D. (2018). Sediment stocks of carbon, nitrogen, and phosphorus in Danish eelgrass meadows. Front. Mar. Sci. 5. doi: 10.3389/fmars.2018.00474
Kirkman H., Kirkman J. (2000). Long-term seagrass meadow monitoring near Perth, Western Australia. Aquat. Bot. 67, 319–332. doi: 10.1016/S0304-3770(00)00097-8
Kupfer J. A. (2012). Landscape ecology and biogeography: rethinking landscape metrics in a post-FRAGSTATS landscape. Prog. Phys. Geogr. 36 (3), 400–420. doi: 10.1177/0309133312439594
Lara M., Padilla C., García C., Espejel J. J. (1992). “Coral reef of Veracruz Mexico I. Zonation and community,” in Proceedings of the 7th International Coral Reef Symposium. 535–544.
Larkum A. W., Orth R. J., Duarte C. M. (2006). Seagrasses: biology, ecology and conservation. Phycologia 45, 5.
Laurance W. F. (2008). Theory meets reality: How habitat fragmentation research has transcended island biogeographic theory. Biol. Conserv. 141, 1731–1744. doi: 10.1016/j.biocon.2008.05.011
Legendre P., Anderson M. J. (1999). Distance-based redundancy analysis: testing multispecies responses in multifactorial ecological experiments. Ecol. Monogr. 69 (1), 1–24.
Legendre P., De Cáceres M. (2013). Beta diversity as the variance of community data: dissimilarity coefficients and partitioning. Ecol. letters 16 (8), 951–963. doi: 10.1111/ele.12141
Lewis C. J., Young M. A., Ierodiaconou D., Baldock J. A., Hawke B., Sanderman, et al. (2020). Drivers and modelling of blue carbon stock variability in sediments of southeastern Australia. Biogeosciences 17, 2041–2059. doi: 10.5194/bg-2019-294
Liaño-Carrera F., Camarena-Luhrs T., Gómez-Barrero A., Martos-Fernández F. J., Ramírez-Macias J. I., Salas-Monreal D. (2019). New coral reef structures in a tropical coral reef system. Lat Amer. J. Aqua. Res. 47, 270–281. doi: 10.3856/vol47-issue2-fulltext-7
Lillesand T. M., Kiefer R. W., Chipman J. W. (1995). Remote sensing and image interpretation (United States of America: John Willey & Sons. Inc).
Lima M. D. A. C., Ward R. D., Joyce C. B. (2020). Environmental drivers of sediment carbon storage in temperate seagrass meadows. Hydrobiologia 847, 1773–1792. doi: 10.1007/s10750-019-04153-5
Long J. A., Nelson T. A., Wulder M. A. (2010). Characterizing forest fragmentation: Distinguishing change in composition from configuration. Appl. Geogr. 30, 426–435. doi: 10.1016/j.apgeog.2009.12.002
MacArthur R. H., Wilson E. O. (2001). The theory of island biogeography Vol. 1 (New Jersey, USA: Princeton university press).
Macreadie P. I., Hindell J. S., Jenkins G. P., Connolly R. M., Keough M. J. (2009). Fish responses to experimental fragmentation of seagrass habitat. Conserv. Biol. 23, 644–652. doi: 10.1111/j.1523-1739.2008.01130.x
Mapel-Hernández M. D., Armstrong-Altrin J. S., Botello A. V., Lango-Reynoso F. (2021). Bioavailability of cd and pb in sediments of the national Park Veracruz reef system, gulf of Mexico. Appl. Geochem. 133, 105085. doi: 10.1016/j.apgeochem.2021.105085
Maritorena S., Morel A., Gentili B. (1994). Diffuse reflectance of oceanic shallow waters: Influence of water depth and bottom albedo. Limnol. oceanogr. 39, 1689–1703. doi: 10.4319/lo.1994.39.7.1689
Mazarrasa I., Lavery P. S., Duarte C. M., Lafratta A., Lovelock C. E., Macreadie P. I., et al. (2021). Factors determining seagrass blue carbon across bioregions and geomorphologies. Global Biogeochem. Cycles 35, 1–17. doi: 10.1029/2021gb006935
Mazarrasa I., Samper-Villarreal J., Serrano O., Lavery P. S., Lovelock C. E., Marbà N., et al. (2018). Habitat characteristics provide insights of carbon storage in seagrass meadows. Mar. pollut. Bull. 134, 106–117. doi: 10.1016/j.marpolbul.2018.01.059
Mcleod E., Chmura G. L., Bouillon S., Salm R., Björk M., Duarte C. M., et al. (2011). A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Front. Ecol. Environ. 9, 552–560. doi: 10.1890/110004
Mellbrand K., Lavery P. S., Hyndes G., Hambäck P. A. (2011). Linking land and sea: different pathways for marine subsidies. Ecosystems 14, 732–744. doi: 10.1007/s10021-011-9442-x
Milner C., Hughes R. E. (1968). Methods for the measurement of the primary productivity of grassland (Oxford: IBP Handbook No. 6 Blackwell Scientific Publications).
Miyajima T., Hamaguchi M. (2019). “Carbon sequestration in sediment as an ecosystem function of seagrass meadows,” in Blue Carbon in Shallow Coastal Ecosystems: Carbon Dynamics, Policy, and Implementation, 33–71. doi: 10.1007/978-981-13-1295-3_2
Montefalcone M., Parravicini V., Vacchi M., Albertelli G., Ferrari M., Morri C., et al. (2010). Human influence on seagrass habitat fragmentation in NW Mediterranean Sea. Estuarine Coast. Shelf Sci. 86, 292–298. doi: 10.1016/j.ecss.2009.11.018
Nagelkerken I. (2009). “Evaluation of nursery function of mangroves and seagrass beds for tropical decapods and reef fishes: patterns and underlying mechanisms,” in Ecological Connectivity among Tropical Coastal Ecosystems. Ed. Nagelkerken I. (Springer Netherlands, Dordrecht), 357–399. doi: 10.1007/978-90-481-2406-0_10
Nordlund L. M., Koch E. W., Barbier E. B., Creed J. C. (2016). Seagrass ecosystem services and their variability across genera and geographical regions. PloS One 11, e0163091. doi: 10.1371/journal.pone.0163091
Nordlund L. M., Unsworth R. K., Gullström M., Cullen-Unsworth L. C. (2018). Global significance of seagrass fishery activity. Fish Fish. 19, 399–412. doi: 10.1111/faf.12259
Oksanen J., Blanchet F. G., Friendly M., Kindt R., Legendre P., McGlinn D., et al. (2020). vegan: Community Ecology Package. R package version 2.5–7.
Olson A. M., Hessing-Lewis M., Haggarty D., Juanes F. (2019). Nearshore seascape connectivity enhances seagrass meadow nursery function. Ecol. Appl. 29, e01897. doi: 10.1002/eap.1897
Oprandi A., Mucerino L., De Leo F., Bianchi C. N., Morri C., Azzola A., et al. (2020). Effects of a severe storm on seagrass meadows. Sci. Total Environ. 748, 141373. doi: 10.1016/j.scitotenv.2020.141373
Oreska M. P., McGlathery K. J., Porter J. H. (2017). Seagrass blue carbon spatial patterns at the meadow-scale. PloS One 12, e0176630. doi: 10.1371/journal.pone.0176630
Ortiz-Lozano L., Gutiérrez-Velázquez A., Aja-Arteaga A., Argüelles-Jiménez J., Ramos-Castillo V. (2021). Distribution, threats, and management of submerged reefs in the north of the reef corridor of the Southwest Gulf of Mexico. Ocean Coast. Manage. 201, 105428. doi: 10.1016/j.ocecoaman.2020.105428
Ortiz-Lozano L., Pérez-España H., Granados-Barba A., González-Gándara C., Gutiérrez-Velázquez A., Martos J. (2013). The Reef Corridor of the Southwest Gulf of Mexico: Challenges for its management and conservation. Ocean Coast. Manage. 86, 22–32. doi: 10.1016/j.ocecoaman.2013.10.006
Palafox-Juárez E. B., de los Ángeles Liceaga-Correa M. (2017). Spatial diversity of a coastal seascape: Characterization, analysis and application for conservation. Ocean Coast. Manage. 136, 185–195. doi: 10.1016/j.ocecoaman.2016.12.002
Patriquin D., Scheibling R. E., Filbee-Dexter K. (2024). Shifts in biodiversity and physical structure of seagrass beds across 5 decades at Carriacou, Grenadines. PloS One 19, e0306897. doi: 10.1371/journal.pone.0306897
Pierrejean M., Cusson M., Rossi F. (2023). Seasonal effects of edge and habitat complexity on eelgrass epifaunal assemblages. Mar. Ecol. Prog. Ser. 718, 39–52. doi: 10.3354/meps14385
Pittman S., Kneib R., Simenstad C., Nagelkerken I. (2011). Seascape ecology: application of landscape ecology to the marine environment. Mar. Ecol. Prog. Ser. 427, 187–302.
Quinn G. P., Keough M. J. (2002). Experimental design and data analysis for biologists (New York, USA: Cambridge university press), 527p.
Ramírez-García P., Pedraza-Venegas K. (2019). “Los Pastos Marinos y la Captura de Carbono Azul una Razón para Estudiarlos en el CASGM,” in Estudios Científicos en el Corredor Arrecifal del Suroeste del Golfo de México, 281.
R Core Team (2018). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.r-project.org.
R Core Team (2023). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.r-project.org.
Ricart A. M., Dalmau A., Pérez M., Romero J. (2015). Effects of landscape configuration on the exchange of materials in seagrass ecosystems. Mar. Ecol. Prog. Ser. 532, 89–100. doi: 10.3354/meps11384
Ricart A. M., Pérez M., Romero J. (2017). Landscape configuration modulates carbon storage in seagrass sediments. Estuarine Coast. Shelf Sci. 185, 69–76. doi: 10.1016/j.ecss.2016.12.011
Ries L., Fletcher R. J. Jr., Battin J., Sisk T. D. (2004). Ecological responses to habitat edges: mechanisms, models, and variability explained. Annu. Rev. Ecol. Evol. Syst. 35, 491–522. doi: 10.1146/annurev.ecolsys.35.112202.130148
Robbins B. D., Bell S. S. (2000). Dynamics of a subtidal seagrass landscape: Seasonal and annual change in relation to water depth. Ecology 81, 1193–1205. doi: 10.1890/0012-9658(2000)081[1193:doassl]2.0.co;2
Salas-Monreal D., Monreal-Jimenez R., Contreras-Tereza V. K., Monreal-Gomez M. A., Salas-de-Leon D. A., Riveron-Enzastiga M. L. (2022). Hydrographic variation in a tropical coral reef system: The Veracruz Reef System, Gulf of Mexico. Oceanologia 64, 473–488. doi: 10.1016/j.oceano.2022.03.002
Santos R., Duque-Núñez N., Santos C. B. D. L., Martins M., Veiga-Pires C. (2019). Superficial sedimentary carbon and nitrogen stocks in coastal vegetated assemblages along a flow velocity gradient. Sci. Rep. 9 (1), 610. doi: 10.1038/s41598-018-37031-6
Samper-Villarreal J., Lovelock C. E., Saunders M. I., Roelfsema C., Mumby P. J. (2016). Organic carbon in seagrass sediments is influenced by seagrass canopy complexity, turbidity, wave height, and water depth: Drivers of seagrass carbon. Limnol. oceanogr. 61, 938–952. doi: 10.1002/lno.10262
Schultz S. T., Kruschel C., Mokos M. (2011). Boat-based videographic monitoring of an Adriatic lagoon indicates increase in seagrass cover associated with sediment deposition. Aquat. Bot. 95, 117–123. doi: 10.1016/j.aquabot.2011.04.004
Serrano O., Mateo M. A., Renom P., Julià R. (2012). Characterization of soils beneath a posidonia oceanica meadow. Geoderma 185, 26–36. doi: 10.1016/j.geoderma.2012.03.020
Serrano O., Lavery P. S., Duarte C. M., Kendrick G. A., Calafat A., York P. H., et al. (2016). Can mud (silt and clay) concentration be used to predict soil organic carbon content within seagrass ecosystems? Biogeosciences 13, 4915–4926. doi: 10.5194/bg-13-4915-2016
Serrano O., Lavery P. S., Rozaimi M., Mateo M. Á. (2014). Influence of water depth on the carbon sequestration capacity of seagrasses. Global Biogeochem. Cycles 28, 950–961. doi: 10.1002/2014gb004872
Short F., Carruthers T., Dennison W., Waycott M. (2007). Global seagrass distribution and diversity: A bioregional model. J. Exp. Mar. Biol. Ecol. 350, 3–20. doi: 10.1016/j.jembe.2007.06.012
Short F. T., Polidoro B., Livingstone S. R., Carpenter K. E., Bandeira S., Bujang J. S., et al. (2011). Extinction risk assessment of the world’s seagrass species. Biol. Conserv. 144, 1961–1971.
Singh S. K., Pandey A. C., Singh D. (2014). Land use fragmentation analysis using remote sensing and Fragstats. Remote Sens. Appl. Environ. Res., 151–176. doi: 10.1007/978-3-319-05906-8_9
Sleeman J. C., Kendrick G. A., Boggs G. S., Hegge B. J. (2005). Measuring fragmentation of seagrass landscapes: which indices are most appropriate for detecting change? Mar. Freshw. Res. 56, 851–864. doi: 10.1071/MF04300
Spivak A. C., Sanderman J., Bowen J. L., Canuel E. A., Hopkinson C. S. (2019). Global-change controls on soil-carbon accumulation and loss in coastal vegetated ecosystems. Nat. Geosci. 12, 685–692. doi: 10.1038/s41561-019-0435-2
Stankovic M., Panyawai J., Khanthasimachalerm N., Prathep A. (2023). National assessment and variability of blue carbon in seagrass ecosystems in Thailand. Mar. pollut. Bull. 197, 115708. doi: 10.1016/j.marpolbul.2023.115708
Stumpf R. P., Holderied K., Robinson J. A., Feldman G., Kuring N. (2003). “Mapping water depths in clear water from space,” in Proceedings of the 13th Biennial Coastal Zone Conference, Vol. 5.
Su L., Huang Y. (2019). Seagrass resource assessment using WorldView-2 imagery in the Redfish Bay, Texas. J. Mar. Sci. Eng. 7, 98. doi: 10.3390/jmse7040098
Sweatman J. L., Layman C. A., Fourqurean J. W. (2017). Habitat fragmentation has some impacts on aspects of ecosystem functioning in a sub-tropical seagrass bed. Mar. Environ. Res. 126, 95–108. doi: 10.1016/j.marenvres.2017.02.003
Tanaya T., Watanabe K., Yamamoto S., Hongo C., Kayanne H., Kuwae T. (2018). Contributions of the direct supply of belowground seagrass detritus and trapping of suspended organic matter to the sedimentary organic carbon stock in seagrass meadows. Biogeosciences 15 (13), 4033–4045. doi: 10.5194/bg-15-4033-2018
Terrados J., Ramírez-García P., Hernández-Martínez Ó., Pedraza K., Quiroz A. (2008). State of Thalassia testudinum Banks ex König meadows in the veracruz Reef System, veracruz, Mexico. Aquat. Bot. 88, 17–26. doi: 10.1016/j.aquabot.2007.08.003
Terrados J., Ramírez-García P. (2011). Cover and edge length to area ratio of seagrass (Thalassia testudinum) meadows in coral reef lagoons (Veracruz reef system, southwest gulf of méxico). Aquat. Conservation: Mar. Freshw. Ecosyst. 21 (3), 224–230. doi: 10.1002/aqc.1188
Trevathan-Tackett S. M., Wessel C., Cebrián J., Ralph P. J., Masqué P., Macreadie P. I. (2018). Effects of small-scale, shading-induced seagrass loss on blue carbon storage: Implications for management of degraded seagrass ecosystems. J. Appl. Ecol. 55, 1351–1359. doi: 10.1111/1365-2664.13081
Uhrin A. V., Turner M. G. (2018). Physical drivers of seagrass spatial configuration: the role of thresholds. Landscape Ecol. 33, 2253–2272. doi: 10.1007/s10980-018-0739-4
Vacchi M., Montefalcone M., Bianchi C. N., Morri C., Ferrari M. (2012). Hydrodynamic constraints to the seaward development of Posidonia oceanica meadows. Estuarine Coast. Shelf Sci. 97, 58–65. doi: 10.1016/j.ecss.2011.11.024
Valadez-Rocha V., Ortiz-Lozano L. (2013). Spatial and temporal effects of port facilities expansion on the surface area of shallow coral reefs. Environ. Manage. 52, 250–260. doi: 10.1007/s00267-013-0098-5
van Der Heide T., Bouma T. J., Van Nes E. H., Van De Koppel J., Scheffer M., Roelofs J. G., et al. (2010). Spatial self-organized patterning in seagrasses along a depth gradient of an intertidal ecosystem. Ecology 91, 362–369. doi: 10.1890/08-1567.1
Van Tussenbroek B. I. (2011). Dynamics of seagrasses and associated algae in coral reef lagoons. Hidrobiológica 21, 293–310.
Vargas-Hernández J. M., Hernández-Gutiérrez A., Carrera-Parra L. F. (1993). “(Eds.),” in Sistema arrecifal veracruzano. Eds. Salazar-Vallejo S. I., González N. E. (CONABIO, Centro de Investigaciones de Quintana Roo, Chetumal: Biodiversidad marina y costera de México), 559–575.
Wang X., Blanchet F. G., Koper N. (2014). Measuring habitat fragmentation: An evaluation of landscape pattern metrics. Methods Ecol. Evol. 5, 634–646. doi: 10.1111/2041-210X.12198
Watanabe K., Kuwae T. (2015). How organic carbon derived from multiple sources contributes to carbon sequestration processes in a shallow coastal system? Global Change Biol. 21, 2612–2623. doi: 10.1111/gcb.12924
Waycott M., Duarte C. M., Carruthers T. J., Orth R. J., Dennison W. C., Olyarnik S., et al. (2009). Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. 106, 12377–12381. doi: 10.1073/pnas.0905620106
Wedding L. M., Lepczyk C. A., Pittman S. J., Friedlander A. M., Jorgensen S. (2011). Quantifying seascape structure: extending terrestrial spatial pattern metrics to the marine realm. Mar. Ecol. Prog. Ser. 427, 219–232. doi: 10.3354/meps09119
Yarnall A. H., Byers J. E., Yeager L. A., Fodrie F. J. (2022). Comparing edge and fragmentation effects within seagrass communities: A meta-analysis. Ecology 103, e3603. doi: 10.1002/ecy.3603
Yeager L. A., Keller D. A., Burns T. R., Pool A. S., Fodrie F. J. (2016). Threshold effects of habitat fragmentation on fish diversity at landscapes scales. Ecology 97, 2157–2166. doi: 10.1002/ecy.1449
Zarnetske P. L., Baiser B., Strecker A., Record S., Belmaker J., Tuanmu M. N. (2017). The interplay between landscape structure and biotic interactions. Curr. Landscape Ecol. Rep. 2, 12–29. doi: 10.1007/s40823-017-0021-5
Keywords: carbon stock, seagrass meadows, spatial configuration, reef lagoons, landscape ecology
Citation: Herrera-Silveira JA, Palafox-Juárez EB, Mendoza-Martínez JE, Montero Muñoz JL and Teutli Hernández C (2025) Landscape configuration in seagrass meadows and its influence on carbon stock in reef lagoon systems. Front. Mar. Sci. 11:1320194. doi: 10.3389/fmars.2024.1320194
Received: 11 October 2023; Accepted: 07 October 2024;
Published: 19 March 2025.
Edited by:
Moritz Müller, Swinburne University of Technology Sarawak Campus, MalaysiaReviewed by:
Briac Monnier, University of Corsica Pascal Paoli, FranceCopyright © 2025 Herrera-Silveira, Palafox-Juárez, Mendoza-Martínez, Montero Muñoz and Teutli Hernández. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juan Enrique Mendoza-Martínez, a2FzdW1vXzYwQGhvdG1haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.