
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Mar. Sci. , 15 April 2024
Sec. Marine Biology
Volume 11 - 2024 | https://doi.org/10.3389/fmars.2024.1235876
This article is part of the Research Topic The Use of Stable Isotope Ecology in Sharks View all 6 articles
 Dalia C. Barragán-Barrera1,2,3*
Dalia C. Barragán-Barrera1,2,3* Carlos J. Polo-Silva4,5
Carlos J. Polo-Silva4,5 Cindy González2,6Miguel Rodríguez2Laura Baldrich2,3Christian Bermúdez-Rivas7
Cindy González2,6Miguel Rodríguez2Laura Baldrich2,3Christian Bermúdez-Rivas7 Ángela Margarita Moncaleano-Niño1*
Ángela Margarita Moncaleano-Niño1* Paco Bustamante8
Paco Bustamante8 Susana Caballero2Andrea Luna-Acosta9
Susana Caballero2Andrea Luna-Acosta9Bonnethead and Caribbean sharpnose sharks frequent coastal waters, rendering them vulnerable to overexploitation, with the bonnethead shark listed as endangered by the IUCN. Marine Protected Areas (MPAs) have been suggested as a management strategy to regulate shark-exploitation. Moreover, it’s essential to identify key areas where nursery grounds (NG) and adult habitats overlap to prioritize conservation zones within MPAs. Along the Caribbean Sea, several NG have been identified for larger shark species, but little is known for smaller sharks. In Bocas del Toro (BDT-Panamanian Caribbean), both bonnethead and Caribbean sharpnose sharks are distributed, with the former appearing to be genetically isolated. The local MPA in BDT doesn't include a NG identified near Solarte Island. In this study, to provide ecological information on bonnethead and Caribbean sharpnose sharks within this potential NG, we evaluated stable isotopes (δ13C and δ15N) and total-mercury (THg) values in their fins. The results revealed a higher proportion of small-adult sharks in the NG, emphasizing the necessity of conducting a comprehensive study to further characterize this area. Moreover, there was no niche-partitioning between the two species and their diets primarily consisted of small prey-items, as evidenced by the significant and negative relationship between isotopic values. Both species exhibited low THg levels in their fins. Based on these initial results and previous genetic data, it appears that BDT plays a critical role as a habitat for sharks. Thus, extending the local MPA to cover the potential NG could effectively ensure conservation of bonnethead and Caribbean sharpnose sharks in BDT.
Sharks constitute one of the most extensively distributed groups of predators, inhabiting both oceanic and coastal regions (Ebert et al., 2013). However, several shark species are currently decreasing in population abundance due mainly to overexploitation (Dulvy et al., 2014, Dulvy et al., 2021; Pacoureau et al., 2021). Therefore, the establishment of marine protected areas (MPAs), where shark fishing is regulated or prohibited, serves as a key strategy to protect sharks (Carlisle et al., 2019).
Despite the establishment of MPAs, shark populations continue to decline significantly worldwide (Dulvy et al., 2014, Dulvy et al., 2021). The bonnethead shark (Sphyrna tiburo) exemplifies this tendency, inhabiting coastal and estuarine waters along the eastern Pacific from California (U.S.A) to Ecuador and throughout the western Atlantic from North Carolina (U.S.A) to southern Brazil (Compagno, 2002; Ebert et al., 2013). Despite this large distribution covering several MPAs (Álvarez Malvido et al., 2021), the species exhibits a declining population trend. Hence, it was recently listed in Appendix II in the last Conference of the Parties (CoP) to CITES (Cop19) and classified as endangered on the IUCN Red List, primarily due to fishing activities and habitat degradation (Pollom et al., 2021). Additionally, genetic and phylogenetic studies suggest the existence of at least one cryptic species in the Atlantic Ocean (e.g., Naylor et al., 2012; Sigovini et al., 2016; Díaz-Jaimes et al., 2021; Gonzalez et al., 2021), with the Caribbean bonnethead shark (S. aff. tiburo) distributed from Belize to southern Brazil (Gonzalez et al., 2021). Therefore, while taxonomic identity of bonnethead sharks remain unresolved in the Atlantic basin, the identification of key areas for inclusion within MPAs is imperative to ensure the protection of their populations as well as other shark species.
Sharks exhibit slow growth rates and a low intrinsic reproduction rate, emphasizing the importance of nursery grounds, where young sharks are born and reside until maturity (DeAngelis et al., 2008). In these areas, both juvenile and adult sharks are frequently encountered, as individuals tend to inhabit these areas for extended periods, often returning over multiple years (Heupel et al., 2007). Nursery grounds are primarily situated in coastal and shallow habitats such as bays, estuaries, and reef lagoons (Heupel et al., 2007). Given their significance, describing these areas is crucial for effective conservation and management of sharks (DeAngelis et al., 2008).
In the Wider Caribbean, several nursery areas have been identified for large sharks such as lemon (Negaprion brevirostris) and blacktip (Carcharhinus limbatus) sharks in coastal waters of the Bahamas, Turks and Caicos Islands, Florida (U.S.A), and the Virgin Islands (Feldheim et al., 2002; DeAngelis et al., 2008; Reyier et al., 2008; Henderson et al., 2010, Henderson et al., 2016). Nevertheless, limited information exists regarding nursery habitats of various shark species in regions along the continental Caribbean where no long-term shark monitoring programs are established, such as in Panama.
Situated along the western Caribbean coast of Panama is the Bocas del Toro Archipelago, where two species of sharks have been identified: the bonnethead shark (S. aff. tiburo) and the Caribbean sharpnose shark (Rhizoprionodon porosus). The Archipelago has a high biodiversity and the features described for a shark nursery ground, like shallow sandy areas fringed with mangrove (DeAngelis et al., 2008; Henderson et al., 2010). Indeed, a nursery area has been suggested for these both species in Solarte Island (Gonzalez et al., 2019); however, this region is not covered by the local MPA (Bastimentos Island National Marine Park, in Spanish Parque Nacional Marino Isla Bastimentos), which protect mainly the Bastimentos Island, Zapatilla Cays and adjacent waters.
Bastimentos Island MPA is in a region declared “particularly sensitive” by the International Maritime Organization and allows for protection of three distinct ecosystems: coral reefs, mangrove stands, and seagrass beds (Guerrón-Montero, 2005). The conservation purpose of this MPA was the protection of a representative portion of local coastal and marine ecosystems, focusing on marine turtles (Mou Sue, 1993), so protection of sharks may be underrepresented. Additionally, several impacts that may imply threats for sharks have been reported within the Archipelago, mainly in areas outside the MPA (Seemann et al., 2013). Contamination, habitat degradation, and overfishing are some of the human impacts affecting local marine fauna (Seemann et al., 2013), decreasing their populations (Del Cid et al., 1997; Seemann et al., 2013), which has also affected the income of local people that depend on these resources (Del Cid et al., 1997; Guerrón-Montero, 2005). Additionally, because of overfishing, fishes in the Archipelago are small-sized, which may also affect local diet for both humans and marine megafauna, as has been reported for common bottlenose dolphins (Tursiops truncatus), whose diet is based mainly on the poor-nutritive dwarf round herring (Jenkinsia lamprotaenia) (Barragán-Barrera et al., 2019).
The lack of information regarding the biology and population status of local sharks raises concerns, especially regarding the endangered status of the bonnethead shark (Pollom et al., 2021), whose population in Bocas del Toro appears to be genetically isolated (Gonzalez et al., 2019). Therefore, obtaining ecological information such as the identification of key areas for sharks in the Archipelago may be crucial for management purposes and should be considered in discussions of MPA expansion, to ultimately guarantee the conservation of sharks and other local marine fauna. For instance, strong baseline data including biological, genetic, and ecological information, revealed that Bocas del Toro Archipelago is a key area for a population of common bottlenose dolphin. This data indicated that the dolphin population living there had genetic isolation, exclusive inshore habits, and has been affected by unregulated tourist boat traffic (e.g., May-Collado et al., 2012; Barragán-Barrera et al., 2013; May-Collado et al., 2014; Sitar et al., 2016; Barragán-Barrera et al., 2017; May-Collado et al., 2017; Barragán-Barrera et al., 2019; Kassamali-Fox et al., 2020). Taking into account these previous studies, local stakeholders began to discuss two strategies to conserve this dolphin population.
The first was increasing the bottlenose dolphin conservation category to endangered in Bocas del Toro, and the second was the possibility of the Bastimentos Island MPA expansion within the Archipelago. Because these conservation strategies are still in discussion, here we intend to provide some ecological evidence supporting the relevance of expanding the area for other marine predators: sharks.
Gathering data on foraging habits and the toxicological status of shark populations is crucial for establishing conservation and management plans (Estupiñán-Montaño et al., 2021). The diet of sharks has been studied primarily via stomach contents and stable isotope analyses (e.g., Plumlee and Wells, 2016; Estupiñán-Montaño et al., 2017; Galindo et al., 2021; Vélez et al., 2021; Branham et al., 2022; Forero, 2022). Stable isotope analyses are a powerful methodology to assess the diet origin and composition by using carbon (δ13C) isotope values, as well as trophic position based on nitrogen (δ15N) measurements (Graham et al., 2010). Similarly, mercury (Hg) concentrations can serve as ecological tracers to determine trophic position, and diet differences related to age, sex, size, habitat use, and migration patterns (e.g., Barragán-Barrera et al., 2023; Estupiñán-Montaño et al., 2021; Graham et al., 2010; Maya M. et al., 2016; Marrugo-Negrete et al., 2018; Méndez-Fernández et al., 2020). However, as top predators like sharks are subject to elevated levels of Hg due to the biomagnification processes exposing them to potential health risks (Ehnert-Russo and Gelsleichter, 2020), monitoring Hg may be crucial for assessing the contamination status of marine habitats. Particularly for sharks, Hg monitoring is very relevant due their consumption in some coastal areas of Central America, including their meat (Clementi et al., 2021).
Here we used shark fins to assess, for the first time in the Bocas del Toro Archipelago, the total Hg (THg), as well as the δ13C and δ15N isotope values of bonnethead and Caribbean sharpnose sharks. Although fins reflect the lowest THg concentrations in comparison to other shark tissues like muscle, kidneys, or liver (O’Bryhim et al., 2017), their use is valuable to provide a first insight and baseline data on shark THg contamination in this region of Panama. Additionally, caution is warranted when interpreting fin stable isotope ratios, as they reflect values over longer periods than a year due their slow turnover rate (Malpica-Cruz et al., 2012). Fins, being appendages rather than tissues, consist of a mixture of several tissues (basal cartilage, blood, connective tissue, muscle, and skin), potentially leading to δ13C values higher than those in muscle (Hussey et al., 2011). However, utilizing the tips of fins enables the examination of shark ecotoxicology through nonlethal methods. This approach is valuable in offering initial first insights into the dietary habits and ecological status of shark populations from the same area trough stable isotope analysis (Hussey et al., 2011; Estupiñán-Montaño et al., 2021). Therefore, in this report, we provide insights about the ecotoxicological status of the Caribbean sharpnose and bonnethead sharks within a potential nursery area situated in the Bocas del Toro Archipelago. It aims to provide crucial baseline data for stakeholders contemplating the expansion of the Bastimentos Island MPA, facilitating the implementation of management and protection measures specifically aimed at conserving the endangered bonnethead sharks.
The Bocas del Toro Archipelago is in the Panamanian Caribbean, on the northwest coast bordering Costa Rica (Figure 1). The Archipelago contains several marine ecosystems including seagrass, sandy bottom, coral reef, and mangrove forest (Guzmán and García, 2002; D’Croz et al., 2005), distributed among seven larger islands and several smaller ones (Guzmán and Guevara, 1998). Shark samples were collected specifically in the coastal area of Solarte Island called Hospital Point, where a potential nursery area for a resident bonnethead shark population has been identified (Gonzalez et al., 2019). Within the Archipelago lies the Bastimentos Island National Marine Park, whose polygon covers a coastal and marine portion over the Bastimentos Island and Zapatilla Cays.
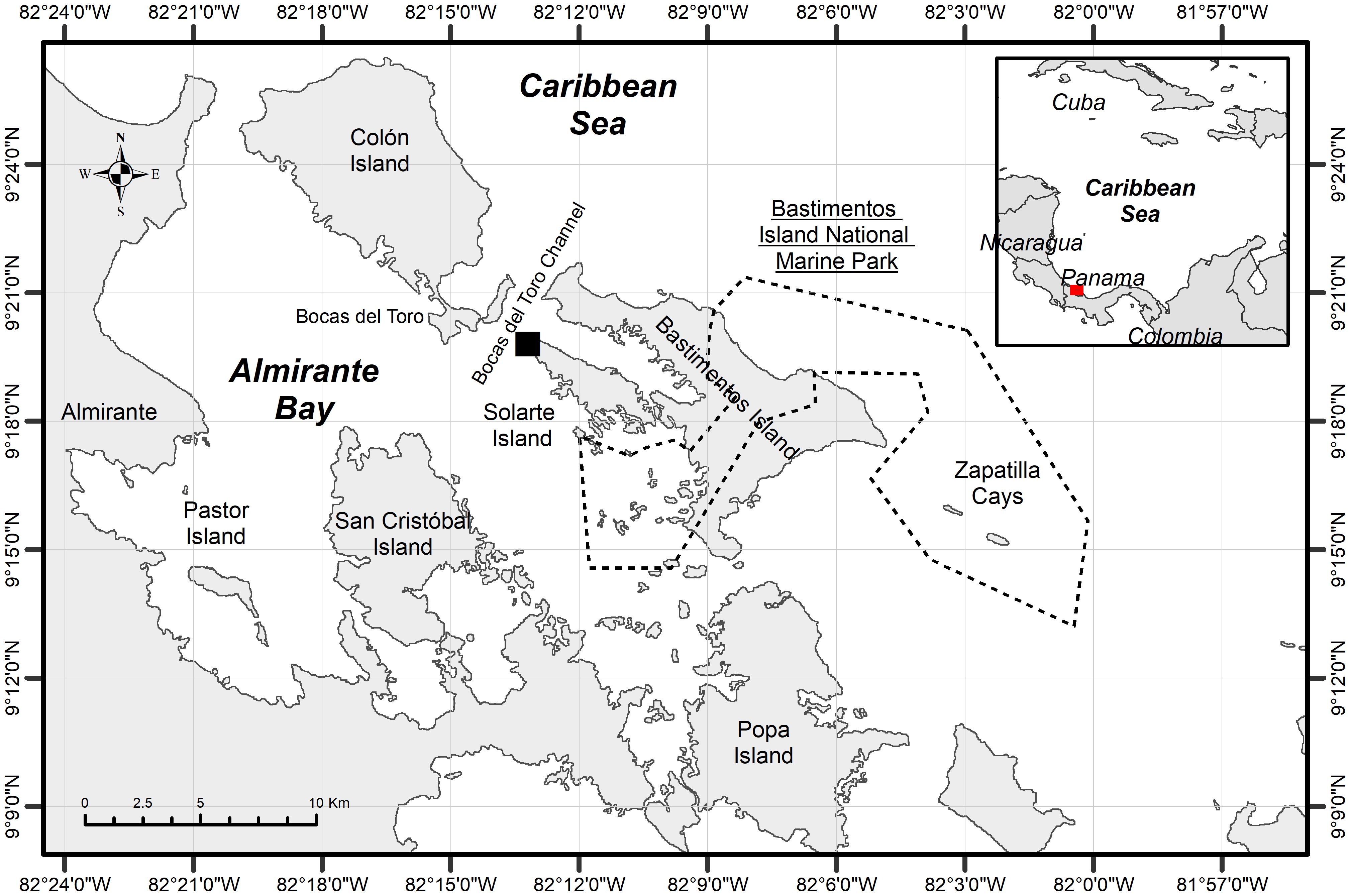
Figure 1 Location of sharks sampling collection area, in Hospital Point (black square), located in the Solarte Island, Bocas del Toro Archipelago, Panamanian Caribbean. The polygon of the Bastimentos Island National Marine Park –Parque Nacional Marino Isla Bastimentos– is represented with a dotted line.
Shark samples were collected with permission from the Ministerio de Ambiente in Panama (permit number 05870- SEX/A-2-17). The methodology for sample collection was approved by the Smithsonian Tropical Research Institute IACUC (Institutional Animal Care and Use Committee; permits number 20676904 and 20676903).
Following Gonzalez et al. (2019), a fin clip sample was collected from six Caribbean sharpnose sharks and 15 bonnethead sharks at Hospital Point (Figure 1) between October 2016 and January 2017. Samples were conserved in 95% ethanol and stored at −20°C for subsequent laboratory analysis. The sex and total length were also registered for each individual, which were released alive after sampling (Gonzalez et al., 2019). Total length was used to distinguish between juveniles and adults based on mean maturity length for both species reported in other Caribbean areas, so values higher than the mean maturity length represented adults. The adult length for Caribbean sharpnose sharks was assessed at 66.9 cm based on 375 specimens collected in Northeastern Region of Venezuela (Medina et al., 2009). For bonnethead sharks, adults were differentiated from juveniles with a length greater than 77.3 cm and 64.5 cm for females and males, respectively, according to a life history study conducted in the western Gulf of Mexico using 913 individuals (N-females= 561, N-males= 352; Frazier et al., 2023).
Shark fin samples were dried, homogenized, and lyophilized prior to analysis as described in Vélez et al. (2021). Prior to isotopic analyses, and because samples are depleted in 13C, we used 4 ml of cyclohexane to extract lipids from approximately 100 mg (or less) of each sample (De Niro and Epstein, 1978). The samples were maintained in constant agitation for 10 min, and then centrifuged at 4,500 rpm for 5 min after which the supernatant containing lipids was discarded. We repeated this process three times, followed by drying the samples at 45°C in an oven for 48 h. Finally, a small portion of the lipid-free sample (0.2 – 0.4 mg) was weighted in a tin cup to conduct stable isotope analyses in a continuous flow mass spectrometer (Delta V Plus with a Conflo IV Interface, Thermo Scientific, Bremen, Germany) coupled to an elemental analyzer (Flash 2000 or EA Isolink, Thermo Scientific, Milan, Italy). The usual δ notation relative to atmospheric N2 for δ15N and Vienna PeeDee Belemnite Standard for δ13C in parts per thousand (‰), was used to report the isotopic results (Méndez-Fernández et al., 2020).
Based on replicate measurements of internal laboratory standards, experimental precision is of ± 0.15 and ± 0.20 ‰ for δ13C and δ15N, respectively. The C:N ratio was assessed to determine if lipid extraction was efficient, by using the percent C and N elemental composition (De Niro and Epstein, 1978). Thus, a good lipid removal was indicated if we obtained C:N ratio between 2.9 – 3.3 (Post et al., 2007; Lesage et al., 2010; Matich et al., 2010; Kim et al., 2012).
Measurements of THg concentrations were performed using an atomic absorption spectrometer AMA-254 (Advanced Altec Mercury Analyzer) following Vélez et al. (2021). Homogenized and lyophilized tissue samples ranging from 1 to 5 mg that did not require chemical treatment were analyzed in duplicates to assess mean and SD of THg concentrations in each sample. To control analytical quality of THg measurements, we ran blanks at the beginning of each analytical session, and we used certified reference material (CRM) DOLT-3 (Dogfish Liver Certified Reference Material, National Research Council of Canada) after running blanks and after every fifth sample run. The mean of concentrations reported for the CRM was 328 ± 0.1 ng g-1 (N= 4) and the percent of recovery was 97.0 ± 0.9%. The detection limit of the AMA was 0.1 ng. The THg concentrations were reported in ng g-1 on a dry weight basis (dw).
Mean isotopic and THg and their standard deviation for each shark species was calculated and plotted. Following Jackson et al. (2011), a Bayesian approach based on multivariate ellipse metrics (SIBER – Stable Stable Isotope Bayesian Ellipses within the R-siar package) was used to assess the overlapping of isotopic niche between bonnethead and Caribbean sharpnose sharks. The degree of niche overlapping was assessed through the overlap in the areas of the corrected standard ellipses (SEAc), by using the OVERLAP command, whose value ranges from 0 (no overlap) to 1 (greater overlap; Jackson et al., 2011).
Because of the low sample size for Caribbean sharpnose shark, these samples were not used to conduct additional statistical analyses. Therefore, Shapiro–Wilk of normality and Levene test of homogeneity of variances were used to test the assumptions of parametric tests on the bonnethead shark data. For data that was not normally distributed, a log-transformation was used. Pearson correlations were conducted among all variables to assess: 1) Hg bioaccumulation in relation to trophic position and age by comparing THg Vs δ15N, and THg Vs total length, respectively (Marrugo-Negrete et al., 2018), and 2) the relationship between δ13C and δ15N values among shark individuals. The existence of significant differences between sex was evaluated using a t-student. All these statistical analyses were performed in R v. 3.4.3.
Fifteen fin samples of bonnethead sharks were sampled, whose total lengths ranged between 35 and 106 cm, including two juveniles and 13 adults (Table 1). The six Caribbean sharpnose shark individuals' total lengths varied between 44 to 95 cm, which corresponded to five juveniles and only one adult (Table 1). Regarding stable isotopes, the C:N ratios reflected an efficient removal of lipids with values ranging from 2.93 to 3.29 (Table 1).
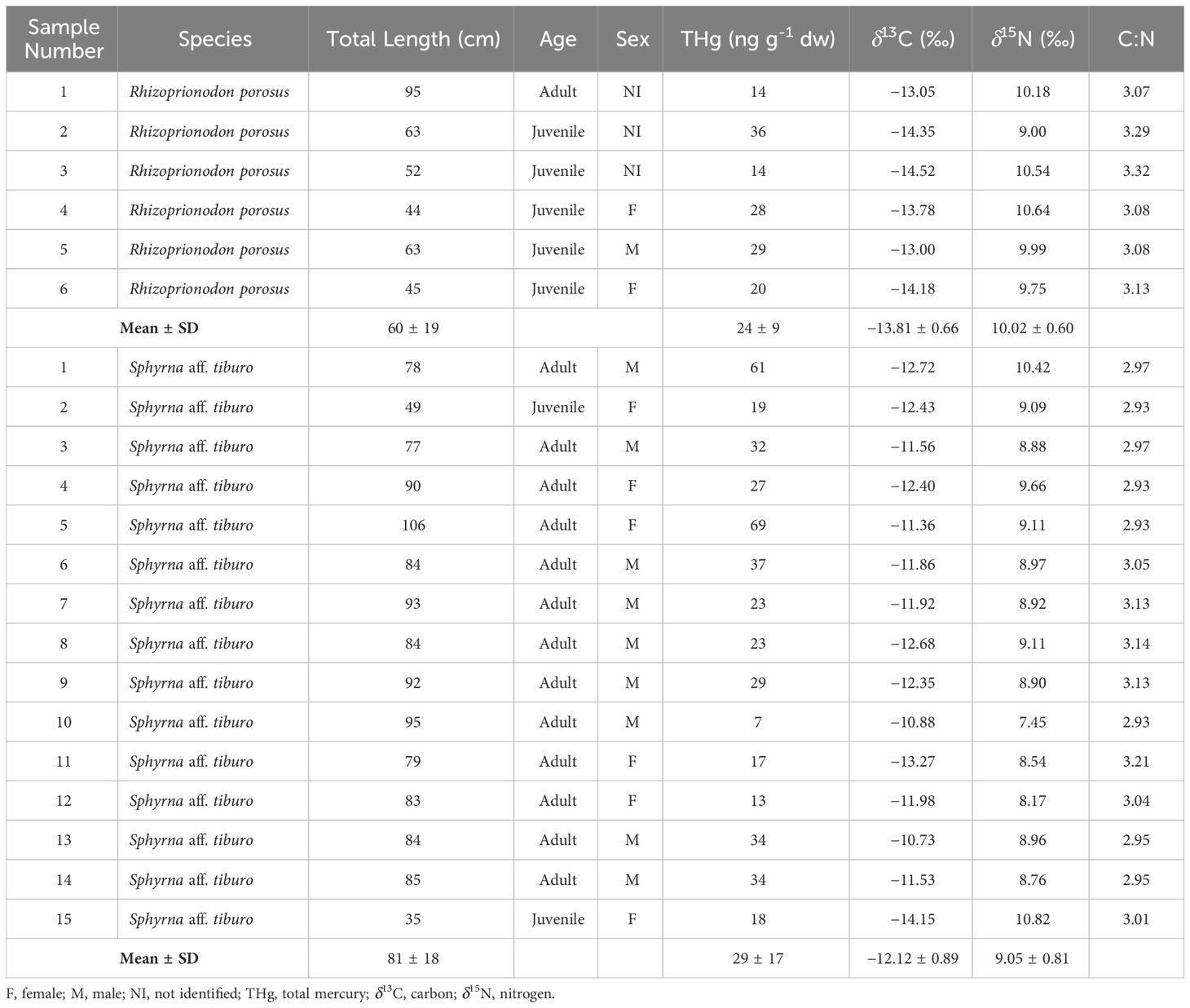
Table 1 Total length, age, sex, total mercury concentration, stable isotope values, and C:N ratio for bonnethead shark (Sphyrna aff. tiburo) and Caribbean sharpnose shark (Rhizoprionodon porosus) in the Bocas del Toro Archipelago, Panamanian Caribbean.
The δ13C values ranged between −14.15 to −10.73 ‰ (mean= −12.12 ± 0.81 ‰) and between −14.52 to −13.00 ‰ (mean= −13.81 ± 0.66 ‰) for bonnethead and Caribbean sharpnose sharks, respectively (Table 1, Figure 2). The δ15N values were similar for both species, which ranged between 7.45 to 10.82 ‰ (mean= 9.05 ± 0.81 ‰) for bonnethead sharks, and between 9.00 to 10.64 ‰ (mean= 10.02 ± 0.60 ‰) for Caribbean sharpnose sharks. Likewise, THg values were also similar between both species, with a range of 7 to 69 ng g-1 dw (mean= 29 ± 17 ng g-1 dw) and 14 to 36 ng g-1 dw (mean= 24 ± 9 ng g-1 dw) for bonnethead and Caribbean sharpnose sharks, respectively (Table 1, Figure 2). However, SIBER analyses based on SEAc values of carbon and nitrogen isotopic signatures showed no niche overlap between both species (Overlap= 0), and the bonnethead shark showed a greater ellipse (SEAc= 1.87) in comparison to the Caribbean sharpnose shark (SEAc= 1.52; Figure 3).
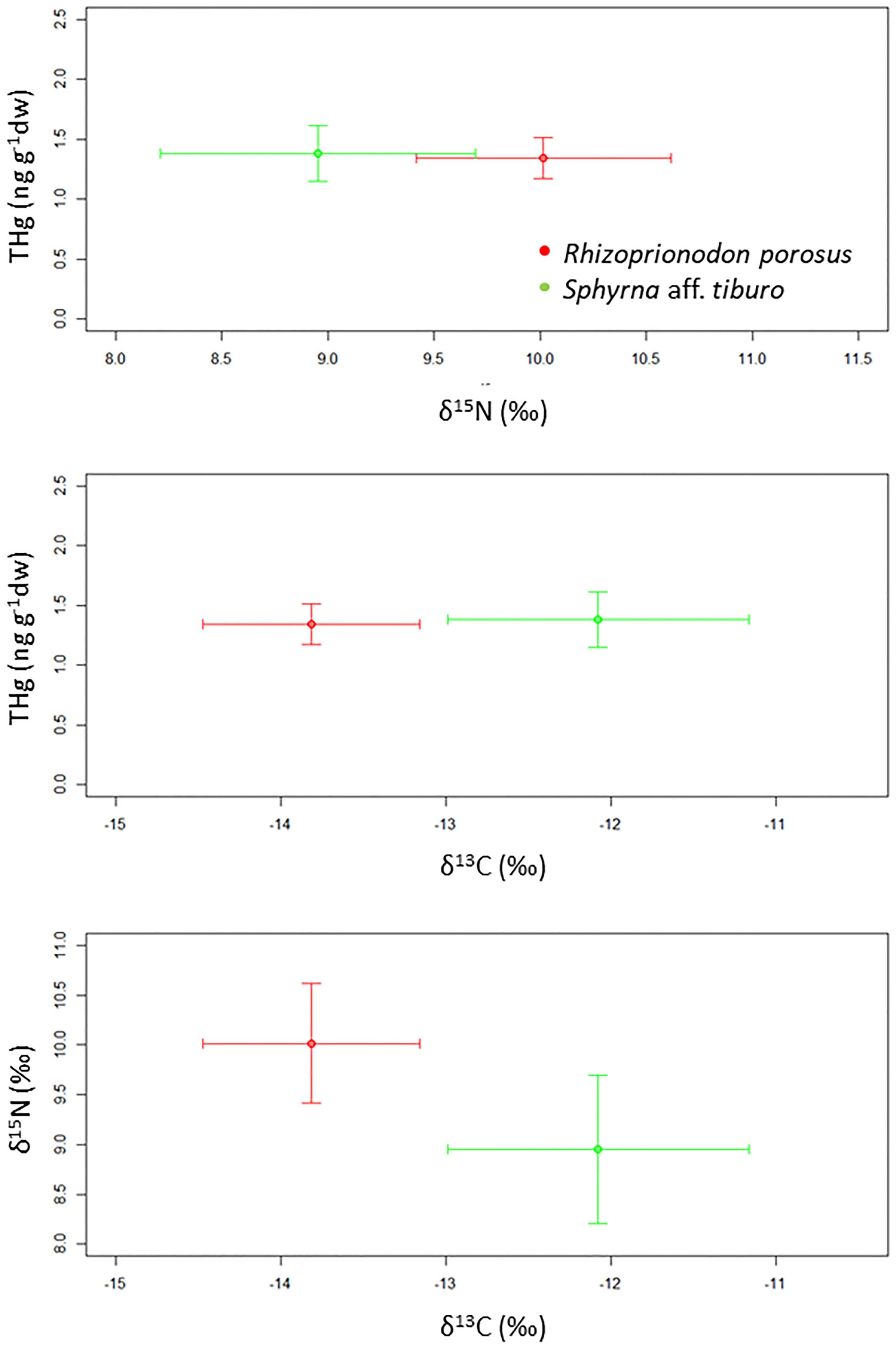
Figure 2 Relationship between carbon (δ13C) and nitrogen (δ15N) stable isotope values, and total mercury (THg) levels of Caribbean sharpnose shark (Rhizoprionodon porosus) and bonnethead shark (Sphyrna aff. tiburo) in the Bocas del Toro Archipelago, Panamanian Caribbean.

Figure 3 Standard ellipses for corrected sample sizes (SEAc) of carbon and nitrogen isotopic signatures in fins for bonnethead shark (Sphyrna aff. tiburo) and Caribbean sharpnose shark (Rhizoprionodon porosus) and in the Bocas del Toro Archipelago, Panamanian Caribbean. Black ellipse: bonnethead shark; Red ellipse: Caribbean sharpnose shark.
For bonnethead sharks, sex comparisons did not show significant differences for THg, δ13C, and δ15N values. The Pearson’s correlation coefficients showed not significant correlations between THg with δ15N and standard length, but was significant for the δ13C and δ15N relationship (p<0.05).
This study provides a first insight into the feeding ecology of the bonnethead and Caribbean sharpnose sharks in a potential nursery area located at the Bocas del Toro Archipelago, Panama, based on THg, δ13C, and δ15N values. Despite most individuals collected at Hospital Point appearing to be adults instead of juveniles, this region maintains features of a nursery ground (e.g., Heupel et al., 2007), in which the presence of both small adults and juveniles suggests that individuals use the region frequently. This preliminary assessment highlights the need for conducting an exhaustive reproductive biology study for both bonnethead and Caribbean sharpnose sharks in the Archipelago, to identify the relevance of this region as a nursery ground for these species in the Caribbean. These results are particularly important for the bonnethead shark, since the population located in Bocas del Toro appears to be genetically isolated (Gonzalez et al., 2019).
The bonnethead shark is widely distributed along estuarine, coastal, and insular areas of the American continent (up 90 m depth) in both Atlantic and Pacific oceans, but limited to warm waters (Ebert et al., 2013). However, this species which likely represents a differentiated linage in the Caribbean and Southwestern Atlantic (Naylor et al., 2012; Sigovini et al., 2016; Díaz-Jaimes et al., 2021; Gonzalez et al., 2021), also shows a restricted distribution in Bocas del Toro, where the population appears to be resident according to genetic data (Gonzalez et al., 2019). Our findings also indicated the absence of niche partitioning between female and male bonnethead sharks within the Archipelago. These findings align with other studies that report no discernible differences in diet between sexes (e.g., Galindo et al., 2021; Branham et al., 2022), particularly in nursery areas where the main goal is to acquire as many prey items as possible for rapid growth (Heithaus, 2007). Conversely, niche partitioning was detected between species, which allows co-existence of the two species in the same region despite both having coastal habits (Maya M. et al., 2016; Estupiñán-Montaño et al., 2017).
Stable carbon isotope compositions suggest that the Caribbean sharpnose shark, whose distribution in the Atlantic Ocean appears to be in deeper waters (i.e., up to 500 m depth; Ebert et al., 2013), prefers items with higher trophic position but specific to the coastline. The trophic niche of this species in Bocas del Toro is narrower in comparison to the bonnethead shark, which showed a wider δ13C range, but with less depleted 13C values, suggesting a wider foraging area. However, variation in δ13C values between both species may be supported mainly by the wide range of isotopically diverse sources that can be found in coastal areas (Bird et al., 2018), and the Archipelago appears to offer a high variety of coastal preys to top predators (Barragán-Barrera et al., 2019). Hence, the partitioning niche found here exhibits diet differences between both shark species. For instance, the diet of the bonnethead shark along the Atlantic waters of Brazil and United States is dominated mostly by portunid crabs (Callinectes spp.; Plumlee and Wells, 2016; Branham et al., 2022), while in the Colombian Caribbean, the diet of the Caribbean sharpnose shark was represented by eight fish species (e.g., Harengula sp., flying gurnard Dactylopterus volitans, Mugil sp., sharptail eel Myrichthys breviceps, among others) and one crustacean penaeid (family Penaeidae), with fish belonging to the Clupeidae family being the most common item in their diet (Forero, 2022). Therefore, the hypothesis of diverse food sources in the Bocas del Toro Archipelago, which had been discussed previously for common bottlenose dolphins, may also be true for sharks. These dolphins have a high philopatry in the Archipelago despite the risk related to continuous boat traffic (May-Collado et al., 2012, May-Collado et al., 2014, May-Collado et al., 2017; Kassamali-Fox et al., 2020), which could only be explained by the good habitat that Bocas del Toro offers to individuals (Barragán-Barrera et al., 2019). Consequently, this area is an important habitat for megafauna in the Caribbean, and deserves special attention for shark conservation, particularly for the bonnethead shark due to its recent IUCN categorization as endangered with a decreasing population trend (Pollom et al., 2021).
In Bocas del Toro, there are potential threats that sharks may face. Hg bioaccumulation may be one of them; however, bonnethead sharks, for instance, did not show higher THg levels in their fins in comparison to other areas throughout its distribution. For example, in the Southeastern United States, bonnethead sharks exhibited THg levels in their fins with a mean of 70 ± 50 ng g-1 dw (N= 18; O’Bryhim et al., 2017), and in the Colombian Pacific, where has been reported high Hg emissions due to gold mining (Massé and McDermott, 2017), this species exhibited THg values in their fins with a mean of 173 ± 161 ng g-1 dw (N=17; Vélez et al., 2021). Nevertheless, this species did not tend to bioaccumulate high THg levels in relation to other oceanic and larger shark species (Vélez et al., 2021). This may be due to the limited size range in our sampling (juveniles and small adults), since Hg is known to bioaccumulate in organisms over their lifespan, older and larger animals typically exhibit higher Hg levels (Neumann and Ward, 1999).
Relationship between δ15N and THg values are also used to assess Hg contamination through diet, in which top predators show a positive correlation due to consumption of high trophic level prey (e.g., Marrugo-Negrete et al., 2018; Barragán-Barrera et al., 2019). However, this pattern was not observed for bonnethead sharks in Bocas del Toro. Stable nitrogen isotope composition showed similar values for both bonnethead and Caribbean sharpnose sharks, but were lower in comparison to the inshore population of common bottlenose dolphins in Bocas del Toro (mean= 10.25 ± 1.48 ‰; tissue: skin; N= 37; Barragán-Barrera et al., 2019), which result in lower THg values through dietary items with lower trophic level.
Diet of both bonnethead and Caribbean sharpnose sharks along Atlantic areas are based on benthic crustacea and fishes (Ebert et al., 2013; Plumlee and Wells, 2016; Branham et al., 2022; Forero, 2022). However, fishes in Bocas del Toro are being overfished (Seemann et al., 2013), so top predators should supply their nutritional demand by consuming smaller fishes or other types of prey (Barragán-Barrera et al., 2019). This may also be represented for sharks by the significant negative relationship between δ13C and δ15N stable isotopes, with a decreasing 15N trend in relation to more depleted 13C values. This relationship suggests a shark diet based on small fishes with both coastal and oceanic habits, like the dwarf round herring, which is widely distributed within the Archipelago, but feeds on oceanic zooplankton during the night (Radakov and Silva, 1974; Friedlander and Beets, 1997). Additionally, the dwarf round herring represents one of the most contributing prey items in the diet of common bottlenose dolphins in Bocas del Toro (Barragán-Barrera et al., 2019), likely because of the lack of larger prey due to local overfishing (Seemann et al., 2013). Consequently, dolphins in the Archipelago and likely sharks may reflect lower δ15N values by acquiring lower trophic level preys in comparison to other areas (e.g., Vélez et al., 2021). Considering this, overfishing (which may be regulated within the local MPA; Seemann et al., 2013), may result in a potential threat that sharks may face in Bocas del Toro.
Our preliminary findings, together with previous genetic data that shows isolation (Gonzalez et al., 2019), suggest the relevance of the Bocas del Toro Archipelago as an important habitat for sharks in the Caribbean basin. However, additional monitoring including more specimens with different age class as well as the complete characterization of the potential nursery ground is needed to confirm this assumption. Nursery delineation is crucial for assessing the significance of this area for shark species and is pivotal for their effective management by integrating it into the local MPA. Furthermore, complementary analyses are needed to effectively assess the feeding ecology of bonnethead and Caribbean sharpnose sharks in the Caribbean Sea. This study provides essential baseline data needed to determine the conservation status of these species in Panama, particularly for the endangered bonnethead shark (Pollom et al., 2021). In order to conserve its populations, urgent measures are needed to determine important areas for this species, with Bocas del Toro emerging as a key location in need of conservation efforts.
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
The animal study was approved by Smithsonian Tropical Research Institute IACUC (Institutional Animal Care and Use Committee; permits number 20676904 and 20676903). The study was conducted in accordance with the local legislation and institutional requirements.
DB-B conceptualized the study. DB-B, CG acquired funding. CG collected samples. SC, AM-N, PB provided logistic support. DB-B conducted lab work. DB-B, MR, LB, CB-R performed statistical analysis. DB-B, CP-S, AL-A validated data. DB-B wrote first draft and reviewed manuscript. CP-S, MR, LB, PC, SC, AL-A reviewed and edited the manuscript draft. All authors contributed to the article and approved the submitted version.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. The Smithsonian Tropical Research Institute through a short-term fellowship and Beneath the Waves Inc supported CG to conduct this study. The Corporation Center of Excellence in Marine Sciences – CEMarin supported this study (DB-B, 2016). The Faculty of Sciences of Universidad de los Andes also provided two Research Grants: “Proyecto Semilla -2016-1 Call for Funding of Research Category: Master and Doctoral students” (CG, 2016), and “Proyecto Semilla -2018-1 Call for Funding of Research Category: Candidates PhD students, project “Isotopic analyses of bottlenose dolphins’ diet in Bocas del Toro: a conservation perspective” (DB-B, 2017). Thanks are due to the CPER (Contrat de Projet Etat-Région) and the FEDER (Fonds Européen de Développement Régional) for funding the AMA and the IRMS of LIENSs laboratory. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
We thank the BDT Research Station—Smithsonian Tropical Research Institute (STRI) for their logistical support, especially to Marlon Smith, Alberto Machuca, Erick Brown, and Deivis Gonzalez. Special thanks to C. Churlaud and M. Brault-Favrou from the Plateforme Analyses Elémentaires of the LIENSs laboratory for facilitating Hg analyses, and G. Guillou from the Plateforme Analyses Isotopiques of the LIENSs laboratory for its support in the stable isotope analyses. Many thanks to Rebecca Regnery for reviewing English in the manuscript. We also want to thank the reviewers whose comments improved the final version of this manuscript. PB is an honorary member of the IUF (Institut Universitaire de France). The Vicerrectoría de Investigaciones from Pontificia Universidad Javeriana is also acknowledged by providing a Postdoctoral Grant (Call 2021-2) to D. Barragán, who also thanks to the Instituto Javeriano del Agua for its support during the Postdoctoral stay.
Author CP-S is a director of Sharky Management & Consulting.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Álvarez Malvido M., Lázaro C., De Lamo X., Juffe-Bignoli D., Cao R., Bueno P., et al. (2021). Informe Planeta Protegido 2020: Latinoamérica y el Caribe (Ciudad de México, Cambridge, Gland, Bogotá: RedParques, UNEP-WCMC, CMAP-UICN, WWF, CONANP, Proyecto IAPA).
Barragán-Barrera D. C., Luna-Acosta A., May-Collado L. J., Polo-Silva C., Riet-Sapriza F. G., Bustamante P., et al. (2019). Foraging habits and levels of mercury in a resident population of bottlenose dolphins (Tursiops truncatus) in Bocas del Toro Archipelago, Caribbean Sea,Panama. Mar. Pollut. Bull. 145, 343–356. doi: 10.1016/j.marpolbul.2019.04.076
Barragán-Barrera D. C., May-Collado L. J., Quiñones-Lebrón S. G., Caballero S. (2013). Population at risk: low genetic diversity in bottlenose dolphins of Bocas del Toro, Panama. Int. Whal. Comm SC/65a/SM15, 1–11.
Barragán-Barrera D. C., May-Collado L. J., Tezanos-Pinto G., Islas-Villanueva V., Correa-Cárdenas C. A., Caballero S. (2017). High genetic structure and low mitochondrial diversity in bottlenose dolphins of the archipelago of Bocas del Toro, Panama: a population at risk? PloS One 12, e0189370. doi: 10.1371/journal.pone.0189370
Barragán-Barrera D. C., Riet-Sapriza F., Negrete J., Curtosi A., Mojica-Moncada D., Bustamante P., et al. (2023). Sex-specific mercury levels in skin samples of southern elephant seals (Mirounga leonina) at Isla 25 de Mayo (King George Island), Antarctic Peninsula. Mar. Mamm. Sci. 40, 108–122. doi: 10.1111/mms.13058
Bird C. S., Veríssimo A., Magozzi S., Abrantes K. G., Aguilar A., Al-Reasi H., et al. (2018). A global perspective on the trophic geography of sharks. Nat. Ecol. Evol. 2, 299–305. doi: 10.1038/s41559-017-0432-z
Branham C. C., Frazier B. S., Strange J. B., Galloway A. S., Adams D. H., Drymon J. M., et al. (2022). Diet of the bonnethead Sphyrna tiburo along the northern Gulf of Mexico and southeastern Atlantic coast of the United States. Anim. Biodivers. Conserv. 45, 257–267. doi: 10.32800/abc.2022.45.0257
Carlisle A. B., Tickler D., Dale J. J., Ferretti F., Curnick D. J., Chapple T. K., et al. (2019). Estimating space use of mobile fishes in a large marine protected area with methodological considerations in acoustic array design. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00256
Clementi G. M., Babcock E. A., Valentin-Albanese J., Bond M. E., Flowers K. I., Heithaus M. R., et al. (2021). Anthropogenic pressures on reef-associated sharks in jurisdictions with and without directed shark fishing. Mar. Ecol. Prog. Ser. 661, 175–186. doi: 10.3354/meps13607
Compagno L. J. V. (2002). Sharks of the world. An annotated and illustrated catalogue of shark species to date, (Rome, Italy: FAO) 4(125), FIR/S125.
D’Croz L., Del Rosario J. B., Góndola P. (2005). The effect of fresh water runoff on the distribution of dissolved inorganic nutrients and plankton in the Bocas del Toro Archipelago, Caribbean Panama. Caribb. J. Sci. 41, 414–429.
DeAngelis B. M., McCandless C. T., Kohler N. E., Recksiek C. W., Skomal G. B. (2008). First characterization of shark nursery habitat in the United States Virgin Islands: evidence of habitat partitioning by two shark species. Mar. Ecol. Prog. Ser. 358, 257–271. doi: 10.3354/meps07308
Del Cid M., Carrión de Samudio J., Valdespinos I. A., Santamaría D. (1997). Evaluación Rural Participativa de las Áreas de Influencia al Parque Nacional Marino Isla Bastimentos y al Humedal San San-Pond Sak, Provincia de Bocas del Toro. Tomo 2: Aspectos Socioeconómicos (Panamá: Asociación Nacional para la Conservación de la Naturaleza (ANCON), 204.
De Niro M. J., Epstein S. (1978). Influence of diet on the distribution of carbon isotopes in animals. Geochim. Cosmochim. Acta 42, 495–506. doi: 10.1016/0016-7037(78)90199-0
Díaz-Jaimes P., Bayona-Vásquez N. J., Escatel-Luna E., Uribe-Alcocer M., Pecoraro C., Adams D. H., et al. (2021). Population genetic divergence of bonnethead sharks Sphyrna tiburo in the western North Atlantic: Implications for conservation. Aquat. Conserv.: Mar. Freshw. 31, 83–98. doi: 10.1002/aqc.3434
Dulvy N. K., Fowler S. L., Musick J. A., Cavanagh R. D., Kyne P. K., Harrison L. R., et al. (2014). Extinction risk and conservation of the world’s sharks and rays. eLife 3, e00590. doi: 10.7554/eLife.00590
Dulvy N. K., Pacoureau N., Rigby C. L., Pollom R. A., Jabado R. W., Ebert D. A., et al. (2021). Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Curr. Biol. 31, 4773–4787. doi: 10.1016/j.cub.2021.08.062
Ehnert-Russo S. L., Gelsleichter J. (2020). Mercury accumulation and effects in the brain of the atlantic sharpnose shark (Rhizoprionodon terraenovae). Arch. Environ. Contam. Toxicol. 78, 267–283. doi: 10.1007/s00244-019-00691-0
Estupiñán-Montaño C., Galván-Magaña F., Tamburín E., Sánchez-González A., Villalobos-Ramírez D. J., Murillo-Bohórquez N., et al. (2017). Trophic inference in two sympatric sharks. Acta Ichthyol. Piscat. 47, 357–364. doi: 10.3750/AIEP/02177
Estupiñán-Montaño C., Tamburin E., Delgado-Huertas A. (2021). Stable isotope evidence for movements of hammerhead sharks Sphyrna lewini, connecting two natural protected areas in the Colombian Pacific. Mar. Biodivers 51, 74. doi: 10.1007/s12526-021-01215-7
Feldheim K. A., Gruber S. H., Ashley. M. V. (2002). The breeding biology of lemon sharks at a tropical nursery lagoon. Proc. Biol. Sci. 269, 1655–1661. doi: 10.1098/rspb.2002.2051
Forero B. (2022). Hábitos tróficos de Carcharhinus falciformis y Rhizoprionodon porosus mediante análisis estomacal e isótopos estables, en el Caribe Colombiano. Santa Marta, Colombia: Fundación Universidad de Bogotá Jorge Tadeo Lozano, 61.
Frazier B. S., Vinyard E. A., Fields A. T., Driggers W. B. III, Grubbs R. D., Adams D. H., et al. (2023). Age, growth and maturity of the bonnethead Sphyrna tiburo in the U.S. Gulf of Mexico. Environ. Biol. Fish. 106, 1597–1617. doi: 10.1007/s10641-023-01439-5
Friedlander A. M., Beets J. P. (1997). Fisheries and life history characteristics of dwarf herring (Jenkinsia lamprotaenia) in the US Virgin Islands. Fish. Res. 31, 61–72. doi: 10.1016/S0165-7836(97)00015-5
Galindo E., Giraldo A., Navia A. F. (2021). Feeding habits and trophic interactions of four sympatric hammerhead shark species reveal trophic niche partitioning. Mar. Ecol. Prog. Ser. 665, 159–175. doi: 10.3354/meps13681
Gonzalez C., Gallagher A. J., Caballero S. (2019). Conservation genetics of the bonnethead shark Sphyrna tiburo in Bocas del Toro, Panama: Preliminary evidence of a unique stock. PloS One 14, e0220737. doi: 10.1371/journal.pone.0220737
Gonzalez C., Postaire B., Domingues R. R., Feldheim K. A., Caballero S., Chapman D. (2021). Phylogeography and population genetics of the cryptic bonnethead shark Sphyrna aff. tiburo in Brazil and the Caribbean inferred from mtDNA markers. J. Fish Biol. 99, 1899–1911. doi: 10.1111/jfb.14896
Graham B. S., Koch P. L., Newsome S. D., McMahon K. W., Aurioles D. (2010). “Using isoscapes to trace the movements and foraging behavior of top predators in oceanic ecosystems,” in Isoscapes: Understanding Movement, Pattern, and Process on Earth Through Isotope Mapping. Eds. West J., Bowen G., Dawson T., Tu K. (Springer, Dordrecht), 299–318. doi: 10.1007/978-90-481-3354-3_14
Guerrón-Montero C. (2005). Marine protected areas in Panama: grassroots activism and advocacy. Hum. Organ. 64, 360–373. doi: 10.17730/humo.64.4.2mx2j6qd0xyg1rqv
Guzmán H. M., García E. M. (2002). Mercury levels in coral reefs along the Caribbean coast of Central America. Mar. Pollut. Bull. 44, 1415–1420. doi: 10.1016/S0025-326X(02)00318-1
Guzmán H. M., Guevara C. A. (1998). Arrecifes coralinos de Bocas del Toro, Panamá: I. Distribución, estructura y estado de conservación de los arrecifes continentales de la Laguna de Chiriquí y la Bahía Almirante. Rev. Biol. Trop. 46, 601–623.
Heithaus M. R. (2007). Nursery areas as essential shark habitats: A theoretical perspective. Am. Fish. Soc Symp. 50, 3–13.
Henderson A. C., Jourdan A., Bell K. (2016). Assessing the incidental value of a marine reserve to a lemon shark Negaprion brevirostris nursery. Aquat. Conserv.: Mar. Freshw. 26, 482–491. doi: 10.1002/aqc.2627
Henderson A., Katherine M., Calosso M. (2010). Preliminary assessment of a possible lemon shark nursery in the Turks & Caicos Islands, British West Indies. Car. J. Sci. 46, 29–38. doi: 10.18475/cjos.v46i1.a5
Heupel M. R., Carlson J. K., Simpfendorfer C. A. (2007). Shark nursery areas: Concepts, definition, characterization and assumptions. Mar. Ecol. Prog. Ser. 337, 287–297. doi: 10.3354/meps337287
Hussey N. E., Chapman D. D., Donnelly E., Abercrombie D. L., Fisk A. T. (2011). Fin-icky samples: an assessment of shark fin as a source material for stable isotope analysis. Limnol. Oceanogr-Meth. 9, 524–532. doi: 10.4319/lom.2011.9.524
Jackson A. L., Inger R., Parnell A. C., Bearhop S. (2011). Comparing isotopic niche widths among and within communities: SIBER - Stable Isotope Bayesian Ellipses. R. J. Anim. Ecol. 80, 595–602. doi: 10.1111/jane.2011.80.issue-3
Kassamali-Fox A., Christiansen F., May-Collado L. J., Ramos E. A., Kaplin B. A. (2020). Tour boats affect the activity patterns of bottlenose dolphins (Tursiops truncatus) in Bocas del Toro, Panama. PeerJ 8, e8804. doi: 10.7717/peerj.8804
Kim S. L., Martínez del Río C., Casper D., Koch P. L. (2012). Isotopic incorporation rates for shark tissues from a long-term captive feeding study. J. Exp. Biol. 215, 2495–2500. doi: 10.1242/jeb.070656
Lesage V., Morin Y., Rioux E., Pomerleau C., Fergusson S. H., Pelletier E. (2010). Stable isotopes and trace elements as indicators of diet and habitat use in cetaceans: Predicting errors related to preservation, lipid extraction and lipid normalization. Mar. Ecol. Prog. Ser. 419, 249–265. doi: 10.3354/meps08825
Malpica-Cruz L., Herzka S. Z., Sosa-Nishizaki O., Lazo J. P. (2012). Tissue-specific isotope trophic discrimination factors and turnover rates in a marine elasmobranch: empirical and modeling results. Can. J. Fish. Aquat. Sci. 69, 551–564. doi: 10.1139/f2011-172
Marrugo-Negrete J. L., Ruiz-Guzmán J. A., Ruiz-Fernández A. C. (2018). Biomagnification of mercury in fish from two gold mining-impacted tropical marshes in Northern Colombia. Arch. Environ. Contam. Toxicol. 74, 121–130. doi: 10.1007/s00244-017-0459-9
Massé F., McDermott J. (2017). Responsible Business Conduct: Due Diligence in Colombia’s Gold Supply Chain Gold Mining in Antioquia (OECD), 44. Available at: https://www.oecd.org/daf/inv/mne/colombias-gold-supply-chain.htm.
Matich P., Heithaus M. R., Layman C. A. (2010). Contrasting patterns of individual specialization and trophic coupling in two marine apex predators. J. Anim. Ecol. 80, 294–305. doi: 10.1111/j.1365-2656.2010.01753.x
Maya M. C. I., Torres R Y. E., Galván M. F., Aguiñiga G. S., Trasviña C. L. D. (2016). Trophic overlap between blue sharks (Prionace glauca) and shortfin makos (Isurus oxyrinchus): trophic linkages between two shark species in the Eastern Pacific Ocean. Food Web. 7, 13–19. doi: 10.1016/j.fooweb.2016.03.002
May-Collado L. J., Amador-Caballero M., Casas J. J., Gamboa-Poveda M. P., Garita-Alpízar F., Gerrodette T., et al. (2017). “Ecology and conservation of cetaceans of Costa Rica and Panama,” in Advances in Marine Vertebrate Research in Latin America. Eds. Rossi-Santos M., Finkl C. (Springer, Berlin), 293–319. doi: 10.1007/978-3-319-56985-7_12
May-Collado L. J., Barragán-Barrera D. C., Quiñones-Lebrón S. G., Aquino-Reynoso W. (2012). Dolphin watching boats impact on habitat use and communication of bottlenose dolphins in Bocas del Toro, Panama during 2004, 2006–2010. Int. Whal. Comm SC/64/WW2, 1–8.
May-Collado L. J., Quiñones-Lebrón S. G., Barragán-Barrera D. C., Palacios J. D., Gamboa-Poveda M. (2014). The dolphin watching industry of Bocas del Toro continues impacting the resident bottlenose dolphin population. Int. Whal. Comm SC/65b/WW6, 1–6.
Medina E., Tavares R., Posada J. M. (2009). Resultados Preliminares sobre la Edad y Crecimiento del Cazón Playón, Rhizoprionodon porosus (Poey 1861), en la Región Nororiental de Venezuela. Proc. 62nd Gulf Caribbean Fisheries Institute GCFI, 441–445.
Méndez-Fernández P., Bustamante P., Bode A., Chouvelon T., Ferreira M., López A., et al. (2020). Population structure of the Atlantic spotted dolphin (Stenella frontalis) inferred through ecological markers. Aquat. Ecol. 54, 21–34. doi: 10.1007/s10452-019-09722-3
Mou Sue L. (1993). “Parque nacional marino isla bastimentos,” in Agenda Ecológica y Social para Bocas del Toro. Ed. Moreno S. H. (Impresora Continental, Panamá), 163–173.
Naylor G. J. P., Caira J. N., Jensen K., Rosana K. A. M., White W. T., Last P. R. (2012). A DNA sequence–based approach to the identification of shark and ray species and its implications for global elasmobranch diversity and parasitology. Bull. Am. Mus. Nat. Hist. 367, 1–262. doi: 10.1206/754.1
Neumann R. M., Ward S. M. (1999). Bioaccumulation and biomagnification of mercury in two warmwater fish communities. J. Freshw. Ecol. 14, 487–497. doi: 10.1080/02705060.1999.9663707
O’Bryhim J. R., Adams D. H., Spaet J. L. Y., Mills G., Lance S. L. (2017). Relationships of mercury concentrations across tissue types, muscle regions and fins for two shark species. Environ. pollut. 223, 323–333. doi: 10.1016/j.envpol.2017.01.029
Pacoureau N., Rigby C. L., Kyne P. M., Sherley R. B., Winker H., Carlson J. K., et al. (2021). Half a century of global decline in oceanic sharks and rays. Nature 589, 567–571. doi: 10.1038/s41586-020-03173-9
Plumlee J. D., Wells R. J. D. (2016). Feeding ecology of three coastal shark species in the northwest Gulf of Mexico. Mar. Ecol. Prog. Ser. 550, 163–174. doi: 10.3354/meps11723
Pollom R., Carlson J., Charvet P., Avalos C., Bizzarro J., Blanco-Parra M. P., et al. (2021). Sphyrna tiburo (amended version of 2020 assessment). The IUCN Red List of Threatened Species. doi: 10.2305/IUCN.UK.2021-3.RLTS.T39387A205765567.en
Post D. M., Layman C. A., Arrington D. A., Takimoto G., Quattrochi J., Montana C. J. (2007). Getting to the fat of the matter: models, methods and assumptions for dealing with lipids in stable isotope analysis. Oecologia 152, 179–189. doi: 10.1007/s00442-006-0630-x
Radakov D. V., Silva A. (1974). Some characteristics of the schooling behavior of Jenkinsia lamprotaenia. J. Ichthyol. 14, 283–286.
Reyier E. A., Adams D. H., Lowers R. H. (2008). First evidence of a high density nursery ground for the lemon shark, Negaprion brevirostris, near Cape Canaveral, Florida. Fla. Sci. 71, 134–148.
Seemann J., González C. T., Carballo-Bolaños R., Berry K., Heiss G. A., Struck U., et al. (2013). Assessing the ecological effects of human impacts on coral reefs in Bocas del Toro, Panama. Environ. Monit. Assess. 186, 1747–1763. doi: 10.1007/s10661-013-3490-y
Sigovini M., Keppel E., Tagliapietra D. (2016). Open nomenclature in the biodiversity era. Methods Ecol. Evol. 7, 1217–1225. doi: 10.1111/2041-210X.12594
Sitar A., May-Collado L. J., Wright A. J., Peters-Burton E., Rockwood L., Parsons E. C. M. (2016). Boat operators in Bocas del Toro, Panama display low levels of compliance with national whale-watching regulations. Mar. Policy. 68, 221–228. doi: 10.1016/j.marpol.2016.03.011
Keywords: Sphyrna aff. tiburo, Rhizoprionodon porosus, trophic ecology, Hg, nursery area, Panama, conservation
Citation: Barragán-Barrera DC, Polo-Silva CJ, González C, Rodríguez M, Baldrich L, Bermúdez-Rivas C, Moncaleano-Niño ÁM, Bustamante P, Caballero S and Luna-Acosta A (2024) First stable isotope and mercury assessment of bonnethead and Caribbean sharpnose sharks from a potential nursery ground in the Archipelago of Bocas del Toro, Panamanian Caribbean. Front. Mar. Sci. 11:1235876. doi: 10.3389/fmars.2024.1235876
Received: 06 June 2023; Accepted: 27 February 2024;
Published: 15 April 2024.
Edited by:
Sm Sharifuzzaman, University of Chittagong, BangladeshReviewed by:
Luiz Drude Lacerda, Federal University of Ceara, BrazilCopyright © 2024 Barragán-Barrera, Polo-Silva, González, Rodríguez, Baldrich, Bermúdez-Rivas, Moncaleano-Niño, Bustamante, Caballero and Luna-Acosta. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dalia C. Barragán-Barrera, ZGFsaWFjLmJhcnJhZ2FuYmFycmVyYUBnbWFpbC5jb20=; Ángela Margarita Moncaleano-Niño, YW1vbmNhbGVhbm9AamF2ZXJpYW5hLmVkdS5jbw==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.