 Max L. E. Gräfnings1,2*
Max L. E. Gräfnings1,2* Ise Grimm1
Ise Grimm1 Stephanie R. Valdez3India Findji1
Stephanie R. Valdez3India Findji1 Tjisse van der Heide1,2,4Jannes H. T. Heusinkveld5Kasper J. Meijer1
Tjisse van der Heide1,2,4Jannes H. T. Heusinkveld5Kasper J. Meijer1 Britas Klemens Eriksson1Quirin Smeele6
Britas Klemens Eriksson1Quirin Smeele6 Laura L. Govers1,2,4
Laura L. Govers1,2,4- 1Conservation Ecology Group, Groningen Institute for Evolutionary Life Sciences (GELIFES), University of Groningen, Groningen, Netherlands
- 2Department of Aquatic Ecology and Environmental Biology, Institute for Water and Wetland Research (IWWR), Radboud University, Nijmegen, Netherlands
- 3Division of Marine Science, Nicholas School of the Environment, Duke University, Beaufort, NC, United States
- 4Department of Coastal Systems, Royal Netherlands Institute for Sea Research (NIOZ), Den Burg, Netherlands
- 5The Fieldwork Company, Groningen, Netherlands
- 6Natuurmonumenten, Paterswolde, Netherlands
Ecological restoration has become an important management-tool to counteract the widespread losses of seagrass meadows and their associated biodiversity. In the Dutch Wadden Sea, long-term restoration efforts have recently led to the successful restoration of annual eelgrass (Zostera marina) at high densities on a local scale. However, it is yet unknown if restored seagrass plants also lead to improved local biodiversity and ecosystem functioning in the intertidal zone. We therefore compared the macrozoobenthos communities of a small-scale restored meadow to 22 naturally occurring intertidal seagrass meadows. Using a taxonomic and trait-based approach we aimed to study 1) how intertidal seagrasses (Zostera marina and Zostera noltii) affect benthic communities and their functional trait distribution and 2) if a restored meadow facilitates benthic communities similar to natural meadows. We found that both natural and restored seagrasses increased abundances of benthic animals and the richness (both taxonomic and functional) of associated benthic communities compared to nearby unvegetated areas. Additionally, the presence of intertidal seagrass shifted benthic community composition both taxonomically and functionally, thus broadening the niche space for species inhabiting tidal flats. Seagrasses especially facilitated epifaunal species and traits associated with these animals. Surprisingly, our results indicate that the mere presence of seagrass aboveground structure is enough to facilitate benthic communities, as neither higher seagrass cover nor biomass increased benthic biodiversity in the intertidal zone. By studying the effect of seagrass restoration on benthic diversity, we found that the restored meadow functioned similarly to the natural meadows after only two years and that the success of our restoration efforts indeed led to local biodiversity enhancements. Our findings contribute to the understanding of the ecological functioning of intertidal seagrasses and can be used to define/refine conservation and restoration goals of these valuable ecosystems.
1 Introduction
Seagrasses are marine flowering plants that form vast underwater meadows in coastal waters worldwide. In addition to being some of the most productive ecosystems in the world (Duarte and Chiscano, 1999), these meadows provide essential ecosystem services and contribute to climate change mitigation. For instance, seagrass meadows provide vital nursery habitats for commercially important species, offer coastal protection, and sequester large amounts of carbon (Nordlund et al., 2016). Additionally, seagrass meadows are generally regarded as diversity hotspots, serving as crucial habitats to a wide array of animals, from charismatic mega herbivores to microscopic critters. However, seagrasses are seriously threatened and during the last century an estimated ~29% of the global seagrass area was lost (Orth et al., 2006; Waycott et al., 2009).
Ecological restoration has become an important management-tool, supplementing traditional passive conservation efforts, to counteract declines of seagrasses and other coastal ecosystems. However, coastal ecosystem restoration is still in its infancy and often challenging in practice (Bayraktarov et al., 2016). The reasons hindering successful coastal restoration are many, ranging from practical methodological problems to how the public perceives marine ecosystem restoration (Suding, 2011; Bayraktarov et al., 2016; Abelson et al., 2020). One specific aspect that is often lacking is how restoration success is defined and measured (Abelson et al., 2020). Success is often defined solely based on how the restoration effort affects the target species (Suding, 2011). Although useful as a short-term indicator of success, this is generally an insufficient approach since ecological restoration is about restoring ecosystem integrity and associated ecosystem functions (Suding et al., 2015). Hence, there is a need for monitoring programs to also measure restoration success beyond the target species. To evaluate the success of a restoration project, The Society of Ecological Restoration (SER, 2004; Gann et al., 2019) recommends assessing nine ecosystem attributes including biodiversity, resilience and self-sustainability. Ideally, restoration projects would follow these guidelines, but in reality, monitoring all the proposed ecosystem attributes requires more funds and long-term commitment than restoration projects usually have available. Thus, measurements that combine feasibility with information about the overall performance of the restored ecosystems are urgently needed.
In the Wadden Sea, seagrasses experienced large declines during the 20th century. Vast subtidal eelgrass (Zostera marina) meadows vanished completely in the 1930s, due to the seagrass wasting disease and coastal development (den Hartog and Polderman, 1975; Giesen et al., 1990). Intertidal seagrass populations persisted, but were heavily diminished by increased eutrophication in the 1970s and 80s (van Katwijk et al., 2010). During the last decades, intertidal meadows have recovered in the northern Wadden Sea (Dolch et al., 2013), but in the south (Netherlands and Lower Saxony) recovery has remained almost entirely absent (Dolch et al., 2017). Today, only intertidal seagrasses, perennial dwarf eelgrass (Zostera noltii) and annual common eelgrass (Z. marina), persist in the Wadden Sea. The loss of seagrass meadows has affected the ecosystem functioning and biodiversity of the sea. For instance, seagrass declines have altered sediment dynamics, increased water turbidity, led to the loss of commercially important nursery habitats and reduced food availability for herbivorous birds (Wolff, 1979; Ganter, 2000; Polte et al., 2005; van der Heide et al., 2007; Eriksson et al., 2010).
During the last decades, seagrass restoration efforts have been performed in the Dutch Wadden Sea (e.g., van Katwijk et al., 2009; van Duren and Van Katwijk, 2015; Govers et al., 2022), with the goal to counteract the widespread seagrass losses and to recover lost ecosystem functions. Recently, the persistent restoration efforts have started to pay off, as we have been able to restore high densities of Z. marina on relatively large areas (2664 m² in 2020 and 1 ha in 2021, Gräfnings et al., 2023 & unpublished results) using a newly developed seeding method (DIS-method; Govers et al., 2022; Gräfnings et al., 2022). However, to properly evaluate if the restoration has been successful, further monitoring is needed to confirm that the efforts have also enhanced ecosystem integrity beyond the target species.
In this paper we investigate how seagrass restoration affects benthic communities as an indicator of the degree of restoration success (Dolbeth et al., 2013; Lefcheck et al., 2017). Macrozoobenthic communities are the foundation of many ecosystem services in benthic systems (Snelgrove, 1999). Macrozoobenthic animals can affect their surroundings in multiple ways, from nutrient recycling and oxygenation of sediments to grazing and stabilization of sediments (see review by Levin et al., 2001). Furthermore, many benthic animals function as crucial food resources for higher trophic levels and can thus offer information about cross-ecosystem dynamics. Hence, studying benthic communities can offer valuable information about how restoration efforts affect local biodiversity and ecosystem functioning, especially if complementary taxonomic and functional diversity approaches are used in parallel (Henseler et al., 2019). Traditional taxonomic diversity approaches base their results on species identities, while functional diversity is usually measured through species traits (Violle et al., 2007) and is considered a good predictor of ecosystem function (Cadotte et al., 2011). However, before the effect of the restored seagrass on benthic communities can be properly evaluated in the Wadden Sea, accurate information is needed about how natural seagrass meadows affect benthic communities in the area. Although the effect of seagrasses on benthic diversity (both taxonomic and functional) is well studied (e.g., Orth et al., 1984; Boström and Bonsdorff, 1997; Lefcheck et al., 2017; Boyé et al., 2019), little is known about how intertidal seagrasses affect benthic diversity (especially functional diversity) in the Wadden Sea or in restored seagrass beds.
In this study, we assess 1) how intertidal seagrass (annual Z. marina and/or perennial Z. noltii) presence affect taxonomic and functional diversity (richness, evenness) and community structure (composition) of macrozoobenthos and 2) whether a recently restored Z. marina meadow affects the associated benthic community similar to natural meadows in the intertidal Wadden Sea. We expect intertidal seagrasses to generally facilitate biodiversity, but that the two seagrass species might affect benthic communities differently due to differences in structural attributes and life cycle. For instance, the yearly turnover of annual Z. marina is expected to affect benthic community composition differently than more stable perennial seagrasses like Z. noltii. We hypothesize that mobile benthic species will benefit the most from the short life cycle of annual Z. marina. Additionally, only Z. noltii forms traditional seagrass ‘meadows’ in the Wadden Sea, while monospecific annual Z. marina populations spread out very sparsely (<1 plant/m²) over large areas (personal observation; Figure 1D). The ability of seagrass meadows to shelter organisms (from both predators and physical disturbance) is often considered one of the most important functions facilitating biodiversity (see review by Boström et al., 2006), and in the absence of a meadow-structure the positive effect of seagrasses on the benthic diversity might be reduced. Finally, we expect the restored seagrass to facilitate benthic biodiversity, but potentially to a lesser degree than natural seagrasses, due the short time span after restoration. With our findings we aim to explain the importance of intertidal seagrass ecosystems for associated benthic communities and their functionality, to evaluate seagrass restoration success beyond the target species, and to provide practical recommendations concerning the conservation and restoration of intertidal seagrasses in the Wadden Sea and globally.
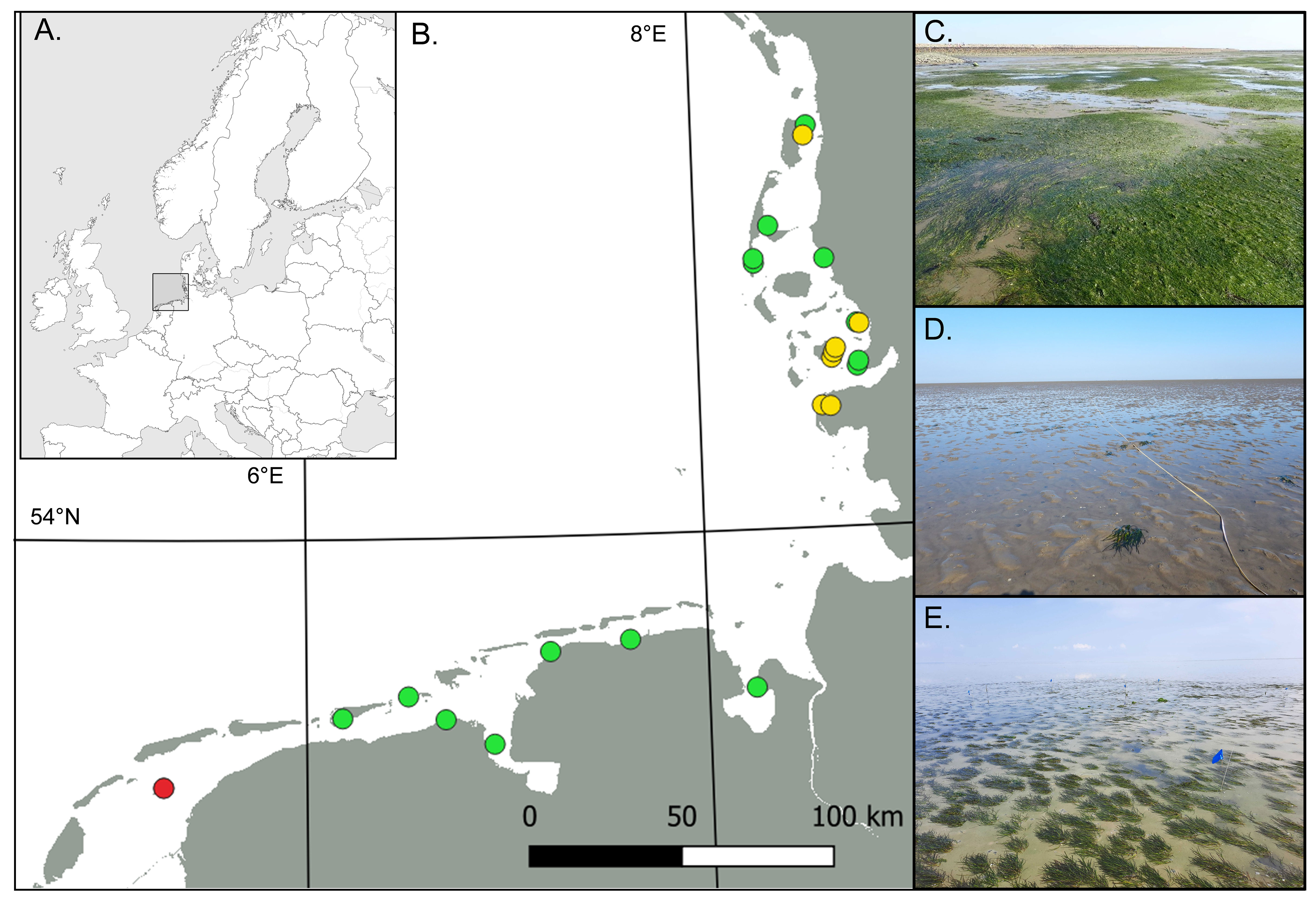
Figure 1 (A) Location of the Wadden Sea in Europe. (B) Locations of sampled seagrass sites in the Wadden Sea where green indicates sites macrozoobenthos was sampled both inside and outside the seagrass and yellow indicates sites macrozoobenthos was sampled only inside the seagrass. The red circle indicates the seagrass restoration site northeast of Griend-island. Pictures on the right show (C) a mixed Zostera noltii & Zostera marina meadow, (D) a monospecific Z. marina meadow and (E) the restoration plot at Griend.
2 Methods
2.1 Study area
The Wadden Sea is a temperate coastal sea that extends from the northwest Netherlands to the southwest coasts of Denmark. The Wadden Sea has a diurnal tidal cycle (range: 1.5 to 4 meters) and during low tide, approximately 50% of the seas total area (~8000 km²) emerges as intertidal flats. These tidal flats provide a hotspot for birds and marine biodiversity.
2.2 Seagrass restoration
Since 2018, eelgrass (Z. marina) restoration trials have been successfully performed northeast of Griend-island (Figure 1; N 53.2692, E 5.2949) in the Dutch Wadden Sea (Gräfnings et al., 2022; Gräfnings et al., 2023). Prior to restoration experiments in 2018, there was no eelgrass growing on the tidal flats surrounding the island. In March 2020, a 400 m² plot was seeded on the Griend sandflat with the DIS-method (Govers et al., 2022; Gräfnings et al., 2022). In the restoration plot, eelgrass seeds were injected to a depth of 3 cm, with ~3 seeds/injection and with a seeding density of 50 injections/m². In July 2020 on average 25.75 eelgrass plants/m² had been established in the 400 m² restoration plot. A year later in July 2021, on average 25 second generation eelgrass plants/m² were counted in the restoration plot.
2.3 Data collection and processing
In the summer (July– August) of 2018, 22 natural seagrass sites were sampled once during low tide across the international Wadden Sea (Figure 1). At each site, we estimated the % cover of Z. marina and Z. noltii by walking three 30 m x 1 m belt transects. At 15 of the sites, we collected samples both from inside and outside (>20 m from the seagrass) of the seagrass. At the remaining 7 sites, samples were only collected inside the seagrass. At each site, three benthic macrofauna cores (top 30 cm & 15.5 cm diameter) were sampled. Cores sampled from within the seagrass meadow always contained seagrass. In the field, the samples were sieved (1 mm), after which any seagrass biomass in the samples was separated and pooled. Macrozoobenthos from the three cores were pooled and stored in 70% ethanol until identification. In July 2021, we sampled six benthic macrofauna cores (top 30 cm & 15.5 cm diameter) inside and outside the seagrass restoration plot at Griend (Figure 1E). Samples were sieved (1 mm) and stored in a 4% formaldehyde solution until identification. The twelve macrofauna cores from Griend were treated as independent samples. In the lab, all individuals were counted and identified to the finest taxonomic level possible under a dissecting microscope. The individuals were identified to 54 different taxonomic groups (35 at species-level, 11 at genus-level (e.g., Eteone sp.), 2 at family-level (e.g., Polynoidae), 3 at order-level (e.g., Actinaria), 2 at phylum-level (e.g., Nematoda) and 1 at class-level (Bivalvia). Once counted and identified, the biomass of individuals of the same taxonomic group was determined per sample. Samples were first dried for 48h at 60°C in a ventilated stove, after which dry weight was measured. Following this, the samples were incinerated for 5 hours at 560°C and then weighed again to obtain the ash free dry mass (AFDM). Seagrass samples were dried for 48 h at 60°C in a ventilated stove, after which the total dry weight of the plants was measured.
2.4 Macrozoobenthos traits
We chose to include traits explaining the size, morphology, mobility, feeding type, burrowing depth and reproduction of the benthic communities (Table 1). These traits were chosen to characterize the basic ecology of the benthic animals and to give an indication how intertidal seagrasses affect benthic ecosystem functioning. All traits were categorical and divided into several modalities (Table 1). The species-specific affinities to all modalities were fuzzy coded within a range from 0 to 3 (0= no affinity, 1= low affinity, 2= moderate affinity and 3= high association of taxon with the trait category). The fuzzy-coded trait values were extracted from two macrozoobenthic biological trait databases (Gusmao et al., 2022; Meijer et al., 2023). To standardize the data between traits with different numbers of modalities, we divided the individual modalities by the total number of modalities of the corresponding trait (see Henseler et al., 2019). As we were not able to identify some bivalves beyond class-level, these individuals were allocated trait information common to the entire class in the Wadden Sea.
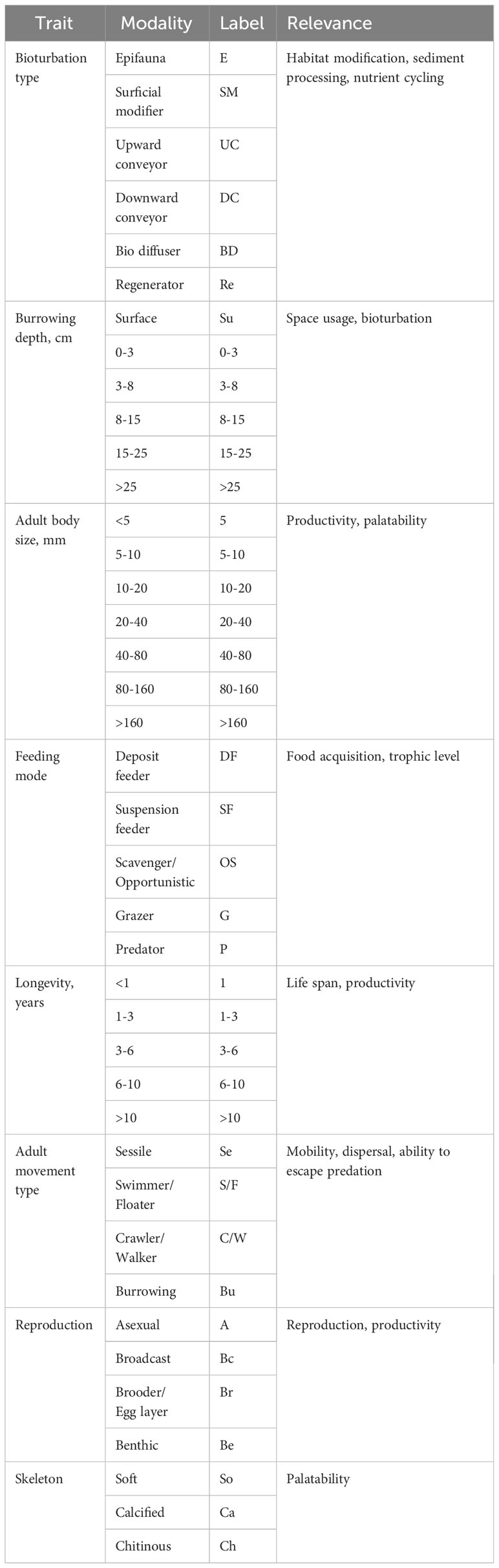
Table 1 List of macrozoobenthos traits and their modalities used in functional trait analysis.
2.5 Statistical analysis
All statistical analyses were performed in R version 4.0.3 (R Core Team, 2020). We investigated the effect of intertidal seagrasses on both taxonomic and functional diversity of macrozoobenthos. To describe taxonomic diversity, we calculated taxonomic richness, Shannon index and Pielou’s evenness using the “vegan”- package (Oksanen et al., 2018). To describe functional diversity, we calculated three trait-based indices (functional richness, evenness and dispersion) corresponding to the previously mentioned taxonomic indices. Functional richness (FRic) expresses the amount of trait space that is occupied by the species of a community (Mason et al., 2005). Functional evenness (FEve), describes how evenly species abundances are distributed between the expressed trait categories (Mason et al., 2005). Functional dispersion (FDis) describes the abundance weighted mean distance of individual species to their weighted group centroid in the multidimensional trait space (Laliberté and Legendre, 2010) and thus measures the spread of the community within the trait space. Hence, functional dispersion can be considered a measure of functional diversity (Laliberté and Legendre, 2010). Additionally, community-level weighted means of trait values (CWM) were computed for each sample. CWMs express trait values weighted by species abundances and can be used to compare how communities differ functionally. Functional diversity indices were calculated with the “FD’’-package (Laliberté and Legendre, 2010; Laliberté et al., 2014). We used count-data (individuals/m²) to calculate the different indices and community-level weighted means. Before calculating the trait-based indices, count-data was log (x+1) transformed to reduce the influence of dominant species without losing the abundance effect.
2.5.1 Analyzing the effect of natural intertidal seagrass on benthic diversity
Differences in benthic diversity indices and abundances (counts and AFDM) between natural seagrass and nearby bare areas were analyzed with linear mixed effect models using the lmer-function in R (package: “lme4”). In the models, we included data from sites where macrozoobenthos was sampled both inside and outside the seagrass (15 sites). No distinction between the two seagrass species was included in these models. “Site” was included as a random factor (random intercept: “1|Site”) in each model. Residuals of the linear mixed effect models were checked for normality and, if necessary, the response variable was transformed to fit model assumptions (log-transformed: Taxonomic richness; sqrt-transformed: FEve & FDis). An ANOVA from the car package (Fox and Weisberg, 2011) was applied to conduct the Wald Chi-Squared test on model outputs.
To investigate if the two seagrass species affect benthic communities differently in the intertidal Wadden Sea, we first divided the sampled seagrass meadows into three categories: Mixed meadows (both seagrass species present; n=5), monospecific Z. marina (n=8) and monospecific Z. noltii (n=9). Data from all sampled seagrass sites (22 sites) were used for the analyses. One-way ANOVAs were performed to analyze if benthic diversity indices differed between seagrass meadow types, after controlling that the data met the assumptions of parametric tests. Additionally, linear regressions were performed to investigate if seagrass cover (%) and biomass (DW, g) affected benthic diversity indices.
2.5.2 Analyzing the effect of restored intertidal seagrass on benthic diversity
Differences in benthic diversity indices (taxonomic richness, Shannon index, Pielou’s evenness FRic, FEve, FDis) and abundances (counts and AFDM) between the restored seagrass plot at Griend (n=6) and nearby bare area (n=6) was analyzed with one-way ANOVAs. Prior to analysis, data were tested to meet the assumptions of parametric tests.
2.5.3 Analyzing the effect of intertidal seagrass on benthic community composition
To assess if benthic communities differed between seagrasses and nearby bare areas, we performed non-metric multidimensional scaling (nMDS) on the taxonomic (based on counts) and functional (based on CWM values) compositions. The replicates from the seagrass restoration site were pooled (seagrass and bare separately) and included in the analysis, to get an indication if restored seagrass affected benthic communities similarly as natural meadows. Bray-Curtis dissimilarity was applied for count-data and the Gower distance for CWM trait values. To statistically test for differences in taxonomic and functional compositions among habitats, we used permutational multivariate ANOVAs (PERMANOVA;9999 permutations; package: “vegan”). Site was included as a random factor in the PERMANOVA analysis. Before PERMANOVAs were performed, we checked with a permutational test of multivariate dispersion (PERMDISP; package: “vegan”) if the observations within groups were spread equally between the habitats. Furthermore, we used Multilevel pattern analysis (package: “indicspecies”) to determine which species and modalities were driving differences between seagrass and bare communities. Multilevel pattern analysis was performed separately for natural meadows (n=15) and the restoration site (n=6).
3 Results
3.1 Effect of (natural) intertidal seagrass on benthic diversity
Benthic taxonomic richness was significantly increased by natural intertidal seagrasses (chi²=7.47, df=1, p=0.006), with seagrasses sheltering on average 33% higher taxonomic richness than nearby bare areas (Figure 2A). The presence of seagrass on average doubled the amount of epifaunal richness (chi²=13.26, df=1, p<0.001), while no significant differences were observed in endobenthic taxonomic richness (Supplementary Figure S1). However, interestingly seagrass presence did not significantly affect either Pielou’s evenness or Shannon index (Figures 2C, E).
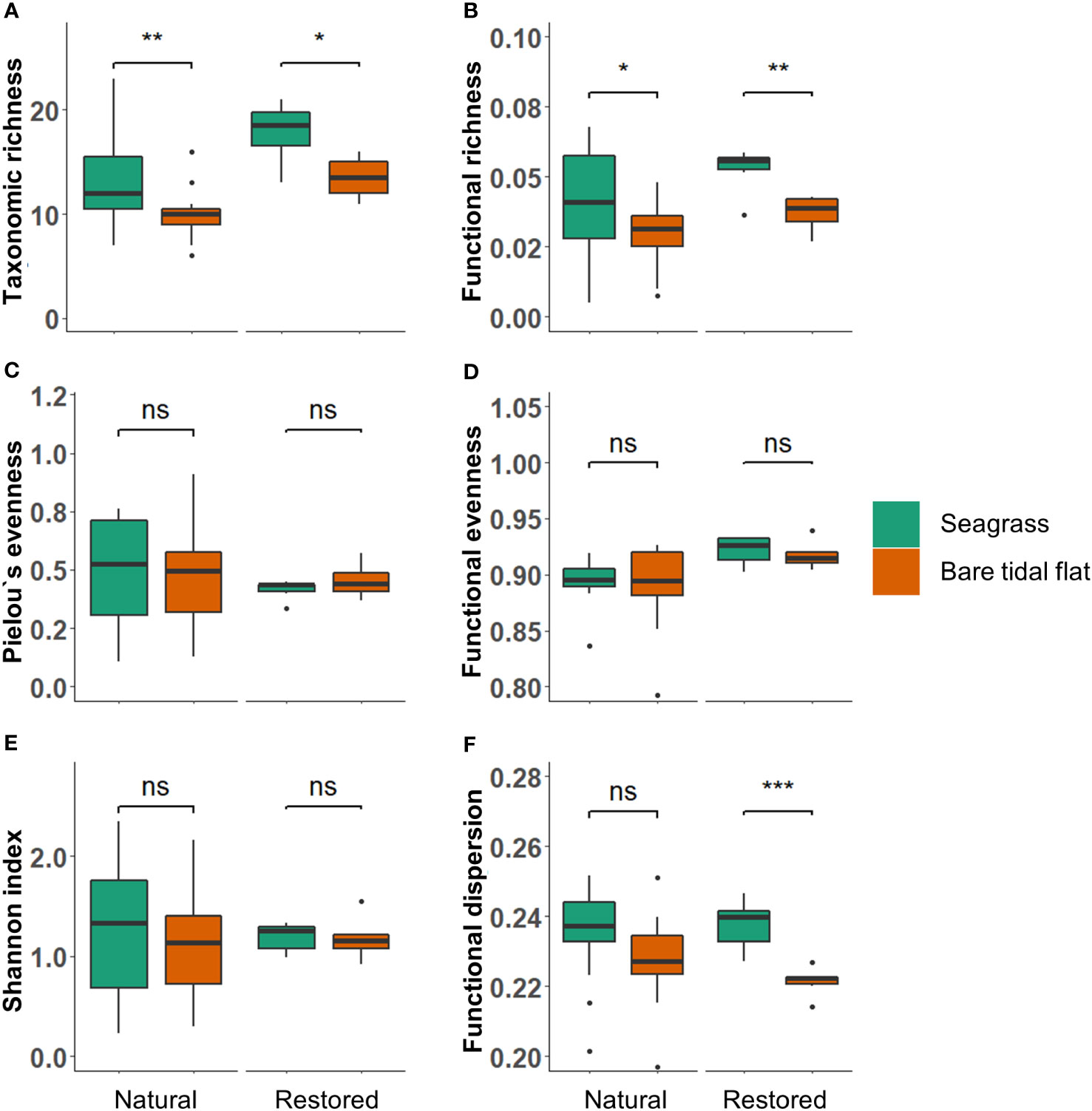
Figure 2 Taxonomic and trait-based indices inside and outside seagrass in natural meadows and in the restored meadow: (A) Taxonomic richness (B) Functional richness, (C) Pielou’s evenness, (D) Functional evenness, (E) Shannon index and (F) Functional dispersion. Boxplots show median (line in box), upper and lower quartile (box), 1.5 x interquartile range (vertical line) and outliers (circle). The indices were calculated with count-data (indviduals/m²). Stars indicate significance of p<0.001 for ***, p<0.01 for ** and p<0.05 for *. Statistically non-significant results are indicated with ns.
The patterns in functional diversity indices largely mirrored their taxonomic counterparts. Functional richness was significantly higher in the presence of intertidal seagrasses (chi²=6.17, df=1, p=0.013; Figure 2B), signifying that the number of expressed traits was increased by seagrass presence. However, neither functional evenness or dispersion of benthic communities differed significantly between seagrasses and bare areas (Figures 2D, F). On average, 170% more benthic individuals inhabited seagrasses than nearby bare areas (chi²=4.87, df=1, p=0.027, Figure 3A), while the animals’ biomass did not differ significantly between the habitats (Figure 3B).
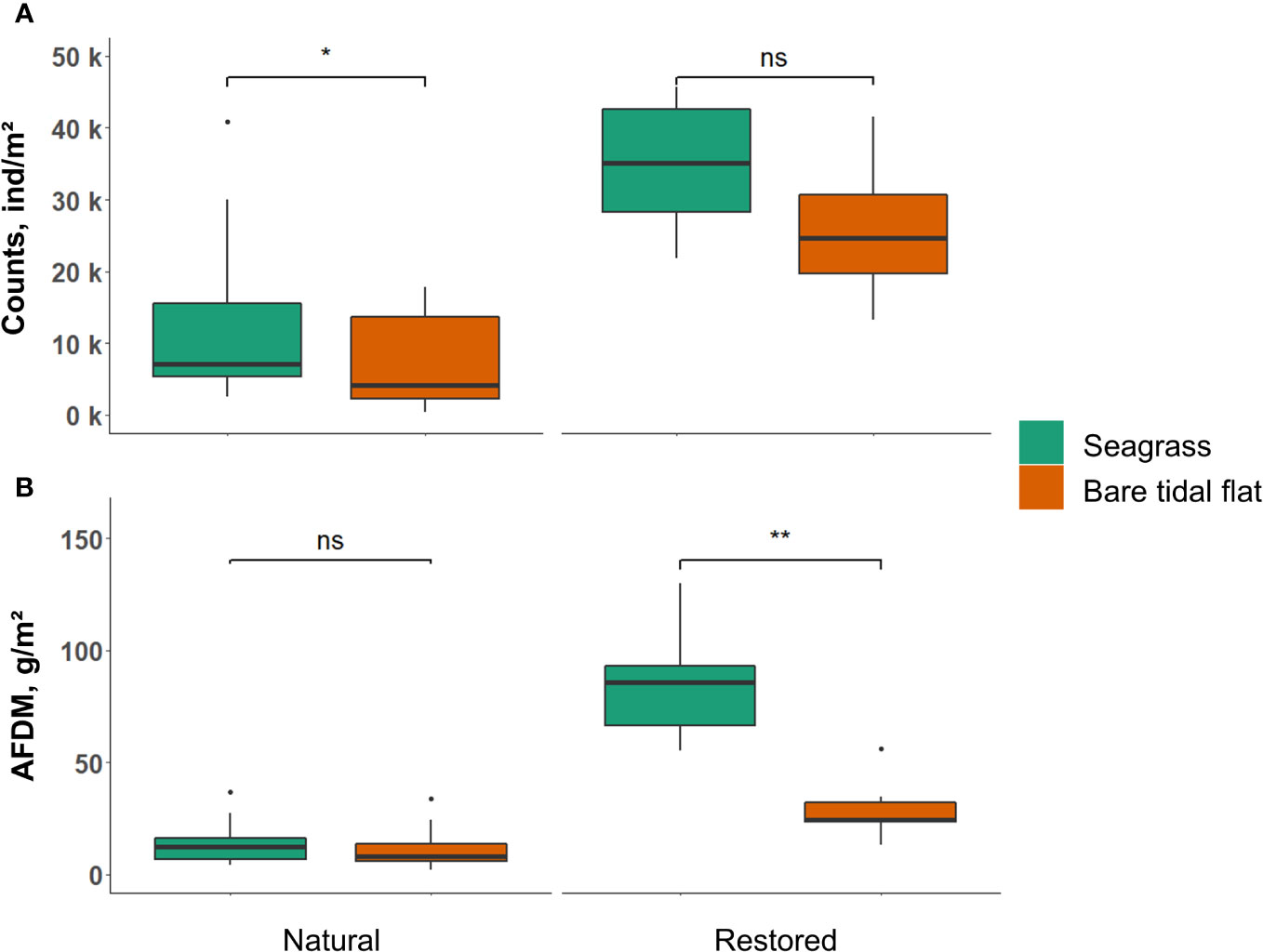
Figure 3 (A) Counts of individuals and (B) AFDM of macrozoobenthos inside and outside seagrass in natural meadows and at the restoration site. Boxplots show median (line in box), upper and lower quartile (box), 1.5 x interquartile range (vertical line) and outliers (circle). Stars indicate significance of p<0.01 for ** and p<0.05 for *. Statistically non-significant results are indicated with ns.
No significant differences were found in macrozoobenthic diversity indices between the three seagrass categories (Mixed meadows, monospecific Z. marina and monospecific Z. noltii; Figure 4). Additionally, no correlation was found between site-wide seagrass cover % and benthic diversity indices (both taxonomic and functional; Supplementary Figures S2A, C, E, G,I, K). Similarly, no correlation was discovered between seagrass biomass (dry weight, g) and benthic diversity indices (both taxonomic and functional; Supplementary Figures S2B, D, F, H, J, L).
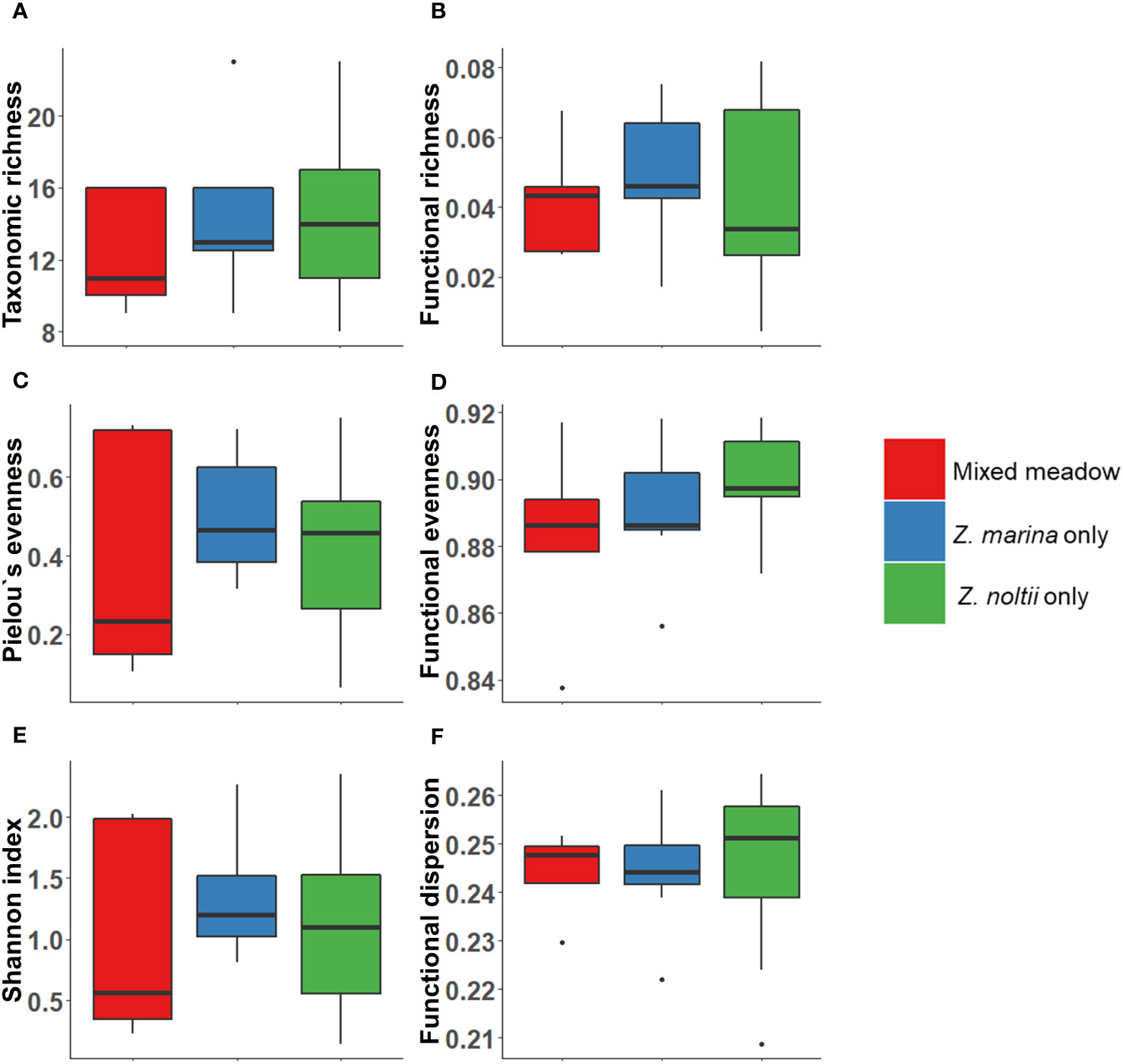
Figure 4 Taxonomic and trait-based indices of different seagrass meadow types [Mixed meadow (both seagrass species present; n=5), monospecific Z. marina (n=8) and monospecific Z. noltii (n=9)] in the Wadden Sea: (A) Taxonomic richness (B) Functional richness, (C) Pielou’s evenness, (D) Functional evenness, (E) Shannon index and (F) Functional dispersion. Boxplots show median (line in box), upper and lower quartile (box), 1.5 x interquartile range (vertical line) and outliers (circle). The indices were calculated with count-data (indviduals/m²). No statistical differences were detected.
3.2 Effect of restored intertidal seagrass on benthic diversity
The recently restored Z. marina meadow at Griend (<2 years) had an almost identical effect on benthic taxonomic diversity as naturally occurring seagrasses in the Wadden Sea. Benthic taxonomic richness was 32% higher in the restored seagrass meadow compared to nearby bare area (F(1,10) = 8.76, p = 0.014; Figure 2A). Similar to natural meadows, the restored seagrass did not affect either Pielou’s evenness or Shannon index significantly (Figures 2C, E). Additionally, the patterns in functional diversity indices largely mirrored their taxonomic counterparts. Similar to natural meadows, functional richness was also higher in the restoration plot compared to nearby bare area (F(1,10) = 13.23, p = 0.004; Figure 2B), signifying that the amount of expressed traits in the benthic community was increased through seagrass restoration. However, unlike in the natural meadows, the restored seagrass also significantly increased functional dispersion of the macrozoobenthos (F(1,10) = 22.23, p < 0.001; Figure 2F), signifying that the diversity of expressed traits was higher in the restoration plots compared to the nearby bare area. Functional evenness of the associated benthic communities did not significantly differ in the restoration plot and nearby bare area (Figure 2D). In contrast to natural seagrasses, we found no difference in animal counts between the restored seagrass and nearby bare area (Figure 3A), while biomass of the benthic community was almost 3x higher in the restored seagrass (F(1,10) = 20.12, p = 0.001; Figure 3B). Mainly three benthic species (mudsnail Peringia ulvae, common cockle Cerastoderma edule and ragworm Hediste diversicolor) were responsible for the higher biomass measured in the restored seagrass.
3.3 Effect of intertidal seagrass on benthic community composition
Benthic community composition was significantly different in the presence of seagrass compared to nearby bare areas (Figure 5), both taxonomically (PERMANOVA; p= 0.04) and functionally (p= 0.001). Interestingly, the restored seagrass meadow seemed to affect the benthic community similarly as natural meadows (Figure 5). Blue mussels (Mytilus edulis), isopods (Idotea spp.) and juvenile shore crabs (Carcinus maenas) functioned as indicator species for seagrass meadows in the Wadden Sea (Multilevel pattern analysis, p<0.05). At the restoration site, blue mussels (Mytilus edulis), mudsnails (Peringia ulvae), nematodes and ragworms (Hediste diversicolor) were indicative of benthic communities in the seagrass, while two polychaeta species (Notomastus latericeus & Heteromastus filiformis) functioned as indicator species for the bare area. Many trait modalities were driving the differences in functional composition between benthic communities in seagrass and bare sites, both on the Wadden Sea scale and at the restoration site (Figure 6). Trait modalities linked to epifaunal species (e.g., sessile, surface living and suspension feeding) were expressed more in seagrass, while the most significant trait modalities telling bare areas apart from seagrasses were: burrowing movement type, deposit feeding and soft skeleton (Figure 6).
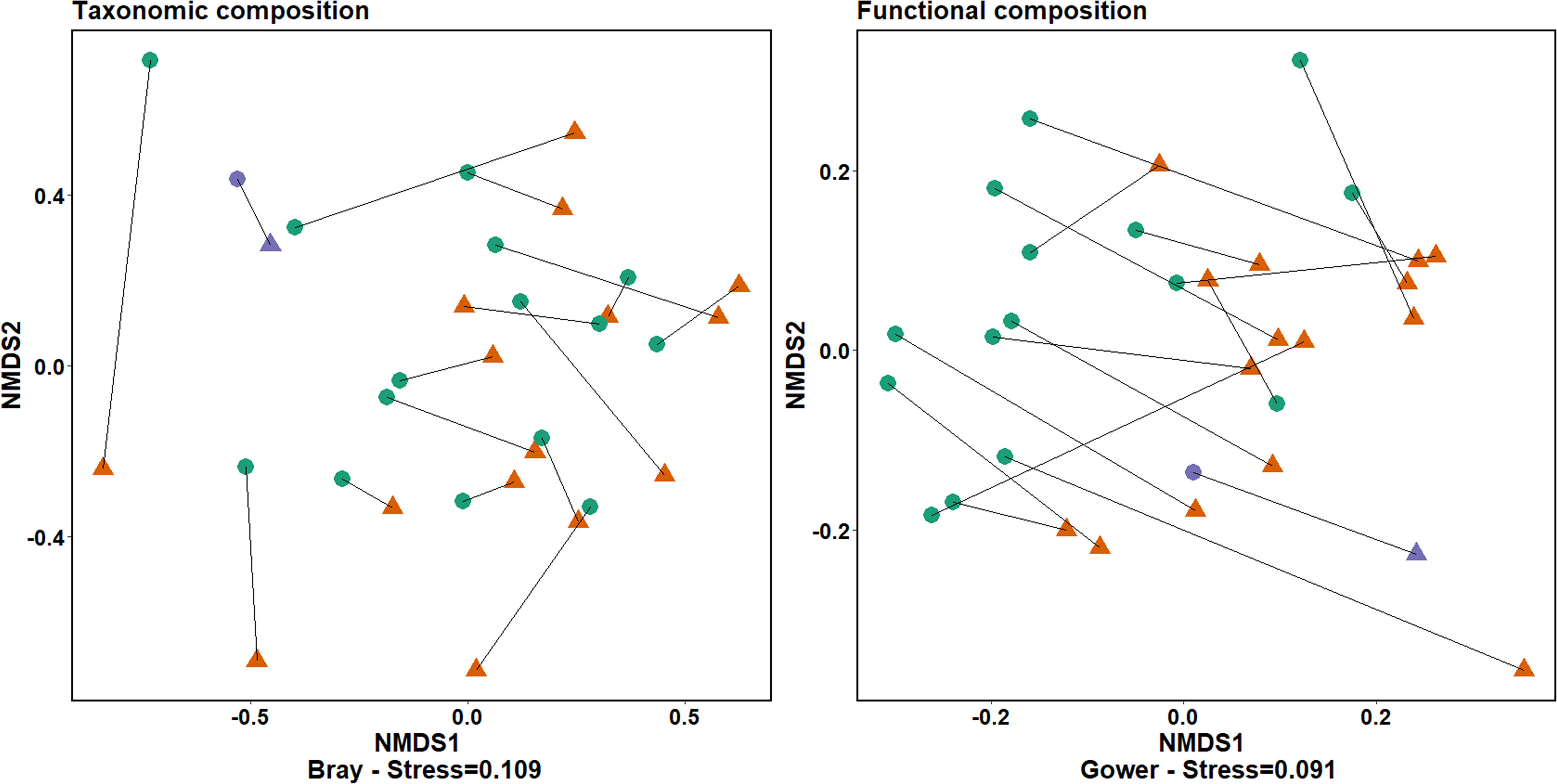
Figure 5 nMDS on taxonomic composition based on count-data and functional composition based on CWM values. Green circles indicate seagrass, orange triangles bare areas and the blue color indicates the two data points of the restoration site. Lines between points signify that data belongs to the same site.
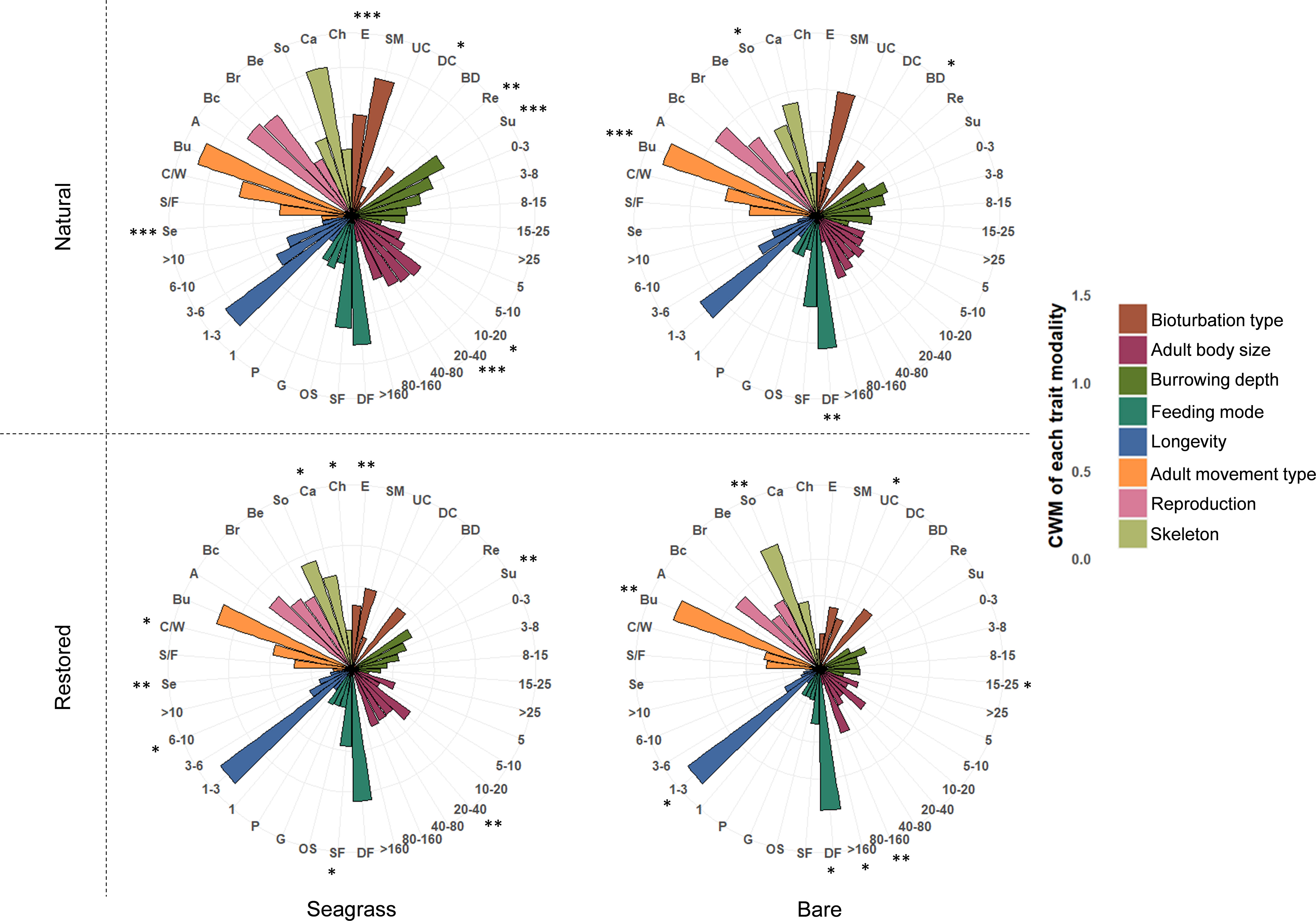
Figure 6 Mean CWM values for benthic communities at seagrass meadows and nearby bare areas in the Wadden Sea and at the restoration site. The stars indicate significant results of the Multilevel pattern analysis highlighting which modalities indicate either seagrass or bare areas. Stars stand for p<0.001 for ***, p<0.01 for ** and p<0.05 for *. For label descriptions, see Table 1.
4 Discussion
Our results show that seagrasses (Zostera marina and Zostera noltii) increase the richness of benthic diversity in the intertidal Wadden Sea. By facilitating epifaunal animals, seagrasses support distinct benthic communities that differ from unvegetated areas both taxonomically and functionally. Importantly, we found that seagrass restoration can quickly (within 2 years) facilitate benthic communities to be taxonomically and functionally similar to communities found in natural seagrass meadows. Hence, we show that the current intertidal seagrass restoration practice in the Dutch Wadden Sea not only leads to restored plants, but also aids in the regeneration of ecosystem integrity through enhancing both taxonomic and functional diversity.
4.1 Intertidal seagrass enhances benthic biodiversity in the Wadden Sea
The effect of seagrasses on biodiversity is well studied and literature shows that the presence of seagrass usually increases the diversity and abundance of benthic fauna (e.g., Orth et al., 1984; Boström et al., 2006). We found that in the intertidal Wadden Sea, seagrasses also support richer (both taxonomically and functionally) and more abundant benthic communities. However, benthic taxonomic richness in the Wadden Sea was lower than what is generally measured in Atlantic seagrass meadows (species richness varying between 10-60; Orth, 1973; Stoner, 1980; Edgar et al., 1994; Blanchet et al., 2004 and references within). This may be the result of challenging living conditions that animals are subjected to in the intertidal Wadden Sea. Seagrasses in the Wadden Sea grow in the upper intertidal zone, which means that the plants and associated communities emerge over the water level for several hours (>5h) each tidal cycle. For most marine species, emergence (even short periods) is very stressful (increased desiccation/temperature) and thus only specialized or resistant benthic species inhabit the area. The ability of seagrasses to modify and ameliorate local conditions is crucial to support rich benthic life in seagrass systems. For instance, the ability of seagrass meadows to provide animals shelter, stabilize sediments, reduce hydrodynamics, aerate sediments and recycle nutrients, are all processes that change living conditions on the bottom of the sea and facilitate macrozoobenthos (Orth et al., 1984; Boström et al., 2006; van der Zee et al., 2016). In the intertidal Wadden Sea, we suggest that seagrasses facilitated species richness by providing shelter (from predators/hydrodynamics/desiccation), food-provisioning and attachment structures to benthic animals.
Interestingly, intertidal seagrasses affected benthic diversity and community compositions very similarly across the Wadden Sea, despite the large spatial scale and differences in seagrass species composition. Worth highlighting is the fact that seagrass presence affected functional compositions more uniformly than their taxonomic counterparts (Figure 5), which suggests that seagrasses facilitate certain traits rather than specific macrozoobenthic species. Functional diversity, and more broadly ecosystem function, is then driven by density shifts in trait representation rather than the presence or absence of traits (e.g., Hewitt et al., 2008). Additionally, we found that higher seagrass cover % and biomass had no detectable influence on benthic diversity. This is somewhat surprising, as the ability of seagrasses to form meadows and modify their environment has previously often been linked with higher benthic richness (Boström et al., 2006 and references within). In our study, monospecific Z. marina meadows with very low cover % (< 1 plant/m²) supported similar benthic communities as dense perennial Z. noltii meadows. Hence our results suggest that the mere presence of seagrass, regardless of species or meadow structure, seems to facilitate benthic biodiversity in the intertidal Wadden Sea. However, seagrasses in the intertidal Wadden Sea provide benthic animals only seasonal hotspots, as during the winter period most aboveground biomass is lost. Thus, benthic communities or at least epifauna need to recolonize the beds each spring. Interestingly, the only benthic species that was exclusively found in samples taken inside the seagrass meadows, was the economically and ecologically important blue mussel (Mytilus edulis). Thus, our results suggest that seagrasses can potentially provide a stepping stone/refuge for mussel spat, facilitating the dispersal of these valuable bivalves.
4.2 Seagrass restoration quickly facilitates benthic communities
Ecological restoration generally aims at restoring ecological integrity by regenerating high biodiversity and lost ecosystem functions. Here, after only two growing seasons a relatively small-scale restoration plot facilitated similar benthic diversity as natural meadows in the Wadden Sea. Our results are especially promising when considering that aquatic restoration efforts often fail to introduce diversity of associated communities to pre-disturbance levels (Rey Benayas et al., 2009). Successful recovery of diversity has most often been observed in systems that are dominated by fast-reproducing organisms with high dispersal ability (Jones and Schmitz, 2009; Duarte et al., 2015). This is most likely also the case in the Wadden Sea, where natural meadows need to be recolonized each spring by epifaunal animals (as discussed above), and one of the main reasons why diversity was able to rebound so quickly at the restoration site. Additionally, most benthic species in this study were found both in seagrass and on the adjacent bare flats, which suggests that intertidal seagrasses do not shelter unique species, but instead provide larger niche spaces with room for richer aggregations. Presumably, mainly by providing aboveground structure and protection, seagrasses facilitate epifaunal animals and traits associated with life above the sediment surface (e.g., sessile, surface living, suspension feeder, Figure 6). Importantly, by investigating the facilitated traits and overall functional diversity of the benthic communities, we could establish that the restored seagrass meadow also functioned very similarly to natural meadows. When comparing restoration efforts over large geographical scales, variation in species assemblages can obscure restoration success, whereas functional trait assemblages may provide a more general indicator of restoration success as they remain more stable over large distances (Bremner, 2008; Hewitt et al., 2008). We concur with previous studies (Dolbeth et al., 2013; Lefcheck et al., 2017) highlighting the usefulness of functional diversity measurements as predictors for seagrass restoration success and urge future restoration projects to incorporate this approach when evaluating restoration success.
4.3 Study limitations
It is important to note the limitations of the study. We only investigated one restoration site, restored with only one of the two native seagrass species and therefore the results of this study cannot be generalized. However, as the included restoration site is the only site where seagrass restoration has been performed successfully in the whole Wadden Sea, we argue that this study should be seen as an important early indication of the value and potential of intertidal seagrass restoration in this area. Additionally, we only investigated the effect of seagrasses on benthic diversity and therefore further research is needed to investigate how the restoration of intertidal seagrass may affect higher trophic levels such as fish and birds. We expect that intertidal seagrasses can also indirectly benefit higher trophic levels (birds and fish) by increasing food availability. For instance, it has been shown that fish diversity is positively influenced by benthic species richness (Lebreton et al., 2012). In light of our results, we expect that especially predators of epifaunal species (e.g., sanderling Calidris alba) and herbivorous birds (e.g., brent geese Branta bernicla) can benefit from seagrass presence in the intertidal Wadden Sea.
4.4 Implications for seagrass conservation and restoration
Our results have important implications for the future of seagrass restoration in the intertidal Wadden Sea. We show that active restoration resulted in the recovery of a threatened marine habitat (intertidal seagrass), restoration of biodiversity and provisioning of habitat. Tangible deliverables like these, can be used to effectively engage stakeholders and to communicate the value of intertidal seagrasses and that restoration of these valuable habitats is possible. Additionally, we argue that dwarf eelgrass (Z. noltii) should be better incorporated in future restoration efforts in the Wadden Sea, as our results show that the smaller seagrass increases benthic diversity to a similar degree as Z. marina. Thus far restoration efforts in the Dutch Wadden Sea have primarily targeted Z. marina, but considering our results we argue that Z. noltii should also get a more visible role in conservation and restoration practices, due to the species ecological importance and ability to create stable perennial meadows.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
MG: Conceptualization, Formal analysis, Investigation, Visualization, Writing – original draft, Writing – review & editing. IG: Formal analysis, Investigation, Writing – review & editing. SV: Investigation, Writing – review & editing. IF: Investigation, Writing – review & editing. TV: Conceptualization, Funding acquisition, Supervision, Writing – review & editing. JH: Conceptualization, Investigation, Writing – review & editing, Funding acquisition. KM: Resources, Writing – review & editing. BE: Resources, Writing – review & editing. QS: Funding acquisition, Writing – review & editing. LG: Conceptualization, Funding acquisition, Investigation, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This project was funded by Waddenfonds grant ‘Sleutelen aan zeegrasherstel’ and European Union’s Horizon 2020 research and innovation program (project: MERCES, Marine Ecosystem Restoration in Changing European Seas; grant agreement #689518).
Acknowledgments
We acknowledge the benthos-lab of NIOZ and especially Loran Kleine Schaars for their help with macrozoobenthos identification. Lisa Bruil is acknowledged for her help in the field. We would like to thank the reviewers for their helpful comments that improved this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2023.1294845/full#supplementary-material
Supplementary Figure 1 | Epi- and endobenthic taxonomic richness inside and outside natural seagrass meadows and the restored meadow. Boxplots show median (line in box), upper and lower quartile (box), 1.5 x interquartile range (vertical line) and outliers (circle). Stars indicate significance of p<0.001 for ***, p<0.01 for ** and p<0.05 for *.
Supplementary Figure 2 | Relationships between seagrass cover % and benthic (A) taxonomic richness, (C) functional richness, (E) Pielou’s evenness, (G) Functional evenness, (I) Shannon index and (K) Functional dispersion in the intertidal Wadden Sea. Relationships between seagrass dry weight (g) and benthic (B) taxonomic richness (D) functional richness, (F) Pielou’s evenness, (H) Functional evenness, (J) Shannon index and (L) Functional dispersion in the intertidal Wadden Sea.
References
Abelson A., Reed D. C., Edgar G. J., Smith C. S., Kendrick G. A., Orth R. J., et al. (2020). Challenges for restoration of coastal marine ecosystems in the anthropocene. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.544105
Bayraktarov E., Saunders M. I., Abdullah S., Mills M., Beher J., Possingham H. P., et al. (2016). The cost and feasibility of marine coastal restoration. Ecol. Appl. 26, 1055–1074. doi: 10.1890/15-1077
Blanchet H., de Montaudouin X., Lucas A., Chardy P. (2004). Heterogeneity of Macrozoobenthic Assemblages within a Zostera noltii Seagrass Bed: Diversity, Abundance, Biomass and Structuring Factors. Estuarine. Coast. Shelf. Sci. 61, 111–123. doi: 10.1016/j.ecss.2004.04.008
Boström C., Bonsdorff E. (1997). Community structure and spatial variation of benthic invertebrates associated with Zostera marina (L.) beds in the northern Baltic Sea. J. Sea. Res. 37 (1), 153–166. doi: 10.1016/S1385-1101(96)00007-X
Boström C., Jackson E., Simenstad C. A. (2006). Seagrass landscapes and their effects on associated fauna: A review. Estuarine. Coast. Shelf. Sci. 68, 383–403. doi: 10.1016/j.ecss.2006.01.026
Boyé A., Thiébaut É., Grall J., Legendre P., Broudin C., Houbin C., et al. (2019). Trait-based approach to monitoring marine benthic data along 500 km of coastline. Diversity Distributions. 25 (12), 1879–1896. doi: 10.1111/ddi.12987
Bremner J. (2008). Species’ traits and ecological functioning in marine conservation and management. J. Exp. Mar. Biol. Ecol. 366, 37–47. doi: 10.1016/j.jembe.2008.07.007
Cadotte M., Carscadden K., Mirotchnick N. (2011). Beyond species: functional diversity and the maintenance of ecological processes and services. J. Appl. Ecol. 48, 1079–1087. doi: 10.1111/j.1365-2664.2011.02048.x
den Hartog C., Polderman P. J. G. (1975). Changes in the seagrass populations of the Dutch Wadden Sea. Aquat. Bot. 1, 141–147. doi: 10.1016/0304-3770(75)90019-4
Dolbeth M. P. C., Grilo T., Raffaelli D., Pardal M. A. (2013). Drivers of estuarine benthic species distribution patterns following a restoration of a seagrass bed: a functional trait analyses. Mar. pollut. Bull. 72, 47–54. doi: 10.1016/j.marpolbul.2013.05.001
Dolch T., Buschbaum C., Reise K. (2013). Persisting intertidal seagrass beds in the northern Wadden Sea since the 1930s. J. Sea. Res. pp, 134–141. doi: 10.1016/j.seares.2012.04.007
Dolch T., Folmer E. O., Frederiksen M. S., Herlyn M., van Katwijk M. M., Kolbe K., et al (2017). Seagrass. In: Wadden Sea Quality Status Report Eds.: Kloepper, S., et al., Common Wadden Sea Secretariat, Wilhelmshaven, Germany. Last updated 21.12.2017. Downloaded 01.06.2023. qsr.waddensea-worldheritage.org/reports/seagrass
Duarte C. M., Borja A., Carstensen J., Elliott M., Krause-Jensen D., Marbà N. (2015). Paradigms in the recovery of estuarine and coastal ecosystems. Estuaries. Coasts. 38, 1202–1212. doi: 10.1007/s12237-013-9750-9
Duarte C. M., Chiscano C. L. (1999). Seagrass biomass and production: A reassessment. Aquat. Bot. 65, 159–174. doi: 10.1016/S0304-3770(99)00038-8
Edgar G. J., Shaw C., Watsona G. F., Hammond L. S. (1994). Comparisons of species richness, size-structure and production of benthos in vegetated and unvegetated habitats in Western Port, Victoria. J. Exp. Mar. Biol. Ecol. 176 (2), 201–226. doi: 10.1016/0022-0981(94)90185-6
Eriksson B. K., van der Heide T., van de Koppel J., Piersma T., van der Veer H. W., Olff H. (2010). Major changes in the ecology of the Wadden Sea: human impacts, ecosys- tem engineering and sediment dynamics. Ecosystems 13, 752–764. doi: 10.1007/s10021-010-9352-3
Fox J., Weisberg S. (2011). An {R} Companion to applied regression. 2nd ed. (Thousand Oaks CA: Sage).
Gann G. D., McDonald T., Walder B., Aronson J., Nelson C. R., Jonson J., et al. (2019). International principles and standards for the practice ofecological restoration. Second. edition. Restor. Ecol. 27, S1–S46. doi: 10.1111/rec.13035
Ganter B. (2000). Seagrass (Zostera spp.) as food for Brent geese (Branta bernicla): an overview. Helgoland. Mar. Res. 54, 63–70. doi: 10.1007/s101520050003
Giesen W. B. J. T., van Katwijk M. M., den Hartog C. (1990). Eelgrass condition and turbidity in the Dutch Wadden Sea. Aquat. Bot. 37, 71–85. doi: 10.1007/BF02743048
Govers L. L., Heusinkveld J. H. T., Gräfnings M. L. E., Smeele Q., van der Heide T. (2022). Adaptive intertidal seed-based seagrass restoration in the Dutch Wadden Sea. PloS One 17 (2), e0262845. doi: 10.1371/journal.pone.0262845
Gräfnings M. L. E., Heusinkveld J. H. T., Hijner N., Hoeijmakers D. J. J., Smeele Q., Zwarts M., et al. (2023). Spatial design improves efficiency and scalability of seed-based seagrass restoration. J. Appl. Ecol 60, 967–977. doi: 10.1111/1365-2664.14405
Gräfnings M. L. E., Heusinkveld J. H. T., Hoeijmakers D. J. J., Smeele Q., Wiersema H., Zwarts M., et al. (2022). Optimizing seed injection as a seagrass restoration method. Restor. Ecol. 31, e13851. doi: 10.1111/rec.13851
Gusmao J. B., Thieltges D. W., Dekker R., Govers L. L., Meijer K. J., Eriksson B. K. (2022). Animal-sediment relationships at the assemblage level: comparing taxonomic and functional trait diversity in marine macrozoobenthos along sediment texture gradient. Ecol. Indicators. 145, 109718. doi: 10.1016/j.ecolind.2022.109718
Henseler C., Nordström M. C., Törnroos A., Snickars M., Pecuchet L., Lindegren M., et al. (2019). Coastal habitats and their importance for the diversity of benthic communities: A species- and trait-based approach. Estuarine. Coast. Shelf. Sci. 226, 106272. doi: 10.1016/j.ecss.2019.106272
Hewitt J. E., Thrush S. F., Dayton P. D. (2008). Habitat variation, species diversity and ecological functioning in a marine system. J. Exp. Mar. Biol. Ecol. 366, 116–122. doi: 10.1016/j.jembe.2008.07.016
Jones H. P., Schmitz O. J. (2009). Rapid recovery of damaged ecosystems. PloS One 4 (5), e5653. doi: 10.1371/journal.pone.0005653
Laliberté E., Legendre P. (2010). A distance-based framework for measuring functional diversity from multiple traits. Ecology 91, 299–305. doi: 10.1890/08-2244.1
Laliberté E., Legendre P., Shipley B. (2014). FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. R package version 1.0-12. http://CRAN.R-project.org/package=FD
Lebreton B., Richard P., Galois R., Radenac G., Brahmia A., Colli G., et al. (2012). Food sources used by sedi- ment meiofauna in an intertidal Zostera noltii seagrass bed: a seasonal stable isotope study. Mar. Biol. 159, 1537–1550. doi: 10.1007/s00227-012-1940-7
Lefcheck J. S., Marion S. R., Orth R. J. (2017). Restored eelgrass (Zostera marina L.) as a refuge for epifaunal biodiversity in mid-western atlantic coastal bays. Estuaries. Coasts. 40 (1), 200–212. doi: 10.1007/s12237-016-0141-x
Levin L. A., Boesch D. F., Covich A., Dahm C., Erseus C., Ewel K. C., et al. (2001). The function of marine critical transition zones and the importance of sediment biodiversity. Ecosystems 4, 430–451. doi: 10.1007/s10021-001-0021-4
Mason N. W. H., Mouillot D., Lee W. G., Wilson J. B. (2005). Functional richness, func- tional evenness and functional divergence: the primary components of functional diversity. Oikos 111, 112–118. doi: 10.1111/j.0030-1299.2005.13886.x
Meijer K. J., Gusmao J. B., Bruil L., Franken O., Grimm I. A., van der Heide T., et al. (2023). The seafloor from a trait perspective. A comprehensive life history dataset of soft sediment macrozoobenthos. Sci. Data 10, 808. doi: 10.1038/s41597-023-02728-5
Nordlund L. M., Koch E. W., Barbier E. B., Creed J. C. (2016). Seagrass ecosystem services and their variability across genera and geographical regions. PloS One 11, e0163091. doi: 10.1371/journal.pone.0163091
Oksanen J., Blanchet F. G., Friendly M., Kindt R., Legendre P., McGlinn D., et al. (2018). Vegan: community ecology package. R package version 2.5.3. https://CRAN.R-project.org/package=vegan.
Orth R. J. (1973). Benthic infauna of eelgrass, Zostera marina, beds. Chesapeake. Sci. 14, 258–269. doi: 10.2307/1350754
Orth R. J., Carruthers T. J., Dennison W. C., Duarte C. M., Fourqurean J. W., Heck K. L., et al. (2006). A global crisis for seagrass ecosystems. Bioscience 56, 987–996. doi: 10.1641/0006-3568(2006)56[987:AGCFSE]2.0.CO;2
Orth R. J., Heck K. L., van Montfrans J. (1984). Faunal communities in seagrass beds: A review of the influence of plant structure and prey characteristics on predator-prey relationships. Estuaries 7 (4), 339–350. doi: 10.2307/1351618
Polte P., Schanz A., Asmus H. (2005). The contribution of seagrass beds (Zostera noltii) to the function of tidal flats as a juvenile habitat for dominant, mobile epibenthos in the Wadden Sea. Mar. Biol. 147, 813–822. doi: 10.1007/s00227-005-1583-z
R Core Team (2020). R: A Language and Environment for Statistical Computing. R Foundation Stat. Computing (Vienna, Austria). https://www.r-project.org/.
Rey Benayas J. M., Newton A. C., Diaz A., Bullock J. M. (2009). Enhancement of biodiversity and ecosystem services by ecological restoration: a meta-analysis. Science 325, 1121–1124. doi: 10.1126/science.1172460
SER, Society for Ecological Restoration International Science & Policy Working Group (2004). Society for ecological restoration international, https://www.ser.org & Tucson: Society for Ecological Restoration International.
Snelgrove P. V. R. (1999). Getting to the Bottom of Marine Biodiversity: Sedimentary Habitats: Ocean bottoms are the most widespread habitat on Earth and support high biodiversity and key ecosystem services. BioScience 49 (2), 129–138. doi: 10.2307/1313538
Stoner A. W. (1980). The role of seagrass biomass in the organization of benthic macrofaunal assemblages. Bull. Mar. Sci. 30 (3), 537–551.
Suding K. N. (2011). Toward an era of restoration in ecology: Success, failures, and opportunities ahead. Annu. Rev. Ecol. Evol. Syst. 42, 465–487. doi: 10.1146/annurev-ecolsys-102710-145115
Suding K., Higgs E., Palmer M., Callicott J.B., Anderson C.B., Baker M., et al. (2015). Committing to ecological restoration. Science 348, 638–640. doi: 10.1126/science.aaa4216
van der Heide T., van Nes E., Geerling G., Smolders A., Bouma T. J., van Katwijk M. (2007). Positive feedbacks in seagrass ecosystems: implications for success in conservation and restoration. Ecosystems 10, 1311–1322. doi: 10.1371/journal.pone.0016504
van der Zee E. M., Angelini C., Govers L. L., Christianen M. J. A., Altieri A. H., van der Reijden K., et al. (2016). How habitat-modifying organisms structure the food web of two coastal ecosystems. Proc. R. Soc. B: Biol. Sci. 283 (1826), 20152326. doi: 10.1098/rspb.2015.2326
van Duren L., Van Katwijk M. M. (2015). Herstelmaatregel groot zeegras in de Nederlandse Waddenzee. 1203892-000. Deltares.
van Katwijk M. M., Bos A. R., de Jonge V. N., Hanssen L. S. A. M., Hermus D. C. R., de Jong D. J. (2009). Guidelines for seagrass restoration: importance of habitat selection and donor population, spreading of risks, and ecosystem engineering effects. Mar. pollut. Bull. 58 (2), 179–188. doi: 10.1016/j.marpolbul.2008.09.028
van Katwijk M. M., Bos A. R., Kennis P., de Vries R. (2010). Vulnerability to eutrophication of a semi-annual life history: a lesson learnt from an extinct eelgrass (Zostera marina) population. Biol. Conserv. 143, 248–254. doi: 10.1016/j.biocon.2009.08.014
Violle C., Navas M.-L., Vile D., Kazakou E., Fortunel C., Hummel I., et al. (2007). Let the concept of trait be functional! Oikos 116, 882–892. doi: 10.1111/j.2007.0030-1299.15559.x
Waycott M., Duarte C. M., Carruthers T. J. B., Orth R. J., Dennison W. C., Olyarnik S., et al. (2009). Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. 106, 12377–12381. doi: 10.1073/pnas.0905620106
Keywords: seagrass, ecological restoration, biodiversity, Wadden Sea, functional diversity, macrozoobenthos
Citation: Gräfnings MLE, Grimm I, Valdez SR, Findji I, van der Heide T, Heusinkveld JHT, Meijer KJ, Eriksson BK, Smeele Q and Govers LL (2024) Restored intertidal eelgrass (Z. marina) supports benthic communities taxonomically and functionally similar to natural seagrasses in the Wadden Sea. Front. Mar. Sci. 10:1294845. doi: 10.3389/fmars.2023.1294845
Received: 15 September 2023; Accepted: 31 December 2023;
Published: 18 January 2024.
Edited by:
Shaochun Xu, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Arnaldo Marín, University of Murcia, SpainJennifer Li Ruesink, University of Washington, United States
Copyright © 2024 Gräfnings, Grimm, Valdez, Findji, van der Heide, Heusinkveld, Meijer, Eriksson, Smeele and Govers. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Max L. E. Gräfnings, bWF4Z3JhZm5pbmdzQGdtYWlsLmNvbQ==