 Simone Panigada1,2*
Simone Panigada1,2* Nino Pierantonio1*
Nino Pierantonio1* Hélder Araújo3
Hélder Araújo3 Léa David4
Léa David4 Nathalie Di-Méglio4
Nathalie Di-Méglio4 Ghislain Dorémus5
Ghislain Dorémus5 Joan Gonzalvo1
Joan Gonzalvo1 Draško Holcer6,7
Draško Holcer6,7 Sophie Laran5
Sophie Laran5 Giancarlo Lauriano8
Giancarlo Lauriano8 Romulus-Marian Paiu9,10
Romulus-Marian Paiu9,10 Morgane Perri11
Morgane Perri11 Dimitar Popov12,13
Dimitar Popov12,13 Vincent Ridoux5,14
Vincent Ridoux5,14 José Antonio Vázquez15
José Antonio Vázquez15 Ana Cañadas16
Ana Cañadas16- 1Tethys Research Institute, Milano, Italy
- 2ACCOBAMS, Monaco, Monaco
- 3Biology Department & ECOMARE, University of Aveiro, Aveiro, Portugal
- 4EcoOcéan Institut, Montpellier, France
- 5Observatoire Pelagis, UAR 3462 La Rochelle Université-CNRS, La Rochelle, France
- 6Blue World Institute of Marine Research and Conservation, Veli Losinj, Croatia
- 7Croatian Natural History Museum, Zagreb, Croatia
- 8Institute for Environmental Protection and Research (ISPRA), Roma, Italy
- 9Mare Nostrum NGO, Constanta, Romania
- 10Faculty of Biology, Bucharest University, Bucharest, Romania
- 11Association AL LARK, Cancale, France
- 12Green Balkans NGO, Plovdiv, Bulgaria
- 13Department of Zoology, Faculty of Biology, Plovdiv University, Plovdiv, Bulgaria
- 14Centre d’Etudes Biologiques de Chizé, UMR 7273 - CNRS - La Rochelle Université, La Rochelle, France
- 15Marine Mammal Group, Oceanographic Research Center of Vigo, Marine Environment Department, Spanish Institute of Oceanography – Spanish National Research Council, Vigo, Spain
- 16Marine Geospatial Ecology Lab, Duke University, Durham, NC, United States
The “ACCOBAMS Survey Initiative” (ASI) is a pilot programme aimed at establishing an integrated and coordinated monitoring system for cetaceans across the Agreement on the Conservation of Cetaceans of the Black Sea, Mediterranean Sea and contiguous Atlantic (hereafter “ACCOBAMS”) area. Conducted in coordination with Mediterranean coastal countries, it supports the implementation of European and regional policies, in particular the EU Marine Strategy Framework Directive and the Ecosystem Approach process. In summer 2018, a synoptic survey was conducted across the Mediterranean Sea and contiguous Atlantic area, combining visual monitoring from aircrafts with visual and passive acoustic monitoring from vessels. Species density and abundance were estimated through design-based approach in a line-transect sampling framework. Based on data arising from the aerial survey only, uncorrected design-based abundance was obtained for striped (N=426,744, CV=0.13), common (N=65,359, CV=0.4), bottlenose (N=63,333, CV=0.17), and Risso´s dolphins (N=26,006, CV=0.3), Cuvier’s beaked whales (N=2,929, CV=0.4) and long-finned pilot whales (N=5,540 CV=0.4). A merged category of either striped or common dolphins resulted in 212,828 individuals (CV=0.26). Fin whales abundance of 1,749 animals (CV=0.3) was corrected for both availability and perception biases and resulted in 3,282 (CV=0.31). The ASI survey offers an overall picture of the distribution and abundance of cetaceans throughout the Mediterranean basin, providing robust estimates to be considered as a baseline for future regional systematic monitoring programmes. The ASI survey is the first step towards establishing a long-term monitoring program across the entire ACCOBAMS area, and, as such, it sets the basis for further future basin-wide monitoring efforts using systematic, shared, coordinated and comparable methods. The information gathered will further enhance knowledge on cetacean status, facilitating the development of informed conservation and mitigation measures, as well as supporting the implementation of international obligations. Furthermore, the outcomes of this survey will support both place- and threat-based conservation efforts in the ACCOBAMS area, through the identification of Important Marine Mammal Areas and Cetacean Critical Habitats. Here the results of the ASI survey are presented and discussed alongside proposed management and conservation actions aimed at ensuring the persistence of cetacean populations in the region.
1 Introduction
The need for monitoring programmes at large spatial and temporal scales aimed at assessing changes in species distributions and abundances, and to predict long-term biological responses to anthropogenic pressures and global changes is widely recognized (Balmford et al., 2005; Green et al., 2005; Pereira and Cooper, 2006). These programmes are also crucial to inform and implement conservation actions and to evaluate the efficacy of management (Grand et al., 2007). However, it is difficult to obtain robust estimates of these population parameters and assess changes for highly mobile and cryptic species, such as cetaceans, whose ranges extend over large areas and occurrence can show strong temporal and spatial patterns (Hughes et al., 2011; Thomas et al., 2015; Guerra et al., 2019; Nykänen et al., 2020).
In the Mediterranean and Black Seas, hotspots of biodiversity (Coll et al., 2010) overlap with high volumes of human activities. The Agreement on the Conservation of Cetaceans of the Black Sea, Mediterranean Sea and Contiguous Atlantic Area (ACCOBAMS) has historically recognized the need for robust baseline data on the conservation status of cetaceans across the Agreement Area, where the overall human impact on the environment is believed to be high (Micheli et al., 2013; Halpern et al., 2015; Stock et al., 2018; Halpern et al., 2019) and where cetacean populations are threatened (Notarbartolo di Sciara, 2016). The ACCOBAMS Secretariat and Scientific Committee therefore proposed and developed the ACCOBAMS Survey Initiative (hereafter ‘ASI’), as the first synoptic survey of the entire ACCOBAMS Region aimed to establish an integrated, collaborative and coordinated monitoring system of the status of cetacean populations across the ACCOBAMS Area (ACCOBAMS Resolutions 6.13, 8.10). The ASI also contained a strong capacity building, sharing and training component that will ultimately strengthen conservation and management in the Region. Whilst primarily developed to further knowledge of cetaceans, the ASI also makes an important contribution to the fulfilment of European Union (EU) Regulations and Directives, specifically the Habitats and the Marine Strategy Framework Directives (Authier et al., 2017), as well as other relevant policy frameworks such as the “Protocol concerning Specially Protected Areas and Biological Diversity in the Mediterranean” of the Barcelona Convention, the “Convention on the Conservation of Migratory Species and Wild Animals” (also known as CMS or Bonn Convention), and the “Berne Convention on the Conservation of European Wildlife and Natural Habitats”.
The ASI officially started at the Sixth Meeting of the Parties to ACCOBAMS (Monaco, 22-25 November 2016) whereas data collection took place in summer 2018 and 2019. Following well-established large-scale monitoring initiatives, such as the Small Cetaceans in European Atlantic waters and the North Sea (SCANS; Hammond et al., 2013), the primary approach was to undertake visual line-transect distance sampling aerial surveys, complemented by ship-based visual and acoustic distance sampling surveys (Boisseau et al., 2023), to maximize survey effort and area coverage. Data were collected for all the cetacean species occurring in the Agreement area (ACCOBAMS, 2021a). Data on other megafauna taxa were also recorded including marine turtles (DiMatteo et al., 2022), and fish (Cañadas et al., 2023). In addition, information on the presence, distribution, and levels of human activities (e.g., floating macro-marine litter, marine traffic, etc.) was collected (Lambert et al., 2020; Nivière et al., 2024).
The ASI is not the first example of such large-scale synoptic surveys, but the inherent and geopolitical complexity of the Mediterranean Sea, with 21 countries bordering its basin, make the ASI a unique case worldwide. Moreover, the fact that the ASI survey spanned across the entire Mediterranean ecoregion, recognized to be a single biogeochemical province (Longhurst, 2007), makes the programme unique in that it attempts to assess and gain knowledge on all cetacean species and their suitable habitats in the entire ecological region.
In this context, the ASI plays an important collaborative role in gathering baseline data for the wide range of cetaceans known to inhabit the Mediterranean Sea, a region where systematic research effort is spatially limited (Mannocci et al., 2018), and gaps exist in the current knowledge of species occurrence, distribution and abundance (Cañadas et al., 2023).
This paper presents uncorrected – apart for fin whales (Balaenoptera physalus) – design-based abundance and density estimates (i.e., estimates are not corrected for perception and availability biases) of cetacean species in the Mediterranean Sea and contiguous Atlantic area (Gulf of Cádiz), based on data collected through the aerial component of the ASI survey. The results are discussed considering their relevance towards informing conservation and management decisions, as well as an example of how a coordinated effort between countries is crucial to raise knowledge and ultimately to achieve and maintain good conservation status of species and habitats in a diverse and heterogeneous biogeographic and geopolitically complex region.
2 Materials and methods
2.1 Survey design
A large portion of the Mediterranean (77% of its entire area totaling 1,9 million Km²) and Gulf of Cádiz was divided in 32 strata based on the best compromise between oceanographic and physiographic characteristics, as well as the overarching political or jurisdictional constraints and logistic issues such as fuel availability, planes’ endurance, location of airports and issuing of flight permits. Equally-spaced zig-zag transects (Buckland, 2001; Strindberg and Buckland, 2004) were designed through the dedicated software Distance 7.3 (Thomas et al., 2010), to obtain an equal coverage probability within each stratum and to optimize effort. Transects were planned to be flown once with the ratio Effort/Area varying between 2.7 and 3.6% coverage, assuming an effective strip width of 0.5 km. The survey design, with transects and strata, is shown in Figure 1. Areas that could not be monitored by plane in the southern basin were instead largely covered by the ship-based component of the ASI and are presented in Boisseau et al. (2024).
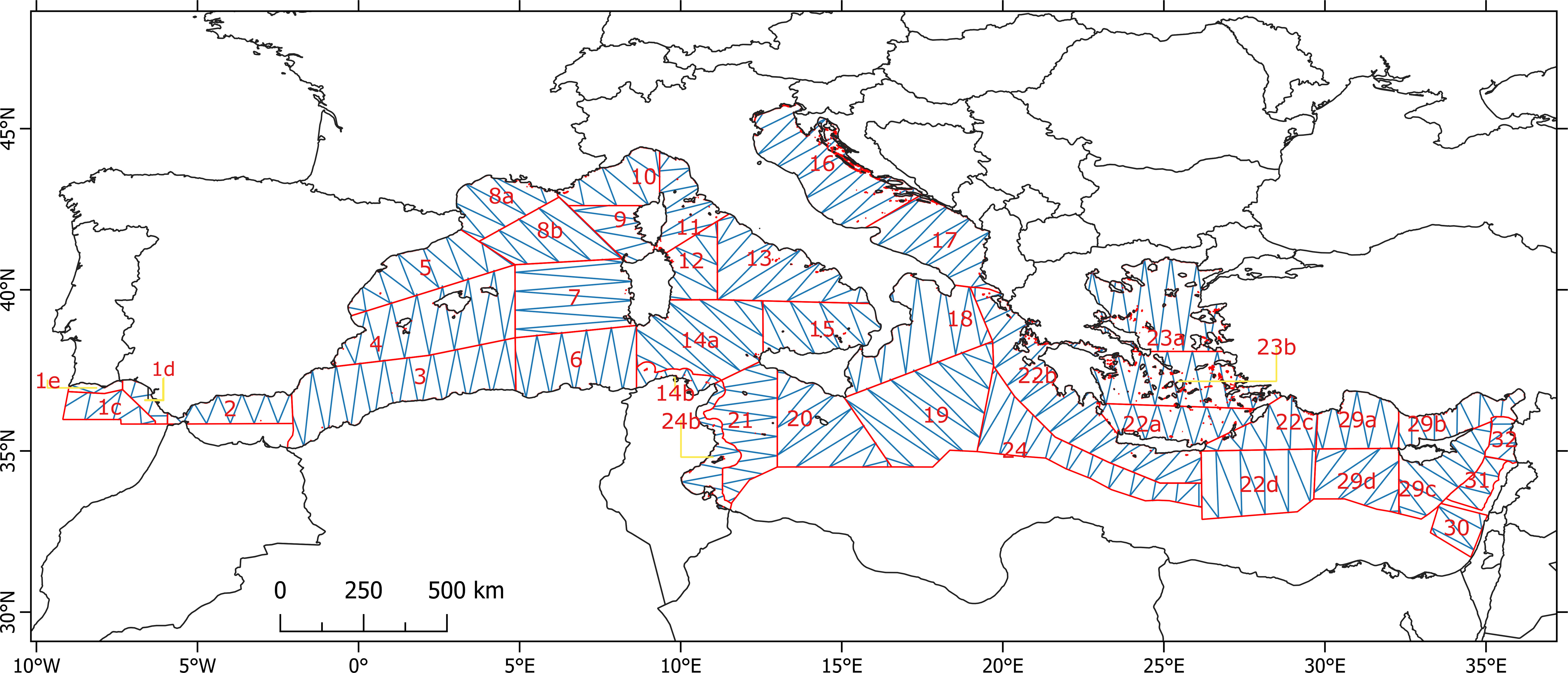
Figure 1 ASI survey design with strata (in red) and planned survey tracks (in blue).
2.2 Data collection
The survey was conducted between June and August 2018. Eight planes of three different models – 4 Partenavia (P68), 2 Britten Norman Islander (BN-2) and 2 Cessna 337 Skymaster O-2 (push-pull) – all equipped with bubble-windows to allow for direct observations on the track-line, were used for the survey. Each aircraft accommodated two primary observers, scanning the sea surface on both sides of the aircraft, and one data operator, in addition to the pilot. Surveys were conducted in passing mode (Dawson et al., 2008), i.e., the plane did not leave the track-line to approach sightings, unless it needed to estimate group size of large groups of animals or to identify species, based on ad-hoc decisions by the observers and cruise leader. The flight altitude was kept constant at 183m (600 feet), in accordance with similar surveys where target species were small cetaceans (Hammond et al., 2013) or marine megafauna (Laran et al., 2017a; Laran et al., 2017b; Pettex et al., 2017; Rogan et al., 2018), and ground speed was maintained at 100 knots (185 km/h).
The software SAMMOA 1.1.21 was used for data collection, storage, validation and management. Teams were created according to previous experience in leading and participating in aerial surveys, with researchers taking part in both theoretical and practical training sessions to prepare for field work activities, and familiarize with protocols.
Data were collected in ‘line transect mode’ by recording the declination angle when the animal or group of animals were abeam to the plane (Buckland, 2001). Sea and weather conditions (i.e., Beaufort wind scale, glare severity and angle, water turbidity, cloud coverage and presence of glint) were recorded at the beginning of each transect and at any time they changed. An overall subjective assessment of the detection conditions (i.e., “excellent”, “good”, “moderate” or “poor”, hereafter referred to as “Sightability score”) for each observer was also recorded where, as an instance, “good” conditions corresponded to an observer perceiving the likelihood of spotting a small delphinid within the searching area (within approx. 300 m from the track line) to be good (e.g., Beaufort wind scale ≤ 2, turbidity < 2 and glare moderate, good or absent). These approximate limits were identified by marks on the bubble-windows corresponding to a theoretical sighting angle of 31°, which at the survey flight altitude would equate to a perpendicular distance of 304.7 m. Survey was halted when conditions were deemed “poor” with at least Beaufort wind scale and water turbidity greater than 3 and 1, respectively. Details on the ‘environmental condition fields and codes’ used during the surveys are presented in the Supplementary Materials.
Data collected during sightings included the species (identified to the lowest possible taxonomic level), school size, number of calves, behavior, swimming direction and possible reaction to the aircraft. The declination angle to the sighted animal/group of animals, measured with a hand-held clinometer, was used to calculate the perpendicular distances to the track line. For sightings with species or school size initially uncertain, primary search effort was stopped, and a specific circling maneuver – similar to the “race-track” when the aircraft circles back to resurvey a defined segment of the transect (Hiby, 1999; Scheidat et al., 2008; Gilles et al., 2009) – was implemented only to gain insight of group size or species identification. Data were validated after each flight, all flight data were collated and merged into a single dataset and sightings were associated to corresponding effort using ArcGIS 10.6 and MGET tool (Roberts et al., 2010) for final review prior to the analysis.
2.3 Data analysis
Before performing density and abundance estimation analysis, data were used to provide explorative descriptive statistics and calculation of encounter rates (sightings per unit of effort), using a grid cell of 50x50 km, through a dedicated plugin PelaSIG. The results of this exploratory analyses and the details on the plugin are presented in Nivière et al. (2024).
2.3.1 Abundance and density estimates
Although the study area was divided into 32 strata (Figure 1), these were later merged into larger sectors referred to as “Sub-Areas” (Figure 2) to better reflect distributional ranges of species, and to prevent having sectors with little effort or too few observations to produce robust estimates. This post-stratification was done in the Distance software by specifying the Sub-Area as stratum within the model definition properties. These larger sectors were also identified to reflect existing spatial extents defined under the MSFD and EcAp framework2 and to facilitate reporting. Although the results of design-based analysis have been performed for each original block (see Supplementary Materials), this paper presents the results as obtained for the above-mentioned Sub-Areas.
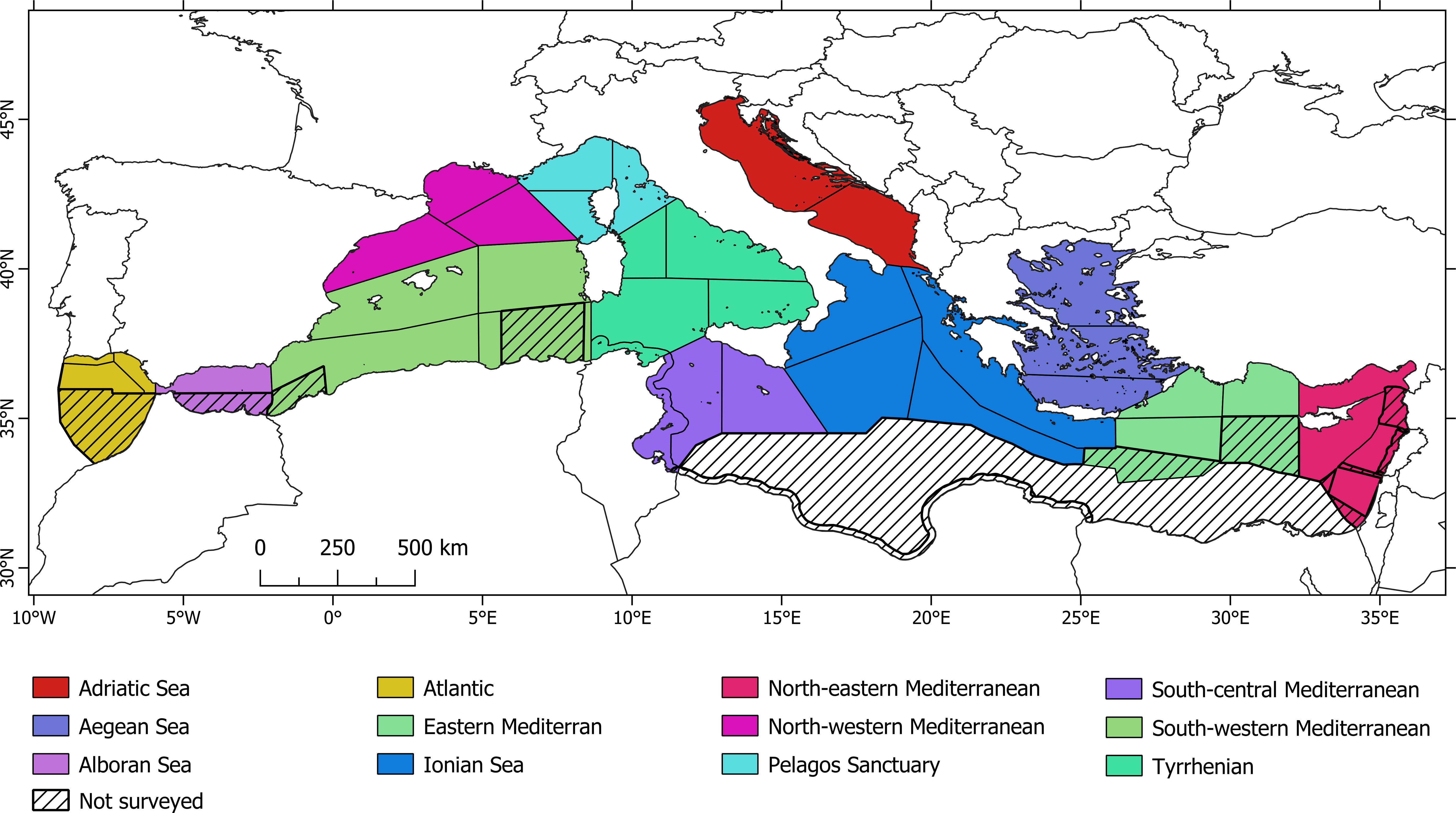
Figure 2 Designation of “Sub-areas” within the ACCOBAMS Region for abundance and density analyses.
2.3.2 Design-based analyses
Analysis, conducted in the software Distance 7.3 Release 1 (Thomas et al., 2010), followed standard multiple covariate distance sampling approach (MCDS; Buckland, 2001; Marques and Buckland, 2004), where additional explanatory variables are considered along with the perpendicular distance to the sightings to estimate the detection function (Buckland et al., 2015). Density of individuals was calculated as:
where is density, n is the number of sighted groups, is mean group size, L is the total length of transect searched, and esw is the effective half strip-width (i.e., width of the strip multiplied by the average probability of detection within that strip), which provides a simple measure of detectability. The quantity 2eswL is thus the area effectively searched. Population density is calculated as the number of individuals/groups counted divided by searched area.
From the estimated density, the abundance is calculated as the product of and the searched area A:
Detection functions were fitted to the perpendicular distance data to estimate the esw. When the number of observations for some species was too low to build a reliable detection function, species with similar characteristics (therefore potentially similar detectability) were pooled together to derive their abundance estimates. The Mark-Recapture Distance Sampling engine (MRDS) available in Distance 7.3 was used with the configuration of “single observer” for these analyses, so estimates could be derived for each species within a guild (when a guild was modelled). However, by using the “single observer” configuration, it worked practically as an MCDS (multiple covariate distance sampling).
A species-specific detection function was obtained for bottlenose, striped and Risso’s dolphins. Considering the wide variation of Delphinidae body lengths, with adults ranging between 1.5 to over 9 m (Jefferson and LeDuc, 2018), and considering the within-species geographical clines in body-length and morphological traits (e.g., van Aswegen et al., 2019), for the purpose of this paper we consider any species larger than striped or common dolphins to be a large Delphinidae. Accordingly, a pooled detection function, mainly based on body size, was calculated for the following groups:
▪ Small dolphins (including common, striped and unidentified small dolphins), to derive estimates for common dolphins.
▪ Large dolphins (including bottlenose and Risso’s dolphins and long-finned pilot whales), to derive estimates for pilot-whales.
▪ Baleen whales (including fin whales, one single minke whale and unidentified Balaenopteridae).
▪ Beaked whales.
For fin whales, estimates were corrected by a factor of 0.538 (CV=0.13) based on an average group size of 1.6 whales from the Western Mediterranean Sea (Panigada et al., 2021). Concerning sperm whales, although size is comparable to other species such as fin and minke whales, their diving-surfacing patterns strongly differ. The differences between these similarly sized species would therefore imply strong differences in the probability of detection and related availability biases. Also, for sperm whales, abundance estimated from visual aerial line-transect distance sampling surveys is likely to be negatively biased because of the long dive times, and passive acoustic approaches might produce more robust estimates (e.g., Barlow and Taylor, 2005; Lewis et al., 2018; Boisseau et al., 2023). Accordingly, sperm whales were not included in the analyses of aerial survey data and estimates are presented, instead, in Boisseau et al. (2024) based on ship-based visual and acoustic surveys.
Covariates used in the MRDS analyses were selected for their potential role in the process of cetacean detection by visual observers from the air (Table 1). The final model selection for each species or guild was done on the base of several parameters, including Delta AIC, K-S and CvM goodness of fit tests, qq-plots, and visual inspection of the shape of the detection function. QQ-plots and detection functions are provided in the Supplementary Materials as well as fit statistics for the selected models (the latter provided as Rdata file with extension *.Rdata).
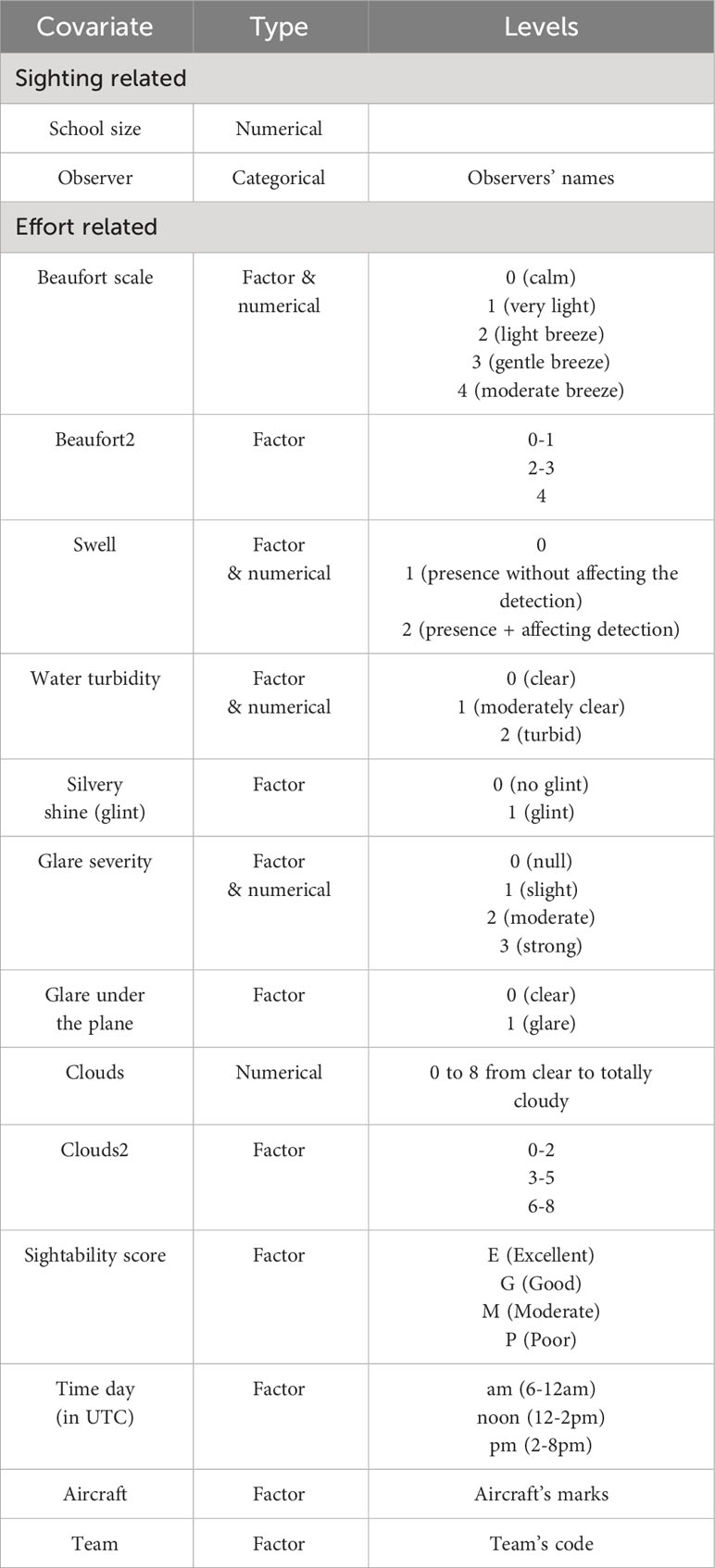
Table 1 Covariates collected during effort and tested in MRDS models and their ranges or factor levels.
3 Results
Overall, more than 55,000 km were monitored along predetermined transects, from the Gulf of Cádiz in the West to the Israeli coast in the far East, over a surface of almost 1.9 million km². Approximately 91% of the planned transects were surveyed (Table 2; Figure 3). Nine species of cetaceans, identified to the lowest taxonomical level, were encountered (Table 3): bottlenose dolphins (Tursiops truncatus), common dolphins (Delphinus delphis), Risso’s dolphins (Grampus griseus), striped dolphins (Stenella coeruleoalba), long- finned pilot whales (Globicephala melas), Cuvier’s beaked whales (Ziphius cavirostris), sperm whales (Physeter macrocephalus), fin whales and minke whales (Balaenoptera acutorostrata), for a total of 577 sightings and 8,348 individuals. For 234 sightings (3,460 individuals) it was not possible to identify the species.
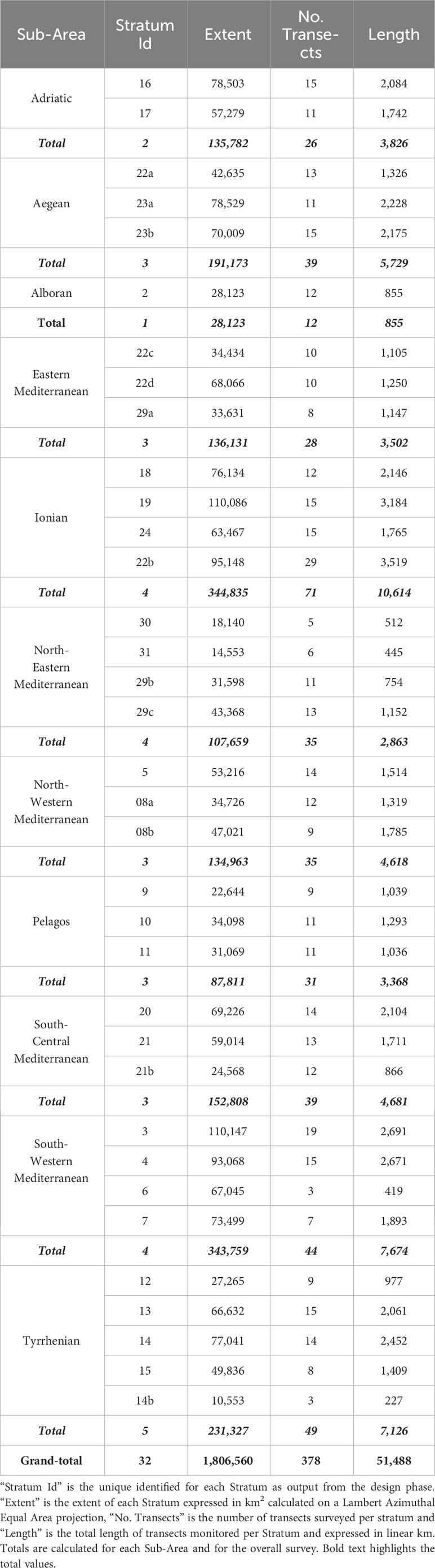
Table 2 Summary of survey effort by Sub-Area.
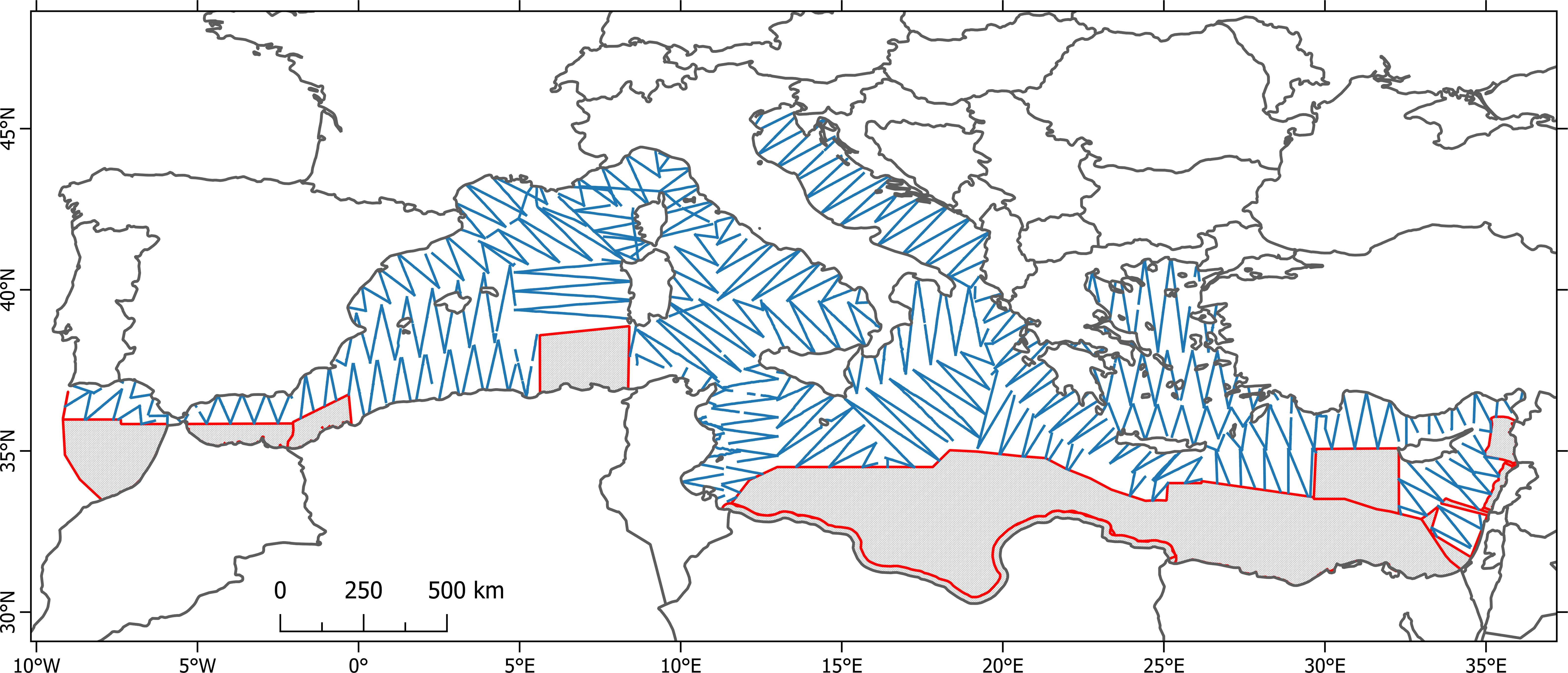
Figure 3 Map with the total realized effort within the ACCOBAMS Region.
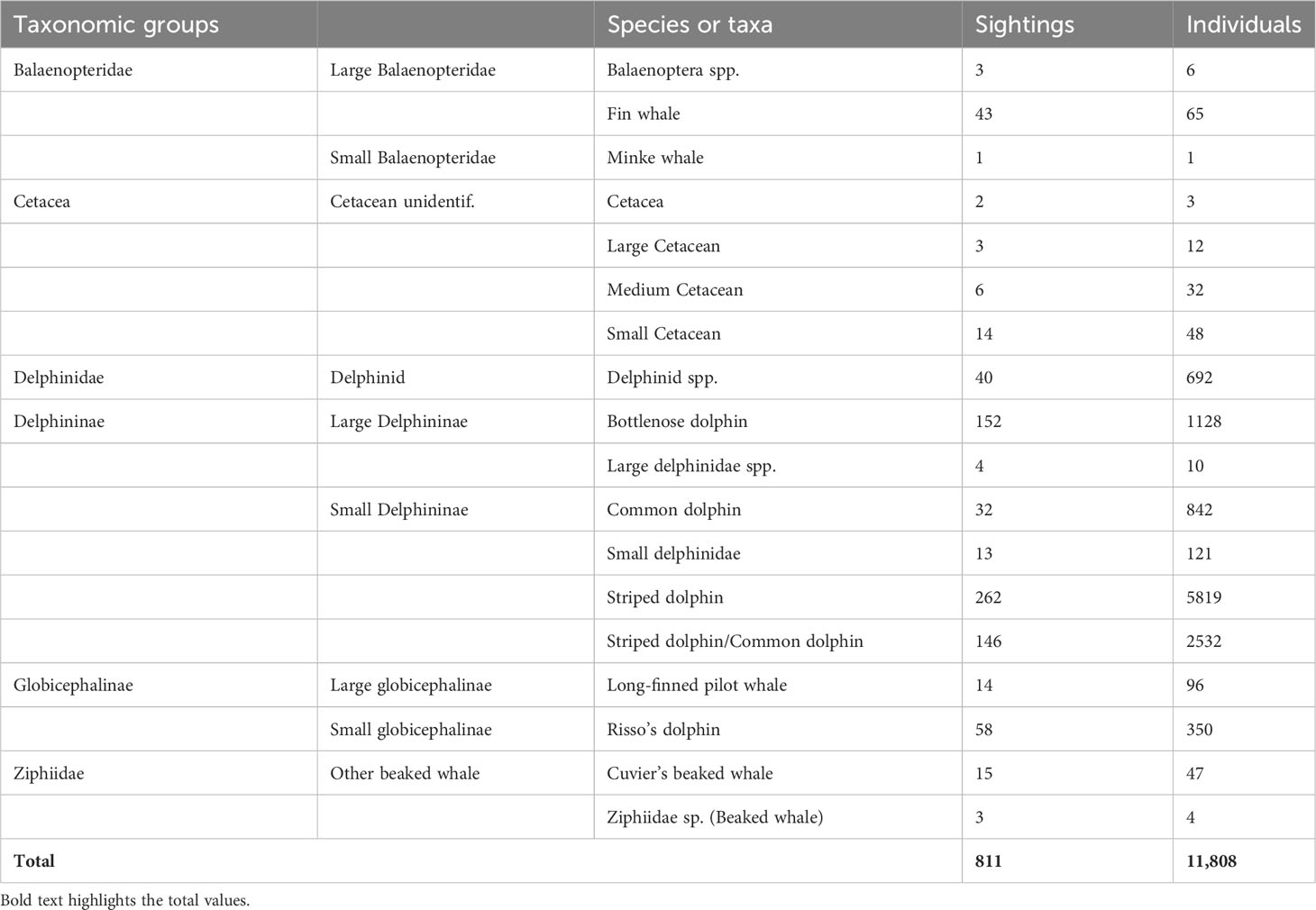
Table 3 Sightings of cetaceans encountered while ‘on effort’ during the aerial survey.
3.1 Search effort and sightings
Table 3 presents a summary of sightings and related information. Striped dolphins were the most observed species in terms of both the number of recorded sightings and the number of observed individuals, followed by bottlenose dolphins. In 18% of the sightings, it was not possible to discriminate between striped and common dolphins.
3.2 Design-based abundance estimates
Uncorrected density and abundance were estimated for all the species with a sufficient number of observations and for 4 guilds, alongside their Coefficients of Variation (CV) and 95% Confidence Intervals (95% CI) (Table 4). The full results of the design-based analyses are presented in the Supplementary Materials, alongside information, for each species/group of species and Sub-areas, on mean group, expected group size, the encounter rate of groups, as well as the parameters and results of the final detection functions and q-q plots.
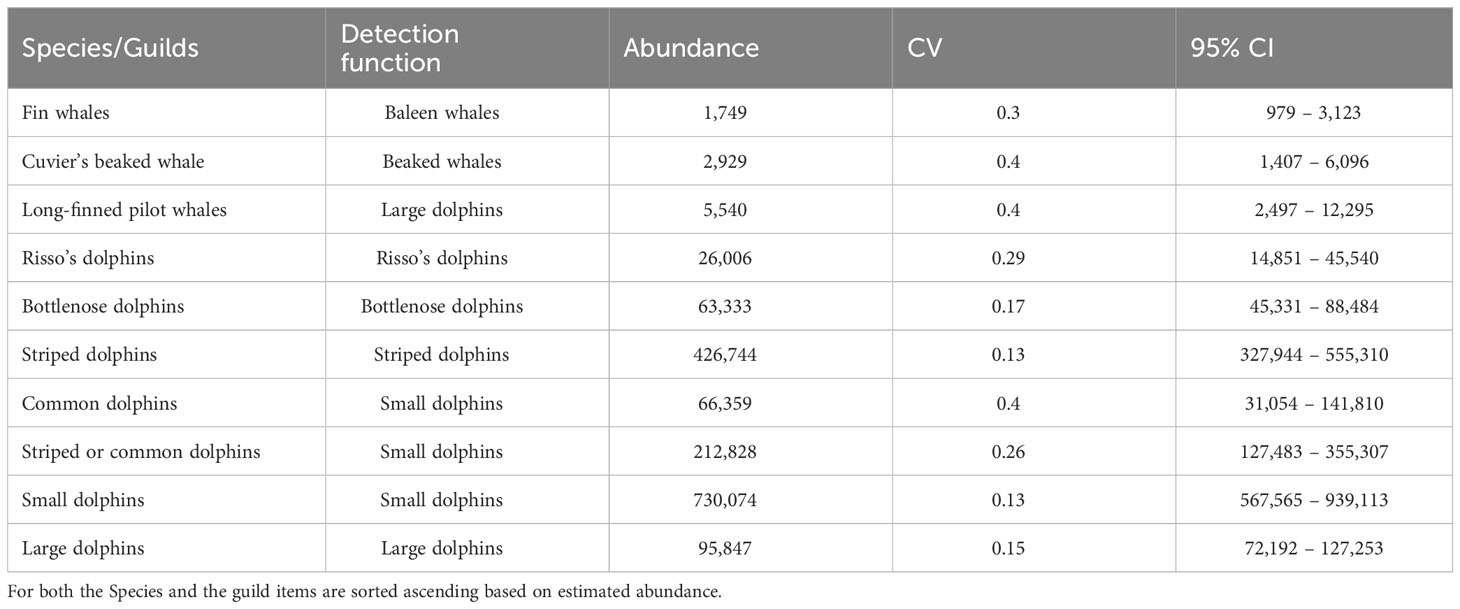
Table 4 Summary of design-based abundance estimates of cetaceans for sub-areas with associated Coefficients of variation (CV) and Confidence intervals at 95% (95% CI).
Truncation distances and goodness-of-fit test results for cetaceans are summarized in Table 5. Only for two species right truncation was not necessary, and the maximum detected distance was used instead.
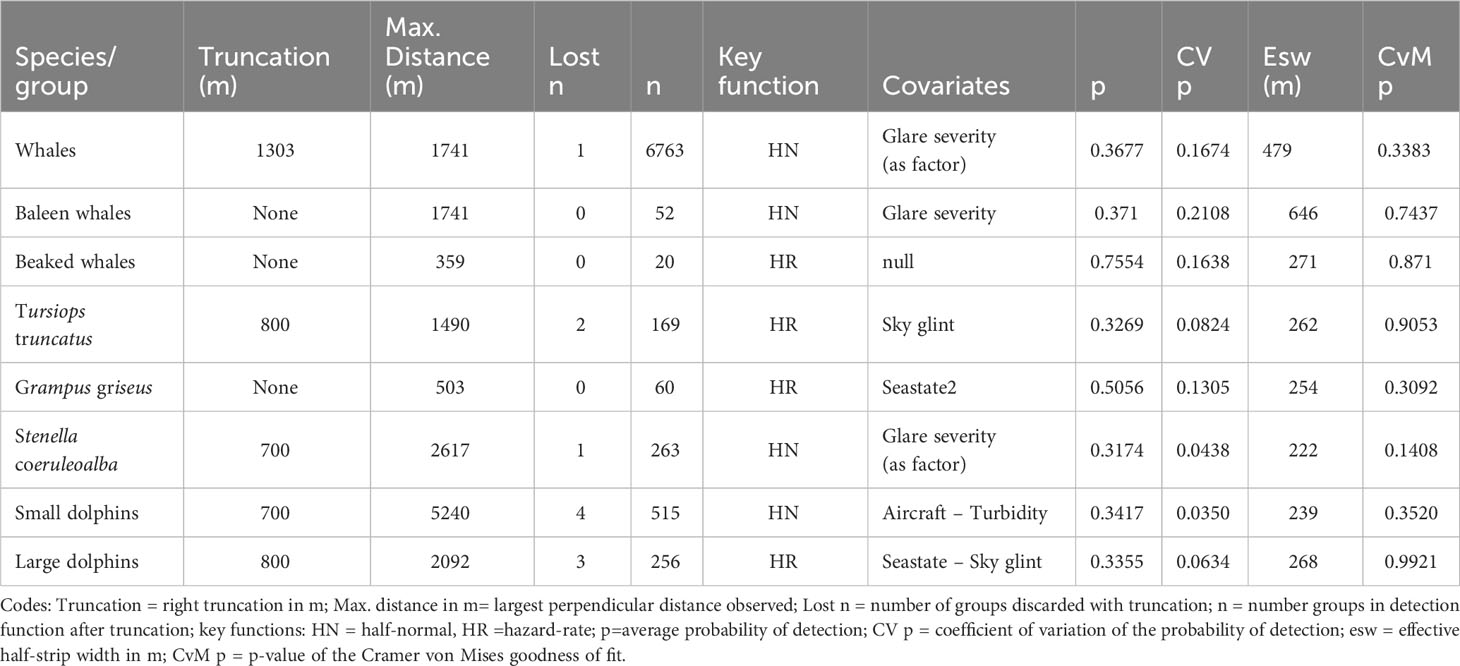
Table 5 Parameters and results of the detection functions for cetaceans.
4 Discussion
4.1 Strengths and weaknesses
The 2018 ASI has provided a first overall picture of the abundance of cetaceans throughout a large portion of the Mediterranean region and the Gulf of Cádiz, providing robust estimates for several species, as well as important information on their occurrence. These represent a baseline for further regional systematic monitoring programmes, coordinated and comparable amongst all areas. They greatly improve current knowledge on cetacean status and facilitate the development of targeted conservation and mitigation measures, as well as facilitate international obligations (EU, UNEP-MAP). Moreover, together with the spatial approach developed by Cañadas et al. (2023), they can contribute towards both place- and threat-based conservation efforts in the Agreement area, with the identification of Important Marine Mammal Areas (IMMA; Hoyt and Notarbartolo di Sciara, 2021; Tetley et al., 2022) and Cetaceans Critical Habitats (CCH), as well as areas of major overlap between cetaceans and human activities in the summer.
It is important to note that this effort represents the first synoptic basin-wide estimates for cetaceans in the Mediterranean Sea and contiguous Atlantic area, thus making quantitative comparisons with existing knowledge more difficult. Most of the past effort in the region has been allocated along coastal areas and – over three decades of research on cetaceans – several portions of the Mediterranean have never or only minimally been surveyed in the past (e.g., Mannocci et al., 2018). The easternmost and southernmost Mediterranean basin remain currently unsurveyed by systematic effort either by plane or vessel. By contrast, coverage of the western Mediterranean Sea has been more substantial during the last decade, with aerial surveys taking place in its central and north-western sectors (e.g., Gómez de Segura et al., 2006; Fortuna et al., 2014; Lauriano et al., 2014; Notarbartolo di Sciara et al., 2015; Laran et al., 2017b; Lauriano et al., 2017; Panigada et al., 2017; Fortuna et al., 2018), as well as wide scale vessel based monitoring (e.g., Forcada et al., 1994; Forcada et al., 1996). Work is ongoing to enable more direct quantitative comparisons between these and the ASI results.
We do acknowledge the need for a comprehensive integration of the design- and model- based estimates (and comparisons with relevant estimates at a more ‘local’ scale, where possible) and the spatial modelling distributional information. This aspect, as well as the issue of stock structure within the region, will be the subject of future dedicated effort. Model-based results and distribution across the Mediterranean Sea are presented and discussed in Cañadas et al. (2023).
With the exception of fin whales, the abundance estimates provided here are underestimates of the true numbers, in that they have not been corrected for availability nor perception biases. Although the perception bias was not estimated, it was assumed to be constant throughout the whole survey given that same protocol and type of planes were used, and survey teams (created during the training phase) remained unchanged throughout the entire survey. Perception and availability biases are not mutually exclusive and can negatively affect the estimates, unless accounted for (e.g., Pollock et al., 2006; Fuentes et al., 2015). Perception bias varies with the target species and can be minimized by using experienced observers and accounted for in a double-platform framework (Buckland et al., 2010; Burt et al., 2014), while availability bias is linked to the activity periods and habitat types of the species of interest (Smolensky and Fitzgerald, 2011) and the integration of tagging data (i.e., data on dive–surfacing patterns) with aerial surveys datasets can help accounting for availability bias in the estimation of abundance (Nykänen et al., 2018). Correcting for such biases, whilst important to obtain estimates of absolute abundance, is not essential for trend analyses, for which the estimates can be treated as indices of abundance, provided that the levels of bias remain constant over time.
For fin whales, estimates have been corrected by a factor 0.538 (CV=0.13), assuming that perception bias for a trained and expert observer to be 1. This assumption is based on the fact that, in general, perception bias for large whales is thought to be relatively small. Hansen et al. (2019) estimated perception bias to range from about 0.86 – 0.99 for fin whales from aerial surveys off Greenland. The same authors provided an availability bias ranging between 0.2 – 0.3. Heide-Jørgensen et al. (2010) estimated a factor of around 0.86 for fin whales from an aerial survey off West Greenland but they did not provide an estimate for availability bias. Here, we used an average group size (g) of 1.6 whales from the Western Mediterranean Sea (Panigada et al., 2021), to obtain a correction factor of 0.538 (CV=0.13).
Generally, cetaceans found in the Mediterranean Sea can be easily distinguished at the species level. However, during aerial surveys the flight altitude and speed, as well as the sea and weather conditions, amongst other factors, can hamper discriminating species when small delphinids are observed. This is exacerbated by the occurrence of sympatric species often in mixed associations. These mixed-species groups mostly include striped and common dolphins (Frantzis and Herzing, 2002; Frantzis et al., 2003; Bearzi et al., 2011b), and to a lesser extent striped, common and Risso’s dolphins (Frantzis and Herzing, 2002). Furthermore, animals with intermediate striped-common dolphin pigmentation have been reported (Frantzis and Herzing, 2002; Bearzi et al., 2011b), providing evidence of introgressive hybridization between these two species (Antoniou et al., 2018). In future campaigns, image acquisition by digital cameras could be used to improve small cetaceans’ species identification (e.g., Žydelis et al., 2019; Garcia-Garin et al., 2020; Raudino et al., 2022; Bigal et al., 2022).
In a line transect distance sampling framework, the probability of detecting an object is a function of many factors other than distance of the object from the track-line, including group-size, species and habitats, as well as sea state (e.g., Holt and Cologne, 1987; Thomas et al., 2012; Fandel et al., 2020). Numerous studies have demonstrated that detection varies among species, over time, and among habitats, and there may be serious consequences when this variability is ignored. Failure to correct for imperfect detection (by observers and/or because of changes in the animal behavior over time or with habitat), as an instance, may result in spurious estimated relationships with ecological covariates and inaccurate abundance estimates that could mask trends and improper selection of indicator species (Kellner and Swihart, 2014). In this context, during the ASI survey, when it was not possible to discriminate between striped and common dolphins, sightings were attributed to a “small dolphin” or “common/striped dolphin” guild. Furthermore, it is important to note that the ASI dataset can be re-analyzed a-posteriori by applying a correction factor related to the observed proportion of one species vs the other (Cañadas, personal communication). While this approach could not be implemented at this stage, the re-classification of ambiguous species detections will be applied during the future replicates of the survey.
4.2 Species accounts
4.2.1 Striped dolphin, common dolphin and striped/common dolphin guild
At the species level, the ASI confirms that the striped dolphin is the most abundant species in the Mediterranean Sea (n= 426,744; 95% CI= 327,944 – 555,310) in agreement with previous research (e.g., Forcada et al., 1994; Forcada et al., 1995; Cotté et al., 2010; Laran et al., 2017b; Panigada et al., 2017). Recently, Laran et al. (2017b) obtained an uncorrected density of striped dolphins for a large portion of the north-western Mediterranean, including the Pelagos Sanctuary (corresponding to ASI strata 8a, 8b, 9, 10 and 11; see Figure 1), for the summer months of 2012, of 0.484 animals/km2. Gómez de Segura et al. (2006), for the waters off eastern central Spain, corresponding to the westernmost portions of ASI strata 3-5, estimated an overall summer density of 0.522 striped dolphins/km2, while the ASI average estimate for these strata is 1 animal/km2 (range= 0.53 – 1.44) individuals/km2. Similarly, Panigada et al. (2017) estimated an uncorrected density of 0.41 animals/km2 from an aerial summer survey conducted in 2010 over an area roughly corresponding to ASI strata 9, 10, 11, 12, 13 and the eastern sectors of strata 8b and 7 (Figure 1). For the Adriatic Sea (ASI area 17 and part of 16) Fortuna et al. (2011) provided an estimate of uncorrected density of 0.208 animals/km2 (N= 15,343, CV=29.8%; 95% CI= 8,545-27,550) from an aerial survey conducted in summer of 2010.
The uncorrected density estimates obtained during the ASI are not directly comparable to these earlier surveys due to the different extent of the surveyed areas, but are within the same range as their results with an average density for those strata of 0.269 animals/km2 (range= 0.0127 – 0.5219).
The Mediterranean sub-population of common dolphin has undergone a drastic reduction in the past decades as a consequence of ever-increasing human pressures on the species range of distribution (Bearzi et al., 2021). Habitat loss and fragmentation, alongside the indirect effects of overfishing, unintentional captures during fishing operations, and take, have strongly contributed to the decline of dolphin numbers across the entire region (e.g., Bearzi et al., 2003; Bearzi et al., 2004; Bearzi et al., 2008; Cañadas and Vázquez, 2017; Mussi et al., 2019). Overall, 32 primary sightings of common dolphins, mostly in the western portion of the basin and in the Strait of Sicily (see Nivière et al., 2024), were recorded during the ASI aerial surveys, with a total estimate of 66,359 individuals (95% CI=31,054 – 141,801).
During the ASI a specific sighting guild of unidentified small dolphins was used when a clear distinction between striped and common dolphins was not possible. A total of 148 sightings were assigned to this category, leading to an overall estimated abundance of 212,828 animals (95% CI=127,483 – 355,307).
4.2.2 Bottlenose dolphin
Bottlenose dolphins were the second most abundant species (n=63,333; 95% CI=45,331 – 88,484) observed during the ASI. The distribution showed ‘patches’ of relatively high density (Cañadas et al., 2023). Lauriano et al. (2014) obtained an overall uncorrected density of 0.006 animals/km2 over a large portion of the central and north-western Mediterranean corresponding to ASI strata 9-13, 14a, 15 and portions of strata 7, 8a and 8b (see Figure 1). The average density of bottlenose dolphins over these strata, obtained during the ASI, was 0.032 animals/km2 (range= 0 – 0.143). Gómez de Segura et al. (2006), for the waters off eastern central Spain, corresponding to the westernmost portions of ASI strata 3-5 (see Figure 1), obtained an uncorrected density estimate for the summer of 0.041 animals/km2 while the average ASI estimated density for these strata is 0.023 (range= 0.003 – 0.063). Fortuna et al. (2018) obtained uncorrected density for the whole Adriatic (2010 and 2013 aerial surveys pooled dataset) of 0.042 animals/km2 corresponding to the strata 16 and 17, with the average ASI estimated density for these strata of 0.074 (range= 0.05 – 0.1). Fortuna et al. (2018) showed there was a notable difference between regions of the three Adriatic sub-basins; North 0.052, Central 0.034 (ASI Strata 16) and South 0.032 (ASI Strata 17). Finally, Laran et al., (2017b) report summer uncorrected density of 0.016 (CV=53%) for an area corresponding to ASI Strata 8a, 8b, 9, 10 and 11. Although also in this case the results are not directly comparable due to differences in the extent of the monitored areas, they agree with previous surveys and show how this species tend to generally occur at low density with patchy areas of higher abundance.
4.2.3 Fin whale
The fin whale was the only species for which a corrected abundance estimate was derived. At 3,282 individuals (CV=0.31), the ASI estimate is comparable to previous estimates of absolute abundance based on shipboard surveys of 3,583 individuals (95% CI=2,130 – 6,027; Forcada et al., 1996). Past abundance and density estimates arising from summer aerial surveys conducted in the central and north-western Mediterranean Sea show that the density of fin whales is higher in the north-western Basin and that abundance decreases at lower latitudes and higher longitudes (Panigada et al., 2017). Laran et al. (2017b), Panigada et al. (2011), Panigada et al. (2017), Bauer et al. (2015) show higher densities of fin whales in the north-western portion of the Mediterranean and this pattern is confirmed by the ASI, as well as by different approaches based on long-term photo-identification and microsatellite genotyping (Zanardelli et al., 2022; Tardy et al., 2023). The north-western Mediterranean is a well-known fin whale summer feeding ground where about 1,000 fin whales aggregate (e.g., Notarbartolo di Sciara et al., 2016; Panigada et al., 2006). The ASI confirms fin whales summer distribution and habitat use and highlights the necessity of seasonal large scale monitoring programmes aimed at understanding spatio-temporal patterns of fin whales occurrence and distribution at the Basin scale to inform conservation and management decisions. In this respect, the ASI has highlighted no appreciable trends in the size of the sub-population of Mediterranean fin whales. It is however necessary to stress that the basin is visited by North-Eastern North Atlantic fin whales (Geijer et al., 2016) and therefore the obtained estimates for true Mediterranean whales might be biased upwards. According to the IUCN, total abundance of fin whales should be greater than 5,200 individuals in order for the sub-population to exceed the limit of 2,500 mature individuals (48% of the total population being mature; Taylor et al., 2007). This, in conjunction with the fact that the sub-population experiences an inferred continuing decline in number of mature individuals, and that all mature individuals are in one subpopulation, further stresses the need to identify and implement targeted measures that would ensure the persistence of the species in the basin (Panigada et al., 2021).
4.2.4 Risso’s dolphin
The Risso’s dolphin of the Mediterranean Sea is one of the least-known cetacean species in the region and it has been the subject of few dedicated studies (Bearzi et al., 2011a). With a total of 58 sightings, abundance for this species has been estimated at 26,006 animals (95% CI=14,851 – 45,540), providing the first abundance estimates for this species at the basin level, in particular for the north-western African coast. Laran et al. (2017b) provide corrected density estimates of 0.006 (CV=47%) for an area corresponding to ASI strata 8a, 8b, 9, 10 and 11. After aerial survey in the Adriatic in 2010, Fortuna et al. (2011) obtained uncorrected density estimate of 0.007 (CV=78.1%) with uncorrected population estimate of 510 individuals (CV=78.1%; 95% CI=124-2,089) present in the Central and South Adriatic (corresponding to ASI strata 17 and southern part of 16). Gómez de Segura et al. (2006), for the waters off eastern central Spain, corresponding to the westernmost portions of ASI strata 3-5 (Figure 1), obtained an uncorrected density estimate of 0.015 animals/km2 while the average ASI estimated density for these strata is 0.044 (range= 0.037 – 0.052). Azzellino et al. (2016) using photographic mark–recapture methods provided a population size estimate for the western Ligurian Sea (corresponding to the north-western ASI stratum 10; Figure 1), for the period from 1998 to 2012, of about 100 individuals (95% CI= 60–220) and detected a decline in population size from an average of about 120–150 from 2000 to 2005, to an average of 70–100 during 2010 to 2014. Furthermore, the same authors highlight how the species occurrence has shifted from coastal and continental slope waters to more pelagic areas. No sightings of Risso’s dolphins were recorded in this stratum during the ASI aerial survey; abundance was estimated for the adjacent southern stratum 9 (Figure 1) at about 200 animals (95% CI= 44 – 894), confirming the possible distributional shift observed by Azzellino et al. (2016). Azzellino et al. (2016) suggest that environmental variability, depletion of resources by fisheries and possibly interspecies competition could have contributed to the changes in Risso’s dolphin habitat use and occurrence in the western Ligurian Sea. Recent studies show that shifts in cetacean distribution can be attributed to changing environmental conditions (Becker et al., 2022) or arising from the impacts of climate changes (van Weelden et al., 2021). However, the correlates to the observed shift in the occurrence of Risso’s dolphins at the Mediterranean level have not been investigated yet.
4.2.5 Pilot and Cuvier’s beaked whales
Cuvier’s beaked whales are known for their elusive behavior, characterized by long diving times and relatively short surfacing periods (e.g., Quick et al., 2020), making them particularly affected by availability bias during aerial surveys (Thomson et al., 2012). To a lesser extent, this also applies to deep-diving pilot whales (Heide-Jørgensen et al., 2002). Overall, sightings of these two species accounted for less than the 2.5% of all cetacean sightings recorded during the ASI, with pilot whales being observed 14 times and beaked whales 15 times. Long-finned pilot whales’ overall abundance was estimated at 5,540 individuals (95% CI= 2,497-12,295). Cuvier’s beaked whales’ abundance was estimated at 2,929 animals (95% CI=1,407 – 6,096), confirming existing knowledge on the basin-wide presence of the species and, at the same time, confirming how Cuvier’s beaked whales typically occur at low densities in relatively small patches. A previous study by Cañadas et al. (2018) estimated the abundance of Cuvier’s beaked whales at 5,600 (CV=0.24, 95% CI= 4807 – 7254). While the ASI aerial component provides useful information on the occurrence, distribution and abundance of deep-diving species in the Mediterranean basin, the results indicate that more targeted studies using different methods may provide more robust estimate abundance of deep diving species.
4.3 Toward a strategy for monitoring cetacean populations in the Mediterranean Sea
Cetacean monitoring programmes aimed at evaluating populations and their distribution in the ACCOBAMS area are implemented within the framework of several initiatives at national and multilateral levels. Member countries of the European Union need to regularly undertake cetacean monitoring programs to collect the information required for their reporting under Article 17 of the Habitats Directive (HD), as well as in relation to the assessment of the Environmental Status under Descriptor 1 (Biodiversity) of the EU Marine Strategy Framework Directive (MSFD).
In the Mediterranean Sea, other cetacean monitoring programmes are expected within the framework of the Integrated Monitoring and Assessment Programme (IMAP) of the Ecosystem Approach process under the framework of the Barcelona Convention. A similar initiative is also expected for the Black Sea within the framework of the Bucharest Convention.
ACCOBAMS is now working to implement a Long-Term Monitoring Programme (LTMP; ACCOBAMS Resolution 8.10), aimed at collecting reliable data to obtain robust estimates of abundance and distribution of cetacean species in the ACCOBAMS area. This effort should also involve the collection of information on human activities at sea (both directly as part of survey protocols, and from relevant organizations collecting such data and indirectly using remote methods) which represent a known or emerging threat to cetaceans (e.g., maritime traffic, leisure boating, fishing, marine litter, chemical pollutants), to facilitate a spatial risk assessment based on pressure data and cetaceans distribution. This information will feed into the ongoing ACCOBAMS process to identify high-risk areas of interaction between cetaceans and human activities, where dedicated conservation measures should be identified and implemented. The ultimate objective is to provide a robust scientific basis for the setting and the regular adjustment of management measures that allow achieving a favorable conservation status for cetaceans in the Agreement area. Through such an achievement, the Parties will fulfil their obligations under ACCOBAMS, as well as their commitments towards other relevant Multilateral Agreements.
The periodic implementation of the ACCOBAMS LTM will allow trend assessments in species population size and distribution and to assess and evaluate conservation measures in place. While we acknowledge that there are high margins of error when detecting trends of small populations or when sample size is small (e.g., Wilson et al., 2011; Porszt et al., 2012), we recognize that the data collected by the ASI in the Mediterranean and Black Seas (see Paiu et al., under review) have established a reliable baseline from which to measure future changes in the monitored parameters.
ACCOBAMS has agreed that conducting synoptic surveys of the whole ACCOBAMS area on a six-year frequency would provide the right balance between monitoring costs and regular updating of information on abundance and distribution of cetacean species and would be in line with the general recommendations from the EU and the Barcelona Convention, as well as with previous large-scale efforts elsewhere in the world (e.g., Hammond et al., 2021).
The main objective of ACCOBAMS is to achieve and maintain a favorable conservation status for cetaceans in the Black Sea, Mediterranean Sea and Contiguous Atlantic Area, and to obtain this, the Agreement requests the Contracting Parties to undertake, individually and through collaborations, a series of conservation and management measures and to develop cooperation for research and monitoring to fully implement them. The development of monitoring effort, such as the one represented by the ACCOBAMS Survey Initiative, is clearly designated by the Agreement among the most recommended ways to enhance the knowledge about the biology, ecology, and population dynamics of cetaceans, in order to identify and implement appropriate conservation measures.
4.4 Final remarks
The ACCOBAMS Survey Initiative provided the first synoptic estimate of abundance of eight cetacean species in the Mediterranean Sea, by implementing an unprecedented collaborative aerial survey in the whole region.
Stakeholders, including Governments, scientists, managers, and NGOs, have benefited from the data arising from the ASI survey to address local, national, and regional conservation objectives, including the negotiation of the adoption of the north-western Mediterranean PSSA (Particularly Sensitive Sea Area) by the IMO, conducting Environmental Impact Assessments, and developing Maritime Spatial Planning plans.
One of the ASI concrete outcomes is the recent re-assessment of the conservation status (sensu International Union for the Conservation of Nature – IUCN) of several of the Mediterranean sub-population of cetaceans. With no sub-populations listed as Data Deficient nor Extinct, the 77% of the resident Mediterranean subpopulations of cetaceans are considered to be threatened with extinction (IUCN, 2023), specifically listed as Vulnerable or Endangered (ACCOBAMS, 2021a). Ten of the 13 endemic sub-populations of cetaceans that are resident in the Mediterranean Sea are assessed as threatened, three as Critically Endangered, six as Endangered and one as Vulnerable. Two sub-populations previously classified as Data Deficient are now assessed as threatened, the Risso’s dolphin and the long-finned pilot whale. Eight of the subpopulations have a declining trend, four have unknown trends, and only striped and bottlenose dolphins have an increasing population trend (IUCN, 2023). This regional assessment sheds light on the conservation status of these sub-populations and highlights the need for more efforts to prevent the likely declining of the cetacean fauna of the Mediterranean Sea.
In light of the ongoing decline in global biodiversity, there is widespread recognition of the need for monitoring programs that can assess changes in species distributions and abundances over large spatial and temporal scales, in order to predict long-term biological responses to human pressures and global changes. Effective monitoring also supports the implementation of national and regional policies by informing a more comprehensive understanding of populations and environmental status. This, in turn, facilitates long-term conservation targets by addressing the challenges posed by global changes in the context of socioeconomic development. Quantifying the distribution and abundance of species, and changes in these parameters, is inherently complex, particularly when dealing with highly mobile and cryptic species such as cetaceans, whose ranges often span across large areas and are affected by the exposure to anthropogenic activities.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://accobams.org/asi-data-access-request/.
Ethics statement
Ethical review and approval was not required for the study on animals in accordance with the local legislation and institutional requirements.
Author contributions
SP: Conceptualization, Investigation, Methodology, Project administration, Supervision, Writing – original draft, Writing – review & editing. NP: Data curation, Investigation, Methodology, Writing – original draft, Writing – review & editing. HA: Investigation, Writing – review & editing. LD: Investigation, Writing – review & editing. ND-M: Investigation, Writing – review & editing. GD: Investigation, Methodology, Writing – review & editing, Software. JG: Investigation, Writing – review & editing. DH: Investigation, Writing – review & editing. SL: Data curation, Formal analysis, Methodology, Software, Writing – review & editing. GL: Investigation, Writing – review & editing. R-MP: Investigation, Writing – review & editing. MP: Investigation, Writing – review & editing. DP: Investigation, Writing – review & editing. VR: Conceptualization, Methodology, Resources, Supervision, Writing – review & editing. JV: Investigation, Writing – review & editing. AC: Conceptualization, Data curation, Formal analysis, Methodology, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The ACCOBAMS Survey Initiative was supported by all riparian countries and has received substantial financial support from the MAVA Foundation, the Prince Albert II of Monaco Foundation, the International Fund for Animal Welfare, the Ministry of Agriculture and Fisheries, Food and Environment of Spain, the French Agency for Biodiversity, the Italian Ministry of the Environment and Protection of Land and Sea, the Government of the Principality of Monaco and from the operational program for the implementation of the European Maritime and Fisheries Fund in the Republic of Slovenia.
Acknowledgments
Special thanks to the members of the ASI Contact Group, the ASI Steering Committee, observers and team leaders, the ACCOBAMS Secretariat and the ACCOBAMS Scientific Committee for their continuous support to the development of the ACCOBAMS Survey Initiative project since its inception. Parts of this manuscript has been released as a final report at https://accobams.org/wp-content/uploads/2021/11/ASI-Med-Report-updated.pdf, (ACCOBAMS, 2021b). Thanks to the government organizations that provided logistical support in the field and/or the necessary authorizations, and to the aircraft companies and their pilots, without whom the ACCOBAMS Survey Initiative would not have been possible. The authors are grateful to the Italian Ministry for the Environment and Energy Security for the voluntary contribution offered to ACCOBAMS in support of the publication of this article. Special thanks also to PELAGIS Observatory (UMS 3462, CNRS-La Rochelle University) for training the observers. Many thanks to Greg Donovan who provided helpful comments on an earlier version of the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2023.1270513/full#supplementary-material
https://github.com/NinoPierantonio/Panigada_etal_ASI_SI_Frontiersin_2023.
Footnotes
- ^ SAMMOA 1.1.2. Système d'Acquisition des données sur la Mégafaune Marine par Observations Aériennes, Software developed by UMS 3462 Pelagis LRUniv-CNRS and Code Lutin (2012-2019).
- ^ https://ec.europa.eu/environment/marine/images/MSFD_regions.jpg
{kind=link}
References
ACCOBAMS. (2021a). Conserving Whales, Dolphins and Porpoises in the Mediterranean Sea, Black Sea and adjacent areas: an ACCOBAMS status report. Eds. Notarbartolo di Sciara G., Tonay Monaco A. M. (ACCOBAMS). Available at: https://accobams.org/wp-content/uploads/2022/03/ACCOBAMS_ConservingWDP_web_2022.pdf.
ACCOBAMS. (2021b). Estimates of abundance and distribution of cetaceans, marine mega-fauna and marine litter in the Mediterranean Sea from 2018-2019 surveys. Eds. Panigada S., Boisseau O., Cañadas A., Lambert C., Laran S., McLanaghan R., Moscrop A. (Monaco: ACCOBAMS - ACCOBAMS Survey Initiative Project). 177 pp.
Antoniou A., Frantzis A., Alexiadou P., Paschou N., Poulakakis N. (2018). Evidence of introgressive hybridization between Stenella coeruleoalba and Delphinus delphis in the Greek Seas. Mol. Phylogenet. Evol. 129, 325–337. doi: 10.1016/j.ympev.2018.09.007
Authier M., Commanducci F. D., Genov T., Holcer D., Ridoux V., Salivas M., et al. (2017). Cetacean conservation in the Mediterranean and Black Seas: Fostering transboundary collaboration through the European Marine Strategy Framework Directive. Mar. Policy 82, 98–103. doi: 10.1016/j.marpol.2017.05.012
Azzellino A., Airoldi S., Gaspari S., Lanfredi C., Moulins A., Podestà M., et al. (2016). Risso’s dolphin, grampus griseus, in the Western Ligurian Sea. Adv. Mar. Biol. (Elsevier) 75, 205–232. doi: 10.1016/bs.amb.2016.08.003
Balmford A., Moore J., Brooks T., Burgess N., Hansen L. A., Williams P., et al. (2005). The convention on biological diversity’s 2010 target. Sci. 307, 212–213. doi: 10.1126/science.1106281
Barlow J., Taylor B. L. (2005). Estimates of sperm whale abundance in the northeastern temperate pacific from a combined acoustic and visual survey. Mar. Mammal Sci. 21, 429–445. doi: 10.1111/j.1748-7692.2005.tb01242.x
Bauer R. K., Fromentin J.-M., Demarcq H., Brisset B., Bonhommeau S. (2015). Co-occurrence and habitat use of fin whales, striped dolphins and atlantic bluefin tuna in the Northwestern Mediterranean Sea. PloS One 10, e0139218. doi: 10.1371/journal.pone.0139218
Bearzi G., Agazzi S., Gonzalvo J., Costa M., Bonizzoni S., Politi E., et al. (2008). Overfishing and the disappearance of short-beaked common dolphins from western Greece. Endang. Species. Res. 5, 1–12. doi: 10.3354/esr00103
Bearzi G., Bonizzoni S., Agazzi S., Gonzalvo J., Currey R. J. C. (2011b). Striped dolphins and short-beaked common dolphins in the Gulf of Corinth, Greece: Abundance estimates from dorsal fin photographs. Mar. Mammal Sci. 27, E165–E184. doi: 10.1111/j.1748-7692.2010.00448.x
Bearzi G., Genov T., Natoli A., Gonzalvo J., Pierce G. J. (2021). Delphinus delphis (Inner Mediterranean subpopulation) (The IUCN Red List of Threatened Species). Available at: https://dx.doi.org/10.2305/IUCN.UK.2021-3.RLTS.T189865869A189865884.en (Accessed 29 July 2023).
Bearzi G., Holcer D., Notarbartolo di Sciara G. N. (2004). The role of historical dolphin takes and habitat degradation in shaping the present status of northern Adriatic cetaceans. Aquat. Conservation: Mar. Freshw. Ecosyst. 14, 363–379. doi: 10.1002/aqc.626
Bearzi G., Reeves R. R., Notarbartolo di Sciara G., Politi E., Cañadas A., Frantzis A., et al. (2003). Ecology, status and conservation of short-beaked common dolphins Delphinus delphis in the Mediterranean Sea. Mammal Rev. 33, 224–252. doi: 10.1046/j.1365-2907.2003.00032.x
Bearzi G., Reeves R. R., Remonato E., Pierantonio N., Airoldi S. (2011a). Risso’s dolphin Grampus griseus in the Mediterranean Sea. Mamm. Biol. 76, 385–400. doi: 10.1016/j.mambio.2010.06.003
Becker E. A., Forney K. A., Miller D. L., Barlow J., Rojas-Bracho L., Urbán R J., et al. (2022). Dynamic habitat models reflect interannual movement of cetaceans within the California current ecosystem. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.829523
Bigal E., Galili O., van Rijn I., Rosso M., Cleguer C., Hodgson A., et al. (2022). Reduction of species identification errors in surveys of marine wildlife abundance utilising unoccupied aerial vehicles (UAVs). Remote Sens. 14, 4118. doi: 10.3390/rs14164118
Boisseau O., Authier M., Bearzi G., Fortuna C., Fossi M., Frantzis A., et al. (2023). Complementing aerial surveys with ship-based visual and acoustic distance sampling surveys for monitoring cetaceans in the Mediterranean Sea. Mar. Policy 126, 104708.
Boisseau O., Reid J., Ryan C., Moscrop A., McLanaghan R., Panigada S. (2024). Acoustic estimates of sperm whale abundance in the Mediterranean Sea as part of the ACCOBAMS Survey Initiative. Front. Mar. Sci. 11. doi: 10.3389/fmars.2024.1164026
Buckland S. T. (Ed.) (2001). Introduction to distance sampling: estimating abundance of biological populations (Oxford ; New York: Oxford University Press).
Buckland S. T., Laake J. L., Borchers D. L. (2010). Double-observer line transect methods: levels of independence. Biometrics 66, 169–177. doi: 10.1111/j.1541-0420.2009.01239.x
Buckland S. T., Rexstad E. A., Marques T. A., Oedekoven C. S. (2015). Distance sampling: methods and applications (Cham: Springer International Publishing). doi: 10.1007/978-3-319-19219-2
Burt M. L., Borchers D. L., Jenkins K. J., Marques T. A. (2014). Using mark–recapture distance sampling methods on line transect surveys. Methods Ecol. Evol. 12, 1180–1191. doi: 10.1111/2041-210X.12294
Cañadas A., Aguilar de Soto N., Aissi M., Arcangeli A., Azzolin M., B-Nagy A., et al. (2018). The challenge of habitat modelling for threatened low density species using heterogeneous data: The case of Cuvier’s beaked whales in the Mediterranean. Ecol. Indic. 85, 128–136. doi: 10.1016/j.ecolind.2017.10.021
Cañadas A., Vázquez J. A. (2017). Common dolphins in the Alboran Sea: Facing a reduction in their suitable habitat due to an increase in Sea surface temperature. Deep Sea Res. Part II: Topical Stud. Oceanography 141, 306–318. doi: 10.1016/j.dsr2.2017.03.006
Cañadas A., Pierantonio N., Araújo H., David L., Di Meglio N., Dorémus G., et al (2023). Distribution patterns of marine megafauna density in the Mediterranean Sea assessed through the ACCOBAMS Survey Initiative (ASI). Front. Marine Sci. 10. doi: 10.3389/fmars.2023.1270917
Coll M., Piroddi C., Steenbeek J., Kaschner K., Lasram F. B. R., Aguzzi J., et al. (2010). The biodiversity of the Mediterranean Sea: estimates, patterns, and threats. PloS One 5, e11842. doi: 10.1371/journal.pone.0011842
Cotté C., Guinet C., Taupier-Letage I., Petiau E. (2010). Habitat use and abundance of striped dolphins in the western Mediterranean Sea prior to the morbillivirus epizootic resurgence. Endang. Species. Res. 12, 203–214. doi: 10.3354/esr00301
Dawson S., Wade P., Slooten E., Barlow J. (2008). Design and field methods for sighting surveys of cetaceans in coastal and riverine habitats. Mammal Rev. 38, 19–49. doi: 10.1111/j.1365-2907.2008.00119.x
DiMatteo A., Cañadas A., Roberts J., Sparks L., Panigada S., Boisseau O., et al. (2022). Basin-wide estimates of loggerhead turtle abundance in the Mediterranean Sea derived from line transect surveys. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.930412
Fandel A. D., Garrod A., Hoover A. L., Wingfield J. E., Lyubchich V., Secor D. H., et al. (2020). Effects of intense storm events on dolphin occurrence and foraging behavior. Sci. Rep. 10, 19247. doi: 10.1038/s41598-020-76077-3
Forcada J., Aguilar A., Hammond P. S., Pastor X., Aguilar R. (1994). Distribution and numbers of striped dolphins in the western Mediterranean Sea after the 1990 epizootic outbreak. Mar. Mammal Sci. 10, 137–150. doi: 10.1111/j.1748-7692.1994.tb00256.x
Forcada J., Aguilar A., Hammond P., Pastor X., Aguilar R. (1996). Distribution and abundance of fin whales (Balaenoptera physalus) in the western Mediterranean sea during the summer. J. Zoology 238, 23–34. doi: 10.1111/j.1469-7998.1996.tb05377.x
Forcada J., Notarbartolo di Sciara G., Fabbri F. (1995). Abundance of fin whales (Balenoptera physalus) and striped dolphins (Stenella coeruloalba) summering in the Corso-Ligurian Basin. Mammalia (France). 51 (9), 127–140.
Fortuna C. M., Cañadas A., Holcer D., Brecciaroli B., Donovan G. P., Lazar B., et al. (2018). The coherence of the European union marine natura 2000 network for wide-ranging charismatic species: A Mediterranean case study. Front. Mar. Sci. 5. doi: 10.3389/fmars.2018.00356
Fortuna C. M., Holcer D., Filidei E. Jr, Donovan G. P., Tunesi. L. (2011). “The first cetacean aerial survey in the Adriatic Sea: summer 2010,” in 7th Meeting of the ACCOBAMS Scientific committee, ACCOBAMS-SC7/2011/Doc06, 16pp.
Fortuna C. M., Kell L., Holcer D., Canese S., Mackelworth P., et al. (2014). Summer distribution and abundance of the giant devil ray (Mobula mobular) in the Adriatic Sea: Baseline data for an iterative management framework. Sci. Mar. 78, 227–237. doi: 10.3989/scimar.03920.30D. C.OMMAJ.R.X.X.X, E. F.
Frantzis A., Alexiadou P., Paximadis G., Politi E., Gannier A., Corsini-Foka M. (2003). Current knowledge of the cetacean fauna of the Greek Seas. J. Cetacean Res. Manage. 5, 219–232. doi: 10.47536/jcrm.v5i3.801
Frantzis A., Herzing D. L. (2002). ). Mixed-species associations of striped dolphins (Stenella coeruleoalba), short-beaked common dolphins (Delphinus delphis), and Risso’s dolphins (Grampus griseus) in the Gulf of Corinth (Greece, Mediterranean Sea). Aquat. Mammals 28, 188–197.
Fuentes M. M. P. B., Bell I., Hagihara R., Hamann M., Hazel J., Huth A., et al. (2015). Improving in-water estimates of marine turtle abundance by adjusting aerial survey counts for perception and availability biases. J. Exp. Mar. Biol. Ecol. 471, 77–83. doi: 10.1016/j.jembe.2015.05.003
Garcia-Garin O., Aguilar A., Borrell A., Gozalbes P., Lobo A., Penadés-Suay J., et al. (2020). Who’s better at spotting? A comparison between aerial photography and observer-based methods to monitor floating marine litter and marine mega-fauna. Environ. Pollut. 258, 113680. doi: 10.1016/j.envpol.2019.113680
Geijer C. K. A., Notarbartolo di Sciara G., Panigada S. (2016). Mysticete migration revisited: are Mediterranean fin whales an anomaly? Mam Rev. 46, 284–296. doi: 10.1111/mam.12069
Gilles A., Scheidat M., Siebert U. (2009). ‘Seasonal distribution of harbour porpoises and possible interference of offshore wind farms in the German North Sea’. Mar. Ecol. Prog. Ser. 383, 295–307. doi: 10.3354/meps08020
Gómez de Segura A., Crespo E. A., Pedraza S. N., Hammond P. S., Raga J. A. (2006). Abundance of small cetaceans in waters of the central Spanish Mediterranean. Mar. Biol. 150, 149–160. doi: 10.1007/s00227-006-0334-0
Grand J., Cummings M., Rebelo L.-M. (2007). Biodiversity, conservation biology, and rational choice. BioScience 57, 329–337. doi: 10.1641/B570412
Green R. E., Balmford A., Crane P. R., Mace G. M., Reynolds J. D., Turner R. K. (2005). A Framework for improved monitoring of biodiversity: responses to the world summit on sustainable development. Conserv. Biol. 19, 56–65. doi: 10.1111/j.1523-1739.2005.00289.x
Guerra C. A., Pendleton L., Drakou E. G., Proença V., Appeltans W., Domingos T., et al. (2019). Finding the essential: Improving conservation monitoring across scales. Global Ecol. Conserv. 18, e00601. doi: 10.1016/j.gecco.2019.e00601
Halpern B. S., Frazier M., Afflerbach J., Lowndes J. S., Micheli F., O’Hara C., et al. (2019). Recent pace of change in human impact on the world’s ocean. Sci. Rep. 9, 11609. doi: 10.1038/s41598-019-47201-9
Halpern B. S., Frazier M., Potapenko J., Casey K. S., Koenig K., Longo C., et al. (2015). Spatial and temporal changes in cumulative human impacts on the world’s ocean. Nat. Commun. 6, 7615. doi: 10.1038/ncomms8615
Hammond P., Lacey C., Gilles A., Viquerat S., Börjesson P., Herr H., et al. (2021). Estimates of cetacean abundance in European Atlantic waters in summer 2016 from the SCANS-III aerial and shipboard surveys. Final project report.
Hammond P. S., Macleod K., Berggren P., Borchers D. L., Burt L., Cañadas A., et al. (2013). Cetacean abundance and distribution in European Atlantic shelf waters to inform conservation and management. Biol. Conserv. 164, 107–122. doi: 10.1016/j.biocon.2013.04.010
Hansen R. G., Boye T. K., Larsen R. S., Nielsen N. H., Tervo O., Nielsen R. D., et al. (2019). Abundance of whales in West and East Greenland in summer 2015. NAMMCOSP 11. doi: 10.7557/3.4689
Heide-Jørgensen M. P., Bloch D., Stefansson E., Mikkelsen B., Ofstad L. H., Dietz R. (2002). Diving behaviour of long-finned pilot whales Globicephala melas around the Faroe Islands. wbio 8, 307–313. doi: 10.2981/wlb.2002.020
Heide-Jørgensen M. P., Laidre K. L., Simon M., Burt M. L., Borchers D. L., Rasmussen M. (2010). Abundance of fin whales in West Greenland in 2007. J. Cetacean Res. Manage. 11, 83–88. doi: 10.47536/jcrm.v11i2.614
Hiby L. (1999). “The objective identification of duplicate sightings in aerial survey for porpoise,” in Marine mammal survey and assessment methods (London: CRC Press).
Holt R. S., Cologne J. (1987). Factors affecting line transect estimates of dolphin school density. J. Wildlife Manage. 51, 836–843. doi: 10.2307/3801749
Hoyt E., Notarbartolo di Sciara G. (2021). Important Marine Mammal Areas: a spatial tool for marine mammal conservation. Oryx 55, 330–330. doi: 10.1017/S0030605321000272
Hughes F. M. R., Stroh P. A., Adams W. M., Kirby K. J., Mountford J.O., Warrington S. (2011). Monitoring and evaluating large-scale, ‘open-ended’ habitat creation projects: A journey rather than a destination. J. Nat. Conserv. 19, 245–253. doi: 10.1016/j.jnc.2011.02.003
IUCN (2023). The conservation status of cetaceans in the Mediterranean Sea: trends and changes after a decade of conservation efforts (Gland, Switzerland: IUCN).
Jefferson T. A., LeDuc R. (2018). “Delphinids, overview,” in Encyclopedia of marine mammals, 3rd ed. Eds. Würsig B., Thewissen J. G. M., Kovacs K. M. (London: Academic Press), 242–246. doi: 10.1016/B978-0-12-804327-1.00101-1
Kellner K. F., Swihart R. K. (2014). Accounting for imperfect detection in ecology: a quantitative review. PloS One 9, e111436. doi: 10.1371/journal.pone.0111436
Lambert C., Authier M., Dorémus G., Laran S., Panigada S., Spitz J., et al. (2020). Setting the scene for Mediterranean litterscape management: The first basin-scale quantification and mapping of floating marine debris. Environ. pollut. 263 (A), 114430. doi: 10.1016/j.envpol.2020.114430
Laran S., Authier M., Van Canneyt O., Dorémus G., Watremez P., Ridoux V. (2017a). A comprehensive survey of pelagic megafauna: their distribution, densities, and taxonomic richness in the tropical Southwest Indian Ocean. Front. Mar. Sci. 4. doi: 10.3389/fmars.2017.00139
Laran S., Pettex E., Authier M., Blanck A., David L., Dorémus G., et al. (2017b). Seasonal distribution and abundance of cetaceans within French waters- Part I: The North-Western Mediterranean, including the Pelagos sanctuary. Deep Sea Res. Part II: Topical Stud. Oceanography 141, 20–30. doi: 10.1016/j.dsr2.2016.12.011
Lauriano G., Pierantonio N., Donovan G., Panigada S. (2014). Abundance and distribution of Tursiops truncatus in the Western Mediterranean Sea: An assessment towards the Marine Strategy Framework Directive requirements. Mar. Environ. Res. 100, 86–93. doi: 10.1016/j.marenvres.2014.04.001
Lauriano G., Pierantonio N., Kell L., Cañadas A., Donovan G., Panigada S. (2017). Fishery-independent surface abundance and density estimates of swordfish (Xiphias gladius) from aerial surveys in the Central Mediterranean Sea. Deep Sea Res. Part II: Topical Stud. Oceanography 141, 102–114. doi: 10.1016/j.dsr2.2017.04.019
Lewis T., Boisseau O., Danbolt M., Gillespie D., Lacey C., Leaper R., et al. (2018). Abundance estimates for sperm whales in the Mediterranean Sea from acoustic line-transect surveys. J. cetacean Res. Manage. 18, 103–117. doi: 10.47536/jcrm.v18i1.437
Mannocci L., Roberts J. J., Halpin P. N., Authier M., Boisseau O., Bradai M. N., et al. (2018). Assessing cetacean surveys throughout the Mediterranean Sea: a gap analysis in environmental space. Sci. Rep. 8, 3126. doi: 10.1038/s41598-018-19842-9
Marques F. F. C., Buckland S. T. (2004). “Covariate models for the detection function,” in Advanced distance sampling. Eds. Buckland S. T., Anderson D. R., Burnham K. P., Laake J. L., Borchers D. L., Thomas L. (United Kingdom: Oxford University Press), 31–47.
Micheli F., Halpern B. S., Walbridge S., Ciriaco S., Ferretti F., Fraschetti S., et al. (2013). Cumulative human impacts on Mediterranean and black sea marine ecosystems: assessing current pressures and opportunities. PloS One 8, e79889. doi: 10.1371/journal.pone.0079889
Mussi B., Vivaldi C., Zucchini A., Miragliuolo A., Pace D. S. (2019). The decline of short-beaked common dolphin (Delphinus delphis) in the waters off the island of Ischia (Gulf of Naples, Italy). Aquat. Conserv: Mar. Freshw. Ecosyst. 31 (S1), aqc.3061. doi: 10.1002/aqc.3061
Nivière M., Blanchard A., Jraifi O., Van Canneyt O., Dorémus G., Spitz J., et al (2024). PelaSIG, a QGIS plugin for marine megafauna census: application to the aerial ACCOBAMS Survey Initiative dataset. Front. Marine Sci. 10. doi: 10.3389/fmars.2023.1270335
Notarbartolo di Sciara G. (2016). “Chapter one - marine mammals in the Mediterranean Sea: an overview,” in Advances in marine biology Mediterranean marine mammal ecology and conservation. Eds. Notarbartolo di Sciara G., Podestà M., Curry B. E. (Amsterdam: Academic Press), 1–36. doi: 10.1016/bs.amb.2016.08.005
Notarbartolo di Sciara G., Castellote M., Druon J.-N., Panigada S. (2016). “Chapter three - fin whales, balaenoptera physalus: at home in a changing mediterranean sea?,” in Advances in marine biology Mediterranean marine mammal ecology and conservation. Eds. Notarbartolo di Sciara G., Podestà M., Curry B. E. (Amsterdam: Academic Press), 75–101. doi: 10.1016/bs.amb.2016.08.002
Notarbartolo di Sciara G., Lauriano G., Pierantonio N., Cañadas A., Donovan G., Panigada S. (2015). The devil we don’t know: investigating habitat and abundance of endangered giant devil rays in the North-Western Mediterranean Sea. PloS One 10 (11), e0141189. doi: 10.1371/journal.pone.0141189
Nykänen M., Jessopp M., Doyle T. K., Harman L. A., Cañadas A., Breen P., et al. (2018). Using tagging data and aerial surveys to incorporate availability bias in the abundance estimation of blue sharks (Prionace glauca). PloS One 13, e0203122. doi: 10.1371/journal.pone.0203122
Nykänen M., Oudejans M. G., Rogan E., Durban J. W., Ingram S. N. (2020). Challenges in monitoring mobile populations: Applying bayesian multi-site mark–recapture abundance estimation to the monitoring of a highly mobile coastal population of bottlenose dolphins. Aquat. Conservation: Mar. Freshw. Ecosyst. 30, 1674–1688. doi: 10.1002/aqc.3355
Panigada S., Gauffier P., Notarbartolo di Sciara G. (2021). IUCN Red List of Threatened Species: Balaenoptera physalus Mediterranean subpopulation (IUCN Red List of Threatened Species). Available at: https://www.iucnredlist.org/species/16208224/50387979 (Accessed January 31, 2022).
Panigada S., Lauriano G., Donovan G., Pierantonio N., Cañadas A., Vázquez J. A., et al. (2017). Estimating cetacean density and abundance in the Central and Western Mediterranean Sea through aerial surveys: Implications for management. Deep Sea Res. Part II: Topical Stud. Oceanography. 141, 41–58 doi: 10.1016/j.dsr2.2017.04.018
Panigada S., Notarbartolo di Sciara G., Panigada M. Z. (2006). Fin whales summering in the Pelagos Sanctuary (Mediterranean Sea): Overview of studies on habitat use and diving behaviour. Chem. Ecol. 22, S255–S263. doi: 10.1080/02757540600720680
Panigada S., Lauriano G., Burt L., Pierantonio N., Donovan G. (2011). Monitoring winter and summer abundance of cetaceans in the Pelagos Sanctuary (Northwestern Mediterranean Sea) through aerial surveys. PLoS ONE 6 (7), e22878. doi: 10.1371/journal.pone.0022878
Pereira H. M., Cooper D. H. (2006). Towards the global monitoring of biodiversity change. Trends Ecol. Evol. 21, 123–129. doi: 10.1016/j.tree.2005.10.015
Pettex E., David L., Authier M., Blanck A., Dorémus G., Falchetto H., et al. (2017). Using large scale surveys to investigate seasonal variations in seabird distribution and abundance. Part I: The North Western Mediterranean Sea. Deep Sea Res. Part II: Topical Stud. Oceanography 141, 74–85. doi: 10.1016/j.dsr2.2016.11.008
Pollock K. H., Marsh H. D., Lawler I. R., Alldredge M. W. (2006). Estimating animal abundance in heterogeneous environments: an application to aerial surveys for dugongs. J. Wildlife Manage. 70, 255–262. doi: 10.2193/0022-541X(2006)70[255:EAAIHE]2.0.CO;2
Porszt E. J., Peterman R. M., Dulvy N. K., Cooper A. B., Irvine J. R. (2012). Reliability of indicators of decline in abundance. Conserv. Biol. 26, 894–904. doi: 10.1111/j.1523-1739.2012.01882.x
Quick N. J., Cioffi W. R., Shearer J. M., Fahlman A., Read A. J. (2020). Extreme diving in mammals: first estimates of behavioural aerobic dive limits in Cuvier’s beaked whales. J. Exp. Biol. 223, jeb222109. doi: 10.1242/jeb.222109
Raudino H. C., Cleguer C., Hamel M. A., Swaine M., Waples K. A. (2022). Species identification of morphologically similar tropical dolphins and estimating group size using aerial imagery in coastal waters. Mamm Biol. 102, 829–839. doi: 10.1007/s42991-021-00214-2
Roberts J. J., Best B. D., Dunn D. C., Treml E. A., Halpin P. N. (2010). Marine Geospatial Ecology Tools: An integrated framework for ecological geoprocessing with ArcGIS, Python, R, MATLAB, and C++. Environ. Model. Software 25, 1197–1207. doi: 10.1016/j.envsoft.2010.03.029
Rogan E., Breen P., Mackey M., Cañadas A., Scheidat M., Geelhoed S. C. V., et al. (2018). Aerial surveys of cetaceans and seabirds in Irish waters: occurrence, distribution and abundance in 2015-2017. Available at: https://research.wur.nl/en/publications/aerial-surveys-of-cetaceans-and-seabirds-in-irish-waters-occurren (Accessed July 19, 2023).
Scheidat M., Gilles A., Kock K.-H., Siebert U. (2008). Harbour porpoise Phocoena phocoena abundance in the southwestern Baltic Sea. Endangered Species Res. 5, 215–223. doi: 10.3354/esr00161
Smolensky N. L., Fitzgerald L. A. (2011). Population variation in dune-dwelling lizards in response to patch size, patch quality, and oil and gas development. Southwestern Nat. 56, 315–324. doi: 10.1894/F03-MLK-21.1
Stock A., Crowder L. B., Halpern B. S., Micheli F. (2018). Uncertainty analysis and robust areas of high and low modeled human impact on the global oceans. Conserv. Biol. 32, 1368–1379. doi: 10.1111/cobi.13141
Strindberg S., Buckland S. T. (2004). Zigzag survey designs in line transect sampling. JABES 9, 443. doi: 10.1198/108571104X15601
Tardy C., Ody D., Gimenez O., Planes S. (2023). Abundance of fin whales (Balaenoptera physalus) in the north-western Mediterranean Sea, using photo-identification and microsatellite genotyping. Mar. Ecol. 44, e12737. doi: 10.1111/maec.12737
Taylor B. L., Chivers S. J., Larese J., Perrin W. F. (2007). Generation length and percent mature estimates for IUCN assessments of Cetaceans (La Jolla, California: NOAA Southwest Fisheries Science Center). Administrative Report LJ-07-01.
Tetley M. J., Braulik G. T., Lanfredi C., Minton G., Panigada S., Politi E., et al. (2022). The important marine mammal area network: a tool for systematic spatial planning in response to the marine mammal habitat conservation crisis. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.841789
Thomas L., Buckland S. T., Burnham K. P., Anderson D. R., Laake J. L., Borchers D. L., et al. (2012). “Distance sampling,” in Encyclopedia of environmetrics. Eds. El-Shaarawi A. H., Piegorsch W. W. (Chichester, West Sussex: Wiley). doi: 10.1002/9780470057339.vad033.pub2
Thomas L., Buckland S. T., Rexstad E. A., Laake J. L., Strindberg S., Hedley S. L., et al. (2010). Distance software: design and analysis of distance sampling surveys for estimating population size. J. Appl. Ecol. 47, 5–14. doi: 10.1111/j.1365-2664.2009.01737.x
Thomas H. L., Hockey P. A., Cumming G. S. (2015). Solving the challenges of monitoring mobile populations: insights from studies of waterbirds in southern Africa. Ostrich 86, 169–178. doi: 10.2989/00306525.2015.1005720
Thomson J. A., Heithaus M. R., Burkholder D. A., Vaudo J. J., Wirsing A. J., Dill L. M. (2012). Site specialists, diet generalists?: Isotopic variation, site fidelity, and foraging by loggerhead turtles in Shark Bay, Western Australia. Mar. Ecol. Prog. Ser. 453, 213–226. doi: 10.3354/meps09637
van Aswegen M., Christiansen F., Symons J., Mann J., Nicholson K., Sprogis K., et al. (2019). Morphological differences between coastal bottlenose dolphin (Tursiops aduncus) populations identified using non-invasive stereo-laser photogrammetry. Sci. Rep. 9, 12235. doi: 10.1038/s41598-019-48419-3
van Weelden C., Towers J. R., Bosker T. (2021). Impacts of climate change on cetacean distribution, habitat and migration. Climate Change Ecol. 1, 100009. doi: 10.1016/j.ecochg.2021.100009
Wilson H. B., Kendall B. E., Possingham H. P. (2011). Variability in population abundance and the classification of extinction risk. Conserv. Biol. 25, 747–757. doi: 10.1111/j.1523-1739.2011.01671.x
Zanardelli M., Airoldi S., Bérubé M., Borsani J. F., Di-Meglio N., Gannier A., et al. (2022). Long-term photo-identification study of fin whales in the Pelagos Sanctuary (NW Mediterranean) as a baseline for targeted conservation and mitigation measures. Aquat. Conservation: Mar. Freshw. Ecosyst. 32, 1457–1470. doi: 10.1002/aqc.3865
Keywords: Mediterranean, density and abundance, aerial surveys, cetacean, conservation, large-scale surveys
Citation: Panigada S, Pierantonio N, Araújo H, David L, Di-Méglio N, Dorémus G, Gonzalvo J, Holcer D, Laran S, Lauriano G, Paiu R-M, Perri M, Popov D, Ridoux V, Vázquez JA and Cañadas A (2024) The ACCOBAMS survey initiative: the first synoptic assessment of cetacean abundance in the Mediterranean Sea through aerial surveys. Front. Mar. Sci. 10:1270513. doi: 10.3389/fmars.2023.1270513
Received: 31 July 2023; Accepted: 29 November 2023;
Published: 07 February 2024.
Edited by:
Michele Thums, Australian Institute of Marine Science (AIMS), AustraliaReviewed by:
Nat Kelly, Australian Antarctic Division, AustraliaHolly Crystal Raudino, Conservation and Attractions (DBCA), Australia
Copyright © 2024 Panigada, Pierantonio, Araújo, David, Di-Méglio, Dorémus, Gonzalvo, Holcer, Laran, Lauriano, Paiu, Perri, Popov, Ridoux, Vázquez and Cañadas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Simone Panigada, cGFuaWdhZGE2OUBnbWFpbC5jb20=; Nino Pierantonio, bmluby5waWVyYW50b25pb0Bwcm90b25tYWlsLmNvbQ==