 Jana Verdura1*†
Jana Verdura1*† Lluc Rehues2†
Lluc Rehues2† Luisa Mangialajo1
Luisa Mangialajo1 Simonetta Fraschetti3,4
Simonetta Fraschetti3,4 Zahira Belattmania5
Zahira Belattmania5 Silvia Bianchelli4,6
Silvia Bianchelli4,6 Aurélie Blanfuné7
Aurélie Blanfuné7 Brahim Sabour5
Brahim Sabour5 Antonia Chiarore8
Antonia Chiarore8 Roberto Danovaro4,6
Roberto Danovaro4,6 Erika Fabbrizzi3,4
Erika Fabbrizzi3,4 Sylvaine Giakoumi9
Sylvaine Giakoumi9 Ljiljana Iveša10
Ljiljana Iveša10 Stelios Katsanevakis11
Stelios Katsanevakis11 Eleni Kytinou11Ina Nasto12
Eleni Kytinou11Ina Nasto12 Athanasios Nikolaou11
Athanasios Nikolaou11 Sotiris Orfanidis13
Sotiris Orfanidis13 Gil Rilov14
Gil Rilov14 Fabio Rindi4,6Marta Sales15
Fabio Rindi4,6Marta Sales15 Maria Sini11
Maria Sini11 Laura Tamburello4,9
Laura Tamburello4,9 Thierry Thibaut7
Thierry Thibaut7 Konstantinos Tsirintanis11
Konstantinos Tsirintanis11 Emma Cebrian16
Emma Cebrian16- 1UMR 7035 ECOSEAS, Université Côte d’Azur, CNRS, Nice, France
- 2GRMAR, Institut of Aquatic Ecology, University of Girona, Girona, Spain
- 3Department of Biology, University of Naples Federico II, Naples, Italy
- 4National Biodiversity Future Centre, Palermo, Italy
- 5Research Unit (RU) Phycology, Blue Biodiversity & Biotechnology, URL CNRST 10 – Laboratory of Plant Biotechnology, Ecology and Ecosystem Valorization, Faculty of Sciences, University Chouaïb Doukkali, El Jadida, Morocco
- 6Dipartimento di Scienze della Vita e dell’Ambiente, Università Politecnica delle Marche, Ancona, Italy
- 7Aix-Marseille University, Mediterranean Institute of Oceanography (MIO), CNRS/INSU, IRD, UM 110, Marseille, France
- 8Department of Integrative Marine Ecology, Stazione Zoologica Anton Dohrn, Naples, Italy
- 9Sicily Marine Centre, Stazione Zoologica Anton Dohrn, Lungomare Cristoforo Colombo (complesso Roosevelt), Palermo, Italy
- 10Center for Marine Research, Ruđer Bošković Institute, Rovinj, Croatia
- 11Department of Marine Sciences, University of the Aegean, Mytilene, Greece
- 12Department of Biology, University “Ismail Qemali”, Vlora, Albania
- 13Fisheries Research Institute, Hellenic Agricultural Organization-Demeter, Kavala, Greece
- 14National Institute of Oceanography, Israel Oceanographic and Limnological Research (ILOR), Haifa, Israel
- 15Institut Menorquí d’Estudis, Maó, Spain
- 16Department Ecologia Marina, Centre d’Estudis Avançats de Blanes-CSIC, Blanes, Spain
The worldwide decline of macroalgal forests is raising major concerns for the potentially negative consequences on biodiversity and ecosystem functions, pushing for the definition of specific conservation and restoration measures. Protecting and restoring these habitats requires detailed information on their distribution, ecological status, and drivers of decline. Here, we provide the most updated available information on the distribution of Mediterranean Cystoseira s.l. forests by conducting a comprehensive bibliographic survey of literature published from 2009 to 2021, complemented by unpublished data. We also provide insights into the ecological status of these forests and the stressors affecting them across the Mediterranean basin. Our results show that most Mediterranean coasts remain un(der)studied and that the available information is concentrated in spatially limited coastal areas, restricted to very few species. When the ecological status is reported, data is highly heterogeneous, making any comparisons problematic, what claims for the description and use of easy and standardized monitoring methods for comparative purposes. Drivers of decline of Cystoseira s.l. forest have been even less investigated and, therefore, still poorly characterized. Our results highlight that our current knowledge is still insufficient to implement effective conservation and restoration strategies at the basin scale but also regionally. We call for the urgent need for mapping and standardized monitoring of Cystoseira s.l. forests to obtain baseline information for future management strategies involving their conservation, the mitigation of the stressors threatening them and the restoration of the degraded forests.
1 Introduction
Marine algal forests are typically dominated by brown canopy-forming macroalgae from the Orders Laminariales, Fucales, Tilopteridales and Desmarestiales. These habitats produce important biomass and create 3D structures of high fractal complexity that host and feed a myriad of associated species (e.g., Cheminée et al., 2013; Teagle et al., 2017). Thus, these forests represent some of the most productive and biodiverse coastal marine ecosystems in temperate and polar regions on the planet (Mann, 1973; Steneck et al., 2002) while also providing essential ecosystem goods and services for human society (Smale et al., 2013). Canopy-forming macroalgal species exhibit a great diversity of growth forms, life strategies and environmental requirements; therefore, forest-forming species vary among regions worldwide. Temperate and Arctic rocky coastlines are mostly dominated by different species of kelps (Laminariales) (Wernberg et al., 2019). In the Mediterranean Sea, different macroalgal (Fucales) species, mainly of the genera Cystoseira, Ericaria and Gongolaria (Orellana et al., 2019; Molinari-Novoa and Guiry, 2020) (hereafter reported as Cystoseira s.l.) shape rocky seascapes from the upper infralittoral to the upper circalittoral zones (Giaccone and Bruni, 1973).
Globally, many brown canopy-forming macroalgal species are increasingly affected in several regions by the cumulative impacts of local and global anthropogenic stressors which raises major concern for their conservation (e.g., Johnson et al., 2011; Smale and Wernberg, 2013; Wernberg et al., 2013; Thibaut et al., 2015; Araújo et al., 2016; Smale, 2020). Their loss often triggers a transition towards less diverse and productive habitats, such as sea urchin barrens or turf-dominated grounds, in which stabilizing feedback processes reinforce their persistence (Thibaut et al., 2005; Ling et al., 2015; Filbee-Dexter and Wernberg, 2018). Specifically, Cystoseira s.l. species are highly vulnerable to several human disturbances. Indeed, widespread declines in their distribution throughout the Mediterranean basin have been reported, mainly linked to habitat destruction, overgrazing and deterioration in water quality (Munda, 1993; Thibaut et al., 2005; Bianchi et al., 2014; Mariani et al., 2019). Some (if not many) species may also be sensitive to global change, for example, ocean warming (e.g., Verdura et al., 2021; Mulas et al., 2022). In response, since the 1980s and 1990s, some efforts aimed at the conservation of Cystoseira s.l. forests have been made, such as stablishing protection status (e.g., being listed as threatened or endangered species in the Annex II of the Barcelona Convention; UNEP/MAP, 2013), or requiring the local mitigation of human pressures (e.g., sewage treatment management plans; Pinedo et al., 2013). However, despite these efforts, the natural recovery of Cystoseira s.l. forests is frequently unachievable. Although isolated instances of natural recolonization have been documented (Iveša et al., 2016; Zavodnik et al., 2020), the overall limited dispersal ability of these species and the scarcity of nearby propagules likely impede this process (Iveša et al., 2016; Zavodnik et al., 2020), suggesting the need for active restoration.
With the increasing awareness of the need for natural ecosystems conservation and concern about human threats, ecological restoration has been included in the policy agenda at global, regional, and national levels, especially following the objectives of different initiatives, such as the UN decade on Ecosystem Restoration (Waltham et al., 2020) or the EU Biodiversity Strategy for 2030 (EC, 2020) and the proposed UE Nature Restoration Law (EC, 2022). Macroalgal restoration attempts are less common than those for other marine habitats (Duarte et al., 2020; Fraschetti et al., 2021). However, during the last decade, macroalgae restoration knowledge has significantly progressed through an increasing number of funded projects in the Mediterranean Sea (e.g., AFRIMED, ROC-POPLife or MERCES EU projects), as well as in other areas of the world (e.g., the Green Gravel or Operation Crayweed projects). These projects have provided different methodologies and protocols leading to the recovery of structural habitat-forming species (e.g., Cystoseira s.l., Phyllospora comosa or Saccharina latissima) as a herald for the recovery of overall associated biodiversity and ecosystem functions, showing active restoration as a potential successful tool to recover marine forests (Layton et al., 2020; Cebrian et al., 2021; Eger et al., 2022; Galobart et al., 2023).
Site prioritization is one of the first challenges for successful restoration outcomes (McDonald et al., 2016). Site selection can be crucial in marine ecosystems to achieving the restoration targets and reducing costs (Bayraktarov et al., 2016; Fraschetti et al., 2021). The primary eligibility criteria for spatial prioritizing restoration sites should be based on the historical presence of the habitat or species targeted for restoration. Equal importance should be given to assessing the suitability of the current and future biotic conditions, such as the presence of potentially harmful invasive species or herbivory pressure, as well as abiotic conditions, such as bottom type, hydrodynamics, or sea-water temperature, within the selected area (Fabbrizzi et al., 2023; Smith et al., 2023). Crucial information to consider thus includes fine-scale data on the past and present distribution of the species, as well as the ecological status of populations. It is also important to identify the causal stressor/s responsible for population declines (Gann et al., 2019). Nonetheless, comprehensive knowledge concerning marine forests is generally limited to a few species and regions with mostly short-time data series, primarily due to the scarcity of long-term monitoring and limited cartographic information about these habitats (Krumhansl et al., 2016).
The Mediterranean Sea is not exempt from this prevailing trend. Remarkable efforts have been dedicated to the compilation of historical and contemporary records in specific regions of France (Thibaut et al., 2005; Thibaut et al., 2014; Thibaut et al., 2015; Thibaut et al., 2016; Blanfuné et al., 2016a, Blanfuné et al., 2019), Catalonia (Mariani et al., 2019), Italy (Tamburello et al., 2022; Rendina et al., 2023), Croatia (Iveša et al., 2014, Iveša et al., 2016) and Montenegro (Mačić and Antolić, 2015). The information acquired holds significant value; however, the process of data acquisition is often arduous due to the need for screening technical reports, monitoring programs, herbarium specimens, scientific literature, and unpublished data from experts and projects. Regrettably, the only studies collecting and mapping all available data (considering historical and present records from 1985 to 2018) regarding the occurrences of Cystoseira s.l. forests across the Mediterranean Sea, indicate that such knowledge is only accessible for approximately 15% of the Mediterranean coasts (Fabbrizzi et al., 2020; Fabbrizzi et al., 2023). In this context, despite having a broad view of the historical and current distribution knowledge, a comprehensive overview of the available and updated knowledge on Cystoseira species’ current distribution, ecological status, and the stressing factors contributing to their decline across the Mediterranean basin is lacking. Furthermore, the distribution and intensity of local and global stressors responsible for population collapse have rarely been validated on case-by-case (Mancuso et al., 2018; Mariani et al., 2019). Finally, this critical knowledge gap across Mediterranean regions hampers our capacity to effectively manage and restore these habitats in a rapidly changing ocean.
In this context, through a systematic scientific literature review and the exploitation of unpublished data together with expert judgment, this work provides a picture of the updated (from 2009) and accessible knowledge about Cystoseira s.l. forests at the Mediterranean scale. More especially, we aim to (1) provide a complete overview of the updated available information on the distribution and ecological status of Cystoseira s.l. forests, (2) identify the drivers of Cystoseira s.l. forests loss, and (3) determine any geographic pattern of these drivers. With the final aim to optimize future management, conservation, and restoration efforts on these ecosystems, our results may be used as an essential baseline to determine which features (e.g., species, stressors, etc.) and areas show major gaps, allowing the identification of where more investments are needed to allocate funds, monitoring and sampling effort (e.g., mapping; Fabbrizzi et al., 2020; Fabbrizzi et al., 2023). Moreover, it can also assist in determining where to focus on impact mitigation and where restoration and conservation plans should be prioritized.
2 Materials and methods
2.1 Information search
2.1.1 Systematic bibliographic assessment
We performed a systematic literature review consisting of (1) relevant article identification, (2) eligibility evaluation and (3) detailed revision of selected articles. The article identification was conducted using three databases: Web of Science (WoS), SCOPUS, and ScienceDirect, limiting the search to peer-reviewed articles published from 2009 to 2021 (cut-off dates 1st January 2009 and 31st December 2021 specifically). The starting cut-off date was decided according to the aim of assessing the current state of Cystoseira s.l. forests, and thus, avoiding older publications that might contain outdated data. WoS facilitates access to different databases and searches the parameters screening in the Title, the Abstract and the Keywords. SCOPUS and ScienceDirect were used to search the parameters within the Title, the Abstract, the Keywords, and the Text. The search query used was the same for the three databases. It was designed to return generic results:
(“Cystoseira” AND “Mediterranean”) OR (“Canopy” AND “Algae” AND “Mediterranean”).
Given the fact that the taxonomy of Cystoseira s.l. has been in constant change during the last few years (Orellana et al., 2019; Molinari-Novoa and Guiry, 2020; Neiva et al., 2022), we performed four consecutive searches each time replacing the word “Cystoseira” by “Carpodesmia”, “Treptacantha”, “Ericaria” and “Gongolaria”. Due to this reinstatement, most of the species’ names used in the revised publications before 2020 are no longer accepted, and thus accepted species names, according to AlgaeBase (Molinari-Novoa and Guiry, 2020), are used in the present study.
The articles identified by the different searches and databases were combined, and duplicates were excluded manually using the EndNote software. After that, we conducted an eligibility evaluation to assess the articles’ suitability for data extraction. This step consisted of screening the abstract, where a first assessment was carried out, and if favourable, an in-depth check of the articles followed. Articles were excluded when: (a) the study area was not within the Mediterranean Sea, (b) the article studied canopy-forming algae other than Cystoseira s.l., (c) no specific geographical information of Cystoseira s.l. populations was provided, and (d) authors used the same sampling site to conduct different studies and published more than one article. When the latter was detected, the articles were regarded as duplicates; hence, only one of the different articles was kept for data extraction. As a result, 138 articles were used for data extraction.
2.1.2 Complementary search and personal communications
Some areas of the Mediterranean Sea were underrepresented in terms of data availability. This unbalance might be due to certain studies being published in local or regional journals (often non-indexed) or/and in the country’s official language and, therefore, may have gone unnoticed by the databases used in the first systematic search. To balance these differences, subsequent searches using Google Scholar and targeting those specific areas were carried out. Citations in the articles reviewed were also checked when considered relevant. This search added 15 articles to those previously selected.
Lastly, all authors contributed data, encompassing extensions of existing datasets from prior publications or technical reports. These data sources encompassed a broad spectrum, including information from monitoring programs and frequently sampled areas from various national and international projects.
2.2 Data extraction
To map the reported knowledge on the distribution, ecological status, and stressors of Cystoseira s.l. forests, key information was extracted from each article when available: Cystoseira s.l. species, geographical information (including coordinates, site name, region, country), ecological status (Good, Moderate, Bad), presence of stressors and stressor type, dynamics of the population (Increased, Stable, Decreased abundance) and sampling year.
When coordinates were not provided, location names were searched on websites and georeferenced using QGIS 3.26 software, allowing the extraction of coordinates with the same program. When sampling points were only provided in a figure map, points were manually georeferenced using QGIS 3.26 software. To identify regional geographical differences, the Mediterranean Sea was divided into eight sub-basins, following the framework proposed by Teruzzi et al. (2011) with a few minor modifications. These adjustments were made considering the regional identification from our bibliographic research, where Adriatic Sea is considered as a single sub-basin and the entire Balearic archipelago is considered into the same sub-basin.
The ecological status of populations was derived from different types of variables. Some articles used numerical variables, while others used categorical ones. Categorical data consisted of descriptions and were highly heterogeneous. As a rule, if a population was described as “forest”, “continuous belt”, or “dominant”, it was classified as Good; if it was described as “patches”, it was classified as Moderate; and if it was described as “Scattered individuals”, “Barren ground”, or “Absent after a population extinction” it was classified as Bad. The georeferenced points reporting the absence of Cystoseira s.l. species but not specifying the reason, were excluded from this analysis. The numerical variables were wet weight per unit area, density (number of thalli per surface), or percentage cover of individuals, the latter being the most common. Different thresholds were set for each of these variables to classify the numeric values into the three established categories in this study. The criteria details used to classify the extracted data into the three categories of ecological status (Good, Moderate, or Bad) are described in Table 1.
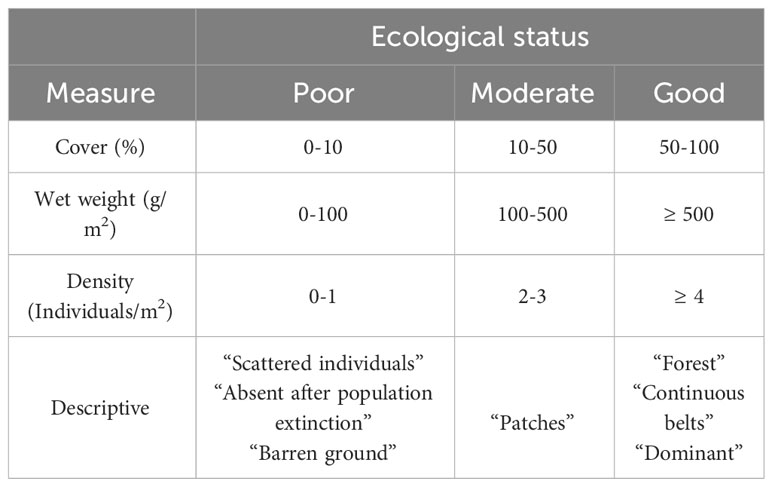
Table 1 Criteria used to classify the different data obtained through different variables within the three ecological states established in this study: Poor, Moderate, and Good.
Information about the presence or absence and the type of stressors affecting Cystoseira s.l. populations were also extracted from the articles, when available. Different types of stressors were classified into different categories, defined in Table 2.
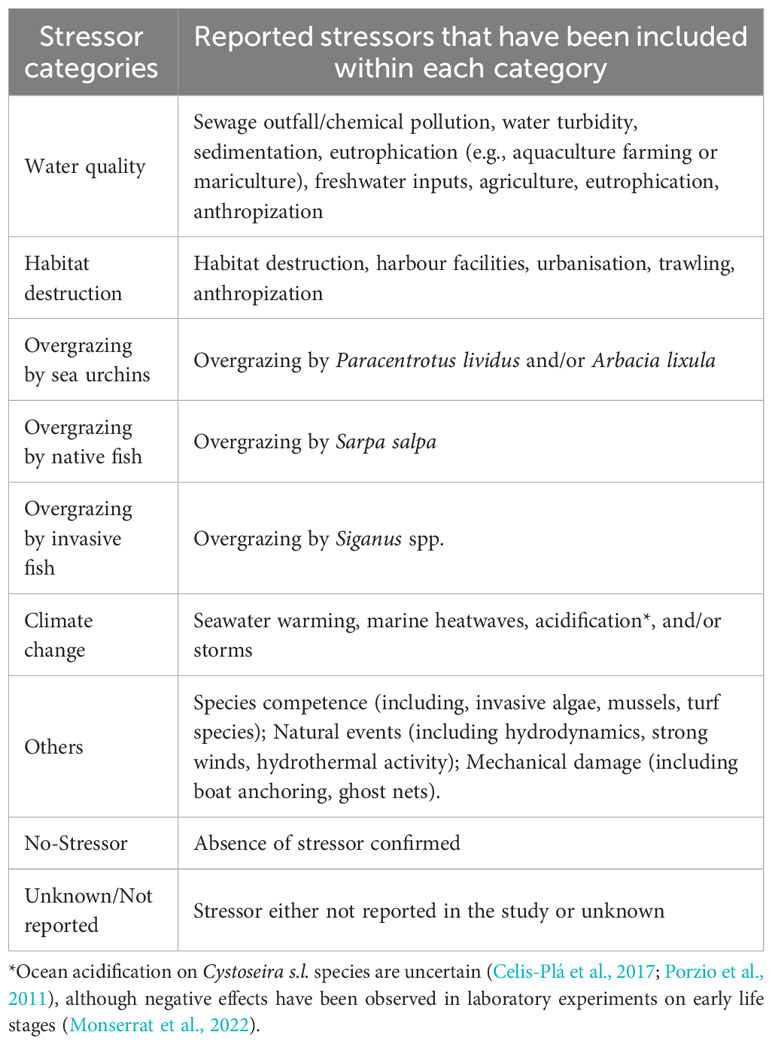
Table 2 Classification of the different reported stressors within the established “stressor categories” in this study.
2.3 Data processing and representation
The resulting database consisted of georeferenced points with different attributes. These points were projected into a map using the Geographic Information System (GIS) software QGIS 3.26. The coordinate representation system used was WGS84 (EPSG: 4326). The spatial data were processed using ArcGIS Pro 3.0.0 software, and all graphics were generated using ggplot2 package (Wickham, 2016) in the statistical environment R (R Core Team, 2019). The spatial analysis consisted of assessing (1) data availability, (2) ecological status, and (3) stressor occurrence.
2.3.1 Data availability
To study the spatial availability of the compiled data, a grid of 10 km x 10 km was projected on a map of the Mediterranean Sea, and then only the cells that intersected with the coastline were kept. Each cell was assigned a distinct colour based on the number of georeferenced points it encompassed (Figure 1). The number of occurrences (presence, absence, and extinction) per Cystoseira s.l. species was also represented in a bar plot (Figure 2).
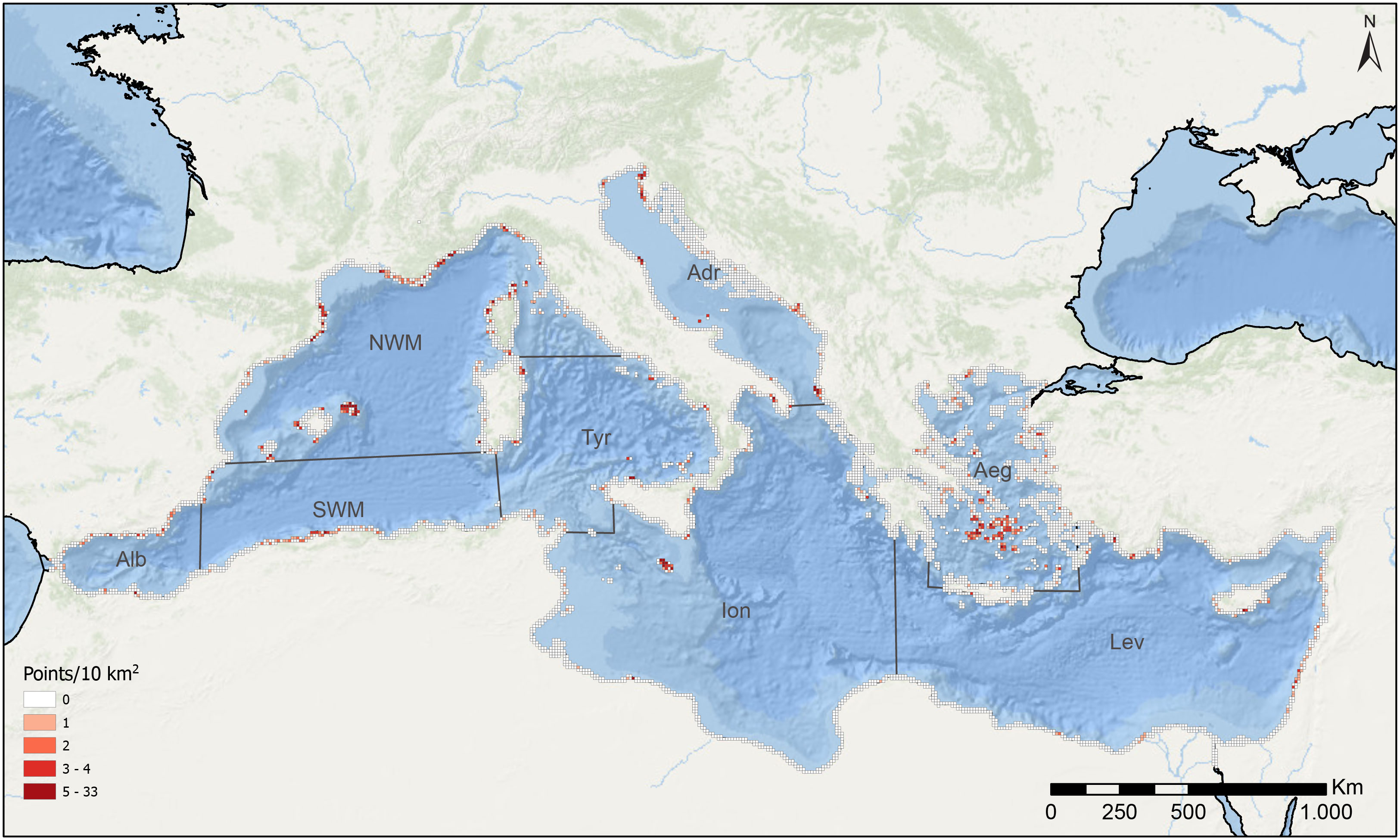
Figure 1 Data availability. Distribution of all sampled points of Cystoseira s.l. populations along the Mediterranean basin that have been obtained in the data extraction process. In the map, each square encompasses an area of 10 km2, the colour of which represents the number of sampling points within the given area. The different divisions represent the Mediterranean sub-basins defined in this study (modified from Terruzi et al., 2011): Alb: Alboran Sea, NWM: North-Western Mediterranean, SWM: South-Western Mediterranean, Tyr: Tyrrhenian Sea, Adr: Adriatic Sea, Ion: Ionian Sea, Aeg: Aegean Sea, Lev: Levantine Sea.
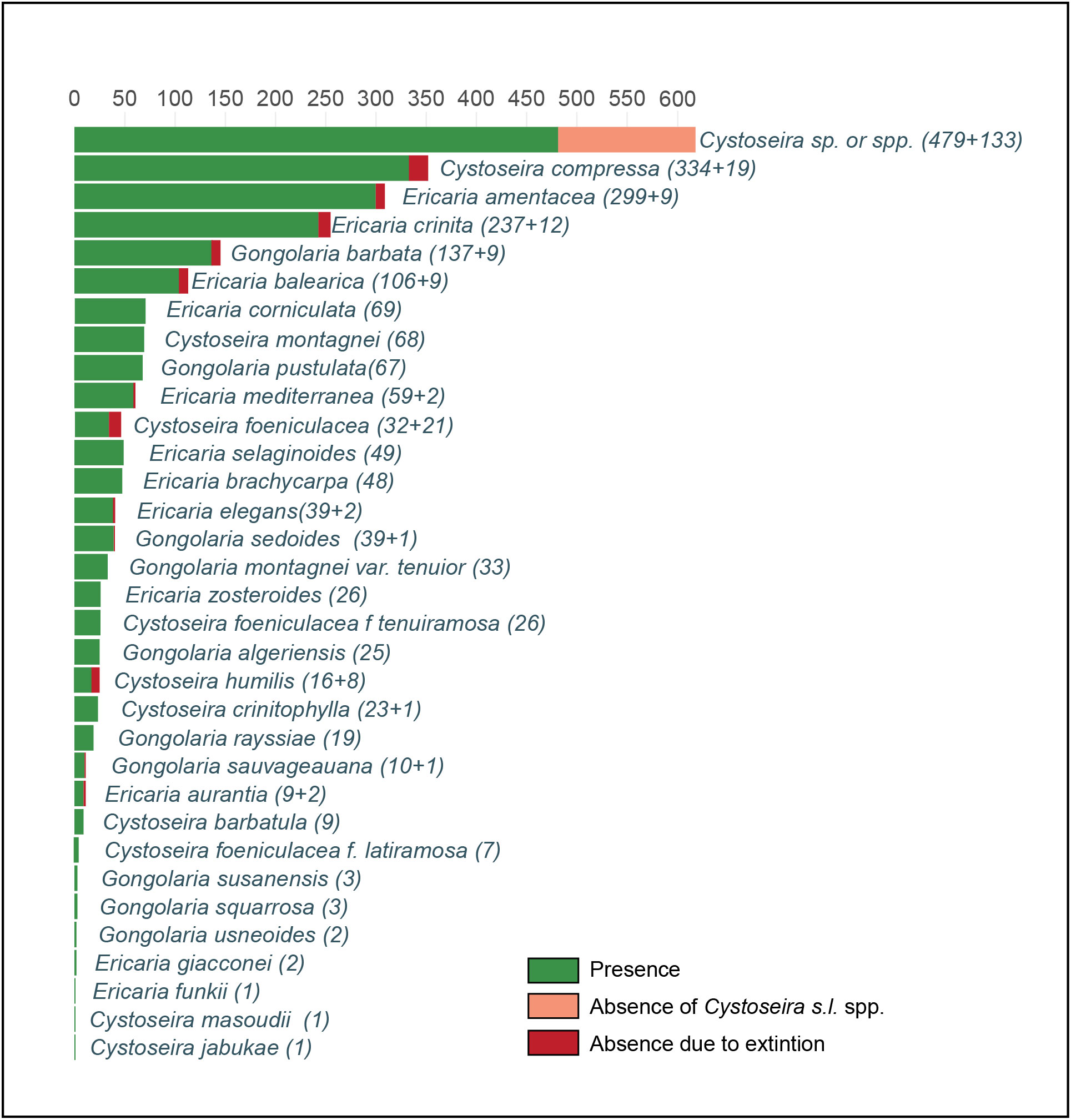
Figure 2 Number of reported occurrences (including past and present presence and absence) of Cystoseira s.l. species, based on the available data.
2.3.2 Ecological status
The points that contained information about the ecological status of Cystoseira s.l. assemblages were classified as Poor, Moderate or Good (Table 1), as explained above. A numerical value was given for each ecological status category; the locations with “Poor” ecological status were given a value of 1, “Moderate” a value of 2, and “Good” a value of 3. Using the same grid described in the Data availability section (2.3.1), the average ecological status was calculated in each cell (Figure 3). The values obtained were rounded to the closest integer. Supplementary Materials S4 contains more detailed maps of occurrences with associated ecological information. Specifically, for regions exhibiting a higher density of points, the ecological value attributed to each individual point is presented. Finally, the number of points of each ecological status category and the number of points without ecological status information (Unknown category) were also represented per sub-basins in a bar plot (Figure 4).
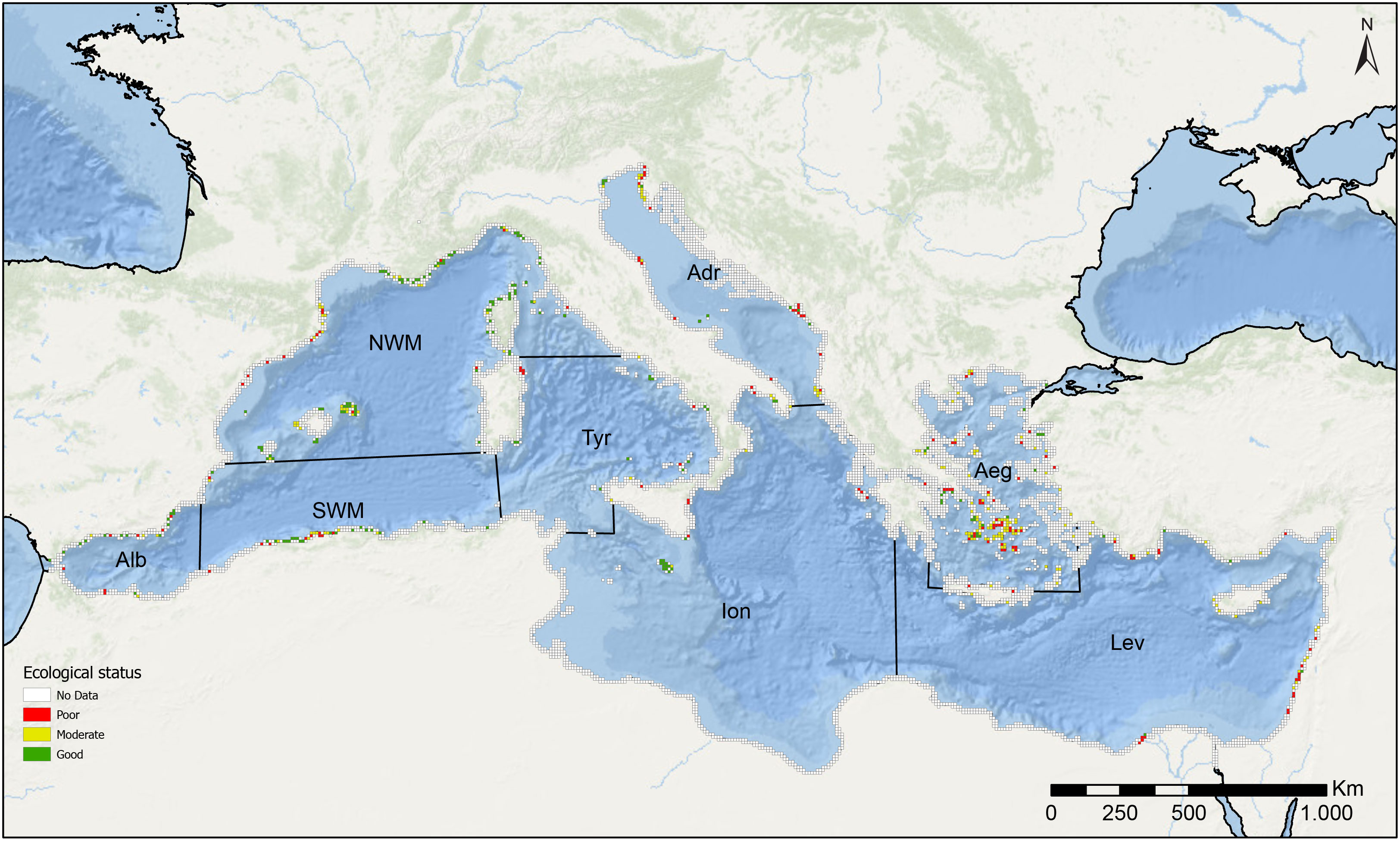
Figure 3 Ecological status. Distribution of the sampling points reporting information regarding the ecological status of Cystoseira s.l. populations along the Mediterranean basin that have been obtained in the data extraction process. Each square encompasses an area of 10 km2, the colour of which shows the mean ecological status of the different points within the given area. The mean ecological status value has been obtained from the average value of the ecological status of the different sampling points that are located within the area of each square. The different divisions represent the Mediterranean sub-basins defined in this study (modified from Terruzi et al., 2011): Alb: Alboran Sea, NWM: North-Western Mediterranean, SWM: South-Western Mediterranean, Tyr: Tyrrhenian Sea, Adr: Adriatic Sea, Ion: Ionian Sea, Aeg: Aegean Sea, Lev: Levantine Sea.
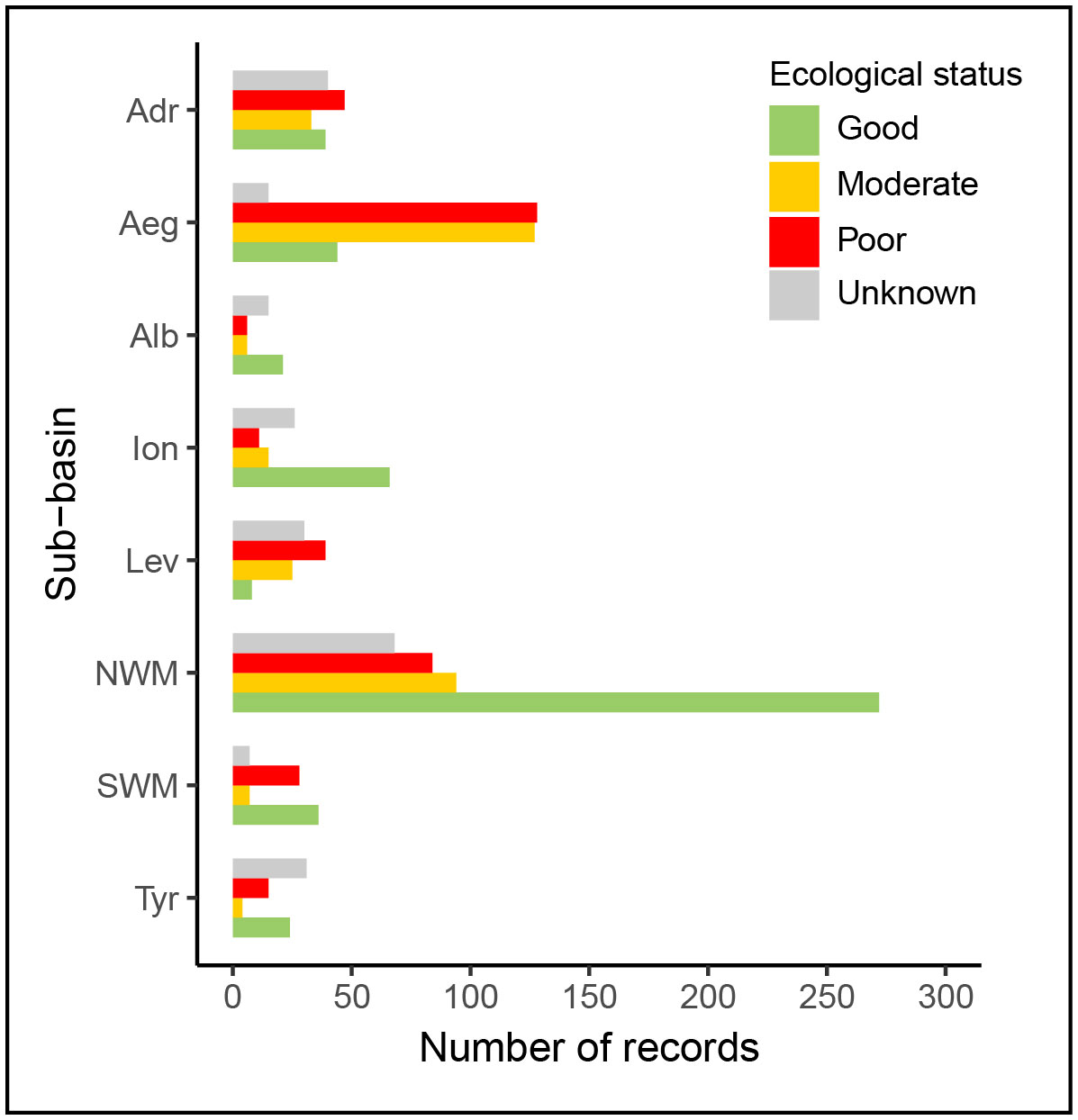
Figure 4 Number of points reporting the past or present presence of Cystoseira s.l. forest in the different sub-basins classified within the different categories of ecological status (Good, Moderate and Poor), based on the extracted data. Those points reporting the presence of Cystoseira s.l. forest but not mentioning the ecological status of the forest are classified within “Unknown” category in the graph. Alb: Alboran Sea, NWM: North-Western Mediterranean, SWM: South-Western Mediterranean, Tyr: Tyrrhenian Sea, Adr: Adriatic Sea, Ion: Ionian Sea, Aeg: Aegean Sea, Lev: Levantine Sea.
2.3.3 Stressor occurrence and expected trends
We identified 590 points reporting information about stressors (or their absence) impacting Cystoseira s.l. populations. It is worth noting that one point could be impacted by more than one stressor category. To study the density of each stressor at a Mediterranean scale, we conducted a Kernel density estimation of each stressor. Kernel density estimation is a technique used to estimate the probability density function of a set of data points (Silverman, 1998). It fits a smoothly curved surface over each point, where the surface value is the highest at the point location and decreases moving away from it until reaching the Search radius (r). The search radius defines the area around each point considered statistically significant for the analysis.
The density at a given location (x,y) is calculated by adding the values of all the kernel surfaces overlaying that location. The result is a continuous surface representing the data points’ density, with higher densities represented by higher surface values (Silverman, 1998).
The analysis was conducted using the “Kernel Density” tool, included in the “Spatial Analyst Tools” package of ArcGIS Pro 3.0.0, which is based on the quartic kernel function described in Silverman, 1998 [(Silverman, 1998), p.76, equation 4.5].
The formula determining the density in a particular location (x,y) is defined as follows:
Where is the distance between point i and the location (x,y). To calculate the search radius around each point, the following formula was used:
Where SD is the standard distance, is the median distance from the mean centre, and n is the number of points.
The Standard Distance is calculated using the following formula:
Where n is the number of points, , are the coordinates for feature i, and , are the coordinates of the mean centre of the group of points.
The representation of the results was restricted to 25 km from the coastline for a better interpretation.
Finally, we illustrate the occurrence of primary stressors (those frequently reported or considered to have substantial implications) in each sub-basin and outline their expected impact trends over the next 50 years (referred to as the mid-term). To do so, we combined the information collected in the present study with expert consensus in each sub-basin. Further details on collecting and evaluating expert perceptions of stressors’ future trends can be found in Supplementary Materials S1.
3 Results
The information search, including the bibliographic search and the personal information from 2009 to 2021, resulted in 1536 geographical points reporting the occurrence of Cystoseira s.l. species. The complete list of the articles reviewed is presented in Supplementary Materials S2. Based on the number of records in the literature and personal information, our results indicate that Cystoseira s.l. forests have been increasingly studied through the years (Supplementary Materials S3). However, our data also show that available information is still scarce, characterized by geographic and taxonomic bias, with a concentration in specific areas and on a limited number of species. For most Mediterranean coasts, there are no available or updated (after 2008) data on the presence or absence of Cystoseira s.l. forests. When available, data are concentrated especially in the Catalan and the Balearic coasts, some regions of the French Mediterranean coast, the north-west coasts of Corsica (NWS sub-basin), the Istrian coast (northern Adriatic Sea), and the coasts of Cyclades Islands (Aegean Sea) and Malta (Ionian Sea). Some information is available in other areas, such as the central region of the Algerian coast, some parts of the Italian, Lebanese and Cyprus coasts, and the southern shores of Spain, although with fewer reports (Figure 1). On the other hand, with few exceptions, such as the Aegean Sea, the Island of Malta, and the Israeli coast, the eastern part of the basin is the most underrepresented, especially the Ionian Sea, and the southern coasts of the Levantine Sea, which present a scarcity of sampled points (Figure 1).
Of the 1536 georeferenced points, 1361 reported the presence of at least one Cytoseira s.l. species. The absence of Cystoseira s.l. species was explicitly mentioned in 175 points. Among these, 50 points specifically indicated the absence of Cystoseira s.l. spp. as a result of local extinction, while the remaining 125 data points do not provide explicit information regarding whether the absence is attributable to natural factors or population decline. Concerning the taxonomic data quality, it is worth noting that 32% of the georeferenced points reporting past or present Cystoseira s.l. presence (450 out of 1411), are not based on species-level identification but are reported simply as Cystoseira sp. or spp. The remaining points (961 out of 1411) reported the presence or absence (due to decline) of at least one Cystoseira s.l. species, with several points reporting the occurrence of more than one species. This resulted in a total of 2382 past or present occurrences. Out of 36 Cystoseira s.l. species present in the Mediterranean Sea (Oliveras-Pla and Gómez-Garreta, 1989), we obtained occurrences for 32 species. Cystoseira compressa, Ericaria amentacea, and E. crinita were the most frequently reported species (with 353, 308 and 249 records, respectively), representing 48% of the total obtained occurrences reporting species-level information (1895 records), while 86% was reached by records of only 14 species (Figure 2). Ericaria funkii, Cystoseira masoudii and C. jabukae were the species least frequently reported, each reported only in one location (Figure 2).
Of the 1411 points reporting the present or past presence of Cystoseira s.l. forests, 1179 (83%) had available information on the ecological status of the Cystoseira s.l. populations (concerning any of the variables previously described, Table 1), among which 510 (36%) were classified as in Good ecological status, 311 (22%) Moderate, and 358 (25%) Poor (Figures 3, 4). Generally, forests in different ecological status categories have been reported in all sub-basins. However, in some sub-basins, such as the Adriatic Sea, the Aegean Sea and the Levantine Sea, the number of forests in poor ecological status predominates (Figure 4). Conversely, in the NWM, the sub-basin containing the highest number of occurrences, forests considered good ecological status are the most reported (Figure 4). For a more comprehensive view with a higher resolution of occurrences reporting ecological information and the ecological value assigned to each individual point, please refer to Supplementary Materials S4.
Information regarding stressors was the least reported in articles or personal communications. Only 590 (38%) of the occurrences (1536) provided specific information on the presence and/or the type of stressors affecting the population, while in only 5% (76) of the cases Cystoseira s.l. populations were recognized as not being threatened by any stressor. Multiple stressors were identified and grouped into different categories (see Table 2). Impacts related to water quality and habitat loss were the most reported, followed by overgrazing by sea urchins and overgrazing by invasive fishes (i.e., Siganus spp.). Despite increasing in recent years, impacts associated with climate change were reported less if compared to the aforementioned stressors, as well as other impacts, such as overgrazing by native fishes (e.g., Sarpa salpa), species competition, strong winds, and mechanical damage.
Certain spatial patterns can be observed when looking at the Kernel Density Estimations (KDE) of the reported stressors (Figure 5). Some stressors, such as water quality and habitat loss, have low-density values and are widespread along the Mediterranean coasts and follow a fairly comparable distribution: their highest occurrence was detected in the western part of the Mediterranean Sea (SWM and NWM sub-basins), but some cases were also detected at the easternmost part of the Mediterranean Sea, and in the western part of the Aegean sub-basin in the case of water quality. Despite being also somewhat scattered, the heat map indicated a higher concentration of reports on overgrazing by sea urchins in some regions of the NWM, southeastern Adriatic Sea, and the southern parts of the Aegean Sea. Finally, stressors related to climate change and overgrazing by invasive species showed a strong zonation pattern, especially the latter, which was restricted to a small area of the Cyclades Islands (Aegean Sea) and the northern Israeli coast. KDE values of overgrazing are not necessarily representative of the true extent of grazing pressure and are associated with the high availability of data in the Cyclades Islands (Sala et al., 2011; Nikolaou et al., 2023). KDE values show that climate change has higher density values in the Levantine part of the Mediterranean Sea, but also in the northern part of the Adriatic Sea, coinciding with the northernmost and coldest part of the Mediterranean Sea (Bianchi and Morri, 2000) and of the distribution of Cystoseira s.l. species. It is imperative to emphasize that in Figure 5, the areas depicted as lacking a specific stressor do not necessarily indicate its absence, as this outcome may also be attributed to the lack of documentation of the said stressor.
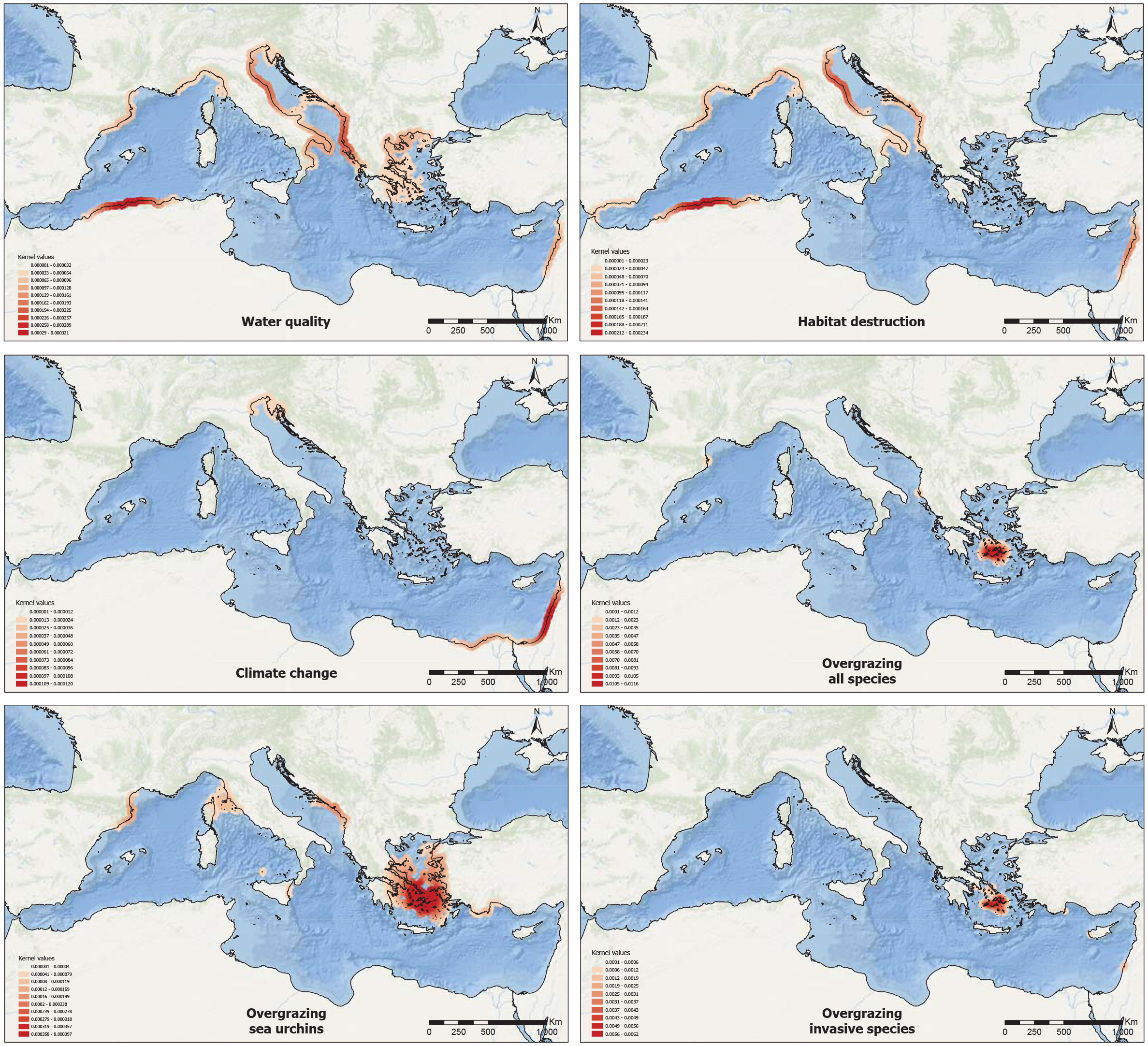
Figure 5 Stressors. Kernel density map of the main reported stressors of Cystoseira s.l. population along the Mediterranean Sea, based on the available data. The red colour on each map indicates the highest density associated with the areas with the highest reporting frequency and intensity of each stressor, from left to right and from top to bottom: Water quality, Habitat destruction, Climate change, Overgrazing by all species, Overgrazing by sea urchins and Overgrazing by invasive fish species. It is important to note that areas depicted as lacking a specific stressor do not necessarily indicate its absence, as this outcome may also be attributed to the lack of documentation of the said stressor.
In terms of stressor presence and their expected impacts, none of the sub-basins is expected to witness an amelioration in any of the considered stressors, except for overgrazing by Sarpa salpa in the Aegean sub-basin, likely due to the out-competence with invasive fish species (Figure 6). Furthermore, there is a prevailing perception that the effects of global stressors will intensify across the entire Mediterranean Sea, with particular emphasis on the easternmost basins (Figure 6).
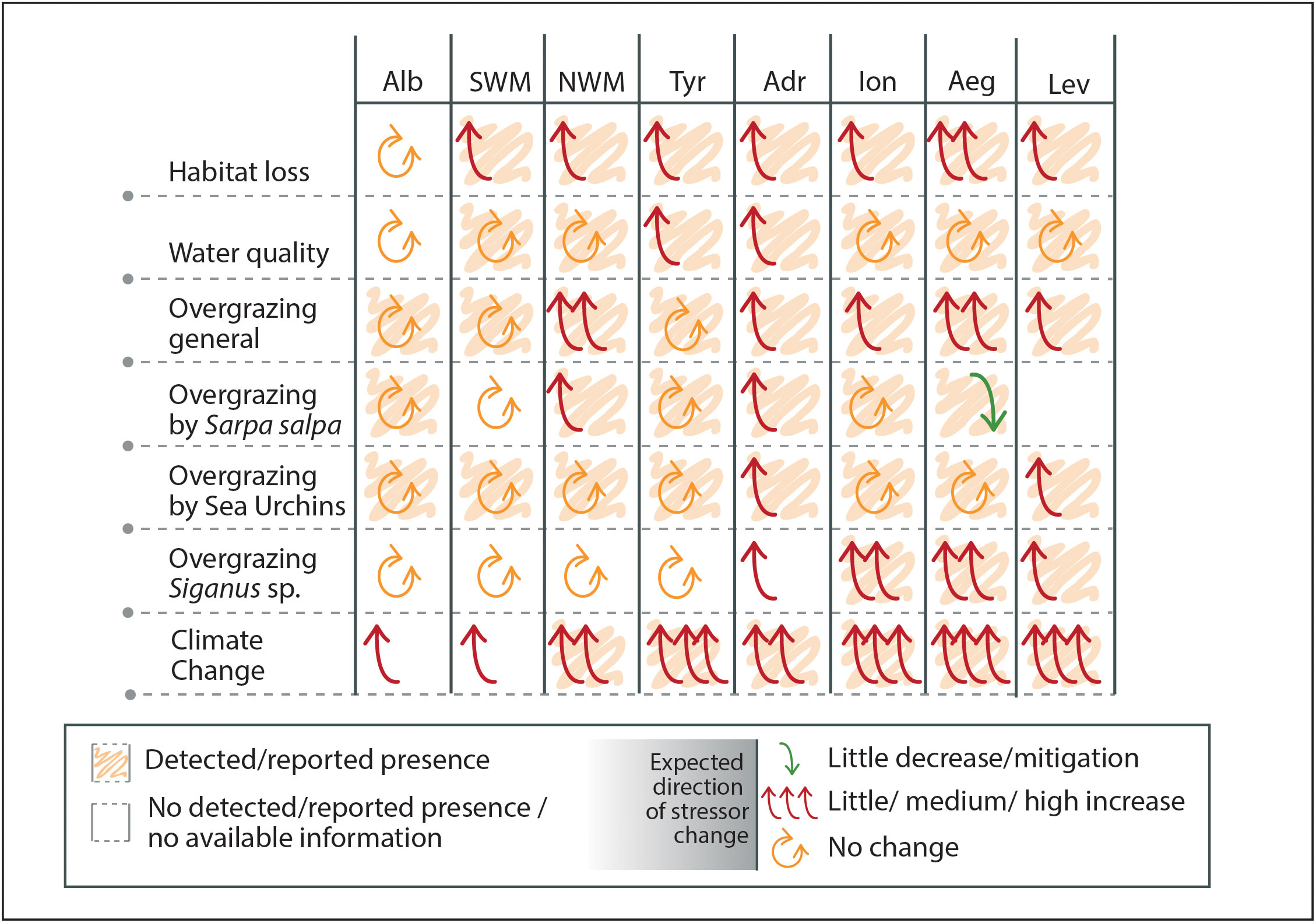
Figure 6 Presence of the reported stressors per sub-basin and expected direction of change based on the extracted data and expert judgment. Alb: Alboran Sea, NWM: North-Western Mediterranean, SWM: South-Western Mediterranean, Tyr: Tyrrhenian Sea, Adr: Adriatic Sea, Ion: Ionian Sea, Aeg: Aegean Sea, Lev: Levantine Sea.
4 Discussion
4.1 The need for cross-basin monitoring efforts
This study highlighted the challenge of establishing reliable baselines of the present distribution, ecological status, and stressors currently threatening Cystoseira s.l. forests in the Mediterranean Sea. Although essential for any regional conservation or restoration action, this information is generally scarce. Our results indicated that knowledge is highly limited, spatially skewed and biased to a few species. Although certain regions have received more attention in terms of research effort [e.g. Italy (Porzio et al., 2020; Tamburello et al., 2022; Rendina et al., 2023), Catalonia (Mariani et al., 2019) and the Mediterranean French coast (Thibaut et al., 2014; Thibaut et al., 2015; Blanfuné et al., 2016a), respective information and long-term monitoring for much of the remaining Mediterranean coastline is lacking (Rilov et al., 2020). Indeed, most available information mainly relies on surveys that provide scattered spatial data without information on the population area/extension (e.g., X, Y coordinates of sampling points; Giakoumi et al., 2012). Frequently, the reported data points exhibited a low spatial resolution. For instance, sampling sites were often represented as dots on image maps of the study area. Furthermore, these data points were frequently derived from studies with objectives unrelated to the distribution of Cystoseira s.l. forests (e.g., Sales et al., 2012).
Additionally, regardless of the overall diversity of Cystoseira s.l. species found in the Mediterranean Sea, a significant portion of the existing information focuses on a few broadly distributed, shallow-water species that are more easily accessible to researchers, such as C. compressa, E. crinita, and E. amentacea (Sales and Ballesteros, 2010; Thibaut et al., 2014; Blanfuné et al., 2016a). A large part of the available information concerning shallow Cystoseira s.l. species, such as the case of E. amentacea, is frequently derived from the widespread implementation of the CARLIT Index throughout the Mediterranean Sea (e.g., Asnaghi et al., 2009; Bermejo et al., 2013; Nikolic et al., 2013; Torras et al., 2016; Blanfuné et al., 2016b; Badreddine et al., 2018; De La Fuente et al., 2018). The CARLIT method uses the cartography of littoral upper-sublittoral macroalgal-dominated communities along rocky shores to assess the ecological status of coastal waters (Ballesteros et al., 2007). Nevertheless, these studies primarily report the obtained ecological status at the surveyed locations, while the detailed data encompassing information on macroalgal communities (e.g., occurrence and abundance) is often not included in the findings (Asnaghi et al., 2009; Nikolic et al., 2013). The creation or the use of existing databases (e.g. EMODnet) should be considered to consolidate data obtained through the widespread application and standardized methodology of this approach. At the same time, public repositories (eventually centralized) would facilitate data sharing, collaboration, and transparency among researchers. By making the database accessible to the public or upon request, it would serve as a valuable resource for the scientific community, policymakers, and stakeholders involved in research, restoration, conservation, and decision-making processes.
Information regarding the ecological status of the targeted populations was obtained from most of the collected occurrences. Overall, we found great variability in the ecological status of the populations within all sub-basins, suggesting that changes or alterations in the ecological status of analyzed populations are mainly attributed to local stressors rather than processes acting at the Mediterranean basin scale. However, these data must be considered carefully since the collected information is sourced from studies with vastly differing objectives, sampling periods, and ecological variables (e.g., density, cover, or descriptive variables), making comparisons difficult. It is also crucial to bear in mind that the ecological status map (Figure 3) does not depict the current ecological status of Cystoseira s.l. forests in the Mediterranean Sea, but instead reflects the restricted scope of knowledge currently made accessible.
4.2 Threats to Cystoseira s.l. Mediterranean forests
Overall, the lack of baseline knowledge on the distribution, health and threats of Cystoseira s.l. forests hinders our ability to detect present or future changes in the ecological status of the populations, such as population decline or recovery. It also makes it challenging to identify the stressors leading to these alterations. This raises the need to develop coordinated and standardized methods, user-friendly, to evaluate and compare these habitats’ ecological status through regions and time. In this line, the limited availability of mid/long-term monitoring studies may account for the fact that only a third of the existing research provides information on stressors affecting Cystoseira s.l. forests. Moreover, even among the studies that do address this aspect, the causal relationship between the identified factors and population declines is generally lacking. As a result, despite efforts to experimentally understand the direct effects of different stressors on Cystoseira s.l. species and life stages (Irving et al., 2009; Sala et al., 2011; de Caralt et al., 2020; Monserrat et al., 2022), in general, cause-effects relationships behind population declines remain unknown, unproven, or poorly understood, with interactive and/or cumulative effects, probably being the most likely drivers of population loss (Tamburello et al., 2022; Cebrian et al., 2021).
Among the available information, as it has been historically mentioned, habitat destruction (e.g., Mangialajo et al., 2008; Rindi et al., 2020), changes in water quality such as sedimentation or pollution (e.g., Pinedo et al., 2015), and overgrazing by native species (e.g., Sala et al., 1998; Vergés et al., 2009; Giakoumi et al., 2012; Gianni et al., 2017), are the most frequently attributed stressors to the deforestation of the Mediterranean Sea (Gros, 1978; Munda, 1982; Thibaut et al., 2005). Such stressors, driven mainly by the lack of optimal local management practices, can result in local-level effects. Nevertheless, as they have been extensively documented across the entire Mediterranean basin, they pose a significant challenge to the basin-wide conservation of these habitats. More recently, overgrazing by invasive species (e.g., Sala et al., 2011; Giakoumi, 2014; Vergés et al., 2014; Nikolaou et al., 2023) has been added to this list, along with the direct effects of climate-driven impacts (e.g., marine heatwaves; Verdura et al., 2021; Mulas et al., 2022). Specifically, the effects of invasive alien species are concentrated in the eastern Mediterranean Sea, where Cystoseira s.l. population declines due to overgrazing by the Lessepsian fishes Siganus luridus and Siganus rivulatus are pronounced. Siganus spp. are thermophilous species; thus, their distribution is restricted by temperature (Ragkousis et al., 2023). In contrast to the extensively documented mass mortality events related to marine heatwaves for benthic invertebrates (Garrabou et al., 2022), reports of temperature-induced collapses of Cystoseira s.l. forests are relatively scarce and primarily reported in the easternmost and warmest regions of the Mediterranean basin, although some isolated cases have also been reported in the northern Adriatic Sea and the Gulf of Lions (NWM) (Verdura et al., 2021; Iveša unpublished obs.). To make predictions on the vulnerability of Cystoseira s.l. species to further ocean warming (and marine heatwaves), experimental work is needed, but so far, this has been extremely scarce (but see Capdevila et al., 2019; Verdura et al., 2021; Monserrat et al., 2022). Recent experimental work did, however, show the high thermal vulnerability of Gongolaria rayssiae, a species endemic only to the coasts of Lebanon and Israel, the hottest and fastest-warming part of the Mediterranean, where this species may become (globally) extinct under business-as-usual climate change scenarios (Mulas et al., 2022).
Although global stressors, such as ocean warming, have received less attention, their interaction with pre-existing local stressors is expected to have far-reaching consequences (Strain et al., 2014). In this context, the effects of marine heatwaves on already locally threatened populations have been identified as a critical driver for Cystoseira s.l. forest collapse, even in the coldest locations of Cystoseira s.l. species distribution (e.g., Verdura et al., 2021; Iveša unpublished obs.). These results are consistent with other studies that suggest global stressors on temperate macroalgal forests can be modulated by other factors or processes at the local scale (Bennett et al., 2015; Krumhansl et al., 2016). Therefore, given the current context of climate change (Lejeusne et al., 2010), studies examining the combined effects of local and global stressors on the resistance and resilience of these species are crucial for preserving these habitats (Gissi et al., 2021; Monserrat et al., 2022).
Based on the compiled expert opinions (Figure 6), there is a prevailing consensus that locally manageable stressors are likely to maintain their current impact levels or experience marginal increases in the near future. This trend is exemplified by the case of water quality, which has witnessed significant improvements in many regions over the past few decades (Soltan et al., 2001; Pinedo et al., 2013; Iveša et al., 2016). Concerning habitat destruction, although coastal urbanization is expected to persist, environmental considerations are foreseen to accompany this process, resulting in a slight increase in its impact. In line with numerous scientific studies (Lejeusne et al., 2010) there is also a consensus that climate change will intensify its effects throughout the entire Mediterranean Sea, particularly in the easternmost basins. The perception of herbivory patterns among the asked experts is not consistently uniform. In certain sub-basins, a slight or medium increase is anticipated, possibly attributed to an increase in herbivore abundance (Gianni et al., 2017) or alterations in herbivory-plant interaction influenced by temperature increase (Mannino et al., 2016; Buñuel et al., 2021). However, no change or even a reduction in herbivory is expected in other sub-basins. Further research should be conducted to understand better the impact of overgrazing on Cystoseira s.l. forests, particularly by Salemas. Moreover, the impact of invasive fish species, such as Siganus spp., is projected to increase in sub-basins where they are already present. In the Adriatic region, where these species are currently uncommon, there is a growing perception that they may become more prevalent in the near future. However, in the western basins, these species are not expected to arrive within the given timeframe. Nevertheless, niche modelling studies provide evidence that Siganus spp. will likely spread across the Mediterranean Sea by the end of the century under different climatic scenarios (Schickele et al., 2021). Therefore, lessons learned from their impacts in the eastern basin should be considered before this almost certain invasion occurs in the west.
The limited available knowledge on Cystoseira s.l. forests could be attributed to several contributing factors. On one hand, despite international agreements, such as the Barcelona Convention (UNEP/MAP, 2013) and European legislation such as the European Union’s Habitats Directive Habitats Directive 1992 (92/43/E; Habitats Directive 1992) and the Marine Strategy Framework Directive (MSFD, 2009), encouraging the mapping and monitoring of priority habitats, Cystoseira s.l. forests were not identified as priority habitats by the Habitat Directive (Verlaque et al., 2019). This fact means that Mediterranean canopy-forming macroalgae (as well as kelp beds in temperate oceanic coasts) are not distinguished from other marine habitats, such as erect macroalgae, turfs or barren grounds, and are all included in the same habitat type: Reefs (Habitat Type 1170). As a result, while existing cartographies are helpful for mapping EU-defined macrophyte-dominated habitats, such as Posidonia oceanica meadows [Habitat type 1120: P. oceanica beds – Posidonion oceanicae; (Telesca et al., 2015)], they do not provide accurate information, nor do they offer high-resolution maps about other habitats, such as Cystoseira s.l. forests. On the other hand, difficulties in taxonomic identification within the Cystoseira s.l. complex (Neiva et al., 2022) may also contribute to the lack of spatial and ecological-related data on these habitats.
By considering both the previously published habitat suitability map for Cystoseira s.l. species (Fabbrizzi et al., 2020) and the updated map of Cystoseira s.l. forest distribution that we present in this study (Figure 1), it becomes apparent that there is a compelling need to explore and investigate understudied areas within the basin in the future. Excluding areas identified as unsuitable for the growth of these forests, such as the Gulf of Lion, Gulf of Venice, and Gulf of Gabes (Fabbrizzi et al., 2020), numerous regions of almost all sub-basins necessitate more comprehensive surveys. Remarkably, there is a pressing need to prioritize thorough investigations in the south Ionian and Levantine sub-basins. Additionally, attention should be given to other areas, including the Eastern Adriatic Sea’s central part, Italy’s western coasts, and Corsica and Sardinia’s eastern coasts. Moreover, significant efforts should be dedicated to mapping and surveying deep-water Cystoseira s.l. species across the Mediterranean basin, as limited information about these species exists (but see, Ballesteros, 1990; Ballesteros et al., 1998; Hereu et al., 2008; Ballesteros et al., 2009; Capdevila et al., 2015).
Only by gathering this knowledge together with an accurate understanding of the stressors currently threatening these populations we will be able to establish ecosystem-based management of marine forests, in which mitigation of both local and global stressors, such as water quality and herbivory pressure, should be a critical step of the process. Appropriate management and threats mitigation are pre-requisite in any conservation and restoration plan, and we can draw inspiration from the few examples of naturally recovered Cystoseira s.l. populations after reduced eutrophication or pollution levels in seawater (Iveša et al., 2016; Ricart et al., 2018) and herbivory (Guarnieri et al., 2020). Climate change already affects some populations and represents a challenging factor that cannot be easily mitigated (Bevilacqua et al., 2019). However, promoting the resilience of healthy Cystoseira s.l. populations, more tolerant species and populations, localizing potential areas that can serve as climate change refugia for thermally sensitive species or determining which areas are expected to be more severely impacted by human activities, are among the leading research challenges in conserving macroalgal forests.
There is a need to allocate efforts and resources to accurately collect distributional, ecological and environmental data for mapping and monitoring Cystoseira s.l. populations and assessing their ecological status. Acquiring this knowledge is crucial for achieving the EU’s Biodiversity Strategy 2030 goals, aiming to protect and restore biodiversity and enhance ecosystem resilience to climate change. As previously highlighted (Verlaque et al., 2019; Tamburello et al., 2022), it is urgent to prioritize Cystoseira s.l. forests as one of the main objectives of the protection and management of the Mediterranean marine environment. We also recommend reconsidering the inclusion of Cystoseira s.l. species in international and European agreements and conventions. Legal protection measures would be critical for implementing conservation and restoration strategies for these canopy-forming macroalgae and their associated marine habitats.
Author contributions
JV: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. LR: Data curation, Formal Analysis, Methodology, Software, Writing – original draft, Writing – review & editing. LM: Funding acquisition, Investigation, Writing – review & editing. SF: Investigation, Writing – review & editing. ZB: Investigation, Writing – review & editing. SB: Investigation, Writing – review & editing. AB: Investigation, Writing – review & editing. BS: Writing – review & editing. AC: Investigation, Writing – review & editing. RD: Funding acquisition, Investigation, Writing – review & editing. EF: Investigation, Writing – review & editing. SG: Investigation, Writing – review & editing. LI: Investigation, Writing – review & editing. SK: Investigation, Writing – review & editing. EK: Investigation, Writing – review & editing. IN: Investigation, Writing – review & editing. AN: Investigation, Writing – review & editing. SO: Investigation, Writing – review & editing. GR: Investigation, Writing – review & editing. FR: Investigation, Writing – review & editing. MSa: Investigation, Writing – review & editing. MSi: Investigation, Writing – review & editing. LT: Investigation, Writing – review & editing. TT: Investigation, Writing – review & editing. KT: Investigation, Writing – review & editing. EC: Conceptualization, Funding acquisition, Investigation, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work is supported by the AFRIMED EU project funded by the Executive Agency for Small and Medium Enterprises (EASME) and European Maritime and Fisheries Fund (EMFF) under grant agreement No. EASME/EMFF/2017/1.2.1.12/S4/01/SI2.789059; by the FoRestA project from the Spanish Ministry of Science and Innovation (Grant/Award No. PID2020-112985GB-I00); and by Biodiversa+ (FORESCUE project), the European Biodiversity Partnership under the 2021-2022 BiodivProtect joint call for research proposals, co-funded by the European Commission (GA N°101052342) and with the funding organisations, Agencia Estatal de Investigación AEI/10.13039/501100011033/NextGenerationEU/PRTR (PCI2022-135070-2, PCI2022-135052-2) and the French Agence Nationale de la Recherche. This research was also funded by the European Union – NextGeneration EU – as part of the MITECO program for the Spanish Recovery, Transformation and Resilience Plan (Recovery and Resilience Facility of the European Union established by the Regulation (EU) 2020/2094), and was entrusted to CSIC, AZTI, SOCIB, and the universities of Vigo and Cadiz. This work has also been supported by the French government, through the UCAJEDI lnvestments in the Future project managed by the Nationall Research Agency (ANR) with the reference number ANR-15-lDEX-01).
Acknowledgments
The authors would thank Enric Ballesteros, Miriam Benali, Lamia Bahbah, Soultana Tsioli and Vasilis Papathanasiou for personal communications and data clarification. This is a contribution from the Consolidated Research Group “Medrecover; www.medrecover.org“, SGR2017-1521 (Catalan Government).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
The reviewer SC declared a shared affiliation with the author IN to the handling editor at the time of review.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2023.1258842/full#supplementary-material
Supplementary Materials S1 | Procedure and set of questions developed to construct, which captures the experts’ perceptions of different stressors in each sub-basin.
Supplementary Materials S2 | List of the reviewed papers.
Supplementary Materials S3 | Number of scientific papers reporting spatial data on Cystoseira s. l. species per year.
Supplementary Materials S4 | Ecological status. Detailed maps of the distribution of the sampling points reporting information regarding the ecological status of Cystoseira s.l. populations along the Mediterranean basin that have been obtained in the data extraction process.
References
Araújo R. M., Assis J., Aguillar R., Airoldi L., Bárbara I., Bartsch I., et al. (2016). Status, trends and drivers of kelp forests in Europe: an expert assessment. Biodivers. Conserv. 25, 1319–1348. doi: 10.1007/s10531-016-1141-7
Asnaghi V., Chiantore M., Bertolotto R. M., Parravicini V., Cattaneo-Vietti R., Gaino F., et al. (2009). Implementation of the European Water Framework Directive: Natural variability associated with the CARLIT method on the rocky shores of the Ligurian Sea (Italy). Mar. Ecol. 30, 505–513. doi: 10.1111/j.1439-0485.2009.00346.x
Badreddine A., Abboud-Abi Saab M., Gianni F., Ballesteros E., Mangialajo L. (2018). First assessment of the ecological status in the Levant Basin: Application of the CARLIT index along the Lebanese coastline. Ecol. Indic. 85, 37–47. doi: 10.1016/j.ecolind.2017.10.006
Ballesteros E. (1990). Structure and dynamics of the assemblage of Cystoseira zosteroides (Turner) C. Agardh (Fucales, Phaeophy- ceae) in the northwestern Mediterranean. Scientia Marina 54, 217–229.
Ballesteros E., Garrabou J., Hereu B., Zabala M., Cebrian E., Sala E. (2009). Deepwater stands of Cystoseira zosteroides C. Agardh (Fucales, Ochrophyta) in the Northwestern Mediterranean: insights into assemblage structure and population dynamics. Estuar. Coast. Shelf Sci. 82, 477–484. doi: 10.1016/j.ecss.2009.02.013
Ballesteros E., Sala E., Garrabou J., Zabala M. (1998). Community structure and frond size distribution of a deep water stand of Cystoseira spinosa (Phaeophyta) in the Northwestern Mediterranean. Eur. J. Phycol. 33 (2), 121–128. doi: 10.1080/09670269810001736613
Ballesteros E., Torras X., Pinedo S., García M., Mangialajo L., de Torres M. (2007). A new methodology based on littoral community cartography dominated by macroalgae for the implementation of the European Water Framework Directive. Mar. pollut. Bull. 55, 172–180. doi: 10.1016/j.marpolbul.2006.08.038
Bayraktarov E., Saunders M. I., Abdullah S., Mills M., Beher J., Possingham H. P., et al. (2016). The cost and feasibility of marine coastal restoration. Ecol. Appl. Ecol. Appl. 26, 1055–1074. doi: 10.5061/dryad.rc0jn
Bennett S., Wernberg T., Joy B. A., De Bettignies T., Campbell A. H. (2015). Central and rear-edge populations can be equally vulnerable to warming. Nat. Commun. 6, 10280. doi: 10.1038/ncomms10280
Bermejo R., de la Fuente G., Vergara J. J., Hernández I. (2013). Application of the CARLIT index along a biogeographical gradient in the Alboran Sea (European Coast). Mar. pollut. Bull. 72, 107–118. doi: 10.1016/j.marpolbul.2013.04.011
Bevilacqua S., Savonitto G., Lipizer M., Mancuso P., Ciriaco S., Srijemsi M., et al. (2019). Climatic anomalies may create a long-lasting ecological phase shift by altering the reproduction of a foundation species. Ecology 100, e02838. doi: 10.1002/ecy.2838
Bianchi C. N., Corsini-Foka M., Morri C., Zenetos A. (2014). Thirty years after; dramatic change in the coastal marine habitats of Kos Island (Greece), 1981-2013. Mediterr. Mar. Sci. 14, 482–497. doi: 10.12681/mms.678
Bianchi C. N., Morri C. (2000). Marine biodiversity of the Mediterranean Sea: Situation, problems and prospects for future research. Mar. pollut. Bull. 40, 367–376. doi: 10.1016/S0025-326X(00)00027-8
Blanfuné A., Boudouresque C. F., Verlaque M., Beqiraj S., Kashta L., Nasto I., et al. (2016b). Response of rocky shore communities to anthropogenic pressures in Albania (Mediterranean Sea): Ecological status assessment through the CARLIT method. Mar. pollut. Bull. 109, 409–418. doi: 10.1016/j.marpolbul.2016.05.041
Blanfuné A., Boudouresque C. F., Verlaque M., Thibaut T. (2016a). The fate of Cystoseira crinita, a forest-forming Fucale (Phaeophyceae, Stramenopiles), in France (North Western Mediterranean Sea). Estuar. Coast. Shelf Sci. 181, 196–208. doi: 10.1016/j.ecss.2016.08.049
Blanfuné A., Boudouresque C. F., Verlaque M., Thibaut T. (2019). The ups and downs of a canopy-forming seaweed over a span of more than one century. Sci. Rep. 9, 5250. doi: 10.1038/s41598-019-41676-2
Buñuel X., Alcoverro T., Romero J., Arthur R., Ruiz J. M., Pérez M., et al. (2021). Warming intensifies the interaction between the temperate seagrass Posidonia oceanica and its dominant fish herbivore Sarpa salpa. Mar. Environ. Res. 165, 105237. doi: 10.1016/j.marenvres.2020.105237
Capdevila P., Hereu B., Salguero-Gómez R., Rovira G. L., Medrano A., Cebrian E., et al. (2019). Warming impacts on early life stages increase the vulnerability and delay the population recovery of a long-lived habitat-forming macroalga. J. Ecol. 107 (3), 1129–1140. doi: 10.1111/1365-2745.13090
Capdevila P., Linares C., Aspillaga E., Navarro L., Kersting D. K., Hereu B. (2015). Recruitment patterns in the Mediterranean deep-water alga Cystoseira zosteroides. Mar. Biol. 162, 1165–1174. doi: 10.1007/s00227-015-2658-0
Cebrian E., Tamburello L., Verdura J., Guarnieri G., Medrano A., Linares C., et al. (2021). A roadmap for the restoration of Mediterranean macroalgal forests. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.709219
Celis-Plá P. S.M., Martínez B., Korbee N., Hall-Spencer J. M., Figueroa F. L. (2017). Ecophysiological responses to elevated CO2 and temperature in Cystoseira tamariscifolia (Phaeophyceae). Climate Change 142, 97-81. doi: 10.1007/s10584-017-1943-y
Cheminée A., Sala E., Pastor J., Bodilis P., Thiriet P., Mangialajo L., et al. (2013). Nursery value of Cystoseira forests for Mediterranean rocky reef fishes. J. Exp. Mar. Biol. Ecol. 442, 70–79. doi: 10.1016/j.jembe.2013.02.003
de Caralt S., Verdura J., Vergés A., Ballesteros E., Cebrian E. (2020). Differential effects of pollution on adult and recruits of a canopy-forming alga: implications for population viability under low pollutant levels. Sci. Rep. 10, 17825. doi: 10.1038/s41598-020-73990-5
De La Fuente G., Chiantore M., Gaino F., Asnaghi V. (2018). Ecological status improvement over a decade along the Ligurian coast according to a macroalgae based index (CARLIT). PloS One 13 (12), e0206826. doi: 10.1371/journal.pone.0206826
Duarte C. M., Agusti S., Barbier E., Britten G. L., Castilla J. C., Gattuso J.-P., et al. (2020). Rebuilding marine life. Nature 580, 39–51. doi: 10.1038/s41586-020-2146-7
EC (2020). “EU Biodiversity Strategy for 2030,” in EU Biodiversity Strategy for 2030 (Brussels). Available at: https://eur-lex.europa.eu/legal-content/EN/TXT/?qid=1590574123338&uri=CELEX:52020DC0380. ed. 380 final. Nature Back into Our Lives. E. Commission.
EC (2022) Proposal for a Regulation of the European Parliament and of the Council on Nature Restoration: E. Commission (Brussels). Available at: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A52022PC0304.
Eger A. M., Marzinelli E. M., Christie H., Fagerli C. W., Fujita D., Gonzalez A. P., et al. (2022). Global kelp forest restoration: past lessons, present status, and future directions. Biol. Rev. 97 (4), 1449–1475. doi: 10.1111/brv.12850
Fabbrizzi E., Giakoumi S., De Leo F., Tamburello L., Chiarore A., Colletti A., et al. (2023). The challenge of setting restoration targets for macroalgal forests under climate changes. J. Environ. Manage. 326, 116834. doi: 10.1016/j.jenvman.2022.116834
Fabbrizzi E., Scardi M., Ballesteros E., Benedetti-Cecchi L., Emma C., Ceccherelli G., et al. (2020). Modeling macroalgal forest distribution at Mediterranean scale: Present status, drivers of changes and insights for conservation and management. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00020
Filbee-Dexter K., Wernberg T. (2018). Rise of turfs: A new battlefront for globally declining kelp forests. BioScience 68, 64–76. doi: 10.1093/biosci/bix147
Fraschetti S., McOwen C., Papa L., Papadopoulou N., Bilan M., Boström C., et al. (2021). Where is more important than how in coastal and marine ecosystems restoration. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.626843
Galobart C., Ballesteros E., Golo R., Cebrian E. (2023). Addressing marine restoration success: evidence of species and functional diversity recovery in a ten-year restored macroalgal forest. Front. Mar. Sci. 10. doi: 10.3389/fmars.2023.1176655
Gann G. D., McDonald T., Aronson J., Nelson C. R., Jonson J., Hallett J. G., et al. (2019). International principles and standards for the practice of ecological restoration (Summary). Second edition: September 2019 (Washington D.C. 20005 U.S.A.: Society for Ecological Restoration). doi: 10.1111/rec.13035
Garrabou J., Gómez- D., Medrano A., Cerrano C., Ponti M., Schlegel R., et al. (2022). Marine heatwaves drive recurrent mass mortalities in the Mediterranean Sea. Glob. Change Biol. 28, 5708–5725. doi: 10.1111/gcb.16301
Giaccone G., Bruni A. (1973). Le Cystoseire e la vegetazione sommersa del Mediterraneo. Atti Istituto Veneto Scienze Lettere ed Arti 131, 59–103.
Giakoumi S. (2014). Distribution patterns of the invasive herbivore Siganus luridus (Rüppell 1829) and its relation to native benthic communities in the central Aegean Sea, Northeastern Mediterranean. Mar. Ecol. 35, 96–105. doi: 10.1111/maec.12059
Giakoumi S., Cebrian E., Kokkoris G. D., Ballesteros E., Sala E. (2012). Relationships between fish, sea urchins and macroalgae: The structure of shallow rocky sublittoral communities in the Cyclades, Eastern Mediterranean. Estuar. Coast. Shelf Sci. 109, 1–10. doi: 10.1016/j.ecss.2011.06.004
Gianni F., Bartolini F., Pey A., Laurent M., Martins G. M., Airoldi L., et al. (2017). Threats to large brown algal forests in temperate seas: The overlooked role of native herbivorous fish. Sci. Rep. 7, 6012. doi: 10.1038/s41598-017-06394-7
Gissi E., Manea E., Mazaris A. D., Fraschetti S., Almpanidou V., Bevilacqua S., et al. (2021). A review of the combined effects of climate change and other local human stressors on the marine environment. Sci. Total Environ. 755, 142564. doi: 10.1016/j.scitotenv.2020.142564
Gros C. (1978). Le genre Cystoseira sur la côte des Albères. Répartition, écologie, morphogénèse PhD thesis., Paris: Université Pierre et Marie Curie.
Guarnieri G., Bevilacqua S., Figueras N., Tamburello L., Fraschetti S. (2020). Large-scale sea urchin culling drives the reduction of subtidal barren grounds in the Mediterranean Sea. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00519
Habitats Directive (1992). Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Union L206, 7–50.
Hereu B., Mangialajo L., Ballesteros E., Thibaut T. (2008). On the occurrence, structure and distribution of deep-water Cystoseira populations in the Port-Cros National Park (North-western Med- iterranean). Eur. J. Phycol. 43, 263–273. doi: 10.1080/09670260801930330
Irving A. D., Balata D., Colosio F., Ferrando G. A., Airoldi L. (2009). Light, sediment, temperature, and the early life-history of the habitat-forming alga Cystoseira barbata. Mar. Biol. 156, 1223–1231. doi: 10.1007/s00227-009-1164-7
Iveša L., Devescovi M. (2004). Distribution and composition of Cystoseira stands along the west Istrian coast (northern Adriatic, Croatia) and comparison with historical data. in 5th Mediterranean Symposium on Marine Vegetation.
Iveša L., Djakovac T., Devescovi M. (2016). Long-term fluctuations in Cystoseira populations along the west Istrian Coast (Croatia) related to eutrophication patterns in the northern Adriatic Sea. Mar. pollut. Bull. 106, 162–173. doi: 10.1016/j.marpolbul.2016.03.010
Johnson C. R., Banks S. C., Barrett N. S., Cazassus F., Dunstan P. K., Edgar G. J., et al. (2011). Climate change cascades: Shifts in oceanography, species’ ranges and subtidal marine community dynamics in eastern Tasmania. J. Exp. Mar. Biol. Ecol. 400, 17–32. doi: 10.1016/j.jembe.2011.02.032
Krumhansl K. A., Okamoto D. K., Rassweiler A., Novak M., Bolton J. J., Cavanaugh K. C., et al. (2016). Global patterns of kelp forest change over the past half-century. Proc. Natl. Acad. Sci. 113, 13785–13790. doi: 10.1073/pnas.1606102113
Layton C., Coleman M. A., Marzinelli E. M., Steinberg P. D., Swearer S. E., Vergés A., et al. (2020). Kelp forest restoration in Australia. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00074
Lejeusne C., Chevaldonné P., Pergent-Martini C., Boudouresque C. F., Pérez T. (2010). Climate change effects on a miniature ocean: the highly diverse, highly impacted Mediterranean Sea. Trends Ecol. Evol. 25, 250–260. doi: 10.1016/j.tree.2009.10.009
Ling S. D., Scheibling R. E., Rassweiler A., Johnson C. R., Shears N., Connell S. D., et al. (2015). Global regime shift dynamics of catastrophic sea urchin overgrazing. Philos. Trans. R. Soc B 370, 20130269. doi: 10.1098/rstb.2013.0269
Mačić V., Antolić B. (2015). Distribution of rare Cystoseira species along the Montenegro coast (South-Eastern Adriatic Sea). Period. Biol. 117, 441–447. doi: 10.18054/pb.2015.117.3.2995
Mancuso F. P., Strain E. M. A., Piccioni E., De Clerck O., Sarà G., Airoldi L. (2018). Status of vulnerable Cystoseira populations along the Italian infralittoral fringe, and relationships with environmental and anthropogenic variables. Mar. pollut. Bull. 129, 762–771. doi: 10.1016/j.marpolbul.2017.10.068
Mangialajo L., Chiantore M., Cattaneo-Vietti R. (2008). Loss of fucoid algae along a gradient of urbanisation, and structure of benthic assemblages. Mar. Ecol. Prog. Ser. 358, 63–74. doi: 10.3354/meps07400
Mann K. (1973). Seaweeds: their productivity and strategy for growth. Science 224, 347–353. doi: 10.1126/science.177.4047.410
Mannino A. M., Vaglica V., Cammarata M., Oddo E. (2016). Effects of temperature on total phenolic compounds in Cystoseira amentacea (C. Agardh) Bory (Fucales, Phaeophyceae) from southern Mediterranean Sea. Plant Biosyst. 150, 152–160. doi: 10.1080/11263504.2014.941033
Mariani S., Cefalì M. E., Chappuis E., Terradas M., Pinedo S., Torras X., et al. (2019). Past and present of Fucales from shallow and sheltered shores in Catalonia. Reg. Stud. Mar. Sci. 32, 100824. doi: 10.1016/j.rsma.2019.100824
McDonald T., Gann G., Jonson J., Dixon K. (2016). International standards for the practice of ecological restoration – including principles and key concepts. Soc. Ecol. Restor. doi: 10.2139/ssrn.3645563
Molinari-Novoa E., Guiry M. (2020). Reinstatement of the genera gongolaria boehmer and ericaria stackhouse (Sargassaceae, phaeophyceae). Notulae Algarum 172, 1–10.
Monserrat M., Comeau S., Verdura J., Alliouane S., Spennato G., Priouzeau F., et al. (2022). Climate change and species facilitation affect the recruitment of macroalgal marine forests. Sci. Rep. 12, 1–12. doi: 10.1038/s41598-022-22845-2
Mulas M., Silverman J., Guy-Haim T., Noè S., Rilov G. (2022). Thermal vulnerability of the Levantine endemic and endangered habitat-forming macroalga, Gongolaria rayssiae: implications for reef carbon. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.862332
Munda I. M. (1982). The effects of organic pollution on the distribution of fucoid algae from the Istrian coast (vicinity of Rovinj). Acta Adriat. 23, 329–337.
Munda I. M. (1993). Changes and degradation of seaweed stands in the Northern Adriatic. Hydrobiologia 260–261, 239–253. doi: 10.1007/BF00049025
Neiva J., Bermejo R., Medrano A., Capdevila P., Milla-Figueras D., Afonso P., et al. (2022). DNA barcoding reveals cryptic diversity, taxonomic conflicts and novel biogeographical insights in Cystoseira s.l. (Phaeophyceae). Eur. J. Phycol. 00, 1–25. doi: 10.1080/09670262.2022.2126894
Nikolaou A., Tsirintanis K., Rilov G., Katsanevakis S. (2023). Invasive fish and sea urchins drive the status of canopy forming macroalgae in the eastern Mediterranean. Biology 12, 763. doi: 10.3390/biology12060763
Nikolic V., Zuljevic A., Mangialajo L., Antolic B., Kuspilic G., Ballesteros E. (2013). Cartography of littoral rocky-shore communities (CARLIT) as a tool for ecological quality assessment of coastal waters in the Eastern Adriatic Sea. Ecol. Indic. 34, 87–93. doi: 10.1016/j.ecolind.2013.04.021
Oliveras-Pla M., Gómez-Garreta A. (1989). Corología del género Cystoseira C. Agardh (Phaeophyceae, Fucales). Anales del Jardín Botánico Madrid 46, 89–97.
Orellana S., Hernández M., Sansón M. (2019). Diversity of Cystoseira sensu lato (Fucales, Phaeophyceae) in the eastern Atlantic and Mediterranean based on morphological and DNA evidence, including Carpodesmia gen. emend. and Treptacantha gen. emend. Eur. J. Phycol. 54, 447–465. doi: 10.1080/09670262.2019.1590862
Pinedo S., Arévalo R., Ballesteros E. (2015). Seasonal dynamics of upper sublittoral assemblages on Mediterranean rocky shores along a eutrophication gradient. Estuar. Coast. Shelf Sci. 161, 93–101. doi: 10.1016/j.ecss.2015.05.004
Pinedo S., Zabala M., Ballesteros E. (2013). Long-term changes in sublittoral macroalgal assemblages related to water quality improvement. Bot. Mar. 56, 461–469. doi: 10.1515/bot-2013-0018
Porzio L., Rech D., Buia M. C. (2020). Long-term changes, (1800–2019) in marine vegetational habitats: Insights from a historic industrialised coastal area. Mar. Environ. Res. 161, 105003. doi: 10.1016/j.marenvres.2020.105003
Ragkousis M., Zenetos A., Souissi J.B., Hoffman R., Ghanem R., Taşkın E., et al. (2023). Unpublished Mediterranean and Black Sea records of marine alien, cryptogenic, and neonative species. BioInvasions Rec. 12 (2), 339–369. doi: 10.3391/bir.2023.12.2.01
R Core Team (2019). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing).
Rendina F., Falace A., Alongi G., Buia M. C., Neiva J., Appolloni L., et al. (2023). The lush fucales underwater forests off the Cilento coast: An overlooked Mediterranean biodiversity hotspot. Plants 12, 1–12. doi: 10.3390/plants12071497
Ricart A. M., García M., Weitzmann B., Linares C., Hereu B., Ballesteros E. (2018). Long-term shifts in the north western Mediterranean coastal seascape: The habitat-forming seaweed Codium vermilara. Mar. pollut. Bull. 127, 334–341. doi: 10.1016/j.marpolbul.2017.12.019
Rilov G., Peleg O., Guy-Haim T., Yeruham E. (2020). Community dynamics and ecological shifts on Mediterranean vermetid reefs. Mar. Environ. Res.Mar. Environ. Res. 160, 105045. doi: 10.1016/j.marenvres.2020.105045
Rindi F., Gavio B., Díaz-Tapia P., Di Camillo C. G., Romagnoli T. (2020). Long-term changes in the benthic macroalgal flora of a coastal area affected by urban impacts (Conero Riviera, Mediterranean Sea). Biodivers. Conserv. 29, 2275–2295. doi: 10.1007/s10531-020-01973-z
Sala E., Boudouresque C. F., Harmelin M. (1998). Fishing, trophic cascades, and the structure of algal assemblages: evaluation of and old but untested paradigm. Oikos 82, 425–439. doi: 10.2307/3546364
Sala E., Kizilkaya Z., Yildirim D., Ballesteros E. (2011). Alien marine fishes deplete algal biomass in the Eastern Mediterranean. PloS One 6, e17356. doi: 10.1371/journal.pone.0017356
Sales M., Ballesteros E. (2010). Long-term comparison of algal assemblages dominated by Cystoseira crinita (Fucales, Heterokontophyta) from Cap Corse (Corsica, North Western Mediterranean). Eur. J. Phycol. 45, 404–412. doi: 10.1080/09670262.2010.498585
Sales M., Ballesteros E., Anderson M. J., Iveša L., Cardona E. (2012). Biogeographical patterns of algal communities in the Mediterranean Sea: Cystoseira crinita-dominated assemblages as a case study. J. Biogeogr. 39, 140–152. doi: 10.1111/j.1365-2699.2011.02564.x
Schickele A., Guidetti P., Giakoumi S., Zenetos A., Francour P., Raybaud V. (2021). Improving predictions of invasive fish ranges combining functional and ecological traits with environmental suitability under climate change scenarios. Glob. Change Biol. 27, 6086–6102. doi: 10.1111/gcb.15896
Silverman B. W. (1998). Density estimation for statistics and data analysis (1st ed.) (Routledge: CRC press). doi: 10.1201/9781315140919
Smale D. A. (2020). Impacts of ocean warming on kelp forest ecosystems. New Phytol. 225, 1447–1454. doi: 10.1111/nph.16107
Smale D. A., Burrows M. T., Moore P., O’Connor N., Hawkins S. J. (2013). Threats and knowledge gaps for ecosystem services provided by kelp forests: A northeast Atlantic perspective. Ecol. Evol. 3, 4016–4038. doi: 10.1002/ece3.774
Smale D. A., Wernberg T. (2013). Extreme climatic event drives range contraction of a habitat-forming species. Proc. R. Soc B: Biol. Sci. 280, 20122829. doi: 10.1098/rspb.2012.2829
Smith C. J., Verdura J., Papadopoulou N., Fraschetti S., Cebrian E., Fabbrizzi E., et al. (2023). A decision-support framework for the restoration of Cystoseira sensu lato forests. Frontiers in Marine Science 10, 1159262.
Soltan D., Verlaque M., Boudouresque C. F., Francour P. (2001). Changes in macroalgal communities in the vicinity of a Mediterranean sewage outfall after the setting up of a treatment plant. Mar. pollut. Bull. 42, 59–70. doi: 10.1016/S0025-326X(00)00116-8
Steneck R., Graham M. H., Bourque B. J., Corbett D., Erlandson J. M. (2002). Kelp forest ecosystems: Biodiversity, stability, resilience and future. Environ. Conserv. 29, 436–459. doi: 10.1017/S0376892902000322
Strain E. M. A., Thomson R. J., Micheli F., Mancuso F. P., Airoldi L. (2014). Identifying the interacting roles of stressors in driving the global loss of canopy-forming to mat-forming algae in marine ecosystems. Glob. Change Biol. 20, 3300–3312. doi: 10.1111/gcb.12619
Tamburello L., Chiarore A., Fabbrizzi E., Colletti A., Franzitta G., Grech D., et al. (2022). Can we preserve and restore overlooked macroalgal forests? Sci. Total Environ. 806, 150855. doi: 10.1016/j.scitotenv.2021.150855
Teagle H., Hawkins S. J., Moore P. J., Smale D. A. (2017). The role of kelp species as biogenic habitat formers in coastal marine ecosystems. J. Exp. Mar. Biol. Ecol. 492, 81–98. doi: 10.1016/j.jembe.2017.01.017
Telesca L., Belluscio A., Criscoli A., Ardizzone G., Apostolaki E. T., Fraschetti S., et al. (2015). Seagrass meadows (Posidonia oceanica) distribution and trajectories of change. Sci. Rep. 5, 12505. doi: 10.1038/srep12505
Teruzzi A., Salon S., Bolzon G., Lazzari P., Campagna S., Ficarelli F., et al. (2011). Operational forecasts of the biogeochemical state of Mediterranean Sea. Mercator Ocean Q. Newslett. 40, 15–25.
Thibaut T., Blanfuné A., Boudouresque C.-F., Cottalorda J.-M., Hereu B., Susini M.-L., et al. (2016). Unexpected temporal stability of Cystoseira and Sargassum forests in Port-Cros, one of the oldest Mediterranean marine National Parks. Cryptogamie Algologie 37, 61–90. doi: 10.7872/crya/v37.iss1.2016.61
Thibaut T., Blanfune A., Boudouresque C., Verlaque M. (2015). Decline and local extinction of Fucales in French Riviera: the harbinger of future extinctions? Mediterr. Mar. Sci. 16, 206–224. doi: 10.12681/mms.1032
Thibaut T., Blanfuné A., Markovic L., Verlaque M., Boudouresque C. F., Bottin L., et al. (2014). Unexpected abundance and long-term relative stability of the brown alga Cystoseira amentacea, hitherto regarded as a threatened species, in the north-western Mediterranean Sea. Mar. pollut. Bull. 89, 305–323. doi: 10.1016/j.marpolbul.2014.09.043
Thibaut T., Pinedo S., Torras X., Ballesteros E. (2005). Long-term decline of the populations of Fucales (Cystoseira spp. and Sargassum spp.) in the Albères coast (France, North-western Mediterranean). Mar. pollut. Bull. 50, 1472–1489. doi: 10.1016/j.marpolbul.2005.06.014
Torras X., Pinedo S., García M., Weitzmann B., Ballesteros E. (2016). “Environmental quality of Catalan coastal waters based on macroalgae: the interannual variability of CARLIT index and its ability to detect changes in anthropogenic pressures over time,” in Experiences from ground, coastal and transitional water quality monitoring: The EU Water Framework Directive implementation in the Catalan river basin district (Part II): 183-199. The Handbook of Environmental Chemistry, vol. 43 . Eds. Munné A., Ginebreda A., Prat N. (Switzerland: Springer International Publishing).
UNEP/MAP (2013). “Protocol concerning specially protected areas and biological diversity in the Mediterranean,” in List of endangered species (Athina: UNEP/MAP).
Verdura J., Santamaría J., Ballesteros E., Smale D. A., Cefalì M. E., Golo R., et al. (2021). Local-scale climatic refugia offer sanctuary for a habitat-forming species during a marine heatwave. J. Ecol. 109, 1758–1773. doi: 10.1111/1365-2745.13599
Vergés A., Alcoverro T., Ballesteros E. (2009). Role of fish herbivory in structuring the vertical distribution of canopy algae Cystoseira spp. in the Mediterranean Sea. Mar. Ecol. Prog. Ser. 375, 1–11. doi: 10.3354/meps07778
Vergés A., Tomas F., Cebrian E., Ballesteros E., Kizilkaya Z., Verg A., et al. (2014). Tropical rabbit fish and the deforestation of a warming temperate sea. J.Ecol 102, 1518–1527. doi: 10.1111/1365-2745.12324
Verlaque M., Boudouresque C., Perret-Boudouresque M. (2019). Mediterranean seaweeds listed as threatened under the Barcelona Convention: A critical analysis. Sci. Rep. Port-Cros Natl. Park 33, 179–214.
Waltham N. J., Elliott M., Lee S. Y., Lovelock C., Duarte C. M., Buelow C., et al. (2020). UN decade on ecosystem restoration 2021–2030—What chance for success in restoring coastal ecosystems? Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00071
Wernberg T., Krumhansl K., Filbee-Dexter K., Pedersen M. F. (2019). “Status and Trends for the World’s Kelp Forests,” in World seas: An environmental evaluation (Amsterdam: Elsevier), 57–78. doi: 10.1016/b978-0-12-805052-1.00003-6
Wernberg T., Smale D., Tuya F., Thomsen M. S., Langlois T. J., de Bettignies T., et al. (2013). An extreme climatic event alters marine ecosystem structure in a global biodiversity hotspot. Nat. Clim. Change 3, 78–82. doi: 10.1038/nclimate1627
Keywords: Cystoseira sensu lato, Mediterranean marine forests, marine threats, macroalgal restoration, macroalgal conservation, macroalgal distribution, ecological status, site prioritization
Citation: Verdura J, Rehues L, Mangialajo L, Fraschetti S, Belattmania Z, Bianchelli S, Blanfuné A, Sabour B, Chiarore A, Danovaro R, Fabbrizzi E, Giakoumi S, Iveša L, Katsanevakis S, Kytinou E, Nasto I, Nikolaou A, Orfanidis S, Rilov G, Rindi F, Sales M, Sini M, Tamburello L, Thibaut T, Tsirintanis K and Cebrian E (2023) Distribution, health and threats to Mediterranean macroalgal forests: defining the baselines for their conservation and restoration. Front. Mar. Sci. 10:1258842. doi: 10.3389/fmars.2023.1258842
Received: 14 July 2023; Accepted: 27 October 2023;
Published: 20 November 2023.
Edited by:
Joao M. Neto, University of Coimbra, PortugalReviewed by:
Sarah Caronni, University of Milan Bicocca, ItalyElisabetta Manea, UMR8222 Laboratoire d’Ecogéochimie des Environnements Benthiques (LECOB), France
Copyright © 2023 Verdura, Rehues, Mangialajo, Fraschetti, Belattmania, Bianchelli, Blanfuné, Sabour, Chiarore, Danovaro, Fabbrizzi, Giakoumi, Iveša, Katsanevakis, Kytinou, Nasto, Nikolaou, Orfanidis, Rilov, Rindi, Sales, Sini, Tamburello, Thibaut, Tsirintanis and Cebrian. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jana Verdura, amFuYS52ZXJkdXJhQHVuaXYtY290ZWRhenVyLmZy
†These authors have contributed equally to this work and share first authorship