 Leonardo Lopes Costa1,2*
Leonardo Lopes Costa1,2* Eduardo Manuel Rosa Bulhões3João Pedro Almeida Caetano1Vitor Figueira Arueira1Daniella Torres de Almeida4Tatiane Bittar Vieira4Leandro Jorge Telles Cardoso5
Eduardo Manuel Rosa Bulhões3João Pedro Almeida Caetano1Vitor Figueira Arueira1Daniella Torres de Almeida4Tatiane Bittar Vieira4Leandro Jorge Telles Cardoso5 Ilana Rosental Zalmon1
Ilana Rosental Zalmon1- 1Laboratório de Ciências Ambientais, Centro de Biociências e Biotecnologia, Universidade Estadual do Norte Fluminense Darcy Ribeiro, Campos dos Goytacazes, Brazil
- 2Instituto Solar Brasil de Desenvolvimento Saúde e Pesquisa, Rio de Janeiro, Brazil
- 3Programa de Pós-graduação em Geografia, Universidade Federal Fluminense, Campos dos Goytacazes, Rio de Janeiro, Brazil
- 4Fundação Projeto Tamar, São João da Barra, Rio de Janeiro, Brazil
- 5Reserva Caruara, São João da Barra, Rio de Janeiro, Brazil
Nest beach selection by sea turtles at a regional scale may depend on upper shore environmental cues. Thus, the variability of sandy beach conditions, including coastal erosion rates and urban development can affect their nesting ecology. Almost half of worldwide sandy beaches are suffering from erosion and most of them are located in highly developed areas. This can lead to shore stretches with coastal squeeze and loss of habitat for beach-dependent species such as sea turtles. Understanding whether an endangered species, such as the loggerhead sea turtle Caretta caretta (Linnaeus, 1758), selects certain beaches to nest is the first step in devising mitigation strategies for human activities and climate change impacts. We predicted that the occurrence of loggerhead nests along 62 km of sandy beaches in southeastern Brazil is negatively related to higher urbanization levels and extreme erosion. A total of 731 nests of the loggerhead sea turtle were recorded in the 2022-2023 nesting season. Only 6% of those nests were found on beach sectors with severe and extreme erosion. The probability of laying a nest was negatively related to erosion rate. Also, approximately 50% of nests were found on low-urbanized beaches; thus, the higher the urbanization, the lower the probability of a turtle laying a nest. Nature-based solutions to recover the beach-dune system are recommended in order to mitigate coastal erosion where loggerheads are avoiding nesting. This should be coupled with management planning of the protected areas where nests are laid and must include nest relocation and monitoring as priority actions.
1 Introduction
Loggerhead sea turtles Caretta caretta (Linnaeus, 1758) undertake transoceanic migrations during their life cycle, and return to their emergence region for nesting after spending years in distant foraging grounds (Reis et al., 2010; Baltazar-Soares et al., 2020). Sea turtle species use multiple cues to navigate, relying on a “magnetic map” for long-distance movements (geomagnetic imprinting hypothesis) and then relying on local signals to pinpoint specific nesting areas (Lohmann and Lohmann, 2019). Although females return to nest within the geographic region of their natal sandy beach (“natal homing”), they may select different nest sites spanning a few hundred kilometers along coastlines (Bjorndal et al., 1983; Bowen et al., 1993; Patino-Martinez et al., 2023). Thus, the variability of sandy beach conditions in the aforementioned scale certainly drives nest site selection in loggerhead sea turtles (Weishampel et al., 2016; Hatase and Omuta, 2018; Patino-Martinez et al., 2023).
Nest site selection has long been one of the main cornerstones of research on the ecology of sea turtles (Miller et al., 2003). As they do not have parental care, nest site selection at different spatial scales is consensually decisive for embryo development, hatchling survival, and reproductive success (Serafini et al., 2009; Hatase and Omuta, 2018; Patino-Martinez et al., 2023). The nesting behavior of sea turtles can be divided into beach selection, emergence of the female, and nest placement (Wood and Bjorndal, 2000). The factors that determine nest placement at fine scales are well-known and represent favorable microhabitats for hatchling success (Miller et al., 2003). In general, female loggerhead sea turtles nest in sparsely vegetated areas with higher elevation (Serafini et al., 2009; Turkozan et al., 2011; Hatase and Omuta, 2018; Patino-Martinez et al., 2023). They usually select the middle of the beach, minimizing the risk of nest flooding close to the tideline, intrusion of plant roots that entangle hatchlings, desiccation, and predation in densely vegetated zones upper shore (Allen et al., 2001; Fujisaki et al., 2018; Martins et al., 2022).
Beach selection and emergence of the female probably depend on shore signals and beach characteristics rather than a magnetic map (Wood and Bjorndal, 2000). The local and landscape features that drive these phases of habitat selection are however controversial. Some authors have found higher nest densities on wider beaches, arguing that they provide more diverse and favorable microhabitats (Mazaris et al., 2006; Hatase and Omuta, 2018). Oppositely, Kikukawa et al. (1999) found that the loggerhead sea turtle selected mainly narrower beaches, although distance to urban settlements was the main predictor of nest abundance. Indeed, anthropogenic factors such as coastal development, artificial light, human-made barriers to movement, human presence, and seawalls have been reported to negatively affect the beach selection by the loggerhead sea turtle (Rizkalla and Savage, 2011; Weishampel et al., 2016; de la Esperanza et al., 2017; Siqueira-Silva et al., 2020). An example is the false crawl, when a sea turtle comes ashore laden with eggs but turns around without nesting, usually because of unfavorable conditions (Miller et al., 2003). The results are not unanimous and other studies did not find negative effects of human stressors on beach selection by the loggerhead sea turtle (Kelly et al., 2017; Stanley et al., 2020).
Although erosion is a natural phenomenon inherent to coastal dynamics, it has been intensified by human activities mainly because of the advance of infrastructure towards the sea and the suppression of dunes and coastal vegetation. The increasing rates of urban development and supporting infrastructure impair the capacity of sandy beaches to provide coastal protection. Coastal infrastructure prevents landward migration of beaches, especially when facing sea-level rise, storms, and flooding events (Almar et al., 2021). Vousdoukas et al. (2020) forecast that coastal retreat might result in a loss of almost half of the world’s sandy beaches by the end of the century. Simultaneously, only 15% of the world’s coastal areas remain undisturbed by human activities (Williams et al., 2021). This imminent and widespread “coastal squeeze” is culminating in the loss of habitat for beach-dependent species such as sea turtles (Mazaris et al., 2009).
As a response from the scientific community, studies assessing the effects of sea-level rise and erosion in sea turtle nesting habitat have emerged. Most of them have modeled how nesting habitat will likely be lost within scenarios of sea-level rise (Fish et al., 2008; Fuentes et al., 2010; Varela et al., 2019; Veelenturf et al., 2020; Sönmez et al., 2021). However, few studies have verified whether current erosion scenarios have already affected the distribution of nests along sandy coasts (Witherington et al., 2009; Spanier, 2010). None of them explicitly measured historical shoreline displacement, and their correlative studies rely on very dynamic short-term erosion proxies such as beach slope and width.
Successful conservation efforts rely upon the ability of managers to understand current threats, and quantify and mitigate them to threatened species. One of the first steps in devising mitigation strategies for human activities and climate change impacts is to understand which are the habitat requirements of endangered species, such as the loggerhead turtle (globally “Vulnerable” according to IUCN, 2022), to ensure they thrive in viable populations. Herein, we aimed to test whether the probability of finding loggerhead sea turtle nests is affected by coastal erosion and urban development along 62 km of coast in one of the most important loggerhead turtle nesting areas in Brazil. To our knowledge, this is the first study that simultaneously evaluated the effect of erosion and urbanization on the frequency of sea turtles nesting on sandy beaches using raster-based predictor variables. We hypothesized that the occurrence of nests is negatively related to higher urbanization levels and extreme erosion rates.
2 Materials and methods
2.1 Study area
Loggerhead sea turtle nests were monitored in the north of Rio de Janeiro state, Southeastern Brazil (Figure 1). This area represents one of the most important loggerhead turtle nesting sites (~20% of all loggerhead nesting) along the Brazilian coast (Marcovaldi and Chaloupka, 2007). In this region, the installation of Açu Port-Industrial Complex (PIC) in 2007 boosted a rapid process of urbanization and economic, social, aesthetic, and environmental changes (Neves et al., 2022). Açu PIC stands out as the largest private port-industry complex in Latin America, comprising a total area of 130 km2 with 90 km2 of retro-area for installation of industries and companies (Neves et al., 2022). Due to its size and impact on the environment, Açú PIC, as part of an environmental compensation, supports the regional monitoring of sea turtles carried out by the Tamar Project Foundation (FPT), the largest and most successful sea turtle conservation program in Brazil. The monitoring program is coordinated by Caruara Reserve with the support of the Açú Pic companies, while the FPT carries out the monitoring activities.
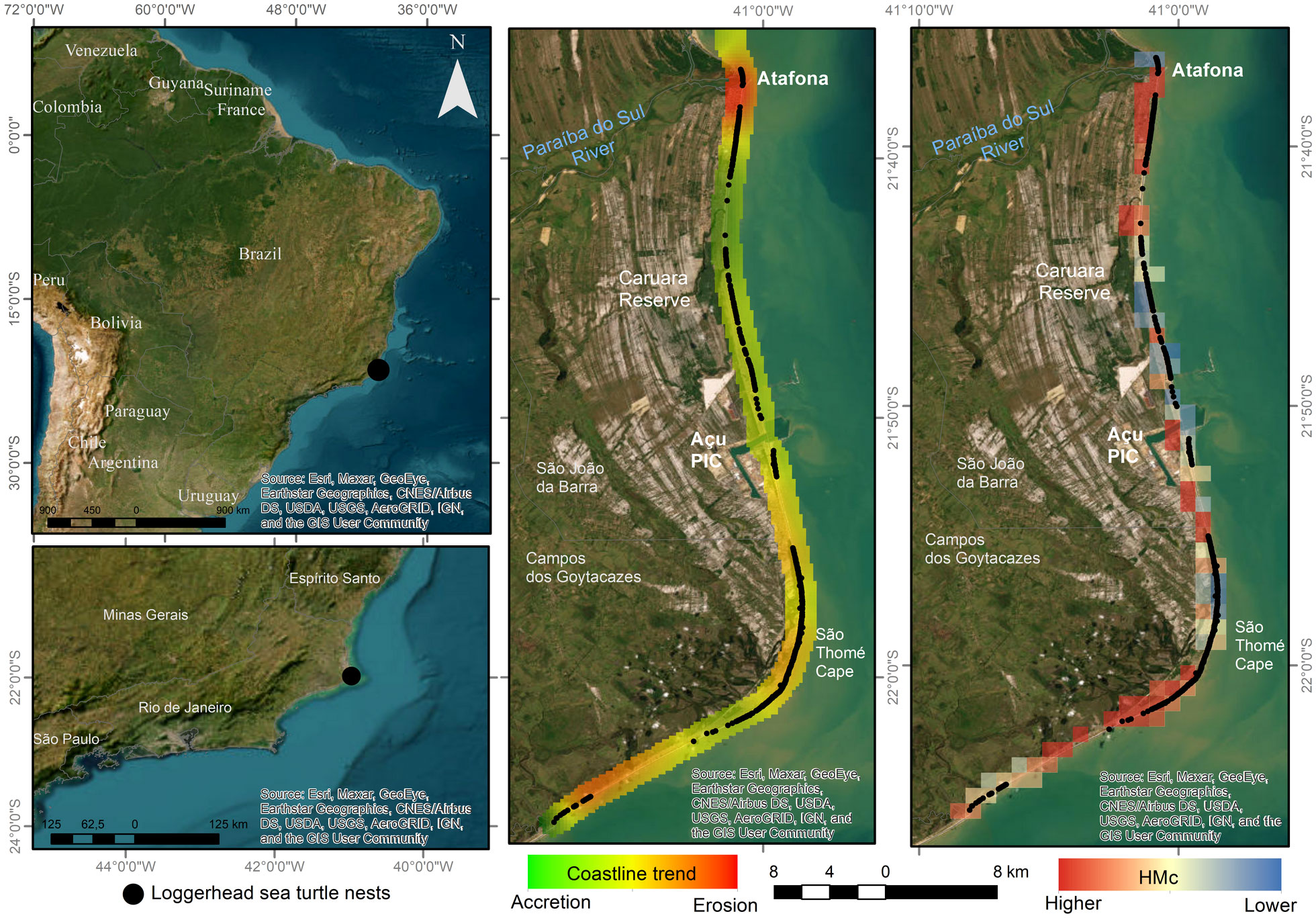
Figure 1 Map of the study area showing the distribution of the loggerhead sea turtle nests (2022/2023 nesting season) along 62 km of sandy beaches. HMc, Human modification metric.
The morphology of the continental shelf of the north of Rio de Janeiro state interferes with the wave propagation pattern, resulting in several beaches with different physical characteristics on a regional scale (Alves et al., 2018). An unconsolidated sandy cape (Cape of São Thomé) marks the transition between regressive barriers to the north and transgressive barriers to the south (Oliveira et al., 2016; Alves et al., 2018). Southward of the cape, sandy barriers are exposed to storm waves and overwash processes that promote the migration of this feature toward the mainland. Oppositely, northward sandy barriers typically have progradation features influenced by recent fluvial sedimentation (Alves et al., 2018).
In the region, beach morphodynamics undergoes high volumetric variation in response to waves, winds, and tides, creating morphological instability with a tendency toward erosion in certain areas. A coastal segment of about 4 km in length located immediately southward from the Paraíba do Sul River mouth (where sea turtle nests are also monitored) has experienced a dramatic erosion rate (up to 5.4 m/year); this has been related to the reduction of river sediment and water discharge due to the construction of dams, groins and jetties, and retention of sand in the lower estuary (Muehe et al., 2010). In this area (named Atafona village), coastal erosion destroyed more than 200 buildings between 1975 and 2016 (Lämmle et al., 2022). On the other hand, northward of Atafona, sedimentary accretion culminates in the progradation of various beaches. Thus, along the 62 km of monitored loggerhead turtle nesting areas, there are urban, peri-urban, and rural beaches, as well as sandy beaches with erosion and accretion dynamics and variable morphodynamics.
2.2 Nest monitoring
The sea turtle monitoring program is coordinated by the Caruara Reserve and supported by Açú Pic companies. The dataset was then provided by the Caruara Reserve for this study of nesting habitat selection. The FPT carried out (as a technical-scientific consultant) the monitoring in the last two nesting seasons.
The loggerhead turtle nesting season begins in September of one year and ends in March of the following year. Daily morning patrols (06:00 to 9:00) took place along 62 km of beach during the entire 2022-2023 nesting season. Once each turtle crawl site is located, the nest is identified and marked by a wood stake. Both nests of the loggerhead sea turtle and detected false crawls (i.e., crawls that do not result in nesting) were georeferenced. Monitoring was done using an off-road vehicle and followed the FIC-ICMBIO patrolling protocol (ICMBIO, 2018).
2.3 Urbanization
The urbanization level surrounding each nest was estimated by the HMc - Human Modification Metric (Kennedy et al., 2019). The HMc measures the degree of human modification across georeferenced lands calculated as the per-pixel product (HMs) of the spatial extent and the expected intensity of impacts including human population density, build-up areas, croplands, livestock, roads, mining, oil wells, wind turbines, and night-time lights through satellite imagery (raster data with 1,000 m resolution). The final HMc value is calculated as:
This fuzzy sum is a function that assumes that the contribution of a given factor decreases as other stressors co-occur. HMc is a continuous gradient of modification ranging from 0 to 1. The HMc values were obtained from the geographic coordinates of each nest at a resolution of 1,000 m using the packages “raster” (Hijmans, 2020) and “rgdal” (Bivand et al., 2021) in R software. This spatial resolution (1,000 m) was chosen because it is closely related to the categorical classification of the level of human disturbance on beaches at local scales (Barboza et al., 2021).
2.3.1 Erosion rates
Coastline displacement data from a global study covering a 32-year period (1984-2016, Luijendijk et al., 2018) was extracted for the study area at 500-meter intervals. This allowed the identification of segments undergoing change rates in accretion (above 0.5m/year), stability (between -0.5 and 0.5 m/year), and erosion (below -0.5 m/year). Negative coastline displacement (erosion) data were further classified into weak erosion (rates between -0.5 and -1.0 m/year), intense erosion (between -1 and -3 m/year), severe erosion (between -3 and -5 m/year), and extreme erosion (above -5 m/year). These data were interpolated using the natural neighbor method to create a raster, enabling an analysis of erosion trends across the study area. Similar to HMc, the coastline displacement values were gathered using the package “raster” in R Software.
2.4 Data analysis
A data matrix containing the location of all nests, HMc, and coastal displacement values was used to build generalized linear models (GLMs) with binomial distribution. Separate GLMs were employed to test whether the presence of nests and false crawls of the loggerhead sea turtle along 62 km of beach are impacted by urbanization level and erosion rate. Nest presence (1), false crawls occurrence (1), and random points (0) were the response variables included in the GLMs, whilst “HMc” and “erosion rate” were tested as predictor variables. The presence of collinearity on predictors was assessed by the variance inflation factor (VIF) using a threshold of VIF ≥ 3 for variable exclusion, but no variable was removed. For the random points (0), HMc and erosion rate values were randomly generated, considering the intervals between the maximum (erosion rate= 4.64; HMc= 0.78) and minimum values (erosion rate= -7.28; HMc= 0.20) of each variable as limits. We decided to randomly generate predictor variables instead of drawing beach sectors to calculate HMc and erosion rates because most of the monitored area still has nests; thus, the variability of the predictors in the “0” treatments would be biased. Apart from models, we randomly selected 731 stretches of beach without nests to further verify the coastal displacement tendency in these areas. The analysis was performed in R software.
3 Results
3.1 Nest beach selection
A total of 731 nests of the loggerhead sea turtle were reported in the 2022-2023 nesting season (between September 2022 and March 2023) in the study area. Only 6% of nests were found on beach sectors with severe and extreme erosion (Figure 2A); among beach sectors without turtle nests (n= 731 random sectors), ~41% are under severe and extreme erosion (Figure 2B). Approximately 55% of nests were found in beach sectors under stability or accretion, and 34% were found in sectors with intense erosion (Figure 2A). The probability of finding a nest was significantly related to the erosion rate (z= 4.681, p< 0.0001). The higher the erosion rate (more negative coastal displacement) the lower the probability of finding a nest (Figure 3A).
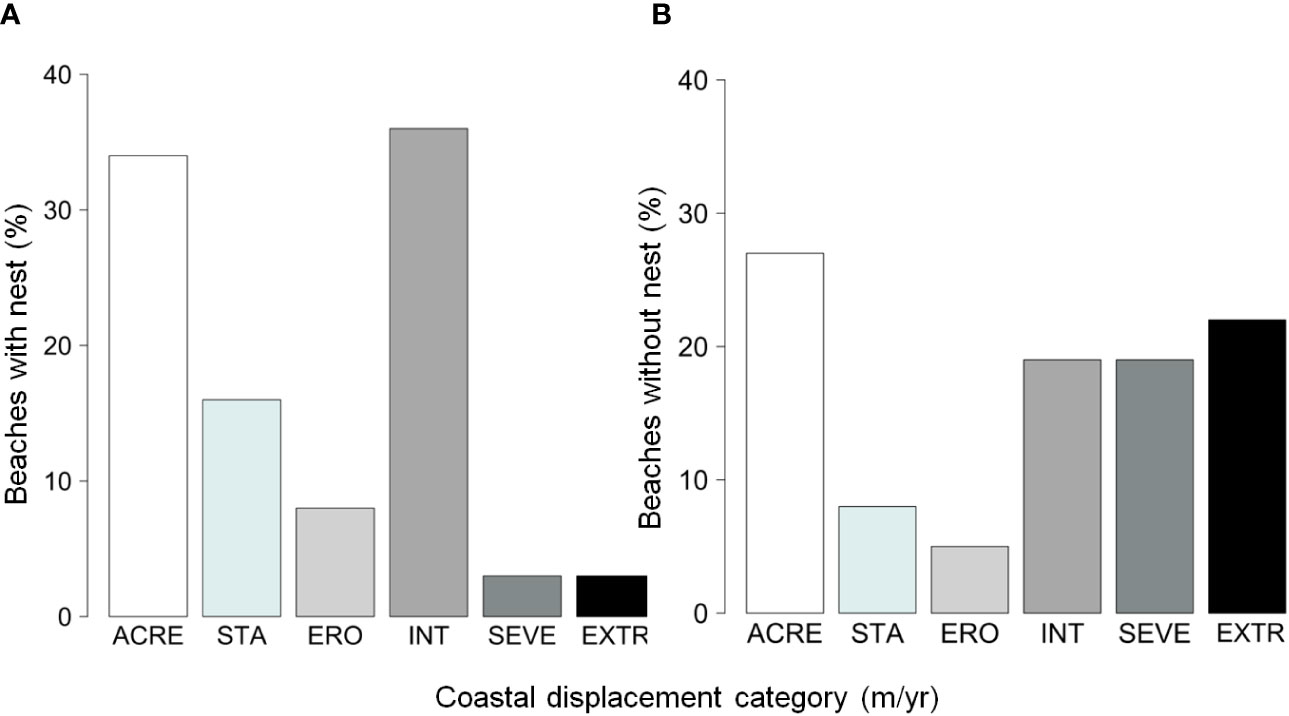
Figure 2 Percentage of beaches with nests (A) and without nests (B) within the different erosion categories. ACRE, accretion (above 0.5 m/year); STA, stability (between -0.5 and 0.5 m/year); ERO, weak erosion (between -0.5 and -1.0 m/year); INT, intense erosion (between -1 and -3 m/year); SEVE, severe erosion (between -3 and -5 m/year); EXTR, extreme erosion (above -5 m/year).
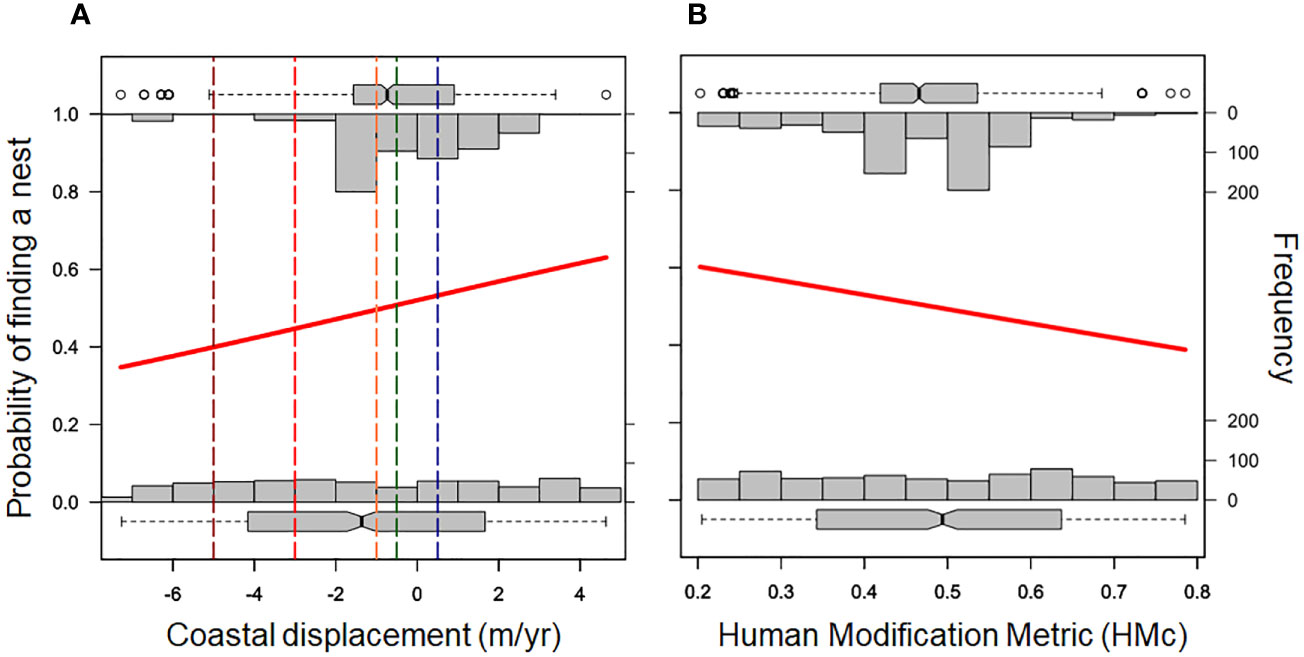
Figure 3 Probability of finding a loggerhead turtle nest along 62 km of beaches in the north of Rio de Janeiro state as a function of coastal displacement trends (A) and urbanization level (1,000 m spatial resolution). The solid red line represents the binomial generalized linear model (GLM). The top bars represent the number of beaches with nests under different conditions of erosion (A) and urbanization (B); the bottom bars represent random values for both of the aforementioned independent variables. The vertical lines in panel A represent erosion rating limits; from left to right: extreme erosion (dark red), severe erosion (light red); intense erosion (orange), stability (green), and accretion (blue).
The binomial GLM also evidenced a negative effect of HMc in the probability of finding a nest on the beaches, although with a lower magnitude than the effect of erosion rates (z= -3.524, p= 0.0004) (Figure 3B). Nearly half of the nests (54%) were laid on beaches with HMc lower than 0.5 (Figure 3B).
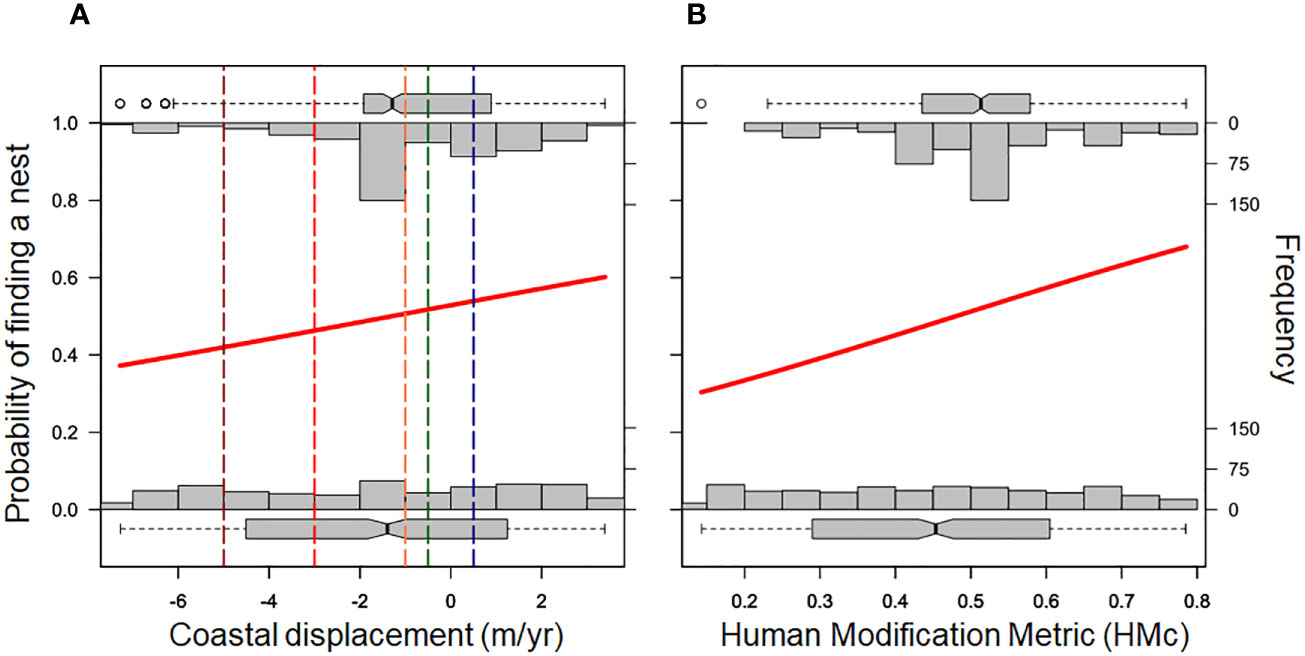
Figure 4 Probability of occurring a sea turtle false crawl along 62 km of beaches in the north of Rio de Janeiro state as a function of coastal displacement trends (A) and urbanization level (1,000 m spatial resolution). The solid red line represents the binomial generalized linear model (GLM). The top bars represent the number of beaches with nests under different conditions of erosion (A) and urbanization (B); the bottom bars represent random values for both of the aforementioned independent variables. The vertical lines in figure A represent erosion rating limits; from left to right: extreme erosion (dark red), severe erosion (light red); intense erosion (orange), stability (green), and accretion (blue).
3.2 False crawls
A total of 528 false crawls of sea turtles were reported in the 2022-2023 nesting season in the study area. False crawls followed the same trend of loggerhead nesting regarding beach erosion, being more frequent in beach sectors without severe and extreme erosion (Figure 4A). Thus, binomial GLM showed that the lower the erosion rate (z= 3.945, p< 0.0001), the higher the probability of occurring a sea turtle false crawl. On the other hand, false crawls were more frequent on beaches with HMc higher than 0.5 (~58%), opposite to the nesting pattern (Figure 4B). Thus, the higher the urbanization level, the higher the probability of occurring a false crawl, according to GLM (z= 6.038, p< 0.0001).
4 Discussion
The findings of this study suggest that the loggerhead sea turtles select beaches that are not undergoing severe and extreme erosive processes to nest, corroborating our hypothesis. This agrees with Spanier (2010) who observed the leatherback sea turtle Dermochelys coriacea (Linnaeus, 1758) nested mainly at locations that were not eroding. Similarly, Witherington et al. (2009) observed local spatial shifts in the loggerheads nesting in Florida, USA as chronic and acute erosion increased over time. According to Pfaller et al. (2009), females with nesting experience select more successful nest sites than females with little experience. In addition, low nesting success of loggerheads is usually found in eroding beaches (Lamont and Houser, 2014). The lower number of nests on beaches with severe and extreme erosion, therefore, might reflect previous unsuccessful nesting experiences (Pfaller et al., 2009). This argument is reinforced by the fact that false crawls occurred more frequently in the same areas with higher nesting frequency, as found by Lamont and Houser (2014). Thus, the loggerhead sea turtle might decide not to nest in eroding beaches before emergence or at the very beginning of the crawl, preventing the expenditure of energy resulting from false crawls in beaches where nesting habitat is being lost (Mazaris et al., 2006). Future studies could tag nesting females or measure their body size (assuming that larger turtles are older) to compare nesting habitat selection between successive nesting seasons and properly test this “nesting experience hypothesis”.
Selecting beaches without extreme erosion is critical to sea turtles, firstly because of imminent loss of nesting habitat, and secondly because when selecting eroding beaches, female sea turtles may have to place their nests in locations where they would not conventionally do. Fujisaki et al. (2018) found temporal shifts in locations where loggerheads placed nests in relation to coastal erosion along a barrier island beach in the northern Gulf of Mexico. They observed that in eroding beaches, loggerheads nested significantly closer to the vegetation line, possibly relying on environmental cues that indicate the inundation risk of nests. However, in eroding beaches, these formerly reliable signals might no longer represent adaptive outcomes. The presence of plant roots upper shore provokes hatchling disorientation and shadow of the elements backing the beach that could alter hatchling sex ratios (Miller et al., 2003). In addition, predation is expected to be more intense in the upper zones of the beach. Wetterer et al. (2007) discussed that three sea turtle species nested further from dune vegetation after beaches were artificially replenished (to contain coastal erosion), being less exposed to predaceous fire ants.
Changing the location of the nest at a fine scale rather than avoiding eroding beaches can be therefore an ecological trap. Similarly, when females nest on highly urbanized coasts they put their offspring at risk because of trampling, vehicles, and artificial light. Similar to Kikukawa et al. (1999); Weishampel et al. (2016), and Price et al. (2018), we verified that the probability of finding loggerhead nests was negatively related to higher urbanization levels. Accordingly, false crawls were more frequent in more developed areas, thus we argue that females regularly give up on laying eggs after encountering highly impacted beaches within urban areas. However, several nests still occur on developed coasts, also because almost half of the beaches in the region suffer erosion; thus, urban beaches in accretion can be selected more frequently for nesting in future nesting seasons. Thus, alternatives for management need to be discussed, including measures to mitigate erosion effects in this hotspot of loggerhead sea turtle nesting.
Sandy beach management in Brazil remains overwhelmingly focused on engineering interventions to mitigate erosion, while ecological components are often neglected. The scenario for the northern coast of the state of Rio de Janeiro is even worse; despite technical determinations, nothing has been done to contain erosion in several beaches, including loggerhead sea turtle nest sites. To mitigate intense and chronic coastal erosion, the placement of artificially pumped compatible sands from the Paraíba do Sul River mouth to rebuild the beach-dune system has been proposed as a suitable nature-based solution (instead of hard engineering intervention). This approach involves concomitant foredune reforestation and conservation measures to preserve the integrity of the ecosystem, which should be locally understood as a fundamental natural structure for coastal defense. This solution was suggested as a method considered optimal for controlling the problem of coastal erosion (Bulhões, 2023). This intervention attempts to mirror the river’s natural role in providing sediment to adjacent beaches, which has been lost because of human impacts in its course. Plantations of non-native vegetation and hard-engineering intervention should be discouraged, because it is inefficient to contain erosion and it has dramatic effects on nesting turtles (Rizkalla and Savage, 2011; de Vos et al., 2019).
Effects of beach nourishment on turtles are controversial, challenging decision-making processes regarding erosion management in loggerhead nesting sites. Beach replenishment provides nesting habitats that otherwise were unavailable (Fujisaki et al., 2018), but in some cases, nourished beaches become less suitable for turtle nesting than natural beaches (Milton et al., 1997; Rumbold et al., 2001). Nourishment can alter natural parameters of natural beaches, such as sand density, shear resistance, moisture content, slope, sand color, grain size, sand shape, and sand mineral content, impacting the reproductive success of sea turtles (Rumbold et al., 2001). However, when erosion is extreme and the nesting area has already been lost, beach restoration becomes an almost indisputable management solution. In such cases, variables such as source sand, application technique, maintenance, and monitoring are decisive in determining the success of nourishment projects with respect to sea turtles nesting and thus should be considered (Milton et al., 1997). Finally, under erosion scenarios in which loggerhead turtles still lay eggs on the beach, nest translocation to beaches in accretion or stability could be a less costly and logistically simpler solution to increase reproductive success (Pike, 2008; Pfaller et al., 2009).
When translocating nests to beaches without predominance of erosive dynamics, coastal development should also be considered. Here we found that lower nesting frequency and more false crawls occurred on beaches with higher urbanization, and thus, human stressors are impacting nest site selection. Indeed, nesting on developed coasts promotes ecological traps (i.e., maladaptive habitat selection). Disoriented hatchlings have been found on roads near urban beaches in the region, causing road-kills. Further research should be carried out to verify spatial factors that increase the risk of beach animals being road-killed, including artificial light and urbanization level (Costa et al., 2020a; Costa et al., 2020b). With these results, it will be possible to find hotspots of car-fauna collisions, from where nests must be translocated or even barriers that could avoid animals emigrating to roads could be built. All these proposed management actions would be facilitated by the recent implementation of coastal marine protection areas (“APA das Dunas e Restingas” and “Private Reserve of Natural Heritage Caruara” - Caruara Reserve), but this does not guarantee their success (Nel et al., 2013). Thus, the creation of management planning focused on the loggerhead sea turtle, including sensitivity maps, is strongly encouraged (Lopez et al., 2015), as well as long-term nesting monitoring. This will certainly promote protective ramifications to sandy beach resident fauna, which is also dramatically affected by loss of habitat associated with erosion, urban development, and consequent coastal squeeze.
5 Conclusion
In conclusion, coastal erosion and urban development threaten loggerhead turtle nesting in one of their most important breeding areas in Brazil. Consequently, the region has several ecological traps that require decision-makers to draw up urgent management and conservation plans. We strongly recommend that nest translocation from extremely eroded to stable areas and from urban to low-urban areas is prioritized, as has already occurred in these 62 km of coast. We also suggest that resident fauna must be embraced in management action and monitoring. The selection of priority areas for conservation in the region may be based on the co-occurrence of loggerhead turtle nests and greater diversity and irreplaceability of the supralittoral and dune vegetation faunal community; beach arthropods are as dependent on the integrity of the beach-dune system as sea turtles. Therefore, sea turtles could function as umbrella species in protecting the biodiversity of sandy beaches.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
LLC and EB led the analyses and writing of the manuscript contributed critically to the drafts and gave final approval for publication. JC, VA, DA, TV, LTC, and IZ contributed critically to the drafts and gave final approval for publication. All authors contributed to the article and approved the submitted version.
Funding
LLC is supported by Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro - FAPERJ (E-26.210.384/2022 and E-26.200.620/2022). IZ is supported by FAPERJ (259920/2021) and the Brazilian Agency for Research Development — CNPq (301203/2019-9). JC is supported by the “Mais Ciência” program coordinated by the secretary of Education, Science and Technology of the municipal government of Campos dos Goytacazes.
Acknowledgments
We thank Porto do Açu Operações (CNPJ: 11.198.242/0001-58) and the management of the Caruara Reserve for providing the data collected through its support to the Marine Turtle Conservation Project (PMTM-Tamar).
Conflict of interest
Authors DA and TV were employed by Fundação Projeto TAMAR and LTC was employed by Reserva Caruara.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Allen C. R., Forys E. A., Rice K. G., Wojcik D. P. (2001). Effects of fire ants (Hymenoptera: Formicidae) on hatching turtles and prevalence of fire ants on sea turtle nesting beaches in Florida. Source: Florida Entomol. 84, 250–253. doi: 10.2307/3496175
Almar R., Ranasinghe R., Bergsma E. W. J., Diaz H., Melet A., Papa F., et al. (2021). A global analysis of extreme coastal water levels with implications for potential coastal overtopping. Nat. Commun. 12, 3775. doi: 10.1038/s41467-021-24008-9
Alves L. D., Bulhões E. M. R., Di Beneditto A. P. M., Zappes C. A. (2018). Ethnoclimatology of artisanal fishermen: Interference in coastal fishing in southeastern Brazil. Mar. Policy 95, 69–76. doi: 10.1016/j.marpol.2018.07.003
Baltazar-Soares M., Klein J. D., Correia S. M., Reischig T., Taxonera A., Roque S. M., et al. (2020). Distribution of genetic diversity reveals colonization patterns and philopatry of the loggerhead sea turtles across geographic scales. Sci. Rep. 10 (1), 18001. doi: 10.1038/s41598-020-74141-6
Barboza C. A., de M., Mattos G., Soares-Gomes A., Zalmon I. R., Costa L. L. (2021). Low densities of the ghost crab Ocypode quadrata related to large scale human modification of sandy shores. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.589542
Bivand R., Keitt T., Rowlingson B. (2021). rgdal: Bindings for the “Geospatial” Data Abstraction Library.
Bjorndal K. A., Meylan A. B., Turner B. J. (1983). Sea turtles nesting at Melbourne Beach, Florida, 1. Size, growth and reproductive biology. Biol. Conserv. 26, 65–77. doi: 10.1016/0006-3207(83)90049-6
Bowen B., Avise J. C., Richardson J. I., Meylan A. B., Margaritoulis D., Hopkins-Murphy S. R. (1993). Population structure of Loggerhead Turtles (Caretta caretta) in the Northwestern Atlantic Ocean and Mediterranean Sea. Conserv. Biol. 7, 834–844. doi: 10.1046/j.1523-1739.1993.740834.x
Bulhões E. M. R. (2023). “Coastal erosion and coastal defense,” in Proceedings of the II International Workshop of Quaternary Geomorphology: Geochronology of Fluvial and Coastal Environments. Ed. Perez F.
Costa L. L., Mothé N. A., Zalmon I. R. (2020a). Light pollution and ghost crab road-kill on coastal habitats. Reg. Stud. Mar. Sci. 39, 101457. doi: 10.1016/j.rsma.2020.101457
Costa L. L., Secco H., Arueira V. F., Zalmon I. R. (2020b). Mortality of the Atlantic ghost crab Ocypode quadrata (Fabricius 1787) due to vehicle traffic on sandy beaches: A road ecology approach. J. Environ. Manage 260, 110168. doi: 10.1016/j.jenvman.2020.110168
de la Esperanza O. A., Arenas Martínez A., Tzeek Tuz M., Pérez-Collazos E. (2017). Are anthropogenic factors affecting nesting habitat of sea turtles? The case of Kanzul beach, Riviera Maya-Tulum (Mexico). J. Coast. Conserv. 21, 85–93. doi: 10.1007/s11852-016-0473-5
de Vos D., Nel R., Schoeman D., Harris L. R., du Preez D. (2019). Effect of introduced Casuarina trees on the vulnerability of sea turtle nesting beaches to erosion. Estuar. Coast. Shelf. Sci. 223, 147–158. doi: 10.1016/j.ecss.2019.03.015
Fish M. R., Côté I. M., Horrocks J. A., Mulligan B., Watkinson A. R., Jones A. P. (2008). Construction setback regulations and sea-level rise: Mitigating sea turtle nesting beach loss. Ocean Coast. Manag. 51, 330–341. doi: 10.1016/j.ocecoaman.2007.09.002
Fuentes M. M. P. B., Limpus C. J., Hamann M., Dawson J. (2010). Potential impacts of projected sea-level rise on sea turtle rookeries. Aquat. Conserv. 20, 132–139. doi: 10.1002/aqc.1088
Fujisaki I., Lamont M., Carthy R. (2018). Temporal shift of sea turtle nest sites in an eroding barrier island beach. Ocean Coast. Manag. 155, 24–29. doi: 10.1016/j.ocecoaman.2017.12.032
Hatase H., Omuta K. (2018). Nest site selection in loggerhead sea turtles that use different foraging areas: do less fecund oceanic foragers nest at safer sites? J. Zool. 305, 232–239. doi: 10.1111/jzo.12557
ICMBIO (2018). Programa de monitoramento de Tartarugas Marinhas do Parque Nacional Marinho dos Abrolhos.
IUCN (2022). The IUCN Red List of Threatened Species. Version 2022-2. https://www.iucnredlist.org. Accessed on [14 june 2023].
Kelly I., Leon J. X., Gilby B. L., Olds A. D., Schlacher T. A. (2017). Marine turtles are not fussy nesters: A novel test of small-scale nest site selection using structure from motion beach terrain information. PeerJ 5, 1–25. doi: 10.7717/peerj.2770
Kennedy C. M., Oakleaf J. R., Theobald D. M., Baruch-Mordo S., Kiesecker J. (2019). Managing the middle: A shift in conservation priorities based on the global human modification gradient. Glob. Chang. Biol. 25, 811–826. doi: 10.1111/gcb.14549
Kikukawa A., Kamezaki N., Ota H. (1999). Factors affecting nesting beach selection by loggerhead turtles (Caretta caretta): A multiple regression approach. J. Zool. 249, 447–454. doi: 10.1111/j.1469-7998.1999.tb01214.x
Lämmle L., Perez Filho A., Donadio C., Arienzo M., Ferrara L., Santos C., et al. (2022). Anthropogenic pressure on hydrographic basin and coastal erosion in the delta of Paraíba do Sul River, southeast Brazil. J. Mar. Sci. Eng. 10, 1585. doi: 10.3390/jmse10111585
Lamont M. M., Houser C. (2014). Spatial distribution of loggerhead turtle (Caretta caretta) emergences along a highly dynamic beach in the northern Gulf of Mexico. J. Exp. Mar. Biol. Ecol. 453, 98–107. doi: 10.1016/j.jembe.2013.11.006
Lohmann K. J., Lohmann C. M. F. (2019). There and back again: Natal homing by magnetic navigation in sea turtles and salmon. J. Exp. Biol. 222, 1–10. doi: 10.1242/jeb.184077
Lopez G. G., Saliés E., de C., Lara P. H., Tognin F., Marcovaldi M. A., et al. (2015). Coastal development at sea turtles nesting ground: Efforts to establish a tool for supporting conservation and coastal management in northeastern Brazil. Ocean Coast. Manag. 116, 270–276. doi: 10.1016/j.ocecoaman.2015.07.027
Luijendijk A., Hagenaars G., Ranasinghe R., Baart F., Donchyts G., Aarninkhof S. (2018). The state of the world’s beaches. Sci. Rep. 8, 6641. doi: 10.1038/s41598-018-24630-6
Marcovaldi M., Chaloupka M. (2007). Conservation status of the loggerhead sea turtle in Brazil: An encouraging outlook. Endanger Species Res. 3, 133–143. doi: 10.3354/esr003133
Martins S., Patrício R., Clarke L. J., de Santos Loureiro N., Marco A. (2022). High variability in nest site selection in a loggerhead turtle rookery, in Boa Vista Island, Cabo Verde. J. Exp. Mar. Biol. Ecol. 556, 151798. doi: 10.1016/j.jembe.2022.151798
Mazaris A. D., Matsinos Y. G., Margaritoulis D. (2006). Nest site selection of loggerhead sea turtles: The case of the island of Zakynthos, W Greece. J. Exp. Mar. Biol. Ecol. 336, 157–162. doi: 10.1016/j.jembe.2006.04.015
Mazaris A. D., Matsinos G., Pantis J. D. (2009). Evaluating the impacts of coastal squeeze on sea turtle nesting. Ocean Coast. Manag. 52, 139–145. doi: 10.1016/j.ocecoaman.2008.10.005
Miller J. D., Limpus C. J., Godfrey M. H. (2003). “Nest site selection, oviposition, eggs, development, hatching, and emergence of loggerhead turtles,” in Loggerhead Turtles. Eds. Alan B., Blair W., 125–143.
Milton S. L., Schulman A. A., Lutz P. L., Schulmant A. A., Lutzt P. L. (1997). The effect of beach nourishment with aragonite versus silicate sand on beach temperature and loggerhead sea turtle nesting success. J. Coast. Res. 13, 904–915.
Muehe D., Sperle M., Tessler M. G., Souza S. R. (2010). Can the trapping of shoreface sand by mud layers induce coastal erosion? The example of the Paraíba do Sul River outlet, northern Rio de Janeiro state. Braz. J. Oceanogr. 58, 65–68. doi: 10.1590/S1679-87592010000600008
Nel R., Punt A. E., Hughes G. R. (2013). Are coastal protected areas always effective in achieving population recovery for nesting sea turtles? PloS One 8, e63525. doi: 10.1371/journal.pone.0063525
Neves R. A. F., Lopes A., Naveira C., Rodrigues N., Silveira R. B., Guimarães T., et al. (2022). Socio-economic impacts of a maritime industrial development area (MIDA) model in Latin America: The case of the Açu Port-Industrial Complex. WMU J. Maritime Affairs 21, 351–368. doi: 10.1007/s13437-021-00261-z
Oliveira P., da C., Di Beneditto A. P. M., Bulhões E. M. R., Zappes C. A. (2016). Artisanal fishery versus port activity in southern Brazil. Ocean Coast. Manag. 129, 49–57. doi: 10.1016/j.ocecoaman.2016.05.005
Patino-Martinez J., Dos Passos L., Amador R., Teixidor A., Cardoso S., Marco A., et al. (2023). Strategic nest site selection in one of the world’s largest loggerhead turtle nesting colonies, on Maio Island, Cabo Verde. Oryx 57, 152–159. doi: 10.1017/S0030605321001496
Pfaller J. B., Limpus C. J., Bjorndal K. A. (2009). Nest-site selection in individual loggerhead turtles and consequences for doomed-egg relocation. Conserv. Biol. 23, 72–80. doi: 10.1111/j.1523-1739.2008.01055.x
Pike D. A. (2008). The benefits of nest relocation extend far beyond recruitment: A rejoinder to Mrosovsky. Environ. Manage 41, 461–464. doi: 10.1007/s00267-006-0434-0
Price J. T., Drye B., DOmangue R. J., Paladino F. V. (2018). Exploring the role of artificial lighting in loggerhead turtle (Caretta caretta) nest-site selection and hatchling disorientation. Herpetol. Conserv. Biol. 13, 415–422.
Reis E. C., Soares L. S., Vargas S. M., Santos F. R., Young R. J., Bjorndal K. A., et al. (2010). Genetic composition, population structure and phylogeography of the loggerhead sea turtle: Colonization hypothesis for the Brazilian rookeries. Conserv. Genet. 11, 1467–1477. doi: 10.1007/s10592-009-9975-0
Rizkalla C. E., Savage A. (2011). Impact of seawalls on loggerhead sea turtle (Caretta caretta) nesting and hatching success. J. Coast. Res. 27, 166–173. doi: 10.2112/JCOASTRES-D-10-00081.1
Rumbold D. G., Davis P. W., Perretta C. (2001). Estimating the effect of beach nourishment on Caretta caretta (Loggerhead Sea Turtle) nesting. Restor. Ecol. 9, 304–310. doi: 10.1046/j.1526-100X.2001.009003304.x
Serafini T. Z., Gilles Lopez G., Luís P., Da Rocha B. (2009). Nest site selection and hatching success of hawksbill and loggerhead sea turtles (Testudines, Cheloniidae) at Arembepe Beach, northeastern Brazil. Phyllomedusa 8, 3–17. doi: 10.11606/issn.2316-9079.v8i1p03-17
Siqueira-Silva I. S., Arantes M. O., Hackradt C. W., Schiavetti A. (2020). Environmental and anthropogenic factors affecting nesting site selection by sea turtles. Mar. Environ. Res. 162, 105090. doi: 10.1016/j.marenvres.2020.105090
Sönmez B., Karaman S., Turkozan O. (2021). Effect of predicted sea level rise scenarios on green turtle (Chelonia mydas) nesting. J. Exp. Mar. Biol. Ecol. 541, 151572. doi: 10.1016/j.jembe.2021.151572
Spanier M. J. (2010). Beach erosion and nest site selection by the leatherback sea turtle Dermochelys coriacea (Testudines: Dermochelyidae) and implications for management practices at Playa Gandoca, Costa Rica. Rev. Biol. Trop. 58, 1237–1246. doi: 10.15517/rbt.v58i4.5408
Stanley T. R., White J. M., Teel S., Nicholas M. (2020). Brightness of the night sky affects loggerhead (Caretta caretta) sea turtle hatchling misorientation but not nest site selection. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00221
Turkozan O., Yamamoto K., Yilmaz C. (2011). Nest site preference and hatching success of green (Chelonia mydas) and loggerhead ( ) sea turtles at Akyatan Beach, Turkey. Chelonian Conserv. Biol. 10, 270–275. doi: 10.2744/CCB-0861.1
Varela M. R., Patrício A. R., Anderson K., Broderick A. C., DeBell L., Hawkes L. A., et al. (2019). Assessing climate change associated sea-level rise impacts on sea turtle nesting beaches using drones, photogrammetry and a novel GPS system. Glob. Chang. Biol. 25, 753–762. doi: 10.1111/gcb.14526
Veelenturf C. A., Sinclair E. M., Paladino F. V., Honarvar S. (2020). Predicting the impacts of sea level rise in sea turtle nesting habitat on Bioko Island, Equatorial Guinea. PloS One 15, e0222251. doi: 10.1371/journal.pone.0222251
Vousdoukas M. I., Ranasinghe R., Mentaschi L., Plomaritis T. A., Athanasiou P., Luijendijk A., et al. (2020). Sandy coastlines under threat of erosion. Nat. Clim. Chang. 10, 260–263. doi: 10.1038/s41558-020-0697-0
Weishampel Z. A., Cheng W. H., Weishampel J. F. (2016). Sea turtle nesting patterns in Florida vis-à-vis satellite-derived measures of artificial lighting. Remote Sens. Ecol. Conserv. 2, 59–72. doi: 10.1002/rse2.12
Wetterer J. K., Wood L. D., Johnson C., Krahe H., Fitchett S. (2007). Predaceous ants, beach replenishment, and nest placement by sea turtles. Environ. Entomol. 36, 1084–1091. doi: 10.1093/ee/36.5.1084
Williams B. A., Watson J. E. M., Beyer H. L., Klein C. J., Montgomery J., Runting R. K., et al. (2021). The global rarity of intact coastal regions. Conserv. Biol. 36 (4), e13874. doi: 10.1111/cobi.13874
Witherington B., Kubilis P., Brost B., Meylan A. (2009). Decreasing annual nest counts in a globally important loggerhead sea turtle population. Ecol. Appl. 19, 30–54. doi: 10.1890/08-0434.1
Keywords: nest ecology, marine turtle, habitat loss, hatchling, nest translocation, urbanization
Citation: Costa LL, Bulhões EMR, Caetano JPA, Arueira VF, de Almeida DT, Vieira TB, Cardoso LJT and Zalmon IR (2023) Do costal erosion and urban development threat loggerhead sea turtle nesting? Implications for sandy beach management. Front. Mar. Sci. 10:1242903. doi: 10.3389/fmars.2023.1242903
Received: 19 June 2023; Accepted: 21 August 2023;
Published: 17 October 2023.
Edited by:
Salvatore Siciliano, Fundação Oswaldo Cruz (Fiocruz), BrazilReviewed by:
Camila Miguel, Instituto Marcos Daniel, BrazilEduardo Cuevas, Universidad Autónoma de Baja California, Mexico
Copyright © 2023 Costa, Bulhões, Caetano, Arueira, de Almeida, Vieira, Cardoso and Zalmon. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Leonardo Lopes Costa, bG9wZXMuYmlvLm1wLnNmaUBwcS51ZW5mLmJy