 Zunli Lu
Zunli Lu Ellen Thomas2,3
Ellen Thomas2,3 Rosalind E. M. Rickaby
Rosalind E. M. Rickaby Wanyi Lu
Wanyi Lu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
GENERAL COMMENTARY article
Front. Mar. Sci., 11 July 2023
Sec. Marine Biogeochemistry
Volume 10 - 2023 | https://doi.org/10.3389/fmars.2023.1221835
A Commentary on
Planktic foraminifera iodine/calcium ratios from plankton tows
by Winkelbauer HA, Hoogakker BAA, Chance RJ, Davis CV, Anthony CJ, Bischoff J, Carpenter LJ, Chenery SRN, Hamilton EM, Holdship P, Peck VL, Poulton AJ, Stinchcombe MC and Wishner KF (2023) Front. Mar. Sci. 10:1095570. doi: 10.3389/fmars.2023.1095570
I/Ca in planktic foraminiferal shells is one of the few proxies proposed for reconstructing past upper ocean oxygenation. Foraminiferal I/Ca values have been measured in globally distributed core-top samples, with low I/Ca values generally indicative of low-oxygen water (Lu et al., 2020a; Lu et al., 2020b). Winkelbauer et al. (2023) observed that “I/Ca values of plankton tow foraminifera from environments with well oxygenated subsurface waters, however, are an order of magnitude lower compared to core-tops from similarly well-oxygenated regions,” and concluded that “planktic foraminifera gain iodine post-mortem, either when sinking through the water column, or during burial.” These tow-derived low I/Ca values add to the information needed to mechanistically understand the planktic I/Ca proxy. However, we highlight the fact that tow-specimens do not represent the same populations as those present in core tops, and we think that further evidence is needed before reinterpreting the core-top and down-core planktic I/Ca records.
We suggest that the use of I/Ca in foraminifera from plankton tows (Winkelbauer et al., 2023) may not easily provide a direct comparison with either the populations or the geochemical signal of foraminifera present in core tops. Core-tops reflect time-averaged populations over hundreds to thousands of years, thus averaging seasonal and shorter-scale variability. Such short-term variability is extensive, because most species of planktic foraminifera have life spans of 2–4 weeks, and the dominant fluxes of tests occur after a reproductive event (e.g., Jonkers et al., 2015). In contrast, plankton tows reflect a snapshot on a much shorter time scale (a few days) than the life span of common species and thus may miss reproductive events. Horizontal tows at the depths of typical habitats where foraminifera live and calcify generally contain shells without the external crust formed during gametogenesis by many taxa, but it is difficult to evaluate exactly what populations are sampled by horizontal tows deeper than the typical habitats. Therefore, such tows cannot capture the average population over time and depth in a manner analogous to the core tops.
Tests without such late stages of calcification appear more often in plankton tows than in core tops, where larger, generally mature specimens are selected for analysis. Selecting fully grown dead shells from the tow materials (when available) introduces the possibility of measuring shells originating from locations that are not representative of the towed area. In addition, it is unclear to what extent the planktic I/Ca signal may be influenced by the chemistry of low-oxygen micro-environments within organic particles (Bianchi et al., 2018) rather than the well-oxygenated surrounding water masses, although it is uncertain how often foraminifera live in such flocs.
We want to emphasize that data from plankton tows, sediment traps and lab culturing all provide unique insights into foraminifera-based proxies and are complementary to core-top data. However, we need to keep in mind that results from such investigations do not necessarily agree with core-top materials because of the complexity of foraminiferal calcification and caveats in individual proxy development. For instance, tow-derived shells without the final stage of calcification have been shown to have higher Mg/Ca values than those in core tops (e.g., Martínez-Botí et al., 2011). In addition, the presence of gametogenic calcite complicates tow-derived foraminiferal δ18O data (Jentzen et al., 2018). Moreover, high productivity may affect shell Mg/Ca composition in sediment trap samples (Gibson et al., 2016). The quantitative relationship between the cadmium partition coefficient and temperature, established in core tops (Rickaby and Elderfield, 1999), does not appear to translate into culture observations (Mashiotta et al., 1997). In our opinion, Winkelbauer et al. (2023) did not fully acknowledge all complexities that should be considered when comparing core-top and plankton tow studies.
Winkelbauer et al. questioned the extent to which seawater iodate is incorporated in planktic foraminifera shells in the core tops, but their iodate data for seawater are spatially and temporally different from the environments sampled in their plankton tows. These spatial and temporal offsets are important because significantly lower iodate concentrations have been reported during seasonal cycles at high latitudes (Chance et al., 2010).
Tow samples (containing living material) and core-top samples are usually preserved/treated differently on board ships before shells are picked. The cleaning method adopted in this study was not validated on potentially more fragile samples from the towed materials, with a potentially unknown impact on the I/Ca signal.
We think that the data from Site 1088 are irrelevant to the discussion. ODP drill cores hardly ever preserve the sediment–water interface because the lowering of the bottom-hole assembly strongly disturbs the sediment/water interface and thus the core top sediments. The analyzed specimens must be considerably older than recently, and the semi-transparent and “white” specimens might not have lived at the same time.
The suggestion that planktic foraminifera might gain iodine post-mortem ‘via abiogenic incorporation shortly after gametogenesis,’ i.e., within the water column during downward transport, is not supported by evidence. The iodine gained in gametogenic calcite is a likely reason for the observed discrepancy. Gametogenic calcite is formed by foraminifera during the late life stages, i.e., at depths around the pycnocline—deep chlorophyll maximum for all species analyzed (Schiebel and Hemleben, 2017).
Winkelbauer et al. alluded to the possibility that the planktic I/Ca signal was mainly generated on the seafloor. This scenario is unlikely, because
1. There are consistently low planktic I/Ca values in Holocene sediments deposited in regions with an oxygen minimum zone above oxygenated bottom waters (Figure 1A, based on the Δδ13C data at the Eastern Equatorial Pacific, Hoogakker et al., 2018; and Benguela, Lu et al., 2020a).
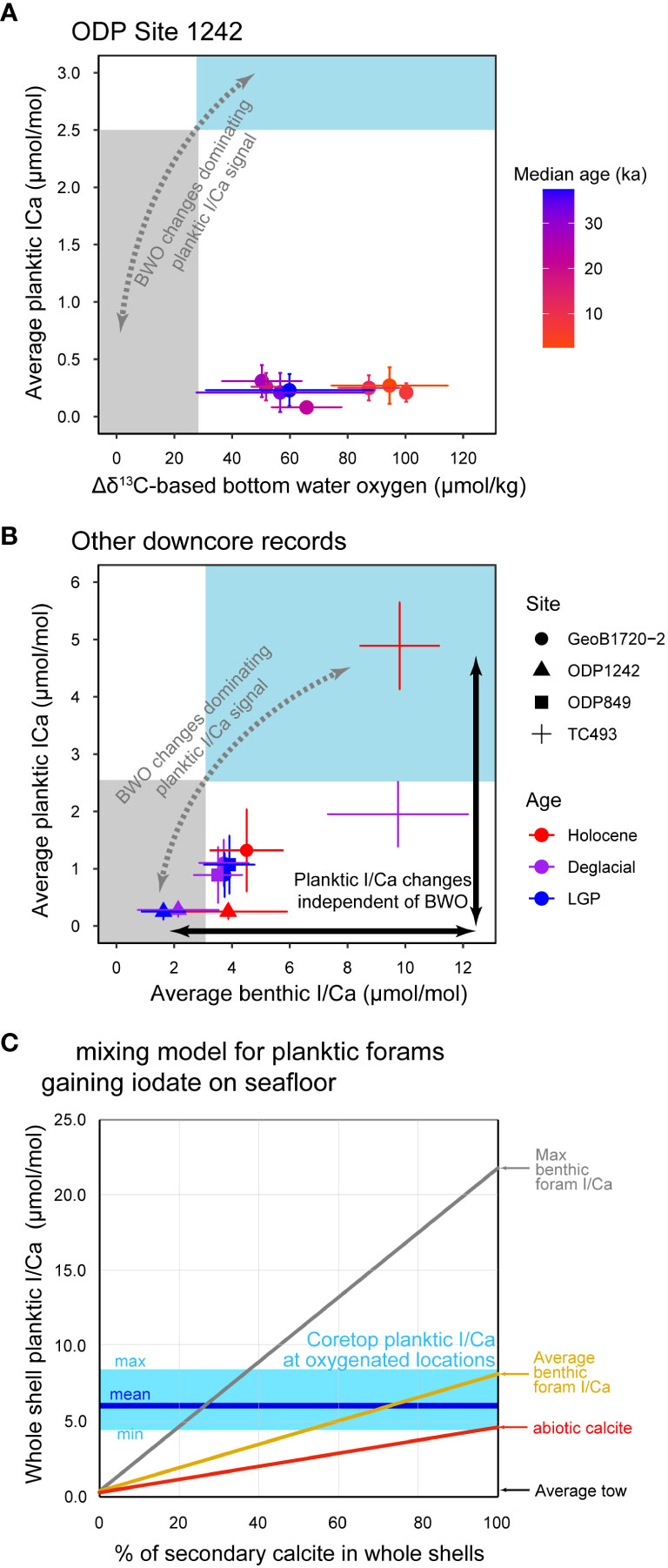
Figure 1 (A) Comparison of planktic I/Ca with Δδ13C-based bottom water oxygen at ODP Site 1242 over the last 40 ka (Hoogakker et al., 2018). Colored boxes indicate the expected I/Ca signal under low (gray) vs. high (blue) oxygen conditions. (B) Comparison of down-core planktic vs. benthic I/Ca at multiple sites (Lu et al., 2020a; Lu et al., 2020b). (C) Endmember mixing calculations for percentages of secondary calcite must be contained in core-top planktic foraminifera if most of their I/Ca signals are gained on the seafloor. The abiotic calcite endmember assumes a partition coefficient Kd (Kd = 10, [I/Ca]/[iodate] with units of [µmol/mol]/[µmol/L] and a normal seawater iodate concentration of 0.45 µmol/L. Benthic foraminiferal core-top I/Ca values from oxygenated bottom waters were used for additional endmember mixing scenarios (Lu et al., 2022).
2. If I/Ca signals in planktic foraminiferal tests were generated on the seafloor, trends in such data should mimic trends in benthic I/Ca and/or other BWO proxies (e.g., Δδ13C). In contrast, the temporal trends of planktic I/Ca are independent of bottom water conditions, as represented by benthic I/Ca and Δδ13C (Figures 1A, B) (Hoogakker et al., 2018; Lu et al., 2020b).
3. It has been observed that the diagenesis of marine carbonate decreases rather than increases the primary I/Ca signal (Hardisty et al., 2017). Adding abiotic calcite precipitated from seawater with an average iodate concentration mathematically cannot explain the observed core-top I/Ca, if the primary I/Ca was indeed as low as reported in the tow samples (Figure 1C). Even when adopting other endmembers (e.g., higher I/Ca measured in core-top benthic foraminifera), core-top shells from oxygenated waters would likely contain >50% secondary calcite, and such heavily crusted shells would have been easily recognized under the microscope and excluded from core-top measurements (Lu et al., 2020a).
Utilizing the chemistry of carbonates as paleoenvironmental proxies benefits from studying materials obtained using as many modes of sample collection as possible. Nonetheless, given that the objective is to understand the sedimentary record, the caveats and differences of each calibration approach must be fully acknowledged and integrated so that the opportunities and limitations of promising proxies, including I/Ca, can be more completely evaluated.
ZL, ET, and RR developed the ideas and wrote the manuscript with assistance from WL and AP. All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
We thank two reviewers for their input. ZL is supported by NSF EAR 2121445.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Bianchi D., Weber T. S., Kiko R., Deutsch C. (2018). Global niche of marine anaerobic metabolisms expanded by particle microenvironments. Nat. Geosci. 11 (4), 263–268. doi: 10.1038/s41561-018-0081-0
Chance R., Weston K., Baker A. R., Hughes C., Malin G., Carpenter L., et al. (2010). Seasonal and interannual variation of dissolved iodine speciation at a coastal Antarctic site. Mar. Chem. 118 (3-4), 171–181. doi: 10.1016/j.marchem.2009.11.009
Gibson K. A., Thunell R. C., Machain-Castillo M. L., Fehrenbacher J., Spero H. J., Wejnert K., et al. (2016). Evaluating controls on planktonic foraminiferal geochemistry in the Eastern tropical north pacific. Earth Planet. Sci. Lett. 452, 90–103. doi: 10.1016/j.epsl.2016.07.039
Hardisty D. S., Lu Z., Bekker A., Diamond C. W., Gill B. C., Jiang G., et al. (2017). Perspectives on proterozoic surface ocean redox from iodine contents in ancient and recent carbonate. Earth Planet. Sci. Lett. 463, 159–170. doi: 10.1016/j.epsl.2017.01.03
Hoogakker B. A., Lu Z., Umling N., Jones L., Zhou X., Rickaby R. E., et al. (2018). Glacial expansion of oxygen-depleted seawater in the eastern tropical pacific. Nature 562 (7727), 410–413. doi: 10.1038/s41586-018-0589-x
Jentzen A., Nürnberg D., Hathorne E. C., Schönfeld J. (2018). Mg/Ca and δ 18 O in living planktic foraminifers from the Caribbean, gulf of Mexico and Florida straits. Biogeosciences 15 (23), 7077–7095. doi: 10.5194/bg-15-7077-2018
Jonkers L., Reynolds C. E., Richey J., Hall I. R. (2015). Lunar periodicity in the shell flux of planktonic foraminifera in the gulf of Mexico. Biogeosciences 12, 3061–3070. doi: 10.5194/bg-12-3061-2015
Lu W., Dickson A. J., Thomas E., Rickaby R. E., Chapman P., Lu Z. (2020a). Refining the planktic foraminiferal I/Ca proxy: results from the southeast Atlantic ocean. Geochimica Cosmochimica Acta 287, 318–327. doi: 10.1016/j.gca.2019.10.025
Lu W., Rickaby R. E., Hoogakker B. A., Rathburn A. E., Burkett A. M., Dickson A. J., et al. (2020b). I/Ca in epifaunal benthic foraminifera: a semi-quantitative proxy for bottom water oxygen in a multi-proxy compilation for glacial ocean deoxygenation. Earth Planet. Sci. Lett. 533, 116055. doi: 10.1016/j.epsl.2019.116055
Lu W., Wang Y., Oppo D. W., Nielsen S. G., Costa K. M. (2022). Comparing paleo-oxygenation proxies (benthic foraminiferal surface porosity, I/Ca, authigenic uranium) on modern sediments and the glacial Arabian Sea. Geochimica Cosmochimica Acta 331, 69–85. doi: 10.1016/j.gca.2022.06.001
Martínez-Botí M. A., Mortyn P. G., Schmidt D. N., Vance D., Field D. B. (2011). Mg/Ca in foraminifera from plankton tows: evaluation of proxy controls and comparison with core tops. Earth Planet. Sci. Lett. 307 (1-2), 113–125. doi: 10.1016/j.epsl.2011.04.019
Mashiotta T. A., Lea D. W., Spero H. J. (1997). Experimental determination of cadmium uptake in shells of the planktonic foraminifera orbulina universa and globigerina bulloides: implications for surface water paleoreconstructions. Geochimica Cosmochimica Acta 61 (19), 4053–4065. doi: 10.1016/S0016-7037(97)00206-8
Rickaby R. E. M., Elderfield H. (1999). Planktonic foraminiferal Cd/Ca: paleonutrients or paleotemperature? Paleoceanography 14 (3), 293–303. doi: 10.1029/1999PA900007
Schiebel R., Hemleben C. (2017). “Chapter 5 reproduction, & 6 ontogeny and test architecture,” in Planktic foraminifers in the modern ocean. (Springer) doi: 10.1007/978-3-662-50297-6
Keywords: I/Ca, foraminifera, core top, plankton tow, oxygen
Citation: Lu Z, Thomas E, Rickaby REM, Lu W and Prow AN (2023) Commentary: Planktic foraminifera iodine/calcium ratios from plankton tows. Front. Mar. Sci. 10:1221835. doi: 10.3389/fmars.2023.1221835
Received: 13 May 2023; Accepted: 26 June 2023;
Published: 11 July 2023.
Edited by:
Khan M. G. Mostofa, Tianjin University, ChinaReviewed by:
Rahul Mohan, National Centre for Polar and Ocean Research (NCPOR), IndiaCopyright © 2023 Lu, Thomas, Rickaby, Lu and Prow. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zunli Lu, enVubGlsdUBzeXIuZWR1
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.