 Evan Tuohy
Evan Tuohy Michelle Schärer-Umpierre
Michelle Schärer-Umpierre Louis Penrod3
Louis Penrod3 Richard Appeldoorn
Richard Appeldoorn- 1Department of Marine Science, University of Puerto Rico, Mayagüez, Puerto Rico
- 2HJR Reefscaping, Boquerón, Puerto Rico
- 3Department of Ocean Engineering and Marine Sciences, Florida Institute of Technology, Melbourne, FL, United States
Understanding animal space use patterns is critical for ecological research and conservation efforts. An organism’s home range territory serves as the fundamental unit of space use and is the area repeatedly used for routine activities. Nassau grouper, Epinephelus striatus, once of high commercial fishery importance, are now on the IUCN’s Red List designated as Critically Endangered due to overexploitation. Known for the formation of large spawning aggregations, information on their movements and space use dynamics outside of the reproductive period are lacking. In this study, we used acoustic telemetry to quantify the non-spawning horizontal and vertical space use patterns of Nassau grouper at the Bajo de Sico seamount, a seasonally closed marine protected area (MPA) in Puerto Rico. Twenty-nine groupers were tagged with acoustic transmitters, of which, fourteen were tracked continuously over a three-year period. Tagged individuals displayed high site fidelity to home reef locations and individuals occupied home ranges of relatively small size that remained constant between years. There was a high degree of overlap of home range territories and in locations where multiple individuals occupied the same home reef, individuals maintained discrete vertical distributions. Nassau groupers were recorded making multiple forays to the spawning site outside of the reported spawning season, and two individuals underwent home reef site relocation. Results indicate relatively low contribution to ecological connectivity during the non-reproductive period, therefore high relative abundances of this species are needed to fulfill their ecological role at the community level. The results also highlight the importance of Bajo de Sico, an isolated seamount of relatively deeper depth, as critical habitat for primary home reef sites of Nassau grouper. These results provide a basis for adapting the current management strategy at this MPA to provide adequate protection to the non-spawning population of Nassau grouper.
1 Introduction
Understanding animal space use patterns is a critical component for ecological research and the development of conservation efforts for threatened species (Horne et al., 2008). An animal’s home range serves as the fundamental unit of space use and is defined as the common area repeatedly used by an individual for routine activities (Burt, 1943; Börger et al., 2008). Although animals that move within home range territories usually exhibit some degree of site fidelity to a particular location (Moffitt et al., 2009), the home range is not static and can change throughout an individual’s life due to ontogeny (Werner and Gilliam, 1984), shifting demands in energetic budgets, habitat selection, predator-prey dynamics, reproduction, competition and ecosystem connectivity (Börger et al., 2008). Defining these space use behaviors is important for understanding fundamental ecological processes underlying the distribution, abundance and community structure of organisms within their environment. Furthermore, defining a species’ home range is critical for the effective design of ecosystem-based management (EBM), which requires a firm understanding of the biology, ecology, and behaviors of target species to fully explore management scenarios (Kramer and Chapman, 1999; Garcia, 2003; Moffitt et al., 2009; Nemeth, 2012).
Groupers (Family Serranidae, Sub-family Epiniphelanae) are medium to large bodied marine piscivores of considerable ecological and commercial importance (Cohen et al., 1990). Considered major predators of the coral reef ecosystem, most groupers are solitary, with high site fidelity to home reef sites. During reproduction, some grouper species can migrate considerable distances to join large aggregations (Cohen et al., 1990; Bolden, 2000; Dahlgren et al., 2016; Stump et al., 2017; Nemeth et al., 2023). Factors such as slow growth, late age at maturity and the tendency to form spatially and temporally predictable spawning aggregations contribute to groupers’ susceptibility to overfishing (Reynolds et al., 2005; Aguilar-Perera, 2006; Sadovy de Mitcheson et al., 2008; Cheung et al., 2013; Sadovy de Mitcheson et al., 2013; Grüss and Robinson, 2015). This susceptibility is best exemplified by the Nassau grouper, Epinephelus striatus (Bloch, 1792), a medium-bodied grouper species found throughout the wider Caribbean. Once considered an icon of the Caribbean as the most commercially important reef fish, Nassau grouper are now severely overfished (Sadovy and Eklund, 1999; Sala et al., 2001; Aguilar-Perera, 2006; Sadovy de Mitcheson et al., 2008; Stallings, 2009; Cheung et al., 2013; Sadovy de Mitcheson et al., 2013; Waterhouse et al., 2020), listed by the International Union for Conservation of Nature (IUCN) as Critically Endangered (Sadovy et al., 2018), and Threatened under the U.S. Endangered Species Act (NOAA, 2016). The Nassau grouper is well-known for historically forming large aggregations, once numbering in thousands to tens-of-thousands; however, information on their social behavior outside of reproductive periods is lacking due to reduced population sizes available for research (Sadovy de Mitcheson et al., 2008; Sadovy de Mitcheson et al., 2012).
Although Nassau grouper have complex sexual and social systems related to spawning, individuals are relatively sedentary and solitary outside of these reproductive periods (Sadovy de Mitcheson et al., 2012). Nassau groupers exhibit high site fidelity and annual residency, returning to the same home reef location each year after spawning (Colin, 1992; Starr et al., 2007; Sadovy de Mitcheson et al., 2012; Blincow et al., 2020). Multiple individuals have been reported to reside within the same reef site, occupying home ranges of several hundred square meters, often with some degree of overlap (Colin, 1992; Bolden, 2001; Keller et al., 2020). In cases where home ranges overlapped, individuals preferentially occupied smaller areas within their home range, with social hierarchy maintained through agonistic or territorial displays as fish compete for the dominant role within a reef site (Colin, 1992; Sadovy and Eklund, 1999). Given that Nassau groupers have the capability of existing within a wide depth range – in some instances occupying depths of >250 m for two months following spawning (Starr et al., 2007) – it has been shown that smaller home range territories are stratified by depth, presumably to reduce or eliminate the degree of overlap among individuals within the same locations (Blincow et al., 2020).
The previous works, however, were limited in scope in that they were either observational, largely focused on reproduction, of small sample sizes, or could not calculate traditional space use metrics. Therefore, a comprehensive multi-year analysis of considerable sample sizes is needed to quantify non-spawning home range sizes and space use metrics. This study focused on the home range space utilization of Nassau grouper during non-spawning times. The purpose of the study was to 1) quantify home range (e.g. site fidelity) and 2) space use patterns in reefs over multiple years of Nassau grouper during non-spawning times as determined by acoustic telemetry off the west coast of Puerto Rico. Of added significance, these fish frequent the only known Nassau grouper spawning aggregation in Puerto Rico, also located within the study area (Schärer et al., 2012; Schärer-Umpierre et al., 2014; Tuohy et al., 2016). This study provides a better understanding of the spatial ecology of this once prevalent species and its contribution to the ecology of coral reef communities, while highlighting important implications for Nassau grouper conservation and marine protected area (MPA) design.
2 Materials and methods
2.1 Study area
Bajo de Sico (BDS), an isolated seamount 27 km of the west coast of Puerto Rico (Figure 1), is characterized by a ridge of highly rugose rock promontories ranging in depths from 25 to 50 m deep. It rises from a mostly flat, gradually sloping shelf covered in rhodolith habitat that extends to a depth of 100 m and encompasses an area of 11.1 km2. Beyond this depth, the shelf slopes deeply into a vertical wall that reaches depths of 200–300 m to the southeast and over 1,000 m to the north. The dominant oceanographic features and its location within the Mona Passage make this area subject to periods of strong (> 2.0 kts), persistent northerly currents (García-Sais et al., 2007; García-Sais et al., 2020).
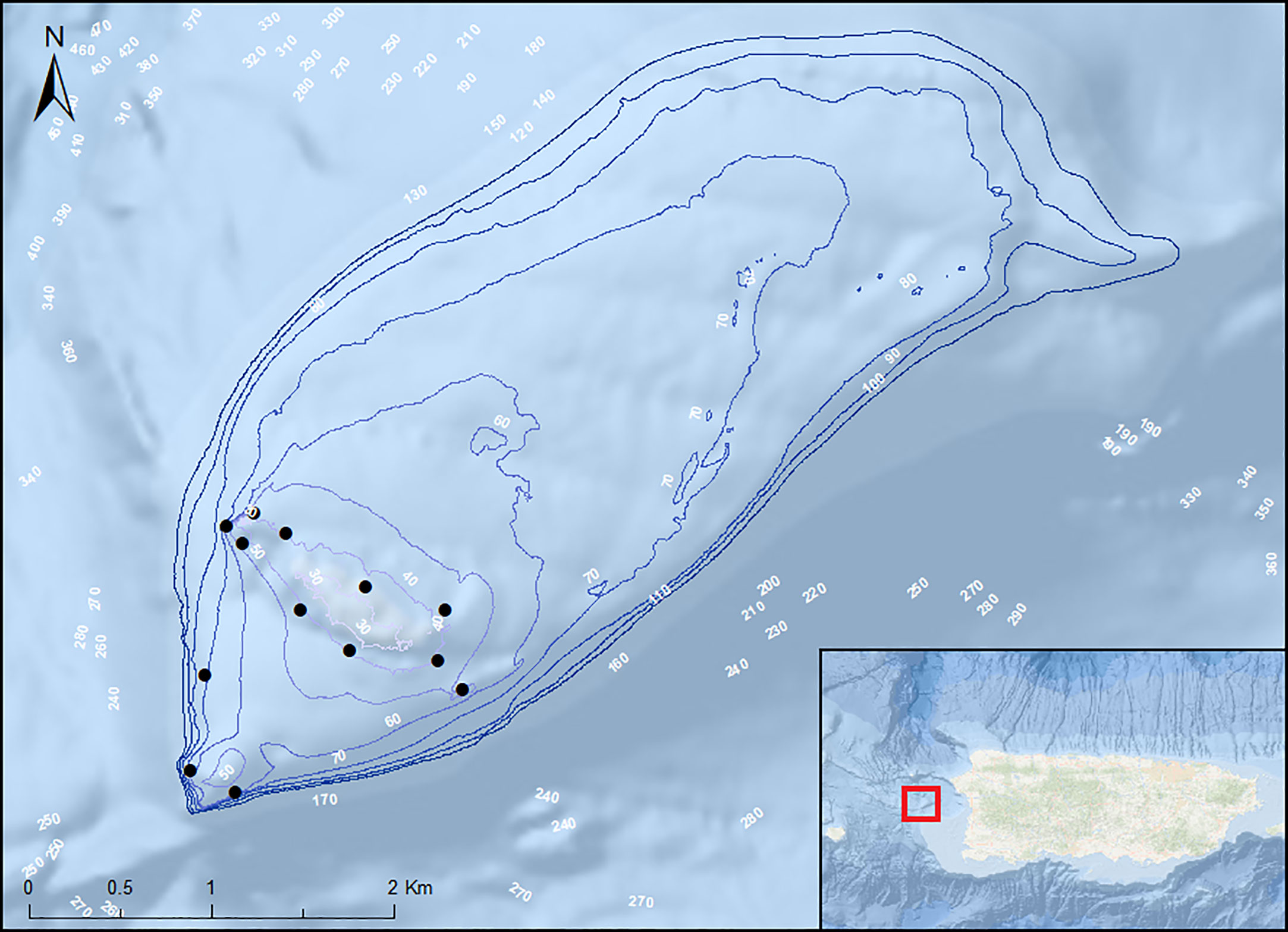
Figure 1 Map of the acoustic array at Bajo de Sico and northwest Puerto Rico shelf. Depth contour at Bajo de Sico in 10 m increments (30-100 m) and black circles indicate relative location of acoustic receivers with receivers in proximity represented by a single circle.
Bajo de Sico is characterized by highly diverse and taxonomically complex benthic and pelagic fish assemblages (García-Sais et al., 2007; García-Sais et al., 2020). Benthic habitats are characterized by colonized pavement reef top (at 25 m) and vertical wall associated with reef promontories (35-50 m), followed by colonized and uncolonized gravel and rhodolith dominated habitats (> 50 m). The sessile-benthic community is comprised primary of benthic algae (55%), sponges (33%) and scleractinain corals (7%; García-Sais et al., 2020). Bajo de Sico is known to harbor multi-species spawning aggregation sites including the only one documented of Nassau grouper in the waters surrounding Puerto Rico (Sadovy, 1994; Schärer et al., 2012; Sanchez et al., 2017). The spawning season for Nassau grouper at this location occurs during the calendar months of January through April, which corresponds to the January-March moons (Tuohy et al., 2016). In the exclusive economic zone (EEZ) waters of BDS there is a seasonal, six-months closure for catching reef fishes from October 1 to March 31 to protect coral reef habitats and fish aggregated to spawn. Management strategies were first implemented to protect the spawning populations of red hind (Epinephelus guttatus) during three months (December 1 through February 28/29), and later expanded to six (October 1 through March 31) to protect the spawning stock of additional species of snappers and groupers, and coral reef habitat (Crabtree, 2009). Although the possession of and fishing for managed reef fish species are prohibited during the six months’ seasonal closure, the regulations allow fishing, scuba diving, and fishing for highly migratory species (HMS) and lobster throughout the entire year. Bottom tending gears such as traps, nets and longlines are prohibited year-round. Despite management regulations, monitoring efforts have indicated low population densities of Nassau grouper at BDS (García-Sais et al., 2007; Schärer-Umpierre et al., 2014; García-Sais et al., 2020).
2.2 Fish tagging
Acoustic tagging of Nassau groupers was conducted at the spawning aggregation site on BDS during the 2013 and 2014 reproductive seasons. Due to the increased depths at which this aggregation occurs (> 40 m) an in situ approach using closed-circuit rebreathers (CCR) was developed in order to decrease barotrauma impacts of tagged individuals and increase the tagging efficiency (Tuohy et al., 2015). Nassau groupers were captured using Antillean arrowhead fish traps (n=4) baited daily with squid and canned cat food. Technical CCR divers descended to the traps and removed the individuals selected for tagging by coercing them into a mesh catch bag modified to include a 30-cm zipper. Divers restrained the fish in an inverted position, inducing a tonic immobility state, and the zipper was positioned over the abdomen to provide access to the incision site. Transmitters (V16-4h and V16p-4h, 69 Hz; dimensions= 16 mm diameter X 54 mm length, 19g weight in air; nominal delay = 60-90 sec; battery life = 1250-1580 days; Innovasea (formerly Vemco), Halifax, NS, Canada) were inserted into the body cavity via a 2cm mid-ventral incision made approximately 5 cm posterior to the pelvic girdle. The incision was closed with 2-3 stainless steel surgical stables (Reflex one skin stapler with 5.7 mm staples). Following surgical procedure, total length (TL) measurements and a fin clip were taken and an external tag (Hallprint T-bar anchor tag type TBA; 7cm) was inserted into the musculature near the base of the dorsal fin. The duration of each procedure lasted 12 ± 2.2 min, after which tagged individuals were released at the site and depth of original capture. Each fish was subsequently observed by divers to ensure that there were no immediate adverse effects following surgical tagging (Tuohy et al., 2015).
2.3 Acoustic receiver array
Tagged Nassau grouper were monitored with an array of 16 omni-directional acoustic receivers (VR2 and VR2W, 69 kHz; Vemco) deployed around the study site at depths ranging from 30-75 m (mean ± SD = 48.05 ± 12.16) (Figure 1). Given the increased depth, limited number of receivers, and adverse weather conditions at BDS, it was not possible to deploy or maintain a larger array that encompassed the entire seamount. Therefore, the array focused on areas in and around the main promontories and areas of high vertical relief that are the preferred habitat for this species (Sadovy and Eklund, 1999). An increased density of receivers (n=4) were placed in an area with high topographic complexity and vertical relief to account for the likelihood of receiver shadow and associated reduction in the probability of detection. Each receiver was deployed on the seafloor moored to a cement base. One receiver was attached to subsurface buoy (hard plastic trawl floats); however, this anchoring system was generally avoided, due to frequent periods of strong currents that caused the receivers to shift in angle and position.
Range testing of acoustic tags was conducted using two different methodologies. First, given the mean depth of receivers (48.05 m) and distance from shore, an in situ range testing at the study site was logistically difficult. Therefore, fine-scale range testing was conducted at an offsite location of similar habitat and shallower (20 m) depth. To range test, a VR2W was moored to a cement base at a depth of 20 m, identical to the mooring used at the study site. A tripod with a range test tag (V16-4h; transmission rate = 5 sec; Vemco) attached 50 cm above the substrate was positioned by divers at 30 m increments from the acoustic receiver. The range tag remained in place for a period of 10 minutes before being moved to the next 30 m distance for a total distance of 300 m. Second, to confirm these results, boat-based drift range tests were conducted by suspending the range test tag from a weighted line attached to the vessel. The weighted line was deployed at the location and depth of an acoustic receiver and the trajectory, distance from the acoustic receiver and time of day was tracked with a handheld GPS as the vessel drifted away from the receiver. A total of 15 drifts were conducted from one receiver during periods of minimal current velocity so that the weighted line remained perpendicular to the surface under the vessel as it drifted. A 70% probability of detection corresponding to distance of 150 m was used for all further analyses (Farmer et al., 2013).
2.4 Data analysis
Non-spawning space use characteristics were calculated for each tagged grouper that resided within the array during years 2014 to 2016. Detection data from years 2013 and 2017 were not included in the analysis due to low sample sizes. Non-spawning months consisted of April-December (2014) and May-December (2015 and 16) with differences related to variation in the observed spawning periodicity of those years (Tuohy et al., 2016). Acoustic receivers were retrieved at intervals of six months to one year, depending on the location. Data were downloaded using VUE software (Vemco) and directly imported into R (R Core Team, 2018). Prior to analysis, data were filtered to remove false or spurious detections defined as single detections within a 24-hour period (Aspillaga et al., 2018; Keller et al., 2020). These detections may occur when two tags emit a signal simultaneously and collide within the detection range of a receiver. Additionally, tag detection records were considered unreliable when a stationary characteristic of the signal or irregular depth were observed, indicating tag loss, mortality, or predation, at which point the tag was no longer included in the analysis.
2.4.1 Home range size
Brownian bridge movement models (BBMM) were used to calculate the utilization distribution (UD) for each tagged individual, The BBMM is a temporally explicit method for estimating the UD based on the probability of an individual being at a location along the movement path conditional to the distance and elapsed time between successive locations. An animal’s UD is defined as the probability distribution representing the total area of space use (Horne et al., 2007). The model not only accounts for the spatial distribution between locations, but also considers their time dependency and assumes a conditional random walk movement pattern between pairs of locations dependent on step length and time interval between successive steps (Horne et al., 2007). Brownian bridge UD estimates also consider the error associated with a given position, which is represented by the estimated detection radius of the acoustic receivers (Horne et al., 2007). The location error used for the models was based on the 70% probability of detection at 150 m and the time step 720 minutes.
An individual’s core area (CA) was defined as the minimum area encompassing the 50% UD volume, and the home range (HR) area was calculated as the minimum area encompassing the 95% UD volume, with CA and HR considered to be the area where individuals spend up to 50% and 95% of their time, respectively. For each tagged individual, the annual non-spawning Brownian bridge HR estimates were calculated for the entire tracking period using the “adehabitatHR” package in R (Calenge, 2006). Kruskall-Wallis H tests were used to test for significant differences in habitat use (CA and HR) between years. The relationship between space use area and fish total length (TL) were tested using a linear regression. To test whether there was an array effect, given the uneven distribution of receivers within the array, differences in home range estimates between individuals with home ranges in areas of high receiver spatial coverages and individuals with home ranges in areas of low receiver spatial coverage were tested using a Wilcoxon rank-sum test.
2.4.2 Vertical distribution
For resident tags that contained pressure sensors (n= 8, V16p-4h), depth measurements were binned into daily and diel periods. Mean depth +/- SE were calculated for the entire non-spawning period and differences in the monthly, daily, and diurnal vertical distributions were tested using Wilcoxon rank-sum test. Additionally, differences in the vertical distribution of fishes that reside within the area of greater receiver density (n=4) were tested with a Kruskal-Wallis H test and a Dunn’s pairwise test to detect differences between individuals.
3 Results
A total of 29 Nassau groupers were tagged at the main spawning area during the 2013 (n=10) and 2014 (n=19) spawning seasons, but only 14 tags were detected throughout the non-spawning season and are included in this analysis (Table 1). The remaining tags were only detected during the spawning season and were removed from this analysis. The 14 individuals ranged in size from 52 to 80 cm TL and were detected for periods ranging from 508 to 1,580 days (mean ± SD = 1192.57 ± 332.63). The total number of detections recorded during non-spawning months was 554,325 with a mean ± SD of 13,520 ± 14,513 (range = 778-77,885) per individual. Three of the 14 tags (21%) were detected for a period greater than four years, seven (50%) were detected between three and four years and four tags (29%) were detected between two and three years (Table 1). The remaining 15 tags were only detected during the spawning season, seven of which were detected for the duration of the study and assumed to reside on BDS, but outside of the detection limits of the array.
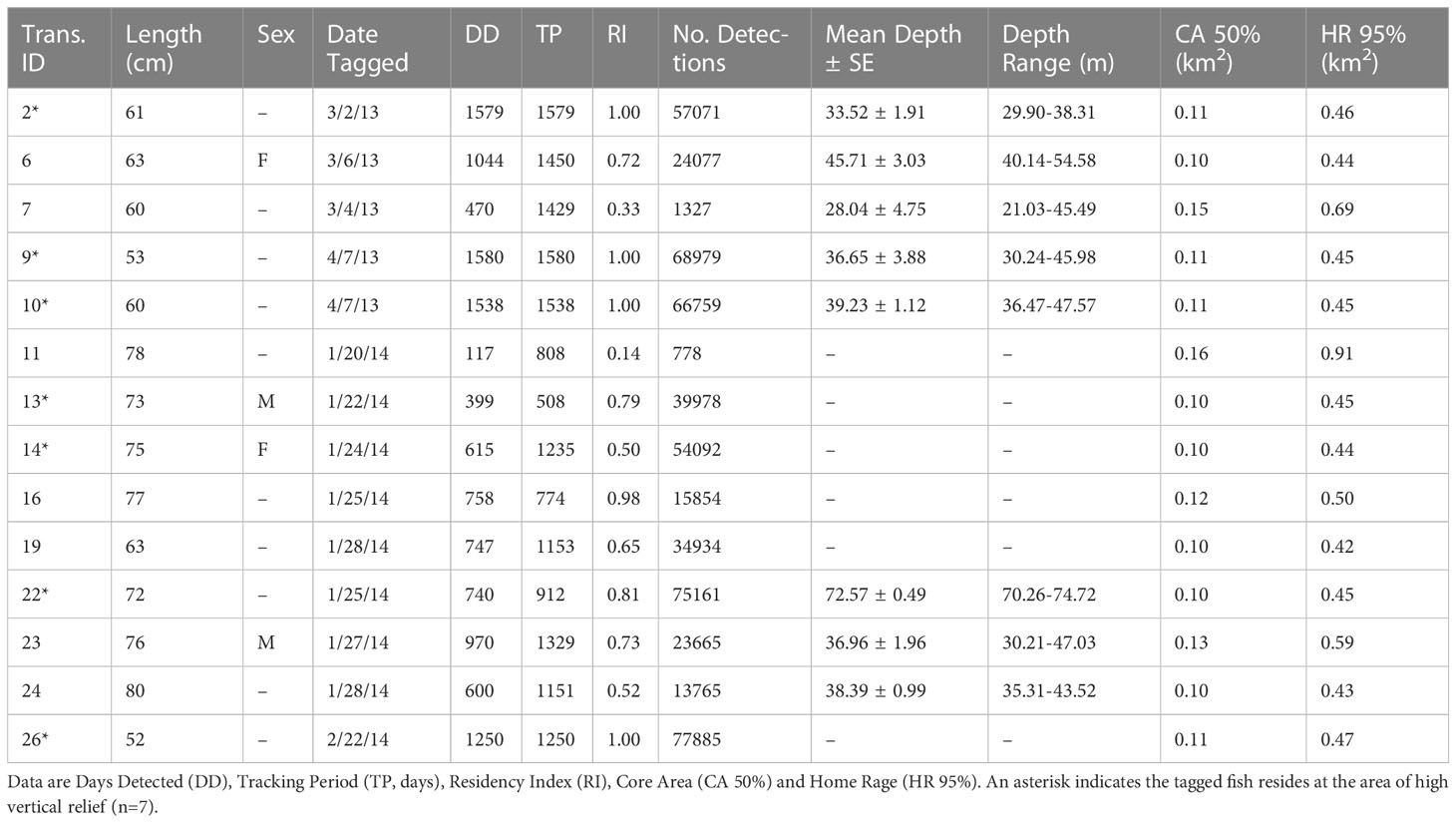
Table 1 Summary of acoustic telemetry data from tagged Epinephelus striatus.
Following each spawning season each fish returned to the same home reef site location. Distances from the fish spawning aggregation (FSA) site to home locations ranged from 0.38 to 1.40 km (mean ± SD = 1.01 ± 0.41 km). Seven of the 14 tagged fish (50%) resided at the area of high topographical complexity and increased receiver density for the entirety of the non-spawning months, with their CA and HR utilization distributions occurring within the array. Some tagged fish occupied HR at or within areas immediately adjacent to the main spawning site with the remaining individuals occupying areas located elsewhere within the array. No tagged fish were recorded to occupy home ranges on a deeper, smaller promontory located at the southern portion of the seamount (Figure 1).
3.1 Home range size and location within acoustic array
The annual non-spawning CA (50% UD) calculated by the BBMM ranged from 0.10 to 0.16 km2 with a mean ± SD of 0.11 ± 0.01 km2. The annual HR (95% UD) ranged from 0.34 to 0.91 km2 with a mean ± SD of 0.48 ± 0.06 km2. For each fish there was no significant difference between the yearly annual CA (χ2(2) = 1.58, p > 0.05) and HR (χ2(2) = 2.09, p > 0.05) utilization distributions. The total combined non-spawning CA and HR for all tagged individuals residing within the array was 0.87 km2 and 2.55 km2 (Table 1; Figure 2). Linear regressions showed no significant relationship between CA (F [1,38]= 0.64, p=0.43, r2=-0.01) and HR (F [1,38]= 1,90, p=0.18, r2 = 0.02) and fish length. All individuals returned to the same home site after each spawning period.
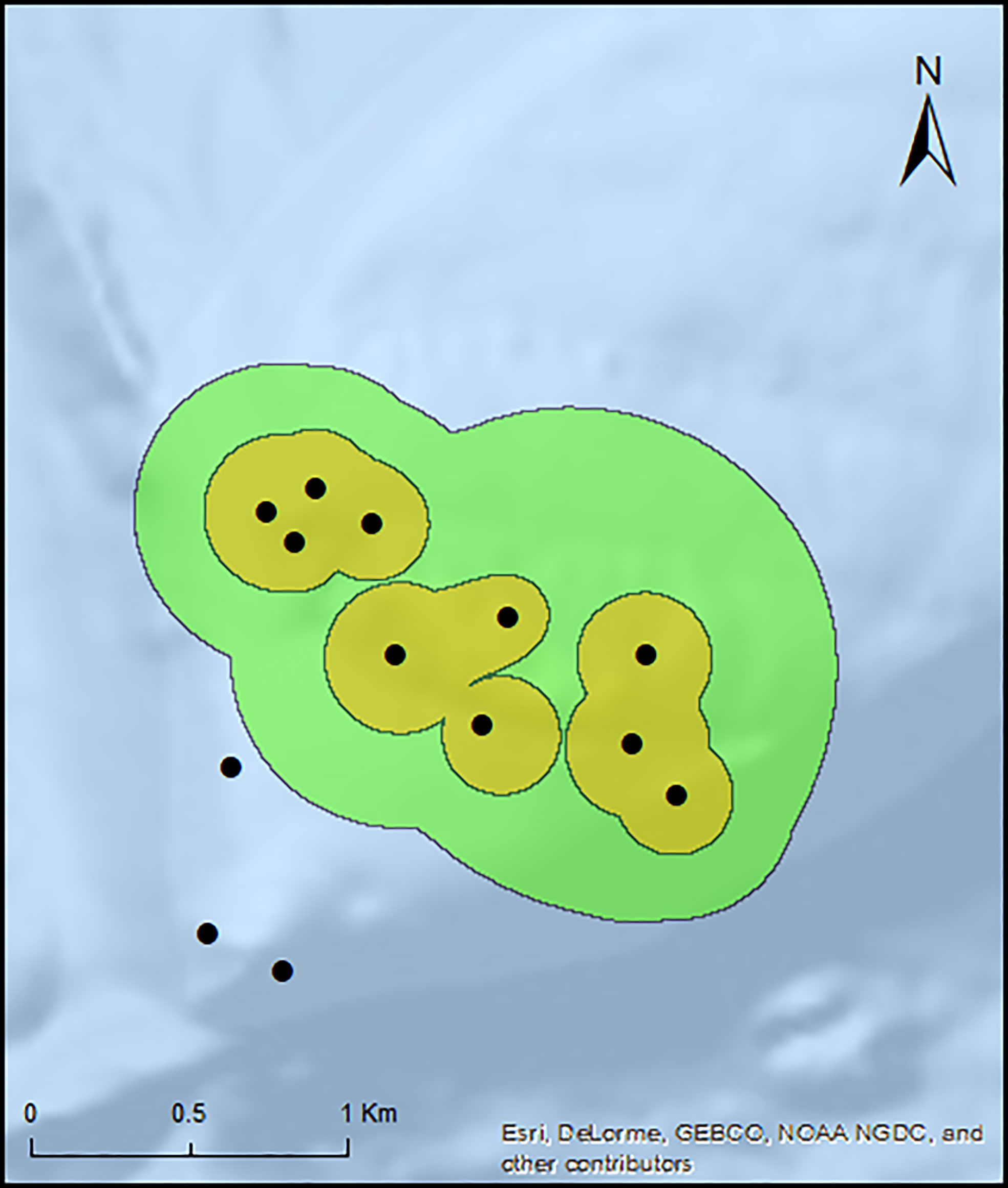
Figure 2 Total non-spawning habitat use for all resident E. striatus tagged within the array (n=14) for the three non-spawning periods. Orange polygon indicates the Core Area (CA 50%) utilization distribution (UD) and green polygon indicates Home range (HR 95%) UD. Black circles indicate relative location of acoustic receivers with receivers in proximity represented by a single circle.
Total space use occupied by all individuals that resided at the location of high receiver density (n=7) was 0.18 km2 (CA) and 0.60 km2 (HR) with overlapping of 50% and 95% UD for all individuals. Individual CA for fish that resided at this location during non-spawning periods ranged from 0.1 to 0.11 km2 (mean ± SD = 0.1 ± 0.003 km2) and the HR ranged from 0.4 to 0.46 km2 (mean ± SD = 0.45 ± 0.01 km2) (Figure 3). Whereas the total space use occupied by groupers that resided at the locations with low receiver density (n=7) was 0.87 km2 (CA) and 2.55 km2 (HR). Groupers CA for fish that resided at this location during non-spawning periods ranged from 0.1 to 0.16 km2 (mean ± SD = 0.12 ± 0.02 km2) and the HR ranged from 0.42 to 0.91 km2 (mean ± SD = 0.57 ± 0.18 km2). A Wilcoxon rank-sum test indicated no significant differences between annual CA (W(2) = 34.0, p > 0.05) and HR (W(2) = 36.0, p > 0.05) utilization distributions between tagged individuals that reside at the area of high and low receiver density, indicating no differences in the home range estimates. Almost all CA estimates for the tagged individuals outside the high complexity site encompassed the same one or two receivers within the array for the duration of the tracking period. Although the receivers characterizing the CAs and HRs of different fish were distributed throughout the array, six of seven tagged fish (86%; IDs # 006, 007, 011, 019, 023, and 024) had overlapping core areas (Figure 4).
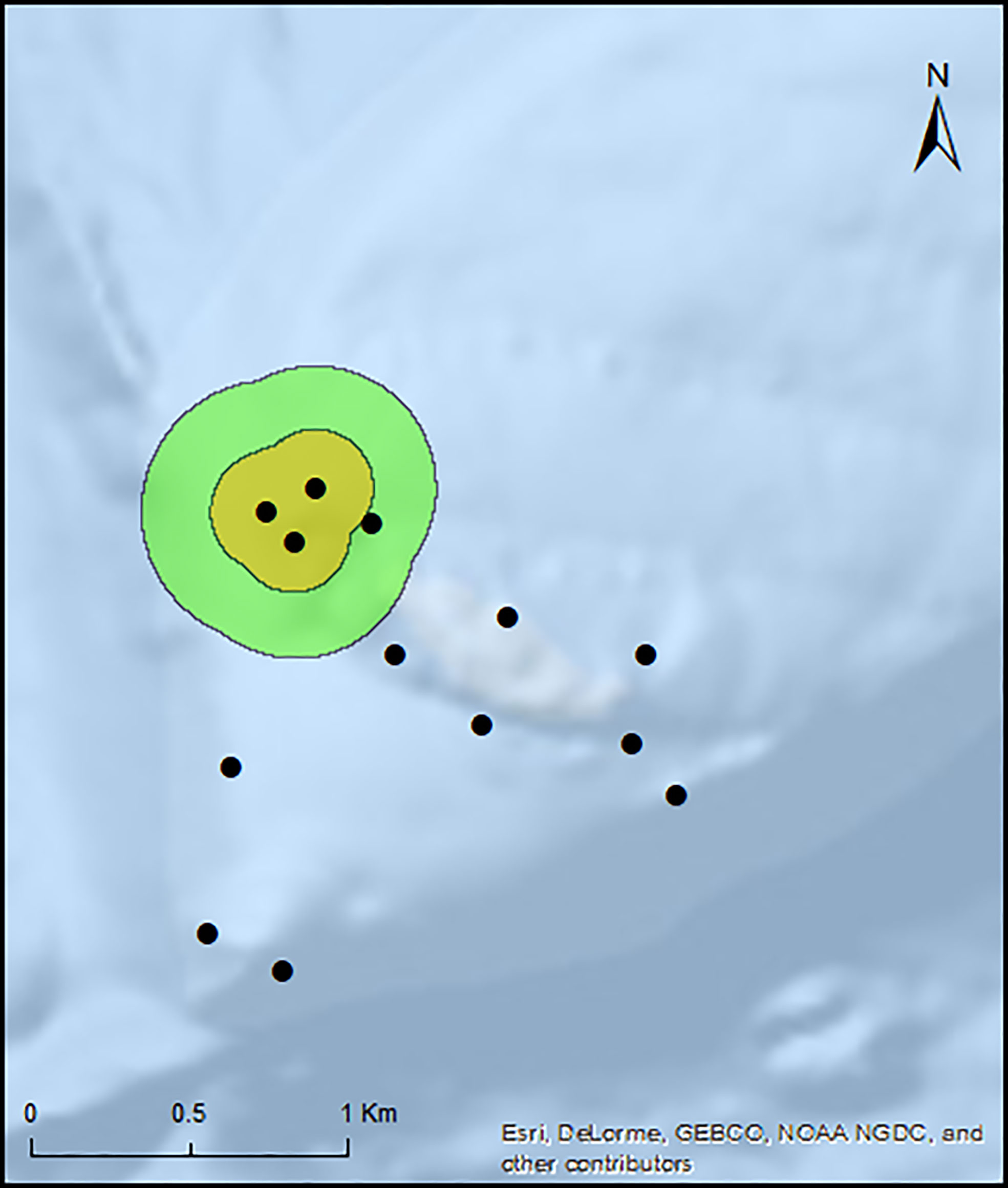
Figure 3 Total non-spawning habitat use for all tagged E. striatus (n=7) residing at the area of high vertical relief for the three non-spawning periods. Orange polygon indicates the Core Area (CA 50%) utilization distribution (UD), and green polygon indicates Home Range (HR 95%) UD. Black circles indicate relative location of acoustic receivers with receivers in proximity represented by a single circle.
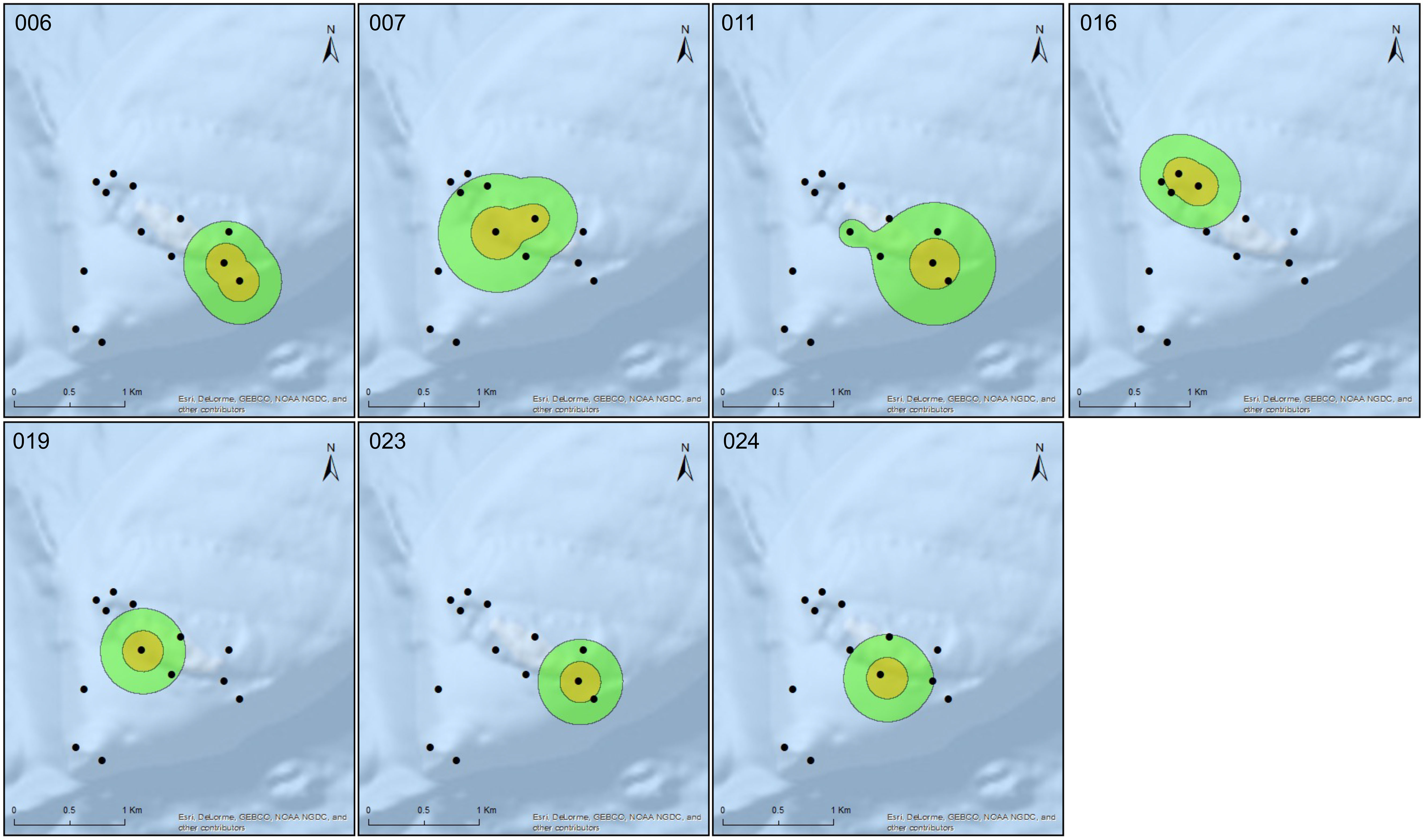
Figure 4 Total non-spawning habitat use for all tagged E. striatus residing outside the area of high relief within the array for the three non-spawning periods. Orange polygon indicates the Core Area (CA 50%) utilization distribution (UD), and green polygon indicates Home Range (HR 95%) UD. Transmitter ID indicated in the upper left-hand corner, and black circles indicate relative location of acoustic receivers with receivers in proximity represented by a single circle.
3.2 Forays and home reef relocation
Three groupers (IDs # 011, 023 and 007) made forays between their home site and the spawning area during non-spawning periods that corresponded to the spawning lunar periodicity observed at this location. In April 2014, grouper #023 migrated seven days after the full moon (DAFM) and remained at the FSA site for 20.5 hrs before returning to the same home site. In May 2014, grouper #011 migrated in a similar way six DAFM and remained at the FSA site for 22.5 hrs before returning. This behavior was not observed in 2015, however it resumed in 2016, when grouper #023 made two forays to the FSA site at seven and nine DAFM in April and remained there for 19.7 and 13.4 hrs, respectively. During the same moon in April 2016, grouper #007 migrated to the FSA site seven DAFM and remained for a period of 6.3 hrs before returning to its home site. Both groupers #023 and #007 underwent forays to and arrived at the FSA site during the same day, however they arrived at the FSA site more than six hours apart, suggesting that the fish migrated separately.
In 2016, grouper #023 shifted the location of its HR to an area of higher vertical relief. This change occurred over two months, July and August, where the fish migrated 1.4 km (minimum linear distance) to the area of high relief on four separate occasions before it remained there for the duration of the tracking period. Migration times ranged from 12.2 to 45.2 hrs and time spent away from its original home reef site ranged from 44 to 282.8 hrs. This fish was detected at multiple receivers while migrating throughout the array, with movements between locations resulting in a similar pattern of detection. The initial migration to this new site took the longest, and subsequent migration times decreased with each visit before the relocation was completed. The increasingly shorter migration time indicate that the fish was moving faster, or that the migration route was more direct leading up to relocation.
3.3 Vertical distribution
During non-spawning months, tagged individuals resided at depths between 21 and 74.7 m (mean ± SD = 40.2 ± 11.9 m; Table 1 and Figures 5A, D). A Kruskal Wallis H test indicated a statistically significant difference in depth among all individuals (χ2(7) = 161.02, p << 0.05). A Wilcoxon rank sum test showed no significant difference between diel vertical distribution among fish (Daily Median = 37.20 m; Nightly Median = 37.16 m; W(2)= 274392, p =0.599), indicating that depth of tagged fish was relatively constant. A Kruskal-Wallis H test showed that there was a statistically significant difference in the mean depth during the non-spawning period between four residents of the high vertical relief area (χ2(3) =75.56, p < 0.001). A pairwise comparison using Dunn’s tests revealed strong evidence (p < 0.05, adjusted using the Bonferroni correction) of differences in depth between fish at this location, indicating that – although there is a high degree of horizontal overlap in home reef sites – individuals occupy different depth ranges (Figure 5B).
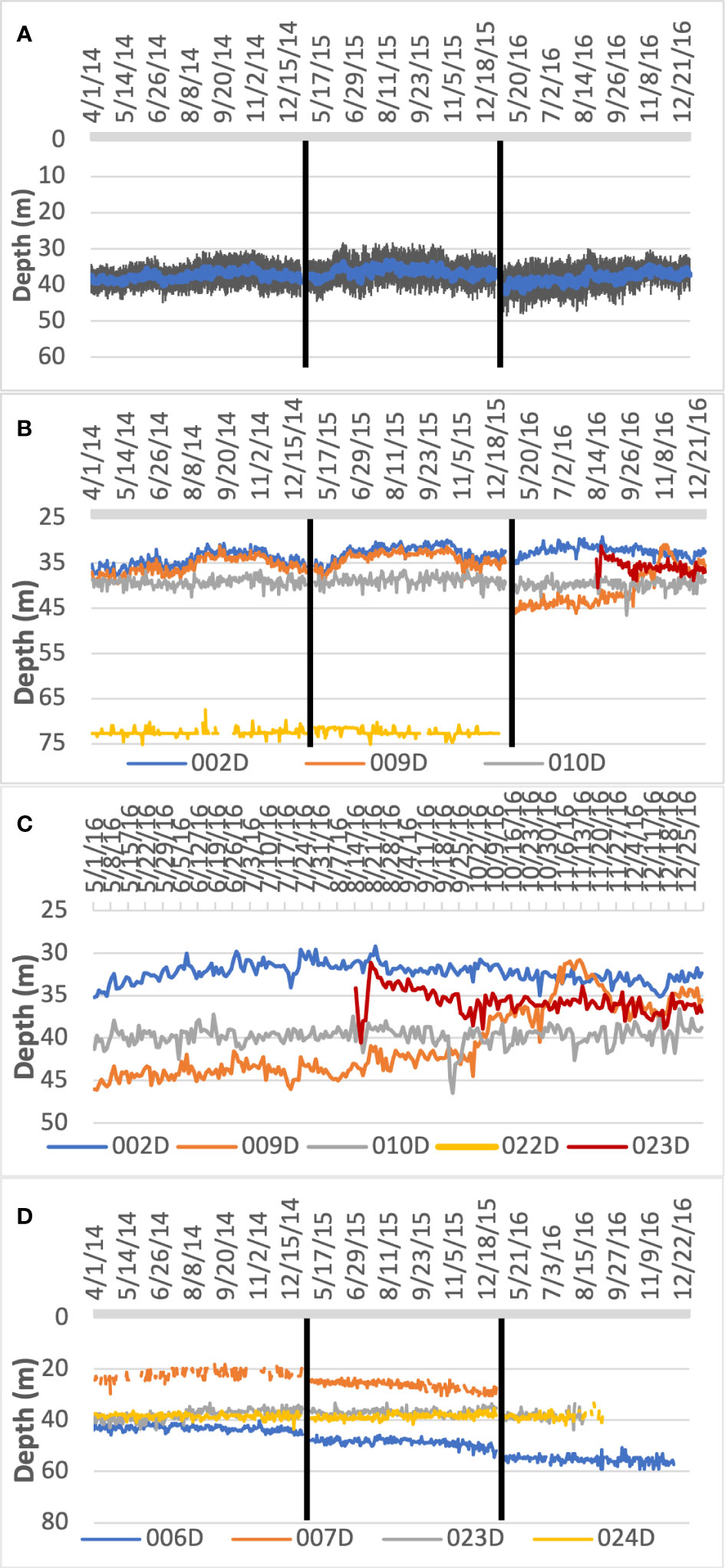
Figure 5 Depth profiles for individuals tagged with pressure sensors for the three non-spawning periods. (A) Daily mean ± SE depth for all individuals tagged with pressure sensors (n=8) for three non-spawning periods. (B) Daily mean depth for tags with pressure sensors residing at the area of increased vertical relief (n= 5) for the entire tracking period. (C) Daily mean depth for tags residing at the area of increased vertical relief habitat for year 2016 (n=4). (D) Daily mean depth for tags with pressure in areas of low complexity (n= 4) for tagged individuals residing at different location within the array.
In 2016, grouper #009 returned to the area of high vertical relief, however in a deeper depth profile (2015 Daily Median= 33.48 m; 2016 Daily Median= 42.07; W(2)= 19976, p < 0.001) that was deeper than the other fish that already resided there. This grouper remained at this depth (43.42 ± 1.32 m) for a period of five months before relocating shallower. However, in August of the same year, when grouper #023 arrived at this location, it was at a depth not occupied by other resident tagged fish at that time, but that was previously occupied by grouper #009 (35.2 ± 1.5 m; Figure 5C). As grouper #009 migrated shallower toward its previously recorded depth, it overlapped in depth with that of fish #023. This movement to shallower habitats occurred over a two-month period and traversed a wide range of depths – including its shallowest recorded - and overlapped with the other individual’s depth profiles within the site (Figure 5C).
4 Discussion
Nassau grouper demonstrated a high degree of residency at Bajo de Sico, with a minimum of 48% of the tagged population (N=29) remaining on the shallowest areas of the seamount throughout the year. However, given the small proportion of BDS covered by this array, the proportion of residents is probably much higher, potentially including all tagged groupers. Consistent with other reports (Starr et al., 2007; Dahlgren et al., 2016; Blincow et al., 2020; Keller et al., 2020), Nassau grouper exhibited high site fidelity to their home reef sites, returning to them after migrating to the nearby spawning site each year. In this study, Nassau grouper were found to occupy relatively small-sized home ranges that remained constant between years, and were similar between individuals, with no relation to fish length although all tagged fish were of adult sizes. Despite the small home range sizes and the limited number of fish tagged, there was a high degree of overlap in home range territories among individuals. Although evidence from this study supports that the location of Nassau grouper home range territories can vary over time (Colin, 1992; Blincow et al., 2020), this study also found that some Nassau grouper underwent home reef site relocation during the three-year tracking period. Tagged individuals occupied significantly different depth ranges throughout the array, and in locations where multiple fish resided at the same home reef, individuals occupied discrete vertical distributions.
In Florida, Nassau grouper home range estimates were larger (core area = 1.58 ± 0.59 km2; home range = 0.78-2.61 km2; Keller et al., 2020) than in the current study. Keller et al.’s (2020) study differs in several important aspects. The home range estimates of Nassau grouper were based on three individuals which were located at a bank reef along a much wider continental shelf with large areas of contiguous habitats. The location and timing of spawning was not known in Florida, but the tracking period encompassed the reported spawning season of the species (Keller et al., 2020). Nassau grouper are transient spawners, migrating long distances to form ephemeral mass spawning aggregations at locations that are outside of their home range (Sadovy and Eklund, 1999; Nemeth, 2012; Kobara et al., 2013; Grüss and Robinson, 2015). Therefore, it is possible that the estimates of Keller et al. (2020) incorporated spawning migrations, which may have inflated the home range estimation. On the other hand, BDS differs in spatial scale and context, as it is a relatively small area of continuous habitat that may explain the reduced home ranges.
Similar to reports from other studies (Colin, 1992; Sadovy and Eklund, 1999; Semmens et al., 2007; Starr et al., 2007; Dahlgren et al., 2016; Blincow et al., 2020; Keller et al., 2020), the tagged fish showed high site fidelity to home reef sites. At BDS all fish migrated back to the same home reef site they occupied prior to the spawning season. The majority (93%) of tagged individuals occupied the same home reef site for the duration of the non-spawning tracking period, and the CA and HR estimates incorporated the same area of a small number of receivers with little to no variation between years.
In 2016, two individuals were recorded to shift to a different home reef site within the array. Following the spawning season, fish #009 returned to the area of high vertical relief and resided at a location deeper than previous years. Later in the year fish #023 also relocated its home reef site to this same location but occupied the then vacant depth range previously occupied by fish #009. In both instances, these individuals relocated to depth ranges not occupied by other tagged fish in the immediate area. The multiple fish residing in this high vertical relief area did so within discrete depth profiles and maintained consistent depth patterns within their home reef site throughout the non-spawning period. However, this was not strictly the case for the two fish that relocated. Both individuals changed depth ranges considerably in the months following their respective relocations (Figures 5B, C). Previous observations have indicated that multiple Nassau grouper can occupy the same home reef site, occurring within a social hierarchy established via competitive interactions among individuals (Colin, 1992; Sadovy and Eklund, 1999). Indeed, fish at BDS also showed a high degree of spatial overlap of home range territories. This may explain this uncharacteristic increase in vertical movement observed over a short period among the two individuals that migrated into new locations. Increased vertical movement would bring these fish into contact with other resident Nassau groupers, thus increasing the opportunity for social interactions and behaviors that led to the establishment of social hierarchies.
Throughout the entire array, depth detection records indicated significant differences in the mean depths among individuals for the duration of the tracking period. Fine scale vertical distributions of tagged individuals showed stationary depth profiles, with no significant differences in diel patterns of vertical distribution, with individuals spending most of the time at 30-40 m. The latter may reflect the biased depth distribution of the receivers, which were limited to the main promontory and shallower portions of BDS. Nevertheless, depth detection data from receivers with detection ranges spanning into deeper habitats indicate that some individuals did reside deeper. For example, in the area were there was as steep vertical slope, one fish (tag #022) was recorded to reside at a depth of approximately 70 m. While Nassau grouper are primarily considered a shallow habitat species, these data reveal that Nassau grouper not only frequent deeper depths temporarily, associated with migrations for reproductive purposes, but they can also reside and maintain home ranges within deeper, mesophotic coral ecosystems. Given that the majority of BDS habitat occurs at deeper depths (> 40 m), and outside of the detection range of the array, this could explain why only half of the tagged fish were detected within the array during the non-spawning period. This further illustrates the importance of documenting the full depth distribution of a species, particularly with respect to the design of spatially based conservation measures and the significance of the seasonal closure within the MPA that contains a multi-species FSA site.
Acoustic tracking from previous studies have recorded multiple periodic, short-term migrations to and from Nassau grouper spawning sites corresponding to peak spawning activity (Starr et al., 2007; Nemeth, 2012; Kobara et al., 2013; Rowell et al., 2015). The spawning season for Nassau grouper has been reported to occur from November through July with the timing of peak spawning activity highly synchronized with the full moon (Colin, 1992; Domeier and Colin, 1997; Whaylen et al., 2004; Starr et al., 2007; Heyman and Kjerfve, 2008; Rowell et al., 2015; Dahlgren et al., 2016; Stump et al., 2017). Variations in timing and duration are reported throughout the region but assumed to be fixed at the site level (Rowell et al., 2015). The Nassau grouper aggregation at BDS corresponds with the January through March new moons (Schärer et al., 2012; Tuohy et al., 2016). However, during the 2014 and 2016 tracking periods, three individuals underwent migrations to the FSA in April and May that correspond to the site’s spawning lunar periodicity. Albeit less than 24 hrs were spent at the aggregation site with no other indication of spawning activity recorded at that time, these movements occurred within the reported spawning season indicating these movements are likely associated with reproductive behaviors. Since its detection, monitoring efforts at BDS indicate relatively low densities of Nassau grouper at peak spawning periods (Schärer-Umpierre et al., 2014). Thus, this movement pattern after the spawning aggregation formation could indicate an individual’s recognition of density dependence for spawning, where individuals leave their home site to the FSA, but low density or lack of individuals in spawning condition encountered upon arrival prompted them to return to their home site. Alternatively, this behavior could be a result of naïve individuals verifying the correct timing of reproduction through trial and error; however, the total length of the individuals at BDS ranged from 60 to 78 cm (Table 1) indicating these fish were well into sexual maturity (Lm = 40-45 cm TL; Sadovy and Eklund, 1999) and may have already spawned at least once.
A largely over-looked component of ecosystem connectivity is the importance of the ecological contribution (trophic interactions, nutrient cycling, nutrient availability, and nutrient translocation) of organisms as they move throughout their environment (Sheaves, 2009; Nemeth, 2012). The implications to ecosystem connectivity associated with large-scale spawning migrations and FSA formations are undoubtedly high (Archer et al., 2015), but they are seasonal and short-lived. Outside of reproduction, Nassau grouper occupy small home range territories of constant depths within the same home reef site that remained relatively consistent among years, indicating a relatively small degree of connectivity at the fine (i.e., individual) scale. A tagged individual was recorded to relocate home reefs making multiple migrations between its original home reef and a potential new site; however, the ecological contribution of such behavior is likely limited given the relatively low proportion of occurrence within the tagged population. Therefore, high relative abundances of this large-bodied marine predator are necessary to maintain their ecological role at the larger (i.e., community) scale, given the limited contribution to ecosystem connectivity throughout its lifespan. Increased abundances would likely lead to increased competition between conspecifics that inhabit that same home reef, but their maintenance of smaller territories of discrete depth ranges could serve as a mechanism to decrease competitive interactions.
Management strategies designed to protect the Nassau grouper have largely focused on protecting FSA sites and spawning populations through seasonal closures, in MPAs or in no-take marine reserves (Starr et al., 2007; Sadovy de Mitcheson et al., 2008; Sadovy de Mitcheson et al., 2012; Schärer et al., 2012; Dahlgren et al., 2016). Thus, understanding their spatial ecology is a critical step to designing effective fisheries management measures or closures during times that offer sufficient protection throughout the year, particularly in areas where reported population densities or ontogenetic connectivity are low. While Nassau grouper are considered relatively sedentary and solitary with high site fidelity to home reef sites (Sadovy de Mitcheson et al., 2012), tagging studies such as this and Blincow et al. (2020) are now documenting the scales at which movements occur. The results of this study highlight the importance of BDS as critical habitat for Nassau grouper not only for the purpose of reproduction, but also as their primary home reef sites. The current six-months seasonal closure at BDS is insufficient for the protection of Nassau grouper during non-spawning time. While the possession and take of Nassau grouper is prohibited year-round in the federal and territorial waters of Puerto Rico, compliance with fisheries regulations is low and a recent survey of commercial small-scale fisheries landings in Puerto Rico (Gedamke et al., 2020) found Nassau grouper to be present in the reported catch. Thus, expanding the fishing prohibition to 12 months of the year would greatly improve the recovery of the Nassau grouper population residing on BDS by protecting all of this sub-population and the spawning corridors within the seamount. For example, in St. Thomas, US Virgin Islands a year-round no-take marine conservation district incorporating the red hind (E. guttatus) FSA has significantly increased their spawning potential ratio (Rosemond et al., 2022). Furthermore, many multi-species FSA sites are used by multiple species at various times throughout the year (Ojeda-Serrano et al., 2007; Heyman and Kjerfve, 2008; Kobara et al., 2013), year-round protection of spawning and home range habits would safeguard the spawning and recruitment dynamics of the highly diverse and taxonomically complex fish species assemblages reported for this location (García-Sais et al., 2007; García-Sais et al., 2020).
Although Nassau grouper are designated as a protected species in Puerto Rico, this does not prevent incidental catch and discard mortality—a likely occurrence at this location due to the depth of the seamount and the depth range profiles of tagged fish. Given the low numbers of Nassau grouper observed at BDS (García-Sais et al., 2007; Schärer-Umpierre et al., 2014; García-Sais et al., 2020), any level of mortality will reduce the probabilities of this population’s recovery. Requiring the possession and use of descending devices by all fishers could potentially reduce release mortality (Benaka et al., 2016). The benefits of requiring such devices would not only increase the chances of survival for released Nassau grouper at BDS, but throughout the extensive Puerto Rico insular shelf. Post-release survival rates observed for red grouper (Epinephelus morio) released with descending devices compared to venting in the Gulf of Mexico provided positive results (Stallings et al., 2023), but these methods still require testing in native Nassau grouper habitats to determine its potential for fisheries management in the Caribbean. Additionally, a continuation and expansion of acoustic telemetry monitoring would be essential to detect the survival of discarded fish as well as any reduction in critical habitats or changes in spawning dynamics due to warming sea surface temperatures as suggested by Asch and Erisman (2018).
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical review and approval was not required for the animal study because this study was conducted before the University of Puerto Rico had an IACUC review board. Instead, this methodology was reviewed and approved at the Federal level by the National Oceanic and Atmospheric Administration (NOAA) and a Letter of Acknowledgment (LOA) was issued granting the approval of this research.
Author contributions
ET, MS-U, and RA contributed to conception, design, and implementation of the study. ET organized and maintained the acoustic telemetry database. ET and LP performed the statistical analysis and modeling. ET wrote the first draft of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
Primary funding for this project was provided by the NOAA SK grant no. NA15NMF4270341, Caribbean Fisheries Management Council (CFMC) and Caribbean Coral Reef Institute (CCRI). Ron Hill and Jennifer Doerr of the Southeast Fisheries Science Center (SEFSC) provided additional funds and supplies to support this effort.
Acknowledgments
The authors would like to thank closed-circuit rebreather divers Michael Nemeth, Ivonne Bejarano and Milton Carlo for assistance with fish tagging and array maintenance. Open-circuit divers Orlando Espinosa, Jaaziel Garcia, Carlos Zayas-Santiago, JP Zegarra, Hanae Spathias, Daniel Mateos, Duane Sanabria, and Phillip Sanchez for assistance with array maintenance and CCR diver support. The crew of the “Orca Too”, Francisco García-Huertas and Carlos Vélez (RIP) were essential in providing vessel support and assistance with fish trapping and dive logistics. Alejandro Acosta and Paul Barbera of the Florida Fish and Wildlife Conservation Commission aided with the development of the in situ tagging methodology and additional acoustic transmitters. The FACT Network assisted with acoustic telemetry analysis and equipment support. The Department of Marine Science of the University of Puerto Rico-Mayagüez for the use of vessel and dive equipment. Primary funding was provided by the NOAA Saltonstall-Kennedy grant No. NA15NMF4270341, as well as the Caribbean Fisheries Management Council (CFMC) and the Caribbean Coral Reef Institute (CCRI). Ron Hill and Jennifer Doerr of the Southeast Fisheries Science Center (SEFSC) provided additional funds and supplies to support this effort.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aguilar-Perera A. (2006). Disappearance of a Nassau grouper spawning aggregation off the southern Mexican Caribbean coast. Mar. Ecol. Prog. Ser. 327, 289–296. doi: 10.3354/meps327289
Archer S. K., Allgeier J. E., Semmens B. X., Heppell S. A., Pattengill-Semmens C. V., Rosemond A. D., et al. (2015). Hot moments in spawning aggregations: implications for ecosystem-scale nutrient cycling. Coral Reefs 34, 19–23. doi: 10.1007/s00338-014-1208-4
Asch R. G., Erisman B. (2018). Spawning aggregations act as a bottleneck influencing climate change impacts on a critically endangered reef fish. Diversity Distrib. 1–17. doi: 10.1111/ddi.12809
Aspillaga E., Bartumeus F., Linares C., Starr R. M., López-Sanz À., Díaz D., et al. (2018). Ordinary and extraordinary movement behaviour of small resident fish within a Mediterranean marine protected area. PloS One 11 (7), e0159813. doi: 10.1371/journal.pone.0159813
Benaka L. R., Sharpe L., Abrams K. H., Campbell M., Cope J. M., Darby F., et al. (2016). Action plan for fish release mortality science. NOAA Technical Report NMFS.
Blincow K. M., Bush P. G., Heppell S. A., McCoy C. M., Johnson B. C., Pattengill-Semmens C. V., et al. (2020). Spatial ecology of Nassau grouper at home reef sites: using acoustic telemetry to track a large, long-lived epinephelid across multiple years, (2005-2008). Mar. Ecol. Prog. Ser. 655, 199–2145. doi: 10.3354/meps13516
Bloch M. E. (1792). Naturgeschichte der ausländischen Fische. Sechster Theil (Berlin:J. Morino & Co).
Bolden S. K. (2000). Long-distance movement of a Nassau grouper (Epinephelus striatus) to a spawning. Fishery Bull. 98 (3-4), 642.
Bolden S. K. (2001). Nassau Grouper (Epinephelus striatus, Pisces: serranidae) movement in the Bahamas, as determined by ultrasonic telemetry (University of Miami).
Börger L., Dalziel B. D., Fryxell J. M. (2008). Are there general mechanisms of animal home range behaviour? a review and prospects for future research. Ecol. Lett. 11 (6), 637–650. doi: 10.1111/j.1461-0248.2008.01182.x
Burt W. H. (1943). Territoriality and home range concepts as applied to mammals. J. Mammal. 24 (3), 346–352. doi: 10.2307/1374834
Calenge C. (2006). The package adehabitat for the R software: tool for the analysis of space and habitat use by animals. Ecol. Model. 197, 1035. doi: 10.1016/j.ecolmodel.2006.03.017
Cheung W. W., de Mitcheson Y. S., Braynen M. T., Gittens L. G. (2013). Are the last remaining Nassau grouper Epinephelus striatus fisheries sustainable? status quo in the Bahamas. Endanger. Species Res. 20 (1), 27–39. doi: 10.3354/esr00472
Cohen D. M., Inada T., Iwamoto T., Scialabba N., Whitehead P. J. P. (1990). FAO species catalogue: vol. 10 gadiform fishes of the world (order gadiformes), an annotated and ilustrated catalogue of Cods. Hakes, grenadiers and other gadiform fishes known to date (No. Sirsi) a324545) (FAO).
Colin P. L. (1992). Reproduction of Nassau grouper, Epinephelus striatus (Pisces: Labroidei) and its relationship to environmental conditions. Environ. Biol. Fish. 34, 357–377. doi: 10.1007/BF00004740
Crabtree R. E. (2009). Regulatory amendment to the fishery management plan for the reef fish fishery of Puerto Rico and the US virgin islands modifying the Bajo de Sico seasonal closure including a regulatory impact review and an environmental assessment. Final Report to Caribbean Fisheries Management Council, Puerto Rico.
Dahlgren C. P., Buch K., Rechisky E., Hixon M. A. (2016). Multiyear tracking of Nassau grouper spawning migrations. Mar. Coast. Fish. 8 (1), pp.522–pp.535. doi: 10.1080/19425120.2016.1223233
Domeier M. L., Colin P. L. (1997). Tropical reef fish spawning aggregations: defined and reviewed. Bull. Mar. Sci. 60 (3), 698–726.
Farmer N. A., Ault J. S., Smith S. G., Franklin E. C. (2013). Methods for assessment of short-term coral reef fish movements within an acoustic array. Movement Ecol. 1 (1), 1–13. doi: 10.1186/2051-3933-1-7
Garcia S. M. (2003). The ecosystem approach to fisheries: issues, terminology, principles, institutional foundations, implementation and outlook (No. 443). Food & Agriculture Org.
García-Sais J. R., Castro R., Sabater-Clavell J., Carlo M., Esteves-Amador R. F., Bruckner A. W., et al. (2007). Characterization of benthic habitats and associated reef communities at Bajo de Sico seamount, Mona Passage, Puerto Rico. Final report.
García-Sais J. R., Williams S., Tuohy E., Sabater-Clavell J., Carlo M. (2020). Monitoring of mesophotic habitats and associated benthic and Fish/Shellfish communities from Abrir la Sierra, Bajo de Sico, Tourmaline, Isla Desecheo, El Seco and Boya 4, 2018-20 survey. Final Report to Caribbean Fisheries Management Council, Puerto Rico.
Gedamke T., Hoenig J. M., Carrera A., Omori K., Gross J., Lastra L., et al. (2020). Puerto Rico Port sampling and catch validation project (August 2017 – December 2019). Final Report to Caribbean Fisheries Management Council, Puerto Rico.
Grüss A., Robinson J. (2015). Fish populations forming transient spawning aggregations: should spawners always be the targets of spatial protection efforts? ICES J. Mar. Sci. 72 (2), 480–497.
Heyman W. D., Kjerfve B. (2008). Characterization of transient multi-species reef fish spawning aggregations at Gladden Spit, Belize. Bull. Mar. Sci. 83 (3), 531–551.
Horne J. S., Garton E. O., Krone S. M., Lewis J. S. (2007). Analyzing animal movements using Brownian bridges. Ecology 88 (9), 2354–2363. doi: 10.1890/06-0957.1
Horne J. S., Garton E. O., Rachlow J. L. (2008). A synoptic model of animal space use: simultaneous estimation of home range, habitat selection, and inter/intra-specific relationships. Ecol. Model. 214 (2-4), 338–348. doi: 10.1016/j.ecolmodel.2008.02.042
Keller J. A., Herbig J. L., Morley D., Wile A., Barbera P., Acosta A. (2020). Grouper tales: use of acoustic telemetry to evaluate grouper movements at Western Dry Rocks in the Florida Keys. Mar. Coast. Fish. 12 (5), 290–307. doi: 10.1002/mcf2.10109
Kobara S., Heyman W. D., Pittman S. J., Nemeth R. S. (2013). Biogeography of transient reef-fish spawning aggregations in the Caribbean: a synthesis for future research and management. Oceanogr. Mar. Biol. Annu. Rev. 51, 281–326.
Kramer D. L., Chapman M. R. (1999). Implications of fish home range size and relocation for marine reserve function. Environ. Biol. Fish. 55, 65–79. doi: 10.1023/A:1007481206399
Moffitt E. A., Botsford L. W., Kaplan D. M., O'Farrell M. R. (2009). Marine reserve networks for species that move within a home range. Ecol. Appl. 19 (7), 1835–1847. doi: 10.1890/08-1101.1
National Oceanic and Atmospheric Administration (2016). Endangered and threatened wildlife and plants: final listing determination on the proposal to list the Nassau grouper as threatened under the Endangered Species Act. Code Federal Regul. 81, 42268–42285.
Nemeth R. S. (2012). “Ecosystem aspects of species that aggregate to spawn,” in Reef fish spawning aggregations: biology, research and management (Dordrecht: Springer Netherlands), 21–55.
Nemeth R. S., Kadison E., Jossart J., Shivji M., Wetherbee B. M., Matley J. K. (2023). Acoustic telemetry provides insights for improving conservation and management at a spawning aggregation site of the endangered Nassau grouper (Epinephelus striatus). Front. Mar. Sci. 10. doi: 10.3389/fmars.2023.1154689
Ojeda-Serrano E., Appeldoorn R., Ruiz-Valentin I. (2007). Reef fish spawning aggregations of the Puerto Rican shelf (Final Report to the CCRI).
R Core Team (2018). R: a language and environment for statistical computing (Vienna: R Foundation for Statistical Computing).
Reynolds J. D., Dulvy N. K., Goodwin N. B., Hutchings J. A. (2005). Biology of extinction risk in marine fishes. Proc. R. Soc. B: Biol. Sci. 272 (1579), 2337–2344.
Rosemond R. C., Nemeth R. S., Heppell S. A. (2022). Demographic recovery of a reef fish population over 30 years of spawning aggregation site protection. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.931409
Rowell T. J., Nemeth R. S., Schärer M. T., Appeldoorn R. S. (2015). Fish sound production and acoustic telemetry reveal behaviors and spatial patterns associated with spawning aggregations of two Caribbean groupers. Mar. Ecol. Prog. Ser. 518, 239–254. doi: 10.3354/meps11060
Sadovy Y. (1994). “Grouper stocks of the western central Atlantic: the need for management and management needs,” in Proceedings of the Gulf and Caribbean Fisheries Institute. 43, 43–64.
Sadovy Y., Aguilar-Perera A., Sosa-Cordero E. (2018). Epinephelus striatus (The IUCN Red List of Threatened Species 2018). T7862A46909843.
Sadovy Y., Eklund A. M. (1999). Synopsis of biological data on the Nassau grouper, Epinephelus striatus (Bloch 1792), and the Jewfish, E. itajara (Lichtenstein 1822). NOAA technical report NMFS-146.
Sadovy de Mitcheson Y. S., Cornish A., Domeier M., Colin P. L., Russell M., Lindeman K. C. (2008). A global baseline for spawning aggregations of reef fishes. Conserv. Biol. 22 (5), 1233–1244. doi: 10.1111/j.1523-1739.2008.01020.x
Sadovy de Mitcheson Y., Craig M. T., Bertoncini A. A., Carpenter K. E., Cheung W. W., Choat J. H., et al. (2013). Fishing groupers towards extinction: a global assessment of threats and extinction risks in a billion dollar fishery. Fish Fish. 14 (2), 119–136. doi: 10.1111/j.1467-2979.2011.00455.x
Sadovy de Mitcheson Y., Heppel S. A., Colin P. L. (2012). “Species case studies,” in Reef fish spawning aggregations: biology, research and management (Dordrecht: Springer), 429–439.
Sala E., Ballesteros E., Starr R. M. (2001). Rapid decline of Nassau grouper spawning aggregations in Belize: fishery management and conservation needs. Fisheries 26 (10), 3–30. doi: 10.1577/1548-8446(2001)026<0023:RDONGS>2.0.CO;2
Sanchez P. J., Appeldoorn R. S., Schärer-Umpierre M. T., Locascio J. V. (2017). Patterns of courtship acoustics and geophysical features at spawning sites of black grouper (Mycteroperca bonaci). Fishery Bull. 115 (2), 186–195. doi: 10.7755/FB.115.2.5
Schärer M. T., Rowell T. J., Nemeth M. I., Appeldoorn R. S. (2012). Sound production associated with reproductive behavior of Nassau grouper Epinephelus striatus at spawning aggregations. Endanger. Species Res. 19 (1), 29–38. doi: 10.3354/esr00457
Schärer-Umpierre M., Nemeth R., Tuohy E., Clouse K., Nemeth M., Appeldoorn R. S. (2014). “Nassau Grouper Epinephelus striatus fish spawning aggregations in the US Caribbean,” in Proceedings of the Gulf and Caribbean Fisheries Institute, Vol. 66. 408–412.
Semmens B. X., Luke K. E., Bush P. G., Pattengill-Semmens C., Johnson B., McCoy C., et al. (2007). “Investigating the reproductive migration and spatial ecology of Nassau grouper (Epinephelus striatus) on little Cayman island using acoustic tags–an overview,” in Proceedings of the Gulf and Caribbean Fisheries Institute, Vol. 56. 1–8.
Sheaves M. (2009). Consequences of ecological connectivity: the coastal ecosystem mosaic. Mar. Ecol. Prog. Ser. 391, 107–115. doi: 10.3354/meps08121
Stallings C. D. (2009). Predator identity and recruitment of coral-reef fishes: indirect effects of fishing. Mar. Ecol. Prog. Ser. 383, 251–259. doi: 10.3354/meps08004
Stallings C. D., Ayala O., Cross T. A., Sauls B. (2023). Post-release survival of red snapper (Lutjanus campechanus) and red grouper (Epinephelus morio) using different barotrauma mitigation methods. Fish. Res. 264, 1–8. doi: 10.1016/j.fishres.2023.106717
Starr R. M., Sala E., Ballesteros E., Zabala M. (2007). Spatial dynamics of the Nassau grouper Epinephelus striatus in a Caribbean atoll. Mar. Ecol. Prog. Ser. 343, 239–249. doi: 10.3354/meps06897
Stump K., Dahlgren C. P., Sherman K. D., Knapp C. R. (2017). Nassau Grouper migration patterns during full moon suggest collapsed historic fish spawning aggregation and evidence of an undocumented aggregation. Bull. Mar. Sci. 93 (2), pp.375–pp.389. doi: 10.5343/bms.2016.1042
Tuohy E., Nemeth M. I., Bejarano I., Schaerer M. T., Appeldoorn R. S. (2015). In situ tagging of Nassau grouper Epinephelus striatus using closed-circuit rebreathers at a spawning aggregation in Puerto Rico. Mar. Technol. Soc. J. 49 (1), 115–123. doi: 10.4031/MTSJ.49.1.9
Tuohy E., Schärer M. T., Appeldoorn R. S. (2016). “Spatio-temporal dynamics of a Nassau grouper spawning aggregation in Puerto Rico,” in Proceedings of the Gulf and Caribbean Fisheries Institute, Vol. 69. 319–320.
Waterhouse L., Heppell S. A., Pattengill-Semmens C. V., McCoy C., Bush P., Johnson B. C., et al. (2020). Recovery of critically endangered Nassau grouper (Epinephelus striatus) in the Cayman islands following targeted conservation actions. Proc. Natl. Acad. Sci. 117 (3), 1587–1595. doi: 10.1073/pnas.1917132117
Werner E. E., Gilliam J. F. (1984). The ontogenetic niche and species interactions in size-structured populations. Annu. Rev. Ecol. Syst. 15 (1), 393–425. doi: 10.1146/annurev.es.15.110184.002141
Whaylen L., Pattengill-Semmens C. V., Semmens B. X., Bush P. G., Boardman M. R. (2004). Observations of a Nassau grouper, Epinephelus striatus, spawning aggregation site in little Cayman, Cayman islands, including multi-species spawning information. Environ. Biol. Fish. 70, 305–313. doi: 10.1023/B:EBFI.0000033341.57920.a8
Keywords: Nassau grouper, acoustic telemetry, home range, habitat use, endangered species, Puerto Rico
Citation: Tuohy E, Schärer-Umpierre M, Penrod L and Appeldoorn R (2023) Home range, space use, and vertical distribution of Nassau grouper (Epinephelus striatus) during non-spawning times in Western Puerto Rico. Front. Mar. Sci. 10:1206070. doi: 10.3389/fmars.2023.1206070
Received: 14 April 2023; Accepted: 19 June 2023;
Published: 11 July 2023.
Edited by:
Salvatore Siciliano, Fundação Oswaldo Cruz (Fiocruz), BrazilReviewed by:
Alfonso Aguilar-Perera, Universidad Autónoma de Yucatán, MexicoJorge Paramo, University of Magdalena, Colombia
Copyright © 2023 Tuohy, Schärer-Umpierre, Penrod and Appeldoorn. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Evan Tuohy, ZXZhbi50dW9oeUB1cHIuZWR1