 Gabriella M. Hancock
Gabriella M. Hancock Katie D. Dudley2
Katie D. Dudley2 Christopher G. Lowe
Christopher G. Lowe- 1Stress and Technology Applied Research Laboratory, Department of Psychology, California State University – Long Beach, Long Beach, CA, United States
- 2Department of Recreation and Leisure Studies, California State University – Long Beach, Long Beach, CA, United States
- 3Department of Economics, California State University – Long Beach, Long Beach, CA, United States
- 4Shark Lab, Department of Biological Sciences, California State University – Long Beach, Long Beach, CA, United States
Keystone species populations, including shark and ray taxonomic groups, are declining due to numerous threats. A better understanding of how individual belief structures inform pro-conservation behavioral intentions is therefore critical to foster support for their conservation and the greater biodiversity of world ecosystems. This study conducted a survey to establish the general public’s cognitive beliefs about the mental and affective capacities of two species groups (sharks and stingrays) known to cause injury to humans, and their effects on different behavioral intentions to support their conservation. Results revealed that a range of different behavioral intentions to support animal conservation are driven by a complex interplay of cognitive beliefs, and some effects were specific to a particular species group while others were not. Beliefs in animals’ cognitive awareness and their emotional abilities were predictive of individuals’ self-reported desire to learn more about the animals and their conservation. Beliefs in animals’ cognitive and emotional capacities were predictive of supporting conservation in terms of voting and donating to their conservation. Finally, beliefs in animals’ reasoning abilities were predictive of different behavioral expressions of tolerance, and support for their conservation via voting only. Despite the potential injurious capabilities of these species, these findings have important implications for educational and conservation programs to emphasize or modify specific beliefs in order to facilitate particular pro-conservation behaviors for maximum benefits to conservation efforts.
1 Introduction
The past fifty years have witnessed a 71% decline in the populations of oceanic sharks and rays (Pacoureau et al., 2021). Such losses not only pose a credible and critical threat to the survival of these individual species groups, but also to the structure, function, and maintenance of the larger ecosystems that rely on them. Furthermore, the greatest dangers to the continued survival of these animals are rooted in human behaviors resulting from overfishing, bycatch, pollution, habitat degradation, and larger climate change considerations (Shiffman et al., 2020). Dulvey et al (2021) observed that the most universal threat was overfishing (affecting all 391 observed threatened species), followed by habitat loss/degradation (31.2%), climate change (10.2%), and pollution (6.9%). As a result, over one-third of Chondrichthyes are now threatened; and the number of threatened species within this class more than doubled over a seven-year time frame (2014-2021). To successfully protect the biodiversity of the ocean, regulatory bodies need to implement significant, scientifically driven programs and legislation to protect wildlife at the regional, national, and international levels (Crossley et al., 2014).
Such large-scale sociopolitical movements, while necessary, are dependent on the perceptions and actions of individuals. Acuña-Marrero et al. (2018) forwarded a psychological framework to illustrate the conceptual relationships among various cognitive and affective precursive factors and conservation behaviors of both individuals and the wider public. Successful shark conservation is consequently predicated on numerous intrinsic (e.g., personal beliefs and emotions) and extrinsic factors (e.g., previous experience interacting with these animals and/or exposure to portrayals of them in popular media). This work focuses on such intrinsic factors, specifically cognitive beliefs, and their effects on humans’ behavioral intentions to support conservation. According to this theoretical model, individuals’ personal beliefs drive behavioral indicators of conservation, including professing a desire to learn more about the animals, expressing tolerance, and providing active support for their conservation. Importantly, the model emphasizes that these relationships can be mediated by increasing knowledge. Understanding how to effectively educate and ensuring that perceptions are based in fact are therefore critical on the individual level, as doing so is a necessary antecedent condition in building conservation support in the general public. This work therefore focuses on individual beliefs pertaining to the cognitive and emotional capacities of separate species groups (sharks and stingrays); meaning, the extent to which humans believe that these animals possess the capacity to process and comprehend information effectively, and their ability to process, experience, and express affective mental states. This work examines the effects of these beliefs on people’s desire to learn more, behavioral tolerance, and willingness to support conservation measures focused on these targeted species groups and their habitats.
Extensive biological research has shown that both sharks and stingrays possess a wide range of cognitive and emotional abilities (Guttridge et al., 2009). Sharks can be both classically and operantly conditioned, demonstrating their capacity to form and maintain mental associations between a stimulus and specific desirable and undesirable outcomes, their ability to discriminate amongst varying stimuli, and to learn from experience (Guttridge et al., 2009; Guttridge and Brown, 2014). They can also remember these learned behavioral patterns for at least a number of weeks, indicating the capacity for functional memory (Kimber et al., 2014). Sharks also possess emotional capabilities as they demonstrate physiological and behavioral responses to stressors (Parsons and Carlson, 1998; Bouyoucos et al., 2017), as well as individual differences in such stress reactivity and indicators of functional coping strategies (Byrnes and Brown, 2016).
Stingrays have also demonstrated sophisticated cognitive abilities. Like sharks, they can also be classically conditioned (Siciliano et al., 2013). They also possess the ability to form and utilize cognitive maps of their environments (i.e., a map-like representation of objective space that provides a stable frame of reference and affords spatial comparisons for successful navigation; Schluessel and Bleckmann, 2005). Using this cognitive map, stingrays demonstrate another cognitive ability (i.e., memory) by successfully remembering fruitful feeding locations and navigating to these target areas from different unique starting points (Schluessel and Bleckmann, 2005; Schluessel et al., 2015). Stingrays have also demonstrated a fundamental degree of emotional processing as they display physiological (Lambert et al., 2018) and behavioral responses to stress (Mickle et al., 2022).
There has been extensive debate as to whether or not fish are capable of feelings. Proponents of this argument point to the fact that fish, like humans, possess nociceptive systems (i.e., mechanisms in the central nervous system that process information about potential harm to the body and translate these signals into a subjective experience of pain; Coghill, 2020) meant to prompt adaptive action to protect against such threats (Chandroo et al., 2004; Braithwaite and Boulcott, 2007). Detractors, on the other hand, make the case that necessary regions of the cerebral cortex associated with the higher-order subjective experience of pain in the human are missing in fish (Rose, 2002), and therefore drawing any direct equivalencies between emotional responses in humans and fish are unfounded and potentially harmful (Rose, 2007). The fundamental case can be made, however, that all vertebrates possess dedicated pain systems (Broom, 2014), and therefore use emotional processing – however rudimentary - to function according to the hedonic principle: approach stimuli that yield pleasant or favorable outcomes and avoid stimuli that produce unpleasant or unfavorable outcomes (Cornwell et al., 2014).
Despite this controversial stance about the veracity and nature of sharks’ and stingrays’ neurophysiological abilities, it is true that humans anthropomorphize animals and often ascribe these cognitive and emotional abilities to them. Moreover, there are individual differences in the extent to which people subscribe to these beliefs (Chin et al., 2004). Furthermore, these perceptions have been found to influence attitudes and behavioral intentions regarding animals’ welfare (Butterfield et al., 2012), and this anthropomorphic tendency has been used for promoting conservation (Chan, 2012). Finally, while this effect of beliefs in animals’ capacities have been observed to promote prosocial behavioral intentions toward terrestrial animals (e.g., dogs; Butterfield et al., 2012), it has not yet been studied in people’s beliefs and intentions towards fish.
Sharks and stingrays were therefore selected as the species groups of interest for three reasons: 1) their key roles within their respective ecosystems, 2) their increased prevalence in California waters, and 3) their shared reputations as “injurious marine life,” capable of causing serious injury to humans. Comparisons between the two species groups are interesting given the contrast in public perceptions of them. Sharks often suffer from a bad reputation; the public holds erroneous preconceived notions about sharks as blood-thirsty man-eaters and clings to these misconceptions even in the face of contradictory evidence (López de la Lama et al., 2018). Stingrays, on the other hand, are not portrayed as significant threats in the media, despite the fact that they pose the greater risk of injury to humans. To provide some perspective, ocean lifeguards treat thousands of stingray-related injuries at Southern California beaches each year (Lowe et al., 2007; Lowe et al. unpub. data), while only 286 shark attacks were reported worldwide during the same four-year time period (International Shark Attack File, n.d.). Humans’ misunderstanding of the true relative risk posed by these respective taxonomic groups can affect their willingness to support their conservation and is often driven by emotion rather than reason, much to the detriment of these animals.
The general public primarily views sharks as a threat and source of danger (Neves et al., 2021). This belief leads to severely overestimating the risk that sharks pose to humans (Acuña-Marrero et al., 2018), which in turn influences humans’ behaviors; and not just any behaviors, but those which are critical to the conservation of sharks, including policy decisions (the Jaws Effect; Neff, 2015), management strategies (Crossley et al., 2014), and voting (Achen and Bartels, 2004). Voting is of particular concern given that it is amongst the most impactful behaviors that an average person can do to promote successful conservation of any species or environment.
While public perceptions of sharks have been studied in the past (i.e., Pepin-Neff and Wynter, 2018; López de la Lama et al., 2018) as well as other marine animals such as dolphins and whales (Naylor and Parsons, 2018), there is little to no literature addressing public attitudes toward stingrays. This is a notable gap in the literature given the prevalence and relative risk to humans posed by these animals, and so this paper consequently addresses it by assessing beliefs about stingrays and their conservation. Again, juxtaposing the public’s impressions of sharks and stingrays is informative considering the differences in their perceptions and relative risks to human well-being given the growing popularity in ocean recreation.
The purpose of this work is therefore to test the Acuña-Marrero theoretical framework by determining the cognitive influences driving behavioral intentions to support animal conservancy at the individual level, with particular focus on the species groups of sharks and stingrays. This work represents the first study to investigate beliefs pertaining to animal cognition as predictors of conservation behaviors, and the first study to investigate attitudes and perceptions pertaining to stingrays. The authors consequently hypothesized that greater personal beliefs in animals’ cognitive and affective capacities would predict greater behavioral intentions to support conservation in different ways (i.e., greater desire to learn more about the animals and their conservation, greater behavioral expressions of tolerance, and greater support for protection in the form of voting and donating funds to dedicated conservation efforts).
2 Materials and methods
2.1 Participants
Four hundred and thirty total participants were recruited. Fifty participants were excluded as they provided consent and completed no further part of the survey. The analytical sample therefore consisted of 380 participants (122 males, 243 females, 15 unidentified) with an average age of 35.3 years (SD = 15.2 years; range 18-82 years; 55 cases wherein age was not disclosed); see Table 1.
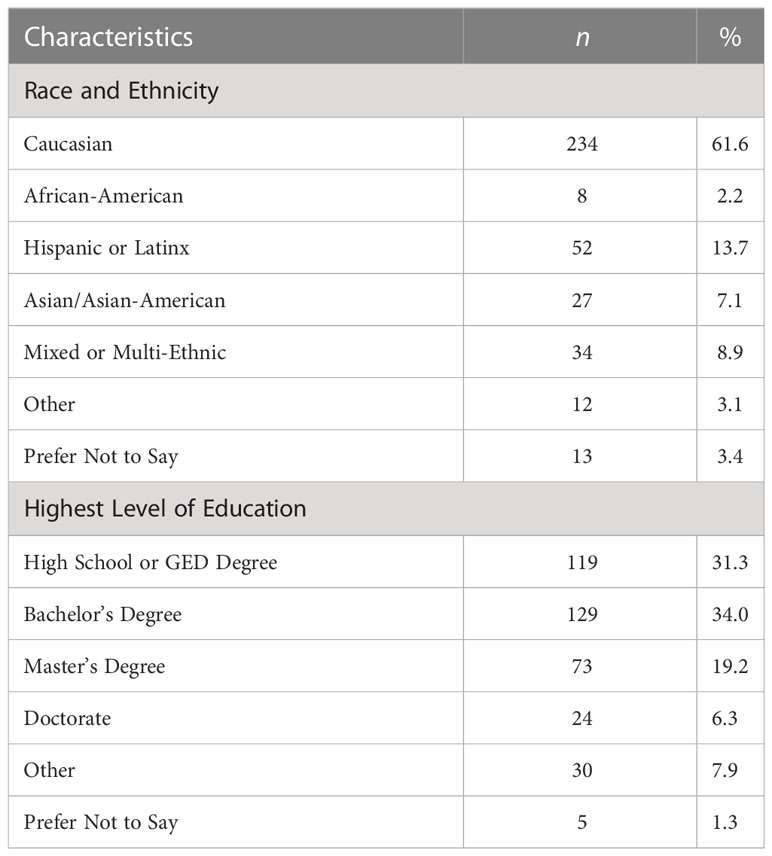
Table 1 Demographic information for survey respondents.
Two hundred and eighty-one (281) participants were California residents. Seventy-three (73) respondents lived in other American states including: Arizona (2), Colorado (2), Connecticut (2), Florida (10), Georgia (3), Hawaii (3), Illinois (6), Kentucky (1), Louisiana (1), Massachusetts (7), Minnesota (6), Nevada (2), New Mexico (2), New York (1), Ohio (8), Oregon (2), Pennsylvania (2), South Carolina (2), Tennessee (1), Utah (1), Virginia (1), Washington (5), West Virginia (2), and Wisconsin (1). Seventeen (17) participants lived outside the United States in the following countries: Argentina (1), Bermuda (1), Canada (4), Egypt (1), France (3), Indonesia (1), Japan (1), the Netherlands (1), Spain (1), and Switzerland (3); and 9 participants did not disclose place of residence.
It should be noted that, per IRB stipulations, participants had the option to refrain from answering any question. Participants were recruited via educational facilities presented along the California coast, via the authors’ lab website and social media accounts, and via internal research programs. In-person recruitment took place at 20 CSULB Shark Shack educational events over the course of three months (June-August 2021) across 18 unique locations on the California coast (e.g., Belmont Shore Pier, Huntington Beach Pier, etc.). Individuals who agreed to participate in the study were given a Quick Response (QR) code to complete the survey on a digital device. The survey, with its associated QR participation code, was also posted to the CSULB Shark Lab’s website landing page and its associated Instagram account to facilitate access. Internal research programs for CSULB students to participate as research volunteers were also used; all of which involved the same recruitment procedure of supplying the study’s unique QR code.
2.2 Experimental design
A novel psychological framework modeling the causal mechanisms underlying animal conservation was put forth within the last five years, and data are consequently necessary for its validation and refinement (Acuña-Marrero et al., 2018). This study was consequently exploratory to determine the validity of inclusion of individual cognitive beliefs within this theoretical model, and to provide data for such justification from a different population from the original dataset (California versus the Galapagos Islands). This study also utilized a different category of cognitive beliefs. The original study focused on beliefs concerning sharks’ conservation status (e.g., how threatened they are perceived to be), aesthetics (positive or negative), and their effects on the economic and ecological landscapes around them (positive or negative; Acuña-Marrero et al., 2018). This work includes cognitive beliefs pertaining to the animals’ mental capacities and cognition, which have not been investigated elsewhere as potential predictors of behavioral intentions of their conservation. Therefore, given the exploratory nature of the study, it was also quasi-experimental. Consequently, no experimental manipulation of these beliefs was implemented; instead, participants’ extant trait beliefs in these taxonomic groups’ capacities were assessed and analyzed. Finally, this study constitutes the first instance in which public attitudes and perceptions relating to stingrays have been assessed.
2.3 Variables
The independent variables were participants’ beliefs in animal minds; the extent to which they perceived a species group as possessing certain cognitive and affective capacities. These beliefs were independently assessed for the taxonomic groups of sharks and stingrays via the Belief in Animal Minds Scale (Hills, 1995; Higgs et al., 2020) described in the following section (i.e., 2.3 Equipment). The participants were not instructed to think of any particular species within this larger taxonomic group, nor were they shown any videos or pictures of particular species in order to avoid priming or biasing their perceptions as attitudes have the ability to vary widely by species. This work was focused on beliefs pertaining to these larger taxonomic groups as a whole, and future work will be dedicated to investigating any inter-species differences experimentally. Participants were therefore asked to respond to the questionnaire items with the broadly labeled taxonomic groups of ‘shark’ and ‘stingray’ with their extant beliefs about their capacities (and see the survey items in Supplementary Material). Similarly, to avoid instilling a belief in the animals’ capacities that may not exist, no evidence pertaining to the specific mental capacities was presented; only the survey items were presented (again, see Supplementary Material).
The dependent variables of interest encompassed the three different behavioral components predicating successful conservation as put forward in the Acuña-Marrero et al. (2018) psychological framework: 1) the desire to know or learn more about the species group and its conservation, 2) tolerance, and 3) intentions to support conservancy. The first dependent variables were participants’ desire to learn more about a species group, assessed separately for sharks and stingrays. The second category of dependent variables was behavioral expressions of tolerance based on the species group and different temporal proximity thresholds (i.e., the closer in time a person would willingly put themselves in the same location as the animal). These variables were operationalized as: willingness to physically go into the water at a beach that had a 1) confirmed shark sighting within the past month, 2) confirmed shark sighting within the past 24 hours, 3) confirmed stingray sighting in the last month, and 4) confirmed stingray sighting in the last 24 hours. The final category of dependent variable concerned the willingness to support conservation as operationalized via different behaviors. These variables included willingness to vote in support of conservation measures targeting a specific species group and their habitat, assessed separately for sharks and stingrays; and participants’ willingness to donate monetary funds to conservation efforts supporting a specific species group and their habitat - again, measured separately for sharks and rays. Each of these dependent measures was presented as a dichotomous (yes/no) choice, envisioned as a one-shot decision for ecological validity wherein one final decision must be made and cannot be revised (e.g., one person cannot vote on the same legislative proposition multiple times in one election).
2.4 Equipment
Qualtrics software (Qualtrics XM; Seattle, WA) was used for the presentation of the survey and all data collection. The Belief in Animal Minds Scale (BAMS; Hills, 1995; Higgs et al., 2020) was used to quantify participants’ perceptions of the two species groups. The scale consists of four items (two items reverse-coded) that measure representative dimensions: animals’ cognitive capacity, emotional capacity, unawareness of self and circumstances (reverse - awareness), and extent to which they function/act on instinct or mechanical functioning (reverse - reasoning). These items have demonstrated high Cronbach’s alpha across studies (α = .96, Hills, 1995; α = 0.958, Higgs et al., 2020). Responses were provided via a 7-point Likert scale, ranging from 1 ‘Strongly Disagree’ to 7 ‘Strongly Agree’; a higher score indicating a greater belief in animals’ minds. The Higgs et al. (2020) 7-point scale was used rather than the original 6-point Hills (1995) version to allow for comparison to more recent research, to provide a greater range of response to reflect the variability of these attitudes, and to afford the opportunity of providing a neutral response. The four questionnaire items are presented in the copy of the survey located in Supplementary Materials.
3 Results
Forward stepwise binary logistic regression was estimated via maximum likelihood in each case, iteratively with a 95% confidence interval. As research has identified sex, age, and education level as robust predictors of conservation behaviors (Kim et al., 2013), these subject variables were included in the first block of each analysis, and the independent variables of interest as they pertain to the species groups of sharks or stingrays were added in the second block. The variables encompassing beliefs regarding the two taxonomic groups (sharks and stingrays) were not input into the same analysis as the test would assume a theoretical relationship amongst these predictors that has not yet been established. Supplementary Table 1 detailing all of the classification summaries for the binary logistic regression models can be found in Supplementary Materials.
3.1 Desire to know more
3.1.1 Self-reported desire to know more about sharks and their conservation
The model for belief in sharks’ minds on individuals’ desire to know more about sharks and their conservation was not significant (Chi-square: 3.455, p = 0.063). Beliefs about sharks’ abilities did not affect people’s desire to learn more about them.
The model of belief in stingrays’ minds was significant (Chi-square: 7.429, p = 0.006, Cox & Snell R2 = 0.029, Nagelkerke R2 = 0.080). The average correct percentage of classification was 94%. Beliefs about stingrays’ capacities therefore influenced people’s desire to learn more about sharks. Specifically, the belief in stingrays’ cognitive awareness was a significant predictor of desire to learn more about sharks and their conservation (B = 0.386, p = 0.004, Exp(B) = 1.472, 95% CI for Exp(B) [1.128, 1.920]). For every one-unit increase signifying a greater belief in stingrays’ cognitive awareness, the odds of expressing a greater desire to know more about shark conservation increased by 47.2%. The more aware people believe stingrays to be, the more they would like to learn about sharks.
3.1.2 Self-reported desire to know more about stingrays and their conservation
The model of belief in sharks’ minds was significant (Chi-square: 9.533, p = 0.009, Cox & Snell R2 = 0.037, Nagelkerke R2 = 0.07). The average correct percentage of classification was 87.8%. People’s beliefs about sharks’ capabilities therefore predicted individuals’ desire to learn more about stingrays. Specifically, the belief in sharks’ cognitive awareness was a significant predictor of desire to learn more about stingrays and their conservation (B = 0.278, p = 0.014, Exp(B) = 1.321, 95% CI for Exp(B) [1.057, 1.650]). For every one-unit increase signifying a greater belief in sharks’ cognitive awareness, the odds of expressing a greater desire to know more about stingray conservation increased by 32.1%. The more aware people believe sharks to be, the more they would like to learn about stingrays. Age was also a significant predictor (B = -0.026, p = 0.025, Exp(B) = 0.975, 95% CI for Exp(B) [0.953,.997]). As illustrated in Figure 1, for every one-year increase in age, participants were 2.5% less likely to express a desire to learn more about stingrays. Younger people were therefore more willing to learn about stingrays.
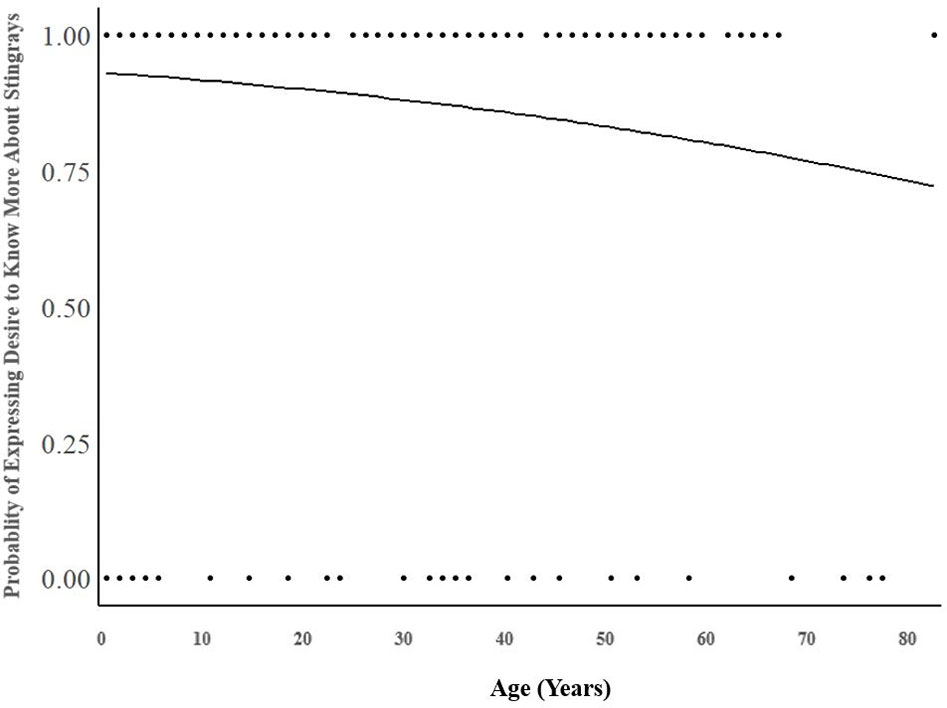
Figure 1 Significant effect of age on the self-reported desire to know more about stingrays and their conservation (B = -0.026, p = 0.025).
The model of belief in stingrays’ minds was significant for predicting the desire to know more about stingrays and their conservation (Chi-square: 8.759, p = 0.003, Cox & Snell R2 = 0.034, Nagelkerke R2 = 0.066). The average correct percentage of classification is 88.1%. Beliefs about stingrays’ abilities influenced the desire to learn more about them and their conservation. Specifically, the belief in stingrays’ emotional capacities was a significant predictor of self-reported desire to learn more about stingrays and their conservation (B = 0.346, p = 0.003, Exp(B) = 1.413, 95% CI for Exp(B) [1.127, 1.773]). For every one-unit increase signifying a greater belief in stingrays’ emotional capacities, the odds of expressing a greater desire to know more about stingray conservation increased by 41.3%. In short, the more people believe stingrays capable of feeling and responding to emotions, the more they want to know about them.
3.2 Behavioral expressions of tolerance
3.2.1 Willingness to enter water at beach with confirmed shark sighting within the last month
The model of belief in sharks’ minds was significant for predicting individuals’ willingness to enter the water at a beach with a confirmed shark sighting within the last month (Chi-square: 8.338, p = 0.004, Cox & Snell R2 = 0.032, Nagelkerke R2 = 0.053). The average correct percentage of classification is 82.4%. Belief in sharks’ reasoning abilities was a significant predictor (B = 0.245, p = 0.004, Exp(B) = 1.277, 95% CI for Exp(B) [1.080, 1.511]). For every one-unit increase signifying a greater belief in sharks’ reasoning abilities, the odds of willingly entering waters wherein a shark had been sighted within the previous month increased by 27.7%. In essence, the more people believe sharks to be capable of reasoning (i.e., thinking logically to guide their behavior in a purpose-driven way), the more willing people are to be in the water with them.
The model of belief in stingrays’ minds was also significant for predicting willingness to go into the water at a beach with a confirmed shark sighting within the previous month (Chi-square: 8.561, p = 0.003, Cox & Snell R2 = 0.033, Nagelkerke R2 = 0.055). The average correct percentage of classification is 82.5%. Belief in stingrays’ reasoning abilities was a significant predictor (B = 0.267, p = 0.004, Exp(B) = 1.306, 95% CI for Exp(B) [1.088, 1.568]). For every one-unit increase signifying a greater belief in stingrays’ reasoning abilities, the odds of willingly entering waters wherein a shark had been sighted within the previous month increased by 30.6%. Believing stingrays to be reasoning creatures also makes one more willing to be in the water with sharks.
3.2.2 Willingness to enter water at beach with confirmed shark sighting within the last 24 hours
The model of belief in sharks’ minds was significant for predicting individuals’ willingness to enter the water at a beach with a confirmed shark sighting within the last 24 hours (Chi-square: 24.116, p <.001, Cox & Snell R2 = 0.091, Nagelkerke R2 = 0.121). The average correct percentage of classification is 62.2%. Belief in sharks’ reasoning abilities was a significant predictor (B = 0.193, p = 0.006, Exp(B) = 1.213, 95% CI for Exp(B) [1.058, 1.392]). For every one-unit increase signifying a greater belief in sharks’ reasoning abilities, the odds of willingly entering waters wherein a shark had been sighted within the previous 24 hours increased by 21.3% (and see Figure 2). Again, the more people believe sharks possess the capacity to reason, the more willing they are to share the same space with them. Sex was also a significant predictor (B = -0.972, p = 0.001, Exp(B) = 0.378, 95% CI for Exp(B) [0.210, 0.682]), indicating that females are 62.2% less likely to be willing to get into the water under these conditions when compared to males. As a result, males reported greater tolerance for sharks than females.
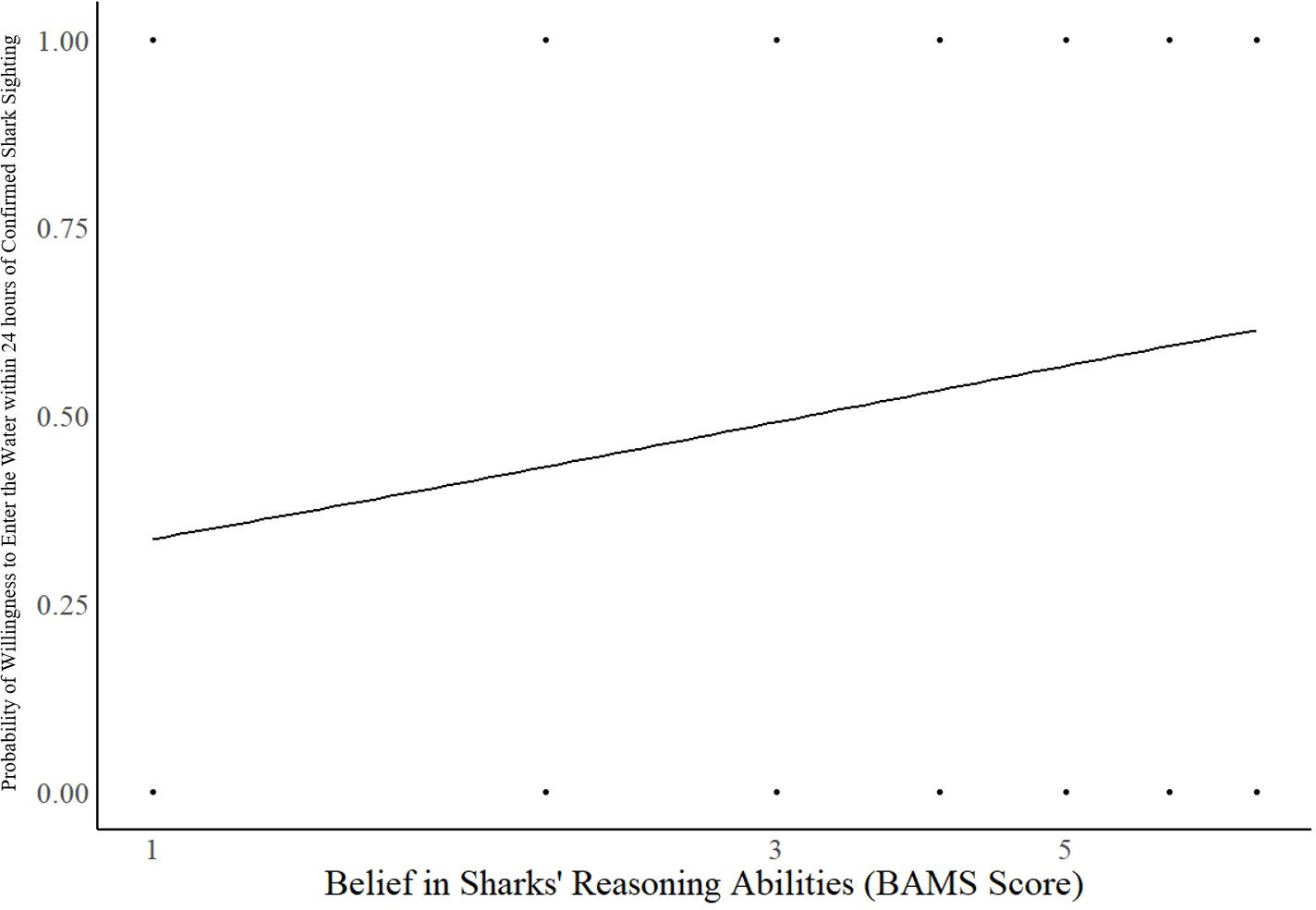
Figure 2 Significant effect of belief in sharks’ reasoning abilities on the willingness to enter the water at a beach with a confirmed shark sighting within the last 24 hours (B = 0.193, p = 0.006). BAMS Score represents the Belief in Animal Minds Scale (BAMS) score for each associated item, in this case the degree of agreement that sharks possess reasoning abilities. The higher the score, the greater the belief in the animal’s capacities.
The model of belief in stingrays’ minds was also significant for predicting willingness to go into the water at a beach with a confirmed shark sighting within the previous 24 hours (Chi-square: 27.141, p <.001, Cox & Snell R2 = 0.102, Nagelkerke R2 = 0.137). The average correct percentage of classification is 63.3%. Belief in stingrays’ reasoning abilities was a significant predictor (B = 0.241, p = 0.001, Exp(B) = 1.273, 95% CI for Exp(B) [1.101, 1.470]). For every one-unit increase signifying a greater belief in stingrays’ reasoning abilities, the odds of willingly entering waters wherein a shark had been sighted within the last 24 hours increased by 27.3%. The more people believe stingrays to be capable of reasoning, the more willing they are to be in the same water with sharks. Sex was also a significant predictor (B = -0.932, p = 0.002, Exp(B) = 0.394, 95% CI for Exp(B) [0.219, 0.709]), indicating that females are 60.6% less likely to be willing to get into the water under these conditions when compared to males. In this model as well, males report being more tolerant of sharks.
3.2.3 Willingness to enter water at beach with confirmed stingray sighting within the last month
The model of belief in sharks’ minds was significant for predicting individuals’ willingness to enter the water at a beach with a confirmed stingray sighting within the last month (Chi-square: 6.468, p = 0.011, Cox & Snell R2 = 0.025, Nagelkerke R2 = 0.045). The average correct percentage of classification is 85.9%. Belief in sharks’ reasoning abilities was a significant predictor (B = 0.235, p = 0.012, Exp(B) = 1.265, 95% CI for Exp(B) [1.054, 1.518]). For every one-unit increase signifying a greater belief in sharks’ reasoning abilities, the odds of willingly entering waters wherein a stingray had been sighted within the previous month increased by 26.5%. The stronger people’s belief in sharks’ ability to reason, the more willing people are to put themselves in the same waters as stingrays.
The model of belief in stingrays’ minds was also significant for predicting willingness to go into the water at a beach with a confirmed stingray sighting within the previous month (Chi-square: 6.651, p = 0.010, Cox & Snell R2 = 0.026, Nagelkerke R2 = 0.047). The average correct percentage of classification is 86.1%. Belief in stingrays’ reasoning abilities was a significant predictor (B = 0.258, p = 0.011, Exp(B) = 1.294, 95% CI for Exp(B) [1.060, 1.580]). For every one-unit increase signifying a greater belief in stingrays’ reasoning abilities, the odds of willingly entering waters wherein a stingray had been sighted within the previous month increased by 29.4%. The more people believed stingrays to be capable of reasoning, the more willing they were to go into the water with them.
3.2.4 Willingness to enter water at beach with confirmed stingray sighting within the last 24 hours
The model of belief in sharks’ minds was significant for predicting individuals’ willingness to enter the water at a beach with a confirmed stingray sighting within the last 24 hours (Chi-square: 16.585, p <.001, Cox & Snell R2 = 0.063, Nagelkerke R2 = 0.087). The average correct percentage of classification is 69.0%. Belief in sharks’ reasoning abilities was a significant predictor (B = 0.159, p = 0.026, Exp(B) = 1.173, 95% CI for Exp(B) [1.020, 1.349]). For every one-unit increase signifying a greater belief in sharks’ reasoning abilities, the odds of willingly entering waters wherein a stingray had been sighted within the previous 24 hours increased by 17.3%. As people’s belief in sharks’ ability to reason increased, so too did their willingness to go into the water with stingrays. Sex was also a significant predictor (B = -1.127, p < 0.001, Exp(B) = 0.324, 95% CI for Exp(B) [0.168, 0.625]), indicating that females are 67.6% less likely to be willing to get into the water under these conditions when compared to males. Males, therefore, expressed greater tolerance for stingrays when compared to females.
The model of belief in stingrays’ minds was also significant for predicting willingness to go into the water at a beach with a confirmed stingray sighting within the previous 24 hours (Chi-square: 19.617, p <.001, Cox & Snell R2 = 0.075, Nagelkerke R2 = 0.104). The average correct percentage of classification is 67.1%. Belief in stingrays’ reasoning abilities was a significant predictor (B = 0.218, p = 0.004, Exp(B) = 1.244, 95% CI for Exp(B) [1.072, 1.442]). For every one-unit increase signifying a greater belief in stingrays’ reasoning abilities, the odds of willingly entering waters wherein a stingray had been sighted within the last 24 hours increased by 24.4%. The more individuals perceived stingrays as reasoning, the more willing they were to go into the water with them. Sex was also a significant predictor (B = -1.090, p = 0.001, Exp(B) = 0.336, 95% CI for Exp(B) [0.174, 0.649]), indicating that females are 66.4% less likely to be willing to get into the water under these conditions when compared to males. Again, males expressed greater tolerance for stingrays when compared to females.
3.3 Support for conservation
3.3.1 Willingness to vote for shark conservation
The model of belief in sharks’ minds was significant for predicting individuals’ willingness to vote in favor of shark conservation (Chi-square: 20.385, p = 0.009, Cox & Snell R2 = 0.077, Nagelkerke R2 = 0.292). The average correct percentage of classification is 96.9%. Belief in sharks’ reasoning abilities was a significant predictor (B = 0.491, p = 0.031, Exp(B) = 1.635, 95% CI for Exp(B) [1.047, 2.553]). For every one-unit increase signifying a greater belief in sharks’ reasoning abilities, the odds of voting in favor of shark conservation increased by 63.5%. The more people believed sharks to have the ability to reason, the more willing they were to vote to protect them. Belief in sharks’ cognitive abilities was also a significant predictor (B = 0.608, p = 0.034, Exp(B) = 1.838, 95% CI for Exp(B) [1.046, 3.230]). For every one-unit increase signifying a greater belief in sharks’ cognitive abilities, the odds of voting in favor of shark conservation increased by 83.8%. The more people believed in sharks’ cognitive abilities (i.e., their capacity to problem-solve, make decisions, etc.), the more willing they were to vote to support their conservation (and see Figure 3).
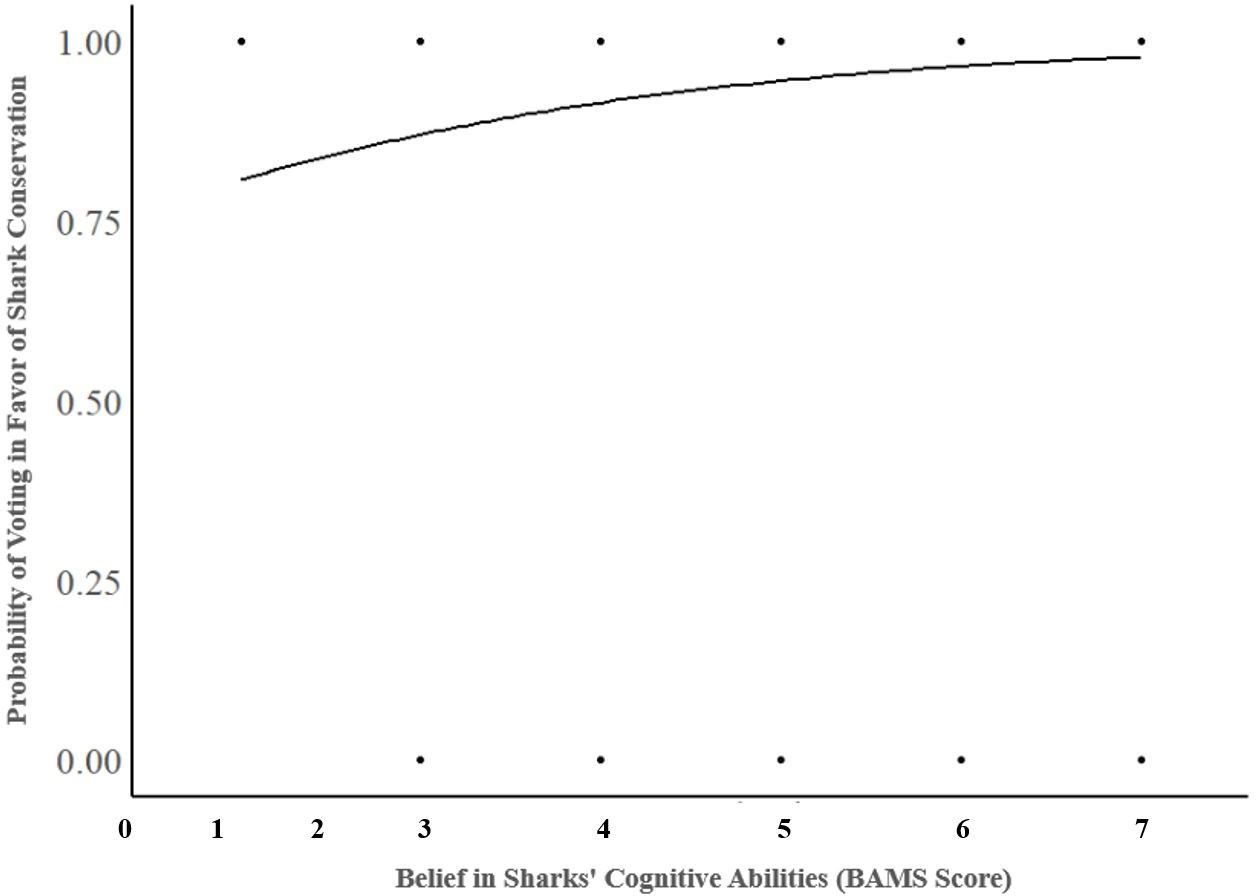
Figure 3 Significant effect of belief in sharks’ cognitive abilities on the willingness to support sharks and their conservation via voting (B = 0.608, p = 0.034). BAMS Score represents the Belief in Animal Minds Scale score for each associated item, in this case the degree of agreement that sharks possess cognitive abilities. The higher the score, the greater the belief in the animal’s capacities.
The model of belief in stingrays’ minds was significant for predicting individuals’ willingness to vote in favor of shark conservation (Chi-square: 16.485, p = 0.021, Cox & Snell R2 = 0.064, Nagelkerke R2 = 0.239). The average correct percentage of classification is 96%. Belief in stingray’s emotional capacities was a significant predictor (B = 0.579, p = 0.012, Exp(B) = 1.785, 95% CI for Exp(B) [1.137, 2.802]). For every one-unit increase signifying a greater belief in stingrays’ emotional capacities, the odds of voting in favor of shark conservation increased by 78.5%. The more people believed stingrays capable of feeling and expressing emotions, the more willing they were to support sharks and shark conservation.
3.3.2 Willingness to vote for stingray conservation
The model of belief in sharks’ minds was significant for predicting individuals’ willingness to vote in favor of stingray conservation (Chi-square: 8.444, p = 0.004, Cox & Snell R2 = 0.033, Nagelkerke R2 = 0.079). The average correct percentage of classification is 92.5%. Belief in sharks’ cognitive abilities was a significant predictor (B = 0.559, p = 0.002, Exp(B) = 1.749, 95% CI for Exp(B) [1.220, 2.507]). For every one-unit increase signifying a greater belief in sharks’ cognitive abilities, the odds of voting in favor of stingray conservation increased by 74.9%. The more people believed sharks capable of cognition (e.g., problem-solving, decision-making), the more willing they were to vote for stingrays and their conservation.
The model of belief in stingrays’ minds was significant for predicting individuals’ willingness to vote in favor of stingray conservation (Chi-square: 13.713, p <.001, Cox & Snell R2 = 0.053, Nagelkerke R2 = 0.128). The average correct percentage of classification is 92.5%. Belief in stingrays’ emotional capacities was a significant predictor (B = 0.517, p <.001, Exp(B) = 1.676, 95% CI for Exp(B) [1.273, 2.207]). For every one-unit increase signifying a greater belief in stingrays’ emotional capacities, the odds of voting in favor of stingray conservation increased by 67.6%. The more individuals believed stingrays are capable of feeling and expressing emotions, the more willing they were to vote in favor of protecting them and their habitat.
3.3.3 Willingness to donate monetary funds to shark conservation
The model of belief in sharks’ minds was significant for predicting individuals’ willingness to donate monetary funds to shark conservation (Chi-square: 14.254, p <.001, Cox & Snell R2 = 0.055, Nagelkerke R2 = 0.077). The average correct percentage of classification is 68.1%. Belief in sharks’ reasoning abilities was a significant predictor (B = 0.176, p = 0.016, Exp(B) = 1.192, 95% CI for Exp(B) [1.034, 1.375]). For every one-unit increase signifying a greater belief in sharks’ reasoning abilities, the odds of donating funds to shark conservation increased by 19.2%. The more people believe sharks are capable of reasoning, the more willing they are to donate funds for their conservation. Belief in sharks’ cognitive abilities was also a significant predictor (B = 0.334, p = 0.012, Exp(B) = 1.396, 95% CI for Exp(B) [1.076, 1.812]). For every one-unit increase signifying a greater belief in sharks’ cognitive abilities, the odds of donating funds to shark conservation increased by 39.6%. The more people believed sharks capable of exercising cognitive abilities, the more willing they were to donate funds for their conservation.
The model of belief in stingrays’ minds was significant for predicting individuals’ willingness to donate monetary funds to shark conservation (Chi-square: 16.282, p <.001, Cox & Snell R2 = 0.063, Nagelkerke R2 = 0.088). The average correct percentage of classification is 68.9%. Belief in stingrays’ reasoning abilities was a significant predictor (B = 0.176, p = 0.016, Exp(B) = 1.192, 95% CI for Exp(B) [1.034, 1.375]). The more people believe stingrays are capable of reasoning, the more willing they are to donate funds for sharks’ conservation. Belief in stingrays’ emotional capacities was a significant predictor (B = 0.357, p <.001, Exp(B) = 1.429, 95% CI for Exp(B) [1.196, 1.709]). For every one-unit increase signifying a greater belief in stingrays’ emotional capacities, the odds of donating funds to shark conservation increased by 42.9%. The more emotionally feeling people believe stingrays to be, the more willing they are to donate funds to shark conservation efforts.
3.3.4 Willingness to donate monetary funds to stingray conservation
The model of belief in sharks’ minds was significant for predicting individuals’ willingness to donate monetary funds to stingray conservation (Chi-square: 20.807, p <.001, Cox & Snell R2 = 0.079, Nagelkerke R2 = 0.107). The average correct percentage of classification is 65%. Belief in sharks’ reasoning abilities was a significant predictor (B = 0.204, p = 0.004, Exp(B) = 1.226, 95% CI for Exp(B) [1.067, 1.408]). For every one-unit increase signifying a greater belief in sharks’ reasoning abilities, the odds of donating funds to stingray conservation increased by 22.6%. The more people believe sharks are capable of reasoning, the more willing they are to donate funds for stingrays’ conservation. Belief in sharks’ cognitive abilities was also a significant predictor (B = 0.406, p = 0.003, Exp(B) = 1.501, 95% CI for Exp(B) [1.151, 1.957]). For every one-unit increase signifying a greater belief in sharks’ cognitive abilities, the odds of donating funds to stingray conservation increased by 50.1%. The more people believe sharks are capable of cognitive abilities, the more willing they are to donate funds for stingrays’ conservation.
The model of belief in stingrays’ minds was significant for predicting individuals’ willingness to donate monetary funds to stingray conservation (Chi-square: 30.812, p <.001, Cox & Snell R2 = 0.116, Nagelkerke R2 = 0.157). The average correct percentage of classification is 66.5%. Belief in stingrays’ reasoning abilities was a significant predictor (B = 0.212, p = 0.007, Exp(B) = 1.236, 95% CI for Exp(B) [1.059, 1.442]). The more people believe stingrays are capable of reasoning, the more willing they are to donate funds for their conservation. Belief in stingrays’ emotional capacities was also a significant predictor (B = 0.334, p <.001, Exp(B) = 1.396, 95% CI for Exp(B) [1.150, 1.695]). For every one-unit increase signifying a greater belief in stingrays’ emotional capacities, the odds of donating funds to stingray conservation increased by 39.6% (and see Figure 4). The more emotionally feeling and expressive people believe stingrays to be, the more willing they are to donate funds to stingray conservation efforts.
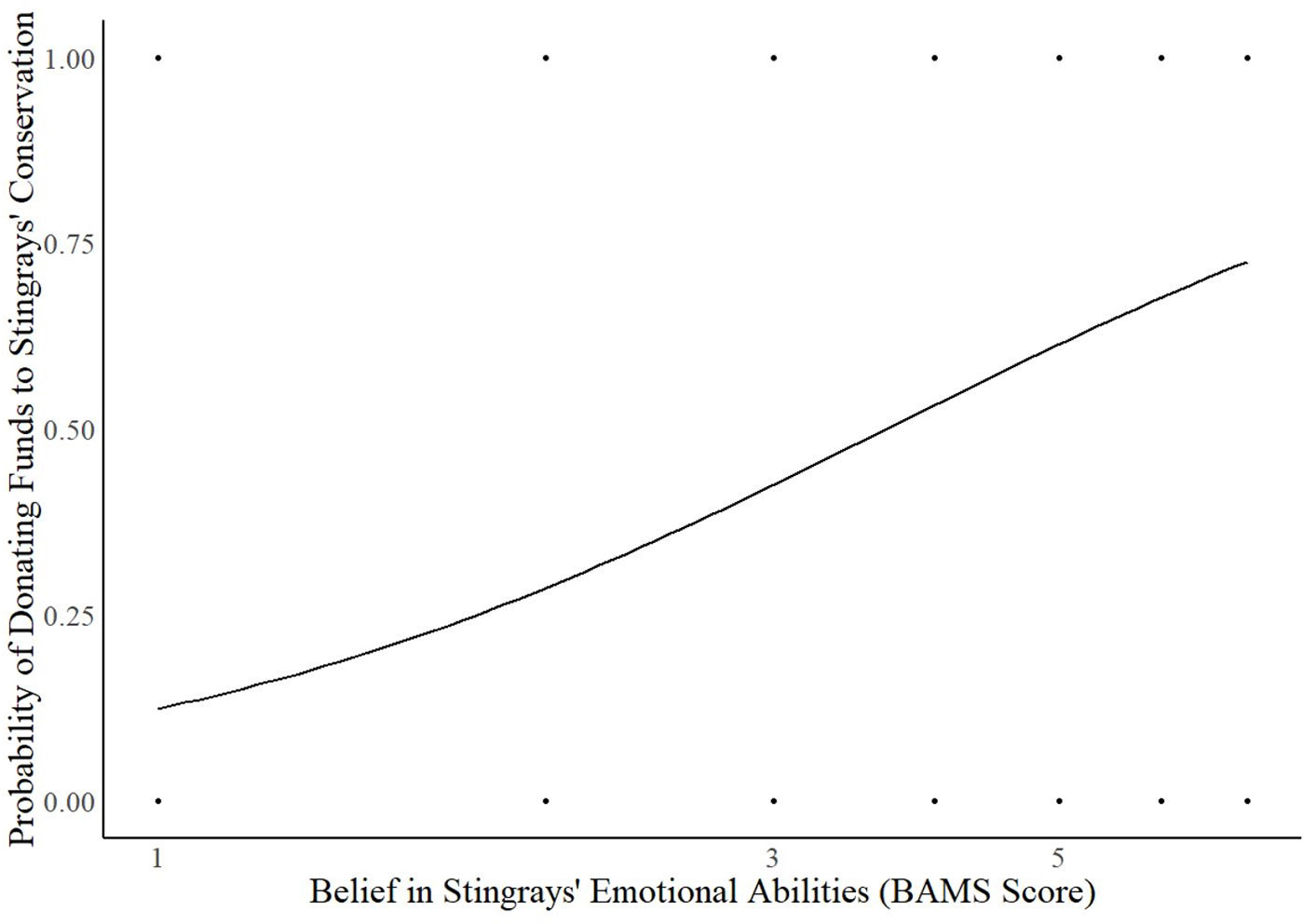
Figure 4 Significant effect of belief in stingrays’ emotional abilities on the willingness to support stingrays and their conservation via donating (B = 0.334, p <.001). BAMS Score represents the Belief in Animal Minds Scale score for each associated item, in this case the degree of agreement that stingrays possess emotional abilities. The higher the score, the greater the belief in the animal’s capacities.
4 Discussion
4.1 Cognitive beliefs and public perceptions
Identifying the personal belief structures that give rise to conservation behaviors is vital for the establishment, promotion, and maintenance of worldwide conservancy efforts that rely on public and legislative support. Acuña-Marrero et al. (2018) suggested that a key precursive factor to successful shark conservation is people’s beliefs about that species group. These results support this hypothesis, demonstrating that a greater belief in animals’ cognitive, reasoning, and emotional capacities led to a greater willingness to support their conservancy across different behaviors (i.e., desire to learn more, behavioral tolerance, supporting conservation via voting and donating). This work represents the first time that beliefs pertaining to animal cognition have been investigated as predictors of pro-conservation behavioral intentions. The interplay between these beliefs was highly complex. Certain beliefs motivated more than one conservation-centric behavior, while others were unique predictors of only a single behavior. Moreover, this work constitutes the first study to investigate beliefs and attitudes pertaining to the taxonomic group of stingrays. These findings will help conservationists identify and strategize as to which perceptions to appeal to or prioritize when promoting conservation efforts. Herein, each respective predictor belief in a specific animal mental capacity (i.e., awareness, cognitive, reasoning, and affective) is discussed in terms of the pro-conservational behavioral intentions that it predicts and influences.
4.1.1 Belief in awareness
Elasmobranchs have an awareness of themselves and their environment. As a basic example, sharks and stingrays react to the introduction of a novel stimulus in their environment, such as the presentation of an electrical stimulus as a simulation of prey (Kajiura and Fitzgerald, 2009; McGowan and Kajiura, 2009). They are, in short, aware of changes in their surroundings as evident by simple and sometimes complex and lasting behavioral responses. As a more complex example, white sharks (Carcharidon carcharias) who became aware of the presence of a predatory pair of killer whales, Orcinas orca, migrated away from their typical aggregation site (Towner et al., 2022). Certain stingray species, as well, such as southern stingrays (Hypanus americanus) will alter their behavior or shift their habitat in response to their awareness of the proximity of predators (Bond et al., 2019).
The belief in animals’ cognitive awareness was predictive of people’s desire to learn more about animals and their conservation. Interestingly, the belief about cognitive awareness was exclusively predictive of desire to learn and no other behavioral components (i.e., tolerance or support). A noteworthy and unexpected dimension about these findings is that the predictive relationship was reversed based on species group: belief in sharks’ cognitive awareness was exclusively predictive of interest in learning more about stingrays, while belief in stingrays’ cognitive awareness exclusively predicted interest in learning more about sharks.
These findings may therefore indicate a self-perceived ceiling effect of individuals’ knowledge about a particular species group. In essence, having already established a set of beliefs about one particular taxonomic group (i.e., the greater belief in the species group’s awareness) does not prompt the desire to learn more about the same species group as said beliefs are already entrenched and robust. These beliefs then instead drive the desire to learn more about a different species group, perhaps to make comparisons between them. This finding is of particular importance, since protection and conservation of batiods (e.g., skates and rays) has lagged behind that of sharks, typically due to lack of notoriety and research (Jorgensen et al., 2022).
4.1.2 Belief in cognitive abilities
Elasmobranchs demonstrate a wide variety of cognitive abilities including discrimination and categorization of stimuli, orientation strategies, social learning, and memory (Schluessel, 2015). For example, in the laboratory, yellow stingrays (Urobatis jamaicensis) can learn behavioral responses based on reward contingencies, indicating the ability to associate a stimulus with a reward and to discriminate among stimuli as to which yield rewards and which do not. They were also able to relearn the task upon modification of the reward structure, and do so at a faster rate than the initial learning curve (Newton and Kajiura, 2020). In the wild, blacktip sharks (Carcharhinus limbatus) integrate input from multiple sensory channels (smell, electroreception, vision, etc.) to navigate over both small- and large-scale distances (Gardiner et al., 2015). Building on a previous example, Towner et al. (2022) observed that not only did white sharks (Carcharodon carcharias) migrate away from an area in light of the presence of predators, but also seemed to remember the potential danger as sightings of the sharks remained scarce in this typically popular aggregation site for the next two and half years (evidence for the presence and use of the cognitive capacity of long-term memory).
People’s belief in sharks’ cognitive abilities (i.e., their ability to solve problems, make decisions, etc.) was predictive of both behaviors to actively support conservation: voting and donating. The more people reported belief in sharks’ cognitive capacities, the greater the odds that they would vote in favor of conservation measures aimed at sharks and their habitat. Interestingly, this effect extended beyond the conservation of a particular taxa as belief in sharks’ cognitive abilities also predicted greater willingness to support the legislative protection of stingrays and donate to stingray conservation. The more individuals perceived sharks as thinking, intelligent creatures, the more willing they were to support both shark and stingray conservation. It should be noted that belief in sharks’ cognitive abilities was not predictive of the other categories of behavioral intentions (i.e., desire to know more or tolerance); it was unique to supporting conservation only. Finally, interestingly, beliefs about stingrays’ cognitive abilities were not predictive of any behavioral intentions. This discrepancy may be due to two potential factors. The first possible consideration could be a relatively greater familiarity with sharks overall given their recognizable, popular status as apex predators, and the greater focus placed on them by the media. This interpretation would support Sergio et al. (2006) proposition of strategically focusing conservation plans on apex predators to yield the greatest benefits for biodiversity across the whole ecosystem. The second possible factor could be the extent to which sharks are perceived to be intentional about their interactions with humans (Pepin-Neff and Wynter, 2018); e.g., whether the shark intends to cause harm to the human. Said intentionality would be the natural product of cognitive processing on the sharks’ part.
4.1.3 Belief in reasoning abilities
Elasmobranch species have also demonstrated a variety of reasoning abilities such as problem-solving, tool use, and using top-down perception to transfer a trained skill (Schluessel, 2015). For example, vermiculate river stingrays (Potamotrygon castexi) have been observed using water as a tool to solve the problem of extracting food from an open-ended tube (Kuba et al., 2009). Fuss et al. (2014) observed that grey bamboo sharks (Chiloscyllium griseum) were capable of using top-down cognitive processes to perceive illusory contours - a visual illusion wherein preconceptions based on memories or neural wiring create the perception of a particular shape which is not in fact present in presented visual stimuli. Having been trained to recognize the complete shapes, thus establishing the memory of said shape being rewarding, the grey bamboo sharks were then presented with Kaniza figures - visual stimuli that portray the illusion of particular shapes through the use of negative space (and see Kaniza, 1974). The sharks were then able to reason that without the exact conditioned stimulus present (the closed square), they should select the closest analog (an open square) to receive the reward, which they did successfully.
Individuals’ belief in animals’ reasoning capacities (i.e., their broad ability to think logically to guide purpose-driven behavior) was predictive of their tolerance, as operationalized behaviorally as willingness to be in close spatial and temporal proximity to these animals in their natural habitat. The effect was not species group-specific; beliefs in sharks’ and stingrays’ reasoning capacities were both predictive of behavioral tolerance for each species group. Interestingly, the belief in animals’ reasoning capacities was the only belief to influence tolerance.
Belief in animals’ reasoning capacities was also predictive of behaviors underlying the support of conservation. Belief in sharks’ and stingrays’ reasoning abilities predicted willingness to donate funds to conservation regardless of species group (both for sharks and rays). However, belief in reasoning capacity was species group-specific for voting and limited only to sharks. The greater the belief in sharks’ reasoning abilities, the more likely to vote in favor of shark conservation only. Voting for stingrays’ conservation was not influenced by the belief in either animals’ reasoning capabilities. Further research is necessary to determine why this belief is so universally predictive for certain behaviors (i.e., all behavioral expressions of tolerance and donating), but is so targeted and species group-specific for other behaviors (i.e., voting).
4.1.4 Belief in emotional abilities
There is evidence to support the assertion that elasmobranchs possess emotional capacities. For example, certain stingray species have been observed consistently socializing in small groups in the wild, most likely in an effort to deter predators (Kanno et al., 2019; Martins et al., 2020). In the laboratory, when given the choice to be alone or to be part of a social pair or group, certain shark species chose to spend time with others (Guttridge et al., 2009). Similarly, in the wild, it has been observed that certain shark species aggregate in social groups (Economakis and Lobel, 1998; Sims et al., 2003), and conform to social hierarchies (Klimley and Nelson, 1981; Jacoby et al., 2010). These traits and examples have been featured in numerous documentaries (i.e., Discovery Channel’s Shark Week, National Geographic’s SharkFest) about sharks, so the public is aware of this research.
Belief in stingrays’ emotional abilities (i.e., their capacity to experience feelings and affects, etc.) was predictive of both behaviors to actively support conservation: voting and donating. Moreover, this effect was not species group-specific; the greater the extent to which stingrays were perceived as feeling and empathetic creatures, the more willing participants were to support both shark and stingray conservation. The belief in stingrays’ emotional capacities was also predictive of participants’ desire to learn more about sharks. Interestingly, this belief influenced two separate behavioral intentions (i.e., desire to know and support of conservation); yet, for one behavior (desire to know), it increased the odds of the outcome for only one animal – and not the corresponding animal. The belief in stingrays’ emotional abilities only increased the desire to learn about sharks, not stingrays themselves.
Furthermore, the belief in sharks’ emotional abilities was not predictive of any behavioral intentions. These findings contrast with recent research which demonstrated that perceptions of sharks as having high communality (i.e., being caring and emotional) were associated with a greater willingness to donate to their conservation (Neves et al., 2021). Such different results could well be due to the populations surveyed in these respective studies. Neves et al. (2021) surveyed a group of 60 undergraduate students, while this study surveyed 380 participants from the general public. Future research is necessary to clarify these equivocal results.
4.2 Subject variables
Age was a significant predictor of participants’ desire to learn more about stingrays. Increases in age were associated with a diminished self-reported desire to learn more about stingrays and their conservation. These findings are interesting as they contrast with previous literature which has found a positive association between age and pro-environmental behavioral intentions (Kim et al., 2013; Masud and Kari, 2015). Such different findings may be due to key differences in sampling and specificity of constructs. For example, Kim et al. (2013) surveyed a narrower age range (18 – 33 years, versus 18 – 82 years in this study) which could therefore arguably be classified as a less representative sample, and asked questions relating to broader climate change, rather than focusing on the conservation of a particular set of animal species groups as was done here. These discrepant findings also suggest that attitudes concerning global conservation versus the conservation of animals may be different, which presents an interesting avenue for future research.
While age was a significant predictor of the desire to know more about stingrays, this was not the case for sharks. This finding could indicate that sharks have a broader appeal as a taxa to all age groups. Such an interpretation is supported by incredibly prevalent and well-publicized shark-centric media events such as National Geographic’s SharkFest and the Discovery Channel’s Shark Week televised programming which occur annually, attracting more than 50 million viewers during Summer 2020, and continuing to attract a growing viewership (Morfoot, 2021).
Sex was a significant predictor of behavioral expressions of tolerance. This effect was observed for tolerance of both groups of animals, and interestingly only at the closer temporal threshold (i.e., 24 hours). Males expressed greater tolerance for both sharks and stingrays when compared to females by reporting a greater willingness to place themselves within the animals’ natural environment (i.e., in the water) within 24 hours of a confirmed sighting of them. These findings are consistent with research concerning sex differences in risk-taking behaviors (Byrnes et al., 1999) and sex differences in self-reported comfort with animals with negative or injurious reputations (Herzog et al., 1991). It is also interesting that sex was exclusively a predictor or tolerance and not for any other behavioral component (i.e., desire to know more, support for conservation).
4.3 Limitations
This work stems from a larger etiological study concerning the stakeholders of California ocean recreation and safety communities. The present analyses are consequently exploratory and quasi-experimental. No previous work has assessed the attitudes of the general public towards the species groups of sharks and stingrays in a population from California. A baseline was consequently needed to afford comparisons across time, as well as across different geographical locales where shark bites or stingray-related injuries are relatively high (e.g., Massachusetts, Hawaii, Florida, Australia, South Africa, etc.). As this first year’s focus was exclusively on establishing a baseline, no experimental manipulation was employed, and the analyses therefore utilized subject variables of interest.
Lower attendance at public beaches in southern California in summer 2021 may have influenced the quantity and composition of the acquired data. Although state-mandated stay-at-home orders due to the COVID-19 Pandemic were lifted prior to the data collection phase, cases of the infectious disease continued to rise in California and remained elevated across the summer months, hampering participant recruitment efforts. As a result of these issues, said baseline admittedly may not constitute a representative sample of the larger population. Moreover, the observed sex differences must also be interpreted with caution given that females outnumbered males in the sample by a 2:1 ratio. The uneven distribution of the sexes is not unexpected as females are more likely to volunteer to participate in medical and social sciences research (Dickinson et al., 2012; Otufowora et al., 2021), especially research utilizing online surveys (Smith, 2008). Future experimental work should recruit participants based on sex and employ equal sample sizes across groups.
Exclusively framing the dependent measures of behavioral intentions as a forced, dichotomous choice was purposeful. As previously mentioned, for certain behaviors, a one-shot decision is an ecologically valid frame for assessing intent to support conservation or not. However, this methodological choice also has the effect of restricting the response range, perhaps curtailing the true amount of variability in intentions. Future work in this program of research incorporates continuous as well as dichotomous responses to better observe variability in behavioral intentions to support conservation.
Finally, certain analyses have missing cases due to the IRB’s stipulation that participants could forgo answering any question. Participants who provided no or insufficient data for analysis of any variable were necessarily excluded for that respective analysis. IRB approval was contingent on offering this non-answer option.
4.4 Recommendations for educational and elasmobranch conservation strategies
Humans’ attitudes concerning animals are a critical prerequisite consideration for the successful promotion of the animals’ conservation and welfare (and see Batt, 2009). This work has extended this understanding to include the cognitive beliefs that underlie these attitudes. These results represent the first occasion wherein extant beliefs about animals’ mental capacities have been shown to influence pro-conservation behavioral intentions. Future research is now needed to specify what factors engender these beliefs about particular taxonomic groups, and specific species within such groups. That being said, certain recommendations can be made here regarding how to go about changing beliefs in a more general sense for the benefit of animal conservation efforts.
Beliefs about sharks are complex, and vary across cultures and time (Neves et al., 2022). Moreover, research shows that these beliefs are often based on misconceptions or inaccurate information (Tsoi et al., 2016). Such may be the case as people are less likely to learn about sharks from their own personal experience than they are via the media (Evans, 2015), which is not held to as rigorous a standard as scientific communication, or - in the case of popular media - can be completely fictitious. Beliefs about stingrays have been neglected in the literature, and this work represents the first time they have been studied. Beliefs therefore vary and can change. The most effective method by which to alter beliefs is through education, ensuring that an individual’s knowledge base is accurate and comprehensive (Murphy and Mason, 2006). Moreover, knowledge is also identified as the only mediating factor in the theoretical framework detailing the psychological mechanisms underlying successful shark conservation (Acuña-Marrero et al., 2018).
To this end, the specific recommendations forwarded here for educational and elasmobranch conservation strategies is to emphasize education and increase knowledge about elasmobranchs, their abilities, roles, and threats. Some specific recommendations for how to modify beliefs (particularly those that may be based on inaccurate or biased information) is to make use of targeted debiasing strategies - methods for presenting and evaluating information to more effectively inform beliefs and consequently guide better judgments (Soll et al., 2015). The first recommendation would be bias inoculation, educating the audience about the nature and prevalence of cognitive biases that affect the general public’s beliefs about elasmobranchs, and providing tools for how to identify and address them (Turk and Salovey, 1986). For example, the base rate fallacy which occurs when an individual places undue weight on the numerator (the specific situation) as opposed to the denominator (the usual state) when making judgments (Bar-Hillel, 1980). In the case of sharks, for example, people become fixated on the number of humans injured or killed by sharks in a year, while neglecting the millions (if not tens of millions) of cases where humans go into the ocean each day and are unharmed.
The second recommendation is to show the consequences of different decisions in a visual form. Data visualization is extremely helpful in terms of presenting large amounts of complex data in a comprehensible way (Sadiku et al., 2016), and has proven an effective method for cognitive debiasing (modifying beliefs; Croskerry et al., 2013). Educational materials for elasmobranch conservation should consequently adopt sparklines (high-resolution infographics with appropriate context for interpretation; Radecki and Medow, 2007) to illustrate the actual risks faced by and posed by different elasmobranch species. Such displays can not only affect beliefs of such elements as they are at present, but can also be used to project certain trends in the future based on conservation decisions made at certain points in time (e.g., comparative estimated population recovery rates based on protective legislation passed today versus 1, 5, or 10 years from now).
The final recommendation would be to favor causal (explanation of why the relationship is true) over non-causal (determination of whether the relationship is true) arguments in educational materials. Research shows that causal arguments are less likely to be discounted (Slusher and Anderson, 1996), result in the greatest change in belief in those individuals whose preexisting beliefs conflict with the advocated argument, and prompt a greater quantity and quality of recall of information (Kardash and Scholes, 1995).
4.5 Conclusions
Behavioral intentions to support conservancy for animals and their habitats through various behaviors (i.e., desiring to know more, tolerance, voting to support and donating to conservation measures) result from a complex interplay of cognitive beliefs, namely the extent to which people perceive a species group as possessing cognitive, reasoning, and affective capacities. This study represents the first work to investigate extant beliefs specifically focused on animals’ mental capacities as predictors of pro-conservation behavioral intentions; moreover, it is also the first study that assessed the general public’s attitudes and beliefs of the stingray species group. This novel approach yielded interesting significant findings. Generally, the desire to learn more was driven by belief in animals’ cognitive awareness and emotional abilities. Tolerance was exclusively predicted by beliefs in animals’ reasoning abilities. Finally, supporting conservation was driven by multiple beliefs, and differed based on the type of supporting behavior (voting or donating). Support via donating was influenced by beliefs in animals’ cognitive and affective capacities, whereas support in the form of voting was affected by beliefs in cognitive and emotional capabilities as well as beliefs in animals’ reasoning abilities. As specified in the results and earlier discussion sections, it is also important to note that these predictive patterns were sometimes consistent across species group (e.g., shark and stingray taxonomic groups’ reasoning abilities consistently predicted tolerance for both species groups), and sometimes were not (e.g., beliefs pertaining to sharks’ emotional capacities and stingrays’ cognitive abilities were not predictive of any behaviors).
Understanding the personal belief structures that underlie conservation behaviors is vital for the formulation, functioning, and continuation of effective, large-scale conservancy efforts to protect the ecosystems of the world. This work represents the first time that such an examination of the effects of attitudes and beliefs on behavioral intentions to support shark and stingray conservation has been conducted on a population from California, an important white shark (Carcharodon carcharias) aggregation site (e.g., Anderson et al., 2021). Findings from this work can aid in the design of educational and promotional materials intended to promote the knowledge of and conservation of animals, especially as it relates to apex predators and other species that humans have been conditioned to fear (Afonso et al., 2020). Research shows that these fears are largely unfounded but persist due to sensationalized popular media portrayals and highly selective media coverage. To be effective, outreach and conservation efforts must therefore concentrate on correcting these misperceptions. Conservationists are consequently encouraged to portray animals factually and with emphasis on their cognitive, reasoning, and emotional capacities (i.e., as thinking, intelligent, feeling, and empathetic) to promote conservation support to the greatest possible extent.
Data availability statement
The datasets presented in this article are not readily available because they are currently restricted per Institutional Review Board stipulations. Access may be granted pending IRB approval. Requests to access the datasets should be directed to R2FicmllbGxhLkhhbmNvY2tAY3N1bGIuZWR1.
Ethics statement
The studies involving human participants were reviewed and approved by California State University, Long Beach Institutional Review Board. The patients/participants provided their informed consent to participate in this study.
Author contributions
GH: This author retains first authorship. KD: This author retains second authorship. DL: This author retains third authorship. CL: This author retains last authorship. All authors contributed to the article and approved the submitted version.
Funding
Support for this work comes from the State of California Shark Beach Safety Program. This funding agency had no involvement in the study design, data collection, analysis or interpretation of data, the writing of this work, or the decision as to the publication outlet.
Acknowledgments
This human-subjects research was approved and conducted under the supervision of the Institutional Review Board of California State University, Long Beach. The authors would like to thank Sydney Roberts, Carmen Machado, Christian Schmitz, Jennifer Truong, Kristen Brown, and Dr. David Illingworth for their assistance in the generation of this work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2023.1178539/full#supplementary-material
References
Achen C. H., Bartels L. M. (2004). Blind retrospection: electoral responses to drought, flu, and shark attacks (Madrid, Spain: Instituto Juan March de Estudios e Investigaciones). doi: 10.1515/9781400888740-007
Acuña-Marrero D., de la Cruz-Modino R., Smith A. N., Salinas-de-León P., Pawley M. D., Anderson M. J. (2018). Understanding human attitudes towards sharks to promote sustainable coexistence. Mar. Policy 91, 122–128. doi: 10.1016/j.marpol.2018.02.018
Afonso A. S., Roque P., Fidelis L., Veras L., Conde A., Maranhão P., et al. (2020). Does lack of knowledge lead to misperceptions? disentangling the factors modulating public knowledge about and perceptions toward sharks. Front. Mar. Sci. 7, 663. doi: 10.3389/fmars.2020.00663
Anderson J. M., Clevenstine A. J., Stirling B. S., Burns E. S., Meese E. N., White C. F., et al. (2021). Non-random co-occurrence of juvenile white sharks (Carcharodon carcharias) at seasonal aggregation sites in southern California. Front. Mar. Sci. 8, 688505. doi: 10.3389/fmars.2021.688505
Bar-Hillel M. (1980). The base-rate fallacy in probability judgments. Acta Psychologica 44 (3), 211–233. doi: 10.1016/0001-6918(80)90046-3
Batt S. (2009). Human attitudes towards animals in relation to species similarity to humans: a multivariate approach. Biosci. Horizons: Int. J. Student Res. 2(2)180–190. doi: 10.1093/biohorizons/hzp021
Bond M. E., Valentin-Albanese J., Babcock E. A., Heithaus M. R., Grubbs R. D., Cerrato R., et al. (2019). Top predators induce habitat shifts in prey within marine protected areas. Oecologia 190, 375–385. doi: 10.1007/s00442-019-04421-0
Bouyoucos I. A., Suski C. D., Mandelman J. W., Brooks E. J. (2017). The energetic, physiological, and behavioral response of lemon sharks (Negaprion brevirostris) to simulated longline capture. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 207, 65–72. doi: 10.1016/j.cbpa.2017.02.023
Braithwaite V. A., Boulcott P. (2007). Pain perception, aversion and fear in fish. Dis. Aquat. organisms 75 (2), 131–138. doi: 10.3354/dao075131
Butterfield M. E., Hill S. E., Lord C. G. (2012). Mangy mutt or furry friend? anthropomorphism promotes animal welfare. J. Exp. Soc. Psychol. 48 (4), 957–960. doi: 10.1016/j.jesp.2012.02.010
Byrnes E. E., Brown C. (2016). Individual personality differences in port Jackson sharks heterodontus portusjacksoni. J. Fish Biol. 89 (2), 1142–1157. doi: 10.1111/jfb.12993
Byrnes J. P., Miller D. C., Schafer W. D. (1999). Gender differences in risk taking: a meta-analysis. Psychol. Bull. 125 (3), 367. doi: 10.1037/0033-2909.125.3.367
Chan A. A. Y. H. (2012). Anthropomorphism as a conservation tool. Biodiver. Conserv. 21, 1889–1892. doi: 10.1007/s10531-012-0274-6
Chandroo K. P., Duncan I. J., Moccia R. D. (2004). Can fish suffer?: perspectives on sentience, pain, fear and stress. Appl. Anim. Behav. Sci. 86 (3-4), 225–250. doi: 10.1016/j.applanim.2004.02.004
Chin M. G., Sims V. K., Clark B., Lopez G. R. (2004). “September. measuring individual differences in anthropomorphism toward machines and animals,” in Proceedings of the human factors and ergonomics society annual meeting, vol. 48. (Sage CA: Los Angeles, CA: SAGE Publications), 1252–1255.
Coghill R. C. (2020). The distributed nociceptive system: a framework for understanding pain. Trends Neurosci. 43 (10), 780–794. doi: 10.1016/j.tins.2020.07.004
Cornwell J. F., Franks B., Higgins E. T. (2014). Truth, control, and value motivations: the “what,””how,” and “why” of approach and avoidance. Front. Syst. Neurosci. 8, 194. doi: 10.3389/fnsys.2014.00194
Croskerry P., Singhal G., Mamede S. (2013). Cognitive debiasing 2: impediments to and strategies for change. Br. Med. J. Qual. Saf. 22 (2), ii65–ii72. doi: 10.1136/bmjqs-2012-001713
Crossley R., Collins C. M., Sutton S. G., Huveneers C. (2014). Public perception and understanding of shark attack mitigation measures in Australia. Hum. Dimensions Wildlife 19 (2), 154–165. doi: 10.1080/10871209.2014.844289
Dickinson E. R., Adelson J. L., Owen J. (2012). Gender balance, representativeness, and statistical power in sexuality research using undergraduate student samples. Arch. Sexual Behav. 41, 325–327. doi: 10.1007/s10508-011-9887-1
Dulvey N. K., Pacoureau N., Rigby C. L., Pollom R. A., Jabado R. W., Ebert D. A., et al. (2021). Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Curr. Biol. 31 (21), 4773–4787. doi: 10.1016/j.cub.2021.08.062
Economakis A. E., Lobel P. S. (1998). Aggregation behavior of the grey reef shark, carcharhibus amblyrhynchos, at Johnston atoll, central pacific ocean. Environ. Biol. Fishes 51, 129–139. doi: 10.1023/A:1007416813214
Evans S. (2015). Shark week and the rise of infotainment in science documentaries. Communication Res. Rep. 32 (3), 265–271. doi: 10.1080/08824096.2015.1052903
Fuss T., Bleckmann H., Schluessel V. (2014). The brain creates illusions not just for us: turns out sharks (Chiloscyllium griseum) can ‘see the magic’ as well. Front. Neural Circuits 8, 24. doi: 10.3389/fncir.2014.00024
Gardiner J. M., Whitney N. M., Hueter R. E. (2015). Smells like home: the role of olfactory cues in the homing behavior of blacktip sharks, carcharhinus limbatus. Integr. Comp. Biol. 55 (3), 495–506. doi: 10.1093/icb/icv087
Guttridge T. L., Brown C. (2014). Learning and memory in the port Jackson shark, heterodontus portusjacksoni. Anim. Cogn. 17, 415–425. doi: 10.1007/s10071-013-0673-4
Guttridge T. L., Myrberg A. A., Porcher I. F., Sims D. W., Krause J. (2009). The role of learning in shark behaviour. Fish Fisheries 10 (4), 450–469. doi: 10.1111/j.1467-2979.2009.00339.x
Herzog H. A. Jr., Betchart N. S., Pittman R. B. (1991). Gender, sex role orientation, and attitudes toward animals. Anthrozoös 4 (3), 184–191. doi: 10.2752/089279391787057170
Higgs M. J., Bipin S., Cassaday H. J. (2020). Man's best friends: attitudes towards the use of different kinds of animal depend on belief in different species' mental capacities and purpose of use. R. Soc. Open Sci. 7 (2), 191162. doi: 10.1098/rsos.191162
Hills A. M. (1995). Empathy and belief in the mental experience of animals. Anthrozoös 8 (3), 132–142. doi: 10.2752/089279395787156347
International Shark Attack File World attack frequency rates. Available at: https://www.floridamuseum.ufl.edu/shark-attacks/trends/frequency-rates/world/.
Jacoby D. M. P., Busawon D. S., Sims D. W. (2010). Sex and social networking: the influence of male presence on social structure of female shark groups. Behav. Ecol. 21, 808–818. doi: 10.1093/beheco/arq061
Jorgensen S. J., Micheli F., White T. D., Van Houtan K. S., Alfaro-Shigueto J., Andrzejaczek S., et al. (2022). Emergent research and priorities for shark and ray conservation. Endangered species Res. 47, 171–203. doi: 10.3354/esr01169
Kajiura S. M., Fitzgerald T. P. (2009). Response of juvenile scalloped hammerhead sharks to electrical stimuli. Zoology 112 (4), 241–250. doi: 10.1016/j.zool.2008.07.001
Kaniza G. (1974). Contours without gradients or cognitive contours? Giornale Italiano di Psicol. 1, 93–113.
Kanno S., Schlaff A. M., Heupel M. R., Simpfendorfer C. A. (2019). Stationary video monitoring reveals habitat use of stingrays in mangroves. Mar. Ecol. Prog. Ser. 621, 155–168. doi: 10.3354/meps12977
Kardash C. M., Scholes H. J. (1995). Effects of preexisting beliefs and repeated readings on belief change, comprehension, and recall of persuasive text. Contemp. Educ. Psychol. 20, 201–221. doi: 10.1006/ceps.1995.1013
Kim S., Jeong S. H., Hwang Y. (2013). Predictors of pro-environmental behaviors of American and Korean students: the application of the theory of reasoned action and protection motivation theory. Sci. Communication 35 (2), 168–188. doi: 10.1177/1075547012441692
Kimber J. A., Sims D. W., Bellamy P. H., Gill A. B. (2014). Elasmobranch cognitive ability: using electroreceptive foraging behaviour to demonstrate learning, habituation and memory in a benthic shark. Anim. Cogn. 17, 55–65. doi: 10.1007/s10071-013-0637-8
Klimley A. P., Nelson D. R. (1981). Schooling of the scalloped hammerhead shark, sphyrna lewini, and subsurface irradiance, temperature, bathymetry, and geomagnetic field. Fishery Bull. 79, 356–360.
Kuba M. J., Byrne R. A., Burghardt G. M. (2009). A new method for studying problem solving and tool use in stingrays (Potamotrygon castexi). Anim. Cogn. 13, 507–513. doi: 10.1007/s10071-009-0301-5
Lambert F. N., Treberg J. R., Anderson W. G., Brandt C., Evans A. N. (2018). The physiological stress response of the Atlantic stingray (Hypanus sabinus) to aerial exposure. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 219, 38–43. doi: 10.1016/j.cbpa.2018.02.009
López de la Lama R., de la Puente S., Riveros J. C. (2018). Attitudes and misconceptions towards sharks and shark meat consumption along the Peruvian coast. PloS One 13 (8), e0202971. doi: 10.1371/journal.pone.0202971
Lowe C. G., Moss G. J., Hoisington G. IV, Vaudo J. J., Cartamil D. P., Marcotte M. M., et al. (2007). Caudal spine shedding periodicity and site fidelity of round stingrays, urobatis halleri (Cooper), at seal beach, California: implications for stingray-related injury management. Bulletin South. California Acad. Sci. 106 (1), 16–26. doi: 10.3160/0038-3872(2007)106[16:CSSPAS]2.0.CO;2
Martins A. P. B., Huepel M. R., Bierwagen S. L., Chin A., Simpfendorfer C. A. (2020). Tidal-diel patterns of movement, activity and habitat use by juvenile mangrove whiprays using towed-float GPS telemetry. Mar. Freshw. Res. 72, 534. doi: 10.1071/MF20078
Masud M. M., Kari F. B. (2015). Community attitudes towards environmental conservation behaviour: an empirical investigation within MPAs, Malaysia. Mar. Policy 52, 138–144. doi: 10.1016/j.marpol.2014.10.015
McGowan D. W., Kajiura S. M. (2009). Electroreception in the euryhaline stingray, dasyatis sabina. J. Exp. Biol. 212, 1544–1552. doi: 10.1242/jeb.025247
Mickle M. F., Pieniazek R., Stasso J. J., Higgs D. M. (2022). Anthropogenic sounds induce escape behaviour in southern stingrays hypanus americanus. Mar. Ecol. Prog. Ser. 694, 125–132. doi: 10.3354/meps14100
Morfoot A. (2021). Inside the shark week vs. sharkfest battle for cable and streaming viewers. Available at: https://variety.com/2021/tv/news/shark-week-vs-sharkfest-battle-cable-streaming-1235015047/.
Murphy P. K., Mason L. (2006). “Changing knowledge and beliefs,” in Handbook of educational psychology. Eds. Alexander P., Winne P. (New York: Simon & Schuster/Macmillan), 305–324.
Naylor W., Parsons E. C. M. (2018). An online survey of public knowledge, attitudes, and perceptions toward whales and dolphins, and their conservation. Front. Mar. Sci. 5, 153. doi: 10.3389/fmars.2018.00153
Neff C. (2015). The jaws effect: how movie narratives are used to influence policy responses to shark bites in Western Australia. Aust. J. Political Sci. 50 (1), 114–127. doi: 10.1080/10361146.2014.989385
Neves J., Giger J. C., Piçarra N., Alves V., Almeida J. (2021). Social representations of sharks, perceived communality, and attitudinal and behavioral tendencies towards their conservation: an exploratory sequential mixed approach. Mar. Policy 132, 104660. doi: 10.1016/j.marpol.2021.104660
Neves J., McGinnies T., Giger J.-C. (2022). Changing trends: beliefs and attitudes toward sharks and implications for conservation. Ethnobiol. Conserv. 11, 1–11. doi: 10.15451/ec2022-05-11.11-1-11
Newton K. C., Kajiura S. M. (2020). The yellow stingray (Urobatis jamaicensis) can use magnetic field polarity to orient in space and solve a maze. Mar. Biol. 167, 1–10. doi: 10.1007/s00227-019-3643-9
Otufowora A., Liu Y., Young H., Egan K. L., Varma D. S., Striley C. W., et al. (2021). Sex differences in willingness to participate in research based on study risk level among a community sample of African americans in north central Florida. J. Immigrant Minority Health 23, 19–25. doi: 10.1007/s10903-020-01015-4
Pacoureau N., Rigby C. L., Kyne P. M., Sherley R. B., Winker H., Carlson J. K., et al. (2021). Half a century of global decline in oceanic sharks and rays. Nature 589 (7843), 567–571. doi: 10.1038/s41586-020-03173-9
Parsons G. R., Carlson J. K. (1998). Physiological and behavioral responses to hypoxia in the bonnethead shark, sphyrna tiburo: routine swimming and respiratory regulation. Fish Physiol. Biochem. 19 (2), 189–196. doi: 10.1023/A:1007730308184
Pepin-Neff C., Wynter T. (2018). Shark bites and shark conservation: an analysis of human attitudes following shark bite incidents in two locations in Australia. Conserv. Lett. 11 (2), e12407. doi: 10.1111/conl.12407
Radecki R. P., Medow M. A. (2007). Cognitive debiasing through sparklines in clinical data displays. Paper presented at AMIA Annu. Symposium Chicago IL 11, 1085.
Rose J. D. (2002). The neurobehavioral nature of fishes and the question of awareness and pain. Rev. fisheries Sci. 10 (1), 1–38. doi: 10.1080/20026491051668
Rose J. D. (2007). Anthropomorphism and ‘mental welfare’ of fishes. Dis. Aquat. organisms 75 (2), 139–154. doi: 10.3354/dao075139
Sadiku M., Shadare A. E., Musa S. M., Akujuobi C. M., Perry R. (2016). Data visualization. Int. J. Eng. Res. Advanced Technol. 2 (12), 11–16.
Schluessel V. (2015). Who would have thought that ‘Jaws’ also has brains? cognitive functions in elasmobranchs. Anim. Cogn. 18, 19–37. doi: 10.1007/s10071-014-0762-z
Schluessel V., Bleckmann H. (2005). Spatial memory and orientation strategies in the elasmobranch potamotrygon motoro. J. Comp. Physiol. A 191, 695–706. doi: 10.1007/s00359-005-0625-9
Schluessel V., Herzog H., Scherpenstein M. (2015). Seeing the forest before the trees–spatial orientation in freshwater stingrays (Potamotrygon motoro) in a hole-board task. Behav. processes 119, 105–115. doi: 10.1016/j.beproc.2015.08.001
Sergio F., Newton I. A. N., Marchesi L., Pedrini P. (2006). Ecologically justified charisma: preservation of top predators delivers biodiversity conservation. J. Appl. Ecol. 43 (6), 1049–1055. doi: 10.1111/j.1365-2664.2006.01218.x
Shiffman D. S., Bittick S. J., Cashion M. S., Colla S. R., Coristine L. E., Derrick D. H., et al. (2020). Inaccurate and biased global media coverage underlies public misunderstanding of shark conservation threats and solutions. Iscience 23 (6), 101205. doi: 10.1016/j.isci.2020.101205
Siciliano A. M., Kajiura S. M., Long J. H. Jr., Porter M. E. (2013). Are you positive? electric dipole polarity discrimination in the yellow stingray, urobatis jamaicensis. Biol. Bull. 225 (2), 85–91. doi: 10.1086/BBLv225n2p85
Sims D. W., Southall E. J., Richardson A. J., Reid P. C., Metcalfe J. D. (2003). Seasonal movements and behaviour of basking sharks from archival tagging: no evidence of winter hibernation. Mar. Ecol. Prog. Ser. 248, 187–196. doi: 10.3354/meps248187
Slusher M. P., Anderson C. A. (1996). Using causal persuasive arguments to change beliefs and teach new information: the mediating role of explanation availability and evaluation bias in the acceptance of knowledge. J. Educ. Psychol. 88 (1), 110–122. doi: 10.1037/0022-0663.88.1.110
Smith W. G. (2008). Does gender influence online survey participation?: a record-linkage analysis of university faculty online survey response behavior. ERIC Document Reprod. Service No. ED 501717 (1), 1–21.
Soll J., Milkman K., Payne J. (2015). “A user’s guide to debiasing,” in Wiley-Blackwell Handbook of judgment and decision making. Eds. Keren G., Wu G. (New York: NY: Blackwell), 924–951.
Towner A. V., Watson R. G. A., Kock A. A., Papstamatiou Y., Sturup M., Gennari E., et al. (2022). Fear at the top: killer whale predation drives white shark absence at south africa’s largest aggregation site. Afr. J. Mar. Sci. 44 (1), 1–14. doi: 10.2989/1814232X.2022.2066723
Tsoi K. H., Chan S. Y., Lee Y. C., Ip B. H. Y., Cheang C. C. (2016). Shark conservation: an educational approach based on children’s knowledge and perceptions toward sharks. PLOSOne 11 (9), e0163406. doi: 10.1371/journal.pone.0163406
Keywords: attitudes, conservation, public perceptions, sharks, stingrays, tolerance
Citation: Hancock GM, Dudley KD, Long D and Lowe CG (2023) An etiological examination of behavioral intentions to support shark and stingray conservancy: the effects of beliefs in elasmobranchs’ cognitive and affective capacities. Front. Mar. Sci. 10:1178539. doi: 10.3389/fmars.2023.1178539
Received: 02 March 2023; Accepted: 19 May 2023;
Published: 08 June 2023.
Edited by:
Yaisel Juan Borrell Pichs, University of Oviedo, SpainReviewed by:
Natascha Wosnick, Federal University of Paraná, BrazilLaura Miralles, University of Oviedo, Spain
Copyright © 2023 Hancock, Dudley, Long and Lowe. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gabriella M. Hancock, R2FicmllbGxhLkhhbmNvY2tAY3N1bGIuZWR1