 Ian V. Hughes
Ian V. Hughes Peter R. Girguis
Peter R. Girguis
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
PERSPECTIVE article
Front. Mar. Sci. , 24 May 2023
Sec. Microbial Symbioses
Volume 10 - 2023 | https://doi.org/10.3389/fmars.2023.1167803
Many bivalves and gastropods from marine reducing environments such as deep ocean hydrothermal vents and seeps host chemosynthetic bacteria in a nutritional symbiosis. Despite their functional similarities, the distribution of chemosymbiosis in these two mollusk classes is surprisingly uneven: the number of bivalve species known to host chemosynthetic symbionts is more than twenty times that of gastropods, and chemosymbiotic bivalves are reported from a far greater diversity of marine habitats. Here we explore the potential drivers for this trend, including but not limited to physiological differences, habitat characteristics, and sampling bias. Sampling bias likely contributes to the magnitude of the observed discrepancy, but we posit that the phenomenon itself reveals how intrinsic (e.g. morphology) and extrinsic (e.g. organic matter availability) factors might have shaped the distribution of extant gastropod and bivalve associations. These observations also serve as an impetus for increasing investigation into gastropods and other mollusks from chemically reducing environments to better understand the evolution and ecology of chemosymbiosis among molluscan hosts.
Chemosynthetic symbioses between eukaryotes and chemoautotrophic microbes have been reported in diverse metazoan phyla and in numerous marine environments. In nutritional chemosynthetic symbiosis, bacterial symbionts oxidize reduced chemical substrates, e.g., sulfide, to harness energy for carbon fixation. Symbionts then translocate fixed organic carbon to the host or are digested by the host (Cavanaugh et al., 2006; Dubilier et al., 2008; Sogin et al., 2020). A large portion of metazoan host diversity is represented by mollusks, specifically gastropods and bivalves. Chemosymbiotic mollusks are abundant in many chemically reducing environments, including deep sea hydrothermal vents and hydrocarbon seeps (Sasaki et al., 2010), wood and whale falls (Smith et al., 1989; Duperron, 2010), continental shelf sediments (Cary et al., 1989), and many shallow water sulfidic sedimented environments (Cavanaugh, 1983).
Notably, bivalves and gastropods have independently acquired chemosynthetic symbionts in multiple lineages. However, and unexpectedly, this adaptation appears unequally distributed between these groups, both with respect to the number of species represented and the diversity of chemosynthetic habitats where they are found. Among Bivalvia, there over 600 described species of chemosymbiotic bivalves, across at least six families (Distel, 1998; Taylor and Glover, 2010) that span a suite of marine reducing habitats, including less conventional chemosynthetic environments, like coral reef sediments (Dekker and Goud, 1994) and sewage outfalls (Burd et al., 2013). Among Gastropoda, the total number of known chemosymbioses is an order of magnitude fewer than in Bivalvia, although there are representative species in four of the six subclasses. Further, unlike their bivalve counterparts, only two potential gastropod chemosymbioses occur in shallow water environments (Smriglio and Mariottini, 2002; Judge and Haszprunar, 2014), while the rest are reported from hydrothermal vents and related deep-water environments. These observed patterns are surprising given that Gastropoda is an extraordinarily diverse clade and contains three-fold more marine species than Bivalvia (species are an incomplete proxy for diversity but provide a means of illustrating the disparities between these two classes; WoRMS Editorial Board, 2023).
Gastropods and bivalves share similarities in basic anatomy, physiology, and life history, which begs the question as to what factor(s) account for this disparity. Here we briefly summarize chemosymbiosis in bivalves, review known gastropod chemosymbioses, then consider differences between bivalves and gastropods that may account for the observed disparities in the diversity of chemosymbioses between these two molluscan classes.
Chemosymbiosis is well documented in many bivalve groups (see Taylor and Glover (2010) for review). The bivalve families that contain chemosymbiotic taxa are Lucinidae, Thyasiridae, Solemyidae, Vesicomyidae, Teredinidae, and Mytilidae (there is circumstantial evidence for chemosymbiosis in several members of Nucinellidae and at least one species in the bivalve family Basterotiidae, though nutritional chemoautotrophy has not yet been demonstrated; Oliver and Taylor 2012; Oliver, 2013). Most recorded bivalve chemosymbionts belong to Gammaproteobacteria (Sogin et al., 2021), however the nature of symbioses varies considerably in metabolism, transmission modes, and host-symbiont nutrient exchange.
Lucinidae is the most species rich group of chemosymbiotic bivalves, with over 400 species described worldwide (WoRMS Editorial Board, 2023). To date, all lucinids are obligately chemosymbiotic (Taylor and Glover, 2010). Although lucinids occur at hydrothermal vents and methane seeps (Sibuet and Olu, 1998; Glover et al., 2004), much of the diversity is found in shallow-water sediments rich in sulfide (Taylor and Glover, 2006).
Thyasirids are the second most species rich group of chemosymbiotic bivalves, however there are considerably fewer studies on the diversity and nature of Thyasiridae symbioses (Taylor et al., 2007). Chemosymbiosis is not obligate for all Thyasiridae (Taylor et al., 2007), and reliance on chemosynthetically fixed carbon varies substantially among the chemosymbioses (Dando and Spiro, 1993). One thyasirid species hosts endosymbionts (Fujiwara et al., 2001), whereas others appear to exclusively host extracellular symbionts on the gill surface (Southward, 1986).
Solemyidae is a unique family of bivalves with three genera (Solemya, Acharax and Petrasma) from diverse reducing habitats (e.g., Stanley, 1970; Neulinger et al., 2006; Coan et al., 2000). Most solemyids supplement symbiosis with heterotrophic filter feeding, though there are exceptions, such as Solemya reidi, that lacks the capacity to filter-feed as an adult (Felbeck, 1983; Stewart and Cavanaugh, 2006). The Nucinellidae are another bivalve family in the order Solemyida with taxa found in a range of marine habitats, including deep sea sediments (Sasaki et al., 2005) and tropical seagrass beds (Glover and Taylor, 2013). Chemosymbiosis has been inferred in this group as some nucinellids have a reduced or absent gut (Kuznetsov, 1984) and bacteriocytes are present in the gill tissue of at least two species (Oliver and Taylor, 2012). However, empirical assessments of chemoautotrophic activity in these taxa are wanting.
Vesicomyidae are another family of chemosymbiotic bivalves, primarily occurring at hydrothermal vents, hydrocarbon seeps, and whale falls (Bennett et al., 1994; von Cosel and Salas, 2001). There are over 100 described species in the family, and all members studied host chemosymbionts (Fisher, 1990; Krylova et al., 2010). Many vesicomyids have been shown to use their foot to uptake reduced sulfur compounds and transport them to symbionts in the gills (Arp et al., 1984).
Teredinidae are well known for their heterotrophic bacterial symbiosis, which aids in breaking down wood (O’Connor et al., 2014), though at least one species hosts sulfide-oxidizing symbionts in the gills. While other teredinids bore into wood, the giant shipworm, Kuphus polythalamia, occurs in muddy sediments and utilizes thioautotrophic symbionts in a nutritional chemosymbiosis (Distel et al., 2017).
Finally, within the family Mytilidae, the subfamily Bathymodiolinae is a group of mussels found in hydrothermal vents, methane seeps, and wood and whale falls (Duperron et al., 2009). Bathymodiolins are primarily epibenthic and are often densely packed in chemosymbiotic habitats. Although the majority of species host chemosymbionts either intracellularly or extracellularly, at least one species in the genus Idas is reported as a predator on other mollusks and appears asymbiotic, likely representing a secondary loss of chemosymbionts (Ockelmann and Dinesen, 2011; Rodrigues et al., 2015). Bathymodiolins host symbionts with a variety of metabolic pathways, including methane, complex hydrocarbons, sulfide, and hydrogen metabolisms (Childress et al., 1986; Duperron et al., 2005; Petersen et al., 2011; Rubin-Blum et al., 2017).
As previously mentioned, evidence for chemosymbiosis has been reported in taxa from four of the six gastropod subclasses, though relatively less work has been done on the evolution and diversity of chemosymbiotic gastropods (reviewed in Table 1). Among caenogastropods, the sister genera Alviniconcha and Ifremeria host chemosynthetic endosymbionts in the gills (Stein et al., 1988; Endow and Ohta, 1989; Windoffer and Giere, 1997). Both taxa are found at hydrothermal vents where they can form large aggregations around vent effluent (Bouchet and Warén, 1991; Desbruyères et al., 1994). Alviniconcha and Ifremeria co-occur around Pacific hydrothermal vents, but their symbionts likely rely on different chemically-reduced compounds that might alleviate niche competition (Henry et al., 2008; Waite et al., 2008; Beinart et al., 2015).
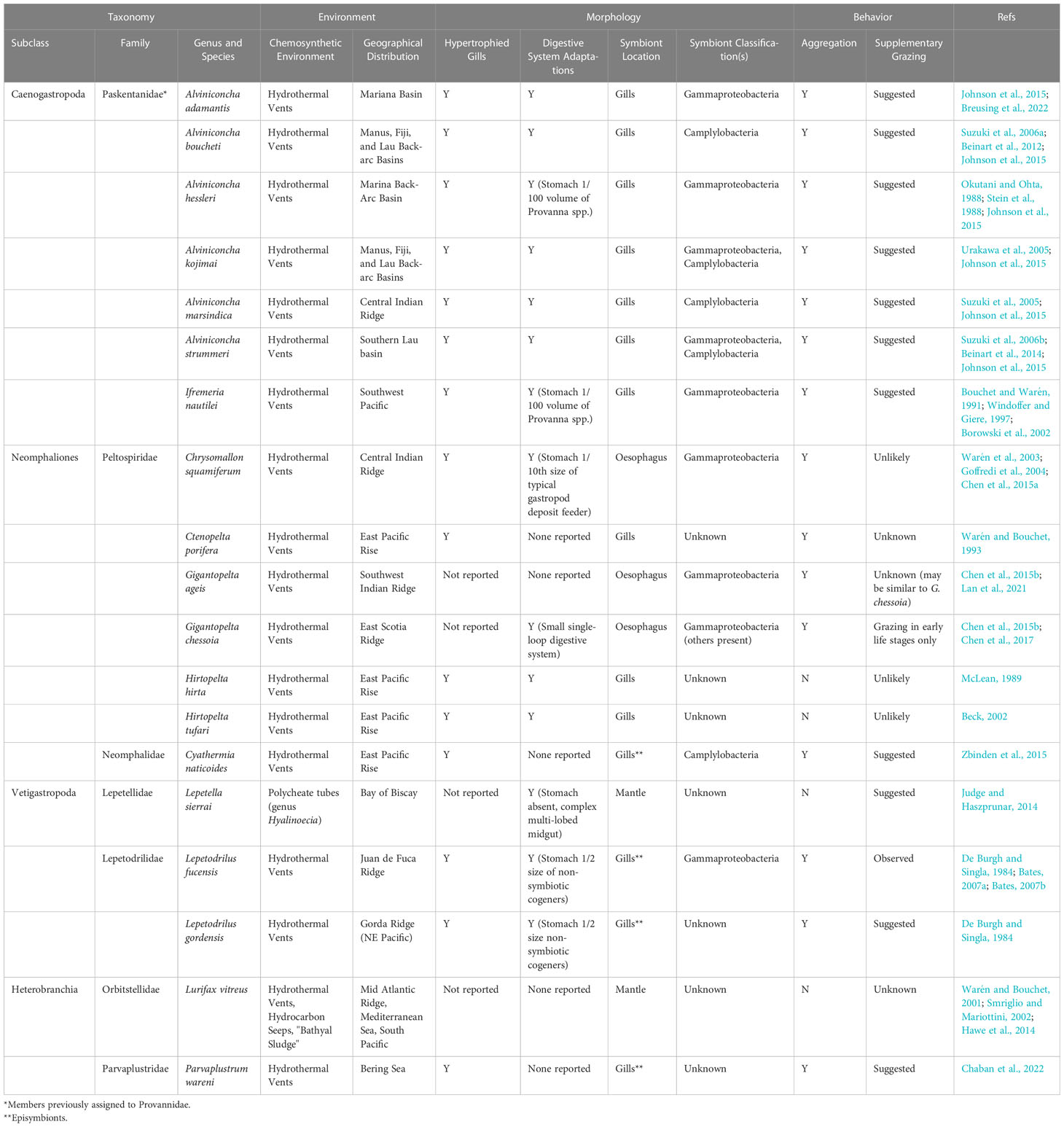
Table 1 Summary of known gastropod chemosymbioses.
The Neomphaliones family Peltospiridae contains several chemosymbiotic taxa from hydrothermal vent sites in the Pacific, Southern and Indian oceans. Perhaps the most striking member of this family is Chrysomallon squamiferum, or the “scaly foot snail”, so named as it secretes iron-rich sclerites on the foot (Warén et al., 2003). C. squamiferum and members of the peltospirid genus Gigantopelta host sulfur-oxidizing bacterial endosymbionts in an enlarged esophagus (Goffredi et al., 2004; Chen et al., 2017). Other peltospirids from vents, namely the Ctenopelta and Hirtopelta, have been suggested as chemosymbiotic due to a reduced digestive tract (Warén and Bouchet, 1993), and, at least in the case of Hirtopelta, hypertrophied gills (Fretter, 1989; McLean, 1989; Warén and Bouchet, 1993). Bacteriocytes have also been observed in the gill tissue of H. tufari, though the identity of these potential symbionts is unknown (Beck, 2002). Finally, another Neomphaliones gastropod suggested as chemosymbiotic is Cyathermia naticoides. This species is endemic to the East Pacific Rise vents and is often found in abundance on Riftia tubes (Warén and Bouchet, 1989). Campylobacteria have been reported extracellularly on gill tissue of this species in association with lysosomes, indicating possible endocytotic digestion (Zbinden et al., 2015).
Lepetodrilus is a Vetigastopoda genus found at a diversity of hydrothermal vent communities. Of the 16 described species, two sister taxa from the Juan de Fuca Ridge are described as chemosymbiotic: L. fucensis and L. gordensis (De Burgh and Singla, 1984; McLean 1988; Johnson et al., 2006). Both species host bacteria on the external surface of the gills, whereas other Lepetodrilus species appear devoid of dense gill microbes (Bates, 2006). L. fucensis was among the first vent gastropods to be described as chemosymbiotic, and De Burgh and Singla (1984) implicate endocytosis and lysosomal digestion of episymbionts as a type of “symbiont farming”. Around hydrothermal vent effluent, Lepetodrilus fucensis can form aggregated stacks of individuals with a density of over 300,000 individuals m-2 (Bates et al., 2005). L. fucensis also filter feed and graze to supplement their nutrition (Bates, 2007a).
Bacteriocytes have also been described in the mantle of Lepetella sierrai, a gastropod found in the Bay of Biscay in association with empty tubes of polychaetes in the genus Hyalinoecia (Dantart and Luoue, 1994). It is hypothesized that these polychaete tubes provide an anoxic, sulfidic environment that supports chemosynthesis (Judge and Haszprunar, 2014). Further molecular or physiological work is needed to confirm the nature of this potential non-vent chemosymbiosis.
Two heterobranch gastropods have also been suggested as chemosymbiotic. The first, Parvaplustrum wareni, occurs at hydrothermal vents in the Bering Sea in densities of up to 31,000 individuals/m-2 (Chaban et al., 2022). Evidence for chemosymbiosis in this taxon comes from the dense aggregates of bacteria that occur externally on gill filaments. Lurifax vitreus is an orbitestellid gastropod originally described from hydrothermal vents on the Mid Atlantic Ridge (Warén and Bouchet, 2001). Bacteriocytes have been identified in the mantle of this gastropod based on TEM microscopy (Hawe et al., 2014). Individuals of L. vitreus have also been described from non-vent environments in Mediterranean Sea (Smriglio and Mariottini, 2002; Giuste and Sbrana, 2012), though these identifications were made from shells collected by dredges, and soft parts were not examined.
As described above, to our knowledge there are at least 600 likely and confirmed bivalve chemosymbioses that span a wide array of chemically reducing environments. In contrast, to our knowledge there are approximately 19 likely and confirmed gastropod chemosymbioses that are found in far fewer chemically reducing environments, namely select hydrothermal vents and, putatively, Polychaetae tubes and Mediterranean sediments (Smriglio and Mariottini, 2002; Judge and Haszprunar, 2014). It is highly likely that sampling bias towards bivalves has a marked influence on the realized disparity. That said, even in chemosynthetic environments where both bivalves and gastropods are present and well-studied (e.g., seagrass beds and hydrothermal vents), chemosymbiotic bivalves are far more diverse. We thus raise the question of what other factors may have collectively resulted in this phenomenon. In the paragraphs below we consider the roles of environmental medium (including sulfide toxicity), physiology, habitat productivity, motility, and our biases in the definition of chemosymbioses.
Chemosymbiosis often occurs at the interface between oxic and anoxic environs (Stewart et al., 2005). At hydrothermal vents, chemically reduced substrates are replete in the vent fluids that often emanate from cracks in the basaltic crust (or emerge from polymetallic sulfide deposits). At mud flats, seagrass sediments, and many other habitats that harbor chemosymbioses, chemically reduced substrates are abundant in the anoxic sediments, resulting from anaerobic microbial processes such as sulfate reduction (Preisler et al., 2007). At such localities, chemosymbiotic hosts must bridge the redox gradient through behavioral and morphological adaptations. Here, bivalves may have an advantage as many are infaunal and use their siphon to access oxygenated surface water while using a highly adapted foot to tap into sulfide-rich sediment porewaters to support their symbionts (Dando and Southward, 1986; Zanzerl and Dufour, 2017). Indeed, much of the diversity of chemosymbiotic bivalves is represented by infaunal, or semi-infaunal groups including the majority of lucinids, thyasirids and solemyids (Taylor and Glover, 2010). The acquisition of chemosynthetic symbionts in infaunal bivalve groups may have facilitated widespread diversification of species and habitats, contributing to the bivalve/gastropod disparity (Seilacher, 1990). However, there are also many gastropod groups that are infaunal and occur in chemosynthetic sediments. For example, burrowing members of Strombidae and Cerithiidae are found in sediments alongside lucinid clams (Ibrahim et al., 2015; Lima et al., 2017), yet chemosymbiosis has not been reported in either gastropod group. Perhaps bivalves are advantaged in this scenario, as they may be better suited to supplement symbiont-derived nutrition with heterotrophic filter feeding when symbionts alone cannot support host metabolism, as the case for the majority of molluscan chemosymbioses (Duplessis et al., 2004; van der Geest et al., 2014).
Chemosymbiotic organisms are faced with the challenges of accessing both oxidants (typically oxygen) and reductants such as sulfide, methane, and hydrogen (Childress and Girguis, 2011). Sulfide is fundamental to many chemosymbioses, however it is toxic to metazoans and presents a major challenge for chemosymbiotic hosts writ large (Powell and Somero, 1986; Grieshaber and Völkel, 1998; Tobler et al., 2016). The bivalve/gastropod disparity begs the question as to whether gastropods face greater physiological or morphological challenges in hosting chemoautotrophic symbionts. One hypothesis is that some gastropods, such as those without an operculum, may be more vulnerable to sulfide toxicity than bivalves and thus are less likely to host intracellular chemosymbionts. However, there are many species of gastropods that lack intracellular symbionts (e.g., Vermeij, 1973; Warén and Bouchet, 2009) and live in environments with elevated sulfide (where sulfide concentrations are comparable to those of hydrothermal vents; Jorgensen and Revsbech, 1983; Gartman et al., 2011). Given the abundance and diversity of gastropods in sulfidic environments, both with and without endosymbionts, it is unlikely that sulfide sensitivity alone is driving this observed trend in chemosymbiotic diversity.
There are also morphological differences that may contribute to the observed gastropod/bivalve disparity. As mentioned, many chemosymbiotic mollusks host symbionts on or within the ctenidia (Table 1). Although gill structure varies considerably within classes (Owen, 1978), many bivalves have large demibranch gills, compared to the relatively smaller gills of many gastropods, which are located beneath the shell. Large gills may be advantageous for bivalve chemosymbioses, as it may provide larger surface area for symbiont colonization. There are a number of extant chemosymbiotic gastropods, however, that successfully host symbionts in hypertrophied gills, and several others that host symbionts in the esophagus (e.g. Gigantopelta chessoia; Chen et al., 2017) or mantle (e.g. Lurifax vitreus; Hawe et al., 2014), indicating that gill morphology does not inherently preclude chemosymbiosis in gastropods.
Many shallow water chemosynthetic environments also harbor a robust photosynthetic community (e.g., mangroves and seagrass beds), which plays a role in driving chemosynthesis via the decay of organic plant material by sulfate-reducing bacteria (Jørgensen, 1977). In these environments, both bivalves and gastropods could take advantage of photosynthetic and chemosynthetic food sources. Motile gastropods have the potential to graze both surface photosynthetic and subsurface chemosynthetic bacteria, without having to expend the extra energy and oxygen required for hosting intracellular symbionts (Childress and Girguis, 2011). Though filter and deposit feeding are both used by many chemosymbiotic bivalves (Allen, 1958; Duplessis et al., 2004; Zanzerl et al., 2019), hosting intracellular chemoautotrophic symbionts can be an efficient way to take advantage of carbon fixed via chemosynthesis for less motile groups. In shallow reducing environments, both bivalves and gastropods exploit carbon fixed by different pathways, but motility in gastropods may allow more efficient heterotrophy without the physiological impacts and costs of hosting chemosynthetic symbionts.
Although most hydrothermal vent ecosystems lack extensive photosynthetic inputs, they can be comparable in productivity to shallow water ecosystems and support microbial mats at high densities (Gaill et al., 1997). Many vent gastropods are motile grazers (Gaudron et al., 2015), but perhaps there are unique attributes of hydrothermal vents that account for the high concentration of gastropod chemosymbioses. Vent flows are highly dynamic, with fluid flow regimes changing over many time scales (from minutes to days; Johnson et al., 1988). This environment may provide highly motile gastropods with a competitive advantage for access to sufficient reductants and oxidants for their symbionts. In western Pacific vents, Alviniconcha and Ifremeria form large aggregations around vent effluent, moving around to access reduced and oxidized substrates as needed (Desbruyères et al., 1994). Chemosymbiotic gastropods also have the ability, to some extent, to move out of environments when they become less suitable (e.g. higher temperatures or lower oxygen or sulfide concentrations). These attributes of vents may contribute to the prominence of motile gastropods in some vent communities, but do not preclude the overall success of bivalves in hydrothermal ecosystems.
We must, of course, consider the impact of sampling bias. The discovery of chemosymbiotic metazoan taxa was relatively recent and the number of documented chemosymbiotic taxa has grown sporadically, particularly among mollusks. For example, the first members of Lucinidae were described by Linnaeus (1758), however it wasn’t until the 1980s that lucinids were found to host chemosynthetic bacteria (Cavanaugh, 1983; Fisher and Hand, 1984). As such, it may be easy to attribute much of gastropod/bivalve disparity to sampling bias in favor of the bivalves. That being said, existing evidence from well-studied sites such as the Eastern Pacific Rise reminds us that this observed disparity may be a biological reality. Nevertheless, despite over four decades of research on chemosynthetic symbioses, some new chemosymbiotic gastropods have been described recently, including two species of Gigantopelta (Chen et al., 2015b), Chrysomallon squamiferum (Chen et al., 2015a), and Parvaplustrum wareni (Chaban et al., 2022). Further, much of the evidence for chemosymbiosis in previously described gastropods has also been reported in the last decade (Hawe et al., 2014; Judge and Haszprunar, 2014; Zbinden et al., 2015).
Finally, we acknowledge the challenges of how we define chemosymbiosis. There is a vibrant discussion in the community about what constitutes a symbiosis (Martin and Schwab, 2013), and to date there is no one simple “litmus test” for determining if an organism is chemosymbiotic. Even among widely accepted chemosymbioses, there is considerable variation in the nature of host/symbiont dynamics. Further investigation into gastropod holobionts from both deep and shallow-water chemosynthetic environments may reveal previously unknown chemosymbiotic relationships.
Chemosymbiosis has evolved multiple times in diverse lineages of bivalve and gastropod mollusks. Despite physiological and functional similarities between these two groups, bivalves have a far higher diversity of chemosymbiotic taxa, both in terms of species richness, and the breadth of marine reducing habitats occupied. This observed trend presents and interesting framework for investigating the evolution and diversification of chemosymbiotic metazoan hosts. If this trend is robust, this disparity reflects evolutionary, ecological, and/or physiological differences between these two lineages that favors chemosymbiosis in extant bivalves. That said, these observations underscore the need for continued sampling to reveal the undiscovered diversity of gastropod hosts, as well as the need to continue to refine our notions of what constitutes a chemosymbiosis.
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
IH and PG conceptualized this study. IH drafted the manuscript, with edits and comments from PG. All authors have read and approve the final manuscript.
This work was supported by a grant to PG from the Gordon and Betty Moore Foundation (grant #9208). This work was also supported by the department of Organismic and Evolutionary Biology at Harvard University.
We thank Stephane Hourdez and members of the Girguis laboratory for helpful discussions and comments. We would also like to thank our reviewer and editor Sébastien Duperron for helpful feedback which improved the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Allen J. A. (1958). On the basic form and adaptations to habitat in the lucinacea (Eulamellibranchia). Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 241 (684), 421–484. doi: 10.1098/rstb.1958.0010
Arp A. J., Childress J. J., Fisher C. R. Jr. (1984). Metabolic and blood gas transport characteristics of the hydrothermal vent bivalve calyptogena magnifica. Physiol. Zoology 57 (6), 648–662. doi: 10.1086/physzool.57.6.30155991
Bates A. E. (2006). Population and feeding characteristics of hydrothermal vent gastropods along environmental gradients with a focus on bacterial symbiosis hosted by lepetodrilus fucensis (Vetigastropoda). University of Victoria, BC, Canada.
Bates A. E. (2007a). Feeding strategy, morphological specialisation and presence of bacterial episymbionts in lepetodrilid gastropods from hydrothermal vents. Mar. Ecol. Prog. Ser. 347, 87–99. doi: 10.3354/meps07020
Bates A. E. (2007b). Persistence, morphology, and nutritional state of a gastropod hosted bacterial symbiosis in different levels of hydrothermal vent flux. Mar. Biol. 152 (3), 557–568. doi: 10.1007/s00227-007-0709-x
Bates A. E., Tunnicliffe V., Lee R. W. (2005). Role of thermal conditions in habitat selection by hydrothermal vent gastropods. Marine Ecol. 305, 1–15. doi: 10.3354/meps305001
Beck L. A. (2002). Hirtopelta tufari sp. n., a new gastropod species from hot vents at the East pacific rise (21 s) harbouring endocytosymbiotic bacteria in its gill (Gastropoda: rhipidoglossa: peltospiridae). Archiv für Molluskenkunde 130, 249–257. doi: 10.1127/arch.moll/130/2002/249
Beinart R. A., Gartman A., Sanders J. G., Luther G. W., Girguis P. R. (2015). The uptake and excretion of partially oxidized sulfur expands the repertoire of energy resources metabolized by hydrothermal vent symbioses. Proc. R. Soc. B: Biol. Sci. 282 (1806), 20142811. doi: 10.1098/rspb.2014.2811
Beinart R. A., Nyholm S. V., Dubilier N., Girguis P. R. (2014). Intracellular O ceanospirillales inhabit the gills of the hydrothermal vent snail a lviniconcha with chemosynthetic, γ-p roteobacterial symbionts. Environ. Microbiol. Rep. 6 (6), 656–664. doi: 10.1111/1758-2229.12183
Beinart R. A., Sanders J. G., Faure B., Sylva S. P., Lee R. W., Becker E. L., et al. (2012). Evidence for the role of endosymbionts in regional-scale habitat partitioning by hydrothermal vent symbioses. Proc. Natl. Acad. Sci. 109 (47), E3241–E3250. doi: 10.1073/pnas.1202690109
Bennett B. A., Smith C. R., Glaser B., Maybaum H. L. (1994). Faunal community structure of a chemoautotrophic assemblage on whale bones in the deep northeast Pacific Ocean. Mar. Ecol. Prog. Ser. 205–223.
Borowski C., Giere O., Krieger J., Amann R., Dubilier N. (2002). New aspects of the symbiosis in the provannid snail ifremeria nautilei from the north Fiji back arc basin. Cahiers biologie Mar. 43 (3-4), 321–324.
Bouchet P., Warén A. (1991). Ifremeria nautilei, nouveau gastéropode d'évents hydrothermaux, probablement associé à des bactéries symbiotiques. Comptes Rendus l’Académie Des. Sciences Paris 312, 495–501.
Breusing C., Klobusnik N. H., Hauer M. A., Beinart R. A. (2022). Genome assembly of the chemosynthetic endosymbiont of the hydrothermal vent snail alviniconcha adamantis from the Mariana arc. G3 12, 10, jkac220. doi: 10.1093/g3journal/jkac220
Burd B., Macdonald T., Bertold S. (2013). The effects of wastewater effluent and river discharge on benthic heterotrophic production, organic biomass and respiration in marine coastal sediments. Mar. Pollut. Bull. 74 (1), 351–363. doi: 10.1016/j.marpolbul.2013.06.029
Cary S. C., Vetter R. D., Felbeck H. (1989). Habitat characterization and nutritional strategies of the endosymbiont-bearing bivalve lucinoma. Mar. Ecol. Prog. Ser. 55, 31–45. doi: 10.3354/meps055031
Cavanaugh C. M. (1983). Symbiotic chemoautotrophic bacteria in marine invertebrates from sulphide-rich habitats. Nature 302 (5903), 58–61. doi: 10.1038/302058a0
Cavanaugh C. M., McKiness Z. P., Newton I. L., Stewart F. J. (2006). Marine chemosynthetic symbioses. Prokaryotes 1, 475–507. doi: 10.1007/0-387-30741-9_18
Chaban E. M., Schepetov D. M., Ekimova I. A., Nekhaev I. O., Chеrnyshev A. V. (2022). The first record of the family parvaplustridae (Gastropoda, heterobranchia) from hydrothermal vent fields of the piip volcano, Bering Sea with the description of a new species of the genus parvaplustrum. Deep Sea Res. Part II: Topical Stud. Oceanography 202, 105135. doi: 10.1016/j.dsr2.2022.105135
Chen C., Linse K., Copley J. T., Rogers A. D. (2015a). The ‘scaly-foot gastropod’: a new genus and species of hydrothermal vent-endemic gastropod (Neomphalina: peltospiridae) from the Indian ocean. J. Molluscan Stud. 81 (3), 322–334. doi: 10.1093/mollus/eyv013
Chen C., Linse K., Roterman C. N., Copley J. T., Rogers A. D. (2015b). A new genus of large hydrothermal vent-endemic gastropod (Neomphalina: peltospiridae). Zoological J. Linn. Soc. 175 (2), 319–335. doi: 10.1111/zoj.12279
Chen C., Uematsu K., Linse K., Sigwart J. D. (2017). By more ways than one: rapid convergence at hydrothermal vents shown by 3D anatomical reconstruction of gigantopelta (Mollusca: neomphalina). BMC Evolutionary Biol. 17 (1), 1–19. doi: 10.1186/s12862-017-0917-z
Childress J. J., Fisher C. R., Brooks J. M., Kennicutt M. C., Bidigare R. A. A. E., Anderson A. E. (1986). A methanotrophic marine molluscan (Bivalvia, mytilidae) symbiosis: mussels fueled by gas. Science 233 (4770), 1306–1308. doi: 10.1126/science.233.4770.1306
Childress J. J., Girguis P. R. (2011). The metabolic demands of endosymbiotic chemoautotrophic metabolism on host physiological capacities. J. Exp. Biol. 214 (2), 312–325. doi: 10.1242/jeb.049023
Coan E. V., Scott P. V., Bernard F. R. (2000). Bivalve seashells of western North America. Santa Barbara, CA: Santa Barbara Museum of Natural History.
Dando P. R., Southward A. J. (1986). Chemoautotrophy in bivalve molluscs of the genus thyasira. J. Mar. Biol. Assoc. United Kingdom 66 (4), 915–929. doi: 10.1017/S0025315400048529
Dando P. R., Spiro B. (1993). Varying nutritional dependence of the thyasirid bivalves thyasira sarsi and t. equalis on chemoautotrophic symbiotic bacteria, demonstrated by isotope ratios of tissue carbon and shell carbonate. Mar. Ecology-Progress Ser. 92, 151–151. doi: 10.3354/meps092151
Dantart L., Luoue A. (1994). Cocculiniformia and lepetidae (Gastropoda: archaeogastropoda) from Iberian waters. J. Molluscan Stud. 60 (3), 277–313. doi: 10.1093/mollus/60.3.277
De Burgh M. E., Singla C. L. (1984). Bacterial colonization and endocytosis on the gill of a new limpet species from a hydrothermal vent. Mar. Biol. 84 (1), 1–6. doi: 10.1007/BF00394520
Dekker H., Goud J. (1994). Review of the living indo-West-Pacific species of divaricella sens auct. with descriptions of two new species and a summary of species from other regions. Vita Marina 42 (4), 115–136.
Desbruyères D., Alayse-Danet A. M., Ohta S. (1994). Deep-sea hydrothermal communities in southwestern pacific back-arc basins (the north Fiji and lau basins): composition, microdistribution and food web. Mar. Geology 116 (1-2), 227–242. doi: 10.1016/0025-3227(94)90178-3
Distel D. L. (1998). Evolution of chemoautotrophic endosymbioses in bivalves. Bioscience 48 (4), 277–286. doi: 10.2307/1313354
Distel D. L., Altamia M. A., Lin Z., Shipway J. R., Han A., Forteza I., et al. (2017). Discovery of chemoautotrophic symbiosis in the giant shipworm kuphus polythalamia (Bivalvia: teredinidae) extends wooden-steps theory. Proc. Natl. Acad. Sci. 114 (18), E3652–E3658. doi: 10.1073/pnas.1620470114
Dubilier N., Bergin C., Lott C. (2008). Symbiotic diversity in marine animals: the art of harnessing chemosynthesis. Nat. Rev. Microbiol. 6 (10), 725–740. doi: 10.1038/nrmicro1992
Duperron S. (2010). “The diversity of deep-sea mussels and their bacterial symbioses,” in The vent and seep biota (Dordrecht: Springer), 137–167.
Duperron S., Lorion J., Samadi S., Gros O., Gaill F. (2009). Symbioses between deep-sea mussels (Mytilidae: bathymodiolinae) and chemosynthetic bacteria: diversity, function and evolution. Comptes rendus biologies 332 (2-3), 298–310. doi: 10.1016/j.crvi.2008.08.003
Duperron S., Nadalig T., Caprais J. C., Sibuet M., Fiala-Médioni A., Amann R., et al. (2005). Dual symbiosis in a bathymodiolus sp. mussel from a methane seep on the Gabon continental margin (Southeast atlantic): 16S rRNA phylogeny and distribution of the symbionts in gills. Appl. Environ. Microbiol. 71 (4), 1694–1700. doi: 10.1128/AEM.71.4.1694-1700.2005
Duplessis M. R., Dufour S. C., Blankenship L. E., Felbeck H., Yayanos A. A. (2004). Anatomical and experimental evidence for particulate feeding in lucinoma aequizonata and parvilucina tenuisculpta (Bivalvia: lucinidae) from the Santa Barbara basin. Mar. Biol. 145 (3), 551–561. doi: 10.1007/s00227-004-1350-6
Endow K., Ohta S. (1989). The symbiotic relationship between bacteria and a mesogastropod snail, alviniconcha hessleri, collected from hydrothermal vents of the Mariana back-arc basin. Bull. Japanese Soc. Microbial Ecol. 3 (2), 73–82. doi: 10.1264/microbes1986.3.73
Felbeck H. (1983). Sulfide oxidation and carbon fixation by the gutless clam solemya reidi: an animal-bacteria symbiosis. J. Comp. Physiol. 152 (1), 3–11. doi: 10.1007/BF00689721
Fisher C. R. (1990). Chemoautotrophic and methanotrophic symbioses in marine invertebrates. Rev. Aquat. Sci. 2, 399–436.
Fisher M. R., Hand S. C. (1984). Chemoautotrophic symbionts in the bivalve lucina floridana from seagrass beds. Biol. Bull. 167 (2), 445–459. doi: 10.2307/1541289
Fretter V. (1989). The anatomy of some new archaeogastropod limpets (Superfamily peltospiracea) from hydrothermal vents. J. Zoology 218 (1), 123–169. doi: 10.1111/j.1469-7998.1989.tb02530.x
Fujiwara Y., Kato C., Masui N., Fujikura K., Kojima S. (2001). Dual symbiosis in the cold-seep thyasirid clam maorithyas hadalis from the hadal zone in the Japan trench, western pacific. Mar. Ecol. Prog. Ser. 214, 151–159. doi: 10.3354/meps214151
Gaill F., Shillito B., Ménard F., Goffinet G., Childress J. J. (1997). Rate and process of tube production by the deep-sea hydrothermal vent tubeworm riftia pachyptila. Mar. Ecol. Prog. Ser. 148, 135–143. doi: 10.3354/meps148135
Gartman A., Yücel M., Madison A. S., Chu D. W., Ma S., Janzen C. P., et al. (2011). Sulfide oxidation across diffuse flow zones of hydrothermal vents. Aquat. Geochemistry 17 (4), pp.583–pp.601. doi: 10.1007/s10498-011-9136-1
Gaudron S. M., Marqué L., Thiébaut E., Riera P., Duperron S., Zbinden M. (2015). How are microbial and detrital sources partitioned among and within gastropods species at e ast p acific r ise hydrothermal vents? Mar. Ecol. 36, 18–34. doi: 10.1111/maec.12260
Giuste F., Sbrana C. (2012). Lurifax vitreus Warén & Bouchet, 2001 (Gastropoda, Orbitestellidae), a new record for deep waters of the Tuscan Archipelago (Tyrrhenian Sea, Italy). Biodivers. J. 3 (1), 91–92.
Glover E. A., Taylor J. D. (2013). A new shallow water species of nucinella from the Philippines (Bivalvia: protobranchia: nucinellidae), member of a tropical seagrass chemosynthetic community. Nautilus 127 (3), 101–106.
Glover E. A., Taylor J. D., Rowden A. A. (2004). Bathyaustriella thionipta, a new lucinid bivalve from a hydrothermal vent on the kermadec ridge, new Zealand and its relationship to shallow-water taxa (Bivalvia: lucinidae). J. Molluscan Stud. 70 (3), 283–295. doi: 10.1093/mollus/70.3.283
Goffredi S. K., Warén A., Orphan V. J., Van Dover C. L., Vrijenhoek R. C. (2004). Novel forms of structural integration between microbes and a hydrothermal vent gastropod from the Indian ocean. Appl. Environ. Microbiol. 70 (5), 3082–3090. doi: 10.1128/AEM.70.5.3082-3090.2004
Grieshaber M. K., Völkel S. (1998). Animal adaptations for tolerance and exploitation of poisonous sulfide. Annu. Rev. Physiol. 60 (1), 33–53. doi: 10.1146/annurev.physiol.60.1.33
Hawe A., Gensler H., Haszprunar G. (2014). Bacteriocytes in the mantle cavity of lurifax vitreus warén & bouchet 2001 (Orbitestellidae): the first case among heterobranch gastropoda. J. Molluscan Stud. 80 (3), 337–340. doi: 10.1093/mollus/eyu018
Henry M. S., Childress J. J., Figueroa D. (2008). Metabolic rates and thermal tolerances of chemoautotrophic symbioses from lau basin hydrothermal vents and their implications for species distributions. Deep Sea Res. Part I: Oceanographic Res. Papers 55 (5), 679–695. doi: 10.1016/j.dsr.2008.02.001
Ibrahim A. R., Fouda M. M., Abdou N. (2015). Macro-invertebrates fauna of mangrove ecosystem in south Sinai, Egypt. Int. J. 3, 1577–1594.
Johnson K. S., Childress J. J., Beehler C. L. (1988). Short-term temperature variability in the rose garden hydrothermal vent field: an unstable deep-sea environment. Deep Sea Res. Part A. Oceanographic Res. Papers 35 (10-11), 1711–1721. doi: 10.1016/0198-0149(88)90045-3
Johnson S. B., Warén A., Tunnicliffe V., Dover C. V., Wheat C. G., Schultz T. F., et al. (2015). Molecular taxonomy and naming of five cryptic species of alviniconcha snails (Gastropoda: abyssochrysoidea) from hydrothermal vents. Systematics Biodiversity 13 (3), 278–295. doi: 10.1080/14772000.2014.970673
Johnson S. B., Young C. R., Jones W. J., Warén A., Vrijenhoek R. C. (2006). Migration, isolation, and speciation of hydrothermal vent limpets (Gastropoda; lepetodrilidae) across the blanco transform fault. Biol. Bull. 210 (2), 140–157. doi: 10.2307/4134603
Jørgensen B. B. (1977). Bacterial sulfate reduction within reduced microniches of oxidized marine sediments. Mar. Biol. 41 (1), 7–17. doi: 10.1007/BF00390576
Jørgensen B. B., Revsbech N. P. (1983). Colorless sulfur bacteria, beggiatoa spp. and thiovulum spp., in O2 and H2S microgradients. Appl. Environ. Microbiol. 45 (4), 1261–1270. doi: 10.1128/aem.45.4.1261-1270.1983
Judge J., Haszprunar G. (2014). The anatomy of lepetella sierrai (Vetigastropoda, lepetelloidea): implications for reproduction, feeding, and symbiosis in lepetellid limpets. Invertebrate Biol. 133 (4), 324–339. doi: 10.1111/ivb.12064
Krylova E. M., Sahling H., Janssen R. (2010). Abyssogena: a new genus of the family vesicomyidae (Bivalvia) from deep-water vents and seeps. J. Molluscan Stud. 76 (2), 107–132. doi: 10.1093/mollus/eyp052
Kuznetsov A. P. (1984). “On gutless protobranchia (Bivalvia),” in Nauchnye doklady vysshei shkoly, biologicheskie nauki, 2, 39–49).
Lan Y., Sun J., Chen C., Sun Y., Zhou Y., Yang Y., et al. (2021). Hologenome analysis reveals dual symbiosis in the deep-sea hydrothermal vent snail gigantopelta aegis. Nat. Commun. 12 (1), 1–15. doi: 10.1038/s41467-021-21450-7
Lima S. F. B., Lucena R. A., Santos G. M., Souza J. W., Christoffersen M. L., Guimarães C. R., et al. (2017). Inventory of mollusks from the estuary of the paraíba river in northeastern Brazil. Biota Neotropica 17. doi: 10.1590/1676-0611-bn-2016-0239
Martin B. D., Schwab E. (2013). Current usage of symbiosis and associated terminology. Int. J. Biol. 5 (1), 32. doi: 10.5539/ijb.v5n1p32
McLean J. H. (1989). New archaeogastropod limpets from hydrothermal vents: new family peltospiridae, new superfamily peltospiracea. Zoologica Scripta 18 (1), 49–66. doi: 10.1111/j.1463-6409.1989.tb00123.x
McLean J. H. (1988). New archaeogastropod limpets from hydrothermal vents; superfamily Lepetodrilacea I. Systematic descriptions. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 319 (1192), 1–32. doi: 10.1098/rstb.1988.0031
Neulinger S. C., Sahling H., Süling J., Imhoff J. F. (2006). Presence of two phylogenetically distinct groups in the deep-sea mussel acharax (Mollusca: bivalvia: solemyidae). Mar. Ecol. Prog. Ser. 312, 161–168. doi: 10.3354/meps312161
Ockelmann K. W., Dinesen G. E. (2011). Life on wood–the carnivorous deep-sea mussel idas argenteus (Bathymodiolinae, mytilidae, bivalvia). Mar. Biol. Res. 7 (1), 71–84. doi: 10.1080/17451001003714504
O’Connor R. M., Fung J. M., Sharp K. H., Benner J. S., McClung C., Cushing S., et al. (2014). Gill bacteria enable a novel digestive strategy in a wood-feeding mollusk. Proc. Natl. Acad. Sci. 111 (47), E5096–E5104. doi: 10.1073/pnas.1413110111
Okutani T., Ohta S. (1988). A new gastropod mollusk associated with hydrothermal vents in the Mariana back-arc basin, Western pacific. Venus (Japanese J. Malacology) 47 (1), 1–9. doi: 10.18941/venusjjm.47.1_1
Oliver P. G. (2013). Description of atopomya dolobrata gen. et sp. nov.: first record of bacterial symbiosis in the saxicavellinae (Bivalvia). J. Conchology 41 (3), 359–367.
Oliver P. G., Taylor J. D. (2012). Bacterial symbiosis in the nucinellidae (Bivalvia: solemyida) with descriptions of two new species. J. Molluscan Stud. 78 (1), 81–91. doi: 10.1093/mollus/eyr045
Owen G. (1978). Classification and the bivalve gill. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 284 (1001), 377–385. doi: 10.1098/rstb.1978.0075
Petersen J. M., Zielinski F. U., Pape T., Seifert R., Moraru C., Amann R., et al. (2011). Hydrogen is an energy source for hydrothermal vent symbioses. Nature 476 (7359), 176–180. doi: 10.1038/nature10325
Powell M. A., Somero G. N. (1986). Adaptations to sulfide by hydrothermal vent animals: sites and mechanisms of detoxification and metabolism. Biol. Bull. 171 (1), 274–290. doi: 10.2307/1541923
Preisler A., De Beer D., Lichtschlag A., Lavik G., Boetius A., Jørgensen B. B. (2007). Biological and chemical sulfide oxidation in a beggiatoa inhabited marine sediment. ISME J. 1 (4), 341–353. doi: 10.1038/ismej.2007.50
Rodrigues C. F., Laming S. R., Gaudron S. M., Oliver G., Le Bris N., Duperron S. (2015). A sad tale: has the small mussel idas argenteus lost its symbionts? Biol. J. Linn. Soc. 114 (2), 398–405. doi: 10.1111/bij.12431
Rubin-Blum M., Antony C. P., Borowski C., Sayavedra L., Pape T., Sahling H., et al. (2017). Short-chain alkanes fuel mussel and sponge cycloclasticus symbionts from deep-sea gas and oil seeps. Nat. Microbiol. 2 (8), 1–11. doi: 10.1038/nmicrobiol.2017.93
Sasaki T., Okutani T., Fujikura K. (2005). Molluscs from hydrothermal vents and cold seeps in Japan: a review of taxa recorded in twenty recent years (1984-2004). Venus (Journal Malacological Soc. Japan) 64 (3-4), 87–133. doi: 10.18941/venus.64.3-4_87
Sasaki T., Warén A., Kano Y., Okutani T., Fujikura K. (2010). “Gastropods from recent hot vents and cold seeps: systematics, diversity and life strategies.” in The Vent and Seep Biota. Topics in Geobiology (Dordrecht: Springer). 33, 169–254. doi: 10.1007/978-90-481-9572-5_7
Seilacher A. (1990). Aberrations in bivalve evolution related to photo-and chemosymbiosis. Historical Biol. 3 (4), 289–311. doi: 10.1080/08912969009386528
Sibuet M., Olu K. (1998). Biogeography, biodiversity and fluid dependence of deep-sea cold-seep communities at active and passive margins. Deep Sea Res. Part II: Topical Stud. Oceanography 45 (1-3), 517–567. doi: 10.1016/S0967-0645(97)00074-X
Smith C. R., Kukert H., Wheatcroft R. A., Jumars P. A., Deming J. W. (1989). Vent fauna on whale remains. Nature 341 (6237), 27–28. doi: 10.1038/341027a0
Smriglio C., Mariottini P. (2002). Lurifax vitreus warén & bouchet 2001 (Gastropoda, orbitestellidae), first report from Western Mediterranean Sea. Bollettino Malacologico 38 (1/4), 45–47.
Sogin E. M., Kleiner M., Borowski C., Gruber-Vodicka H. R., Dubilier N. (2021). Life in the dark: phylogenetic and physiological diversity of chemosynthetic symbioses. Annu. Rev. Microbiol. 75, 695–718. doi: 10.1146/annurev-micro-051021-123130
Sogin E. M., Leisch N., Dubilier N. (2020). Chemosynthetic symbioses. Curr. Biol. 30 (19), R1137–R1142. doi: 10.1016/j.cub.2020.07.050
Southward E. C. (1986). Gill symbionts in thyasirids and other bivalve molluscs. J. Mar. Biol. Assoc. United Kingdom 66 (4), 889–914. doi: 10.1017/S0025315400048517
Stanley S. M. (1970). Relation of shell form to life habits of the bivalvia (Mollusca). Geological Soc. America 125. doi: 10.1130/MEM125-p1
Stein J. L., Cary S. C., Hessler R. R., Vetter R. D., Felbeck H., Ohta S., et al. (1988). Chemoautotrophic symbiosis in a hydrothermal vent gastropod. Biol. Bull. 174 (3), 373–378. doi: 10.2307/1541963
Stewart F. J., Cavanaugh C. M. (2006). Bacterial endosymbioses in solemya (Mollusca: bivalvia)–model systems for studies of symbiont–host adaptation. Antonie Van Leeuwenhoek 90 (4), 343–360. doi: 10.1007/s10482-006-9086-6
Stewart F. J., Newton I. L., Cavanaugh C. M. (2005). Chemosynthetic endosymbioses: adaptations to oxic–anoxic interfaces. Trends Microbiol. 13 (9), 439–448. doi: 10.1016/j.tim.2005.07.007
Suzuki Y., Kojima S., Sasaki T., Suzuki M., Utsumi T., Watanabe H., et al. (2006a). Host-symbiont relationships in hydrothermal vent gastropods of the genus alviniconcha from the southwest pacific. Appl. Environ. Microbiol. 72 (2), 1388–1393. doi: 10.1128/AEM.72.2.1388-1393.2006
Suzuki Y., Kojima S., Watanabe H., Suzuki M., Tsuchida S., Nunoura T., et al. (2006b). Single host and symbiont lineages of hydrothermal-vent gastropods ifremeria nautilei (Provannidae): biogeography and evolution. Mar. Ecol. Prog. Ser. 315, 167–175. doi: 10.3354/meps315167
Suzuki Y., Sasaki T., Suzuki M., Nogi Y., Miwa T., Takai K., et al. (2005). Novel chemoautotrophic endosymbiosis between a member of the epsilonproteobacteria and the hydrothermal-vent gastropod alviniconcha aff. hessleri (Gastropoda: provannidae) from the Indian ocean. Appl. Environ. Microbiol. 71 (9), 5440–5450. doi: 10.1128/AEM.71.9.5440-5450.2005
Taylor J. D., Glover E. A. (2006). Lucinidae (Bivalvia)–the most diverse group of chemosymbiotic molluscs. Zoological J. Linn. Soc. 148 (3), 421–438. doi: 10.1111/j.1096-3642.2006.00261.x
Taylor J. D., Glover E. A. (2010). “Chemosymbiotic bivalves,” in The vent and seep biota (Dordrecht: Springer), 107–135.
Taylor J. D., Williams S. T., Glover E. A. (2007). Evolutionary relationships of the bivalve family thyasiridae (Mollusca: bivalvia), monophyly and superfamily status. J. Mar. Biol. Assoc. United Kingdom 87 (2), 565–574. doi: 10.1017/S0025315407054409
Tobler M., Passow C. N., Greenway R., Kelley J. L., Shaw J. H. (2016). The evolutionary ecology of animals inhabiting hydrogen sulfide–rich environments. Annu. Rev. Ecology Evolution Systematics 47, 239–262. doi: 10.1146/annurev-ecolsys-121415-032418
Urakawa H., Dubilier N., Fujiwara Y., Cunningham D. E., Kojima S., Stahl D. A. (2005). Hydrothermal vent gastropods from the same family (Provannidae) harbour ε-and γ-proteobacterial endosymbionts. Environ. Microbiol. 7 (5), 750–754. doi: 10.1111/j.1462-2920.2005.00753.x
van der Geest M., Sall A. A., Ely S. O., Nauta R. W., Van Gils J. A., Piersma T. (2014). Nutritional and reproductive strategies in a chemosymbiotic bivalve living in a tropical intertidal seagrass bed. Mar. Ecol. Prog. Ser. 501, 113–126. doi: 10.3354/meps10702
Vermeij G. J. (1973). Molluscs in mangrove swamps: physiognomy, diversity, and regional differences. Systematic Zoology 22 (4), 609–624. doi: 10.2307/2412965
von Cosel R., Salas C. (2001). Vesicomyidae (Mollusca: bivalvia) of the genera vesicomya, waisiuconcha, isorropodon and callogonia in the eastern Atlantic and the Mediterranean. Sarsia 86 (4-5), 333–366. doi: 10.1080/00364827.2001.10425523
Waite T. J., Moore T. S., Childress J. J., Hsu-Kim H., Mullaugh K. M., Nuzzio D. B., et al. (2008). Variation in sulfur speciation with shellfish presence at a lau basin diffuse flow vent site. J. Shellfish Res. 27 (1), 163–168. doi: 10.2983/0730-8000(2008)27[163:VISSWS]2.0.CO;2
Warén A., Bengtson S., Goffredi S. K., Van Dover C. L. (2003). A hot-vent gastropod with iron sulfide dermal sclerites. Science 302 (5647), 1007–1007. doi: 10.1126/science.1087696
Warén A., Bouchet P. (1989). New gastropods from East pacific hydrothermal vents. Zoologica Scripta 18 (1), 67–102. doi: 10.1111/j.1463-6409.1989.tb00124.x
Warén A., Bouchet P. (1993). New records, species, genera, and a new family of gastropods from hydrothermal vents and hydrocarbon seeps. Zoologica Scripta 22 (1), 1–90. doi: 10.1111/j.1463-6409.1993.tb00342.x
Warén A., Bouchet P. (2001). Gastropoda And monoplacophora from hydrothermal vents and seeps; new taxa and records. veliger 44, 116–231.
Warén A., Bouchet P. (2009). New gastropods from deep-sea hydrocarbon seeps off West Africa. Deep Sea Res. Part II: Topical Stud. Oceanography 56 (23), 2326–2349. doi: 10.1016/j.dsr2.2009.04.013
Windoffer R., Giere O. (1997). Symbiosis of the hydrothermal vent gastropod ifremeria nautilei (Provannidae) with endobacteria-structural analyses and ecological considerations. Biol. Bull. 193 (3), 381–392. doi: 10.2307/1542940
Zanzerl H., Dufour S. C. (2017). The burrowing behaviour of symbiotic and asymbiotic thyasirid bivalves. J. Conchology 42, 299–308.
Zanzerl H., Salvo F., Jones S. W., Dufour S. C. (2019). Feeding strategies in symbiotic and asymbiotic thyasirid bivalves. J. Sea Res. 145, 16–23. doi: 10.1016/j.seares.2018.12.005
Keywords: Bivalvia, chemosynthesis, Gastropoda, Mollusca, symbiosis
Citation: Hughes IV and Girguis PR (2023) A molluscan class struggle: exploring the surprisingly uneven distribution of chemosymbiosis among two major mollusk groups. Front. Mar. Sci. 10:1167803. doi: 10.3389/fmars.2023.1167803
Received: 16 February 2023; Accepted: 12 May 2023;
Published: 24 May 2023.
Edited by:
Sébastien Duperron, Muséum National d’Histoire Naturelle, FranceReviewed by:
Suzanne Dufour, Memorial University of Newfoundland, CanadaCopyright © 2023 Hughes and Girguis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ian V. Hughes, aWh1Z2hlc0BnLmhhcnZhcmQuZWR1; Peter R. Girguis, cGdpcmd1aXNAb2ViLmhhcnZhcmQuZWR1
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.