 Xiaoling Liu1
Xiaoling Liu1 Guangmei Wang
Guangmei Wang Guangxuan Han
Guangxuan Han- 1CAS Key Laboratory of Coastal Environmental Processes and Ecological Remediation, Yantai Institute of Coastal Zone Research (YIC), Chinese Academy of Sciences (CAS), Shandong Key Laboratory of Coastal Environmental Processes, YICCAS, Yantai, China
- 2Key Laboratory of Forest Ecosystem Process of Beijing, Beijing Forestry University, Beijing, China
- 3Administration Bureau of the Yellow River Delta National Nature Reserve, Dongying, China
The availability and stoichiometry ratio of nitrogen (N) and phosphorus (P) play vital roles in plant trophic dynamics and primary production. However, the responses of these plant traits to varying N and P supplies remain largely unclear for supratidal wetland herbs. Here, we conducted a 4-year field manipulation experiment in a supratidal wetland in the Yellow River Delta. The changes in aboveground biomass, leaf N and P concentrations and N:P ratios of two dominant herbs (Suaeda glauca and Phragmites australis) were examined at 3 overall nutrient supply levels (low, medium and high) combined with 3 N:P supply ratios (5:1, 15:1 and 45:1). The results showed that the leaf trophic dynamics of the two dominant species rely on the overall supply level as well as on the N:P supply ratio, while the aboveground biomass of both species was only significantly influenced by the overall supply level. With the increase in supply level, S. glauca gained an advantage over P. australis in aboveground biomass competition. The leaf N and P concentrations of both species raised with the respective increasing nutrient inputs, and N:P improved with the increasing supply ratio. The leaf stoichiometry of S. glauca was more strongly influenced by the various N and P supplies than that of P. australis. Specifically, the gap of nutrient contents between the two species widened as nutrient availability improved, with the dominance of S. glauca increasing while that of P. australis decreasing. This species-specific response may explain the altered aboveground biomass of the two species. Our findings suggested that changing the N and P supply can potentially influence primary productivity by changing leaf nutrient status, indirectly affecting the shifts in plant dominance and community composition in supratidal wetland ecosystems.
1 Introduction
Nitrogen (N) and phosphorus (P) are essential nutrients controlling plant growth and primary production (Elser et al., 2007; Sardans and Peñuelas, 2012). In terrestrial ecosystems, the increase in N deposition due to fossil fuel burning and anthropogenic fertilizer input has not been paralleled by a proportional increase in P inputs (Venterink and Güsewell, 2010; Peñuelas et al., 2012), leading to changes in the ambient N and P supply status from both aspects of overall supply level and relative supply ratio (Elser et al., 2009; Carnicer et al., 2015). Normally, plants respond to these changes at metabolic and physiological levels (Elser et al., 2010; Wang et al., 2018; Chen et al., 2020) and further alter the elemental composition and growth rate of plants (Sardans and Peñuelas, 2012). As responses are frequently species-specific (Mayor et al., 2014), patterns of plant competition and dominant species in the plant community may be changed (Bobbink et al., 2010).
Plants can optimize nutrient acquisition and conservation strategies to acclimate to altered nutrient availability and elemental stoichiometry (Yan et al., 2014; Huang et al., 2018a; Jin et al., 2020). Accordingly, the concentrations and stoichiometric ratio of N and P in plants are widely used to reflect the plant nutrient status and nutrient supply of ecosystems (Sardans and Peñuelas, 2012). Notably, many previous studies have focused on green leaves (Mayor et al., 2014; Yan et al., 2018; Liu et al., 2021). Leaf nutrient dynamics are a more reliable indicator of plant adaptive strategies than total nutrient concentrations (Yang, 2018; Xu et al., 2021); moreover, their species-specific responses could explain the changes in community structure (Wan et al., 2020). To date, the effects of altered absolute nutrient availability on leaf N and P stoichiometry have been well demonstrated (Lü et al., 2013; Jing et al., 2017; Huang et al., 2018b). Evidence from grasslands, wetlands, and forests indicates that enriched N and P availability generally increased the respective nutrient concentrations in the leaves (Yuan and Chen, 2015), which will in turn have important feedbacks on productivity (Li et al., 2016; Huang et al., 2018b). The N:P supply ratio is also perceived as a key factor affecting leaf nutrient traits, which could influence the trade-offs in the nutrient allocation of different species and may explain their distinct performances under competition (Venterink and Güsewell, 2010; Yuan et al., 2013); however, relatively little attention has been given to it. Considering that the environmental N and P status had been changed from both the overall supply level and the supply ratio, it is necessary to distinguish the respective and interactive effects of the two aspects on the leaf nutrient status thus to predict future changes in the productivity and community composition.
Wetlands are experiencing an increase in reactive N and P loadings globally (Jordan et al., 2011; Wolf et al., 2013; Hu et al., 2017), and the plant trophic dynamics under the changing external environment have been extensively studied in wetland ecosystems (Yuan and Chen, 2015). Nonetheless, the response of plant stoichiometry to varying N and P supplies, and its role in vegetation community structure shifts, are still elusive due to different climate, vegetation and soil properties (Yue et al., 2017; Gao et al., 2018; Wan et al., 2020). Given these uncertainties, more studies are urgently needed to further reveal the plant ecological stoichiometry responses to altered nutrient supply conditions in wetlands.
The Yellow River Delta wetland, the most efficiently conserved and youngest wetland ecosystem in the warm temperature zone in China, is regarded as vulnerable to climate change and anthropogenic impacts (Yu et al., 2016). During the last several decades, jointly influenced by atmospheric deposition and regional economic development, the region has been undergoing increasing N and P inputs, which has significant impact on plant communities (Yu et al., 2014; Li et al., 2017). Generally, the adaptive capacity of plants to environmental change varies with species evolutionary history, and may lead a change in interspecific competition, thus shifting the community composition (Peñuelas et al., 2008; Sardans and Peñuelas, 2014; Sardans et al., 2015). Suaeda glauca and Phragmites australis are two dominant species in the supratidal wetland of this area but have received little attention in previous studies. Comparatively, S. glauca is an annual forb and is restricted to the upland of the Yellow River Delta (He et al., 2012), while P. australis is a perennial and widespread wetland grass (Guo et al., 2018). Although our previous study based on a field-stimulated experiment showed how the two species respond to various N and P supplies (Liu et al., 2019), it mainly focused on the variation in their dominance, while the species-specific responses of leaf nutrient status and plant biomass remain unclear.
In this study, we examined the effects of nutrient supply level and N:P supply ratio on the aboveground biomass and leaf nutrient stoichiometry of P. australis and S. glauca using the same field-stimulated experiment (Liu et al., 2019). The primary objectives of this study were to (i) clarify the main and interactive effects of nutrient supply level and supply ratio on the aboveground biomass and leaf nutrient stoichiometry of two dominant species, (ii) assess the linkage between aboveground biomass and leaf nutrient stoichiometry in two dominant species, and (iii) compare the plant growth strategies employed by two dominant species in response to various N and P supplies. Our previous findings suggest that the plant community composition in this area was only affected by the overall nutrient supply level, in which S. glauca became increasingly dominant with increasing supply level and thus suppressed other species, including P. australis. In addition, the N:P supply ratio and overall supply level both significantly affected the concentrations of soil inorganic N and available P and the N:P ratio, with the soil resource availability being more affected by the supply level and the soil N:P ratio being overridingly influenced by the N:P supply ratio (Liu et al., 2019). We therefore hypothesized that (i) the aboveground biomass of both species was only significantly influenced by the overall supply level, while the leaf nutrient stoichiometry relied on the overall supply level as well as on the N:P supply ratio (H1); (ii) the aboveground biomass of S. glauca was positively associated with the altered leaf nutrient contents, while the performance of P. australis was the opposite, and there was no significant relationship between the N:P and aboveground biomass of both species (H2); and (iii) S. glauca exhibited relatively greater plasticity in plant growth strategies than P. australis, leading to its dominance in the plant community under increased nutrient inputs (H3).
2 Materials and methods
2.1 Experimental site
This study was conducted in the Yellow River Delta Ecology Research Station of Coastal Wetland (37°45’52” N, 118°58’52” E), Chinese Academy of Sciences. The study site has a warm-temperate and continental monsoon climate with a mean annual temperature and precipitation of 12.9°C and 560 mm, respectively. This area is periodically flooded from July to September, and nearly 70% of the annual precipitation is concentrated in this period. The soil type is a saline-alkali soil with a pH of 7.20–7.89 and conductivity of 1.64–3.15 ms cm-1. The dominant plant species at the experimental site are S. glauca, P. australis and Suaeda salsa, comprising more than 90% of the total aboveground biomass.
2.2 Experimental design
In July 2014, a N and P addition experiment was installed to simulate future nutrient supply changes in this wetland. Detailed information about the experimental design has been reported in a previous study (Liu et al., 2019). Briefly, there were nine fertilization treatments combined with 3 overall nutrient supply levels (low, medium, high) and 3 N:P supply ratios (5:1, 15:1, 45:1) with four replicates for each treatment, and one control treatment (CK) was also set up with six replicates, for a total of 42 plots (Figure 1). Each plot was 3.5 × 3.5 m in size and was laid out separated by 1 m aisles. Starting in 2015, fertilizer was applied twice each year with 50% of the supply in early April and 50% in late June. N and P were supplied as urea and dihydrogen phosphate (NaH2PO4) dissolved in 6 L water, respectively, and the CK plots received an equal amount of water.
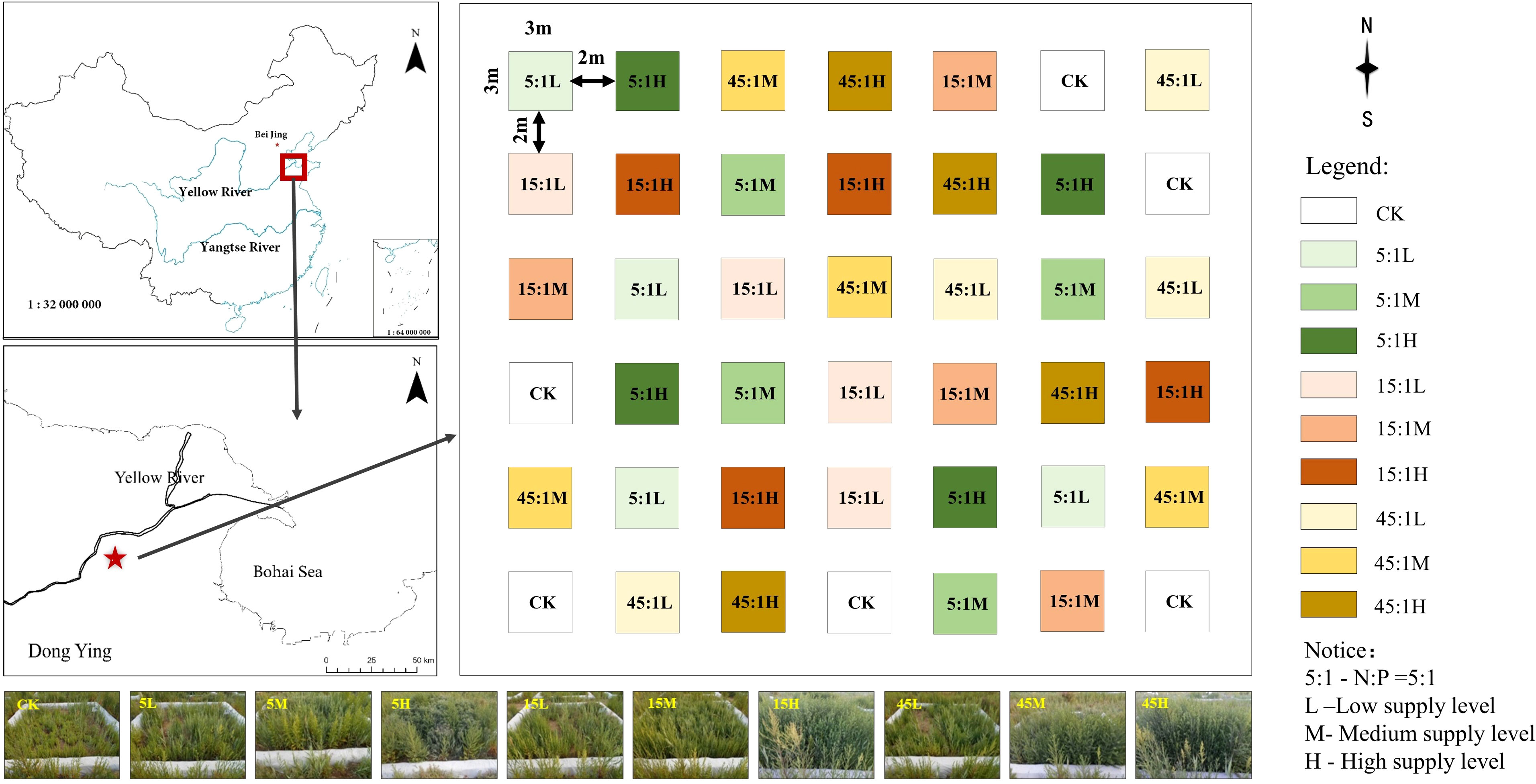
Figure 1 Location and layout of the experimental plots.
2.3 Field sampling and measurement
In late July 2018, peak aboveground biomass was estimated by clipping at ground level in three 0.3 m × 0.3 m random quadrats within each plot and sorting the biomass into different species. After clipping, living biomass was oven-dried at 75 °C for 48 h to a consistent weight and then weighed and ground. Dominant species S. glauca and P. australis were sampled in all experimental plots (Figure 2), while another dominant species S. salsa was not sampled in 3/4 of the high nutrient supply experimental plots due to its low occurrence. Therefore, we only measured the N and P concentrations of S. glauca and P. australis. Total N concentrations in plant leaf were analyzed by a C/N analyzer (Vario Micro, Germany). For total P concentration, powders of plant leaf samples were digested with H2SO4-H2O2 and then determined by the molybdenum blue method on a continuous-flow autoanalyzer (AA3, Seal Analytical, Germany).
Figure 2 S. glauca is a C3 annual forb with fibrous roots; P. australis is a C4 perennial rhizome grass.
2.4 Statistical analysis
Data were tested for normality using Levene’s test. Two-way ANOVA was employed to examine the effects of supply level, supply ratio and their interactions on aboveground biomass, N concentration, P concentration and the N:P ratio of the two species. The ANOVAs were followed by S-N-K post hoc tests to determine the difference level (if necessary). Independent samples t-test was used to compare the mean values of the parameters between the two species under different N and P supplies. The Pearson coefficient was calculated to detect the relationship between the community biomass and aboveground biomass of the two species and the correlation among the N and P concentrations, the N:P ratio and the aboveground biomass of the two species. Structural equation model (SEM) was further constructed with Amos 21.0 software (SPSS Inc., IBM Co., Armonk, NY, USA) to examine direct and indirect hypothetical relationships among supply level, supply ratio, plant N and P concentrations, plant N:P ratios and plant biomass. In these analyses, the data were fit to the model using maximum likelihood estimation. All statistical analyses were performed with SPSS 17.0 (SPSS Inc., Chicago, IL, USA). The significance level was set as 0.05.
3 Results
3.1 Effects on aboveground biomass
The increase in aboveground biomass of S. glauca (BSg) occurred in all N and P addition plots compared to the control plots (Figure 3A). In contrast, the aboveground biomass of P. australis (BPa) decreased with N and P addition, except for the 5:1 M and 45:1 M treatments (Figure 3B). Both BSg and BPa were significantly affected by the N and P supply level rather than the supply ratio, and no significant supply level × supply ratio interaction was observed (Table 1). Compared with the control treatment, BSg increased strongly at the medium and high supply level, while BPa decreased significantly at the low and high supply level (Figure 4A). With the rise of the overall supply level, BSg increased significantly between all gradual supply levels. For BPa, there was no difference between the low and medium supply level, while it decreased significantly at the high supply level. Relative to BPa, BSgwas lower at the low supply level, nearly equal at the medium supply level, and higher at the high supply level (Figure 4A).
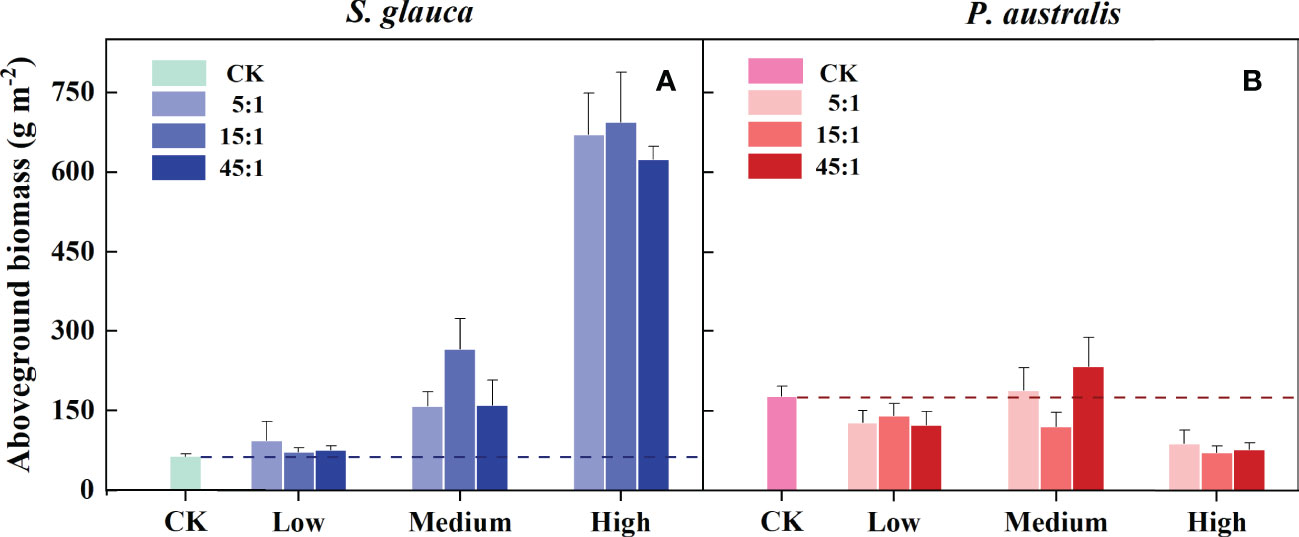
Figure 3 Effects of N and P addition on aboveground biomass of S. glauca (A) and P. australis (B). CK=control. Different bars indicate the mean value (± SE) for each treatment (N and P addition, n=4; CK, n=6).
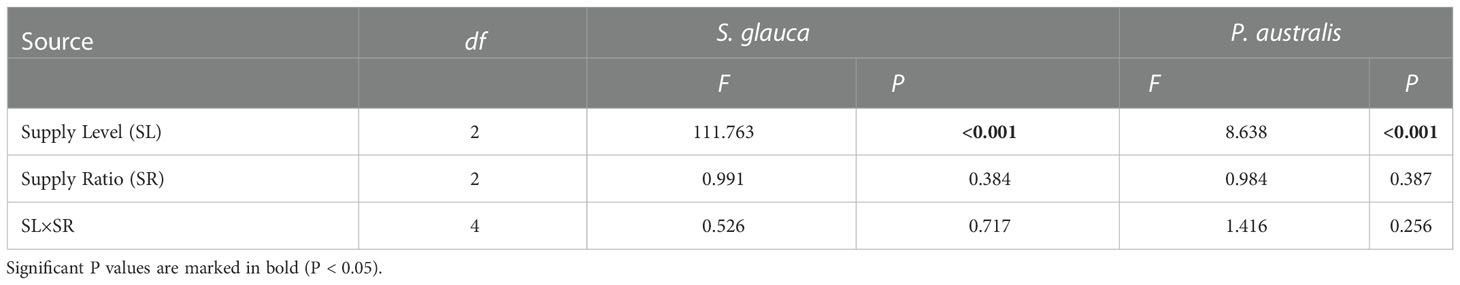
Table 1 Two-way ANOVA for the aboveground biomass of S. glauca and P. australis using the overall supply level (SL) and N:P supply ratio (SR) as the main effects. P-values in bold indicate significant differences at P < 0.05.
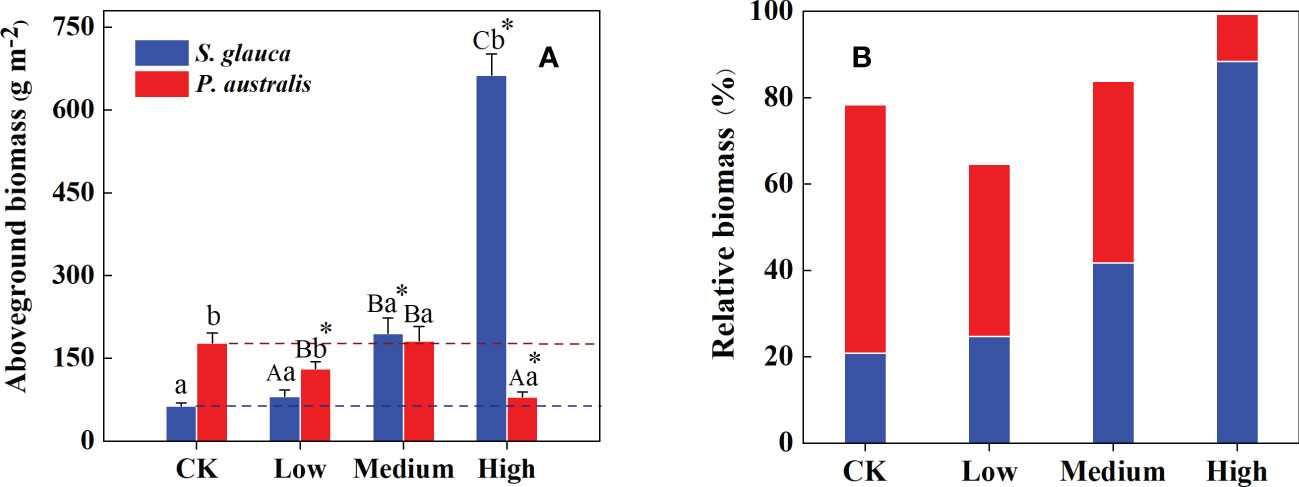
Figure 4 Mean aboveground biomass (A) and relative biomass (B) of S. glauca and P. australis under different treatments. Data show the mean ± SE (supply level treatment, n=12; control treatment, n=6). Different capital letters represent significant differences in plant biomass among the three supply level treatments at the 0.05 level. Different lowercase letters represent significant differences between S. glauca and P. australis under the same treatments at the 0.05 level. * denotes a significant difference between the control treatment and the supply level treatment at the 0.05 level.
The plant community structure in the control plots was co-dominated by S. glauca (20.8%) and P. australis (57.6%) with over 75% of the total aboveground biomass (Figure 4B). However, these two dominant species showed opposite patterns in relative aboveground biomass along the nutrient supply gradient. When the supply level reached a high level, S. glauca became the single-dominant species, accounting for 88.4% of the total aboveground biomass, while P. australis deceased to only 10.9%. Further linear regression analysis showed that the community biomass response was positively correlated with BSg (P<0.001, Supplementary Figure S1) and negatively correlated with BPa(P=0.005, Supplementary Figure S1).
3.2 Effects on N and P stoichiometry
For both S. glauca and P. australis, all of the N and P addition treatments had greater N and P concentrations than the control treatment (Figures 5A–D). However, foliar nutritional traits responded differently to changing N and P supply conditions between S. glauca and P. australis. Two-way ANOVA showed that the effects of supply level and supply ratio on the N and P concentrations of S. glauca were significant, and there was a significant supply level × supply ratio interaction. For P. australis, the P concentration was significantly affected by the supply level and supply ratio, while the N concentration was only significantly affected by the supply level, and no significant supply level × supply ratio interaction was observed (Table 2). In terms of the N:P ratio, the supply ratio rather than the supply level had a significant effect on both species, and a significant supply level × supply ratio interaction was observed only for S. glauca. The N and P concentrations of the two species tended to increase with increasing supply level. With the increase in the supply ratio, the N concentration and N:P ratio also increased, while the P concentration declined (Figures 5E–F, Supplementary Figure S2).
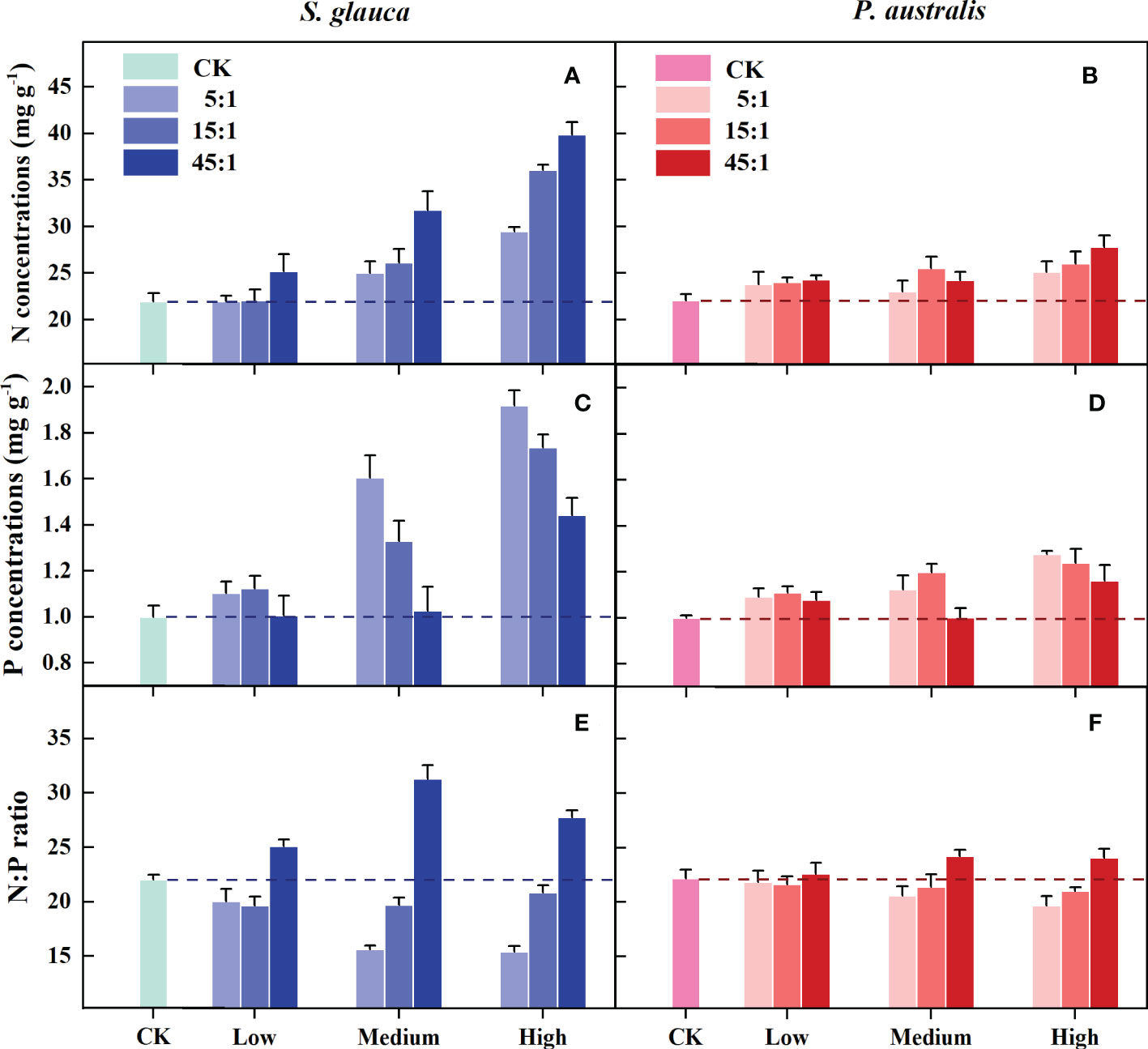
Figure 5 Effects of N and P addition on the N (A, B) and P concentrations (C, D) and the N:P ratio (E, F) of S. glauca and P. australis. Different bars indicate the mean value (± SE) for each treatment (N and P addition, n=4; CK, n=6). Green and red indicate positive and negative relationships corrected to red and green indicate positive and negative relationships.
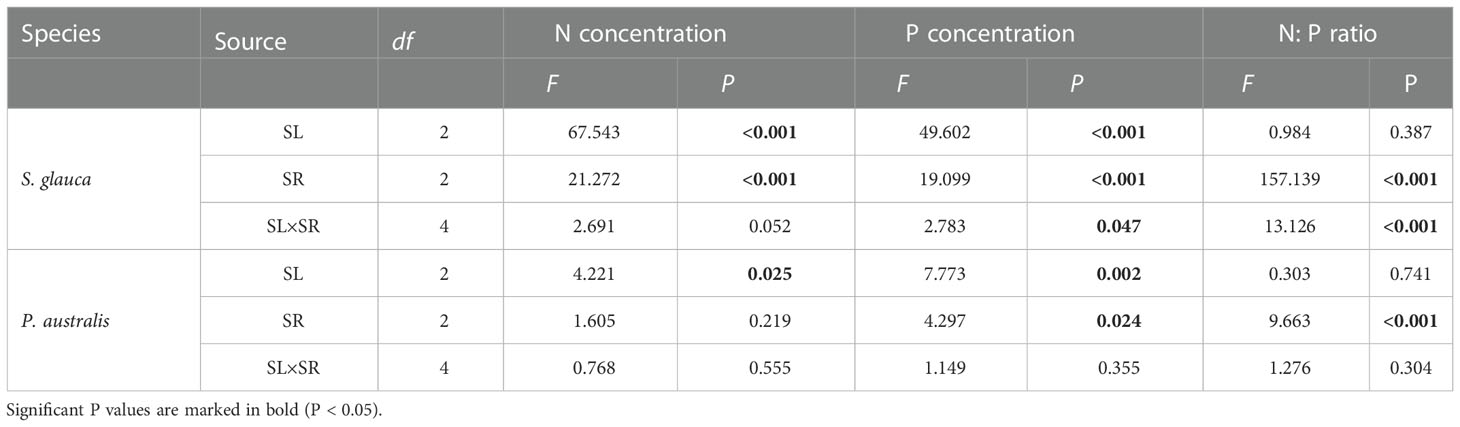
Table 2 Two-way ANOVA for N and P concentrations and the N:P ratio of S. glauca and P. australis using the overall supply level (SL) and N:P supply ratio (SR) as the main effects. P-values in bold indicate indicate significant differences at P < 0.05.
Overall, changing the supply conditions affected the foliar nutritional traits of S. glauca more than those of P. australis, which could be indicated by the F value (Table 2). Correspondingly, the N and P concentrations of S. glauca among the 3 supply levels and 3 supply ratios were all significantly different, and there were significant differences in N:P among the 3 supply ratio gradients. Otherwise, the N and P concentrations of P. australis increased significantly only at the high supply level, and only the high supply ratio significantly increased the N:P ratio and decreased the P concentration. At the medium and high supply level, the N and P concentrations of S. glauca were significantly higher than those of P. australis. Furthermore, the N concentration of S. glauca was significantly higher than that of P. australis at the 45:1 supply ratio, and the P concentration was significantly higher at the 5:1 and 15:1 supply ratio. Compared with the N:P ratio of P. australis, that of S. glauca was higher at the 45:1 supply ratio and lower at the 5:1 and 15:1 supply ratio (Supplementary Figure S2).
3.3 Linkages between nutrient stoichiometry and aboveground biomass
The SEM results indicated that the supply level had a significant positive direct effect on the leaf N and P concentrations of S. glauca and P. australis (Figure 6). In addition, the supply ratio had a significant negative direct effect on the leaf P concentrations of S. glauca and P. australis, whereas it had a significant positive direct effect on the leaf N concentration of S. glauca. The leaf N concentrations of S. glauca and P. australis showed significant positive correlations with the N:P ratios of the corresponding species, which is contrary to the observed relationship between leaf P concentration and the N:P ratio. Regression analysis suggested that the variation in the N:P ratio was primarily determined by foliar P for both species (Supplementary Figure S3). The two dominant species showed opposite patterns in the relationship between leaf nutrient concentrations and aboveground biomass. The N and P concentrations of S. glauca had a positive direct effect on the biomass of S. glauca, but the N and P concentrations of P. australis had a negative direct effect on the biomass of P. australis (Figure 6, Supplementary Figure S4). However, there were no significant correlations between the aboveground biomass of the two species and the N:P ratio of the corresponding species.
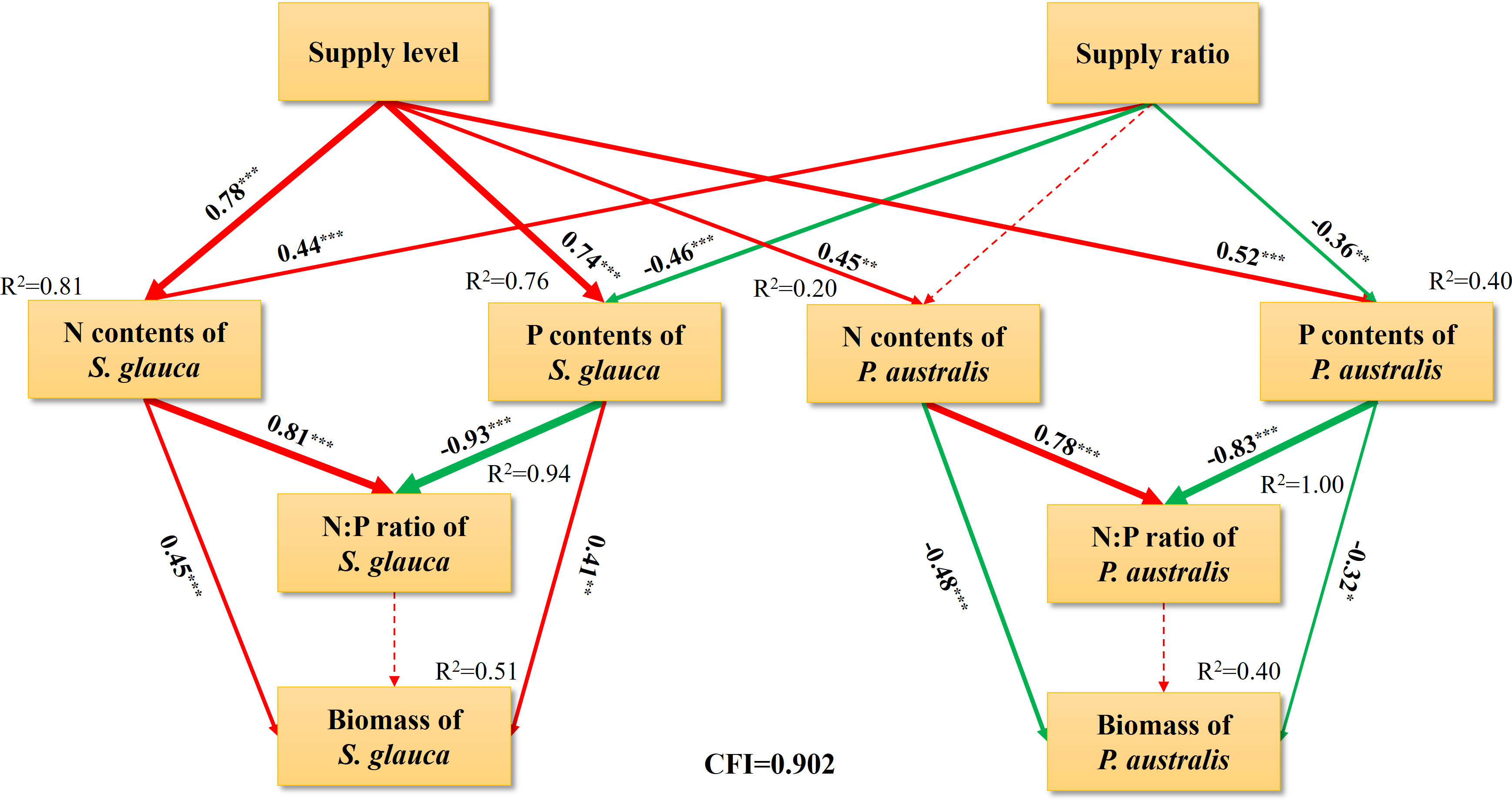
Figure 6 Structural equation model (SEM) considering the underlying causal relationships among leaf N and P contents, N:P ratio and biomass of S. glauca and P. australis under various N and P supplies. CFI, confirmatory fit index. Bold and dashed lines indicate significant (P<0.05) and nonsignificant (P>0.05) pathways, respectively. The width of arrows indicates the strength of the causal effect, while arrows colored red and green indicate positive and negative relationships respectively. The numbers listed within arrows are the standardized path coefficients. The R2 values represent the proportion of the variance explained for each variable. Significance levels are as follows: *P < 0.05, **P < 0.01, and ***P < 0.001.
4 Discussion
4.1 Effects of the overall supply level and supply ratio on plant aboveground biomass
The responses of S. glauca and P. australis to N and P supply were inconsistent from the perspective of plant biomass. Overall, N and P supply had a positive effect on the biomass of S. glauca and a negative effect on that of P. australis (except for the 5:1 M and 45:1 M treatments). The results correspond principally with our first hypothesis (H1), the biomass of both species was mainly affected by the supply level rather than either the supply ratio or the interactive effect. Correlation analysis of species biomass and soil properties demonstrated that BSg was significantly correlated with soil available P (R2= 0.309, n =36, P<0.01) and the available N:P ratio (R2= 0.130, n =36, P=0.031), while BPa was negatively correlated with soil conductivity (R2= 0.185, n =36, P=0.009). Correspondingly, our previous analysis showed that the supply level affected the soil available P, available N:P ratio and soil conductivity more than the supply ratio in the 4th study year (Liu et al., 2019), which could help explain the stronger effects of the supply level on plant aboveground biomass.
As frequently observed in previous studies, grasses and forbs may have contrasting biomass responses to nutrient inputs (Xia and Wan, 2008; Li et al., 2019). Grasses often have high resource use ability and consequently become increasingly dominant with nutrient supply (Zhang et al., 2018a; Su et al., 2020). However, the response of the grass species P. australis in our study is some different. The dominance shifted from P. australis in low nutrient condition to S. glauca in high nutrient condition, indicating S. glauca was well acclimated to nutrient enrichment. Shifts in species dominance may favor acquisitive rather than conservative resource use strategies (Lan and Bai, 2012). P. australis, as a C3 species with conservative resource-use strategies, might perform poorer than S. glauca (C4 species) when exposed to the same changing nutrient conditions (Wang et al., 2015; Zhang et al., 2018b). That is, S. glauca, with faster-growing traits, tends to have a high aboveground competitive ability following nutrient enrichment, which indirectly causes a reduction of P. australis dominance. Additionally, the enhancement of S. glauca led to the positive response of plant community biomass to nutrient addition (Supplementary Figure S1), which confirmed that the growth of S. glauca was the main driver of changes in the plant community (Liu et al., 2019).
4.2 Effects of the overall supply level and supply ratio on plant N and P stoichiometry
Generally, the varying supplies of N and P had significant effects on plant N and P uptake (Fujita et al., 2010; Chen et al., 2020), which could further mediate plant stoichiometry (Zhan et al., 2017). For instance, a pot-culture study showed that the N:P supply ratio had a significant effect on the N and P concentrations and the N:P ratio of wetland grasses through a direct effect on nutrient uptake, whereas the supply level only affected the N:P ratio with a slight effect (Güsewell, 2005). Partly consistent with hypothesis 1 (H1), our results indicated that the leaf N and P concentrations of S. glauca and P. australis were affected more by the supply level than by the supply ratio, while the plant N:P ratio only varied as a function of the supply ratio. Given the positive feedback between soil nutrient availability and plant nutrient contents (Fan et al., 2015; Lü et al., 2015; Su et al., 2021), the inconsistent result with the previous study (Liu et al., 2019) was probably caused by the fact that supply level mainly affected soil resource availability, while the N:P supply ratio overridingly influenced the variations of the soil N:P. Moreover, we found that supply level and supply ratio significantly interacted to affect the N and P concentrations and N:P of S. glauca. This observation highlights the importance of the synergistic effects of supply level and supply ratio on plant stoichiometry (Güsewell and Bollens, 2003; Venterink and Güsewell, 2010), although the underlying mechanism remains unclear.
From the perspective of plant stoichiometry, species with distinct resource use strategies may exhibit different responses to the same changes in environmental conditions (Lü et al., 2015; Li et al., 2019; Qi et al., 2015). Consistent with previous results (Mao et al., 2016; Huang et al., 2018a), the N and P concentrations of both S. glauca and P. australis were significantly enhanced with corresponding increasing N and P input amounts. Theoretically, P. australis is a clonal rhizome grass with strong root reproduction and could benefit from relatively rapid extensive growth to have more chances to access different available nutrients (Cao et al., 2021). Unlike the extensiveness of P. australis root growth, S. glauca is a forb with fibrous roots, and its forage range for obtaining soil available nutrients is relatively limited. Unexpectedly, in contrast to the pattern observed with S. glauca, P. australis exhibited a lower variability in N and P concentrations with the input of corresponding elements, suggesting the relatively stable access of P. australis to N and P despite the nutrient supply gradients (Li et al., 2014; Lü et al., 2015). The nutrient supply had a greater effect intensity on the leaf nutritional traits of S. glauca than that of P. australis after fertilization annually for four years. By comparing variations in the N:P ratio of the two species along the supply ratio gradient, we found that P limitation was transformed to N-P colimitation in S. glauca under decreased N:P supply ratios, whereas P. australis remained P limited (Güsewell, 2004) (Supplementary Figure S2). These results supported the idea that grasses with stronger homeostasis are generally less sensitive to varying N and P supplies than forbs (Yu et al., 2011; Luo et al., 2016). Accordingly, leaf stoichiometry of P. australis spanned a much smaller range than the nutrient availability gradient due to the plant’s regulatory control (Qi et al., 2015), and that in S. glauca could better reflect variation in the N and P supply.
4.3 Plant growth strategies under varying supplies of N and P
In this study, variation in leaf N:P ratios was primarily determined by the response of leaf P concentration, which emphasized the important role of P in affecting plant growth in the supratidal wetland in the Yellow River Delta (Liu et al., 2019). Concretely, the leaf N:P ratios of the two dominant species improved with increasing N concentrations and decreasing P concentrations. Based on this, both the leaf N and P concentrations of the two species increased with increasing supply level, and the leaf N:P ratio remained relatively constant, indicating a higher access of the species to both nutrients and no change in its relative access to N and P (Lü et al., 2015). Alternatively, with an increasing N:P supply ratio, the leaf N concentration improved, while the P concentration exhibited an opposite change trend thus directly determined the positive response of the leaf N:P ratio to the supply ratio. In accordance with the growth rate hypothesis (Elser et al., 2000), changes of plants N:P ratio under changing environments would directly affect biomass accumulation, while our results do not support this view. Our observations suggest that the aboveground biomass of the two dominant species was significantly affected by the supply level rather than the supply ratio, while the leaf N:P ratio was significantly affected by the supply ratio rather than the supply level, which may indirectly result in no significant correlations between the aboveground biomass and the leaf N:P ratio of the two species.
Although species-level stoichiometry exhibited a similar response trend under varying nutrient statuses, the altered leaf nutrients of the two species in turn have divergent feedbacks to plant aboveground biomass (Huang et al., 2018b). Consistent with our expectation (H2), the aboveground biomass of S. glauca increased with increasing leaf N and P concentrations, while that of P. australis increased with reducing leaf N and P concentrations, which may be partially attributed to the differences in their growth strategies. With acquisitive resource-use strategies, S. glauca showed high leaf nutrient contents, corresponding to high aboveground competitive ability (Lan and Bai, 2012). In contrast, P. australis with conservation resource-use strategies exhibited disadvantages in competition, and even increased leaf nutrient contents were insufficient to increase biomass accumulation. Rather, a previous study found that the nutrient contents of grasses affected their biomass positively, while forbs showed an opposite trend (Li et al., 2019). Considering that our results are contrary to the typical expectations (Fujita et al., 2014; Luo et al., 2016), more empirical evidence is needed to clarify the competitive strategies between these two functional groups.
Together, these results supported our third hypothesis (H3) that S. glauca exhibited relatively greater plasticity in growth strategies than P. australis. Our SEM consistently showed that S. glauca has stronger positive feedback under nutrient enrichment (Figure 6) and therefore contributes to increasingly dominance when facing elevated N and P inputs. This finding might provide an underlying mechanism for the changes in the plant community structure from the S. glauca and P. australis co-dominant community to the S. glauca single-dominant community under N and P enrichment.
5 Conclusion
Unbalanced N and P supply as well as absolute nutrient status significantly influenced the leaf stoichiometry of both S. glauca and P. australis, however, the aboveground biomass of the two species was only significantly affected by the overall supply level. Nutrient enrichment promoted the growth of S. glauca but decreased the biomass of P. australis in the coexist community, owing to the strong competitiveness of S. glauca to resources. There was both convergence and divergence in the responses of the leaf stoichiometry of S. glauca and P. australis to various N and P supplies. The plasticity of nutrient resorption in S. glauca was much higher than that of P. australis, although the leaf N and P concentrations of the two species were both significantly enhanced with the increase in corresponding nutrient availability. Furthermore, the altered nutrient concentrations of the two species in turn have contrasting feedbacks to aboveground biomass due to the substantial differences in growth strategies. Comparatively, S. glauca with acquisitive resource-use strategies might become increasingly dominant under increasing nutrient inputs in the supratidal wetland in the Yellow River Delta. These species-specific responses are contrary to the typical expectations of competitive outcomes between grasses and forbs in nutrient supply conditions. The results can provide a scientific basis for adaptive management of wetlands under evolving nutrition change. As the prescribed fertilization was only performed for four years in this study, continuous monitoring is needed to test whether the results still be constant under longer-term fertilization.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
XL and GW designed the project and wrote the manuscript with contributions from GH and AW. HZ, KL and XL performed the experiment and data analysis. All authors contributed to the article and approved the submitted version.
Funding
This work was funded by Joint Funds of the National Natural Science Foundation of China (U2243207), the Strategic Priority Research Program of the Chinese Academy of Sciences, China (XDA23050202), International Science Partnership Program of the Chinese Academy of Sciences (121311KYSB20190029) and the Habitat and Bird Dynamics Monitoring Project of the Yellow River Delta National Reserve (Y991021021).
Acknowledgments
We would like to thank the support from Yellow River Delta Ecological Research Station of Coastal Wetland, CAS, and thank Changli Yang for field work. We thank the reviewers for helpful comments on revision of this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2023.1113629/full#supplementary-material
References
Bobbink R., Hicks K., Galloway J., Spranger T., Alkemade R., Ashmore M., et al. (2010). Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecol. Appl. 20, 30–59. doi: 10.1890/08-1140.1
Cao J. R., Yang L. Y., Pang S., Yang J. J., Hu Y. C., Li Y. C., et al. (2021). Convergent nitrogen uptake patterns and divergent nitrogen acquisition strategies of coexisting plant species in response to long-term nitrogen enrichment in a temperate grassland. Environ. Exp. Bot. 185, 104412. doi: 10.1016/j.envexpbot.2021.104412
Carnicer J., Sardans J., Stefanescu C., Ubach A., Bartrons M., Asensio D., et al. (2015). Global biodiversity, stoichiometry and ecosystem function responses to human-induced C–N–P imbalances. J. Plant Physiol. 172, 82–91. doi: 10.1016/j.jplph.2014.07.022
Chen Z. F., Xiong P. F., Zhou J. J., Lai S. B., Jian C. X., Wang Z., et al. (2020). Photosynthesis and nutrient-use efficiency in response to n and p addition in three dominant grassland species on the semiarid loess plateau. Photosynthetica 58, 1028–1039. doi: 10.32615/ps.2020.056
Elser J. J., Andersen T., Baron J. S., Bergström A.-K., Jansson M., Kyle M., et al. (2009). Shifts in lake N:P stoichiometry and nutrient limitation driven by atmospheric nitrogen deposition. Sci. (80-.). 326, 835–837. doi: 10.1126/science.1176199
Elser J. J., Bracken M. E. S., Cleland E. E., Gruner D. S., Harpole W. S., Hillebrand H., et al. (2007). Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 10, 1135–1142. doi: 10.1111/j.1461-0248.2007.01113.x
Elser J. J., Fagan W. F., Kerkhoff A. J., Swenson N. G., Enquist B. J. (2010). Biological stoichiometry of plant production: metabolism, scaling and ecological response to global change. New Phytol. 186, 593–608. doi: 10.1111/j.1469-8137.2010.03214.x
Elser, O’Brien, Dobberfuhl and Dowling (2000). The evolution of ecosystem processes: growth rate and elemental stoichiometry of a key herbivore in temperate and arctic habitats. J. Evol. Biol. 13, 845–853. doi: 10.1046/j.1420-9101.2000.00215.x
Fan H. B., Wu J. P., Liu W. F., Yuan Y. H., Hu L., Cai Q. K. (2015). Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations. Plant Soil 392, 127–138. doi: 10.1007/s11104-015-2444-2
Fujita Y., de Ruiter P. C., Wassen M. J., Heil G. W. (2010). Time-dependent, species-specific effects of N:P stoichiometry on grassland plant growth. Plant Soil 334, 99–112. doi: 10.1007/s11104-010-0495-y
Fujita Y., Venterink H. O., van Bodegom P. M., Douma J. C., Heil G. W., Hölzel N., et al. (2014). Low investment in sexual reproduction threatens plants adapted to phosphorus limitation. Nature 505, 82–86. doi: 10.1038/nature12733
Gao Y. H., Cooper D. J., Zeng X. Y. (2018). Nitrogen, not phosphorus, enrichment controls biomass production in alpine wetlands on the Tibetan plateau, China. Ecol. Eng. 116, 31–34. doi: 10.1016/j.ecoleng.2018.02.016
Guo X., Yu T., Li M. Y., Guo W. H. (2018). The effects of salt and rainfall pattern on morphological and photosynthetic characteristics of Phragmites australis (Poaceae) 1. J. Torrey Bot. Soc 145, 212–224. doi: 10.3159/TORREY-D-17-00003.1
Güsewell S. (2004). N : P ratios in terrestrial plants: variation and functional significance. New Phytol. 164, 243–266. doi: 10.1111/j.1469-8137.2004.01192.x
Güsewell S. (2005). Responses of wetland graminoids to the relative supply of nitrogen and phosphorus. Plant Ecol. 176, 35–55. doi: 10.1007/s11258-004-0010-8
Güsewell S., Bollens U. (2003). Composition of plant species mixtures grown at various N:P ratios and levels of nutrient supply. Basic Appl. Ecol. 4, 453–466. doi: 10.1078/1439-1791-00174
He Q., Cui B. S., Bertness M. D., An Y. (2012). Testing the importance of plant strategies on facilitation using congeners in a coastal community. Ecology 93, 2023–2029. doi: 10.1890/12-0241.1
Huang J. Y., Wang P., Niu Y. B., Yu H. L., Ma F., Xiao G. J., et al. (2018a). Changes in C:N:P stoichiometry modify n and p conservation strategies of a desert steppe species glycyrrhiza uralensis. Sci. Rep. 8, 12668. doi: 10.1038/s41598-018-30324-w
Huang J. Y., Yu H. L., Liu J. L., Luo C. K., Sun Z. J., Ma K. B., et al. (2018b). Phosphorus addition changes belowground biomass and C:N:P stoichiometry of two desert steppe plants under simulated n deposition. Sci. Rep. 8, 3400. doi: 10.1038/s41598-018-21565-w
Hu M. J., Wilson B. J., Sun Z. G., Ren P., Tong C. (2017). Effects of the addition of nitrogen and sulfate on CH4 and CO2 emissions, soil, and pore water chemistry in a high marsh of the Min river estuary in southeastern China. Sci. Total Environ. 579, 292–304. doi: 10.1016/j.scitotenv.2016.11.103
Jing H., Zhou H. X., Wang G. L., Xue S., Liu G. B., Duan M. C. (2017). Nitrogen addition changes the stoichiometry and growth rate of different organs in pinus tabuliformis seedlings. Front. Plant Sci. 8, 1922. doi: 10.3389/fpls.2017.01922
Jin X. M., Yang L. X., Yang X. G., Guan Q. X., Ma Z. S., Pan J., et al. (2020). Effects of n and p fertilization on the biomass and ecological stoichiometric characteristics of agropyron michnoi in sandy grasslands. Chem. Ecol. 36, 938–952. doi: 10.1080/02757540.2020.1821672
Jordan S. J., Stoffer J., Nestlerode J. A. (2011). Wetlands as sinks for reactive nitrogen at continental and global scales: A meta-analysis. Ecosystems. 14, 144–155. doi: 10.1007/s10021-010-9400-z
Lan Z. C., Bai Y. F. (2012). Testing mechanisms of n-enrichmentinduced species loss in a semiarid inner mongolia grassland: Critical thresholds and implications for long-term ecosystem responses. Philos. Trans. R. Soc B Biol. Sci. 367, 1606. doi: 10.1098/rstb.2011.0352
Li X. Y., Chen H. T., Jiang X. Y., Yu Z. G., Yao Q. Z. (2017). Impacts of human activities on nutrient transport in the yellow river: The role of the water-sediment regulation scheme. Sci. Total Environ. 592, 161–170. doi: 10.1016/j.scitotenv.2017.03.098
Li Y., Niu S. L., Yu G. R. (2016). Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: A meta-analysis. Glob. Change Biol. 22, 934–943. doi: 10.1111/gcb.13125
Liu Y. L., Li L., Li X. Y., Yue Z. W., Liu B. (2021). Effect of nitrogen and phosphorus addition on leaf nutrient concentrations and nutrient resorption efficiency of two dominant alpine grass species. J. Arid Land. 13, 1041–1053. doi: 10.1007/s40333-021-0080-7
Liu X. L., Wang G. M., Ran Y. N., Qi D. H., Han G. X., Guan B., et al. (2019). Overall supply level, not the relative supply of nitrogen and phosphorus, affects the plant community composition of a supratidal wetland in the yellow river delta. Sci. Total Environ. 695, 133866. doi: 10.1016/j.scitotenv.2019.133866
Li J. J., Yang C., Liu X. L., Shao X. Q. (2019). Inconsistent stoichiometry response of grasses and forbs to nitrogen and water additions in an alpine meadow of the qinghai-Tibet plateau. Agric. Ecosyst. Environ. 279, 178–186. doi: 10.1016/j.agee.2018.12.016
Li L. P., Zerbe S., Han W. X., Thevs N., Li W. P., He P., et al. (2014). Nitrogen and phosphorus stoichiometry of common reed (Phragmites australis) and its relationship to nutrient availability in northern China. Aquat. Bot. 112, 84–90. doi: 10.1016/j.aquabot.2013.08.002
Lü X. T., Freschet G. T., Kazakou E., Wang Z. W., Zhou L. S., Han X. G. (2015). Contrasting responses in leaf nutrient-use strategies of two dominant grass species along a 30-yr temperate steppe grazing exclusion chronosequence. Plant Soil 387, 69–79. doi: 10.1007/s11104-014-2282-7
Luo X., Mazer S. J., Guo H., Zhang N., Weiner J., Hu S. J. (2016). Nitrogen:phosphorous supply ratio and allometry in five alpine plant species. Ecol. Evol. 6, 8881–8892. doi: 10.1002/ece3.2587
Lü X. T., Reed S., Yu Q., He N. P., Wang Z. W., Han X. G. (2013). Convergent responses of nitrogen and phosphorus resorption to nitrogen inputs in a semiarid grassland. Glob. Change Biol. 19, 2775–2784. doi: 10.1111/gcb.12235
Mao R., Chen H. M., Zhang X. H., Shi F. X., Song C. C. (2016). Effects of P addition on plant C : N : P stoichiometry in an N-limited temperate wetland of northeast China. Sci. Total Environ. 559, 1–6. doi: 10.1016/j.scitotenv.2016.03.158
Mayor J. R., Wright S. J., Turner B. L. (2014). Species-specific responses of foliar nutrients to long-term nitrogen and phosphorus additions in a lowland tropical forest. J. Ecol. 102, 36–44. doi: 10.1111/1365-2745.12190
Peñuelas J., Sardans J., Ogaya R., Estiarte M. (2008). Nutrient stoichiometric relations and biogeochemical niche in coexisting plant species: Effect of simulated climate change. Polish J. Ecol. 56, 613–622.
Peñuelas J., Sardans J., Rivas-Ubach A., Janssens I. A. (2012). The human-induced imbalance between C, N and P in earth’s life system. Glob. Change Biol. 18, 3–6. doi: 10.1111/j.1365-2486.2011.02568.x
Sardans J., Janssens I. A., Alonso R., Veresoglou S. D., Rillig M. C., Sanders T. G. M., et al. (2015). Foliar elemental composition of European forest tree species associated with evolutionary traits and present environmental and competitive conditions. Glob. Ecol. Biogeogr. 24, 240–255. doi: 10.1111/geb.12253
Sardans J., Peñuelas J. (2012). The role of plants in the effects of global change on nutrient availability and stoichiometry. Plant Physiol. 160, 1741–1761. doi: 10.1104/pp.112.208785
Sardans J., Peñuelas J. (2014). Climate and taxonomy underlie different elemental concentrations and stoichiometries of forest species: the optimum “biogeochemical niche”. Plant Ecol. 215, 441–455. doi: 10.1007/s11258-014-0314-2
Su Y., Le J. J., Ma X. F., Li K. H., Gong Y. M., Ahmed Z., et al. (2020). Plant community composition altered by long-term nitrogen addition has minor contribution to plant nutrient status at the community level. Appl. Ecol. Environ. Res. 18, 6469–6484. doi: 10.15666/aeer/1805_64696484
Su Y., Ma X. F., Le J. J., Li K. H., Han W. X., Liu X. J. (2021). Decoupling of nitrogen and phosphorus in dominant grass species in response to long-term nitrogen addition in an alpine grassland in central Asia. Plant Ecol. 222, 261–274. doi: 10.1007/s11258-020-01103-3
Venterink H. O., Güsewell S. (2010). Competitive interactions between two meadow grasses under nitrogen and phosphorus limitation. Funct. Ecol. 24, 877–886. doi: 10.1111/j.1365-2435.2010.01692.x
Wang W. Q., Sardans J., Wang C., Zeng C. S., Tong C., Asensio D., et al. (2015). Ecological stoichiometry of c, n, and p of invasive phragmites australis and native cyperus malaccensis species in the minjiang river tidal estuarine wetlands of China. Plant Ecol. 216, 809–822. doi: 10.1007/s11258-015-0469-5
Wang J. Y., Wang J. N., Guo W. H., Li Y. G., Wang G. G., Wu T. G. (2018). Stoichiometric homeostasis, physiology, and growth responses of three tree species to nitrogen and phosphorus addition. Trees - Struct. Funct. 32, 1377–1386. doi: 10.1007/s00468-018-1719-7
Wan S. Z., Yang G. S., Mao R. (2020). Responses of leaf nitrogen and phosphorus allocation patterns to nutrient additions in a temperate freshwater wetland. Ecol. Indic. 110, 105949. doi: 10.1016/j.ecolind.2019.105949
Wolf K. L., Noe G. B., Ahn C. (2013). Hydrologic connectivity to streams increases nitrogen and phosphorus inputs and cycling in soils of created and natural floodplain wetlands. J. Environ. Qual. 42, 1245–1255. doi: 10.2134/jeq2012.0466
Xia J. Y., Wan S. Q. (2008). Global response patterns of terrestrial plant species to nitrogen addition. New Phytol. 179, 428–439. doi: 10.1111/j.1469-8137.2008.02488.x
Xu L. C., Xing A. J., Du E. Z., Shen H. H., Yan Z. B., Jiang L., et al. (2021). Effects of nitrogen addition on leaf nutrient stoichiometry in an old-growth boreal forest. Ecosphere 12, e03335. doi: 10.1002/ecs2.3335
Yang H. (2018). Effects of nitrogen and phosphorus addition on leaf nutrient characteristics in a subtropical forest. Trees - Struct. Funct. 32, 383–391. doi: 10.1007/s00468-017-1636-1
Yan Z. B., Kim N., Han W. X., Guo Y. L., Han T. S., Du E. Z., et al. (2014). Effects of nitrogen and phosphorus supply on growth rate, leaf stoichiometry, and nutrient resorption of arabidopsis thaliana. Plant Soil. 388, 147–155. doi: 10.1007/s11104-014-2316-1
Yan Z. B., Li X. P., Tian D., Han W. X., Hou X. H., Shen H. H., et al. (2018). Nutrient addition affects scaling relationship of leaf nitrogen to phosphorus in Arabidopsis thaliana. Funct. Ecol. 32, 2689–2698. doi: 10.1111/1365-2435.13219
Yuan Z. Y., Chen H. Y. H. (2015). Negative effects of fertilization on plant nutrient resorption. Ecology 96, 373–380. doi: 10.1890/14-0140.1
Yuan Y. F., Guo W. H., Ding W. J., Du N., Luo Y. J., Xu F., et al. (2013). Competitive interaction between the exotic plant Rhus typhina L. and the native tree Quercus acutissima Carr. in northern China under different soil N : P ratios. Plant Soil 372, 389–400. doi: 10.1007/s11104-013-1748-3
Yue K., Fornara D. A., Yang W. Q., Peng Y., Li Z. J., Wu F. Z., et al. (2017). Effects of three global change drivers on terrestrial C:N:P stoichiometry: a global synthesis. Glob. Change Biol. 23, 2450–2463. doi: 10.1111/gcb.13569
Yu Q., Elser J. J., He N. P., Wu H. H., Chen Q. S., Zhang G. M., et al. (2011). Stoichiometric homeostasis of vascular plants in the inner Mongolia grassland. Oecologia. 166, 1–10. doi: 10.1007/s00442-010-1902-z
Yu J. B., Ning K., Li Y. Z., Du S. Y., Han G. X., Xing Q. H., et al. (2014). Wet and dry atmospheric depositions of inorganic nitrogen during plant growing season in the coastal zone of Yellow River Delta. Sci. World J. 2014, 1–8. doi: 10.1155/2014/949213
Yu J. B., Zhan C., Li Y. Z., Zhou D., Fu Y. Q., Chu X. J., et al. (2016). Distribution of carbon, nitrogen and phosphorus in coastal wetland soil related land use in the modern yellow river delta. Sci. Rep. 6, 2–10. doi: 10.1038/srep37940
Zhang H. X., Gao Y. Z., Yohannis B. L., Baskin J. M., Baskin C. C., Lü X. T., et al. (2018a). Divergent responses to water and nitrogen addition of three perennial bunchgrass species from variously degraded typical steppe in Inner Mongolia. Sci. Total Environ. 647, 1344–1350. doi: 10.1016/j.scitotenv.2018.08.025
Zhang J. J., Yan X. B., Su F. L., Li Z., Wang Y., Wei Y. N., et al. (2018b). Long-term N and P additions alter the scaling of plant nitrogen to phosphorus in a Tibetan alpine meadow. Sci. Total Environ. 625, 440–448. doi: 10.1016/j.scitotenv.2017.12.292
Keywords: N and P supply, dominant species, aboveground biomass, leaf nutrient stoichiometry, supratidal wetland
Citation: Liu X, Wang G, Zhang H, Han G, Li K and Wang A (2023) Inconsistent stoichiometry and growth responses of two coexisting dominant species to various N and P supplies in a supratidal wetland of the Yellow River Delta. Front. Mar. Sci. 10:1113629. doi: 10.3389/fmars.2023.1113629
Received: 08 December 2022; Accepted: 04 January 2023;
Published: 19 January 2023.
Edited by:
Laibin Huang, University of California, Davis, United StatesReviewed by:
Jiang Bao Xia, Shandong Key Laboratory of Eco-Environmental Science for the Yellow River Delta, Binzhou University, ChinaHao Chen, Institute of Oceanology, Chinese Academy of Sciences (CAS), China
Copyright © 2023 Liu, Wang, Zhang, Han, Li and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guangmei Wang, Z213YW5nQHlpYy5hYy5jbg==