 Free Espinosa1*
Free Espinosa1* Manuel Maestre1
Manuel Maestre1 José Carlos García-Gómez1María Isabel Cotaina-Castro2Carmen Pitarch-Moreno2Juan Manuel Paramio2
José Carlos García-Gómez1María Isabel Cotaina-Castro2Carmen Pitarch-Moreno2Juan Manuel Paramio2 Patricia Fort-Santa María3Natalia García-Estévez3
Patricia Fort-Santa María3Natalia García-Estévez3- 1Laboratorio de Biología Marina, Departamento de Zoología, Facultad Biología, Universidad de Sevilla, Sevilla, Spain
- 2División de Medio Ambiente y Prevención de Riesgos Laborales, Departamento de Infraestructuras, Autoridad Portuaria de Melilla, Melilla, Spain
- 3Departamento de Medio Ambiente. Acciona Ingeniería, Madrid, Spain
The Anthropocene era is characterized by a biodiversity crisis, where many species are pushing to extinction, causing alteration of the stability in the ecosystems and loss of their services. In this context, the translocation of endangered species is a powerful resource in conservation as long as its goals and appropriateness are clearly stated and it matches unambiguous criteria. Patella ferruginea is one of the most endangered marine species in the Mediterranean, and several translocations have been made with limited results in terms of survival. A methodology that mimics the natural topography of the substrate in the donor population based on Artificial Inert Mobile Substrates (AIMS, 40x20x10 cm) is proposed for restocking purposes. Three different treatments were tested: on substrate, slope, and cantilever depending on the position in which the AIMS were deployed. A total of 660 AIMS were installed within the donor area. The installation of such AIMS did not negatively affect either donor or receiving populations. A total of 188 specimens were translocated in three different attempts in 2018, 2019, and 2020, and no mortality was observed during the translocation process that lasted 24 h. Survival through the medium and long-term for the translocated specimens ranged from 80.6 to 91.5% after one month, 55.5 to 80.9% after eight months, and 48.4 to 76.6% after one year, with survival in the control population being 91.6% after four months and 87.5% after one year. Overall, recruits showed higher survival values than adults, whereas substrate treatment hosted more specimens (both recruits and adults) than slope and cantilever ones, while recruits were more abundant on cantilever treatment. The methodology has been proven to be useful for restocking and reintroduction purposes between donor and receiving areas.
Introduction
A large mass extinction debt exists today due to habitat disruption by humans within a new era called Anthropocene (Spalding and Hull, 2021). Species perform critical functions in ecosystem dynamics and provide resilience under ever-changing environments (Tilman et al., 2014; Reyers et al., 2018). Therefore, biodiversity erosion induces alteration of the stability in the ecosystems, loss of ecosystem services, and affects human life and socioeconomic systems. In this regard, habitat destruction and overexploitation have been the most relevant threats to biodiversity, being drivers to push so many species to extinction thresholds (Primack and Ros, 2002).
A key tool to offset this biodiversity crisis caused by human impacts on populations and ecosystems has been species translocations (see Pérez et al., 2012 and references therein). However, even though it could be a powerful resource in conservation biology, is not free of criticism due to its use without clear goals, lack of comprehensive analysis of root causes of population decline (Meffe, 1992), and unambiguous criteria to assess if a given project is going to be successful and its implementation is appropriate (Pérez et al., 2012). Translocations involve considerable risks and, consequently, must rely on the best practice guidelines for further policy application (Swan et al., 2016).
Translocations have been conducted elsewhere and have focused on a wide range of species but mainly on terrestrial vertebrates and birds (Seddon et al., 2005, Seddon et al., 2007), with marine species scarcely mentioned in conservation translocation literature reviews (Swan et al., 2016). Moreover, most marine translocations have focused on ecosystem restoration and catch augmentation for economic purposes, with the translocations focused on single species of conservation concern for preventing their extinction being very scarce (Swan et al., 2016). Therefore, there is a paucity of knowledge about the techniques, success rates, and outcomes in the translocation of marine invertebrates in particular. For instance, out of 174 translocation projects in Spain, none were focused on marine species (Pérez et al., 2012). In general, the term translocation includes three different concepts: introduction (the release of specimens outside of its historically known native range), reintroduction (the release of specimens within its historically known native range from which the species was extirpated), and restocking (the release of specimens within an area where the species is found in order to rebuild its stock).
Patella ferruginea (Mediterranean ribbed limpet) is considered the most endangered marine invertebrate in the Mediterranean along with the noble pen shell Pinna nobilis (Ramos, 1998; Vázquez-Luis et al., 2017). Its distributional range originally encompassed the entire Western Mediterranean basin but has become restricted to a few areas, being almost extinct in continental Europe (Laborel-Deguen and Laborel, 1991a; Espinosa et al., 2013). It is a giant limpet that lives within the upper intertidal and shows homing behavior, reaching 10 cm in length with remarkable ribs and a star-shaped shell (Espinosa and Rivera-Ingraham, 2017). Due to its conspicuous shell, intertidal habitat, and great size, the species has been harvested by humans since the Palaeolithic Age (Fa et al., 2016; Espinosa and Rivera-Ingraham, 2017). Both human harvesting and habitat destruction by coastal development have been the main drivers to put the species at risk of extinction (Espinosa et al., 2018). Currently, there are only dense and well-preserved populations in scattered points along the Northern African coast: Ceuta, Melilla, and Chafarinas Islands (Spanish enclaves); Rachgoun, Habibas, and Plane Islands (Western Algeria); and Zembra Island (Tunisia). Smaller populations can be also found in Corsica and Sardinia (Espinosa et al., 2013). Out of these places, only isolated specimens have been recently discovered on the Ligurian coast in mainland Italy (Ferranti et al., 2019) or in Egadi and Pantelleria islands (Espinosa et al., 2013). Focusing on the Iberian Peninsula, the species is only found in the Southernmost part from Palos Cape to the Strait of Gibraltar but in low density and showing a fragmented distribution (Espinosa et al., 2013; https://www.juntadeandalucia.es/medioambiente/portal/landing-page-documento/-/asset_publisher/jXKpcWryrKar/content/informes-regionales-sobre-gesti-c3-b3n-sostenible-del-medio-marino-andaluz-2008-2018-/20151). Within this area, the most important population is settled in the Algeciras Bay where hundreds to a few thousands of scattered specimens have been reported (Fernández-Casado et al., 2017). Furthermore, García-Gómez et al. (2011; 2015) pointed out that some artificial substrates hosted important populations of this species (and other endangered species as well), and those sites with limited access could be considered for management and conservation of endangered species as a new figure of protection named Artificial Marine Micro Reserves (AMMRs). The inner part of Gibraltar Bay hosted several specimens settled in the breakwater of ‘La Línea’ harbor where the access has been restricted by fences, being a location with the potential to develop a reproductive population in this context. It is worth noting that the species is a broadcast spawner, showing protandrous hermaphroditism, and fecundity is highly dependent on size (Espinosa et al., 2006a). Consequently, a minimum number of closely adjacent specimens, with different size classes are required for successful fertilization.
The increasing interest in the conservation of this species has prompted previous translocations for conservation purposes or derived from engineering works in which repair of existing breakwaters or deployment of new ones have threatened local populations (Laborel-Deguen and Laborel, 1991b; Espinosa et al., 2008; APM (Autoridad Portuaria de Melilla), 2014; Fa et al., 2018; Zarrouk et al., 2018). Unfortunately, high mortality rates have often been observed, although the translocations made with the specimens attached to their own substrate have been proven most successful (Fa et al., 2018). Nevertheless, this approach based on moving entire boulders in a breakwater where the limpets are settled seems difficult to use for many individuals or over long distances. Consequently, its applicability in reinforcement or reintroduction of this or similar endangered species would be limited. Moreover, substrate heterogeneity has been shown as a main driver that influences recruitment in limpets (Martins et al., 2010; Espinosa et al., 2011), and, therefore, it should be taken into consideration in any further translocation procedure. Furthermore, Potet et al. (2021) indicated that the surface for recruitment of the European flat oyster (Ostrea edulis) must be irregular at the scale of mm.
The goals and criteria to assess the outcomes and appropriateness of translocation must be clearly established. In this sense, Pérez et al. (2012) pointed out that any translocation must be properly justified in terms of necessity, risk evaluation, and technical suitability, providing ten major criteria to be considered prior to undertaking any intervention. The present translocation experiment complies with these requirements, and therefore, the likelihood of success was maximized, and the utility of the study was properly justified from a conservation perspective (see Table 1 for details).
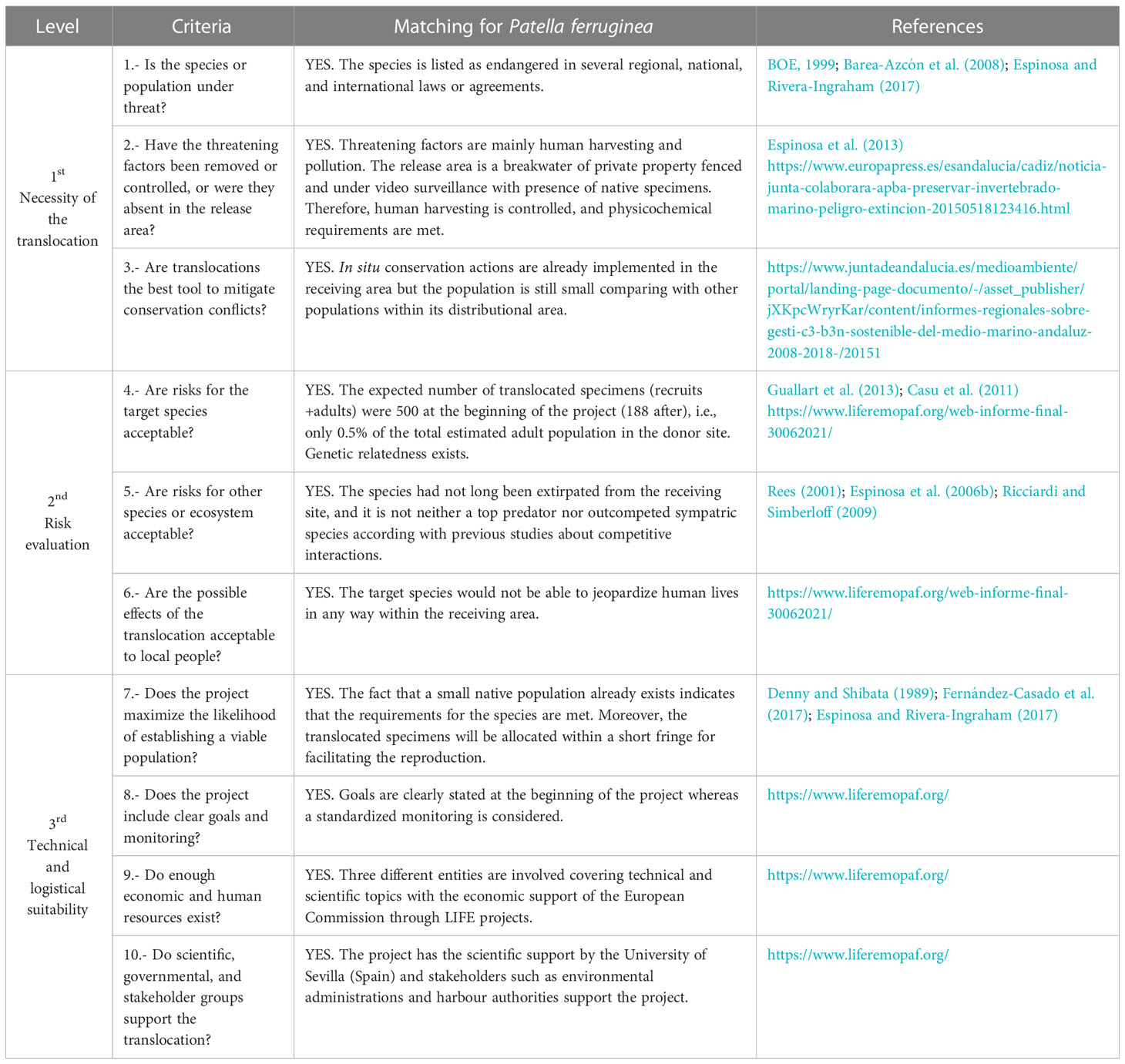
Table 1 Matching criteria for translocations (adapted from Pérez et al., 2012 based on IUCN guidelines and already published literature).
Our aim was to provide a valid protocol to translocate specimens of Patella ferruginea to re-stock depleted or repopulated areas by mimicking the original habitat of the donor population. The specific objective was to test the influence of position and impact on the natural substrate on the efficacy of deployment of Artificial Inert Mobile Substrates (AIMS) in translocation,
Material and methods
Study area
The donor population was located in the autonomous city of Melilla, a Spanish enclave in North Africa, where a dense population estimated at 32,821 adults (>3 cm) was found (Guallart et al., 2013). According to these authors, most of the population (68.1%) was on the exterior breakwater of the Melilla’s harbor where the specimens were collected for translocation and the mean density of adults (per linear length of the breakwater) was 17.6 ind./m with maximum values up to 66.5 ind./m. Melilla Harbor is exposed to strong wave action and has a low risk of marine spills, being considered moderately pristine in terms of environmental quality due to the efficiency of management measures in place, which comprises environmental management, eco-efficiency, and environmental quality (see Fernández-Macho, 2016; González-Laxe et al., 2017).
In contrast, the population to be re-stocked was located in the exterior breakwater of the harbor in La Línea in the inner part of the Algeciras Bay (Figure 1), where a scattered population has been reported with a mean density below 1 ind./m (https://www.juntadeandalucia.es/medioambiente/portal/landing-page-documento/-/asset_publisher/jXKpcWryrKar/content/informes-regionales-sobre-gesti-c3-b3n-sostenible-del-medio-marino-andaluz-2008-2018-/20151). This population was within an enclosed area under surveillance with restriction to public access (Fernández-Casado et al., 2017), being appropriate to undertake restocking for conservational purposes as has been noted previously. The area nearby the restocked population showed slight pollution levels (see Dissanayake and Bamber, 2010; Rojo-Nieto et al., 2011; Rojo-Nieto and Perales, 2015), with moderate wave exposure.
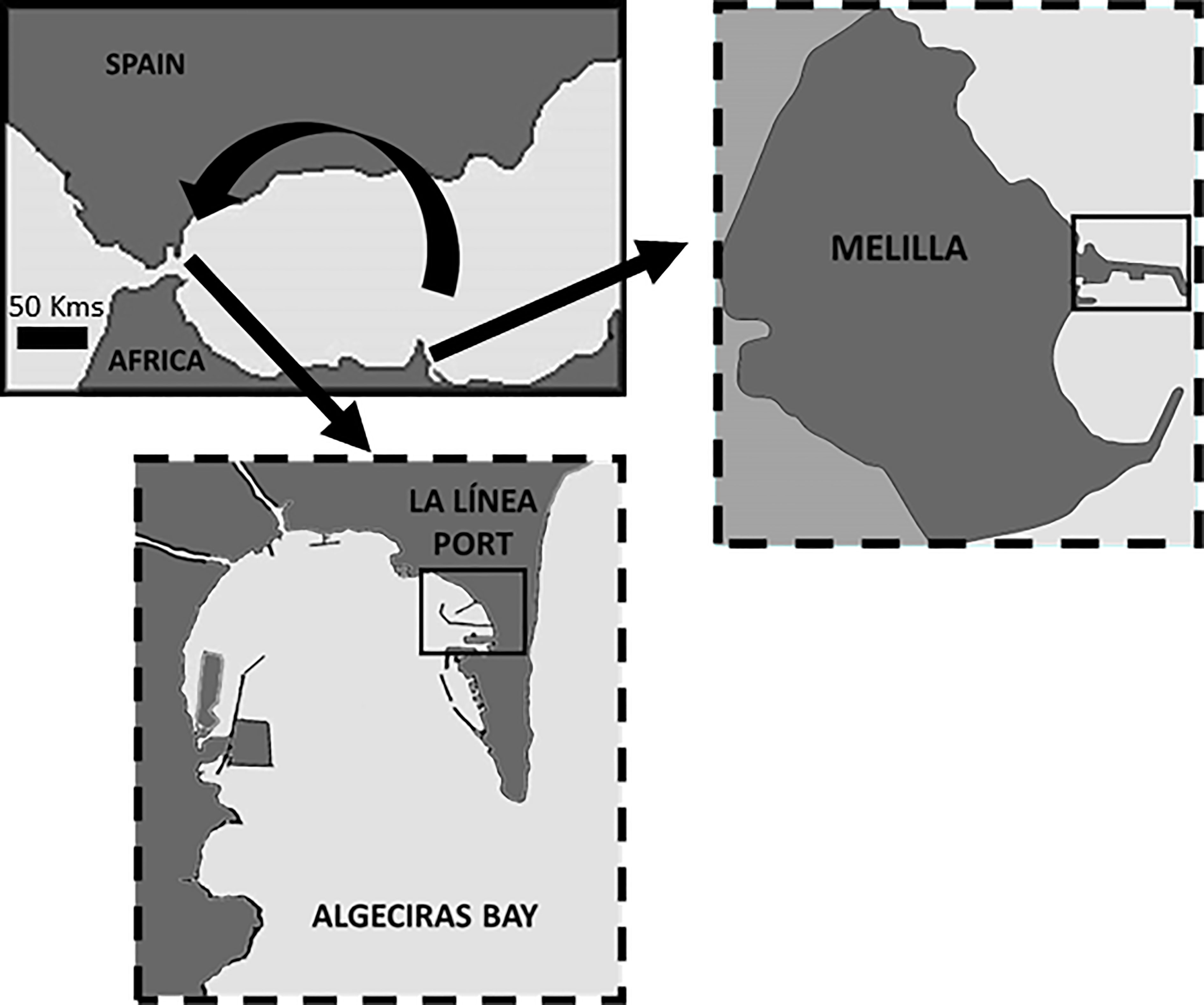
Figure 1 Map of the study area showing the donor and receptor sites located in Melilla and La Línea ports, respectively.
Artificial inert mobile substrates design
The specimens were transplanted using artificial plates designed to mimic the topography of the substrate where the donor population was settled. For it, the surface of the rock was recorded using different 360° cameras and the images were digitalized. The electronic file was used to create an ABS – FDM (Acrylonitrile Butadiene Styrene - Fused Deposition Modeling) mold using 3D printing technology by the Materialise® company that was subsequently allocated on conventional concrete (type HA-35/F/12/IIIc+Qb) with internal reinforced steel to print a ‘negative’ that recreated the microtopography of the original substrate. The plates could be easily screwed and unscrewed from the rocks, having a dimension of 40 x 20 x 10 cm (length, width, and height), and showed resistance and durability for 25 years according to the calculations made considering the forces from the wave action in the area (Acciona Ingeniería, unpublished data), covering the life expectancy of the species (see Espinosa et al., 2008).
To test the effect of the AIMS position on Mediterranean ribbed limpet recruitment rate as well as their impact on the intertidal zone, three different treatments were used: Onto the substrate (OS), in which the AIMS were directly fixed on the rock surface so that continuity of hard substrate was maximized, while their impact on the natural substrate through occupation was high; Sloped (S), in which the AIMS were fixed to the rocky substrate at an angle to the natural substrate so that the impact of the manipulation on the natural substrate was largely reduced, while habitat continuity was still substantial; and Cantilever (C), where the AIMS were fixed above the rock surface without direct contact with it, so that the impact of the experiment on the natural substrate was reduced to a minimum at the cost of very reduced hard substrate continuity for limpets (Figure 2). Therefore, it would be expected that colonization and the impact within the intertidal habitat (both of major conservation concern) followed the pattern OS>S>C, according to the decreasing contact with the surrounding environment.

Figure 2 Artificial Inert Mobile Substrates (AIMS) used for translocating individuals of Patella ferruginea. The three different treatments considered are shown: on substrate, slope, and cantilever. The solid line indicates the lower limit of the habitat of Patella ferruginea just above the macroalgal turf (note that deployed AIMS were installed out of the habitat of Patella ferruginea). The dashed line indicates the corresponding tidal level of the AIMS surface that relied on the tidal height where the habitat of Patella ferruginea is found.
Temporal monitoring of limpet populations
To test if the installation of the AIMS could affect either donor or receiving populations, monitoring was conducted between September 2017 and June 2021. In the donor population, a total of seven 10 m transects were set up along the breakwater and were considered as ‘Controls’. Moreover, additional seven transects were interspersed with controls and there, a total of six AIMS were placed per transect (two for each type of position: OS, S, and C), being those transects considered as ‘AIMS’. Within each transect, all the adult specimens (>3 cms) were counted and measured using a caliper, recording the longitudinal axis of the shell approximately every four months (Espinosa et al., 2008). Similarly, in the receiving population, three control and three AIMS transects were also set up and monitored since the restocking was performed. The AIMS were deployed in fewer numbers than in the donor population due to its smaller area. Data were recorded similarly (every four months) for the receiving population and the donor population groups, although there was a gap in the first months of 2020 due to logistical constraints derived from the Covid-19 pandemic.
Apart from the status of both populations in donor and receiving areas, a community assessment was carried out within the intertidal habitat where Patella ferruginea was located. Five sites were randomly distributed within the donor and receiving populations and five quadrats of 25 x 25 cm were photographed per site either at low or high tidal levels (+25 and +75 cm above zero tidal level, respectively). Each photograph was analyzed digitally by adding a grid of 25 subquadrants to obtain the coverage of each species in % (Guerra-García et al., 2006).
In addition to biological data, several environmental parameters (pH, temperature, salinity, and turbidity) were recorded in situ by taking water samples in the vicinity of the transects. All the parameters were measured using a multiparameter probe CRISON MM40© and turbidimeter WTW 335IR©.
Translocation methodology
A total of 660 AIMS were deployed in the donor population several months prior to the recruitment period of the species (late Autumn-early Winter) to allow the development of the typical microfilm on which the species feeds (Burgos-Rubio et al., 2015). Due to logistical constraints in terms of fabrication, transporting, and deploying in the donor population area within the precise tidal level, two different installation events were undertaken. A pilot attempt was done in July-August 2017 when a total of 150 AIMS were deployed, whereas the other 510 AIMS were deployed between June-September 2018. Three different translocation events were undertaken in October 2018, 2019, and September 2020. The AIMS were monitored annually in early September and the specimens settled in them were registered for subsequent translocation. Once the AIMS with recruits and adults were identified, they were unscrewed at low tide, the limpets measured in length, and marked using epoxy putty (Espinosa et al., 2008). Such extracted AIMS were replaced with new ones after the first and second translocations, so the total number of AIMS in the donor population remained constant at 660. The AIMS were then introduced into a plastic container with a rubber base to avoid vibration, since it has been indicated that vibration could detach limpets (Fa et al., 2018). Finally, they were covered with towels soaked with marine water, introduced in a refrigerated truck at 16°C, and transported to the receiving zone. All the AIMS were screwed on the receiving zone at low tide within the next 24h.
Limpets were monitored 24h and 48h after transplantation, weekly during the first month, monthly during the first four months, and every four months afterward. The position of the limpets was recorded to see if they had left the AIMS to settle on the rock and all the transplanted specimens were also measured in length at the different monitoring times to obtain growth rate values.
Statistical analyses
A repeated‐measures ANOVA (RM‐ANOVA) was carried out to test whether the mean size or density of adults significantly varied through time using all transects (intrasubject factor ‘Time’ and intersubject factor ‘Treatment’ with two levels: Control vs. AIMS). Mauchly’s test of sphericity was used to test the assumption that variance in the differences between all possible pairs of groups is equal. The Huynh-Feldt correction was used when an absence of sphericity was detected.
Possible differences in survival among the three transplantations (2018, 2019, and 2020) through time as well as among recruits and adults were tested using a Chi-squared test. A one-way ANOVA test was used to test differences in growth rates.
A matrix of the community structure based on species composition and their coverages was employed to test if the intertidal community changed through the monitored time using the SIMPROF test. The analysis was run on a triangular similarity matrix derived from the values of the Bray-Curtis similarity on fourth root transformed coverage data. A similar analysis was performed with environmental variables using Euclidean distance instead of Bray-Curtis similarity.
Univariate analyses (Chi-squared test, ANOVA, and RM-ANOVA) were performed using SPSS© software, and multivariate analyses (SIMPROF test) were conducted using PRIMER v.6+PERMANOVA package (Clarke and Gorley, 2006).
Results
Monitoring in donor and receiving populations
During the two years of monitoring (from September 2017 to September 2019) mean size values ( ± SE) of the donor population in Melilla port slightly decreased in both control and AIMS transects (from 4.63 ± 0.11 to 4.33 ± 0.06 cm and from 4.95 ± 0.08 to 4.42 ± 0.04 cm, respectively) (Figure 3A). RM-ANOVA indicated significant differences over time in mean size values (Table 2), but not between treatments (control vs. AIMS). Moreover, density values fluctuated similarly for both control and AIMS transects (Figure 3A) Nevertheless, the AIMS transects showed an increase from 22.3 to 27.03 ind./m after two years, whereas the control ones remained with similar values after that period (19.21 to 18.57 ind./m), showing fluctuations over the monitored time. In fact, the RM-ANOVA also showed significant differences in density through time (Table 2) but not between treatments (control vs. AIMS).
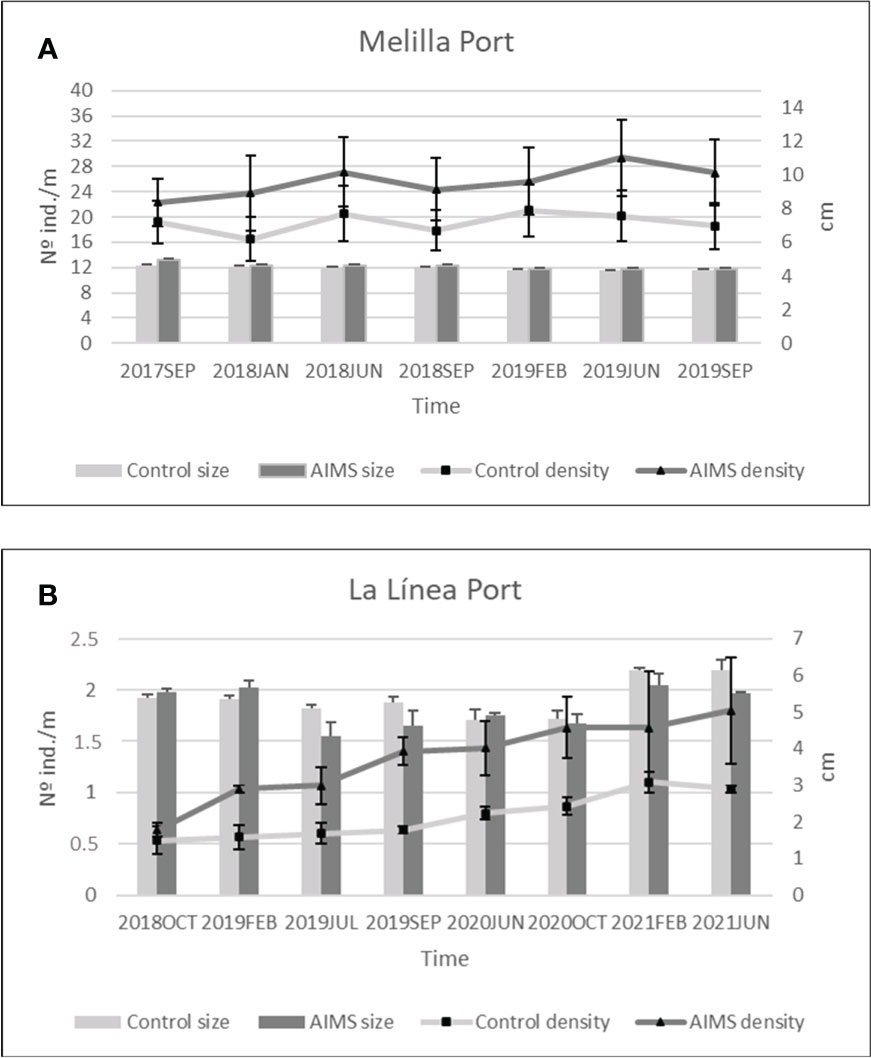
Figure 3 Density and size mean values ( ± SE) during the monitoring period in the Control and AIMS transects. (A): Melilla; (B): La Línea.

Table 2 Results of a repeated-measured analysis of variance (RM-ANOVA) on the mean size and density of adults (>3 cm) of the monitored transects in the donor population (Melilla).
Regarding the receiving population in La Línea port, an increase in mean size values was recorded for control and AIMS transects (from 5.39 to 6.15 cm and from 5.54 to 5.5 cm, respectively) (Figure 3B). The RM-ANOVA also indicated significant differences over time in mean size values (Table 3) but not between treatments (control vs. AIMS). Similarly, density values showed an increase from 0.63 to 1.8 ind./m within the AIMS transects and from 0.53 to 1.03 ind./m within the control ones (Figure 3B). The results of the RM-ANOVA were like those recorded for size values (Table 3) or even for the donor population: differences over time but not between the considered treatments.
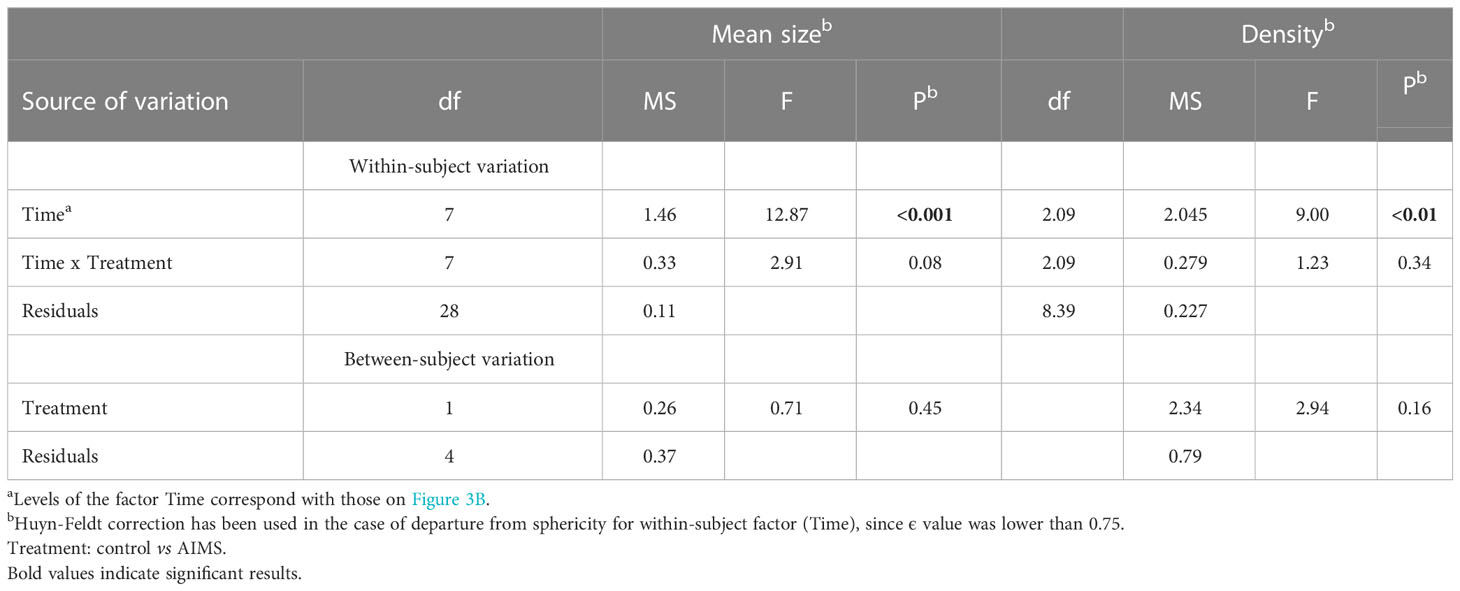
Table 3 Results of a repeated-measured analysis of variance (RM-ANOVA) on the mean size and density of adults (>3 cm) of the monitored transects in the receiving population (La Línea).
The intertidal assemblages in the donor (Melilla port) and receiving populations (La Línea port) did not show differences in structure composition over the monitored time according to the SIMPROF test. Furthermore, environmental variables showed similar values throughout the monitoring period, excepting the usual seasonal changes in temperature, and recording low levels of turbidity (0.2-1.65 ntu in Melilla and 0.7-1.6 ntu in La Línea) (Figures 4A, B).
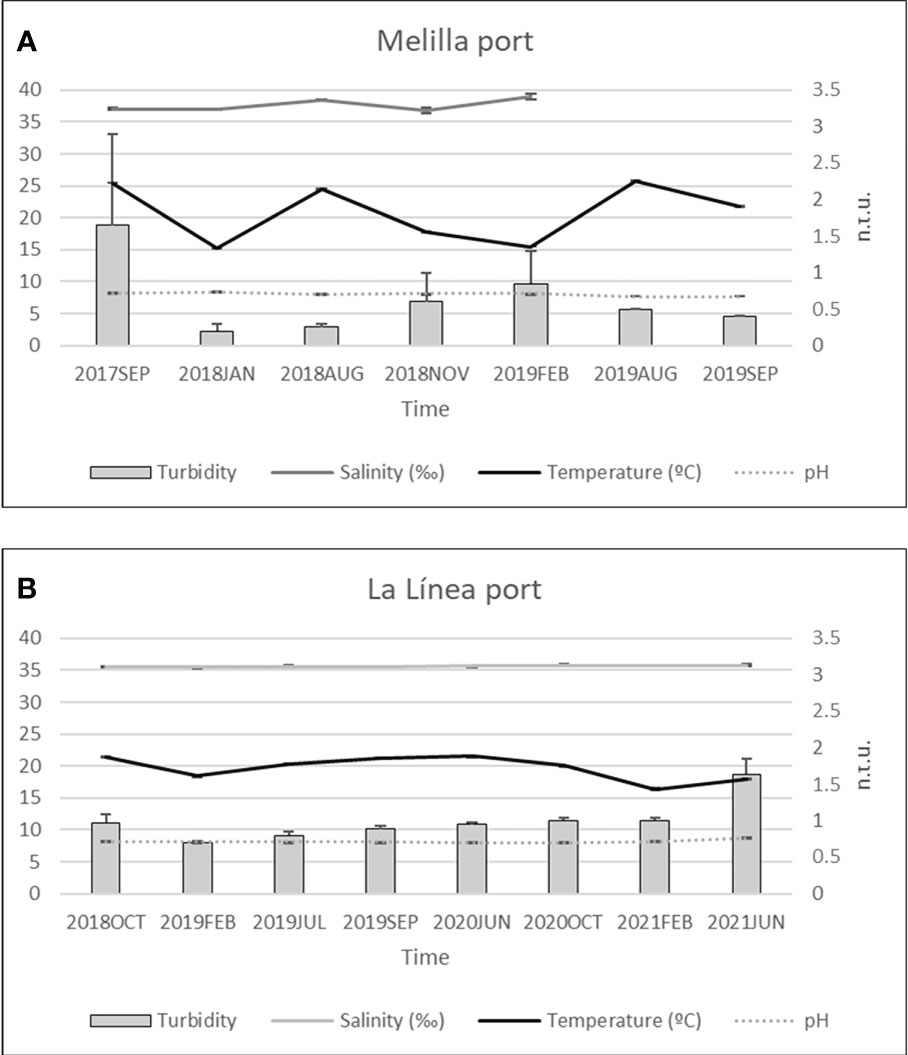
Figure 4 Physicochemical variables during the monitoring period. (A) Melilla; (B) La Línea. Turbidity is represented by the right Y axis (ntu units), whereas salinity, temperature, and pH are all represented by the left Y axis. Salinity values for August and September 2019 in Melilla were not available due to an error with the probe.
Translocation experiments
A total of 188 specimens were translocated in three different translocation events: 31, 47, and 110 respectively (Figure 5). The number of recruits (<3 cm) was higher than adults for the first translocation, but adults outnumbered recruits globally. Besides, the number of AIMS translocated was 20, 40, and 65 for each of the translocation events performed for a total of 125 AIMS translocated to the receiving population during the whole study (Figure 5). The percentage of AIMS with specimens of Patella ferruginea settled was 3%, 6%, and 10% for each translocation event, with the total recruited being almost 19% of such experimental structures deployed in the donor population. Regarding the capacity for settlement of P. ferruginea individuals on the experimental structures (AIMS), the mean number of individuals per AIMS was similar among the three translocation events (1.55, 1.18, and 1.69 individuals/AIMS, respectively), but the mean number per square meter showed a marked increase from 1.04 to 3.7 ind./m2 similar to that observed for the mean number of settled individuals in the whole set of AIMS (Figure 6).
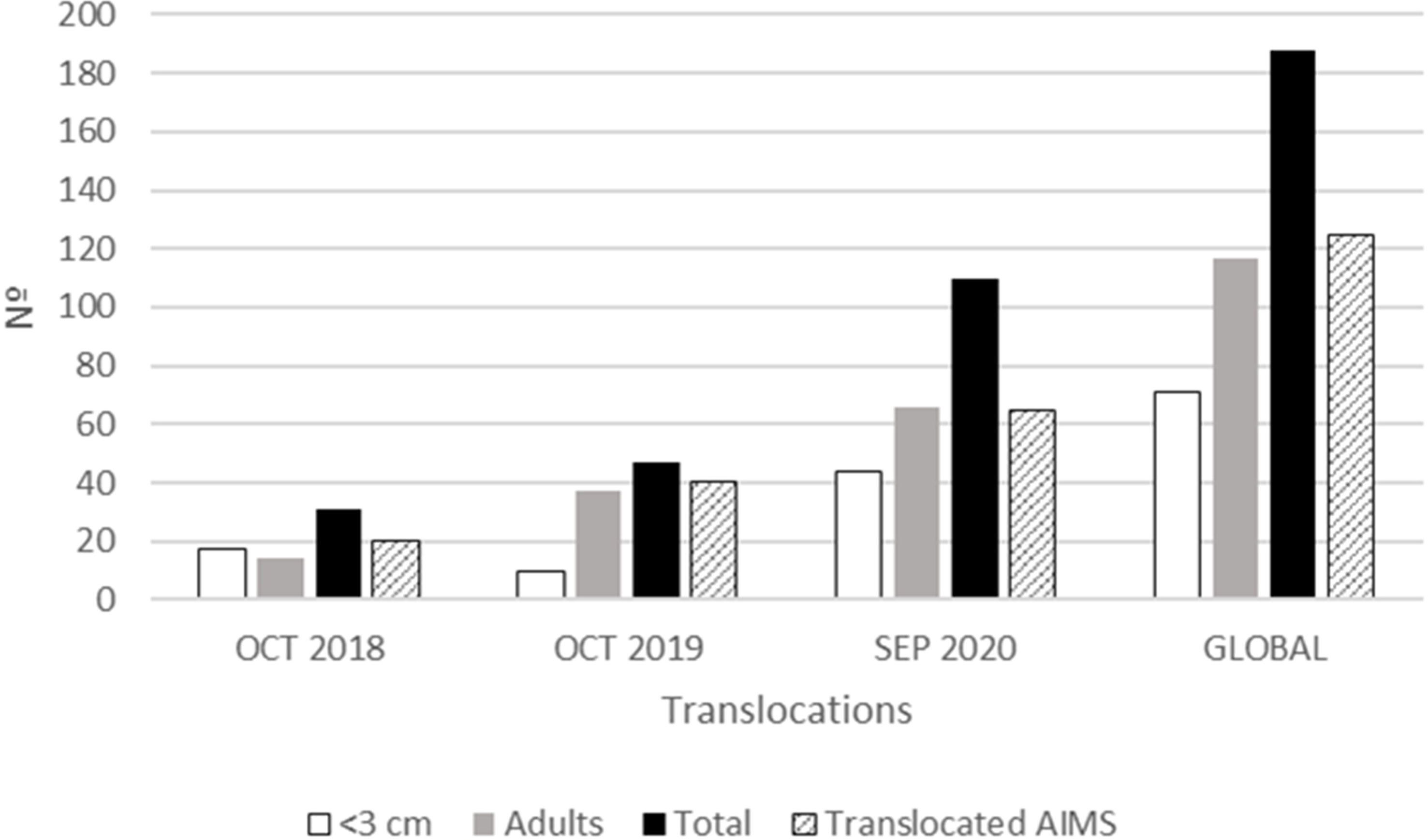
Figure 5 Number of individuals (juveniles, adults, and total) translocated per translocation event and at the end of the study (whole). The number of AIMS translocated is also shown.
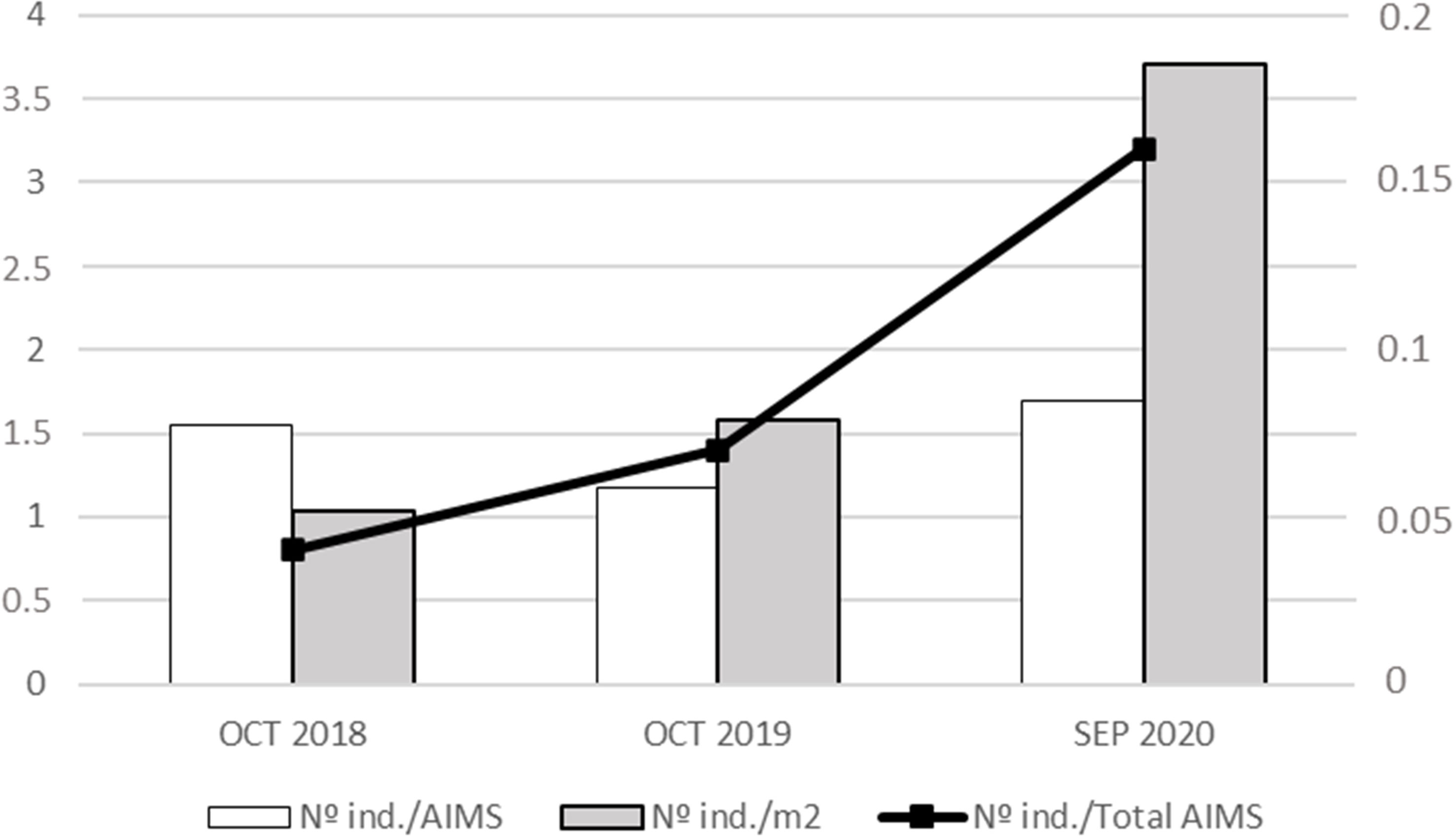
Figure 6 Density values per translocation event. No. individuals/AIMS: mean number of individuals per each of the translocated AIMS. No. ind./m2: mean number of individuals per square meter, considering the whole set of deployed AIMS, which were the available surface for settlement (660 AIMS = 29.7 m2). No. ind./Total AIMS: mean number of individuals settled considering the whole set of deployed AIMS (660). No. ind./AIMS and No. ind./m2 are represented by the left Y axis, whereas No. ind./Total AIMS is represented by the right Y axis.
Considering the different treatments used (AIMS deployed directly on substrate, sloped, or cantilever) the treatment that supported a greater number of Patella ferruginea individuals was onto the substrate, followed by slope and cantilever, and this pattern was constant for each of the three translocations (Figures 7A–D). The results indicated that the distribution of individuals among the three treatments clearly differed from a random one (Chi-square test: 60.1; df: 2; p<0.001). Nevertheless, recruits showed slight differences among translocations (Figures 7A–C), with a greater number on cantilever compared with slope AIMS on the second translocation. Moreover, the distribution of recruits and adults among treatments showed significant differences (Chi-square test: 26.9; df: 2; p<0.001).
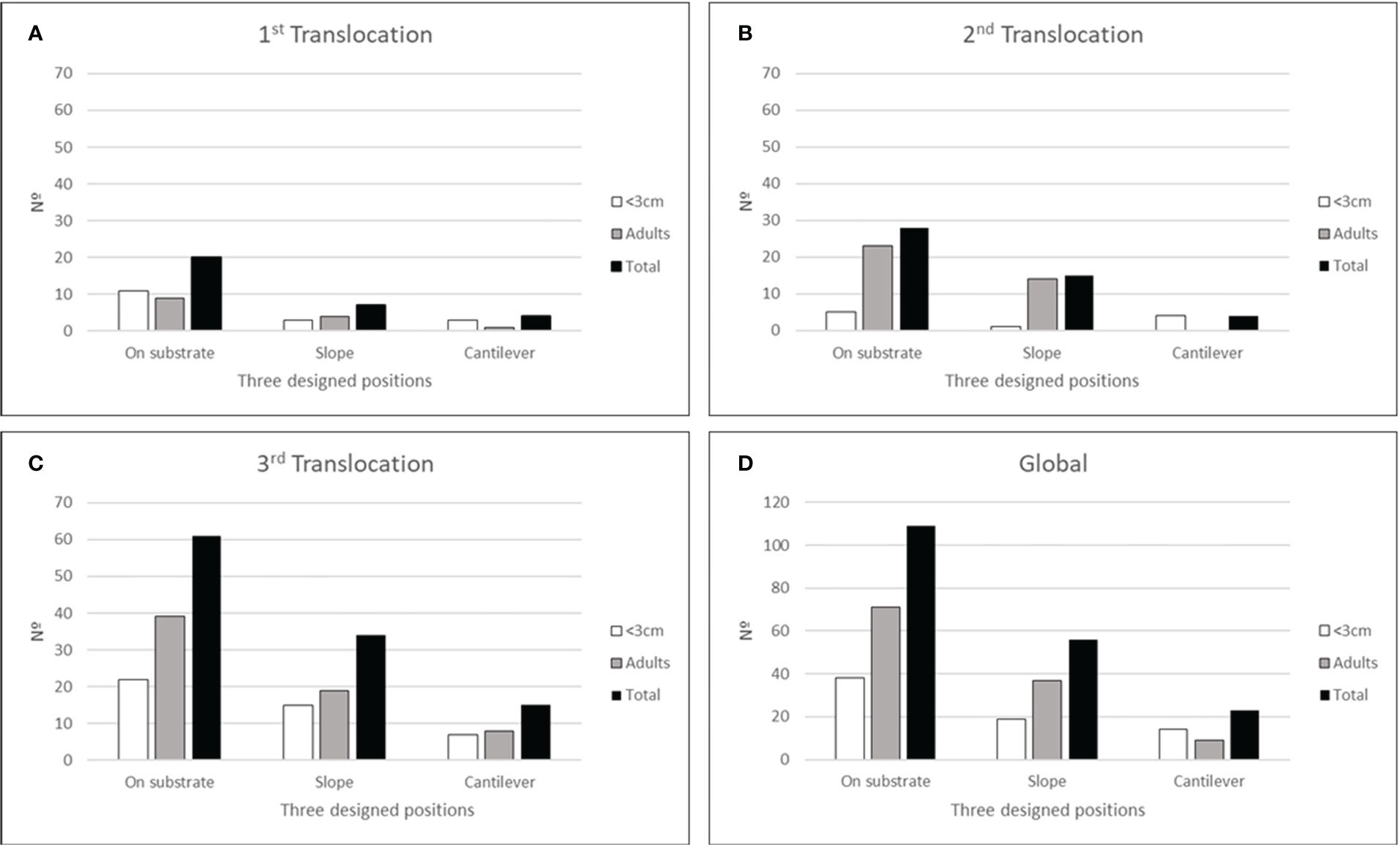
Figure 7 Number of individuals (<3cm, adults and total) settled on the three different treatments used (on substrate, slope, and cantilever). (A) first translocation; (B) second translocation; (C) third translocation; (D) the whole number of individuals after the three translocation events.
During the translocation procedure (from extracting in the donor population, transportation, and reallocation in the receiving population within 24 hours) no specimen died in any of the three different attempts. However, the survival of translocated specimens was progressively reduced through time, with differences among the three translocations (Figures 8A–C). Considering all translocations, the mean survival of the three translocations was 87.1% after one month, 75% after four months, and 68% after eight months (Figure 8D). Interestingly, differences were observed between the survival of adults and recruits for all the translocations, with it being greater for recruits than for adults, but significant differences were only recorded in the first translocation (Chi-square test: 1457.9; df: 9; p<0.05). When the three translocations were compared in terms of survival values they showed similar results for the first four months after the translocation, but the differences increased later (Figure 9), ranging from 55.5 to 80.9% after eight months, 48.4 to 76.6% after one year, and 41.9 to 66% after 20 months. Nevertheless, significant differences were only recorded between the second and the third translocations (Chi-square test: 13.4; df: 2; p<0.05). Survival of control individuals in the receiving population showed higher values, being 91.6% after four months, 87.5% after one year, and 83.3% after two years and at 32 months, the end of the monitored period, whereas these values fell to 75%, 62.5%, 41.9%, and 32.3%, respectively, for translocated individuals (Figure 9). The mean observed survival values of the three translocations differed from the expected control values (Chi-square test: 123.7; df: 8; p<0.001).
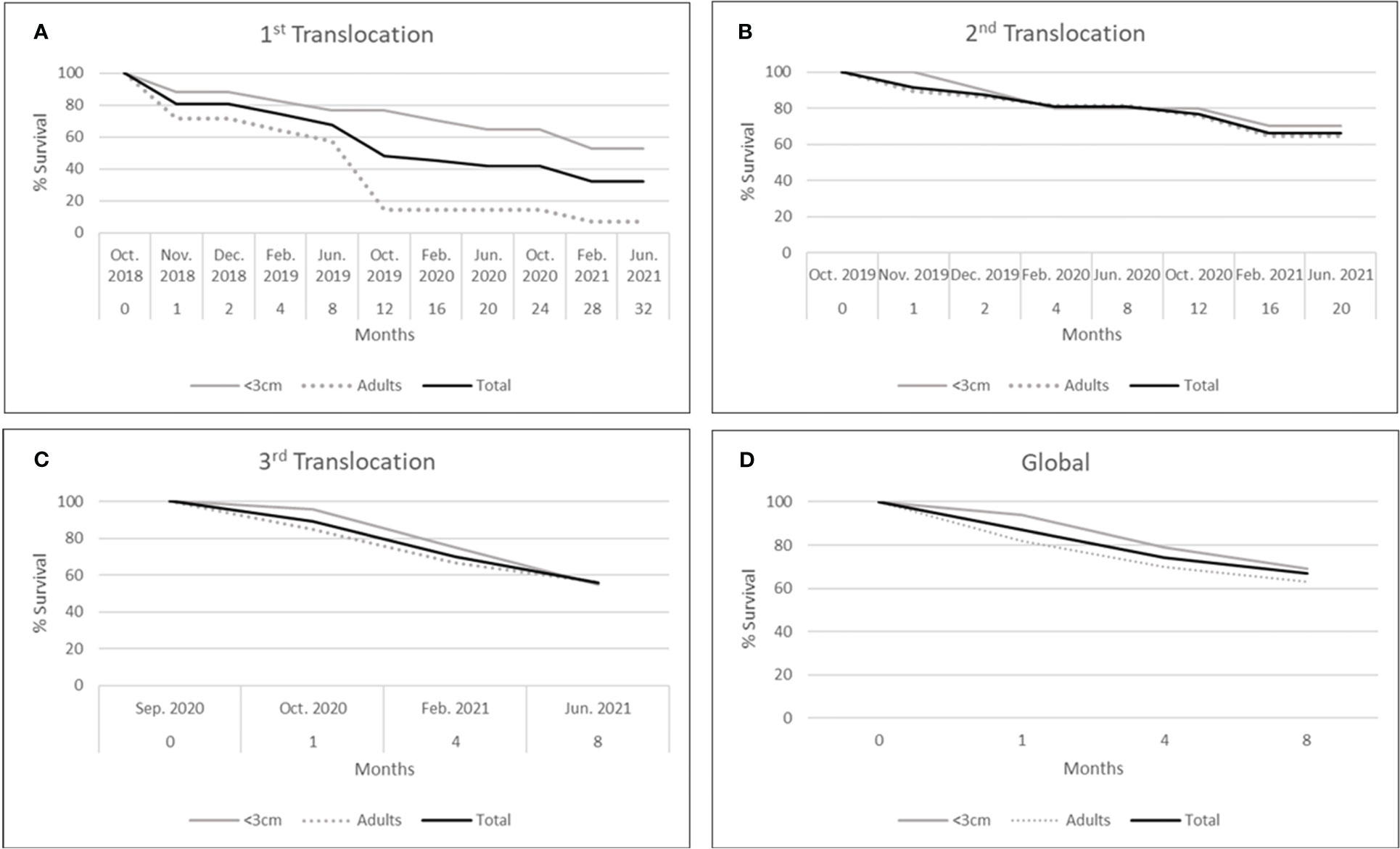
Figure 8 Survival percentage for each translocation done and size class (<3 cm: recruits, >3 cm: adults). (A): first translocation (2018); (B): second translocation (2019); (C): third translocation (2020); (D): mean values of the three translocations.
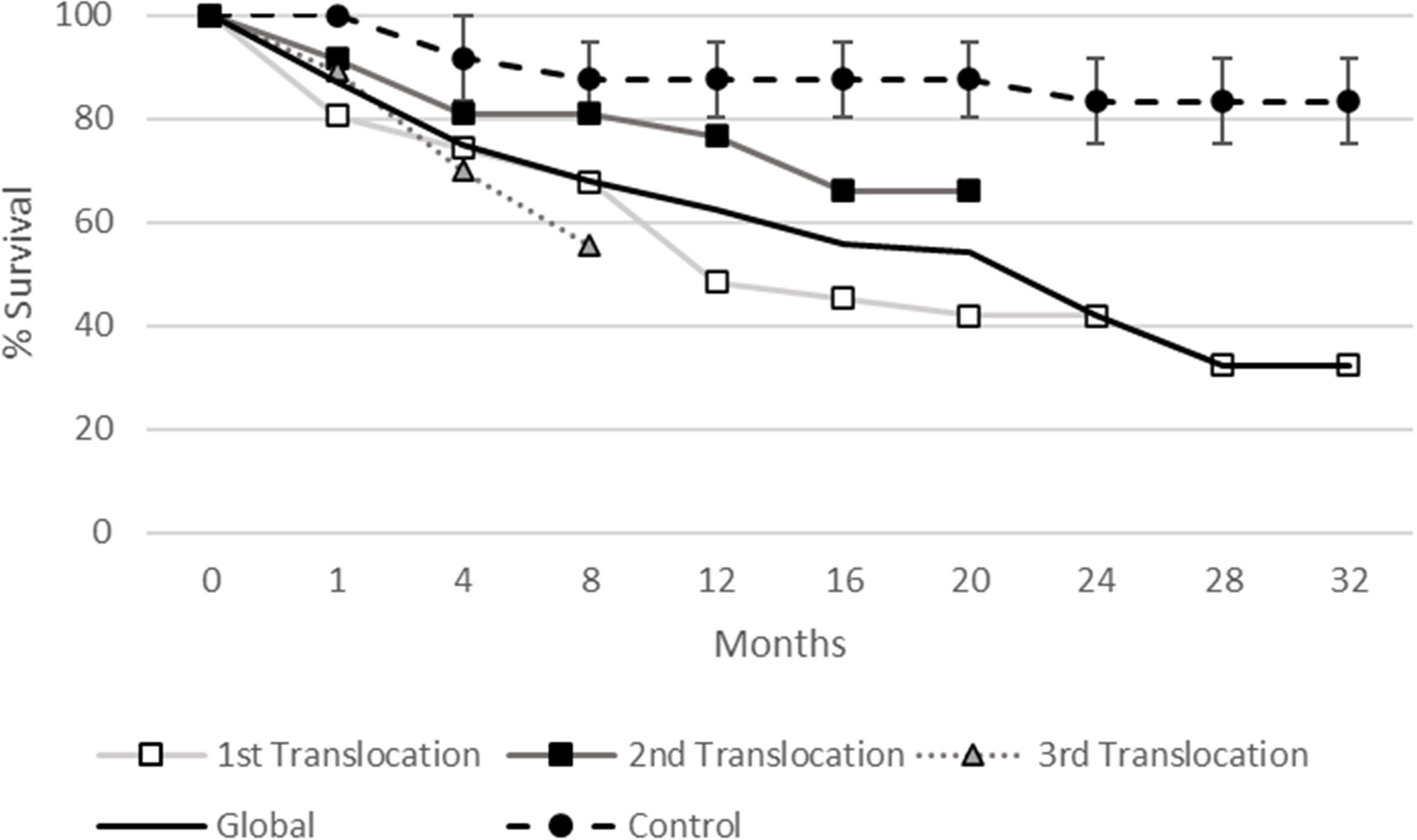
Figure 9 Survival percentage for each translocation and the mean of the three performed (total) over the whole monitoring period. The survival percentage of the control individuals in the receiving population located in La Línea port is also shown for comparison. Three 10 m transects were monitored and the mean ± standard error of their survival values is shown for the control threshold.
Growth of translocated individuals was consistently higher for recruits, but also showed differences for the time passed from translocation for the first and second attempts (Figure 10). In this sense, the growth rate was higher in the first month after translocation but decreased over time. The values ranged from 2.09 to 4.19 mm/month for recruits, 0.5 to 1.85 mm/month for adults, and 0.65 to 2.69 mm/month for the total of individuals translocated (recruits+adults). It is worth noting the great increase in the size of some individuals: number 7 during the first translocation increased in size from 2.2 cm to 8.1 cm in 31 months, and numbers 21 and 27 increased in size from 1 cm to 7.7 in the same period. In contrast, the number 36 during the second translocation only increased in size from 6.7 cm to 7.6 cm in 19 months and many others showed similar slow growth. Moreover, high variability in growth rates was recorded among individuals of similar size. In this sense, during the third translocation, specimen number 57 increased from 1.7 cm to 5.5 cm in eight months, whereas number 63 only increased from 1.6 cm to 2.3 cm in the same period.

Figure 10 Mean growth rates ( ± SE) for recruits (<3cm), adults, and the total individuals translocated for each of the translocation events. (A): first translocation (Oct. 2018). ***p<0.001 (ANOVA test). Letters indicate homogeneous groups according to a posteriori SNK test; (B): second translocation (Oct. 2019). ***p<0.001, **p<0.01 (ANOVA test); and (C) third translocation (Sep. 2020). ***p<0.001, **p<0.01, ns, not significant (ANOVA test). Data have been calculated by rating the increase in size (mm) by the months that have passed between time intervals in which size measurements were recorded. The individuals were considered adults if their size was greater than 3 cm at the beginning of the calculated period.
Discussion
Translocations of individuals for conservation purposes have played a central role in conserving endangered species and routinely yielded the intended benefits without producing unintended harm (Novak et al., 2021). From 1,014 taxa reviewed for 125 years, only a single case of biodiversity loss was reported by these authors in conservation-based governance translocations. Nevertheless, other authors, such as Strayer (2022), have challenged this optimistic view querying the validity of assessments of the costs and benefits of translocations. The viewpoint is that well-planned translocations could generate ecosystem benefits and should be weighed against the cost of inaction in conservation strategies.
The implemented translocation methodology proved to be feasible in terms of logistical constraints and achievements reached. First of all, the method proposed has been proven not to affect the donor or receiving population according to the results of the monitoring program carried out. The AIMS deployed did not affect the donor or receiving population accordingly with the temporal trends in terms of density and sizes. According to Kleiman et al. (1994), in any translocation, the survival of a wild population of an endangered species must never be jeopardized by such a conservation tool. There were differences in survival between translocation events, with the values of the 2019 translocation being very similar to the control population after one year (76.6% vs. 87.5%), whereas the first translocation, done in 2018, only achieved survival of 48.4% after one year. Different values of survival have been already obtained in previous studies, as well as relevant differences in growth rates, sex ratios, or food intake among specimens and populations (Rivera-Ingraham et al., 2011a; Espinosa and Rivera-Ingraham, 2017; Espinosa et al., 2021), indicating high intraspecific variability in life history traits that would be able to affect the specimen’s adaptation to different environmental conditions. The same findings had been already reported for other limpets elsewhere (Branch, 1981; Espinosa and Rivera-Ingraham, 2017). Nevertheless, the reported differences can also be a result of a mismatch among different researchers regarding the identification of the relevant temporal and spatial scales of measurement of survival rates in the focused species.
Notwithstanding the lack of broadly accepted criteria for assessing translocation success (Seddon, 1999; Fischer and Lindenmayer, 2000; Moehrenschlager et al., 2013; Robert et al., 2015), survival of released specimens is a main indicator (Swan et al., 2016). In this sense, assessments of success depend on time, and it is controversial because some authors report that the outcomes of long-term monitoring have much higher failure rates (Dalrymple et al., 2011), whereas others pointed out that the highest proportion of failures occurred within the first four years (Bubac et al., 2019). In fact, according to the review by these authors, most studies that conducted post-released monitoring lasted between 1-4 years, which is the range of monitoring of the present study. For Patella ferruginea, several translocations can be found in the literature. Summarizing, two different approaches have already been implemented: one in which the specimens were directly dislodged from the rock surface and reattached into a new one, and a second approach in which specimens were translocated with the substrate itself. The former has reported high mortality values in the first days or even hours after the translocation, due to the difficulty of adapting the shell edge to a new topography, provoking water loss and desiccation or increasing the chance of predation. In this sense, Laborel-Deguen and Laborel (1991b) translocated a total of 222 specimens in Corsica recording a survival of 50% after the first day, 25% after one year, and 10% after two years. Similarly, Espinosa et al. (2008) translocated 420 specimens in Ceuta (Strait of Gibraltar) reporting a survival of 50% after a few days, approximately 20% after one year, and 10% after two years, but survival in control specimens was also low: approximately 60% after one year and 30% after two years. Guallart et al. (2014) translocated ten specimens that were stored in aquariums after a captive breeding project to Hormigas Islands (Southern Spain), reporting a survival of 60% after 24 hours and 10% after two years. More recently, Zarrouk et al. (2018) used cages to protect the translocated specimens in Zembra Islands (Tunisia) and the survival was clearly improved (approximately 60% for caged specimens vs. 25% for non-caged specimens after almost two years). The survival rate of those specimens protected by cages was like the mean survival rate recorded in the present study after one year (approximately 60% in both studies), but higher after two years (60% vs. 41%). Survival of control specimens was slightly higher than 80% after one and two years in that study, similar to the survival obtained in the present study for the control specimens. Cages were removed after 43 days when the specimens had already fixed their new home scars and were not maintained the whole monitoring period. Consequently, the differences in survival between both studies cannot be attributed to this fact and were probably due to the different environmental and ecological conditions between donor and receiving sites that could trigger long-term mortality when the translocation is done to a faraway receiving area. In this sense, Zarrouk et al. (2018) translocated the specimens to a very close area (5 km between donor and receiving sites), but in that study, the authors also performed a long-distance translocation (185 km) in which the survival was very low (69% after two days and 18% after approximately one and two years of monitoring). Considering the second type of translocations reported in the literature (limpets translocated with their substrate), great differences exist between studies. For instance, due to maintenance works, a total of 301 specimens were translocated attached to the boulder rocks of a breakwater from a small harbor within the Chafarinas Islands (North Africa). Survival was only 37% after three months, even though the distance between donor and receiving sites was in the range of tens of meters (APM (Autoridad Portuaria de Melilla), 2014). However, also due to breakwater repair works, Fa et al. (2018) performed a similar translocation procedure to a very close receiving area in Gibraltar (Southern Iberian Peninsula), reporting survival of 87% after ten months for 97 specimens
Transplantation of other limpet species, such as Patella vulgata has been reported as a very difficult process indeed (Jenkins and Hartnoll, 2001), similar to the results obtained for P. ferruginea according to the information already mentioned. In this sense, home scars confer mechanical advantage primarily in shear resistance (Smith, 1992). Therefore, on one hand, the translocations performed better when the specimens were moved with the substrate in which they had already fixed their home scars, such as in the present study, and on the other hand, short-term translocations (<10 km) also performed better than long-term ones (>100 km). The ultimate reasons would be the importance of establishing the home scar for intertidal limpets, gaining an advantage for facing environmental stress and predation, whereas changes in the physicochemical composition of the water, different availability of trophic resources (in terms of quality and quantity), changes in competition intensity, and predation risk would be affecting the long-term survival for the translocated specimens. Despite being a long-distance translocation, the results of the present study, in terms of survival, are among the best outcomes reported in the literature, not only in the short term (100% of survival during translocation and almost 90% after one month) but also in the long term (62.5% after one year but 75% if the natural mortality is considered in the receiving site).
In addition, recruitment in the donor area and survival in the receiving area were not negatively affected by environmental factors in terms of great fluctuations, considering the stability of physicochemical parameters in the water column together with the maintenance of the ecological structure within intertidal assemblages. In this sense, profound changes in physicochemical parameters such as an increase in turbidity or a decrease in pH values are commonly associated with polluted waters within the study area (Estacio et al., 1997), whereas the ecological composition of the intertidal assemblages in the area has been proved to be very useful for monitoring studies and reflects their common history (Fa et al., 2002; Guerra-García et al., 2006). The decrease in mean size in the donor population during the monitoring period was not, consequently, attributable to any negative environmental issue. On the contrary, it could be explained by the fact that the density increased through the monitoring period mainly due to good recruitment events (both % of colonized AIMS and the density of specimens per m2 within them increased yearly, being a proxy for recruitment in the area), and the new recruits were progressively censused when they became adults, reducing the mean size of the whole population.
The success of recruitment on the AIMS increased from the first to the third translocation and it could be related to the maturity of the intertidal assemblages that were developed over time on the experimental structures and the presence of conspecifics of Patella ferruginea already settled nearby. Moreover, chemical cues either from conspecifics or from intertidal assemblages have been proposed as attractors for recruitment in limpets and particularly in P. ferruginea (see Rivera-Ingraham et al., 2011b, Rivera-Ingraham et al., 2015). Therefore, AIMS should be deployed at least one or two years before any translocation attempt is undertaken for maximizing the recruitment. In addition, recruitment has been greater on the AIMS deployed directly over the substrate, collecting either adults or recruits and it would be preferable for restocking since a greater number of specimens could be collected. However, provided the physical impact on the intertidal habitat and the greater mortality shown by adults the use of cantilever disposition could be interesting for preventing potential impacts on the intertidal habitat (in some areas of special concern) and for selecting recruits instead of adults. Bottari et al. (2017) also found greater mortality in translocated adults of the endangered mollusk Pinna nobilis compared with juveniles. Even though some authors have advocated against the translocation of adults especially during the reproductive season to avoid additional stress (Guallart, 2014), from a restocking perspective in conservation biology it could be very useful to increase genetic diversity in the receiving population, at least for those species that, in previous translocation experiences, have registered low adult mortality during the first critical months of adaptation to their new environment. Such specimens would release their gametes just after translocation and even if they died after, the translocation would be successful from a reproductive perspective. In this sense, Coates et al. (2013) indicated that restoring abalone populations (Haliotis corrugata) by releasing juveniles could limit the success because they are more susceptible to predation, show propensities for dispersal, and those reared in captive conditions may not exhibit some behaviors that promote survival. Furthermore, these authors stated that creating aggregations by transplanting adults may be an alternative to releasing juveniles due to the higher reproductive output. Delgado et al. (2004) pointed out that reproductive specimens of the queen conch (Strombus gigas) that were translocated spawned three months after, highlighting that it could be the key to expediting the recovery of this species in the Florida Keys. Similarly, Romero-Barón (2017) indicated that adult colonies of coral should be translocated to accelerate the recruitment process because those colonies would release larvae in the short term. Consequently, the mixed approach used in the present study could be more beneficial from a conservation perspective, although further studies would be required. Furthermore, the donor population from Melilla is adapted to warmer conditions than those found in the Strait of Gibraltar (see temperature map of the Mediterranean in Coll et al., 2010), and the introduction of alleles adapted to such conditions would enhance the resilience of the population in the receiving area against the impact of SST increase derived from climate change. In this sense, Garrabou et al. (2022) pointed out that mass mortality events in several taxa of invertebrates within the Mediterranean will be usual because of marine heatwaves, with the increase in genetic diversity being even more relevant.
The differences observed in growth rates between recruits and adults are concordant with similar findings in previous studies (see Espinosa et al., 2008) since the species shows a decrease in growth rate through the life span. Moreover, the decrease observed in growth rates through the monitoring period is an expected outcome, because the specimens reduce their growth rates progressively.
Moreover, the AIMS used have shown resistance against the intense shear forces generated by waves greater than 15 m, which were recorded within the donor area through the study (REMOPAF, 2021), nevertheless, the whole set of the AIMS remained fixed. That, coupled with its great versatility in terms of unscrewing from donor sites, transportation, and screwing on receiving sites, makes them a suitable procedure in translocation attempts. It is important to highlight that the AIMS could be reutilized when the translocated specimens had already fixed their home scars on the receiving substrate, being a feasible method for persistent restocking over time. In fact, Seddon et al. (2007) pointed out that technical aspects were being promoted in translocations for improving biological outcomes. Mimicking the natural substrate by using 3D printing technology has recently been proposed in marine restoration with promising results (see Evans et al., 2021), with the translocation procedure being used at the forefront of the knowledge in this field.
Many important populations of Patella ferruginea can be found settled on artificial substrates such as breakwaters or ripraps of harbor facilities or elsewhere (Espinosa et al., 2008; Guallart et al., 2013; CAGPDS, 2020). Therefore, artificial substrates could play a key role in conservation, and Artificial Marine Micro Reserves (AMMRs) have been proposed by García-Gómez et al. (2011) as a new tool in marine conservation Accordingly, discrete portions of breakwaters with dense populations of endangered species, such as P. ferruginea, could be fenced and monitored. Human harvesting should be effectively excluded within such areas since it has proven to be the main threat to the endangered Mediterranean ribbed limpet, and many other limpets (Branch and Odendaal, 2003; Espinosa and Rivera-Ingraham, 2017). These AMMRs should be integrated into a network of interconnected sites, constituting an Artificial Marine Micro Reserves Network (AMMRNs) (see García-Gómez et al., 2015). Finally, this type of figure would be within the concept of ‘Other Effective Area-Based Conservation Measures’ (OECMs) recently proposed by the IUCN (IUCN-WCPA Task Force on OECMs, 2019), as has been properly discussed by Ostalé-Valriberas et al. (2022). In this context, some of the AMMRs could be donor populations with a permanent system of AIMS deployed that would be translocated periodically to appropriate receiving sites. Therefore, the great number of AIMS that should be deployed for recruiting enough specimens would be located within artificial substrates, with the impact over the intertidal habitat (mainly visual) not being a major concern.
Furthermore, Pérez et al. (2012) pointed out that only 4% of translocations reviewed fully complied with translocation criteria, independently of the outcomes obtained for the target species. Considering that the present study complies with all the criteria already proposed by these authors, and the outcomes obtained, it could be considered not only adequately designed but also recommended for future translocations. In this sense, species of endangered intertidal or even shallow subtidal species within the Mediterranean basin, such as the limpet Patella candei from Macaronesia, the vermetid Dendropoma spp., or the orange coral Astroides calycularis, could potentially benefit (they are already included within endangered species list, see https://www.miteco.gob.es/es/biodiversidad/temas/conservacion-de-especies/especies-proteccion-especial/ce-proteccion-listado-situacion.aspx).
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
FE contributed to the conception, design, acquisition, analysis and interpretation of data. MM contributed to the conception, design, acquisition, analysis and interpretation of data. JG-G contributed to the conception, design and interpretation of data. MC-C contributed to the acquisition of data. CP-M contributed to the conception, design and acquisition of data. JP contributed to the conception and design. PF-SM contributed to the design, acquisition and analysis of data. NG-E contributed to the conception and design. All authors contributed to the article and approved the submitted version.
Funding
This study has been financially supported (60% of total eligible costs) by the European Commission through the LIFE project ReMoPaf (Recovery of the endangered Mollusc Patella ferruginea) with code LIFE15/NAT/ES/000987. The authors express their gratitude to ‘Autoridad Portuaria Bahía de Algeciras’ for providing support and collaboration during the project.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
APM (Autoridad Portuaria de Melilla) (2014). “Informe de seguimiento de los ejemplares de patella ferruginea afectados en el proyecto “Rehabilitación del embarcadero de chafarinas”,” in Informe técnico (no publicado) para la dirección general de sostenibilidad de la Costa y del mar, ministerio de agricultura (Madrid, España: Alimentación y Medio Ambiente), 55.
Barea-Azcón J. M., Ballesteros-Duperón E., Moreno D. (2008). Libro rojo de los invertebrados de andalucía. Consejería. Medio. Ambiente. Junta. Andalucía. Sevilla. 4.
BOE (1999). Boletín oficial del estado. 22 junio 1999, n° 148 (España: Ministerio de la Presidencia), 23921–23922.
Bottari T., Spinelli A., Busalacchi B., Rinelli P., Giacobbe S. (2017). Transplant trials of the fan mussel Pinna nobilis inside the coastal lagoon of capo peloro (Central Mediterranean, Italy). J. Shellfish. Res. 36 (1), 3–8. doi: 10.2983/035.036.0102
Branch G. M., Odendaal F. (2003). The effects of marine protected areas on the population dynamics of a south African limpet, Cymbula oculus, relative to the influence of wave action. Biol. Conserv. 114 (2), 255–269. doi: 10.1016/S0006-3207(03)00045-4
Bubac C. M., Johnson A. C., Fox J. A., Cullingham C. I. (2019). Conservation translocations and post-release monitoring: Identifying trends in failures, biases, and challenges from around the world. Biol. Conserv. 238, 108239. doi: 10.1016/j.biocon.2019.108239
Burgos-Rubio V., de la Rosa J., Altamirano M., Espinosa F. (2015). The role of patellid limpets as omnivorous grazers: a new insight into intertidal ecology. Mar. Biol. 162 (10), 2093–2106. doi: 10.1007/s00227-015-2739-0
CAGPDS (2020). Programa de gestión sostenible del medio marino andaluz. informe regional 2020 (Junta de Andalucía: Consejería de Agricultura, Ganadería, Pesca y Desarrollo Sostenible (CAGPDS).
Casu M., Rivera-Ingraham G. A., Cossu P., Lai T., Sanna D., Dedola G. L., et al. (2011). Patterns of spatial genetic structuring in the endangered limpet patella ferruginea: implications for the conservation of a Mediterranean endemic. Genetica 139 (10), 1293–1308. doi: 10.1007/s10709-012-9631-3
Coates J. H., Hovel K. A., Butler J. L., Klimley A. P., Morgan S. G. (2013). Movement and home range of pink abalone Haliotis corrugata: implications for restoration and population recovery. Mar. Ecol. Prog. Ser. 486, 189–201. doi: 10.3354/meps10365
Coll M., Piroddi C., Steenbeek J., Kaschner K., Ben Rais Lasram F., Aguzzi J., et al. (2010). The biodiversity of the Mediterranean Sea: estimates, patterns, and threats. PloS One 5 (8), e11842. doi: 10.1371/journal.pone.0011842
Dalrymple S. E., Stewart G. B., Pullin A. S. (2011) Are re-introductions an effective way of mitigating against plant extinctions? collaboration for environmental evidence review 07–008 (SR32). Available at: www.environmentalevidence.org/SR32.html.
Delgado G. A., Bartels C. T., Glazer R. A., Brown-Peterson N. J., McCarthy K. J. (2004). Translocation as a strategy to rehabilitate the queen conch (Strombus gigas) population in the Florida keys. Fishery. Bull. 102 (2), 278.
Denny M. W., Shibata M. F. (1989). Consequences of surf-zone turbulence for settlement and external fertilization. Am. Nat. 134, 859–889. doi: 10.1086/285018
Dissanayake A., Bamber S. D. (2010). Monitoring PAH contamination in the field (South west Iberian peninsula): Biomonitoring using fluorescence spectrophotometry and physiological assessments in the shore crab carcinus maenas (L.)(Crustacea: Decapoda). Mar. Environ. Res. 70 (1), 65–72. doi: 10.1016/j.marenvres.2010.03.003
Espinosa F., González A. R., Maestre M. J., Fa D., Guerra-García J. M., García-Gómez J. C. (2008). Responses of the endangered limpet Patella ferruginea to reintroduction under different environmental conditions: survival, growth rates and life-history. Ital. J. Zool. 75 (4), 371–384. doi: 10.1080/11250000801887740
Espinosa F., Guerra-García J. M., Fa D., García-Gómez J. C. (2006b). Effects of competition on an endangered limpet patella ferruginea (Gastropoda: Patellidae): Implications for conservation. J. Exp. Mar. Biol. Ecol. 330 (2), 482–492. doi: 10.1016/j.jembe.2005.09.020
Espinosa F., Guerra-Garcia J. M., FA D., Garcia-Gomez J. C. (2006a). Aspects of reproduction and their implications for the conservation of the endangered limpet, Patella ferruginea. Invertebrate. Reprod. Dev. 49 (1-2), 85–92. doi: 10.1080/07924259.2006.9652197
Espinosa F., Pavón-Paneque A., García-Gómez J. C. (2021). Disentangling the impact of artificial substrata on the trophic ecology of the highly endangered marine invertebrate Patella ferruginea gmelin 1791. Molluscan. Res. 41 (1), 41–45. doi: 10.1080/13235818.2021.1882925
Espinosa F., Rivera-Ingraham G. A. (2017). Biological conservation of giant limpets: the implications of large size. Adv. Mar. Biol. 76, 105–155. doi: 10.1016/bs.amb.2016.10.002
Espinosa F., Rivera-Ingraham G., García-Gómez J. C. (2011). Influence of habitat structure and nature of substratum on limpet recruitment: Conservation implications for endangered species. Estuarine. Coast. Shelf. Sci. 94 (2), 164–171. doi: 10.1016/j.ecss.2011.06.005
Espinosa F., Rivera-Ingraham G. A., Maestre M., González A. R., Bazairi H., García-Gómez J. C. (2013). Updated global distribution of the threatened marine limpet Patella ferruginea (Gastropoda: Patellidae): An example of biodiversity loss in the Mediterranean. Oryx 48, 266–275. doi: 10.1017/S0030605312000580
Espinosa F., Rivera-Ingraham G. A., Ostalé-Valriberas E., García-Gómez J. C. (2018). Predicting the fate of the most endangered marine invertebrate of the Mediterranean: The power of long-term monitoring in conservation biology. Aquat. Conserv.: Mar. Freshw. Ecosyst. 28 (6), 1283–1293. doi: 10.1002/aqc.2944
Estacio F. J., García-Adiego E. M., Fa D. A., García-Gómez J. C., Daza J. L., Hortas F., et al. (1997). Ecological analysis in a polluted area of algeciras bay (Southern spain): external ‘versus’ internal outfalls and environmental implications. Mar. pollut. Bull. 34 (10), 780–793. doi: 10.1016/S0025-326X(97)00046-5
Evans A. J., Lawrence P. J., Natanzi A. S., Moore P. J., Davies A. J., Crowe T. P., et al. (2021). Replicating natural topography on marine artificial structures–a novel approach to eco-engineering. Ecol. Eng. 160, 106144. doi: 10.1016/j.ecoleng.2020.106144
Fa D. A., Finlayson J. C., Finlayson G., Giles-Pacheco F., Rodríguez-Vidal J., Gutiérrez-López J. M. (2016). Marine mollusc exploitation as evidenced by the gorham's cave (Gibraltar) excavations 1998–2005: The middle–upper Palaeolithic transition. Quaternary. Int. 407, 16–28. doi: 10.1016/j.quaint.2015.11.148
Fa D. A., Finlayson C., García-Adiego E., Sánchez-Moyano J. E., García-Gómez J. C. (2002). Influence of some environmental factors on the structure and distribution of the rocky shore macrobenthic communities in the bay of Gibraltar: Preliminary results. Almoraima 28, 73–88.
Fa D. A., Finlayson G., Sempere-Valverde J., García-Gómez J. C. (2018). ‘Impossible’re-introduction of the endangered limpet Patella ferruginea gmelin 1791? progress in resolving translocation mortality. Endangered. Species. Res. 37, 219–232. doi: 10.3354/esr00921
Fernández-Casado M., Arroyo M. C., Vivas M. S. (2017). Caracterización de las poblaciones de la lapa ferruginosa (Patella ferruginea gmelin 1971) de la costa norte del estrecho de gibraltar”. Almoraima. Rev. Estudios. Campogibraltareños. 47, 181–202.
Fernández-Macho J. (2016). Risk assessment for marine spills along European coastlines. Mar. pollut. Bull. 113 (1-2), 200–210. doi: 10.1016/j.marpolbul.2016.09.015
Ferranti M. P., Monteggia D., Asnaghi V., Dagnino A., Gaino F., Moretto P., et al. (2019). Distribution of the Mediterranean ribbed limpet Patella ferruginea gmelin 1791 along the ligurian coast and implications for conservation actions. Mediterr. Mar. Sci. 20 (3), 496–501. doi: 10.12681/mms.20209
Fischer J., Lindenmayer D. B. (2000). An assessment of the published results of animal relocations. Biol. Conserv. 96 (1), 1–11. doi: 10.1016/S0006-3207(00)00048-3
García-Gómez J. C., Guerra-García J. M., Espinosa F., Maestre M. J., Rivera-Ingraham G., Fa D., et al. (2015). Artificial m arine m icro-r eserves n etworks (AMMRN s): an innovative approach to conserve marine littoral biodiversity and protect endangered species. Mar. Ecol. 36 (3), 259–277. doi: 10.1111/maec.12167
García-Gómez J. C., López-Fé C. M., Espinosa F., Guerra-García J. M., Rivera-Ingraham G. A. (2011). Marine artificial micro-reserves: A possibility for the conservation of endangered species living on artificial substrata. Mar. Ecol. 32 (1), 6–14. doi: 10.1111/j.1439-0485.2010.00409.x
Garrabou J., Gómez-Gras D., Medrano A., Cerrano C., Ponti M., Schlegel R., et al. (2022). Marine heatwaves drive recurrent mass mortalities in the Mediterranean Sea. Global Change Biol. 28, 5708–5725 doi: 10.1111/gcb.16301
González-Laxe F., Martín-Bermúdez F., Martín-Palmero F., Novo-Corti I. (2017). Assessment of port sustainability through synthetic indexes. application to the Spanish case. Mar. pollut. Bull. 119 (1), 220–225. doi: 10.1016/j.marpolbul.2017.03.064
Guallart J. (2014). Condicionantes de la biología de la lapa ferruginosa (Patella ferruginea) en vistas a potenciales traslados de ejemplares (Ministerio de Agricultura, Alimentación y Medio Ambiente, España: Informe Técnico (no publicado), 98.
Guallart J., Luque Á.A., Acevedo I., Calvo M. (2013). Distribución y censo actualizado de la lapa ferrugínea (Patella ferruginea gmelin 1791) en el litoral de melilla (Mediterráneo suroccidental). Iberus 31 (1), 21–51.
Guallart J., Templado J., Acevedo I., Calvo M., Machordom A., Luque A. A., et al. (2014). Informe de la reintroducción de ejemplares de la lapa patella ferruginea (catalogada “en peligro de extinción”) en la isla hormiga (Murcia) (Museo Nacional de Ciencias Naturales-CSIC: Informe técnico), 15.
Guerra-García J. M., Maestre M. J., González A. R., García-Gómez J. C. (2006). Assessing a quick monitoring method using rocky intertidal communities as a bioindicator: a multivariate approach in algeciras bay. Environ. Monit. Assess. 116 (1), 345–361. doi: 10.1007/s10661-006-7572-y
IUCN-WCPA Task Force on OECMs (2019). Recognising and reporting other effective area-based conservation measures (Gland, Switzerland: IUCN).
Jenkins S. R., Hartnoll R. G. (2001). Food supply, grazing activity and growth rate in the limpet Patella vulgata l.: a comparison between exposed and sheltered shores. J. Exp. Mar. Biol. Ecol. 258 (1), 123–139. doi: 10.1016/S0022-0981(01)00211-8
Kleiman D. G., Price M. R. S., Beck B. B. (1994). “Criteria for reintroductions,” in Creative conservation: interactive management of wild and captive animals. Eds. Olney P. J. S., Mace G. M., Feistner A. T. C. (London, UK: Chapman & Hall).
Laborel-Deguen F., Laborel J. (1991a). “Statut de patella ferruginea gmelin en méditerranée,” in Les Espèces marines à protéger en méditerranée. Eds. Boudouresque C. F., Avon M., Gravez V. (Marseille: GIS Posidonie Publishers), 91–103.
Laborel-Deguen F., Laborel J. (1991b). “Une tentative de reintroduction de patella ferruginea gmel. (Gastropoda) dans le parc national de port-cros (Var, France),” in Les Espèces marines à protéger en méditerranée. Eds. Boudouresque C. F., Avon M., Gravez V. (Marseille: GIS Posidonie Publishers), 129–132.
Martins G. M., Thompson R. C., Neto A. I., Hawkins S. J., Jenkins S. R. (2010). Enhancing stocks of the exploited limpet Patella candei d’Orbigny via modifications in coastal engineering. Biol. Conserv. 143 (1), 203–211. doi: 10.1016/j.biocon.2009.10.004
Meffe G. K. (1992). Techno-arrogance and halfway technologies: salmon hatcheries on the pacific coast of north America. Conserv. Biol. 6, 350–354. doi: 10.1046/j.1523-1739.1992.06030350.x
Moehrenschlager A., Shier D. M., Moorhouse T. P., Stanley Price M. R. (2013). Righting past wrongs and ensuring the future: challenges and opportunities for effective reintroductions amidst a biodiversity crisis. Key. Topics. Conserv. Biol. 2, 405–429. doi: 10.1002/9781118520178.ch22
Novak B. J., Phelan R., Weber M. (2021). US Conservation translocations: Over a century of intended consequences. Conserv. Sci. Pract. 3 (4), e394. doi: 10.1111/csp2.394
Ostalé-Valriberas E., Sempere-Valverde J., Pavón-Paneque A., Coppa S., Espinosa F., García-Gómez J. C. (2022). Artificial marine micro-reserves as a new ecosystem-based management tool for marine conservation: The case of Patella ferruginea (Gastropoda, patellidae), one of the most endangered marine invertebrates of the Mediterranean. Mar. Policy 136, 104917. doi: 10.1016/j.marpol.2021.104917
Pérez I., Anadón J. D., Díaz M., Nicola G. G., Tella J. L., Giménez A. (2012). What is wrong with current translocations? a review and a decision-making proposal. Front. Ecol. Environ. 10, 494–501. doi: 10.1890/110175
Potet M., Fabien A., Chaudemanche S., Sebaibi N., Guillet T., Gachelin S., et al. (2021). Which concrete substrate suits you? Ostrea edulis larval preferences and implications for shellfish restoration in Europe. Ecol. Eng. 162, 106159. doi: 10.1016/j.ecoleng.2021.106159
Primack R. B., Ros J. (2002). Introducción a la biología de la conservación (Barcelona, Spain: Grupo Planeta (GBS)).
Ramos M. A. (1998). Implementing the habitats directive for mollusc species in Spain. J. Conchol. Special. Publ. 2, 125–132.
Rees P. A. (2001). Is there a legal obligation to reintroduce animal species into their former habitats? Oryx 35, 216–223. doi: 10.1046/j.1365-3008.2001.00178.x
REMOPAF (2021). “Recovery of endangered mollusc patella ferruginea population by artificial inert mobile substrates in Mediterranean Sea,” in Madrid, Acciona Ingeniería Final report. LIFE project n° LIFE15 NAT/ES/000987, 102.
Reyers B., Folke C., Moore M. L., Biggs R., Galaz V. (2018). Social-ecological systems insights for navigating the dynamics of the anthropocene. Annu. Rev. Environ. Resour. 43, 267–289. doi: 10.1146/annurev-environ-110615-085349
Ricciardi A., Simberloff D. (2009). Assisted colonization is not a viable conservation strategy. Trends Ecol. Evol. 24, 248–253. doi: 10.1016/j.tree.2008.12.006
Rivera-Ingraham G. A., Espinosa F., García-Gómez J. C. (2011a). Environmentally mediated sex change in the endangered limpet patella ferruginea (Gastropoda: Patellidae). J. Molluscan. Stud. 77 (3), 226–231. doi: 10.1093/mollus/eyr007
Rivera-Ingraham G. A., Espinosa F., Garcia-Gomez J. C. (2011b). Effect of γ-amino butyric acid on limpet populations: towards the future management and conservation of endangered patellid species. J. Chem. Ecol. 37 (1), 1–9. doi: 10.1007/s10886-010-9884-1
Rivera-Ingraham G. A., Espinosa F., Krock B. (2015). Presence of gamma-aminobutyric acid (GABA) in the pedal mucus of the critically endangered species patella ferruginea. J. Chem. Ecol. 41 (5), 501–504. doi: 10.1007/s10886-015-0579-5
Robert A., Colas B., Guigon I., Kerbiriou C., Mihoub J. B., Saint-Jalme M., et al. (2015). Defining reintroduction success using IUCN criteria for threatened species: a demographic assessment. Anim. Conserv. 18, 397–406. doi: 10.1111/acv.12188
Rojo-Nieto E., Garrido-Pérez C., Anfuso-Melfi G., Lopez-Aguayo F., Sales-Marquez D., Perales-Vargas-Machuca J. A. (2011). The zoning of semi-enclosed bodies of water according to the sediment pollution: The bay of algeciras as a case example. Estuaries. costas. 34, 1129–1139. doi: 10.1007/s12237-011-9389-3
Rojo-Nieto E., Perales J. A. (2015). Estimating baseline toxicity of PAHs from marine chronically polluted sediments and bioaccumulation in target organs of fish hypothetically exposed to them: a new tool in risk assessment. Environ. Sci.: Processes. Impacts. 17 (7), 1331–1339. doi: 10.1039/c5em00111k
Romero-Barón M. F. (2017). La restauración de arrecifes de coral; una alternativa de conservación en el sistema arrecifal mesoamericano (SAM) (Université de Sherbrooke: Tesis Profesional en Ecología Internacional).
Seddon P. J. (1999). Persistence without intervention: assessing success in wildlife reintroductions. Trends Ecol. Evol. 14, 503–503. doi: 10.1016/S0169-5347(99)01720-6
Seddon P. J., Armstrong D. P., Maloney R. F. (2007). Developing the science of reintroduction biology. Conserv. Biol. 21, 303–312. doi: 10.1111/j.1523-1739.2006.00627.x
Seddon P. J., Soorae P. S., Launay F. (2005). Taxonomic bias in reintroduction projects. Anim. Conserv. 8, 51–158. doi: 10.1017/S1367943004001799
Smith A. M. (1992). Alternation between attachment mechanisms by limpets in the field. J. Exp. Mar. Biol. Ecol. 160, 205–220. doi: 10.1016/0022-0981(92)90238-6
Spalding C., Hull P. M. (2021). Towards quantifying the mass extinction debt of the anthropocene. Proc. Biol. Sci. 288, 20202332. doi: 10.1130/abs/2021AM-369486
Strayer D. L. (2022). Comment: Novak et al.(2021) overestimated the successes of species translocations and minimized their risks. Conserv. Sci. Pract. 4 (7). doi: 10.1111/csp2.12694
Swan K. D., McPherson J. M., Seddon P. J., Moehrenschlager A. (2016). Managing marine biodiversity: the rising diversity and prevalence of marine conservation translocations. Conserv. Lett. 9 (4), 239–251. doi: 10.1111/conl.12217
Tilman D., Isbell F., Cowles. J. M. (2014). Biodiversity and ecosystem functioning. Annu. Rev. Ecol. Evol. Syst. 45, 471–493. doi: 10.1146/annurev-ecolsys-120213-091917
Vázquez-Luis M., Álvarez E., Barrajón A., García-March J. R., Grau A., Hendriks I. E., et al. (2017). SOS Pinna nobilis: a mass mortality event in western Mediterranean Sea. Front. Mar. Sci. 4, 220. doi: 10.3389/fmars.2017.00220
Keywords: Patella ferruginea, translocation, conservation, endangered species, limpets
Citation: Espinosa F, Maestre M, García-Gómez JC, Cotaina-Castro MI, Pitarch-Moreno C, Paramio JM, Fort-Santa María P and García-Estévez N (2023) Joining technology and biology to solve conservation problems through translocation in the endangered limpet Patella ferruginea. Front. Mar. Sci. 10:1100194. doi: 10.3389/fmars.2023.1100194
Received: 16 November 2022; Accepted: 16 March 2023;
Published: 29 March 2023.
Edited by:
Stephen John Hawkins, University of Southampton, United KingdomReviewed by:
Gustavo Martins, University of the Azores, PortugalTomas Vega Fernandez, Anton Dohrn Zoological Station Naples, Italy
Copyright © 2023 Espinosa, Maestre, García-Gómez, Cotaina-Castro, Pitarch-Moreno, Paramio, Fort-Santa María and García-Estévez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Free Espinosa, ZnJlZUB1cy5lcw==