 Cristina Cedeño-Posso*
Cristina Cedeño-Posso* Martha Vides-CasadoVenus Rocha
Martha Vides-CasadoVenus Rocha Giomar H. Borrero-Pérez
Giomar H. Borrero-Pérez Andrea Polanco F.
Andrea Polanco F. David Alonso
David Alonso- Marine and Coastal Research Institute, INVEMAR, Santa Marta, Colombia
Corales de Profundidad National Natural Park (CPNNP), at the central Colombian Caribbean margin, has an extension of 142.195 ha with depths ranging from 34 to 1,234 m. The CPNNP’s essential ecological value is Madracis spp. as potential structure-forming deep-water coral; this work represents the first footages of these unique habitats for the Colombian Caribbean. In 11 sectors, remote video surveys are conducted, based on a detailed digital elevation model. From these sectors, four cover mesophotic zones (46–169m depth), and seven cover aphotic zones (up to 354m depth). After still images’ description and interpretation, a guide was generated with nine types of macrohabitats surrounding the coral formations of the CPNNP: soft substrate (mud/sand), soft substrate/rubble/rock with solitary corals, coral and shell rubble, rock/ledges with attached fauna, rugged stones with attached fauna, hard substrate with attached fauna, sandstone with attached fauna, soft substrate with burrows, and pockmark with attached fauna. Video transect analysis confirms the occurrence of Madracis spp., with coral patches as the main framework builder of the CPNNP. These patches comprise many clumped 50 to 80 cm high coral colonies that developed over slow slopes, with seafloor elevations between 107 and 233 m and textured soils of soft and not very compact ridges, associated with the macrohabitat sandstone with attached fauna. This information is being used for the management and monitoring of this unique protected area.
1. Introduction
The major deep-sea habitats or megahabitats according to Roberts et al. (2009) have dimensions of kilometers, like the abyssal plains, mid-ocean ridges, hydrothermal vents, trenches, and the ones located in the continental margins that include sedimentary slopes, submarine canyons, cold seeps, mud volcanoes, pockmarks, and oxygen minimum zones (Ramirez-Llodra, 2020). Some of them have already been described in different regions for the Colombian continental margin, such as cold seeps (Gracia and Ardila, 2010; Gracia et al., 2011; Digby et al., 2016; Cosel and Gracia, 2018; Dueñas et al., 2021). On another scale are macrohabitats, defined as 1 to 10 m, including soft bottoms, rocky outcrops, biogenic habitats such as sponges, coral carbonate mounds, and cold-water corals (Roberts et al., 2009). Adding to the above, three-dimensional structures of cheilostome bryozoans have also been recognized as essential bio-builders, providing structures and spaces for the establishment of other organisms (Flórez-Romero et al., 2007).
Cold-water corals are nowadays known to be almost globally distributed (Levin et al., 2019), and despite having evidence of 18 deep-sea azooxanthellate corals in the Colombian Caribbean, based on the material collected by the R/V Albatross, R/V Oregon in the 1960s, and R/V Pillsbury in 1972 campaigns (Cairns, 1979; Reyes et al., 2009), it was not until the years 1998 and 2002 that the presence of coral banks in the Colombian Caribbean became evident (Reyes et al., 2005; Flórez and Santodomingo, 2010). The collection of species of cup corals and other azooxanthellate corals along three sites in the Colombian Caribbean was the result of systematic bottom trawl surveys, between 20 and 520 m depth, conducted by INVEMAR researchers on “Macrofauna” cruises (1998–2002) (Reyes et al., 2005; Santodomingo et al., 2007; Santodomingo et al., 2013). One of those findings, in the San Bernando locality, across the Sinú Accretionary Prism offshore Colombia (Vinnels et al., 2010) triggered the designation of the Corales de Profundidad National Natural Park (CPNNP) in 2013 (Category II—IUCN), becoming the first Colombian underwater marine National Natural Park aiming to conserve the deep-sea coral formations, especially Madracis spp., as a primary habitat-forming species (Alonso et al., 2015; Alonso et al., 2021). The Park covers an extension of 142.195 ha with depths ranging from 34 to 1,234 m.
Mapping marine habitats makes it possible to visualize their spatial distribution and the quality and quantity of deep-sea resources, with the ultimate goal of generating a better understanding of the environment and thus having inputs for management, planning, policy, and research, in support of conservation and sustainable development. To describe and define the extent of CPNNP’s benthic macrohabitats, scientific exploration was carried out in 2015, with the mission of conducting a non-disturbing video sampling. The collection of biological information was done through video surveys with a remotely operated vehicle (ROV), which made it possible to cover a large area in less time and to incorporate more information about the area that could complement the collections carried out between 1998 and 2002. In this work, we address the variation of macrohabitats in the CPNNP and their variability between mesophotic and aphotic environments and discuss the distribution of Madracis spp. coral patches in the National Natural Park.
2 Materials and methods
2.1 Data acquisition and processing
Expedition PNN Corales de Profundidad was conducted aboard the R/V Los Angeles between 6 and 18 October 2015. The first step was to select the sectors that were evaluated; this was done taking into account the analysis performed with a digital elevation model and resolution of 10 m per pixel, obtained from the bathymetric data collected in a campaign performed by the Maritime General Directorate (DIMAR) (Morales et al., 2017) and the collection of information from the study area (Reyes et al., 2005; Santodomingo et al., 2007; Santodomingo et al., 2013). Eleven sectors (S) were chosen, which cover nine geoforms to make video surveys with an ROV Diavolo II (E384–E394) (Figure 1A and Supplementary Table S1).
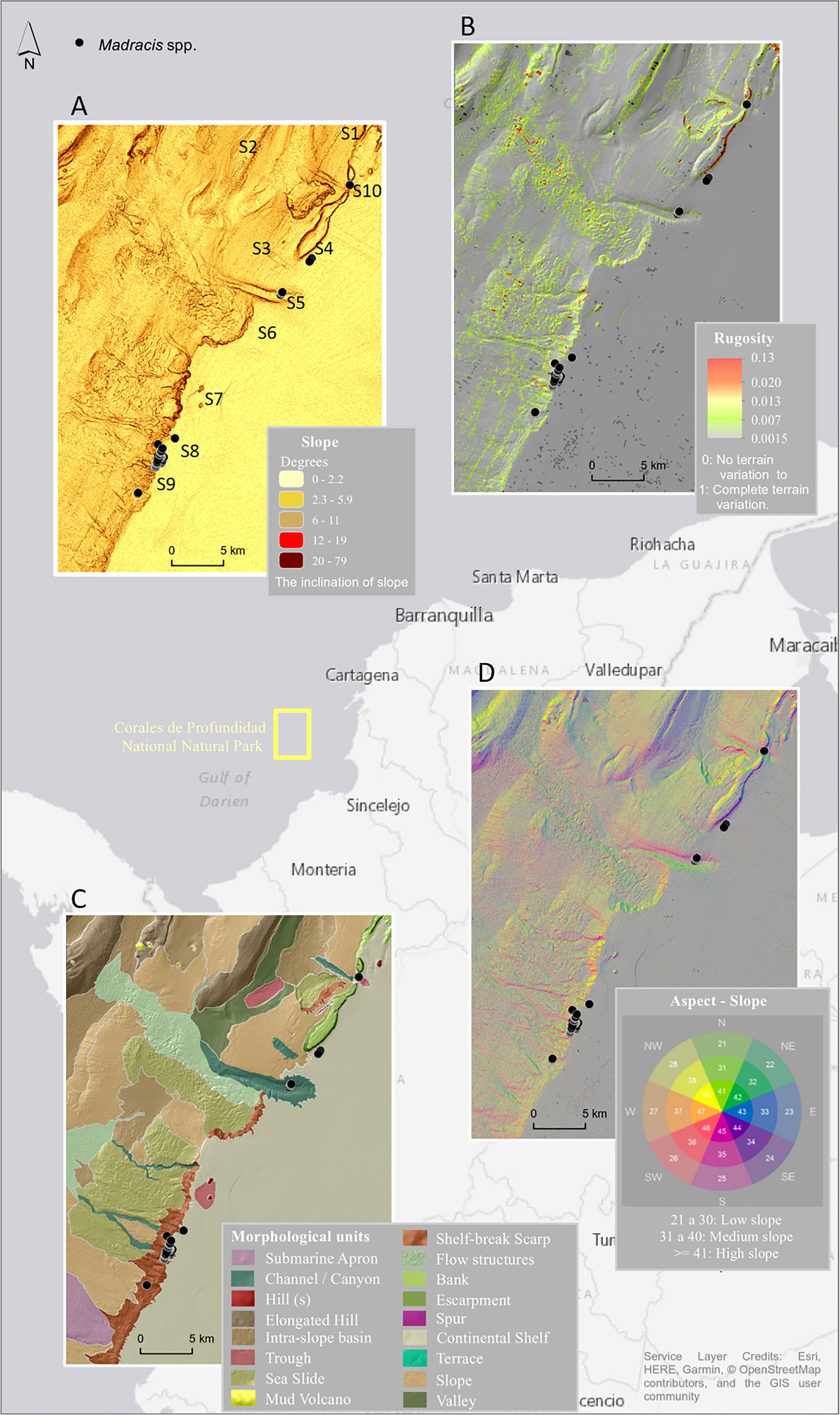
Figure 1 Study areas and sectors (S) where video transects were carried out; black dots are the location of Madracis spp. coral patch occurrences obtained from the analysis of the ROV video transects. (A) Slope map. (B) Rugosity. (C) Morphological units. (D) Aspect map. The background image shows the area of the Corales de Profundidad National Natural Park in the Colombian Caribbean. ROV, remotely operated vehicle.
ROV linear video transects were made at each sector, in water depths between 46 and 354 m, to characterize qualitatively the benthic macrohabitats and their associated macrobiota and fishes. The equipment had a front camera (¼″ CCD 380 I TV-lines, 0.1 Lux/F = 1.2) pointed ~30° to view near the bottom and far the horizon, with 10 cm parallel lasers for scale. Positioning was estimated using an underwater acoustic positioning method like an ultra-short baseline (USBL) mounted on the vehicle, which gave a geographic position every 30 s approximately. In each sector, ROV dives ranged from 1 to 4 h in length, covering approximately length 1 km (0.54 nm), sometimes doubling the time when necessary depending on the benthic complexity. The ROV tracks (navigation data like coordinates and depth) were projected to UTM Zone 18N using the Geographic Information System ArcGIS 10.3.1 software.
2.2 Video analysis
During the video analysis with the GOM player program, still images (photogram) were taken every minute (JPEG) (reducing autocorrelation between the same video transect) (Henry et al., 2014; Howell et al., 2014). Blurred or dark video and images out of focus were excluded; if the camera has risen too high from the background to identify the macrohabitat type, the photogram was recorded as undetermined (N/A). Each photogram had an on-screen display (OSD) with ROV heading and depth; this information was entered in a Microsoft Excel spreadsheet file, in which the observation of the macrohabitat or associated macrobiota was included.
Macrohabitats defined as 1 to 10 m in scale, according to Roberts et al. (2009), were classified and modified, in each photogram, taking into account the classification of deep-sea macrohabitats by Southeastern United States Deep-Sea Corals (SEADESC) developed by the National Oceanic and Atmospheric Administration (NOAA) Deep-Sea Coral Program (Partyka et al., 2007), which is based on the classification of deep-sea habitats by Greene et al. (1999) (Cedeño-Posso et al., 2015).
The observed macrobiota occurring during the video analysis were identified and separated into broad groups, according to Collaborative and Annotation Tools for Analysis of Marine Imagery and video (CATAMI), based on a taxonomic and morphological hierarchy (CATAMI Technical Working Group, 2014; Althaus et al., 2015). The identification at the lowest possible taxa was carried out by the group’s taxonomist, from the Museum of Marine Natural History of Colombia-Makuriwa, only if the diagnostic characteristics of the species are visible and distinguishable in the photogram (Hill and Wilkinson, 2004). Some macrobiota were identified as genera or species, based on morphology and consulting with experts (see Acknowledgments), but many could only be identified to a higher taxonomic level such as family, order, or even class. Other cases, such as Echinoderms, were identified based on the samples collected in 1998 and 2002.
2.3 Madracis spp. macrohabitat characterization
To establish the distribution of Madracis spp. complex (Madracis myriaster, Madracis asperula, and Madracis brueggemanni) (Santodomingo et al., 2007) along the 11 sectors surveyed on the CPNNP, during the video analysis, every time Madracis spp. coral colony or patch was visualized in the video transects, time was recorded, and thus, its position was later on established. The use of an underwater acoustic positioning method (USBL) during the recording of the images allowed the positioning of the video transects. This information was recorded in the same initial Microsoft Excel spreadsheet file and incorporated into the ROV video tracks to relate them to geographic position and depth. Substratum type was the only environmental variable determined from the ROV video transects and was classified into four classes: mud/sand, coral-shell rubble, rock/ledges, and sandstone. Terrain variables were derived from the digital elevation model (resolution of 10 m per pixel) to help support the analysis of the characterization of the coral patches.
3 Results
3.1 Classification of benthic macrohabitats
A total of 14 ROV dives were surveyed in 11 sectors; 35.9 h of video was recorded, and 1,215 photograms were analyzed. Nine benthic macrohabitat classifications were established for the area and defined as follows: A) soft substrate (mud/sand), B) soft substrate/rubble/rock with solitary corals, C) coral and shell rubble, D) rock/ledges with attached fauna, E) rugged stones with attached fauna, F) hard substrate with attached fauna, G) sandstone with attached fauna, H) soft substrate with burrows, and I) pockmark with attached fauna) (Figure 2).

Figure 2 Deep-sea macrohabitats from the Corales de Profundidad National Natural Park (Modified from Partyka et al., 2007). (A) Soft substrate (mud/sand). (B) Soft substrate/rubble/rock with solitary corals. (C) Coral and shell rubble. (D) Rock/ledges with attached fauna. (E) Rugged stones with attached fauna. (F) Hard substrate with attached fauna. (G) Sandstone with attached fauna. (H) Soft substrate with burrows. (I) Pockmark with attached fauna.
3.1.1 Soft substrate (mud/sand)
Homogeneous, unconsolidated, and not colonized habitats (Figure 2A), present in eight of the eleven sampling sectors of the CPNNP. Sector 4 (Frijol or Barú Bank, low part on the platform), Sector 2 (elongated hill), Sector 3 (slope), Sector 5 (Angeles channel), Sector 6 (continental shelf), Sector 8 (shelf-break scarp), Sector 9 (shelf-break scarp), and Sector 11 (platform with pockmarks). Five taxa were identified associated with this macrohabitat belonging to three phyla: Echinodermata with ophiuroids, Arthropoda with squat lobsters from the family Munididae, and fishes from the families Serranidae, Lutjanidae, and Carangidae (Supplementary Table S2).
3.1.2 Soft substrate/rubble/rock with solitary corals
Homogeneous habitat, with either rubble or rock colonized by solitary corals (Figure 2B). Present in three sampling sectors: Sector 8 (shelf-break scarp), Sector 9 (shelf-break scarp), and Sector 11 (platform with pockmarks). Sixteen taxa were identified in this macrohabitat from four phyla: Cnidaria with scleractinians like cup corals from the families Cariophylliidae, Dendrophylliidae, and Flabelliidae and Madracis spp complex. Octocorals and antipatharians, one echinoderm belonging to the class Crinoidea, Democrinus cf. conifer, arthropods like crabs, hermit crabs, arrow crabs, and fishes from the families Priacanthidae and Carangidae (Supplementary Table S2).
3.1.3 Coral and shell rubble
Unconsolidated habitat, formed by broken pieces of rock, corals, or shells; can be colonized by bas-relief macrofauna (Figure 2C). Present in two sampling sectors of the CPNNP: Sector 7 (trough and hills) and Sector 8 (shelf-break scarp). Eight taxa were identified in this macrohabitat from three phyla: Cnidaria with various types of octocorals and cup corals, some echinoderms belonging to Crinoidea and Echinoidea classes (Figure 3G), from the orders Comatulida (possibly several species) and Clypeaster euclastus (order Clypeasteroida), and the Carangidae fish Seriola rivoliana (Figure 3L and Supplementary Table S2).
3.1.4 Rock/ledges with attached fauna
Heterogeneous habitat composed of soft terrain interrupted by large rocks or boulders colonized by macrofauna, usually sponges and octocorals (Figure 2D). Present in four sampling sectors of the CPNNP: Sector 1 (Calamarí Bank, on the slope), Sector 2 (summit of the elongated hill), Sector 6 (continental shelf), and Sector 8 (shelf-break scarp). Twenty-seven taxa were identified in this macrohabitat from four phyla: Cnidaria with representatives from five orders Anthoathecata, Actiniaria, Scleractinia, Alcyonacea, and Antipatharia; echinoderms from the classes Crinoidea (order Comatulida), Asteroidea (one unidentified species), Ophiuroidea (orders Ophiacanthida and Ophiurida), and Echinoidea (order Arbacioida); and fishes from the families Holocentridae, Carangidae, Chaetodontidae, Serranidae, Gerreidae, Pomacanthidae, and Scorpaenidae. It is necessary to highlight the presence of the glass sponges, Class Hexactinellida (Figure 2C), at depths between 309.6 and 325 m (Supplementary Table S2).
3.1.5 Rugged stones with attached fauna
Heterogeneous soft substrate or gravel and hard bottom habitat areas colonized by macrofauna, mainly sponges and octocorals (Figure 2E). Present in three sampling sectors of the CPNNP: Sector 8 (shelf-break scarp), Sector 10 (trough), and Sector 11 (platform with pockmarks). Twenty-two taxa from five phyla were identified in this macrohabitat: Cnidaria with representatives of four orders Anthoathecata, Scleractinia, Alcyonacea, and Antipatharia; echinoderms from the class Crinoidea (order Comatulida); sponges from the orders Agelasida, Tetractinellida, Dictyoceratida, and Haplosclerida; and fishes from the families Holocentridae, Carangidae, Chaetodontidae, and Priacanthidae (Supplementary Table S2).
3.1.6 Sandstone with attached fauna
Unconsolidated pavement colonized by macrofauna habitat (such as corals or anemones) (Figure 2F) present in five sampling sectors: Sector 3 (slope), Sector 6 (continental shelf), Sector 8 (shelf-break scarp), Sector 9 (shelf-break scarp), and Sector 10 (trough). Twenty-eight taxa from five phyla were identified in this macrohabitat: Cnidaria with representatives from six orders Spirularia (ceriantarians), Actiniaria (anemones), Alcyonacea, Pennatulacea, Antipatharia, and Scleractinia, with Madracis spp. as the framework builder coral; echinoderms from the classes Crinoidea (order Comatulida), Ophiuroidea (order Euryalida), and Echinoidea (orders Echinothurioida and Clypeasteroida); sponges such as Xestospongia muta and Agelasida specimens; arthropods such as Brachyura crabs; and fishes from the families Chlorophthalmidae, Synodontidae, Caproidae, Carangidae, Serranidae, Sciaenidae, Polymixiidae, and Scorpaenidae (Supplementary Table S2).
3.1.7 Hard substrate with attached fauna
Homogeneous, consolidated, and mainly flat habitat, colonized by calcareous and fleshy algae and other encrusting fauna (Figure 2G). Present in three sampling sectors: Sector 1 (Calamarí Bank, between 65 and 71 m), Sector 4 (Frijol or Barú Bank, between 48 and 51 m), and Sector 7 (trough and hills, between 86 and 112 m). Twenty-five taxa from five phyla were identified along this macrohabitat: Cnidaria with representatives from four orders Anthoathecata, Scleractinia, Alcyonacea, and Antipatharia; echinoderms from Crinoidea class (Nemaster grandis); sponges from the orders Agelasidae, Aplysinidae, and Petrosiidae; and fishes from the families Acanthuridae, Carangidae, Chaetodontidae, Pomacanthidae, Sphyraenidae, Pomacentridae, Scorpaenidae, Monacanthidae, and Balistidae. It is the only macrohabitat with the presence of Halimeda copiosa and Lobophora variegata macroalgae, located at the mesophotic zones between 48.5 and 51.9 m (Supplementary Table S2).
3.1.8 Soft substrate with burrows
Unconsolidated habitat with bottom depressions easily identifiable, some of which could be produced by a variety of fishes; sometimes, it was possible to observe fish inhabiting them (Figure 2H). Present in two sampling sectors: Sector 5 (Angeles channel) and Sector 11 (platform with pockmarks). In this macrohabitat, the only organism possible to be identified is the fish S. rivoliana (Supplementary Table S2).
3.1.9 Pockmark with attached fauna
Loose sediment habitat with one or more sinks colonized by macrofauna (sponges and corals) (Figure 2I). Present in two sampling sectors of the CPNNP: Sector 8 (shelf-break scarp) and Sector 9 (shelf-break scarp). Only two phyla were identified in this macrohabitat: octocorals (Cnidaria) and echinoderms from the Crinoidea class (order Comatulida) (Supplementary Table S2).
The biotic assemblage of the 11 sectors of the Corales de Profundidad National Natural Park, comprised 88 taxa, including algae, sponges, hydrozoans, anthozoans, echinoderms, and fishes (Supplementary Table S2). Corales de Profundidad NNP brings together mesophotic environments ranging from 30 to 150 m depth, and aphotic from 150 to the abyssal zone (Rocha et al., 2018; Denis et al., 2019). No large deep-sea coral formation was observed other than hard and soft substrates covered with macrofauna. The macrohabitats with the highest richness are the rocky ones (rock/ledges with attached fauna, rugged stones with attached fauna, and hard substrate with attached fauna).
3.2 Madracis spp. coral patch distribution
Linear extension of the video transects mapping effort totaled 32.4 km measured in a GIS of which 67% was mud/sand, 15% rock/ledges, 15% sandstone, and 4% coral-shell rubble. The video transects analysis confirms the occurrence of Madracis spp. as the framework builder of the CPNNP. These patches (observed on 101 photograms of 1,215) are associated with the macrohabitat sandstone with attached fauna (Figure 4).
Combining bathymetry, the digital elevation model derivatives, and the geomorphological unit’s map (Morales et al., 2017) (Figure 1) with ground-truthing from video transects has proven to be particularly efficient in showing the distribution of the Madracis spp. coral patches. Terrain variables allowed the association of the patches with three morphological units: shelf-break scarp (112 and 232 m depth), continental shelf (107 and 169 m), and channel/canyon between 197 and 233 m depth (Figure 1C). Occurrences could not be related to a particular steepness of the terrain (ranging from 0° to 23°). However, 44% of the occurrences were associated with a low slope, while 31% and 21% of the points belong to a medium and a flat terrain. Only 2% were observed on a high slope (Figure 1D). The spatial scale of the estimation of rugosity failed to aid the assessment of the topographic structural complexity of the area (Figure 1B).
Madracis spp. coral patches were identified in five sectors: in Sector 10, small and scattered colonies were recorded over a mud/sand located on the north escarp of the Frijol or Barú Bank at 169 m depth. In Sector 4, 20 cm high colonies were observed between 163 and 167 m depth, scattered on mud/sand substrate. In Sector 6, the patches were recorded at depths between 197 and 233 m, on mud/sand substrate (Supplementary Figures S3–S5). In Sectors 8A, extensive and dense Madracis spp. patches of approximately 50–80 cm high were recorded on mud/sand substrate between 118 and 197 m depth (Figure 4), and in Sectors 8B and 9A, small and scattered colonies were recorded on the soft bottom 107 and 230 m depths (Figure 4).
Although several attempts were made to generate a suitable habitat mapping for the coral patches, the environmental data available for the area lacked the spatial resolution needed to have an accurate model map.
4 Discussion
The mesophotic zones, of the Corales de Profundidad National Natural Park, range from 46 up to 169 m depth; aphotic ranges from 108 to 354 m depth (the deepest ROV video transect). The overlap in depths between these zones and the changes in their community structure is driven by the light penetration conditions in each sector, as suggested by Tamir et al. (2019), and by environmental and ecological drivers that vary from sector to sector (Castellan et al., 2022).
The mesophotic zones distribute between the summits of two banks and two hills of diapir origin (Morales et al., 2017) on Sectors 1, 4, and 7; these elevations are characterized by hard substrate and rocky macrohabitats (Figures 2D, E, G). This characteristic provides an adequate environment for rhodolites (calcareous red algae) and gravel with high algae cover, such as L. variegata and H. copiosa (Figures 3A, B). It also supports the settlement of sessile macrofauna (plate corals, octocorals, and sponges), like large barrel sponges (X. muta), and erect sponges (Aplysina cauliformis). The other mesophotic sector, Sector 11, is a zone of the less consolidated bottom, with the presence of some large rocks, which also serve as an aggregation structure for organisms. The fact that more varieties of fish and invertebrates are observed in the mesophotic sectors is explained by the fact that in areas of hard substrate where there is no presence of massive reefs, the microhabitats that form between the small rocks and rhodoliths serve as a refuge for lower organisms; the ichthyofauna is reduced to small demersal fish and a reduced range of demersal predators such as crossbows and barracudas (García-Sais et al., 2011).

Figure 3 Example of the fauna found in the Corales de Profundidad National Natural Park (CPNNP). (A) Agaricia sp.; Lobophora variegata and Stegastes partitus. (B) Halimeda copiosa. (C) Glass sponges and Comatulida spp. 2. (D) Ceriantheopsis sp. (E, F) Liponema spp. (G) Araeosoma cf. fenestratum. (H) Comatulida spp. 2 and Ophiacanthida sp. (I) Nemaster grandis. (J) Percophidae, Bembrops sp. (K) Epinephelus sp. (L) Seriola rivoliana.
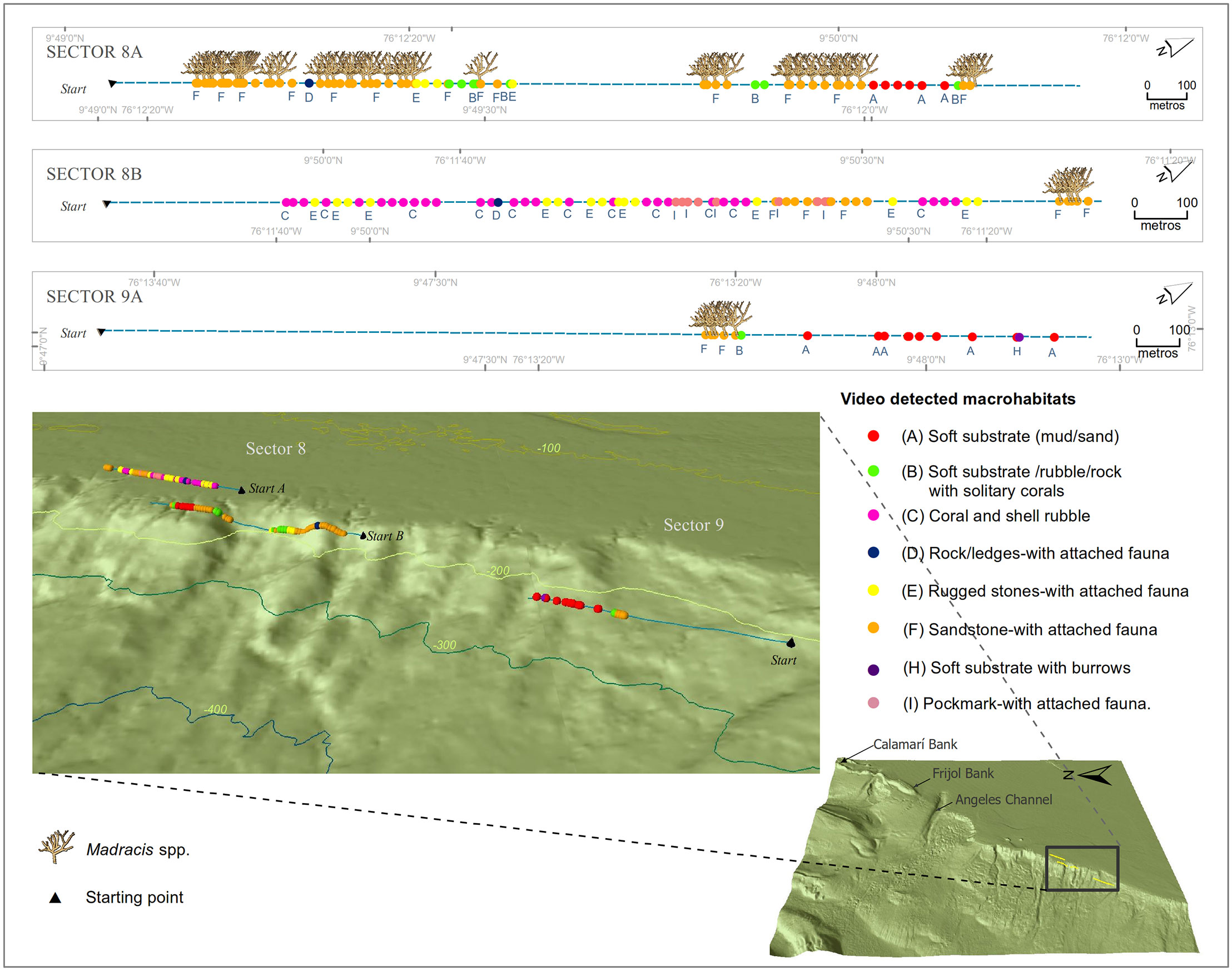
Figure 4 Distribution of the macrohabitats identified in Sectors 8 and 9 of the Corales de Profundidad National Natural Park; area superimposed on the 3D bathymetry map. The occurrence of Madracis spp. coral patches is also shown.
The aphotic zones correspond to one hill, one channel, and an escarpment and basin (Morales et al., 2017). These last two occupy a large section of the sampled platform where the soft substrate macrohabitats (Figures 2A, B, F, H) are constant. Solitary fauna inhabits these macrohabitats dominated by crinoids (sea lilies) and cnidarians (cup corals, sea fans, whip corals, black corals, anemones, tube anemones, and sea feathers). Hard coral Madracis spp. complex is the most representative. Sponges are among the most critical structural elements of the CPNNP, providing a refuge habitat for mobile fish and invertebrates, very like to what occurs on the continental shelf of Puerto Rico (Lutz and Ginsburg, 2007). The fish fauna in soft substrates is composed of lizardfishes (Synodontidae), squirrel fishes (Holocentridae), and butterflyfishes (Chaetodontidae), among other demersal dwellers. It is relevant to remark on the observation of some individuals of deeper groups as the polymixides and caproids being some of the groups with representative species found as more abundant in the community structure of the deep-sea fishes in the Central Caribbean (Benavides-Morera and Campos-Calderón, 2019) and duckbills (Percophidae) (Figure 3J) with some quick appearances of active swimmers predators in front of the camera like jacks and groupers (Figures 3K, L).
Deep-sea macrohabitats (mesophotic or aphotic) with hard substrata components hold the highest richness of fauna observed; species richness is related to the heterogeneity of the seabed that offers shelter and resources obtained from the community of sessile organisms with different attachment mechanisms (Vertino et al., 2010). For example, cold-water corals, gorgonians (excluding Isididae), black corals, and sponges need hard substrates to be attached and are sensitive to sedimentation. Bamboo corals have branched root-like structures to anchor themselves in muddy bottoms. Pennatulids use a basal bulb that swells when buried in the sediment (De la Torriente et al., 2018). Echinoderms such as Comatulida spp. include both types of species, inhabiting large rocks and located near the bottom (Figures 3C, H, I).
The presence of demersal predators such as jacks, snappers, barracudas, and groupers are characteristic of these areas. Other groups associated with rocky areas are triggerfishes (Balistidae), damselfishes (Pomacentridae), angelfishes (Pomacanthidae), butterflyfishes (Chaetodontidae), filefishes (Monacanthidae), surgeonfishes (Acanthuridae), and squirrel fishes (Holocentridae). Even at depths reaching 150 m, the presence of the invasive lionfish (Pterois volitans) associated with sponges or rock ledges was common (Henao-Castro et al., 2016; Chasqui-Velasco and González, 2019; Sanjuan-Muñoz et al., 2022). Fish’s distribution pattern is typical of mesophotic habitats with the presence of large predators targeted by fishermen and different groups of reef-associated fishes, as previously described for Puerto Rico (García-Sais et al., 2012; Bejarano et al., 2014).
The dominance of Madracis spp. complex (M. myriaster, M. asperula, and M. brueggemanni) on sandy mud bottoms over the shelf break of CPNNP is of particular importance since it has not been identified as a dominant structuring species in other regions of the world (Reyes et al., 2005; Lutz and Ginsburg, 2007; Santodomingo et al., 2007). In the Caribbean, sightings of Madracis cf. myriaster have been made on a rocky substrate, with ROV and manned submersibles, between 200 and 366 m, at Bermuda (Stefanoudis et al., 2019), Pichincho Wall East at Mona Passage, East Vieques Island (Wagner et al., 2019), and Dog and Conrad Seamounts at Anegada Passage (Auscavitch et al., 2020). Thus, Madracis spp. coral patches continue to give the area absolute uniqueness and increased value of conservation priority (Alonso et al., 2015; Alonso et al., 2021).
Although macrobenthic species footage recognition from ROV video transects may not be the most appropriate method for identification, it is one of the methods that can provide us with direct observations of a deep-sea habitat where a diver cannot reach (Parry et al., 2002). Many small-sized organisms tend to be underestimated, and only those with sessile characteristics are taken into account. Examples of the difficulty of identification are Cnidarian and Echinodermata phyla. Cnidarian identification at the species level was not possible due to the video resolution of the camera mounted on the ROV (Diavolo II). Further identification of the specimens will need still images that allow a close-up of the polyps. Echinoderms’ further identification of taxonomic levels was only possible due to the previous knowledge of several species with clear diagnostic characteristics collected in the same area between 1998 and 2002 (Reyes et al., 2005; Benavides-Serrato et al., 2011; Borrero-Perez et al., 2012); for this particular case, the images provided details of morphology that had been destroyed by the collection method of previous years.
Analysis of videos and photograms becomes a bottleneck in the habitat classification process due to a large amount of acquired material needing to be reviewed. Although automatic or semi-automatic software can be used for the detection and classification of fauna and substrate (Lutz and Ginsburg, 2007; Schoening et al., 2012; Gomes-Pereira et al., 2016; Romero-Ramirez et al., 2016), the software needs initial training with a vast inventory of facies, structures, and species that are still unknown for the area. Habitat suitability at scales relevant for mapping requires detailed environmental data that may guide the natural expression of the elements to be mapped. There is still a large information gap in the data needed for habitat mapping. Cost-effective methods for the acquisition of oceanographic and physicochemical variables will better establish the divisions and extend among observed macrohabitats. Even so, sampling restricted to megafauna, large species, epi-benthic, and without evasive behavior allows sampling the seabed at a scale of centimeters to kilometers, making it possible to have interpolation of the data and an estimation of abundance and distribution on inaccessible areas (Zawada et al., 2008; Romero-Ramirez et al., 2016).
All fauna identified in this work were previously recorded in earlier studies (Reyes et al., 2005), except for the Cnidarian (ceriatharian) Ceriantheopsis sp. (Figure 3D), found at 225.5 m in sandstone with attached fauna macrohabitat. Moreover, two morphotypes of an anemone from the family Liponematidae (Figures 3E, F) are the first records for the Colombian Caribbean, a rose-colored recorded between 188.9 and 197.3 m found in sandstone with attached fauna macrohabitat and a red one between 310.1 and 332.4 m found in rugged stones with attached fauna macrohabitat. All echinoderms classes were recorded in the CPNNP except for Holothuroidea, due to the sampling method, although previous studies only registered the species Holothuria (Cystipus) occidentalis in this area (Reyes et al., 2005).
Our study contributes to refine the scale of identification of deep water macrohabitats in the Colombian Caribbean specifically for the Corales de Profundidad NNP. The distinctive macrohabitats: hard substrate with attached fauna and the rock/ledges with attached fauna are distributed on three big geoforms of the CPNNP: Hills, Angeles channel, Calamari Bank, Frijol or Barú Bank; the last one is a mesophotic zone studied in the last few years using rebreathers (Chasqui-Velasco and González, 2019; Sánchez et al., 2019), but the others are geoforms that have not yet been described in detail. It must begin with the study of these areas that may provide suitable environmental conditions (food availability and bottom currents) for deep-sea corals to grow. Future studies might discover new deep-sea macrohabitats. Thanks to these results, three of the macrohabitats have been analyzed in more detail and are now represented in the Atlas of the Coral Areas of Colombia (INVEMAR-MINAMBIENTE, 2020), as the following microhabitats or biotic units: the macrohabitat sandstone with attached fauna, where patches of coral Madracis spp. complex are present, are now known as the biotic unit Madracis spp. (Mad); the macrohabitat soft substrate/rubble/rock with solitary corals that has a higher cover of cup corals is now known as the biotic unit cup corals (CorCop); and finally, the macrohabitat hard substrate with attached fauna that has a higher cover of rhodoliths is now known as the biotic unit Rhodoliths (Rod).
Efforts should concentrate on implementing a research strategy to address the management of this marine protected area, given the limited knowledge and understanding of the ecological and physical processes. Research topics should use higher-resolution ROV video surveys with in situ collections or the use of emerging tools like environmental DNA (eDNA), address the distribution of corals, and other bio-builders, and understand the physical and geological processes that control their presence, community structure, patterns, and abundance.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, Further inquiries can be directed to the corresponding author.
Author contributions
CC-P did the video analysis of the expedition. CC-P, AP, and GB identified the macrofauna of the videos. CC-P, AP, and GB wrote the biological component of the article. MV-C and VR contributed to spatial analysis. DA, CC-P, MV-C, and VR contributed to the project research and conceived the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported and funded by INVEMAR through the projects PNN Corales de Profundidad (Convenio 291-14 ANH-INVEMAR) co-funded by Parques Nacionales Naturales, Manchester University and Agencia Nacional de Hidrocarburos.
Acknowledgments
Thanks to the Institute of Marine and Coastal Research-INVEMAR for co-financing Agencia Nacional de Hidrocarburos Anadarko (ANH), Parques Nacionales Naturales, Manchester University. We thank SERPORT and the crew of the R/V Los Angeles and MARISCOPE and Diavolo II ROV pilot, Juan Manuel Casal, during the surveys in 2015. Additional taxonomic experts who contributed to the further identification of some organisms are as follows: Sven Zea, Nadiezhda Santodomingo, and Sergio Stampar. Thanks to the Taxonomy Research Group of INVEMAR for their collaboration and good friendship. Contribution No. 1347 of the Marine and Coastal Research Institute – INVEMAR.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.995419/full#supplementary-material
References
Alonso D., Vides-Casado M., Arias-Isaza F., Zambrano H., Rodriguez E., Rocha-Gutierrez V., et al. (2021). Behind the scenes for the designation of the corales de profundidad national natural park of Colombia. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.567438
Alonso D., Vides M., Cedeño-Posso C., Marrugo M., Henao A., Sanchez J. A., et al. (2015). Parque nacional natural corales de profundidad: descripción de comunidades coralinas y fauna asociada (Santa Marta: Serie de Publicaciones Generales del Invemar No. 88), 20p.
Althaus F., Hill N., Ferrari R., Edwards L., Przeslawski R., Schönberg C. H. L., et al. (2015). A standardised vocabulary for identifying benthic biota and substrata from underwater imagery: The CATAMI classification scheme. PloS One 10 (10), e0141039. doi: 10.1371/journal.pone.0141039
Auscavitch S. R., Lunden J. J., Barkman A., Quattrini A. M., Demopoulos A. W. J., Cordes E. E. (2020). Distribution of deep-water scleractinian and stylasterid corals across abiotic environmental gradients on three seamounts in the anegada passage. Peer J. 8, e9523. doi: 10.7717/peerj.9523
Bejarano I., Appeldoorn R. S., Nemeth M. (2014). Fishes associated with mesophotic coral ecosystems in la parguera. Puerto Rico Coral Reefs 33 (2), 313–328. doi: 10.1007/s00338-014-1125-6
Benavides-Morera R., Campos-Calderón F. (2019). Abundancia, biomasa y estructura de la ictiofauna demersal del mar caribe de américa central. Uniciencia 33 (1), 1–17. doi: 10.15359/ru.33-1.1
Benavides-Serrato M., Borrero-Pérez G. H., Díaz-Sánchez C. M. (2011). Equinodermos del caribe colombiano i. crinoidea, asteroidea y ophiuroidea (Santa Marta, Colombia: Serie de Publicaciones Especiales, INVEMAR No. 22), 384 p.
Borrero-Pérez G. H., Benavides-Serrato M., Diaz-Sanchez y C. M. (2012) Equinodermos del Caribe colombiano II: Echinoidea y Holothuroidea. Serie de Publicaciones Especiales de Invemar No. 30. (Santa Marta, Colombia), 250.
Cairns S. D. (1979). The deep-water scleractinia of the Caribbean Sea and adjacent waters. studies on the fauna of curaçao and other Caribbean islands, Vol. 57. 341 pp.
Castellan G., Angeletti L., Montagna P., Taviani. M. (2022). Drawing the borders of the mesophotic zone of the Mediterranean Sea using satellite data. Sci. Rep. 12, 5585. doi: 10.1038/s41598-022-09413-4
CATAMI Technical Working Group (2014). CATAMI classification scheme for scoring marine biota and substrata in underwater imagery. version 1.4 (Canberra, Australia: National Environmental Research Program, Marine Biodiversity Hub), 24pp. (Technical Report). doi: 10.25607/OBP-11
Cedeño-Posso C., Alonso D., Vides M., Chasqui L., Aguirre J., Ballesteros D., et al. (2015) Caracterización de las comunidades coralinas del parque nacional natural corales de profundidad en el caribe colombiano: una aproximación a la conservación de su biodiversidad. informe técnico final. INVEMAR, PNN, u. Manchester y ANH. convenio interinstitucional 291-14 entre el instituto de investigaciones marinas y costeras “José benito vives de andréis”– INVEMAR y la agencia nacional de hidrocarburos. Available at: https://n2t.net/ark:/81239/m9rh3g (Accessed January 15, 2020).
Chasqui-Velasco L., González J. (2019). Fishes found in mesophotic environments of bajo frijol, the shallowest portion of the corales de profundidad natural national park, using CCR technical diving. Bol. Invest. Mar. Cost. 48, 89–101. doi: 10.25268/bimc.invemar.2019.48.1.759
Cosel R., Gracia A. (2018). A new mussel species, Gigantidas samario (Bivalvia: Mytilidae: Bathymodiolinae), from hydrocarbon seep communities off the Caribbean coast of Colombia. Archiv. für Molluskenkunde 147 (2), 173–179. doi: 10.1127/arch.moll/147/173-179
De la Torriente A., Serrano A., Fernández-Salas L. M., García M., Aguilar R. (2018). Identifying epibenthic habitats on the seco de los olivos seamount: Species assemblages and environmental characteristics. Deep Sea Res. Part I. 135, 9–22. doi: 10.1016/j.dsr.2018.03.015
Denis V., Soto D., De Palmas S., Vicky Lin Y. T., Benayahu Y., Huang Y. M., et al. (2019). “Taiwan,” in Mesophotic coral ecosystems. Eds. Loya Y., Puglise K. A., Bridge T. C. L. (New York: Springer) 249–264
Digby A., Puentes V., León J. (2016). Cold seeps associated with structured benthic communities: More accurate identification and evaluation using a new multibeam survey methodology in the offshore southern Colombian Caribbean. Int. J. Geosci. 07, 761–774. doi: 10.4236/ijg.2016.75058
Dueñas A., Puentes V., León J., Herrera S. (2021). Fauna associated with cold seeps in the deep Colombian Caribbean. Deep Sea Res. Part I: Oceanogr. Res. Papers 173, 103552. doi: 10.1016/j.dsr.2021.103552
Flórez-Romero P., Montoya-Cadavid E., Reyes-Forero J., Santodomingo N. (2007). Briozoos cheilostomados del caribe colombiano. Bol. Invest. Mar. Cost. 36, 229–250. doi: 10.25268/bimc.invemar.2007.36.0.208
Flórez P., Santodomingo N. (2010). “Cnidaria: Corales escleractineos, antipatarios, anémonas, zoantideos, octocorales e hidroides. 151-178 pp,” in Biodiversidad del margen continental del caribe colombiano (Santa Marta, Colombia: Serie de publicaciones especiales, Invemar No. 20) (Santa Marta, Colombia), 458p.
Freiwald A., Fosså J., Grehan A., Koslow T., Roberts J. M. (2004) Cold-water coral reefs: Out of sight – no longer out of mind (Cambridge, UK: UNEP-WCMC). Available at: http://www.unep-wcmc.org/resources/publications/UNEP_WCMC_bio_series/22.htm (Accessed March 5, 2019).
García-Sais J., Sabater-Clavell J., Esteves R., Capella J., Carlo M. (2011). Characterization of benthic habitats and associated mesophotic coral reef communities at El seco, southeast vieques, Puerto Rico. 102 p. doi: 10.13140/RG.2.1.2752.1361
García-Sais J. R., Sabater-Clavell J., Esteves R., Carlo M. (2012). Fishery-independent survey of commercially exploited fish and shellfish populations from mesophotic reefs within the Puerto Rican exclusive economic zone (San Juan, Puerto Rico: Caribbean Fisheries Management Council).
Gomes-Pereira J. N., Auger V., Beisiegel K., Benjamin R., Bergmann M., Bowden D., et al. (2016). Current and future trends in marine image annotation software. Prog. Oceanogr. 149, 106–120. doi: 10.1016/j.pocean.2016.07.005
Gracia A., Ardila N. (2010). “Moluscos de la plataforma y talud superior del caribe colombiano (20-940 m):12 años de investigación. 181-227pp,” in Biodiversidad del margen continental del caribe colombiano. Ed. INVEMAR (Santa Marta, Colombia: Serie de publicaciones especiales, Invemar No. 20), 458p.
Gracia A., Rangel-Buitrago N., Sellanes J. (2011). Methane seep molluscs from the sinu´ – San jacinto fold belt in the Caribbean Sea of Colombia. J. Mar. Biol. Assoc. United Kingdom, 92 (6), 1367–1377. doi: 10.1017/S0025315411001421.
Greene H., Yoklavich M., Starr R., O’Connell V., Wakefield W., Sullivan D., et al. (1999). A classification scheme for deep seafloor habitats. Oceanologia Acta, 22, 663–678
Henao-Castro A., Marrugo M., Molina M. P. (2016). Invasión del pez león (Pterois volitans) en ecosistemas profundos del caribe colombiano: Parque nacional natural corales de profundidad. La Timonera 25, 61–63.
Henry L., Vad J., Findlay H., Murillo J., Milligan R., Roberts J. M. (2014). Environmental variability and biodiversity of megabenthos on the Hebrides terrace seamount (Northeast Atlantic). Sci. Rep. 4, 5589. doi: 10.1038/srep05589
Hill J., Wilkinson C. (2004). Methods for ecological monitoring of coral reefs. version 1 (Australia: Australian Institute of Marine Science), 117 p.
Howell K. L., Huvenne V., Piechaud N., Robert K., Ross R. E. (2014). Analysis of biological data from the JC060 survey of areas of conservation interest in deep waters off north and west Scotland (Peterborough: JNCC).
INVEMAR-MINAMBIENTE (2020) Atlas de Áreas coralinas de Colombia. Available at: https://areas-coralinas-de-colombia-invemar.hub.arcgis.com.
Levin L. A., Bett B. J., Gates A. R., Heimbach P., Howe B. M., Janssen F., et al. (2019). Global observing needs in the deep ocean. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00241
Lutz S. J., Ginsburg R. N. (2007). State of deep coral ecosystems in the Caribbean region: Puerto Rico and the U.S. Virgin Islands, in: Lumsden, S.E. et al. (Ed.) The state of deep coral ecosystems of the United States. NOAA Technical Memorandum, CRCP-3 , 307–365 p.
Morales D., Rocha V., Posada B. (2017). Geomorfología de los fondos submarinos del parque nacional natural corales de profundidad, mar caribe colombiano. Bull. Mar. Coast. Res. 46 (2), 73–90. doi: 10.25268/bimc.invemar.2017.46.2.727
Parry D., Nickell L., Kendall M., Burrows M., Pilgrim D., Jones M. (2002). Comparison of abundance and spatial distribution of burrowing megafauna from diver and remotely operated vehicle observations. Mar. Ecol. Prog. Ser. 244, 89–93. doi: 10.3354/meps244089
Partyka M., Ross S., Quattrini A., Sedberry G., Birdsong T., Potter J., et al. (2007). Southeastern united states deep-Sea corals (SEADESC) initiative: A collaborative effort to characterize areas of habitat-forming deep-Sea corals (Silver Spring, MD: NOAA Technical Memorandum OAR OER 1).
Ramirez-Llodra E. (2020). “Chapter 2 deep-Sea ecosystems: Biodiversity and anthropogenic impacts,” in The law of the seabed (Leiden, The Netherlands: Brill | Nijhoff). doi: 10.1163/9789004391567_004
Reyes J., Santodomingo N., Cairns S. (2009). Caryophylliidae (Scleractinia) from the Colombian Caribbean. Zootaxa 2262, 1–39. doi: 10.11646/zootaxa.2262.1.1
Reyes J., Santodomingo N., Gracia A., Borrero-Pérez G., Navas G., Mejía-Ladino L. M., et al. (2005). “Southern Caribbean azooxanthellate coral communities off colombia. p. 309-330,” in Cold-water corals and ecosystems. Eds. Freiwald A., Roberts J. M. (Berlin Heidelberg: Springer-Verlag), 1243 p.
Roberts J.M., Wheeler A., Freiwald A., Cairns S. (2009). Cold-water corals: The biology and geology of deep-sea coral habitats (UK: Cambridge University Press), 334 p.
Rocha L. A., Pinheiro H. T., Shepherd B., Papastamatiou Y. P., Luiz O. J., Pyle R. L., et al. (2018). Mesophotic coral ecosystems are threatened and ecologically distinct from shallow water reefs. Science 361 (6399), 281–284. doi: 10.1126/science.aaq1614
Romero-Ramirez A., Grémare A., Bernard G., Pascal L., Maire O., Duchêne J. (2016). Development and validation of a video analysis software for marine benthic applications. J. Mar. System. 162, 4–17. doi: 10.1016/j.jmarsys.2016.03.003
Sánchez J. A., Dueñas L. F., Rowley S. J., González F. L., Vergara D. C., Montaño-Salazar S. M., et al. (2019). “Gorgonian corals (39),” in Mesophotic coral ecosystems coral reefs of the world. Eds. Loya Y., Puglise K. A., Bridge T. C. L. (Cham: Springer Nature Switzerland AG), 727–745. doi: 10.1007/97
Sanjuan-Muñoz A., Bustos-Montes D., Polo-Silva C., Henao-Castro A., Marrugo M., Delgado-Huertas A., et al. (2022). Biología y ecología del pez león (Pterois volitans) en el parque nacional natural corales de profundidad, caribe colombiano. Bol. Invest. Mar. Cost. 51 (1), 75–98. doi: 10.25268/bimc.invemar.2022.51.1.1087
Santodomingo N., Reyes J., Flórez P., Chacón-Gómez I. C., van Ofwegen L. P., Hoeksema B. W. (2013). Diversity and distribution of azooxanthellate corals in the Colombian Caribbean. Mar. Biodiversity 43, 7–22. doi: 10.1007/s12526-012-0131-6
Santodomingo N., Reyes J., Gracia A., Martínez A., Ojeda G., García C. (2007). Azooxanthellate Madracis coral communities off San Bernardo and Rosario islands (Colombian Caribbean). Bull. Mar. Sci. 81 (1), 273–287.
Schoening T., Bergmann M., Ontrup J., Taylor J., Dannheim J., Gutt J., et al. (2012). Semi-automated image analysis for the assessment of megafaunal densities at the Arctic deep-Sea observatory HAUSGARTEN. PloS One 7 (6), e38179. doi: 10.1371/journal.pone.0038179
Stefanoudis P. V., Rivers M., Smith S. R., Schneider C. W., Wagner D., Ford H., et al. (2019). Low connectivity between shallow, mesophotic and rariphotic zone benthos. R. Soc. Open Sci. 6 (9), 1–13. doi: 10.1098/rsos.190958
Tamir R., Eyal G., Kramer N., Laverick J. H., Loya Y. (2019). Light environment drives the shallow-to-mesophotic coral community transition. Ecosphere; 10, e02839. doi: 10.1002/ecs2.2839
Vertino A., Savini A., Rosso A., Di Geronimo I., Mastrototaro F., Sanfilippo R., et al. (2010). Benthic habitat characterization and distribution from two representative sites of the deep-water SML coral province (Mediterranean). Deep Sea Res. Part II: Topical Stud. Oceanogr. 57 (5–6), 380–396. doi: 10.1016/j.dsr2.2009.08.023
Vinnels J. S., Butler R. W. H., McCaffrey W. D., Paton D. A. (2010). Depositional processes across the sinú accretionary prism, offshore Colombia. Mar. Petroleum Geol. 27 (4), 794–809. doi: 10.1016/j.marpetgeo.2009.12.008
Wagner D., Sowers D., Williams S. M., Auscavitch S., Blaney D., Cromwell M. (2019). Océano profundo 2018: exploring deep sea habitats off Puerto Rico and the US virgin islands (Silver Spring: Office of Ocean Exploration and Research, NOAA), 1–171.
Keywords: macrohabitat, deep-sea coral, national natural park, megafauna, new records
Citation: Cedeño-Posso C, Vides-Casado M, Rocha V, Borrero-Pérez GH, Polanco F A and Alonso D (2022) Benthic macrohabitat classification and Madracis spp. coral patch distribution in a deep-sea marine protected area of Colombia. Front. Mar. Sci. 9:995419. doi: 10.3389/fmars.2022.995419
Received: 15 July 2022; Accepted: 14 October 2022;
Published: 15 November 2022.
Edited by:
Pedro A. Ribeiro, University of Bergen, NorwayReviewed by:
Ivan Hernández-Ávila, Universidad Autónoma del Carmen, MexicoDiogo Antonio Tschoeke, Federal University of Rio de Janeiro, Brazil
Copyright © 2022 Cedeño-Posso, Vides-Casado, Rocha, Borrero-Pérez, Polanco F and Alonso. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cristina Cedeño-Posso, Y3Jpc3RpbmEuY2VkZW5vQGludmVtYXIub3JnLmNv