 Paula Suarez-Bregua1*
Paula Suarez-Bregua1* Miguel Álvarez-González2
Miguel Álvarez-González2 Kim M. Parsons3
Kim M. Parsons3 Josep Rotllant1
Josep Rotllant1 Graham J. Pierce1
Graham J. Pierce1 Camilo Saavedra2
Camilo Saavedra2- 1Institute of Marine Research, Spanish National Research Council (IIM-CSIC), Vigo, Spain
- 2Centro Oceanográfico de Vigo, Instituto Español de Oceanografía (IEO-CSIC), Vigo, Spain
- 3Northwest Fisheries Science Center, National Marine Fisheries Service (NOAA), Seattle, WA, United States
Monitoring marine mammal populations is essential to permit assessment of population status as required by both national and international legislation. Traditional monitoring methods often rely on visual and/or acoustic detections from vessels and aircraft, but limitations including cost, errors in the detection of some species and dependence on taxonomic expertise, as well as good weather and visibility conditions often limit the temporal and spatial scale of effective, long-term monitoring programs. In recent years, environmental DNA (eDNA) has emerged as a revolutionary tool for cost-effective, sensitive, non-invasive species monitoring in both terrestrial and aquatic realms. eDNA is a rapidly developing field and a growing number of studies have successfully implemented this approach for the detection and identification of marine mammals. Here, we review 21 studies published between 2012 and 2021 that employed eDNA for marine mammal monitoring including single species detection, biodiversity assessment and genetic characterization. eDNA has successfully been used to infer species presence (especially useful for rare, elusive or threatened species) and to characterize the population genetic structure, although additional research is needed to support the interpretation of non-detections. Finally, we discuss the challenges and the opportunities that eDNA could bring to marine mammal monitoring as a complementary tool to support visual and acoustic methods.
Introduction
Marine mammals play a crucial ecological role in the world’s oceans and seas. They influence the structure and function of marine ecosystems through predation, as prey to other animals, and by promoting nutrient transfer and distribution through the release of fecal plumes, urine and carcasses (Roman and Estes, 2018). Most marine mammals occupy high trophic levels (i.e., they are top predators and secondary consumers) and play an important role in the stability of the food web through “top-down” effects (Estes et al., 2011). However, marine mammal populations are currently threatened by a multitude of anthropogenic stressors, including fisheries bycatch (Reeves et al., 2013), ship strikes (Schoeman et al., 2020), prey depletion caused by overfishing (DeMaster et al., 2001), climate change (Albouy et al., 2020), habitat degradation (Todd et al., 2014) and chemical and acoustic pollution (Desforges et al., 2016; Nabi et al., 2018). Nearly 40% of marine mammal species have been listed as threatened (i.e., as vulnerable, endangered and critically endangered species) by the International Union for Conservation of Nature’s (IUCN) Red List (Schipper et al., 2008). To protect marine mammals and mitigate the effect of human activities, a number of laws and directives have been implemented, including the EU Marine Strategy Framework Directive (MSFD), EU Habitats Directive, US Marine Mammal Protection Act (MMPA), US Endangered Species Act (ESA), as well as numerous international agreements/organizations (CITES, ASCOBANS, ACCOBAMS, OSPAR, and the International Whaling Commission (IWC) among others), all of which require the monitoring, assessment and management of marine mammal populations.
Monitoring programs are needed to assess both population status and anthropogenic threats to marine mammal populations. Distribution and abundance, essential parameters for population monitoring, have traditionally been characterized using visual methods including line transect distance sampling (Zerbini et al., 2007; Dick and Hines, 2011; Hammond et al., 2013) and photo-identification (Urian et al., 2015). Visual monitoring relies on dedicated based-boat and aerial surveys that are costly and time-consuming, as well as challenging for widely dispersed species with prolonged periods of sub-surface activities (Booth et al., 2020). Additionally, visual methods require experienced observers, and are restricted to both daylight hours and good weather conditions to minimize detection bias due to environmental effects (Barlow et al., 2006). These factors often limit the collection of reliable data over long time periods, whereas long-term biomonitoring is needed for effective population assessment and conservation. Because of these limitations, recent years have seen increasing interest in novel data collection methods to support effective marine mammal monitoring.
The application of environmental DNA (eDNA) approaches to marine systems has emerged as a promising tool for both single species detection and biodiversity monitoring (Thomsen and Willerslev, 2015). The term eDNA refers to intracellular and extracellular DNA that can be extracted from environmental samples (e.g. water, soil or air) (Taberlet et al., 2012). All muticellular organisms naturally shed cellular material from skin, feces, urine and/or gametes into their surroundings, thus leaving a molecular signature in the environment they inhabit (Barnes et al., 2014). For example, the blow exhaled by cetaceans when they surface to breath is composed of a mixture of cells, mucus and fluids from the respiratory tract that are deposited on the water surface. The “footprint” that cetaceans leave behind in surface waters when breathing and diving includes genetic material from skin and potentially also mucus and/or feces (Figure 1). eDNA analysis offers a potentially more accessible, cost-effective, non-invasive, sensitive approach to monitoring, with less reliance on field-based taxonomic expertise compared to traditional visual survey methods (Port et al., 2016; Pikitch, 2018). In aquatic environments, eDNA can be collected by filtration, precipitation or centrifugation of water samples (Ficetola et al., 2008; Tsuji et al., 2019) and then amplified using assays targeting specific DNA sequences (Figure 1). Thus, eDNA can be used to detect the presence of single species via PCR amplification using species-specific primers (Ficetola et al., 2008; Foote et al., 2012; Baker et al., 2018; Parsons et al., 2018) while multiple species can be identified simultaneously through eDNA metabarcoding using universal primers coupled with next-generation sequencing (NGS) (Thomsen et al., 2012; Miya et al., 2015; Valsecchi et al., 2020; Xie et al., 2021). In addition, multiple primer pairs targeting different species or taxa can be combined in a single PCR reaction (i.e., multiplex PCR assay) (Ramón-Laca et al., 2021). Primers are often designed to amplify short fragments (<300bp) from mitochondrial DNA (mtDNA) genes, such as the control region (D-loop) (Baker et al., 2018; Parsons et al., 2018; Pinfield et al., 2019), 12s rRNA gene (12s) (Riaz et al., 2011; Foote et al., 2012; Miya et al., 2015; Valentini et al., 2016; Taberlet et al., 2018; Valsecchi et al., 2020), 16s rRNA gene (16s) (Valsecchi et al., 2020) or cytochrome b (cyt b) (Ma et al., 2016). Mitochondrial (mtDNA) genes are frequently targeted because of the high mtDNA copy number in eukaryotic cells (Clay Montier et al., 2009) and the availability of annotated and curated nucleotide sequence data from a wide range of species (Ross, 2003; Benson et al., 2013).
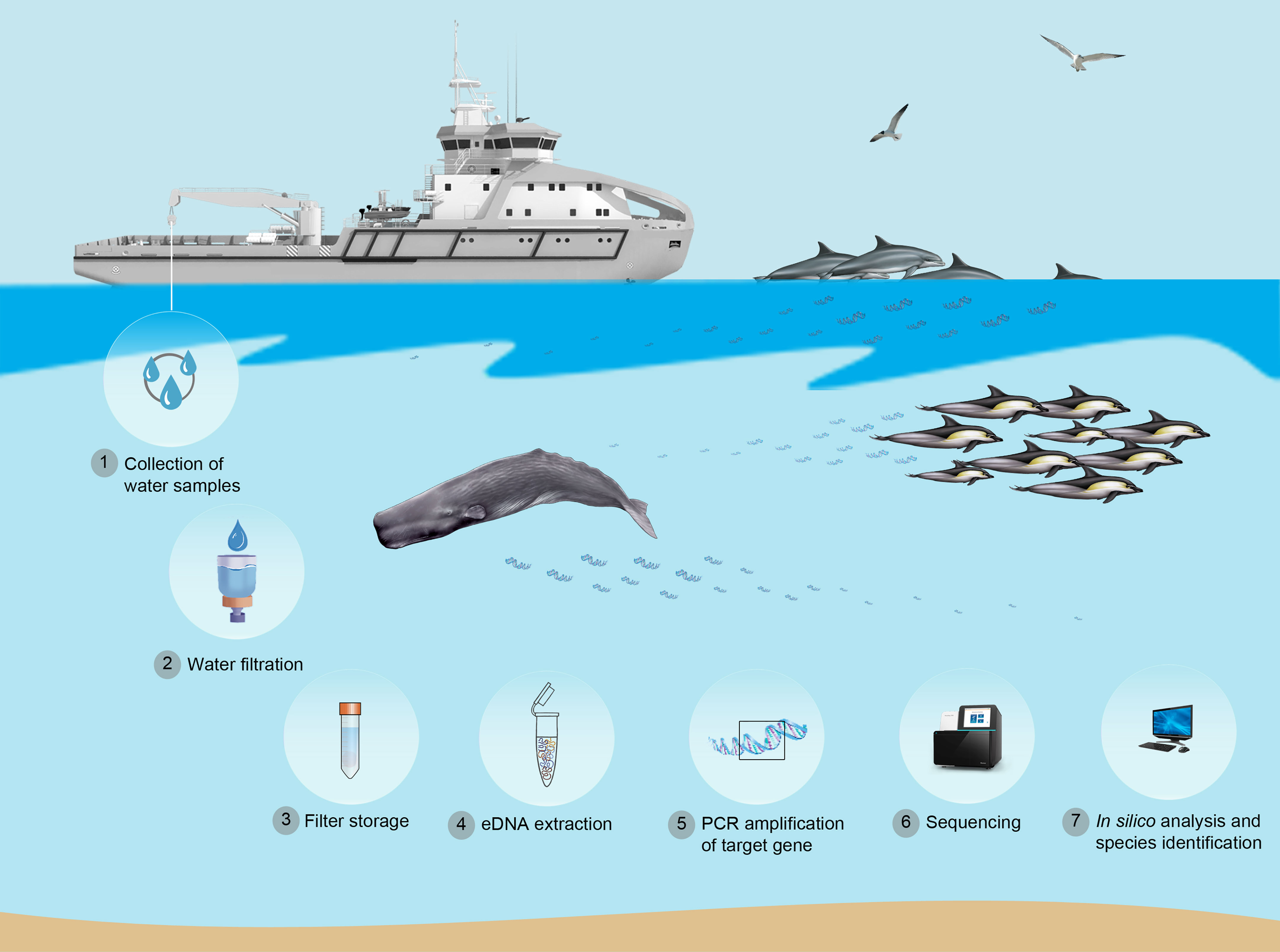
Figure 1 Application of eDNA methodology for marine mammals monitoring (single species, biodiversity and genetic characterization).
In recent years, eDNA studies in seawater have been used to monitor the diversity of marine vertebrate communities, primarily focusing on teleost fish and elasmobranchs (Thomsen et al., 2012; Miya et al., 2015; Andruszkiewicz et al., 2017; Truelove et al., 2019). However, eDNA studies on marine mammals are scarce and, to date, have mainly focused on the detection and identification of single species (Baker et al., 2018; Valsecchi et al., 2022), especially elusive or rare species (Foote et al., 2012; Ma et al., 2016), rather than the monitoring of marine mammals diversity. This is due, in part, to the lack of efficient taxon-specific metabarcoding primers. Previous studies evaluating marine vertebrate biodiversity identified some groups of marine mammals (Kelly et al., 2014; Andruszkiewicz et al., 2017; Closek et al., 2019) using universal vertebrate (Riaz et al., 2011) or fish primers (Miya et al., 2015). More recently, novel universal primers for marine vertebrates have been specifically designed and optimized to increase the efficiency of detection of marine mammals through eDNA analysis in seawater samples (Valsecchi et al., 2020; Valsecchi et al., 2021).
The molecular detection of marine mammals through eDNA obtained from water samples offers a non-invasive approach to not only to identify which species are present and to infer their temporal and spatial distribution, but also to characterize intraspecific genetic diversity (Parsons et al., 2018; Székely et al., 2021).
In this paper, we present a review of the use of eDNA for monitoring marine mammals. We focus on three different eDNA approaches that have been used to date: single species detection, biodiversity assessment and genetic characterization. Finally, we discuss the challenges and limitations associated with eDNA methodology and marine mammal assessment as well as the opportunities and benefits that eDNA could bring to this field.
Systematic literature review
We conducted a Google Scholar search (https://scholar.google.com) for articles related to environmental DNA and marine mammals from 2012 to 2021. The literature search included the terms “environmental DNA OR eDNA” AND “marine mammals” in the document title or text. The search yielded 893 publications and were manually filtered according to the following criteria: a) titles and abstracts were consistent with the search objective; b) they were peer-reviewed studies (excluding preprints); and c) they were published in international journals. We excluded review papers, media posts, opinions, perspectives, letters, conference abstracts and technical reports. A total of 21 papers was identified as having met these criteria.
Single species detection
Several eDNA studies targeting marine mammals have adopted a species-specific assay, demonstrating this to be useful for detecting some rare, cryptic, elusive and endangered species. The harbor porpoise Phocoena phocoena (Foote et al., 2012) and Yangtze finless porpoise (Neophocaena asiaeorientalis) (Ma et al., 2016; Qu and Stewart, 2019) were both detected in separate studies using eDNA analysis of water samples collected in both controlled and natural environments. For harbor porpoise detection, Foote et al. (2012) used qPCR and specific primers targeting 12s rRNA gene. In addition to harbor porpoise, long-finned pilot whale (Globicephala melas) was also identified at natural sites by using additional PCR primer sets (D-loop and cyt b) when amplicons did not match the porpoise reference sequence (Foote et al., 2012). This study of harbor porpoise was the first study to highlight the potential for eDNA isolated from surface seawater samples to detect both detected and undetected marine mammal species at a specific location, and also highlighted the importance of the primer design and optimization to obtain an accurate target species identification. Similarly, primers targeting the D-loop, designed to specifically detect humpback whale (Megaptera novaengliae) eDNA by qPCR assay, showed cross-amplification with minke whale (Balaenoptera acutorostrata) and gray whale (Eschrichtius robustus) (Andruszkiewicz et al., 2020). Both studies provide compelling evidence for the need for careful validation of qPCR assays both in silico and using vouchered DNA samples to ensure assay specificity, capture intraspecific variants and avoid off-target amplification. On the other hand, harbor porpoise eDNA analysis by Foote et al. (2012) in a controlled environment (four harbor porpoises maintained in a sea pen in a sheltered port area) showed that the animals could be detected at distances of up to 10 meters from the sea pen (Foote et al., 2012). In contrast, seawater sampling at distances of around 20 meters from killer whales (Orcinus orca) in both inshore and offshore waters failed to yield positive detections, emphasizing that differences in assay sensitivity as well as numerous abiotic and biotic factors could influence eDNA persistence in the marine environment and, consequently, the probability of detection (Pinfield et al., 2019). Baker et al. (2018) also highlighted marine environment dynamics as a likely causative factor underlying the stochastic detection of killer whales. Nevertheless, they were successful in identifying the target species up to 2h after the killer whales encounters using droplet digital PCR (ddPCR) technology (Baker et al., 2018). Other assays based on ddPCR and quantitative PCR (qPCR) were shown to be highly efficient to detect manatee species (Trichechus manatus, T. inunguis and T. senegalensis) using genus-specific primers (Hunter et al., 2018).
Biodiversity monitoring
Recent advances in metagenomics have demonstrated the successes of efforts to characterize marine vertebrate communities via eDNA metabarcoding using NGS. Initial studies focused primarily on investigating fish diversity using universal vertebrate (Riaz et al., 2011) or fish (Miya et al., 2015) primers, while also resolving several marine mammal species (Table 1). Kelly et al. (2014) unintentionally detected sea otters (Enhydra lutris) when analyzing the fish fauna from aquarium tanks using vertebrate 12S rRNA gene primers (termed 12S-V5) (Riaz et al., 2011), providing evidence of eDNA from marine mammals inhabiting the nearshore environment, which probably entered the system through the aquarium seawater intake (Kelly et al., 2014). Subsequently, Miya et al. (2015) designed universal primers (termed MiFish) to amplify a hypervariable region of the 12S rRNA gene from various fish species. These primers also amplified non-fish vertebrates present in mesocosm tanks, such as pantropical spotted dolphin (Stenella attenuata) (Miya et al., 2015), and have successfully generated metabarcoding sequence data for marine mammals in other studies including Gold et al. (2021a; 2021b) and Monuki et al. (2021). Both 12S-V5 and MiFish universal primer sets were used for marine vertebrate eDNA monitoring, identifying species across all four marine mammal taxonomic groups (i.e., fissipeds, pinnipeds, cetaceans and sirenians) (Table 1). Studies in Monterey Bay (California) detected sea otters, seals and sea lions associated with kelp forest and other marine microhabitats, in concordance with visual observations (Port et al., 2016; Djurhuus et al., 2017). Although no visual data were recorded in open waters by Port et al. (2016), metabarcoding analysis detected eDNA from California sea lions (Zalophus californianus) and Risso’s dolphins (Grampus griseus) (Port et al., 2016). Other studies in the same geographical area identified pinniped and cetacean species, representing the families Balaenopteridae, Delphinidae, Otariidae (Andruszkiewicz et al., 2017; Gold et al., 2021b) and Phocidae (Andruszkiewicz et al., 2017). Seawater sampling in the central California current (including Monterey Bay station among others) also revealed eDNA from Eschrichtiidae and Phocoenidae (Closek et al., 2019). These studies highlight the power of eDNA metabarcoding for detecting and monitoring marine mammals in both time and space, as for other vertebrate and invertebrate taxa (O’Donnell et al., 2017; Jeunen et al., 2019; Fraija‐Fernández et al., 2020). However, marine mammals in general represent very rare ‘targets’ in large open water systems and the use of more generalized barcodes, rather than more taxonomically specific primers, may limit detectability. Closek et al. (2019) found that eDNA metabarcoding did not return as many marine mammal taxa detections as obtained by traditional (dedicated annual visual survey) observation, with the exception of bottlenose dolphin (Tursiops truncatus) which was only detected by eDNA analysis (Closek et al., 2019).
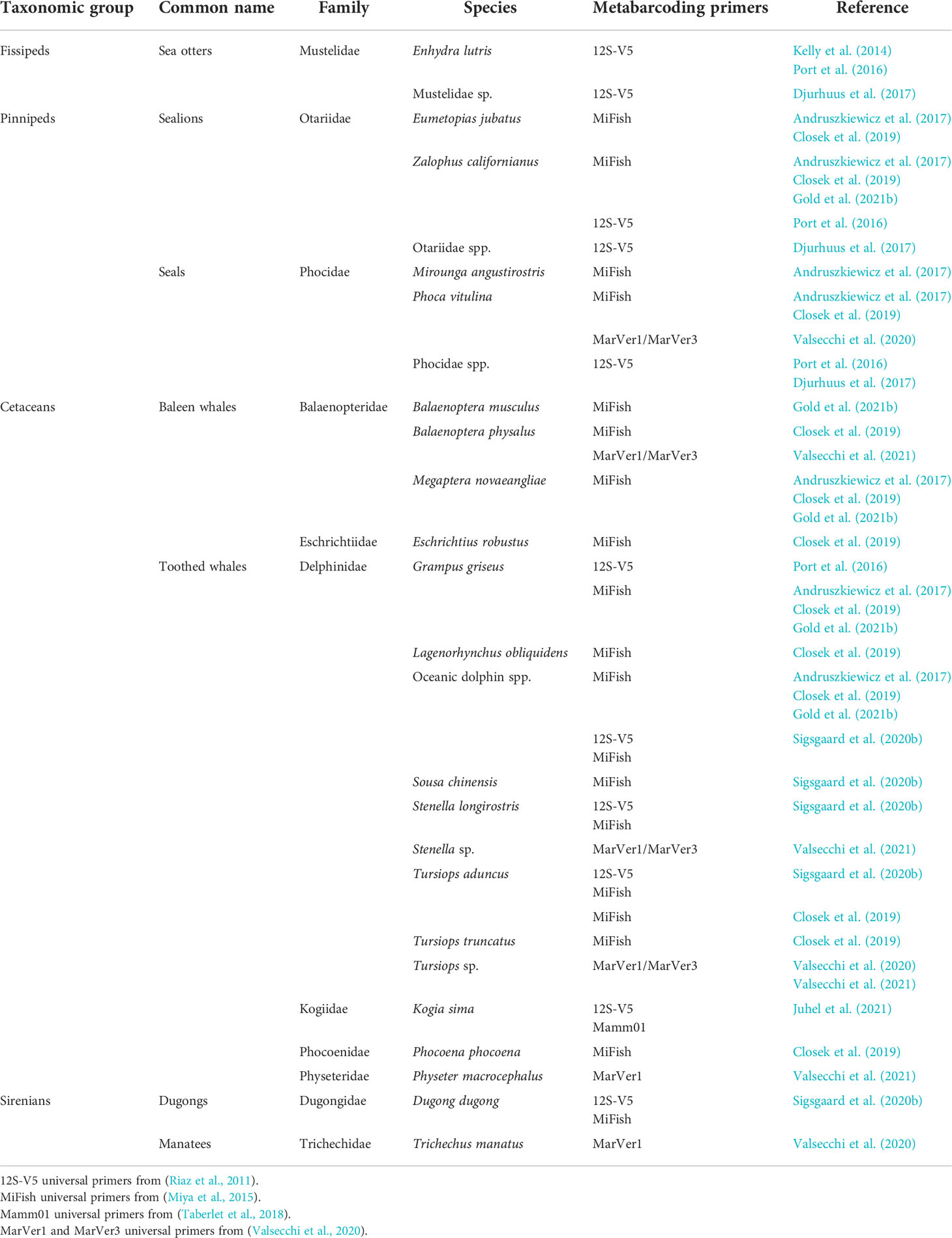
Table 1 Marine mammal species identified by eDNA metabarcoding studies using published vertebrate universal primers.
Inconsistencies in detections captured by eDNA metabarcoding and traditional methods were also found in fish studies. On the one hand, false-negative detections by eDNA metabarcoding have been attributed to the low abundance of certain species, sequencing errors, or an incomplete reference database (Fraija‐Fernández et al., 2020). On the other hand, eDNA metabarcoding has often reported more species than trawling and visual census methods, possibly due to the difficulty of using traditional methods to catch or detect certain pelagic species, cryptic species or small individuals (Yamamoto et al., 2017; Fraija‐Fernández et al., 2020)1
Bottlenose dolphin eDNA was detected by amplification of MiFish universal primers followed by nested PCR using GoFish species-specific primers, as a strategy to evaluate presence/absence in a time-series of water samples (Stoeckle et al., 2018). Diversity biomonitoring in the Persian Gulf using vertebrate-specific and MiFish metabarcoding primer sets identified some dolphin species in open waters and, interestingly, dugong (Dugong dugon) eDNA was detected at seagrass and sand bottom sites (Sigsgaard et al., 2020b). Another eDNA metabarcoding study, using 12S-V5 (Riaz et al., 2011) and mammal-specific (Mamm01) (Taberlet et al., 2018) universal primers, identified dwarf sperm whale (Kogia sima), which is an elusive and hard-to-sight species around Malpelo island (Columbia) (Juhel et al., 2021) (Table 1).
Recently, novel 12S and 16S rRNA metabarcoding primers (MarVer1 and MarVer3, respectively), designed to target marine vertebrates, have been specifically optimized to maximize the detection probability and taxonomic resolution of marine mammal groups (Valsecchi et al., 2020). These universal primer sets were shown to be effective for in silico identification of most families of cetaceans (Balaenopteridae, Delphinidae, Ziphiidae and Phocoenidae), Pinnipeds (Phocidae and Otariidae) and Sirenians (Dugongidae and Trichechidae). However, unresolved species were found within the Delphinidae, specifically between Tursiops truncatus and Sousa chinensis, and between Tursiops aduncus and Delphinus capensis1 (Valsecchi et al., 2020). The MarVer primers were also used to perform systematic eDNA surveys of the Mediterranean Sea taking advantage of commercial ferry routes. Two species (Balaenoptera physalus and Physeter macrocephalus) were unambiguously identified but two other species (Tursiops truncatus and Stenella coeruleoalba) were erroneously assigned to non-resident congeneric species (Tursiops aduncus and Stenella frontalis or Stenella longirostris, respectively) due to low taxonomic resolution for delphinids at this locus (Valsecchi et al., 2021) (Table 1).
Genetic characterization
Genetic analysis to gain information about haplotype diversity and differentiation of marine mammal populations often relies on direct sampling methods including biopsy darting (reviewed in Noren and Mocklin, 2012). The use of remote biopsy dart sampling to collect tissue samples (e.g., skin and blubber) from wild animals, in addition to being an invasive technique, poses skill and logistic challenges, which are exacerbated for fast-swimming, small, and elusive cetaceans. Recently, genetic characterization via eDNA sampling has emerged as an alternative or complementary method to traditional tissue sampling methods, given its potential for generating valuable genetic information (Sigsgaard et al., 2020a). Killer whale eDNA analysis led to haplotype identification in some high quality mtDNA sequences and, thus, confirmed the ecotype of the detected whales (Baker et al., 2018). High-throughput sequencing of eDNA captured from harbor porpoise “footprints” generated mtDNA haplotypes for the Alaska southeast population across sampled geographic regions, thus contributing to the characterization of its genetic diversity and assessments of stock structure based on genetic differentiation (Parsons et al., 2018). Recently, a study of bowhead whales (Balaena mysticetus) revealed that mitochondrial that haplotype frequencies obtained from eDNA samples, collected from “footprints”, were similar to those recorded by biopsy-darting over the years (Székely et al., 2021).
Discussion
The last decade has seen dramatic advances in the application of eDNA for marine organisms. Environmental DNA has been used to detect single or multiple taxa, in addition to characterizing intraspecific genetic diversity and differentiation. Some studies have obtained successful and encouraging results, while others have highlighted how challenging the eDNA technique can become in the vast ocean. Although eDNA promises to be a useful approach to support the monitoring and conservation of marine mammals, it is important to address the strengths and limitations, given the current state of knowledge, to inform choice of methods and facilitate appropriate ecological interpretations of eDNA data.
eDNA can be an accessible, non-invasive and cost-effective strategy. Environmental DNA has been used successfully to detect and identify single, target species of interest (Foote et al., 2012; Baker et al., 2018) as well as broader groups of marine mammals (Miya et al., 2015; Valsecchi et al., 2020; Valsecchi et al., 2021) including threatened, rare, cryptic and elusive species which are frequently overlooked in traditional visual monitoring. Moreover, eDNA can offer increased accuracy in taxonomic assignment for species that are difficult to sight or difficult to differentiate visually (Juhel et al., 2021). Some studies have also demonstrated the ability of eDNA to provide data that contributes to the knowledge of the genetic structure of populations (Parsons et al., 2018; Székely et al., 2021) as well as their temporal and spatial distribution patterns (Port et al., 2016; Djurhuus et al., 2017). Obtaining time-series data indicating the presence of key species is critical for assessment and conservation of marine mammal species, and in this context, eDNA methods could be implemented in periodic oceanographic surveys or in surveys based on Platforms Of Opportunity (POOs) (Valsecchi et al., 2021). Thus, eDNA offers a complementary approach that could be integrated into ongoing surveys to increase the data available from traditional visual monitoring for marine mammals.
The current limitations of eDNA for generating detections and sequence data for marine mammals can be defined broadly as those associated with assays design, and those attributable to the ‘ecology’ of eDNA (Barnes et al., 2014). eDNA assays including critical steps such as collection of water sample and amplication of the genetic material should be designed depending on the purpose of the research and the nature of water samples. Water sampling to detect the presence of single species or monitoring marine mammals biodiversity implies collection of water from multiple stations within a study area. In contrast, the genetic characterization of marine mammals populations usually entails sampling water directly from the animals "footprints" when they are sighted (Parsons et al., 2018; Székely et al., 2021). Although some coastal waters studies (Foote et al., 2012; Ma et al., 2016; Andruszkiewicz et al., 2017), and some studies based on collecting water from "footprints" (Székely et al., 2021), have isolated eDNA from small volumes of water (≤ 1 l), it has recently been recommended that 4-5 l of seawater should be sampled to assure efficient eDNA capture in both inshore and offshore waters (Valsecchi et al., 2021). Typically, eDNA amplification assays are designed to achieve a balance between the amplicon size and the number of polymorphic sites (Baker et al., 2018; Valsecchi et al., 2020). Due to potential environmental degradation of DNA over time (Barnes et al., 2014; Collins et al., 2018), studies have focused on amplification of short fragments (< 300bp) to maximize the detection rate of target eDNA. However, shorter amplicons may not contain sufficient nucleotide variation to avoid ambiguous taxonomic assignment, as found in previous studies targeting single species (Foote et al., 2012; Andruszkiewicz et al., 2020) and in metabarcoding studies, in which unequivocal identification of some delphinid species continues to be challenging (Closek et al., 2019; Sigsgaard et al., 2020b; Valsecchi et al., 2021). A recent study using Teleo01 metabarcoding primers (Valentini et al., 2016) demonstrated significant inconsistency in the marine mammals identified, resulting in the erroneous assignment of three non-endemic species to the Black Sea (Zhang et al., 2020b). This underscores the caution warranted in the interpretation of eDNA results. Using longer amplicons could be an alternative to obtain sufficient sequence information for species identification (Zhang et al., 2020a). Additionally, the detection of longer eDNA fragments could be used to infer more recent presence of animals in the field (Bista et al., 2017), due to their higher rate of degradation compared to that of short fragments (Jo et al., 2017). In recent years, amplicon length in eDNA metabarcoding studies has been constrained by the limitations in read length of commonly used short-read sequencing platforms (Illumina or BGI sequencers), although such platforms have provided greater accuracy and higher quality data compared to long-read sequencing technology (Pacific Biosciences or Oxford Nanopore) (Goodwin et al., 2016; Amarasinghe et al., 2020). However, a recent long-read eDNA metabarcoding study targeting fish retrieved and sequenced 2kb amplicons, showing enhanced taxonomic resolution using a new bioinformatics pipeline to generate reliable sequences from Nanopore sequencing (Doorenspleet et al., 2021).
Finally, the use of metabarcoding primers targeting a broader taxonomic group (i.e., fish, vertebrates) to study marine mammal species might produce false negative detections if other taxonomic groups are present in greater abundance in the eDNA sample (Miya et al., 2020).
Little is currently known about how environmental factors affect the spatial and temporal persistence of eDNA in aquatic systems. It has been shown that the quality and quantity of detectable eDNA can vary based on numerous biotic and abiotic factors, which influence DNA persistence in a different manner depending on its molecular state (extracellular, intracellular, free or adsorbed to particles) (Barnes et al., 2014; Strickler et al., 2015). One of main factors affecting eDNA persistence is the microbial community, which has been reported to utilize DNA as a source of carbon, nitrogen, phosphorus, and nucleic acid precursors (Vorkapic et al., 2016). Abiotic factors including temperature, UV radiation, salinity, pH and dissolved oxygen could affect eDNA degradation rates directly but also indirectly by influencing microbial metabolism (Barnes et al., 2014; Strickler et al., 2015).
By its very nature, the highly dynamic marine environment (currents, tides, waves, etc.) contributes to eDNA dispersion, dilution and degradation beyond detectability. Therefore, non-detections could be due to degradation or transport of target eDNA out of sampled areas as well as to low organism density or low quantity of cellular material shed into the environment (i.e. they could represent false negatives). In addition, hydrodynamic phenomena in marine ecosystems can transport eDNA from connected habitats leading to false positive detections (Collins et al., 2018). Nevertheless, studies have shown that the eDNA signal can be linked to discrete nearshore and open water habitats, suggesting relatively localized vertical and horizontal dispersal (Port et al., 2016; O’Donnell et al., 2017; Collins et al., 2018; Kelly et al., 2018; Jeunen et al., 2019; Monuki et al., 2021). Modeling studies of how oceanographic variables and degradation rates affect the temporal and spatial resolution of the eDNA signal in a given marine habitat are needed to support accurate ecological interpretations of eDNA detections and non-detections.
Conclusions and future directions
eDNA offers enormous potential for monitoring marine mammals that is complementary to traditional visual surveys, while simultaneously filling some of the persistent knowledge gaps for many threatened and endangered species. Despite their limitations, to date eDNA methods have resulted in the detection of marine mammals in areas where they were unreported or poorly reported in previous visual surveys, highlighting the sensitivity of genetic assays. However, in-depth investigations are still needed to understand how environmental factors (e.g., temperature, currents, tides, etc.) affect eDNA detection in the marine environment, to both optimize sampling and support interpretation of eDNA detections and non-detections. Implementing eDNA sampling in oceanographic surveys and in surveys based on POOs globally would benefit periodic data collection in the long-term. Finally, the use of eDNA as a complimentary tool, along with other traditional monitoring methods, will contribute to achieve improved monitoring programs for marine mammal assessment and conservation.
Author contributions
PS-B, MÁ-G, KMP, JR, GJP, and CS wrote and revised the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the Biodiversity Foundation project (NuTEC - BM2019/40) and Spanish Ministry for the Ecological Transition and the Demographic challenge, (MITECO) through the Commission [28-5307] for “Technical Scientific Advice for the Protection of the Marine Environment: Assessment and Monitoring of Marine Strategies, Monitoring of Marine Protected Areas of State Competence (2018-2021)”. PS-B was supported by NuTEC project contract.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
- ^ In light of recent evidence, Delphinus capensis has been recognized as Delphinus delphis by the Committee on Taxonomy of the Society for Marine Mammalogy (https://marinemammalscience.org/speciesinformation/list-marine-mammal-species-subspecies/).
References
Albouy C., Delattre V., Donati G., Frölicher T. L., Albouy-Boyer S., Rufino M., et al. (2020). Global vulnerability of marine mammals to global warming. Sci. Rep. 10, 1–12. doi: 10.1038/s41598-019-57280-3
Amarasinghe S. L., Su S., Dong X., Zappia L., Ritchie M. E., Gouil Q. (2020). Opportunities and challenges in long-read sequencing data analysis. Genome Biol. 21, 1–16. doi: 10.1186/s13059-020-1935-5
Andruszkiewicz E. A., Starks H. A., Chavez F. P., Sassoubre L. M., Block B. A., Boehm A. B. (2017). Biomonitoring of marine vertebrates in Monterey bay using eDNA metabarcoding. PloS One 12, 1–20. doi: 10.1371/journal.pone.0176343
Andruszkiewicz E. A., Yamahara K. M., Closek C. J., Boehm A. B. (2020). Quantitative PCR assays to detect whales, rockfish, and common murre environmental DNA in marine water samples of the northeastern pacific. PloS One 15, e0242689. doi: 10.1371/journal.pone.0242689
Baker C. S., Steel D., Nieukirk S., Klinck H (2018). (eDNA) from the wake of the whales: Droplet digital PCR for detection and species identification. Front. Mar. Sci. 5. doi: 10.3389/fmars.2018.00133
Barlow J., Ferguson M., Errin W. F. P., Ballance L., Gerrodette T., Joyce G. G, et al. (2006). Abundance and densities of beaked and bottlenose whales (family ziphiidae). J. Cetacean Res. Manage. 7 (3), 263–270.
Barnes M. A., Turner C. R., Jerde C. L., Renshaw M. A., Chadderton W. L., Lodge D. M. (2014). Environmental conditions influence eDNA persistance in aquatic systems. Environ. Sci. Technol. 48, 1819–1827. doi: 10.1371/journal.pone.0185043
Benson D. A., Cavanaugh M., Clark K., Karsch-Mizrachi I., Lipman D. J., Ostell J., et al. (2013). GenBank. Nucleic Acids Res. 41, D36–D42. doi: 10.1093/nar/gks1195
Bista I., Carvalho G. R., Walsh K., Seymour M., Hajibabaei M., Lallias D., et al. (2017). Annual time-series analysis of aqueous eDNA reveals ecologically relevant dynamics of lake ecosystem biodiversity. Nat. Commun. 8, 14087. doi: 10.1038/ncomms14087
Booth C. G., Sinclair R. R., Harwood J. (2020). Methods for monitoring for the population consequences of disturbance in marine mammals: A review. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00115
Clay Montier L. L., Deng J. J., Bai Y. (2009). Number matters: control of mammalian mitochondrial DNA copy number. J. Genet. Genomics 36, 125–131. doi: 10.1016/S1673-8527(08)60099-5
Closek C. J., Santora J. A., Starks H. A., Schroeder I. D., Andruszkiewicz E. A., Sakuma K. M., et al. (2019). Marine vertebrate biodiversity and distribution within the central California current using environmental DNA (eDNA) metabarcoding and ecosystem surveys. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00732
Collins R. A., Wangensteen O. S., O’Gorman E. J., Mariani S., Sims D. W., Genner M. J. (2018).Persistence of environmental DNA in marine systems. J. Commun. Biol. 1, 1–11. doi: 10.1038/s42003-018-0192-6
DeMaster D. P., Fowler C. W., Perry S. L., Richlen M.F. (2001). Predation and competition: the impact of fisheries on marine-mammal populations over the next one hundred years. J. Mammal. 82, 641–651. doi: 10.1644/1545-1542(2001)082<0641:PACTIO>2.0.CO;2
Desforges J. P. W., Sonne C., Levin M., Siebert U., De Guise S., Dietz R. (2016). Immunotoxic effects of environmental pollutants in marine mammals. Environ. Int. 86, 126–139. doi: 10.1016/j.envint.2015.10.007
Dick D. M., Hines E. M. (2011). Using distance sampling techniques to estimate bottlenose dolphin (Tursiops truncatus) abundance at turneffe atoll, Belize. Mar. Mammal Sci. 27, 606–621. doi: 10.1111/j.1748-7692.2010.00435.x
Djurhuus A., Port J., Closek C. J., Yamahara K. M., Romero-Maraccini O., Walz K. R., et al. (2017). Evaluation of filtration and DNA extraction methods for environmental DNA biodiversity assessments across multiple trophic levels. Front. Mar. Sci. 4. doi: 10.3389/fmars.2017.00314
Doorenspleet K., Jansen L., Oosterbroek S., Bos O., Kamermans P., Janse M., et al. (2021). High resolution species detection: accurate long read eDNA metabarcoding of north Sea fish using Oxford nanopore sequencing. bioRxiv 2021, 11.26.470087. doi: 10.1101/2021.11.26.470087
Estes J. A., Terborgh J., Brashares J. S., Power M. E., Berger J., Bond W. J., et al. (2011). Trophic downgrading of planet earth. Science. (80-. ). 333, 301–306. doi: 10.1126/science.1205106
Ficetola G. F., Miaud C., Pompanon F., Taberlet P. (2008). Species detection using environmental DNA from water samples. Biol. Lett. 4, 423–425. doi: 10.1098/rsbl.2008.0118
Foote A. D., Thomsen P. F., Sveegaard S., Wahlberg M., Kielgast J., Kyhn L. A., et al. (2012). Investigating the potential use of environmental DNA (eDNA) for genetic monitoring of marine mammals. PloS One 7, 2–7. doi: 10.1371/journal.pone.0041781
Fraija-Fernández N., Bouquieaux M., Rey A., Mendibil I., Cotano U., Irigoien X., et al. (2020). Marine water environmental DNA metabarcoding provides a comprehensive fish diversity assessment and reveals spatial patterns in a large oceanic area. Ecol. Evol. 10, 7560–7584. doi: 10.1002/ece3.6482
Gold Z., Curd E. E., Goodwin K. D., Choi E. S., Frable B. W., Thompson A. R., et al. (2021a). Improving metabarcoding taxonomic assignment: A case study of fishes in a large marine ecosystem. Mol. Ecol. Resour. 21, 2546–2564. doi: 10.1111/1755-0998.13450
Gold Z., Sprague J., Kushner D. J., Zerecero Marin E., Barber P. H. (2021b). eDNA metabarcoding as a biomonitoring tool for marine protected areas. PloS One 16, e0238557. doi: 10.1371/journal.pone.0238557
Goodwin S., McPherson J. D., McCombie W. R. (2016). Coming of age: Ten years of next-generation sequencing technologies. Nat. Rev. Genet. 17, 333–351. doi: 10.1038/nrg.2016.49
Hammond P. S., Macleod K., Berggren P., Borchers D. L., Burt L., Cañadas A., et al. (2013). Cetacean abundance and distribution in European Atlantic shelf waters to inform conservation and management. Biol. Conserv. 164, 107–122. doi: 10.1016/j.biocon.2013.04.010
Hunter M. E., Meigs-Friend G., Ferrante J. A., Kamla A. T., Dorazio R. M., Diagne L. K., et al. (2018). Surveys of environmental DNA (eDNA): A new approach to estimate occurrence in vulnerable manatee populations. Endanger. Species Res. 35, 101–111. doi: 10.3354/esr00880
Jeunen G., Knapp M., Spencer H. G., Lamare M. D., Taylor H. R., Stat M., et al. (2019). Environmental DNA (eDNA) metabarcoding reveals strong discrimination among diverse marine habitats connected by water movement. Mol. Ecol. Resour. 19, 426–438. doi: 10.1111/1755-0998.12982
Jo T., Murakami H., Masuda R., Sakata M. K., Yamamoto S., Minamoto T. (2017). Rapid degradation of longer DNA fragments enables the improved estimation of distribution and biomass using environmental DNA. Mol. Ecol. Resour. 17, e25–e33. doi: 10.1111/1755-0998.12685
Juhel J. B., Marques V., Polanco Fernández A., Borrero-Pérez G. H., Mutis Martinezguerra M., Valentini A., et al. (2021). Detection of the elusive dwarf sperm whale (Kogia sima) using environmental DNA at malpelo island (Eastern pacific, Colombia). Ecol. Evol. 11, 2956–2962. doi: 10.1002/ece3.7057
Kelly R. P., Gallego R., Jacobs-Palme E. (2018). The effect of tides on nearshore environmental DNA. PeerJ 2018, e4521. doi: 10.7717/peerj.4521
Kelly R. P., Port J. A., Yamahara K. M., Crowder L. B. (2014). Using environmental DNA to census marine fishes in a Large mesocosm. PloS One 9, e86175. doi: 10.1371/journal.pone.0086175
Ma H., Stewart K., Lougheed S., Zheng J., Wang Y., Zhao J. (2016). Characterization, optimization, and validation of environmental DNA (eDNA) markers to detect an endangered aquatic mammal. Conserv. Genet. Resour. 8, 561–568. doi: 10.1007/s12686-016-0597-9
Miya M., Gotoh R. O., Sado T. (2020). MiFish metabarcoding: a high-throughput approach for simultaneous detection of multiple fish species from environmental DNA and other samples. Fish. Sci. 86, 939–970. doi: 10.1007/s12562-020-01461-x
Miya M., Sato Y., Fukunaga T., Sado T., Poulsen J. Y., Sato K., et al. (2015). MiFish, a set of universal PCR primers for metabarcoding environmental DNA from fishes: Detection of more than 230 subtropical marine species. R. Soc Open Sci. 2, 1–33. doi: 10.1098/rsos.150088
Monuki K., Barber P. H., Gold Z. (2021). eDNA captures depth partitioning in a kelp forest ecosystem. PloS One 16, e0253104. doi: 10.1371/journal.pone.0253104
Nabi G., McLaughlin R. W., Hao Y., Wang K., Zeng X., Khan S., et al. (2018). The possible effects of anthropogenic acoustic pollution on marine mammals’ reproduction: an emerging threat to animal extinction. Environ. Sci. Pollut. Res. 25, 19338–19345. doi: 10.1007/s11356-018-2208-7
Noren D. P., Mocklin J. A. (2012). Review of cetacean biopsy techniques: Factors contributing to successful sample collection and physiological and behavioral impacts. Mar. Mammal Sci. 28, 154–199. doi: 10.1111/j.1748-7692.2011.00469.x
O’Donnell J. L., Kelly R. P., Shelton A. O., Samhouri J. F., Lowell N. C., Williams G. D. (2017). Spatial distribution of environmental DNA in a nearshore marine habitat. PeerJ 2017, e3044. doi: 10.7717/peerj.3044
Parsons K. M., Meredith E., Marilyn D., Linda P. (2018). Water, water everywhere: environmental DNA can unlock population structure in elusive marine species. R. Soc Open Sci. 5, 180537. doi: 10.1098/rsos.180537
Pikitch E. K. (2018). A tool for finding rare marine species. Science 360, 1180–1182. doi: 10.1126/science.aao3787
Pinfield R., Dillane E., Runge A. K. W., Evans A., Mirimin L., Niemann J., et al. (2019). False-negative detections from environmental DNA collected in the presence of large numbers of killer whales ( orcinus orca ). Environ. DNA 1, 316–328. doi: 10.1002/edn3.32
Port J. A., O’Donnell J. L., Romero-Maraccini O. C., Leary P. R., Litvin S. Y., Nickols K. J., et al. (2016). Assessing vertebrate biodiversity in a kelp forest ecosystem using environmental DNA. Mol. Ecol. 25, 527–541. doi: 10.1111/mec.13481
Qu C., Stewart K. A. (2019). Evaluating monitoring options for conservation: comparing traditional and environmental DNA tools for a critically endangered mammal. Sci. Nat. 106, 1–9. doi: 10.1007/s00114-019-1605-1
Ramón-Laca A., Wells A., Park L. (2021). A workflow for the relative quantification of multiple fish species from oceanic water samples using environmental DNA (eDNA) to support large-scale fishery surveys. PloS One 16, e0257773. doi: 10.1371/journal.pone.0257773
Reeves R. R., McClellan K., Werner T. B. (2013). Marine mammal bycatch in gillnet and other entangling net fisherie to 2011. Endanger. Species Res. 20, 71–97. doi: 10.3354/esr00481
Riaz T., Shehzad W., Viari A., Pompanon F., Taberlet P., Coissac E. (2011). EcoPrimers: Inference of new DNA barcode markers from whole genome sequence analysis. Nucleic Acids Res. 39, e145. doi: 10.1093/nar/gkr732
Roman J., Estes J. A. (2018). “Ecology,” in Encyclopedia of marine mammals (Elsevier), 299–303. doi: 10.1016/B978-0-12-804327-1.00114-X
Ross H. A. (2003). DNA Surveillance: Web-based molecular identification of whales, dolphins, and porpoises. J. Hered. 94, 111–114. doi: 10.1093/jhered/esg027
Schipper J., Chanson J. S., Chiozza F., Cox N. A., Hoffmann M., Katariya V., et al. (2008). The status of the world’s land and marine mammals: diversity, threat, and knowledge. Sci. (80-. ). 322, 225–230. doi: 10.1126/science.1165115
Schoeman R. P., Patterson-Abrolat C., Plön S. (2020). A global review of vessel collisions with marine animals. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00292
Sigsgaard E. E., Jensen M. R., Winkelmann I. E., Møller P. R., Hansen M. M. (2020a). Population-level inferences from environmental DNA–current status and future perspectives. Evol. Appl. 13, 245–262. doi: 10.1111/eva.12882
Sigsgaard E. E., Torquato F., Frøslev T. G., Moore A. B. M., Sørensen J. M., Range P., et al. (2020b). Using vertebrate environmental DNA from seawater in biomonitoring of marine habitats. Conserv. Biol. 34, 697–710. doi: 10.1111/cobi.13437
Stoeckle M. Y., Das Mishu M., Charlop-Powers Z. (2018). GoFish: A versatile nested PCR strategy for environmental DNA assays for marine vertebrates. PloS One 13, e0198717. doi: 10.1371/journal.pone.0198717
Strickler K. M., Fremier A. K., Goldberg C. S. (2015). Quantifying effects of UV-b, temperature, and pH on eDNA degradation in aquatic microcosms. Biol. Conserv. 183, 85–92. doi: 10.1016/j.biocon.2014.11.038
Székely D., Corfixen N. L., Mørch L., Knudsen S. W., McCarthy M., Teilmann J., et al. (2021). Environmental DNA captures the genetic diversity of bowhead whales (Balaena mysticetus) in West Greenland. Environ. DNA 3, 248–260. doi: 10.1002/edn3.176
Taberlet P., Bonin A., Zinger L., Coissac E. (2018). Environmental DNA: For biodiversity research and monitoring (Oxford: Oxford University Press). doi: 10.1093/oso/9780198767220.001.0001
Taberlet P., Coissac E., Hajibabaei M., Rieseberg L. H. (2012). Environmental DNA. Mol. Ecol. 21, 1789–1793. doi: 10.1111/j.1365-294X.2012.05542.x
Thomsen P. F., Kielgast J., Iversen L. L., Møller P. R., Rasmussen M., Willerslev E. (2012). Detection of a diverse marine fish fauna using environmental DNA from seawater samples. PloS One 7, e41732. doi: 10.1371/journal.pone.0041732
Thomsen P. F., Willerslev E. (2015). Environmental DNA - an emerging tool in conservation for monitoring past and present biodiversity. Biol. Conserv. 183, 4–18. doi: 10.1016/j.biocon.2014.11.019
Todd V. L. G., Todd I. B., Gardiner J. C., Morrin E. C. N., MacPherson N. A., DiMarzio N. A., et al. (2014). A review of impacts of marine dredging activities on marine mammals. ICES J. Mar. Sci. 72, 328–340. doi: 10.1093/icesjms/fsu187
Truelove N. K., Andruszkiewicz E. A., Block B. A. (2019). A rapid environmental DNA method for detecting white sharks in the open ocean. Methods Ecol. Evol. 10, 1128–1135. doi: 10.1111/2041-210X.13201
Tsuji S., Takahara T., Doi H., Shibata N., Yamanaka H. (2019). The detection of aquatic macroorganisms using environmental DNA analysis–a review of methods for collection, extraction, and detection. Environ. DNA 1, 99–108. doi: 10.1002/edn3.21
Urian K., Gorgone A., Read A., Balmer B., Wells R. S., Berggren P., et al. (2015). Recommendations for photo-identification methods used in capture-recapture models with cetaceans. Mar. Mammal Sci. 31, 298–321. doi: 10.1111/mms.12141
Valentini A., Taberlet P., Miaud C., Civade R., Herder J., Thomsen P. F., et al. (2016). Next-generation monitoring of aquatic biodiversity using environmental DNA metabarcoding. Mol. Ecol. 25, 929–942. doi: 10.1111/mec.13428
Valsecchi E., Arcangeli A., Lombardi R., Boyse E., Carr I. M., Galli P., et al. (2021). Ferries and environmental DNA: Underway sampling from commercial vessels provides new opportunities for systematic genetic surveys of marine biodiversity. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.704786
Valsecchi E., Bylemans J., Goodman S. J., Lombardi R., Carr I., Castellano L., et al. (2020). Novel universal primers for metabarcoding eDNA surveys of marine mammals and other marine vertebrates. Environ. DNA 00, 1–17. doi: 10.1002/edn3.72
Valsecchi E., Coppola E., Pires R., Parmegiani A., Casiraghi M., Galli P., et al. (2022). A species-specific qPCR assay provides novel insight into range expansion of the Mediterranean monk seal (Monachus monachus) by means of eDNA analysis. Biodivers. Conserv. 31, 1175–1196. doi: 10.1007/s10531-022-02382-0
Vorkapic D., Pressler K., Schild S. (2016). Multifaceted roles of extracellular DNA in bacterial physiology. Curr. Genet. 62, 71–79. doi: 10.1007/s00294-015-0514-x
Xie R., Zhao G., Yang J., Wang Z., Xu Y., Zhang X., et al. (2021). eDNA metabarcoding revealed differential structures of aquatic communities in a dynamic freshwater ecosystem shaped by habitat heterogeneity. Environ. Res. 201, 111602. doi: 10.1016/j.envres.2021.111602
Yamamoto S., Masuda R., Sato Y., Sado T., Araki H., Kondoh M., et al. (2017). Environmental DNA metabarcoding reveals local fish communities in a species-rich coastal sea. Sci. Rep. 7, 1–12. doi: 10.1038/srep40368
Zerbini A. N., Waite J. M., Durban J. W., LeDuc R., Dahlheim M. E., Wade P.R (2007). Estimating abundance of killer whales in the nearshore waters of the gulf of Alaska and Aleutian islands using line-transect sampling. Mar. Biol. 150, 1033–1045. doi: 10.1007/s00227-006-0347-8
Zhang Y., Pavlovska M., Stoica E., Prekrasna I., Yang J., Slobodnik J., et al. (2020b). Holistic pelagic biodiversity monitoring of the black Sea via eDNA metabarcoding approach: From bacteria to marine mammals. Environ. Int. 135, 105307. doi: 10.1016/j.envint.2019.105307
Keywords: environmental DNA, eDNA, marine mammals, monitoring, assessment, conservation
Citation: Suarez-Bregua P, Álvarez-González M, Parsons KM, Rotllant J, Pierce GJ and Saavedra C (2022) Environmental DNA (eDNA) for monitoring marine mammals: Challenges and opportunities. Front. Mar. Sci. 9:987774. doi: 10.3389/fmars.2022.987774
Received: 06 July 2022; Accepted: 05 September 2022;
Published: 23 September 2022.
Edited by:
David M. P. Jacoby, University of Lancaster, United KingdomReviewed by:
Chloe Victoria Robinson, Ocean Wise, CanadaJason Doyle, Australian Institute of Marine Science (AIMS), Australia
Copyright © 2022 Suarez-Bregua, Álvarez-González, Parsons, Rotllant, Pierce and Saavedra. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paula Suarez-Bregua, cGF1bGFzdWFyZXpAaWltLmNzaWMuZXM=