 Sara Königson
Sara Königson Rahmat Naddafi
Rahmat Naddafi Sven-Gunnar Lunneryd1
Sven-Gunnar Lunneryd1 Andreas C. Bryhn
Andreas C. Bryhn Kelly Macleod
Kelly Macleod Peter Ljungberg
Peter Ljungberg
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 20 December 2022
Sec. Marine Fisheries, Aquaculture and Living Resources
Volume 9 - 2022 | https://doi.org/10.3389/fmars.2022.985431
This article is part of the Research TopicInnovations in Fishing Technology Aimed at Achieving Sustainable FishingView all 23 articles
Small-scale fisheries along the Swedish west coast are often operated by using small vessels, targeting multiple species by means of pots and gillnets. Fisheries using pots targeting shellfish such as European lobster (Homarus gammarus), and edible crab (Cancer pagurus) along the Swedish west coast have a relatively high economic value. However, gillnet fisheries targeting demersal fish are currently diminishing primarily due to depleted fish stocks and increased seal depredation. Small-scale fisheries are considered sustainable fisheries due to the use of selective gears and low energy consumption. To be able to retain and promote a sustainable small-scale fishery, there is a need to develop an economically viable fishery, where selective, seal-safe and sustainable gear is utilized. One potential way forward could be to develop a pot to be used for multiple target species traditionally caught in pots and gillnets. Since both shellfish and cod (Gadus morhua) can be caught in pots, the objective of this work was to develop a multispecies pot targeting lobster, edible crab and cod. Seven multi-species pots were developed and tested off the west coast of Sweden between 2015 and 2017. The catch rate, defined as catch per pot per day (CPUE) of lobster, edible crab and cod, was evaluated taking into regard fisheries-related variables such as pot type, bait, soak time, seal damage and abundance of species in the pot. The relative CPUE of lobster and cod was highest in larger pots with two chambers and three open entrances. The highest CPUE for lobster was 0.24 individuals per pot and the highest CPUE for cod was 0.17 individuals per pot. Pots with entrances equipped with funnels, preventing cod from escaping, also had a high cod CPUE (0.23 individuals per). The CPUE of crabs was not affected by pot type. For cod, lobster and crab, the CPUE significantly decreased with increasing soak time. Seal damage only occurred when cod were trapped in the pots and the CPUE of cod was higher in pots subjected to seal damage, indicating that seals raid pots specifically when cods are trapped inside.
The small-scale fishery along the west coast of Sweden has been subjected to major changes during the past 30 years. Atlantic cod (Gadus morhua) used to be one of the major species targeted by fisheries along the coast. However, since the late 20th century, the cod stocks have declined (Hentati Sundberg, 2017; Bergenius et al., 2018). Nowadays, the main target species in small-scale fisheries include European lobster (Homarus gammarus), edible crab (Cancer pagurus), Norway lobster (Nephrops norvegicus), mackerel (Scomber scombrus), plaice (Pleuronectes platessa), witch flounder (Glyptocephalus cynoglossus) and to a certain extent cod. Small-scale fisheries along the Swedish west coast are often described as a fishery where boats are operated individually, using vessels (vessels less than 12 meters), targeting multiple species and where the catch is sold directly to the customer or it is refined, thereby increasing its value. Smallscale fisheries primarily use pots and gillnets. Pot fisheries targeting shellfish such as crab and Norway lobster, have increased along the Swedish west coast over the past 15 years. In contrast, the small-scale gillnet fishery mainly targeting cod has decreased (Bergenius et al., 2018), primarily due to depleted fish stocks but also increased seal depredation which adds economic costs to the fishers. The pot fishery targeting European lobster is important along much of the coast of Europe (De Wit et al., 2021). In Sweden along the west coast, European lobster is the species with the highest annual landing value (10,000,000 SEK) followed by mackerel, labrids, edible crab (landing value of 5,000,000 SEK) and cod (landing value of 3,750,000 SEK) (Bergenius et al., 2018).
Pots are a potential alternative to gillnets targeting cod. Pots are considered to be a fuel-efficient type of gear that has a minimal impact on the bottom substrate (Suuronen et al., 2012) and can also be made species and size selective, decreasing bycatch of non-target species or undersized target species. Cod, similar to lobster and edible crab, can be lured into and thereafter retained in baited pots. In the southern Baltic Sea, cod pots have been successfully developed as an alternative to the seal damaged gillnet fisheries for cod (Königson et al., 2015). The main reason for this pot development in the Baltic Sea is the severe depredation, mainly by grey seal (Halichoerus grypus) but also to some extent by seabirds, in gillnet cod fisheries. Pots that can be made seal-safe because the catch is kept in a closed compartment framed by a solid construction and in materials withstanding seal damage. In recent decades, the grey seal population in the Baltic Sea has increased significantly, resulting in a growing conflict between seals and small-scale fisheries (Lunneryd et al., 2005; Varjopuro, 2011). On the west coast of Sweden, the most common seal species is harbor seal (Phoca vitulina) and their abundance has also increased dramatically to a population size that is larger or equivalent to that in the beginning of the 20th century (Havs och Vattenmyndigheten, 2014; Lunneryd and Königson, 2017). As in the Baltic Sea, the increasing harbor seal population has led to an increasing seal and fisheries conflict, with mainly gillnet fisheries along the west coast being subjected to depredation by harbor seal (Lundström et al., 2010).
Even though the small-scale fisheries off western Sweden are diminishing due to low fish stock biomasses and increased seal depredation (Lunneryd and Königson, 2017), there is a need for finding sustainable and seal-safe gear for a future small-scale fishery, as this provides valuable income to coastal communities. Small-scale pot fisheries are considered to be more sustainable than net fisheries, as they are more selective with regards to reducing the amount of bycatch of mammals and birds and have smaller catches, rather than excessive catches that can be associated with net fisheries (Suuronen et al., 2012). Therefore, developing small-scale pot fisheries as an alternative to net fisheries could have positive benefits for both the exploited stocks and fishers. However, a sustainable small-scale fishery also needs to be economically viable and the effectiveness of an alternative pot gear needs to be demonstrated.
Small-scale fishers are dependent on fishing for multiple target species, having different fishing seasons with different fishing gear. Since both shellfish and cod can be caught in pots, there is a potential to develop gear that can target both shellfish and fish. A multispecies pot fishery would also potentially be a more cost-effective fishery as the same gear can be used to catch different target species, which in turn can eventually prolong the fishing season. If the fishing season is prolonged, there are greater possibilities for adaptive management, taking into account the species life cycle as part of a sustainable fishery. Therefore, to further improve the small-scale fisheries, efforts should be set on developing a pot that can be used for fishing multiple target species such as European lobster, edible crab and cod.
The most common index of abundance in fisheries work is catch per unit effort (CPUE) and it is often assumed that CPUE is proportional to overall population abundance (Stoner, 2004). However, variation in CPUE is affected by multiple factors, including fish availability to the gear (distribution over time and space), the catchability of the gears (intended as the fraction of an available resource that is captured by one unit of effort) and fishing effort (Engås and Løkkeborg, 1994; Arreguín-Sánchez, 1996; Chiarini et al., 2022).
Biotic and abiotic factors influence the catchability of baited gears (He, 2010). Biotic factors can include the abundance of both target species and suitable prey. Abiotic factors include water current speed and direction, water temperature, underwater topography, soak time, stimuli provided to attract the fish, and various features of the pot design. All of these factors may impact pot catchability through their impact on the target species´ behavior, activity levels, feeding motivation and ability to detect, locate or consume the bait (Stoner, 2004). Cod behavior in relation to pots and factors affecting the pot catches have been studied in the southern Baltic (Hedgärde et al., 2016; Ljungberg et al., 2016). Cod catchability in pots can also be explained by cod entry and exit rate (Hedgärde et al., 2016; Ljungberg et al., 2016). Hedgärde et al. (2016) found that the entry rate decreased with an increasing number of cod already in the pot, and the exit rate also increased with the number of cod in the pot. Observations carried out off the coast of New Hampshire, United States, on American lobster (Homarus americanus) behavior around pots, showed that only 6% of the American lobsters that entered the pot were retained and the rest of them escaped mainly through the entrance, suggesting that the design of the pot entrance is important to be able to retain the catch of lobster (Jury et al., 2002). Lobster catch in pots is affected by pot size, bait quantity and quality, soak time, and preventing escape through the entrance (Miller, 1990). However, the highest lobster catches have been observed by facilitating entry and preventing the effect of gear saturation (animals inside traps preventing those outside from entering) (Miller, 1990). A saturation effect occurs when the present catch in a gear reduces its potential for additional catch including both escapement (increased exit rate) and reduced entry (Ovegård et al., 2011). One way of preventing saturation is to include a fish holding chamber within the pot, a so called parlor, keeping the catch away from the entry chamber where the bait bag is most commonly situated. Modifying pots with selection panels or escape vents enabling small fish and lobsters to escape can also prevent saturation since it will only keep the large individuals inside the pot (Ovegård et al., 2011).
The aim of the present study was to compare the catch rate of traditional and newly developed pot design targeting multiple species (i.e European lobster, edible crab and cod), as well as to evaluate biotic and abiotic factors affecting the catch rate. The main question of interest regarding the catch rate (defined as catch per pot per day, CPUE) of the different pots whether CPUE was affected by: i) the type of bait (salted/fresh), ii) the effects of the abundance of one or other of the target species (lobster/edible crab) in the pot on the CPUE of the other target species (lobster, cod and crab), iii) the soak time of the pots, iv) the length (in consecutive days) of the fishing season. Additionally, the analysis seeks to understand whether seal interactions with the pots were related to higher CPUE of one or multiple target species. Quantitatively understanding the variables related to higher catch rates could inform the management of the fishery, in regard to fishing gears to be used and fishing practices.
The pot types used during the trials carried out in 2015, 2016 and 2017 included a traditional lobster pot and seven newly developed multispecies pots. Three of the new pot types were used in 2015 and the remaining four in 2016 and 2017. The new multispecies pot types were developed in collaboration with fishers, researchers at Swedish University of Agricultural Sciences, Department of Aquatic Resources, and a fishing gear manufacturer. The characteristics of the pots are described in Table 1. The names of the pots are based on the characteristics of the pot types (Table 1). The traditional lobster pot (C22OW) was from the Swedish manufacturer Carapax, with a volume of 166 dm3. This pot was used in all areas and in all trials. The frame is made of galvanized round steel covered with a polyethylene black net material with a mesh size of 33 mm. It has two open entrances and a parlor to prevent escapement. The volumes of the newly developed multispecies pots vary from 198 dm3 to 621 dm3. Frames are constructed of galvanized round steel with a covering net material of polyethylene. Mesh size varies from 22 mm to 35 mm between knots. Pot entrances are either open or closed. The closed entrances include a funnel placed vertical or horizontal (Figures 1A–D). Pots with open entrances were equipped with a parlor, which is a second chamber in the pot with entrance from the entrance chamber. None of the pots was equipped with escape openings, in order to allow for analysis of the total unselected catch.
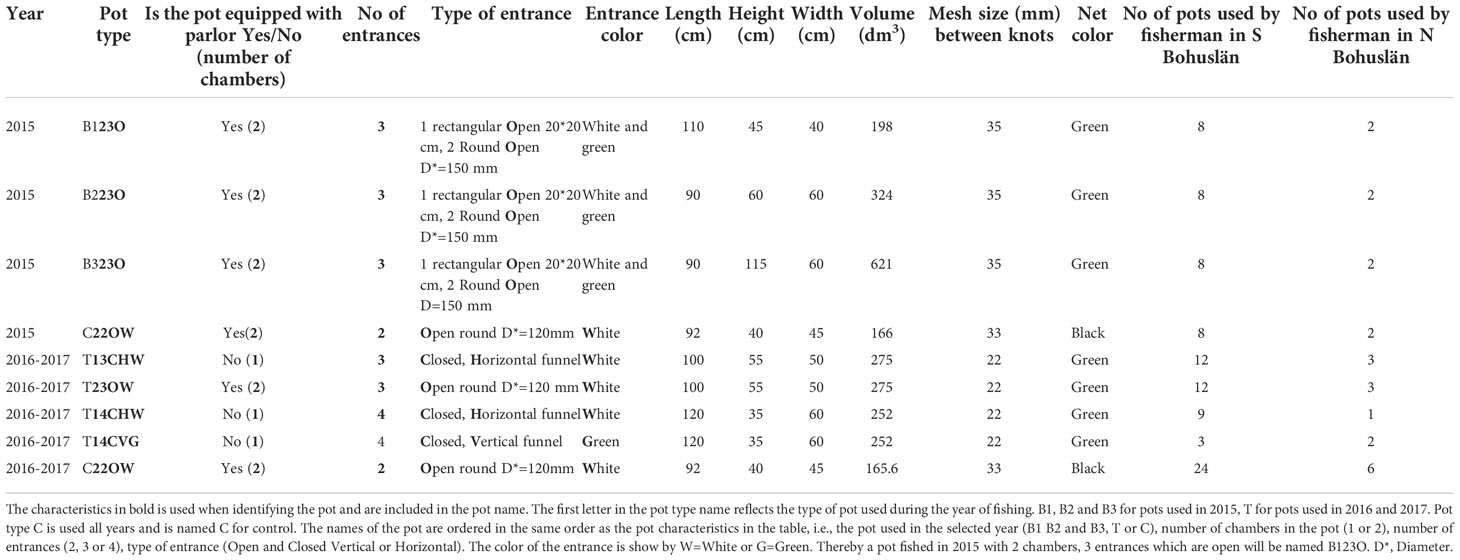
Table 1 The different characteristics of the pots evaluated in the study.

Figure 1 (A) From the left in figure A there is a traditional lobster pot C22OW, the newly developed multispecies pots B123O, B223O and B323O, used in 2015. (B) Pot T13CHW with a closed funnel entrance in white net material. (C) Pot T23OW that has three open entrances. (D) Pot T14CHW, which have four closed, green, vertically placed funnel entrances.
The study was carried out in two areas along the Swedish west coast (Figure 2). In one of these areas (Northern [N] Bohuslän), fishing was conducted by researchers from the Swedish University of Agricultural Sciences, Department of Aquatic Resources, an area just south of the islands of Koster (58°49’ N, 11°02’ E). The second area (Southern [S] Bohuslän) is situated outside the island of Hamburgö (58°32’ N, 11°14’ E), an area with similar conditions as in N Bohuslän in regard to bathymetry and bottom substrate (Figure 2). In this area, the study was undertaken in collaboration with commercial lobster fishers willing to test the newly developed multi-species pot types targeting European lobster, edible crab and cod. The fishing was conducted with the aim of maximizing lobster catch as is normally done in commercial lobster fisheries.
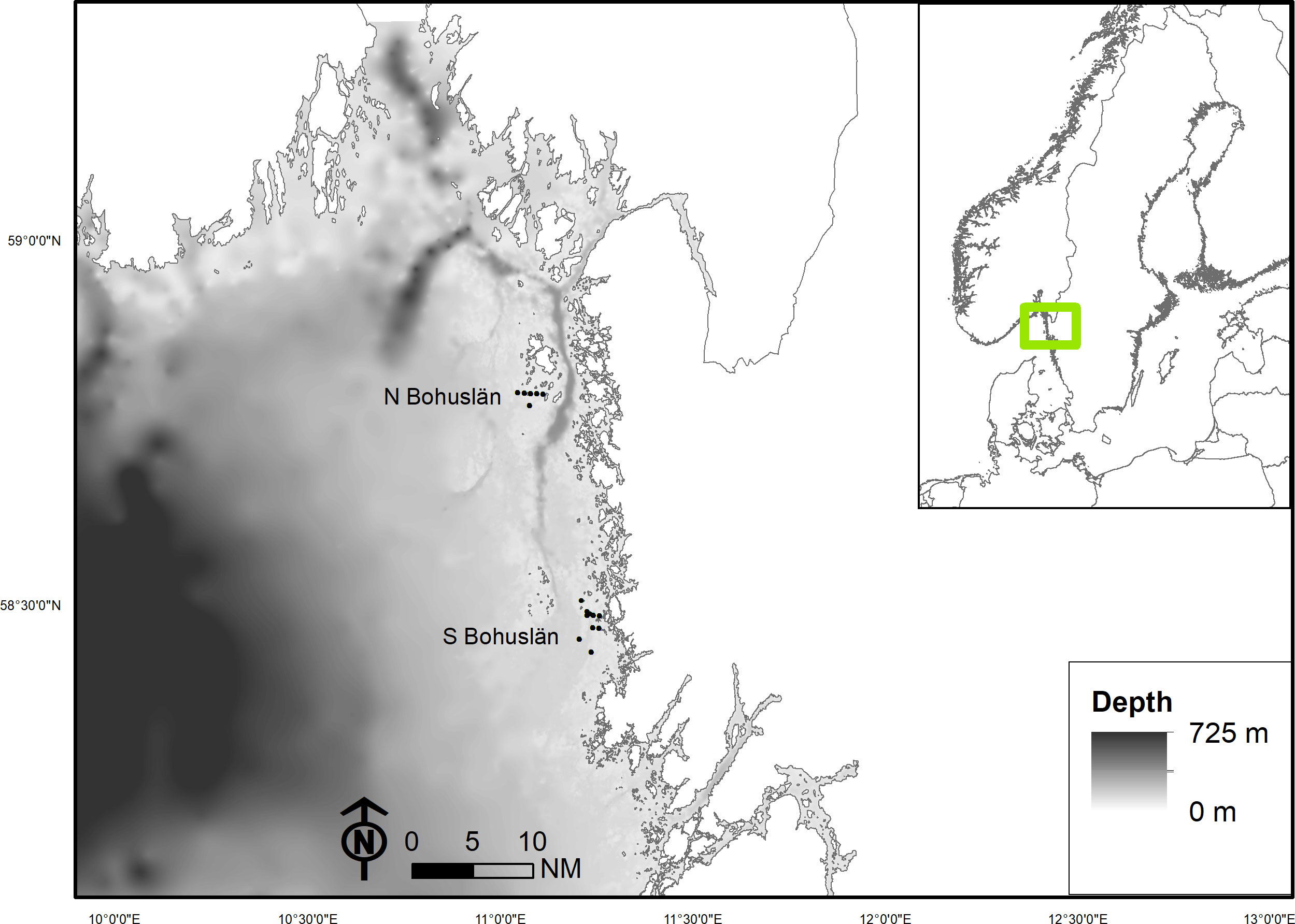
Figure 2 A bathymetric map of the study areas along the west coast of Sweden including the fishing positions. Dots on the map represent the majority of locations where pots were set out.
In N Bohuslän, test fishing started in late September in 2015 and 2016 and continued for two months. In S Bohuslän, the trials also started in late September in both 2015 and 2016 and continued into spring 2016 and 2017, respectively. In 2015, fishing was carried out using four types of pots (one traditional lobster pot and three multispecies pots, Table 1 and Figure 1A). In 2016, four newly developed pot types and a traditional lobster pot were used (Table 1 and Figures 1B–D).
Since fishing was not conducted by commercial fishers in N Bohuslän, the number of pots allowed was restricted to 15 pots (of four different types) in 2015, and 14 (of four different types) in 2016.
In S Bohuslän, in 2015, one of each type of pot was placed in the vicinity of each other. In 2016, five pots of at least four different types were placed in the same area. At every fishing occasion, two traditional lobster pots were placed in the same area.
To increase lobster CPUE, single pots were placed in the zone between soft and hard seabed sediment types as well as on irregular hard bottom types, at an average depth of 20.3 m (95 C.I ± 0.32). All pots were baited with 400 g of cut fresh or salted herring and crushed edible crab. The pots were emptied on average every fourth day (95% C.I ± 0.15). The catch of all cod and lobster was counted and each lobster carapace and cod length was measured to ensure they could be legally retained. Only cod of 30 cm or more in length can be legally retained and for European lobster the minimum carapace length is 90 mm. In addition, female lobster with roe cannot be landed. Since lobster, cod and edible crab constituted the main part of the catch (79% of the catch in N Bohuslän) and were the commercial species of interest, they were the only species reported in S Bohuslän, whereas in N Bohuslän, all caught species were counted and reported. Female lobsters and those smaller than 90 mm in carapace length were released after measuring. Seal damaged fish were also counted; however, only cod was found damaged by seal except for one observation of a damaged sculpin (Cottoidea). Soak time, position, type of bait and seal damage evident on gear or caught fish were noted.
A Generalized Additive Model (GAM) approach was used to analyze the catch rate (CPUE) of different pots in relation to environmental and fisheries variables. GAM is a nonparametric method that allows investigating non-linear relationships between dependent and independent variables (Zuur et al., 2007). GAMs employ a class of equations called smoothers: algorithms which attempt to generalize data into smooth curves by local fitting to subsections of the data (Beck and Jackman, 1998). GAMs have been successfully applied to analyze environmental effects using both categorical and continuous variables (Barton et al., 2020). Similarly, the approach was used in previous studies to explain the variability in catches of cod (Königson et al., 2015) and for investigating the effect of pot type (as a categorical variable) on the pots’ CPUE (Hedgärde et al., 2016)
In our models, the response variable was the mean CPUE of European lobster, cod, or edible crab, and it was treated as a function of several explanatory variables. The explanatory variables chosen were pot type, area where fishing was carried out, soak time, number of days after the experiment started and number of crabs or lobsters in the pot. Different pot designs can affect catches of both target species; soak time has been shown to influence catches and the variation in abundance in different areas can also affect catches (Bennett, 1974; Miller, 1990; Addison, 1995; Königson et al., 2015; Hedgärde et al., 2016). The number of days after the experiment started was chosen to reflect seasonal changes. The number of days after the experiment started can also indicate if there is a possibility to decrease abundance of resident species by continuous fishing in the area. Number of lobster and/or edible crab were included because they are known scavengers and in other studies intraspecific as well as interspecific interactions in pots have shown to affect catch (Addison, 1995). Edible crab are numerous in the area and can potentially fill up the pots.
Multicollinearity of covariates were checked first through the examination of correlation matrix and then by the Variance Inflation Factor (VIF) of the variables. Correlation value above 0.7 (Pallant, 2010; Aminu and Shariff, 2014) and a VIF level above 2.5 (Johnston et al., 2018) were suggested as a threshold for multicollinearity among covariates. In the first step, only temperature was excluded from the model due to its high VIF value that caused multicollinearity (see results). The number of days after the experiment had also a high VIF value but it was preferred to use in the model rather than temperature (see above).
In the next step, for each species, CPUE was modelled as a function of pot type (categorical factor, eight levels), bait (categorical factor, two levels: fresh bait/salted bait), seal damage (only for the cod model, categorical factor, two levels: occurred/not occurred), area (categorical factor, two levels: S Bohuslän/N Bohuslän), and sampling period (categorical factor, four levels: fall 2015/fall 2016/spring 2016/spring 2017), as well as the smoothing function of soak time, number of days after the experiment started, and number of lobster and/or edible crab in the pot.
CPUE data for each species contained many zero observations. Thus, a GAM with a Tweedie distribution was used in our study (Dunn and Gordon, 2005) since this distribution is a potential candidate to suppress the over dispersion of many occasions with zero catch (Shono, 2005). To avoid overfitting the models and to obtain ecologically relevant responses that were easier to interpret, the final models were kept simple (Lehmann et al., 2002; Sandman et al., 2008), with the maximum number of knots for each of the smoothers limited to four (k = 4), allowing the smoother to divide the response from each explanatory variable into a maximum of three parts. A subset of predictors that provided the best fit to the observed data was identified with the Akaike information criterion (AIC; Quinn and Keough, 2002).
Smaller values of AIC indicate more parsimonious models (Quinn and Keough, 2002). The differences in AIC (ΔAIC) among candidate models (Burnham and Anderson, 2004); was computed and the best model will have a ΔAIC of zero (Burnham and Anderson, 2004). For the final model, the percentage of deviance explained by a single smoothing parameter was extracted(xi) (i.e. each covariate) by calculating the difference in deviance explained between models with s(xi) (for example for the first smoothing item, m < gam (y ~ a + s(x1) + s(x2) + s(x3) + s(x4)), and without it (m1 < gam (y ~ a + s(x2) + s(x3) + s(x4), sp=m$sp[-1])), provided that the model parameters in both models are identical. A generalized linear model (GLM) was also performed to test the factors that best predicted the catch efficiency of different pots. For each species, the GLM and GAM models were then compared using adjusted R2, the amount of deviance explained, and plot of the residuals of the each model. The analyses were performed using the mgcv package (Wood, 2001) in the software R (R-Development-Core-Team, 2008).
A total of 2,395 pot emptyings were undertaken during four periods (Table 2). In S Bohuslän, most pots were emptied during the trials in the fall. The mean CPUE was also the highest during these trials for all three species. Edible crab was the most commonly caught species in the pots and thus had the highest mean CPUE (mean 1.05 SD ± 1.70 edible crab per pot) while the mean CPUEs for cod were particularly low in both areas (Table 2). Catches of cod were generally low (mean 0.11 SD ± 0.29 cod per pot) and 65% of the cod were released and had a length less than 30 cm (mean length 26.8 95% CI ± 1.1) length. With regard to European lobster (mean 0.16 SD ± 0.39 lobster per pot), 40% of the catch consisted of individuals over the minimum carapace length of 90 mm and 23% of the catch were females with roe.
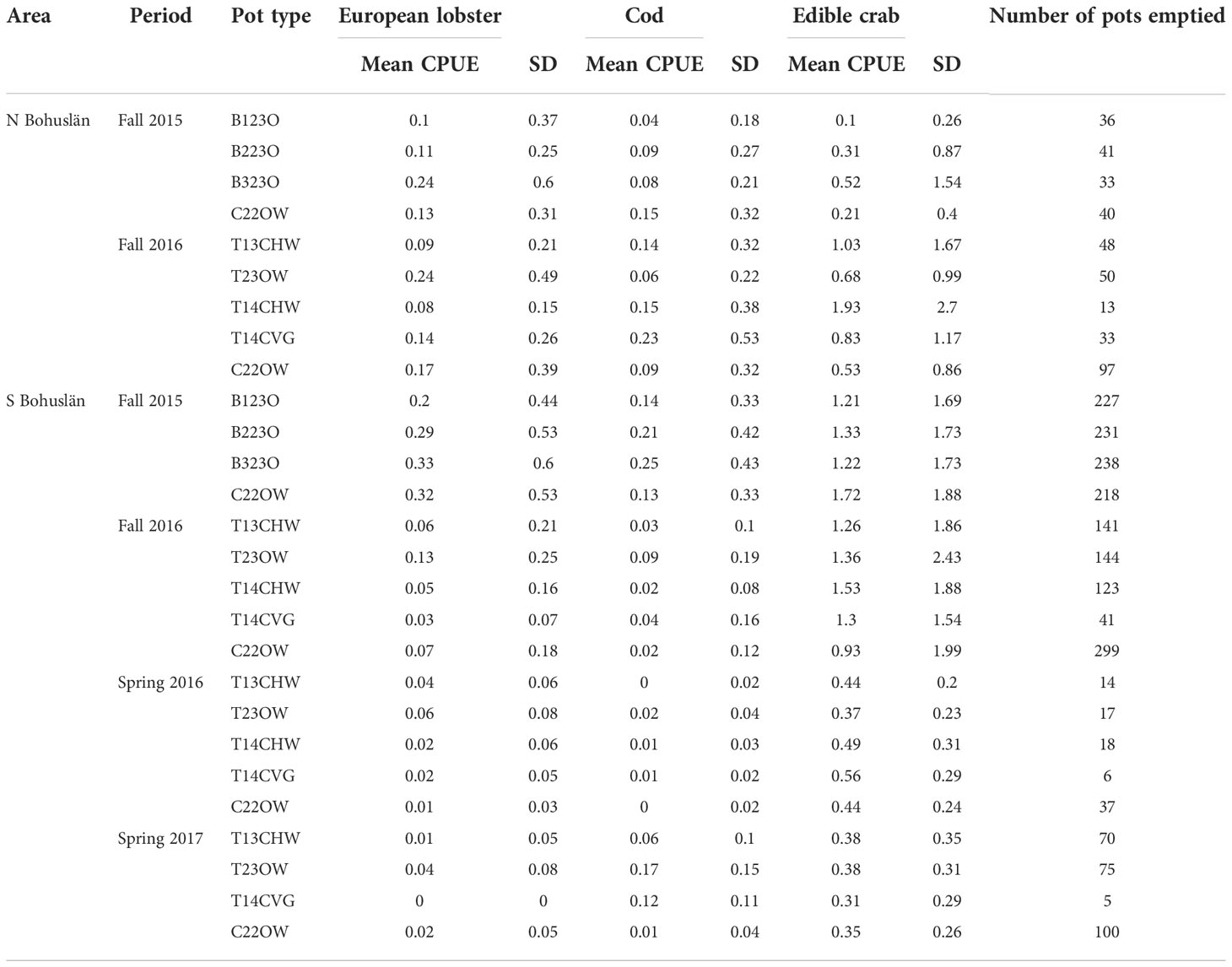
Table 2 CPUE of species caught per pot type for each area and period.
Because the correlation value among covariates was below the threshold value of 0.7, there was no concern about the observed correlation (Table 3). Nevertheless, both temperature and number of days after the experiment had high VIF levels (VIF > 2.5) indicating the existence of multicollinearity. After excluding temperature, VIF values were smaller than 2.5 suggesting that there was weak collinearity among covariates (Table 4).
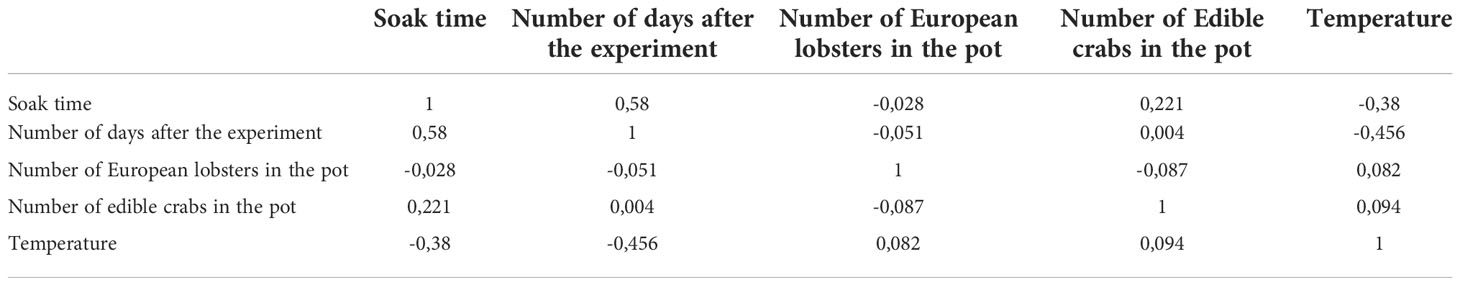
Table 3 Correlation among covariates. Soak time, Number of days after the experiment, Number of European lobsters in the pot, Number of Edible crabs in the pot, Temperature, n = 2395.
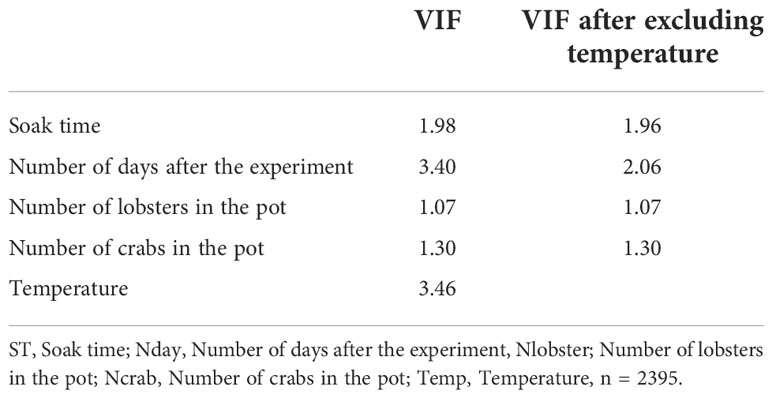
Table 4 Multicollinearity test based on Variance Inflation Factor (VIF).
GAM yielded 8-10 models with different subsets of predictors for CPUE of lobster, cod or edible crab (Table 5). The models with the following predictors had a ΔAIC of zero and provided the best fit to the observed data for each of the target species (Table 5):

Table 5 Best-supported generalized additive models for factors affecting CPUE of cod/European lobster/edible crab.
CPUE of lobster ~ pot type + sampling period + s(soak time)+ s(number of days after the experiment) + s(number of crabs in the pot) (model 1-1)
CPUE of cod~ pot type + bait + seal damage+ sampling period + s(soak time) + s(number of days after the experiment) + s(number of lobsters in the pot) + s(number of crabs in the pot) (model 1-2)
CPUE of crab ~ pot type + area + bait + sampling period + s(soak time) + s (number of days after the experiment) + s(number of lobsters in the pot) (model 1-3)
CPUE of European lobster slightly decreased with increasing number of days after the experiments, soak time, and number of edible crabs in the pot (Figures 3A-C). Both area and bait were excluded from the final model (Table 5). The highest catch rate of European lobster was observed for the T23OW pot (Figure 3D). CPUE of European lobster for B123O was very close to that for T13CHW, T14CHW, and T14CVG but lower than that for T23OW, B223O, B323O, and C22OW (Figure 3D). European Lobster CPUE in fall 2015 was similar to CPUE in spring 2016 but was significantly higher than CPUE in fall 2016 (p < 0.001) and spring 2017 (p < 0.001) (Figure 3E). CPUE in the sampling period of spring 2017 was lower than that in the other three sampling periods (Figure 3E). The final model with selected explanatory predictors, the explained deviance in CPUE data for lobster was 24% (Table 6). Among covariates, the percentages of soak time and number of days after the experiment were higher than “number of crabs in the pot” in explaining the variation in European lobster CPUE (Table 7).
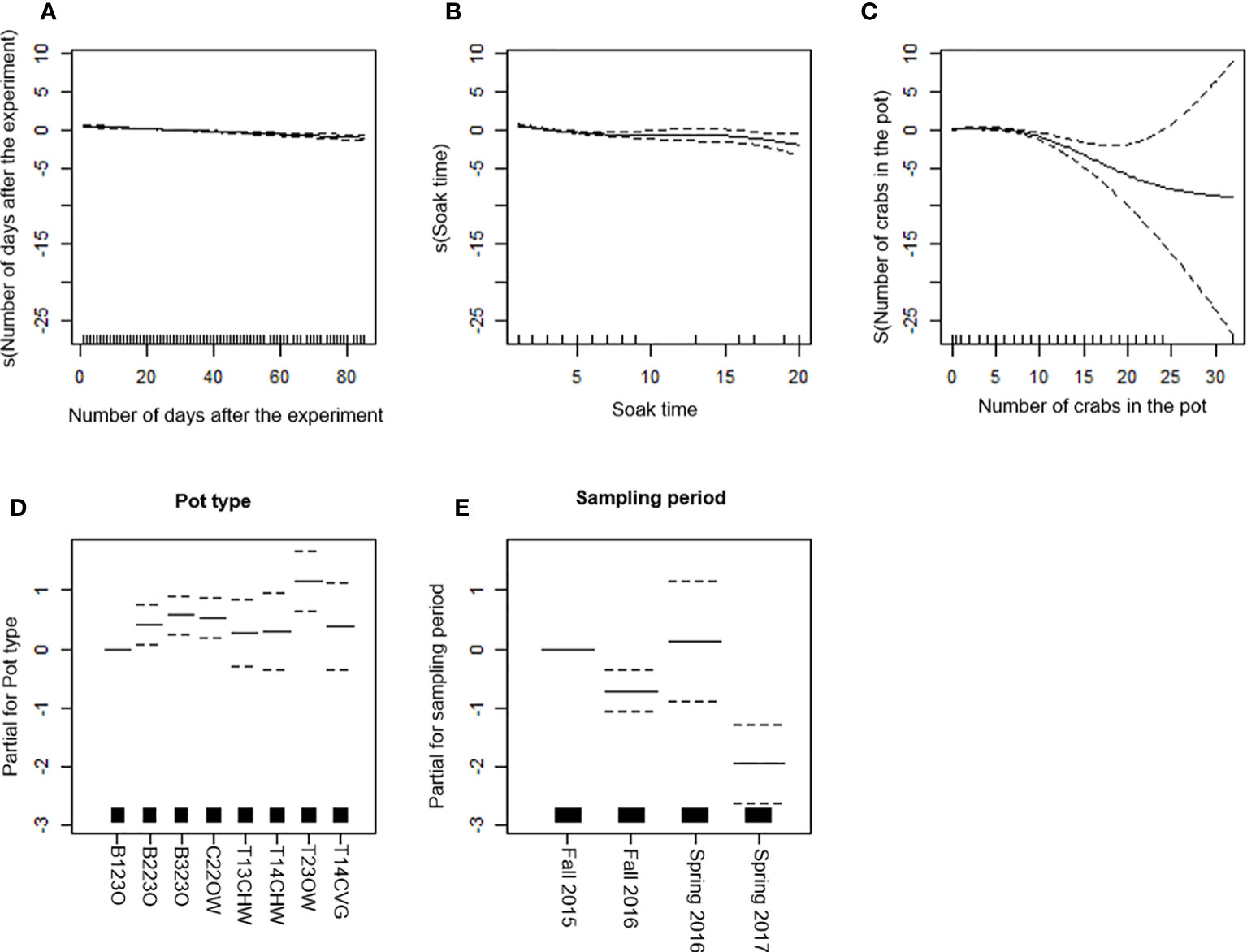
Figure 3 Generalized additive model (GAM) partial dependence plots for European lobster. (A-C) Partial response curves: the x-axis represents the value of the model independent variable whereas the y-axis represents the additive contribution of the variable to the nonparametric GAM smoothing function. Values below zero indicate a negative effect of the variable on the CPUE. (D, E) Partial effects of pot type and sampling period on the CPUE. The dotted line (A–E) represents the 95% confidence interval. The black lines on the x-axis represents sample size.
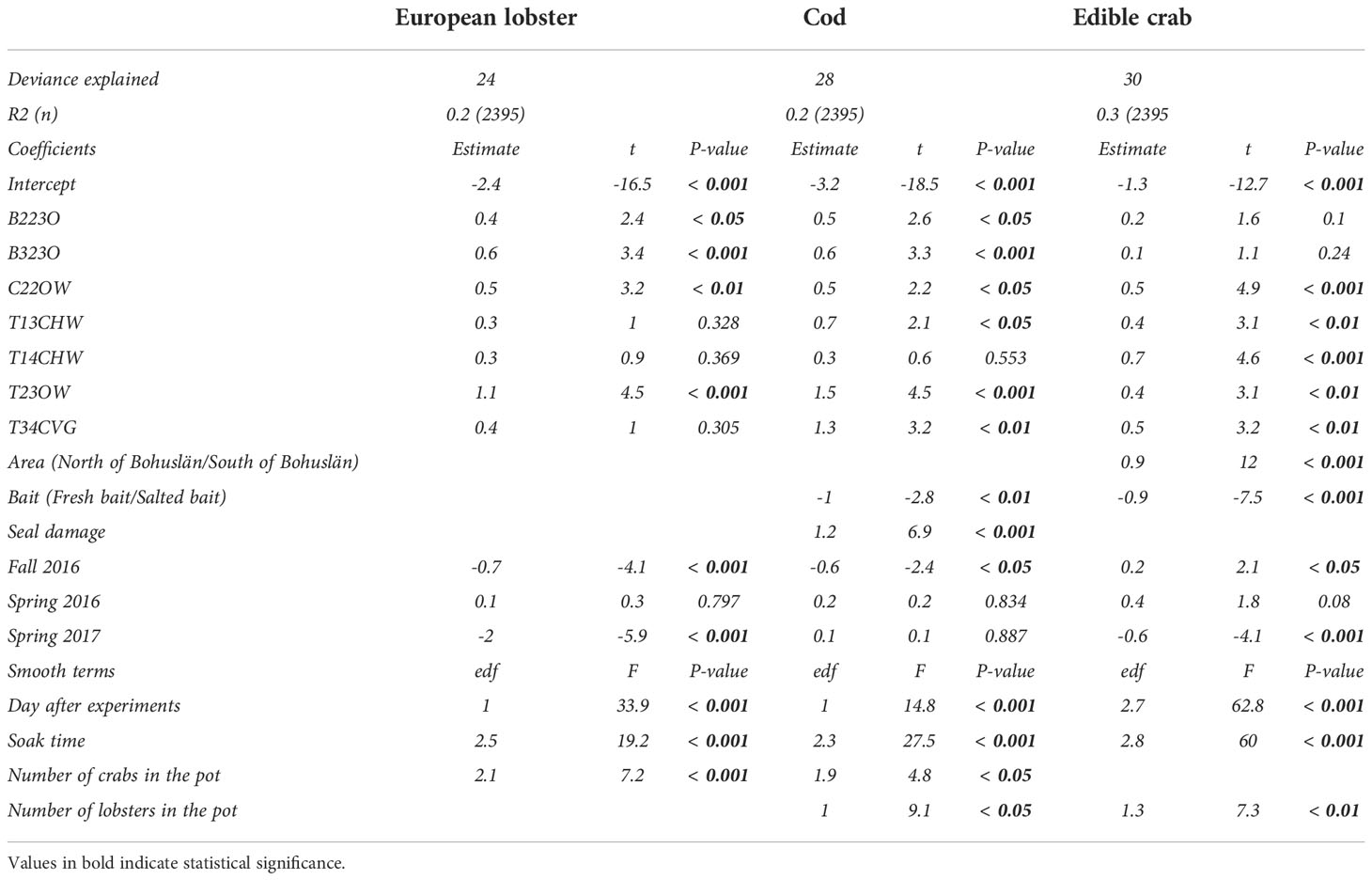
Table 6 Results of the best generalized additive model for CPUE of European lobster, cod and edible crab.

Table 7 Percentage of deviance explained by each covariates in the final generalized additive model for CPUE of lobster, cod, and crab.
Catch rate (CPUE) for cod decreased with increasing number of days after the experiments, soak time, and number of edible crabs and lobsters in the pot (Figures 4A-D). Area was dropped from the final model (Table 5). Among pots, cod CPUE was the highest in T23OW. Pot B123O had significantly lower cod CPUE than all pots except T14CHW (Table 6 and Figure 4E). Catches of cod was higher in pots with fresh bait than in pots with salted bait (Figure 4F). Seal damage to the pot occurred more often when there were more cod in the pot (Figure 4G). Cod CPUE was significantly higher only in the fall of 2015 than the fall of 2016 (Figure 4H). The final model with selected explanatory predictors, the explained deviance in CPUE data for cod was 28% (Table 6). Among covariates, soak time explained the highest percentage of the variation in cod CPUE (Table 7).
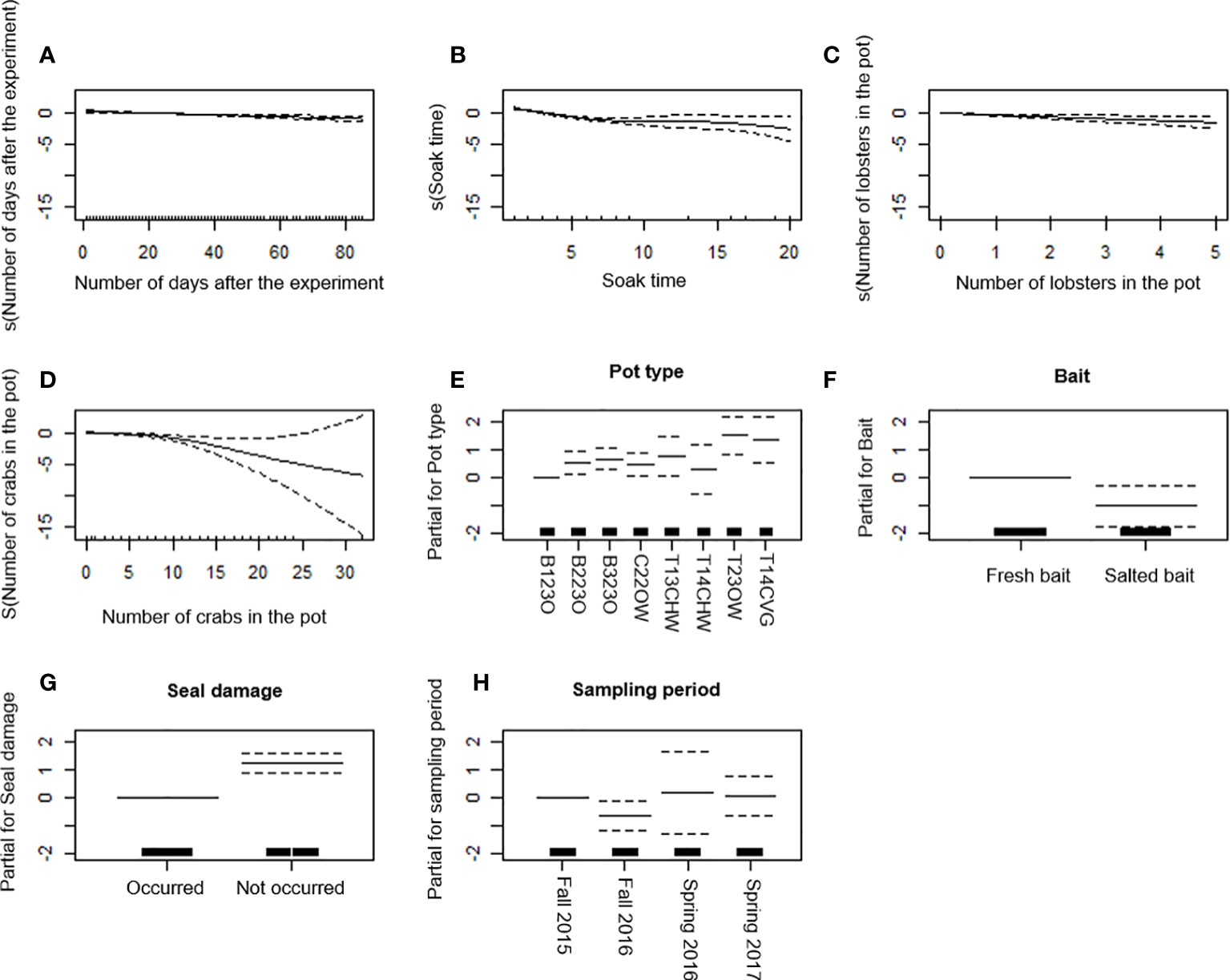
Figure 4 Generalized additive model (GAM) partial dependence plots for cod. (A–D) Partial response curves: the x-axis represents the value of the model independent variable whereas the y-axis represents the additive contribution of the variable to the nonparametric GAM smoothing function. Values below zero indicate a negative effect of the variable on the CPUE. (E–H) Partial effects of pot type, bait, seal damage, and sampling period on the CPUE. The dotted line (A–H) represents the 95% confidence interval. The black lines on the x-axis represents sample size.
There was a strong decreasing trend in CPUE for edible crab with the number of days after the experiment, soak time, and number of lobster in the pot (Figures 5A–C). The T14CHW pot caught the highest number of edible crabs and had a significantly higher CPUE than B123O, B223O and B323O (p < 0.001; Table 6 and Figure 5D). Also, T23OW and T14CVG had a significantly higher CPUE than B123O. The CPUE of edible crab was higher in S Bohuslän than in N Bohuslän (Figure 5E) and higher for pots with fresh bait than the pots with salted bait (Figure 5F). CPUE of edible crab was lower in spring 2017 than during other periods (Table 6 and Figure 5G). The final model with all explanatory predictors, the explained deviance in CPUE for edible crab was 30%. Among covariates, the percentages of soak time and number of days after the experiment were higher than “number of lobsters in the pot” in explaining the variation in edible crab CPUE (Table 7).
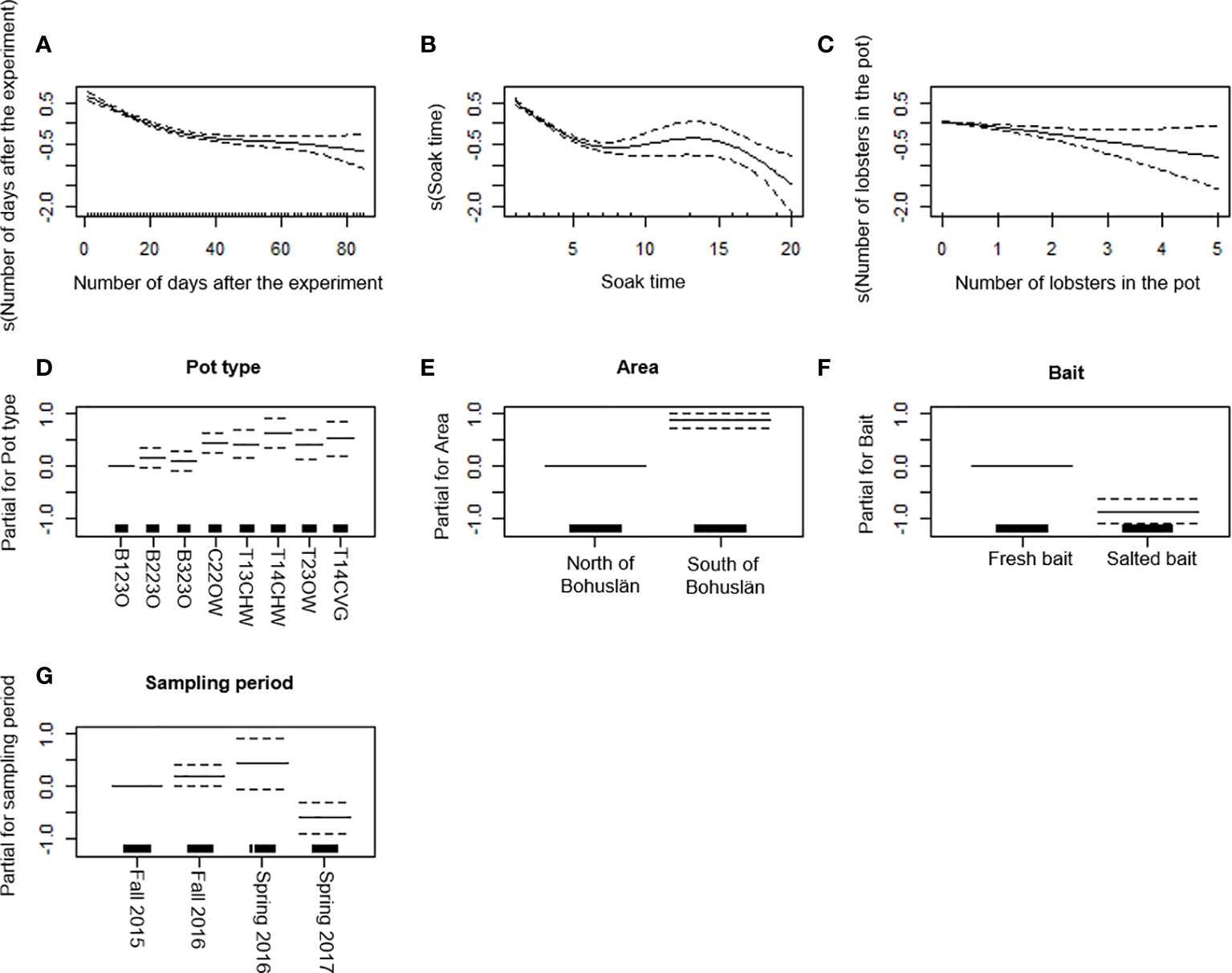
Figure 5 Generalized additive model (GAM) partial dependence plots for edible crab. (A-C) Partial response curves: the x-axis represents the value of the model independent variable whereas the y-axis represents the additive contribution of the variable to the nonparametric GAM smoothing function. Values below zero indicate a negative effect of the variable on the CPUE. (D-G) Partial effects of pot type, area, bait, and sampling period on the CPUE. The dotted line (A-G) represents the 95% confidence interval. The black lines on the x-axis represents sample size.
In the GLM model, adjusted R2 and amount of deviance explained were 0.115 and 12% for European lobster, 0.165 and 17% for cod, and 0.162 and 17% for edible crab, respectively. These values were lower than the corresponding values in the GAM model for all three species (Table 6). In addition, residual plots of each model for each species indicated that the data deviated drastically from the line (unhealthy residual plots) in the GLM model compared to the GAM model (Appendix 1). These results revealed that GAM was a better suited modelling approach than GLM for this dataset.
Commonly, the development of new fisheries focuses on a single target species using one type of gear. However, this study investigated the potential to develop a sustainable commercial pot fishery targeting multiple species such as European lobster, cod and edible crab. Results from this study show that European lobster, cod and crab can indeed be caught in the same gear when the gear is properly designed. Moreover, this study also revealed information on environment and fisheries related parameters that affect the catch of European lobster, cod and edible crab, providing vital information, which is needed when developing a sustainable pot fishery.
The catch of European lobster and cod is affected by pot size, bait quantity and quality, and prevention of escape through the entrance (Miller, 1990). The pot with highest catch rate of European lobster and cod (T23OW) is larger than many of the other pots tested and the parlor of the pot seems to prevent European lobsters from escaping. However, this pot indeed has an open entrance, in contrast to the funnel entrance which effectively prevents escape. Results showed that the pot with open entrances and a parlor increased catch rates compared to similar pots with no parlor. This indicates that there can be a saturation effect, as defined in the introduction. Parlors do not only prevent caught individuals from escaping, but they also prevent species from taking up space in the entrance chamber of the pots. Lovewell et al. (1988) also showed that using two-chambered pots increase the catch efficiency of lobster.
In the pot with highest catch rate of European lobster and cod (T23OW), the open entrance is much smaller than open entrances in the pots used in 2015 (B123O, B223O, B323O) where the entrance had a diameter of 150 mm compared to 120 mm. The participating fishers hypothesized that the entrances in the pots used in 2015 were too large and that the catch easily escaped even though all pots had a parlor. The results also showed that the smallest pot used in 2015 (B123O), with a 150 mm diameter open entrance, had lower catches than a majority of the other pot types with smaller entrances. Furevik and Løkkeborg (1994) found that when pots are equipped with a wider entrance, the entry rates of cod increases but so does also the escape rate.
The medium sized pot with high catches of cod and edible crab has funnels in the entrances preventing the catch to escape (pot T14CVG). Ljungberg et al. (2016), demonstrated that funnel entrances caused cod to hesitate when entering the pot by turning before the funnel. However, the same study also showed that pots equipped with funnel entrances increased catch by preventing escape. The funnel but also the mesh size of the net used in the gear can thus prevent not only the target catch but also undersized catch from escaping.
Munro (1974) showed that the size of a pot can affect the catch with an escape rate being inversely proportional to the volume of the pot when the size of the entrance is being constant. Larger pots generally show a higher CPUE of cod (Furevik and Løkkeborg, 1994; Kindt-Larsen et al., 2022) and the larger the pot, the more increase in saturation effect which has a marked effect on the catch (Miller, 1979). The results in this study also showed that the pot with a rather small volume (198 dm3), a parlor and a large open entrance (diameter of 150 mm) (pot B123O), yielded the lowest catches for all three species. On the other hand, for the largest pot in thestudy (pot B323O), the CPUE of all three species was not higher compared to other pots. This could potentially be explained by a high entrance/escape rate due to large entrance rather than a decreased saturation effect.
The results showed increased CPUE for cod in two pots with comparable size, number of entrances and with no parlor (T14CVG compared to T14CHW), but no increase for lobster and edible crab. The difference between pots was the color and orientation of the entrance, with T14CHW having white vertical entrances. The color of the entrance might have affected the catch rate. White entrances can appear as a contrast in relation to the surroundings. However, single colors become less visible with increasing depth. Regarding the wave length of the light and the depth where the pots were placed (at around 20 meters) there might not be enough light available enabling the fish to distinguish contrasts or certain colors. The orientation of the funnel on the other side can affect the behavior of the catch and in particular fish. A funnel placed horizontally forces the fish to push up the weight of the upper part of the funnel when entering the pot. Ljungberg et al. (2016) showed that funnels can prevent cod from entering the pot due to the close contact with the pot material. A funnel placed vertically enables the fish to enter the pot without having net material in front of the fish obstructing its entry. The results from this study, increased catches of cod in pots with vertical funnels compared to horizontal funnels, supports that observation.
For edible crab, the pot type characteristics do not seem to affect CPUE. All pots, except the two large pots used in 2015 with large open entrances (B223O and B323O), had a higher CPUE of edible crab than B123O. There was no difference in CPUE of edible crabs among the other pot types. The limited maneuverability of the edible crabs prevented them from escaping when they had entered the pot. Thus, having an open entrance, a funnel entrance or a parlor did not affect the entering or escaping of edible crab.
Regarding all three species, CPUE decreased or was stable with increasing soak time, day after the experiment and number of edible crab or European lobster in the pot. CPUE for all species decreased already after the first soak day. Since the CPUE was measured in catch per pot and day (soak time), it is not surprising that CPUE decreased with increased soak time nor that the extracted percentage of deviance explained by a single smoothing parameter indicated that soak time is more important factor explaining the variation in CPUE for all species than the other evaluated factors. Bennett (1974) showed that the maximum catches of lobster (analyzing catch per pot) occurred after a soak time of one day. The CPUE of crabs, however, increased with pot immersion, indicating that the pots continued to yield catches. The difference can be explained by the ability of the lobsters to maneuver compared to crabs and therefore the escape rate of edible crab is most likely not as high as for European lobster. The lower CPUE after an extended soak time observed in the current study could be due to the bait becoming less effective over time; as the scent of the bait diminishes with time. Alternatively, in pots with open entrances, lobsters and cod most likely escaped if they were unable to find their way into the parlor. Königson et al. (2015) found that the optimal soak time in cod pots in the Baltic Sea was six days. In that study, however, CPUE was measured as catch per pot, not taking into account catch per pot and day. If soak time would have been accounted for i.e. catch per pot and day, CPUE would most likely have been continuous the first 6 days in contrast to the findings from this study. In additional studies, carried out in Norwegian waters, no correlation was found between catch per pot of cod and soak time; i.e., the catch rate did not increase with increased soak time (Bjordal and Furevik, 1988; Furevik and Skeide, 2003). In addition, saturation can start limiting catches after only a few hours and its importance in limiting non target species such as edible crab catches in pots is most likely larger than generally appreciated (Miller, 1980).
Regarding the decrease of CPUE of European lobster, cod and edible crab, with increasing numbers of edible crab in the pot (for European lobster and cod) and number of European lobsters in the pot (for cod and edible crab), these results indicate interspecific interactions between species. Addison (1995) studied interspecific interaction between European lobster and edible crab, revealing that lobsters deter edible crabs from entering pots. Similar to the results from this study, decrease in CPUE of European lobster with increasing number of edible lobster in the pot, Addison (1995) also found a negative correlation between the number of European lobsters caught in individual pots and the number of edible crabs in the pot. However, that study could not determine whether it was the increased number of crabs that affected the lobster CPUE or the use of different bait. Importantly, the results from this study lend support to the findings of Addison (1995) that an increased number of edible crab in the pot affects the catch of lobsters negatively and it is not dependent on the bait that is used.
All pots had high catches of edible crab, a scavenger and predator species, and the results also showed a decrease of cod CPUE with increased abundance of edible crab or lobster. This observation can probably be explained by a lack of motivation for cod to enter a pot occupied by predators or scavengers.
The reduction in CPUE for all species over the consecutive days after the experiment started is probably due to the progress of the season and likely fish not being present in the area any more. Water temperatures, which tend to decrease from summer to winter, can affect the presence of the species. For cod, low temperatures reduce cod activity (Brown et al., 1989), thereby reducing the possibility to enter pots and their chances of swimming into the odor stream of the bait. Biological processes of lobster such as being active and thereby becoming accessible to pot fisheries and molting into a harvestable size class (Mills et al., 2017) are processes tightly related to water temperature (Cooper and Uzmann, 1971; Aiken, 1973; Aiken and Waddy, 1975; Ennis, 1984; Crossin et al., 1998). Also, female edible crab are known to migrate over large areas while the males move around more locally searching for females, new habitats, prey or a response to seasonal temperature changes (Ungfors et al., 2007) and this can give seasonal variations in catches. However, even though it was not possible to include temperature in the GAM due to multicollinearity, the changes of temperature can also be reflected in the consecutive days after the experiment started.
The use of bait can have an important role with regard to catch success in creel fisheries (Chapman and Smith, 1978; Siikavuopio et al., 2017). Even so, the type of bait did not have a significant effect on lobster CPUE. However, fresh bait seemed to attract more cod and edible crabs than salted bait. These results are in line with earlier bait choice experiment studies were it has been concluded that that fresh bait is chosen over stale bait (Miller, 1990). However, salted herring is mainly used as bait when fishing with pots targeting European lobster. In pot fisheries targeting cod, fresh herring have shown to be the most effective bait compared to shrimp and squid when used in the Baltic Sea cod pot fishery (Ljungberg, 2007). Both fresh and salted bait was added with crushed edible crab since it is known among fishers that baiting with edible crab reduces the catch of edible crab. Since the fishery was conducted during the main lobster fishing season, a reduction in edible crabs in the pot was preferred to increases in catches of lobster even though edible crab can also be seen as a commercial target catch. The knowledge of fishers regarding edible crab as bait relates well with the results of Moore and Howarth (1996) who found that added crushed and substantially fractured shore crab (Carcinus maenas) significantly reduced the capture of conspecifics. Similarly, Chapman and Smith (1978) revealed a reduction in catches of edible crab when pots were baited with crushed edible crab.
Only on a few occasions, pots were subjected to seal damage. Seal damages were more common in pots with a relatively high CPUE of cod and did not occur in pots with no cod or low cod CPUE, indicating that seals prefer cod to the other target species and raid pots more often if there are cod inside the pot compared to when there is no or little cod in the pot. Preference among harbor seals to raid fishing gear containing favorable prey for seals have been found in the eel fishery with fyke nets (Königson et al., 2006).
The environment and fisheries related parameters explained the variance in CPUE by 24 to 30% depending on species. This is similar to the explanatory power of GAMs used in other similar studies; for example Mitchell et al. (2014) reported that environmental variables explained 30% of the deviance in a model testing relationships between environmental variables and CPUE of the blue shark Prionace glauca in the western English Channel. Other factors that could affect pot catches include water current, pot placement in relation to the current, possible prey abundance in the area, and individual behavior. These factors, and possibly other unknown factors, could affect catch rate but have not been quantified and were not available for inclusion in the model.
To be able to implement the widespread use of a new, more sustainable gear it is important to first evaluate if the relevant fishery can be cost effective using the new gear. Therefore, this study has focused on evaluating pot catch rate for multiple species. Increasing pot catch rates, decreasing seal depredation, using the same gear for multiple species, as well as prolonging the fishing season, can help increase the uptake of new sustainable pots in fisheries. However, prolonging the fishing season can potentially have other consequences (e.g. extending fishing into spawning periods when lobsters are more vulnerable) and should be thoroughly evaluated before being considered as an option to compensate for reduced in-season catches that may result from using a new gear. For lobster and for crab, CPUE was most strongly related to the number of days after the experiment started. This can indicate that species abundance might actually decrease with continuous fishing pressure in the area, thereby prolonging the fishing season could have a negative impact on the species abundance. Extending the fishing season can also be detrimental to other life cycle stages but this can partly be solved by requiring all pots to have escape openings or panels dependent on the targeted species for the time. Today European lobster fisheries are regulated by: seasonal opening and closure of the fishery, limitations on gear (must have escape openings), size and condition limits (minimum size of 90 mm and no roe) as well as limitation on the number of gears that can be used. Cod fisheries along the Swedish west coast are also limited by quotas, time and area closures and regulation of gears by only allowing certain mesh sizes at specific depths.
For this study, the CPUE of the total number of European lobster, cod and crab was chosen as a variable in the GAM, and size of the species was not taken in consideration. However, when evaluating the catch composition, the caught cod was found to be mainly undersized, which indicates that there is a need for including escape panels to prevent undersized cod from being caught. If needed, in pot fisheries, it is also possible to restrict the entry of large cod by limiting the size of the entrance.
Altogether, results from the study shows the possibility for a fishery targeting several species using the same multispecies pots. The fishing season could technically be prolonged over several fishing seasons, however this might increase the impact on populations of target species. The study also revealed information on which pot characteristics could influence CPUE of the three target species. If for example when targeting European lobster and cod, a large pot with two open entrances and a parlor generates the highest CPUE. Pots with entrances equipped with funnels, preventing cod from escaping, also yielded a high cod CPUE. Instead edible crab CPUE, was not affected by pot type. The results from this study could be used to further improve and develop an adaptive and sustainably small-scale fishery using selective pots.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The study was carried out in collaboration with a commercial and recreational fishery and therefore there is no ethic statement needed.
SK and S-GL did the conception and design of the study. RN performed the statistical analyses. PL wrote sections in the manuscript. KM and AB revised the manuscript. All authors contributed to the article and approved the submitted version.
Special thanks to Henrik Björklund who has been an excellent and collaborative fisher carrying out test fishing with the multispecies pots. Special thanks to Lachlan Fetterplace for reviewing the manuscript and the reviewers who provided many helpful comments, which considerably improved the paper. The study was funded by Program Seals and Fisheries, Swedish Agency for Marine and Water Management and EU.
Author KM was employed by HiDef Aerial Surveying Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.985431/full#supplementary-material
Addison J. T. (1995). “Influence of behavioural interactions on lobster distribution and abundance as inferred from pot-caught samples,” in ICES marine science symposia (. Copenhagen, Denmark: International Council for the Exploration of the Sea), 294–300.
Aiken D. (1973). Proecdysis, setal development, and molt prediction in the American lobster (Homarus americanus). J. Fisheries Board Canada 30, 1337–1344. doi: 10.1139/f73-214
Aiken D., Waddy S. (1975). Temperature increase can cause hyperecdysonism in American lobsters (Homarus americanus) injected with ecdysterone. J. Fisheries Board Canada 32, 1843–1845. doi: 10.1139/f75-218
Aminu I. M., Shariff M. (2014). Strategic orientation, access to finance, business 500 environment and SMEs performance in Nigeria: Data screening and preliminary analysis. Eur. J. Business Manage. 6 (35), 124–132. doi: 10.5539/ass.v11n4p2981
Arreguín-Sánchez F. (1996). Catchability: A key parameter for fish stock assessment. Rev. Fish Biol. Fisheries 6, 221–242. doi: 10.1007/BF00182344
Barton N. A., Farewell T. S., Hallett S. H. (2020). Using generalized additive models to investigate the environmental effects on pipe failure in clean water networks. NPJ Clean Water 3, 31. doi: 10.1038/s41545-020-0077-3
Beck N., Jackman S. (1998). Beyond linearity by default: Generalized additive models. Am. J. Political Sci. 42, 596–627. doi: 10.2307/2991772
Bennett D. (1974). The effects of pot immersion time on catches of crabs, cancer pagurus l. and lobsters, homarus gammarus (L.). ICES J. Mar. Sci. 35, 332–336. doi: 10.1093/icesjms/35.3.332
Bergenius M., Ringdahl K., Sundelöf A., Carlshamre S., Wennhage H., Valentinsson D. (2018). Atlas över svenskt kust- och havsfiske 2003-205. Aquareports 2018, 3.
Bjordal Å., Furevik D. M. (1988). Full scale fishing trials for tusk (Brosme brosme) and cod (Gadus morhua) with a collapsible fish trap (ICES) 1988/B:33 (1988), p. 10.
Brown J. A., Pepin P., Methven D. A., Somerton D. C. (1989). The feeding, growth and behavior of juvenile cod, gadus-morhua l in cold environments. J. Fish Biol. 35, 373–380. doi: 10.1111/j.1095-8649.1989.tb02989.x
Burnham K. P., Anderson D. R. (2004). Multimodel inference - understanding AIC and BIC in model selection. Sociological Methods Res. 33, 261–304. doi: 10.1177/0049124104268644
Chapman C., Smith G. (1978). Creel catches of crab, cancer pagurus l. using different baits. ICES J. Mar. Sci. 38, 226–229. doi: 10.1093/icesjms/38.2.226
Chiarini M., Guicciardi S., Angelini S., Tuck I. D., Grilli F., Penna P., et al. (2022). Accounting for environmental and fishery management factors when standardizing CPUE data from a scientific survey: A case study for nephrops norvegicus in the pomo pits area (Central Adriatic Sea). PloS One 17, e0270703. doi: 10.1371/journal.pone.0270703
Cooper R. A., Uzmann J. R. (1971). Migrations and growth of deep-sea lobsters, homarus americanus. Science 171, 288–290. doi: 10.1126/science.171.3968.288
Crossin G. T., Al-Ayoub S. A., Jury S. H., Howell W. H., Watson W. (1998). Behavioral thermoregulation in the American lobster homarus americanus. J. Exp. Biol. 201, 365–374. doi: 10.1242/jeb.201.3.365
De Wit P., Svanberg L., Casties I., Eriksson S., Sundell K., André C., et al. (2021). Single nucleotide polymorphisms are suitable for assessing 525 the success of restocking efforts of the European lobster (Homarus gammarus, l.). 526 Conserv. Genet. Resour. 14, 47–52. doi: 10.1007/s12686-021-01234-3
Dunn P. K., Gordon K. S. (2005). Series evaluation of tweedie exponential dispersion model densities. Stat. Computing 15, 267–280. doi: 10.1007/s11222-005-4070-y
Engås A., Løkkeborg S. (1994). “Abundance estimation using bottom gillnet and long-line — the role of fish behaviour”, in Marine fish behaviour in capture and abundance estimation vol. 221 . Eds. Ferno A., Olsen S. (Oxford: Fishing News Books).
Ennis G. (1984). Small-scale seasonal movements of the American lobster homarus americanus. Trans. Am. Fisheries Soc. 113, 336–338. doi: 10.1577/1548-8659(1984)113<336:SSMOTA>2.0.CO;2
Furevik D. M., Løkkeborg S. (1994). Fishing trials in Norway for torsk (Brosme brosme) and cod (Gadus morhua) using baited commercial pots. Fisheries Res. 19, 219–229. doi: 10.1016/0165-7836(94)90040-X
Furevik D., Skeide R. (2003). Fishing for cod (Gadus morhua), ling (Molva molva) and tusk (Brosme brosme) using two-chamber pots along the Norwegian coast. Fangstseksjonen, Havforskningsinstituttet. Fisken og Havet 9.
Havs och Vattenmyndigheten (2014). “Sälpopulationernas tillväxt och utbredning samt effekterna av sälskador i fisket,” in Redovisning av ett regeringsuppdrag. havsoch vattenmyndighetens rapport 2014 (Swedish).
He P. (2010). Behavior of marine fishes: Capture processes and conservation challenges (Oxford, UK: Wiley-Blackwell).
Hedgärde M., Berg C. W., Kindt-Larsen L., Lunneryd S. G., Königson S. (2016). Explaining the catch efficiency of different cod pots using underwater video to observe cod entry and exit behaviour. J. Ocean Technol. 11, 67–90.
Hentati Sundberg J. (2017). Svenskt fiske i historiens ljus en historisk fiskeriatlas. Aqua reports (Sveriges lantbruksuniversitet: Institutionen förakvatiska resurser, Lysekil) 2017, 4.
Johnston R., Jones K., Manley D. (2018). Confounding and collinearity in regression analysis: a cautionary tale and an alternative procedure, illustrated by studies of British voting behaviour. Qual Quant 52 (4), 1957–1976. doi: 10.1007/s11135-017-0584-6
Jury S. H., Howell H., O'Grady D. F., Watson I. I. I., W. H. (2002). Lobster trap video: In situ video surveillance of the behaviour of homarus americanus in and around traps. Mar. Freshw. Res. 52, 1125–1132. doi: 10.1071/MF01096
Kindt-Larsen L., Willestofte Berg C., Hedgärde, Königson S. (2022). Avoiding grey seal depredation in the baltic sea while increasing catch rates of cod. Accepted Fisheries Res.
Königson S. J., Fredriksson R. E., Lunneryd S.-G., Strömberg P., Bergström U. M. (2015). Cod pots in a Baltic fishery: are they efficient and what affects their efficiency? ICES J. Mar. Science: J. du Conseil 72, 1545–1554. doi: 10.1093/icesjms/fsu230
Königson S., Lundström K., Hemmingsson M., Lunneryd S.-G., Westerberg H. (2006). Feeding preferences of harbour seals (Phoca vitulina) specialised in raiding of fishing gear aquatic mammals, Vol. 32. 152–156.
Lehmann A., Overton J. M., Leathwick J. R. (2002). GRASP: generalized regression analysis and spatial prediction. Ecol. Model. 157, 189–207. doi: 10.1016/S0304-3800(02)00195-3
Ljungberg P. (2007). Evaluation of baited pots in the fishery for cod (Gadus morhua) within the southeast Baltic (Lund, Schweden: Department of Biology, Lund University).
Ljungberg P., Lunneryd S.-G., Lövgren J., Königson S. (2016). Including cod (Gadus morhua) behavioural analysis to evaluate entrance type dependent pot catch in the Baltic Sea. J. Ocean Technol. 11, 48–63.
Lovewell S. R., Howard A. E., Bennett D. B. (1988). The effectiveness of parlour pots for catching lobsters (homarus gammarus (L.)) and crabs (cancer pagurus l.). ICES J. Mar. Sci. 44, 247–252. doi: 10.1093/icesjms/44.3.247
Lundström K., Lunneryd S.-G., Königson K., Hemmingsson M. (2010). Interactions between harbour seals (Phoca vitulina) and coastal fisheries along the Swedish west coast: an overview (Tromsö: NAMMCO Scientific Publications) 8, 8. doi: 10.7557/3.2697
Lunneryd S. G., Hemmingsson M., Tärnlund S., Fjälling A. (2005). A voluntary logbook scheme as a method of monitoring the by-catch of seals in Swedish coastal fisheries (Köpenhagen, Denmark: International Council for the Exploration of the Sea), 10.
Lunneryd S.-G., Königson S. (2017). Hur löser vi konflikten mellan säl och kustfiske. Program Sälar och Fiskes verksamhet från and 1994 till 2017. Aqua reports (Drottningholm Lysekil Öregrund: Sveriges lantbruksuniversitet, Institutionen för akvatiska resurser), 9.
Miller R. J. (1979). Saturation of crab traps - reduced entry and escapement. J. Du Conseil. 38, 338–345. doi: 10.1093/icesjms/38.3.338
Miller R. J. (1980). Design criteria for crab traps. J. Du Conseil 39, 140–147. doi: 10.1093/icesjms/39.2.140
Miller R. J. (1990). Effectiveness of crab and lobster traps. Can. J. Fisheries Aquat. Sci. 47, 1228–1251. doi: 10.1139/f90-143
Mills K. E., Pershing A. J., Hernández C. M. (2017). Forecasting the seasonal timing of maine's lobster fishery. Front. Mar. Sci. 4. doi: 10.3389/fmars.2017.00337
Mitchell J. D., Collins K. J., Miller P. I., Suberg L. A. (2014). Quantifying the impact of environmental variables upon catch per unit effort of the blue shark prionace glauca in the western English channel. J. Fish Biol. 85, 657–670. doi: 10.1111/jfb.12448
Moore P., Howarth J. (1996). Foraging by marine scavengers: effects of relatedness, bait damage and hunger. J. Sea Res. 36, 267–273. doi: 10.1016/S1385-1101(96)90795-9
Munro J. L. (1974). The mode of operation of antillean fish traps and the relationships between ingress, escapement, catch and soak. ICES J. Mar. Sci.35 pp, 337–350.
Ovegård M., Königson S., Persson A., Lunneryd S. G. (2011). Size selective capture of Atlantic cod (Gadus morhua) in floating pots. Fisheries Res. 107, 239–244. doi: 10.1016/j.fishres.2010.10.023
Pallant J. (2010). SPSS Survival manual: A step by step guide to data analysis using SPSS. 4th ed (Australia: Allen & Unwin Book Publishers).
Quinn G. P., Keough M. J. (2002). Experimental design and data analysis for biologists (Cambridge: Cambridge University Press).
R-Development-Core-Team (2008). “R: A language and environment for statistical computing,” in R foundation for statistical computing (Vienna, Austria).
Sandman A., Isaeus M., Bergstrom U., Kautsky H. (2008). Spatial predictions of Baltic phytobenthic communities: Measuring robustness of generalized additive models based on transect data. J. Mar. Syst. 74, S86–S96. doi: 10.1016/j.jmarsys.2008.03.028
Shono H. (2005). Is model selection using akaike's information criterion appropriate for catch per unit effort standardization in large samples? Fisheries Sci. 71, 978–986. doi: 10.1111/j.1444-2906.2005.01054.x
Siikavuopio S. I., Dragøy Whitaker R., Martinsen G., Saether B. S., Stormo S. K. (2017). Testing baits prepared from by-product of the shrimp and snow crab industry in the pot fishery for gadus morhua (Linnaeus 1758) and pollachius virens (Linnaeus 1758). J. Appl. Ichthyology 33, 1153–1157. doi: 10.1111/jai.13468
Stoner A. W. (2004). Effects of environmental variables on fish feeding ecology: implications for the performance of baited fishing gear and stock assessment. J. Fish Biol. 65, 1445–1471. doi: 10.1111/j.0022-1112.2004.00593.x
Suuronen P., Chopin F., Glass C., Løkkeborg S., Matsushita Y., Queirolo D., et al. (2012). Low impact and fuel efficient fishing–looking beyond the horizon. Fisheries Res. 119-120, 135–146. doi: 10.1016/j.fishres.2011.12.009
Ungfors A., Hallbäck H., Nilsson P. (2007). Movement of adult edible crab (Cancer pagurus l.) at the Swedish West coast by mark-recapture and acoustic tracking. Fisheries Res. 84, 345–357. doi: 10.1016/j.fishres.2006.11.031
Varjopuro R. (2011). Co-Existence of seals and fisheries? adaptation of a coastal fishery for recovery of the Baltic grey seal. Mar. Policy 35, 450–456. doi: 10.1016/j.marpol.2010.10.023
Keywords: pot fishery, sustainable fishery, Atlantic cod, edible crab, European lobster, catch rate, Generalized Additive Model (GAM), Catch per unit effort CPUE
Citation: Königson S, Naddafi R, Lunneryd S-G, Bryhn AC, Macleod K and Ljungberg P (2022) Effects of fishery and environmental factors on a novel multispecies pot targeting European lobster (Homarus gammarus), Atlantic cod (Gadus morhua) and edible crab (Cancer pagurus). Front. Mar. Sci. 9:985431. doi: 10.3389/fmars.2022.985431
Received: 03 July 2022; Accepted: 22 November 2022;
Published: 20 December 2022.
Edited by:
Valentina Melli, Technical University of Denmark, DenmarkReviewed by:
Michela Martinelli, National Research Council (CNR), ItalyCopyright © 2022 Königson, Naddafi, Lunneryd, Bryhn, Macleod and Ljungberg. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sara Königson, c2FyYS5rb25pZ3NvbkBzbHUuc2U=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.