
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Mar. Sci. , 07 October 2022
Sec. Coral Reef Research
Volume 9 - 2022 | https://doi.org/10.3389/fmars.2022.952199
This article is part of the Research Topic Stony Coral Tissue Loss Disease in the Caribbean View all 50 articles
 Nikki Traylor-Knowles1*
Nikki Traylor-Knowles1* Andrew C. Baker1
Andrew C. Baker1 Kelsey M. Beavers2
Kelsey M. Beavers2 Neha Garg3
Neha Garg3 Jeffrey R. Guyon4
Jeffrey R. Guyon4 Aine Hawthorn5
Aine Hawthorn5 Nicholas J. MacKnight2Mónica Medina6
Nicholas J. MacKnight2Mónica Medina6 Laura D. Mydlarz2Esther C. Peters7Julia Marie Stewart6
Laura D. Mydlarz2Esther C. Peters7Julia Marie Stewart6 Michael S. Studivan8,9
Michael S. Studivan8,9 Joshua D. Voss10
Joshua D. Voss10Coral disease has progressively become one of the most pressing issues affecting coral reef survival. In the last 50 years, several reefs throughout the Caribbean have been severely impacted by increased frequency and intensity of disease outbreaks leading to coral death. A recent example of this is stony coral tissue loss disease which has quickly spread throughout the Caribbean, devastating coral reef ecosystems. Emerging from these disease outbreaks has been a coordinated research response that often integrates ‘omics techniques to better understand the coral immune system. ‘Omics techniques encompass a wide range of technologies used to identify large scale gene, DNA, metabolite, and protein expression. In this review, we discuss what is known about coral immunity and coral disease from an ‘omics perspective. We reflect on the development of biomarkers and discuss ways in which coral disease experiments to test immunity can be improved. Lastly, we consider how existing data can be better leveraged to combat future coral disease outbreaks.
Since the 1970s, the rate, intensity, and recurrence of coral diseases have been well documented, particularly in the Caribbean, where coral cover has declined by at least 66–80% since the 1970s from absolute coral cover of ~50% to 10–17% (Gardner et al., 2003; Schutte et al., 2010; Jackson et al., 2014). While the causative agents and the primary mechanisms of these diseases are unknown, local stressors, such as increasing nutrients, pollution, and global stressors, such as increasing temperatures, have likely contributed to the increase in the frequency and severity of disease outbreaks (Harvell et al., 2002; Peters, 2015; Vega Thurber et al., 2020). These compounded stressors, many caused by human impacts, have created challenging and often inhospitable environments for corals to thrive. This has led to the need for more diagnostic tools to assess reef health to assist coral restoration practitioners and managers. The relatively recent development of ‘omics techniques such as transcriptomics (gene expression), proteomics (protein expression), and metabolomics (metabolite expression) hold promise for the development of such diagnostic tools. However, in practice, the development of useful biomarkers to assess coral health is challenging, and the speed with which new diseases are emerging and spreading has motivated researchers to identify alternative approaches to assist in assessing coral health.
Stony coral tissue loss disease (SCTLD) is an example of a newly emerged disease whose severity and rapid spread have caused researchers to seek novel ‘omics approaches to understand, diagnose, and mitigate this deadly outbreak. First documented in 2014 on corals near the Port of Miami dredging project, it spread quickly throughout Florida’s Coral Reef (Precht et al., 2016; NOAA, 2018; Dobbelaere et al., 2020; Muller et al., 2020; Roth et al., 2020; Sharp et al., 2020) and subsequently to other parts of the northern Caribbean, including Mexico, the U.S. Virgin Islands, Puerto Rico, and Turks and Caicos (Alvarez-Filip et al., 2019; Estrada-Saldívar et al., 2020; Roth et al., 2020; Dahlgren et al., 2021; Heres et al., 2021; Meiling et al., 2021). SCTLD is known to affect more than 22 different coral species, and its rapid spread through the Caribbean is suspected to be linked in part to ballast-water releases and human activities (Rosenau et al., 2021). The ongoing spread of SCTLD, the impacts of which have yet to be incorporated into region-wide assessments of coral cover, illustrates the rapidly changing disease seascape to which coral reefs are exposed and the dire ecological consequences as new disease spreads across multiple species.
Biomarkers are critical tools used to assess and monitor health in a myriad of organisms. Typically used for early detection and diagnosis, prognosis, and monitoring of disease treatments (Duan et al., 2005; Henry and Hayes, 2012; Kourou et al., 2015; Meyer et al., 2021), their use has been a cornerstone of most modern medical interventions. Within coral ‘omics research, the use of biomarkers is often proposed as one of the primary goals to understand coral disease, but their full utility is still evolving. Corals are holobionts that consist of coevolved relationships with the coral host, dinoflagellates, bacteria, viruses, fungi, archaea, and other macro-eukaryotes (Rohwer et al., 2002). The relationship between the coral hosts and the different holobiont components is considered complex because (1) there are many different partnerships-most of which are not well characterized, (2) there is variability in the types of micro-organisms involved that varies depending on the coral species, the historical environmental conditions, and current environmental conditions, and (3) we do not know how these relationships are maintained, but the immune system is hypothesized to play a role in this. ‘Omics methods such as transcriptomics, proteomics, and metabolomics of the coral host have been developed into to assess the range of factors involved in coral disease, including current and past environmental conditions, the variable holobiont composition, and overall coral host health (Montilla et al., 2019; Vega Thurber et al., 2020) (Table 1), but experimental limitations remain a challenge in identifying the most appropriate and informative biomarkers of coral disease. This process is particularly impacted by the many pathways and molecular responses involved in coral immunity. Critical efforts to connect coral immune function with its response to disease have begun (Vega Thurber et al., 2020), but challenges identifying signatures of disease at a gene or protein level are still present.
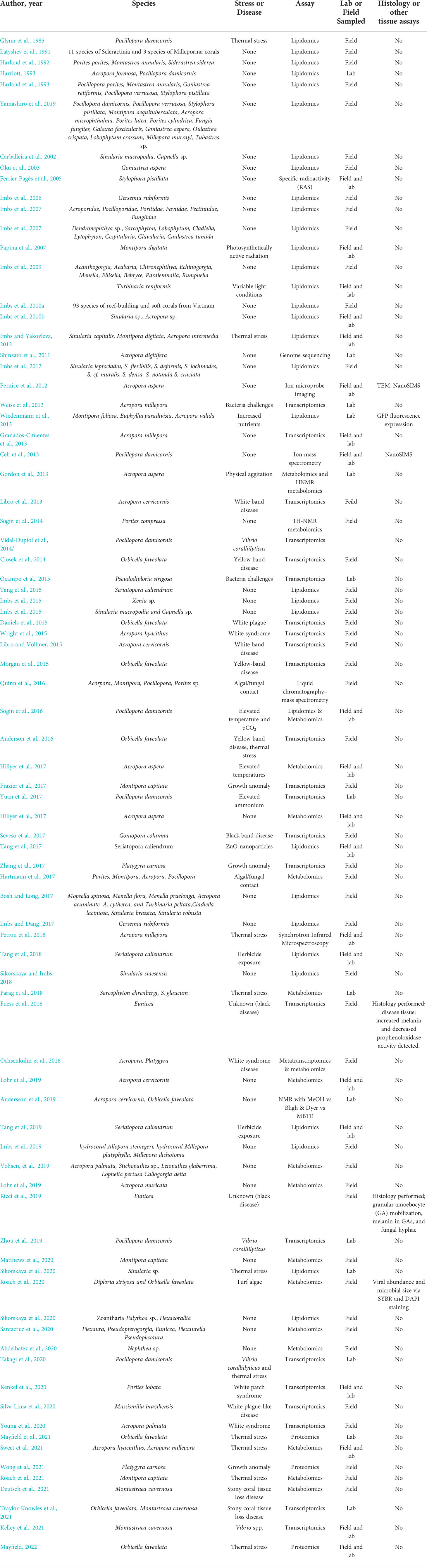
Table 1 Summary of previous 'omics studies done on soft corals, octocorals, and stony corals.
Coral biomarker development has largely focused on host gene or protein-level responses or identification of bacterial strains because they represent a single measurable unit suitable for deployment of disease diagnostics in a natural environment (Downs et al., 2000; Downs et al., 2005; Downs et al., 2009; Kenkel et al., 2014; Ushijima et al., 2014; Louis et al., 2017; Ushijima et al., 2020). However, there are many limitations to this approach. For example, consistency in the statistical significance of these types of biomarkers has been elusive (Parkinson et al., 2020). The intrinsic complexity of coral species as holobionts has proven a limitation to underpin specific mechanisms that either define a disease or are shared between different diseases. This is further compounded by the fact that corals span a wide range of evolutionary timescales and therefore may have very different strategies for dealing with different pathogens and subsequent diseases (Rowan, 2004; Bourne et al., 2009; Pinzón C. et al., 2014). In addition, within species, transcriptomic approaches have proven to be extremely variable, making biomarker development challenging (Traylor-Knowles and Palumbi, 2014; Louis et al., 2017; Parkinson et al., 2020). Lastly, technical hurdles, such as sample and library preparation, sequencing, and the type of bioinformatic packages used in the analysis can yield variability within outcomes. However, despite these complications, a rapid biomarker would be beneficial to monitor coral health.
Given the rapid emergence of deadly diseases like SCTLD, understanding the immune response of corals to disease has become more important than ever. Innate immunity is a non-specific immune process by which organisms combat foreign invaders (Palmer and Traylor-Knowles, 2012; Palmer and Traylor-Knowles, 2018). For corals, the first line of defense is the surface mucus layer, a critical boundary for trapping foreign particles and microbes (Palmer and Traylor-Knowles, 2012; Palmer and Traylor-Knowles, 2018). If this boundary is breached then an innate immune response can occur. This response is generally characterized in organisms as having the steps of (1) recognition of non-self and (2) activation of signaling pathways that lead to (3) an effector response. Like all invertebrates, corals possess pathogen recognition receptors (PRRs) that perceive microbe- or pathogen-associated molecular patterns (MAMPs, PAMPs) in different microbial communities. When compared to other metazoans, however, corals have a complex recognition system that possesses a highly redundant and abundant PRR network associated with many downstream signaling pathways (Emery et al., 2021). Much of what we understand of the coral innate immune response comes from measurements of different effector responses including phagocytosis (Palmer et al., 2011b; Snyder et al., 2021), the melanin response (Mydlarz et al., 2008; Palmer et al., 2011a; Pinzón C. et al., 2014), production of antimicrobial peptides (van de Water et al., 2018), reactive oxygen species (Mydlarz and Jacobs, 2006) and other antioxidants (Mydlarz and Harvell, 2007). Now with the use of ‘omics, the recognition and signaling mechanisms that initiate these immune effector responses are being further discovered, characterized, and connected to phenotypic outcomes. However, with this progress comes more questions, due to the complicated holobiont interactions with coral hosts. These organisms are non-self, yet they bypass the coral immune system. Is the disease caused from an in hospite organism, or a foreign pathogen invading from the outside environment, a combination of both?
Corals are holobionts; they are host to many different microorganisms including bacteria, the dinoflagellate photosymbiont (Symbiodiniaceae), viruses, micro-eukaryotes, archaea, and fungi. The most well characterized of these symbiotic relationships is between the coral host and the dinoflagellates that live in its gastrodermis, an epithelium lining the interior of the polyps and coenenchyme (Brown et al., 1995; Brown, 1997; Weis, 2008). Dysbiosis between these two partners results in bleaching and is a hallmark of many coral diseases (Brown et al., 1995; Brown, 1997; Weis, 2008). Due to the complex symbiotic nature of the coral holobiont, the causative agent(s) of many coral diseases have been elusive to identify (Vega Thurber et al., 2020). Furthermore, the resulting organismal and cellular responses can be challenging to link to a specific pathogen(s). Coral health is tightly associated with the environment where dysbiosis, or the breakdown of microbial homeostasis often caused by environmental disturbances, can initiate disease pathologies including tissue lesions, bleaching, and tissue loss (Brandt and McManus, 2009; Vega Thurber et al., 2020). This has led to a focus in coral research to monitor the coral host and its immune response in response to disease outbreaks or environmental stressors (Mydlarz and Harvell, 2007; Pollock et al., 2014; Fuess et al., 2017; Vega Thurber et al., 2020). Changes in gene expression; protein, lipid, and carbohydrate synthesis; and alterations in metabolic cascades can be studied to understand the underlying disease processes. However, this approach is not without challenges. The complexity of the holobiont and the immune response has led to limitations in understanding large-scale patterns of change because results from different ‘omics datasets can emphasize different findings that reflect the variation in the holobiomes of the affected corals. Lastly, immune outcomes can be varied depending on different abiotic stressors and the type of immune measurements being used. For example, Montipora capitata found in different pCO2 environments had contrasting catalase and prophenoloxidase activity that correlated with environmental history and temperature stress, but melanin, another marker for immunity, only correlated with environmental history (Wall et al., 2018). This example highlights the complexity of the innate immune response within individual corals. The response is varied, and influenced by the type of stress exposure.
In this review, we examine the current use of ‘omics approaches as a tool for understanding coral disease and in the development of biomarkers. We focus on diseases suspected to be biotic rather than abiotic (e.g. coral bleaching in response to thermal stress). We acknowledge that biotic and abiotic stressors are intricately linked but have chosen to focus on biotic diseases because less is understood about these processes. We start by discussing the current state of coral disease ‘omics in relation to coral immunity. We then highlight considerations for experimental design for doing large scale disease ‘omics studies, and we consider how to move forward from disease ‘omics studies to biomarker development. Lastly, we discuss the integration of ‘omics into effective conservation and restoration strategies. This review can serve as a platform to push forward the development of biomarkers and expand our understanding of coral disease outbreaks and coral immune function (Figure 1).
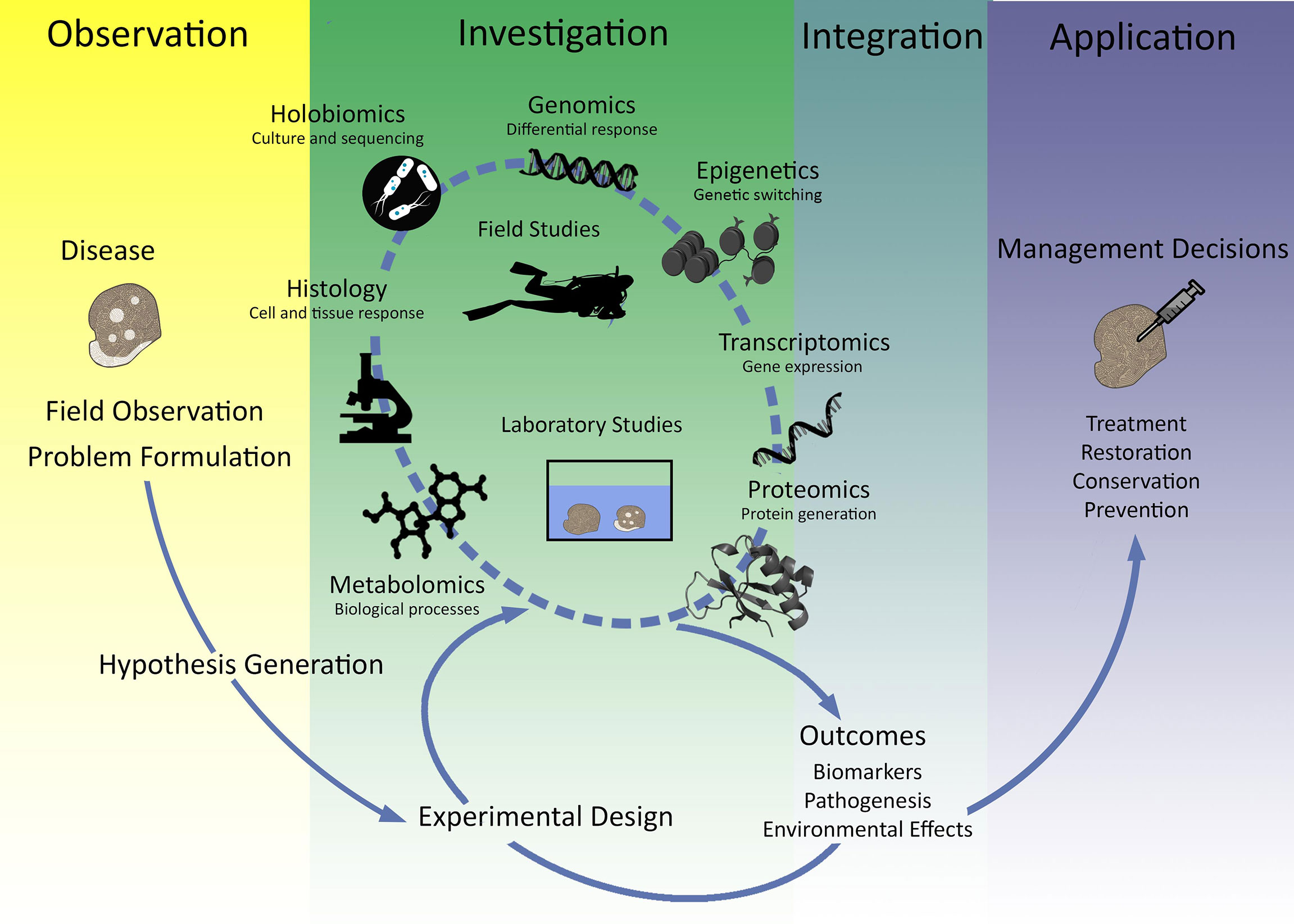
Figure 1 Integrative summary of the use of 'omics technologies for coral disease outbreaks. Summary figure of how hypothesis driven experimentation to investigate immune function as measured by 'omics technology is critical for application of marine management and conservation outcomes.
Here, we summarize what is known about the ‘omics immune response from each coral disease described to date, including what genes or proteins have been identified as critical to the response. We also provide, where possible, the histological context for each disease (Table 1). We start this section with gorgonians, which are not stony corals, but are critical for the background of coral disease because this is where the coral immunity field began, and thus gives context to the growth in this field.
One of the first models for coral disease was the sea fan (Gorgonia ventalina) aspergillosis pathosystem. This model was tractable due to easy collection of specimens and clear identification of purple lesions that often revealed the site of pathogen infection and coral immune response driven by melanization and cell repair mechanisms (Petes et al., 2003; Mydlarz et al., 2008; Mydlarz et al., 2009). Based on this seminal research, it became evident that the visual signs of disease were not only caused by the aspergillosis, but also by the host immune response. Histopathological examination of diseased sea fans helped to initially characterize cellular outcomes. For example, pleomorphic amoebocytes were found to function as the principal cells of the innate immune system in Cnidaria (Hyman, 1940; Meszaros and Bigger, 1999; Menzel et al., 2015). In gorgonians, these cells are relatively large and contain abundant acidophilic granules dispersed in the mesoglea of the coenenchyme (Figure 2; Bayer, 1954; Peters, 2015). The abundance of these acidophilic granular amoebocytes (AGrAs) varied with the condition of the colony, and focal aggregations of these cells have been seen in response to putative pathogenic microorganisms, such as Aspergillus and bacteria. AGrAs line up at the surface of the exposed mesoglea to form a barrier epithelium (Mydlarz et al., 2009; Tracy et al., 2020), and can produce a thick deposition of melanin-like material to prevent the fungi from infiltrating the gorgonian polyps and coenenchyme (Mullen et al., 2004).
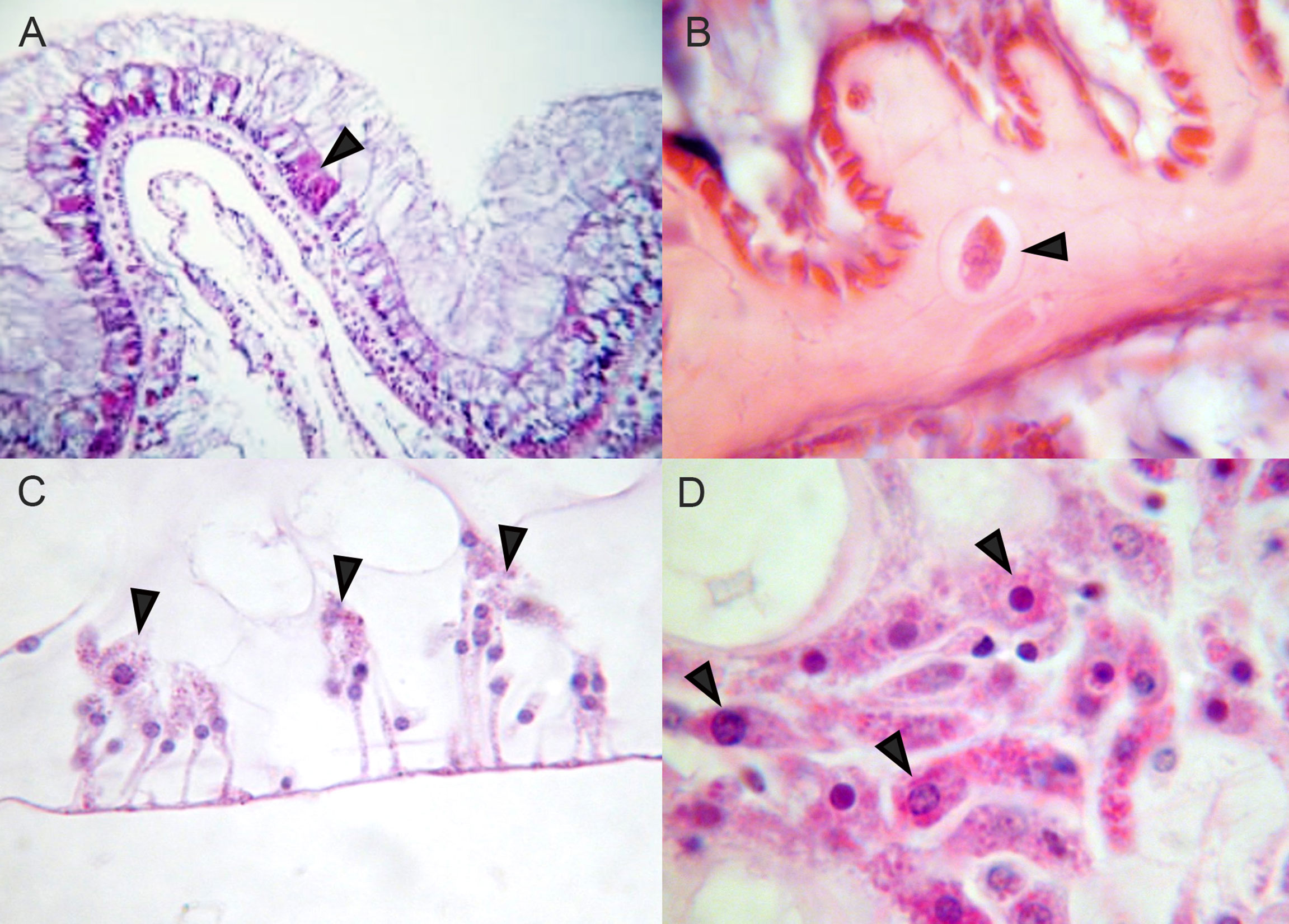
Figure 2 Amoebocytes in Hexacorals and Octocorals (A) Scleractinian hexacoral Orbicella faveolata, arrowhead indicates eosinophilic granular amoebocytes in surface epithelium. (B) Scleractinian hexacoral Pseudodiploria strigosa, arrowhead indicates amoebocyte transiting through mesoglea. (C) Octocoral Plexaurella fusifera, arrowheads indicate eosinophilic granular cells on surface. (D) Octocoral sea fan Gorgonia ventalina, arrowheads indicate eosinophilic granular amoebocytes in mesoglea.
Transcriptomic examination of diseased sea fans identified genes involved in pattern recognition, antimicrobial peptide production, and wound repair (Burge et al., 2013). These included immune-related genes such as tachylectin-5A, arenicin-2, and uromodulin (Burge et al., 2013). Likewise, Tracy et al. (2021) identified increased expression from several candidate immune genes based on the results of Burge et al. (2013) when sea fans were exposed to copepods and Aspergillus in both field and laboratory experiments. They found higher densities of AGrAs in treatments with copepods only and copepods with Aspergillus co-infection indicating that copepods, based on histopathological examination.
The first proteomic study of diseased soft corals examined Sinularia tissue-loss disease (STLD) (Gochfeld et al., 2015). Examination of differentially abundant proteins revealed a significant increase in fluorescent protein expression in diseased tissue. Fluorescent proteins are known to act as antioxidants, and this finding indicates a stimulated antioxidant immune response of the coral host (Palmer et al., 2009; Gochfeld et al., 2015). Proteins associated with symbiont photosynthesis and stress responses (e.g., heat shock proteins) were also up regulated in diseased tissue. Histological examination of Sinularia-affected tissue samples revealed increased densities of AGrAs in response to unknown foreign cells and hyperplasia of gastrodermal cells, as well as necrosis (Slattery et al., 2013).
In another gorgonian species, Eunicea calyculate, histological studies of tissue affected by Eunicea black disease also revealed increased densities of AGrAs, most containing melanin granules, and fungal hyphae penetrating the coral tissue (Ricci et al., 2019). Proteomic analysis of diseased E. calyculata identified differentially abundant proteins associated with immunity, reproduction, inflammation, and antimicrobial responses (Ricci et al., 2019). There were four up-regulated proteins in diseased samples that correlated to significantly differentially abundant transcripts quantified using transcriptomics in an earlier study (Fuess et al., 2018): a C-type lectin, a histone, a heat shock cognate 70, and a carbonic anhydrase. One of the highest upregulated proteins in diseased corals was arachidonate 5-lypoxygenase, an enzyme involved in the inflammatory response and possible recruitment of amoebocytes or melanocytes to sites of tissue damage in coral (Fuess et al., 2018; Ricci et al., 2019).
In contrast to gorgonians, histopathological examinations of scleractinian corals have shown they lack large AGrAs in the mesoglea, which is relatively thin compared to that in the octocorals, although the microscopic anatomy of only a few species has been studied (Peters, 2015). Scleractinian corals have external and internal contiguous layers of cuboidal to columnar cells that contain ciliated supporting cells and mucocytes forming a protective surface mucopolysaccharide layer. Some species have small amoebocytes with small or larger acidophilic granules between epithelial cells in the epidermis (external) and gastrodermis (internal), whereas others are found only between mucocytes in mesenteries (Figure 2). Amoebocytes may also produce melanin granules in some species. It is therefore crucial to compare the ‘omics of octocorals with hexacorals along with their histology to give context to the 'omics outcomes that are measured.
One of the first studies to use transcriptomics to investigate the immune response of diseased stony corals was done in Acropora cervicornis affected by white-band disease (WBD) on Panama reefs (Libro et al., 2013). The main differentially expressed pathways identified in the response to WBD were immunity, apoptosis, arachidonic acid metabolism, calcification and calcium homeostasis, cell growth and remodeling, and lipid and fatty acid metabolism. Libro et al. (2013) found 69 immune-related differentially expressed transcripts, including pattern recognition receptors (namely C-type lectins), mediators of phagocytosis, producers of reactive oxygen species (ROS), antioxidants, and general stress response genes. However, no differential expression of toll-like (TLR) receptor signaling, complement, and prophenoloxidase (PPO) innate immune pathways were identified. The authors concluded that WBD-induced apoptosis of coral host cells, which subsequently mounted a powerful immune response driven by phagocytic engulfment and degradation of apoptotic cells. No WBD-affected or apparently healthy samples were collected at the same time for histology; however, samples were processed and examined for other A. cervicornis samples from Bocas del Toro, Panama, presenting WBD (Casas et al., 2004), later confirming the infection of mucocytes by a novel Rickettsiales bacterium, Candidatus Aquarickettsia rohweri (Miller et al., 2014; Klinges et al., 2019). Cell degradation was evident; however, “phagocytic engulfment” of apoptotic cells was not observed. In a study several years later on Acropora palmata and A. cervicornis, histological evidence discovered two different etiologies for what was presumed to be WBD. One was described as true WBD and he other was described as rapid tissue loss (RTL) (Miller et al., 2019). RTL is characterized by acute patchy sloughing off of tissue from the skeleton with increased mucus production. At a cellular level, changes to the calicodermis, along with necrosis and/or apoptosis within the basal and surface body walls, were observed. For WBD, the histopathology varied, but similar changes were also observed in other WBD-affected acroporid (Miller et al., 2019). Subsequent studies of these same A. palmata samples found that a strong immune response was initiated in affected corals, but that the Symbiodiniaceae gene expression was not modulated, indicating that this disease may be one of the coral host rather than the algal symbionts (Young et al., 2020).
In the Indo-Pacific, white syndromes (WS) affect a variety of acroporid coral species. In the field, WS is characterized by variable and rapid tissue-loss lesions that leave the coral skeleton bare and exposed (Sussman et al., 2008). In the coral Acropora hyacinthus, the main signaling pathways identified in the response to WS using transcriptomics were those involved in the immune response, lipid and carbohydrate metabolism, oxidative stress, tissue regeneration, and calcification (Wright et al., 2015). The immune response to WS was also found to be similar to WBD with up-regulation of C-type lectins, complement factors, and transcripts potentially involved in lysosomal degradation via ROS production and proteolytic enzymes (Libro et al., 2013; Young et al., 2020). Pairwise comparisons between three distinct health states (disease lesion, apparently healthy tissues of diseased tissues, and healthy tissues) found no significant differences between apparently healthy tissues of diseased tissues and healthy tissues, indicating that the response to WS is limited to the immediate area affected by the advancing tissue loss (Wright et al., 2015). Although no transcriptomic signatures of the algal symbionts were reported in this study, the authors noted that the upregulation of lipid-based metabolism in diseased tissues may indicate a decline in carbohydrate exchange by Symbiodiniaceae, possibly due to stress-induced symbiont loss (Wright et al., 2015). While histological outcomes of the samples sequenced by Wright et al. (2015) are not known, a previous study on WS-affected tissue in A. hyacinthus found apoptosis present, as measured by TUNEL assay (Ainsworth et al., 2007).
Caribbean yellow band disease/yellow blotch disease (CYBD) is a tissue-loss disease characterized by a pale, yellow-colored band of tissue that surrounds the tissue-loss margin along an exposed, denuded skeleton in Orbicella faveolata (Reeves, 1994; Bruckner and Bruckner, 2006). Using a multiomics approach to examine coral host immune responses, CYBD affected tissue was found to have significant upregulation of various inflammation-related genes, suppression of mitochondrial associated genes, and a reduction in antimicrobial peptide expression (Closek et al., 2014). Overall gene expression patterns supported distinct changes between apparently healthy and diseased conditions; however, the visually healthy tissues of diseased corals included some differentially expressed genes that responded similarly compared to diseased samples, whereas some were more similar to trends seen in healthy samples. These patterns indicate the challenges in using differential gene expression (DGE) analysis to determine biomarkers in corals that appear healthy (Closek et al., 2014).
In another transcriptomic study of CYBD, authors focused on identifying immune pathways in diseased tissue as well as the coral immune repertoire (Anderson et al., 2016). The authors found evidence of novel homologues of the wnt and notch protein family, nod-like receptors, and dicer-like proteins within the disease transcriptome. Taken together these studies show that O. faveolata is having an immune response, but that there are nuances and challenges to using DGE as a tool for biomarker identification. This is exemplified by the fact that corals that looked visually healthy had a lot of variation in their DGE. DGE in corals is very sensitive and rapid (Traylor-Knowles, et al. 2017).
Growth anomalies (GA) affect more than 40 different species of coral, and are described as abnormal skeletal growths characterized by hyperplasia of the basal body wall, reduced oocyte development, and reduced symbiont densities (Sutherland et al., 2004; Domart-Coulon et al., 2006; Burns et al., 2011; Work et al., 2015; Zhang et al., 2017). Off the coast of Hawai'i, high prevalence of GA has been documented on colonies of Montipora capitata in areas of dense human populations; however the precise cause of the disease is still not understood (Takabayashi et al., 2010; Aeby et al., 2011). To further understand the molecular drivers of this disease, Spies and Takabayashi (2013) used quantitative real-time-polymerase chain reaction (qRT-PCR) methods to quantify the DGE of several previously identified immune and onco-genes including galaxin, murine double minute 2 (MDM2), tumor necrosis factor (TNF), tyrosine protein kinase (TPK), and βγ-crystallin (BGC). TNF was unchanged in GA compared with healthy and non-GA areas, whereas galaxin, a coral calcification gene, was upregulated in non-GA areas (Spies and Takabayashi, 2013). Later, a transcriptomic analysis of M. capitata affected by GA was performed and several genes involved in collagen, osteogenesis, and skeletal matrix production were significantly upregulated in GA-affected tissues (Frazier et al., 2017). An ortholog for the human bone morphogenetic protein 1 (HBMP1) was upregulated, as well as tumor necrosis receptor-associated factors (TRAFs), both of which are involved in the immune response. While these two studies have some contradictions in gene expression patterns, it does appear that both immune and calcification genes play an important role in GA (Andersson et al., 2020; Andersson et al., 2021).
Like M. capitata, transcriptomic and proteomic studies of GA on Platygyra carnosa both showed differential expression of genes related to immune responses, including several TRAFs (Zhang et al., 2017). Both P. carnosa and M. capitata GA also showed similar patterns in reduced expression of genes involved in calcification, and increased expression of fluorescent proteins in GA tissues. The proteomic analysis also found a suite of proteins associated with amino acid synthesis and lysosomes downregulated in the GA-affected sample that may indicate malnutrition and disruption of protein catabolism. Additionally, an increased abundance of ribosomal proteins and heat shock proteins in GA were identified. Reduced abundances of putative pattern recognition molecules and lysosomal proteins were also identified leading to the hypothesis that the stress response in GA was not triggered by microbial invasion. While the cause of these GA is still not well understood, it does seem to elicit an immune response that is primarily driven by interactions with TRAF signaling and changes in calcification.
The pathology of stony coral tissue loss disease (SCTLD) is characterized by the presence of multifocal lytic necrosis that starts within the gastrodermis and progresses to the outer epithelial layer (Aeby et al., 2019; Landsberg et al., 2020). First reported in 2014, this disease affects at least 24 different Caribbean coral species, is waterborne, and the causative agent is suspected to be microbial in origin (NOAA, 2018; Dobbelaere et al., 2020; Landsberg et al., 2020; Muller et al., 2020; Work et al., 2021). While the causative agent of the disease is still unknown, there is growing evidence that algal symbionts are tightly implicated in the etiology of this disease based on (1) histological studies showing rapid deterioration of gastrodermal cells (Aeby et al., 2019; Landsberg et al., 2020), (2) differential susceptibility of corals manipulated to host different Symbiodiniaceae (Dennison et al., 2021), (3) reduced incidence in bleached corals with reduced symbiont loads (Neely et al., 2020), and (4) correlations between SCTLD disease signs and the presence of viruses in associated Symbiodiniaceae (Work et al., 2021; Veglia et al., 2022). Metabolomic approaches have tended to support these conclusions, with different Symbiodiniaceae-related lipid classes (including betaine lipids, glycolipids, and tocopherols) being differentially expressed between apparently healthy and diseased corals (Deutsch et al., 2021). This is very similar to what was previously found during coral bleaching where changes in lipids and glycolipids within specific genera of Symbiodiniaceae are associated with thermotolerant or thermosensitive corals (Tchernov et al., 2004; Rosset et al., 2019; Sikorskaya et al., 2020; Sikorskaya et al., 2021; Deutsch et al., 2021; Roach et al., 2021). At a transcriptomic level, SCTLD is associated with the expression of many different stress genes, immunity, and apoptosis genes including peroxidases, bax-like, fibrinogen-like, protein tyrosine kinase, and transforming growth factor beta (Traylor-Knowles et al., 2021). Both the metabolomic and transcriptomic study of SCTLD reveal changes in lipid production and a concomitant immune response in the coral host, reaffirming the tight interaction between the holobiont members (Deutsch et al., 2021; Traylor-Knowles et al., 2021). As this disease is one of the newest described coral diseases, ‘omics studies are currently limited in number, but are increasing rapidly due to reduced costs of sequencing approaches and improved annotation of ‘omics references (e.g., genomes, transcriptomes, proteomes). Previous studies of other coral diseases described above laid the foundation for examination of coral immunity, and specifically emphasized the need for consistency among approaches and implementation of multiomics pipelines to better characterize immune responses and pathways.
The use of ‘omics approaches to study the immune system reaction to coral diseases has generated several converging themes. From a broad perspective, overwhelmingly we can see that the coral immune system is reacting to disease. Genes and proteins from many different immune signaling pathways are differentially regulated in disease-exposed versus unexposed (i.e., apparently healthy) tissues. An often-discussed hypothesis that corals are immunocompromised does not appear to be supported by these previous studies, in fact one could hypothesize that the opposite could be true; an overactive immune state may be present in corals. While comparative studies between coral species could shed some light on this (Fuess et al., 2017), other cnidarian models show evidence of immune memory, supporting the hypothesis that immune activity in anthozoans can be modulated based on prior disease exposure (Brown and Rodriguez-Lanetty, 2015). Future studies on coral disease could benefit from focusing on the possible hyper-activity and hyper-responsivity of the coral immune system, and the possible etiologies of disease or resistance that could result from each state (Wright et al., 2017).
While each of the 'omics studies discussed above present their data in different ways, there does appear to be convergence on several immune-related processes including extracellular matrix stability, mucus production, and markers of inflammation, and apoptosis. Many of these processes and resulting biomarker candidates are likely indicative of late-stage tissue degradation as a direct result of disease presence. Early time sampling before visual signs of disease could elucidate more about genetic mechanisms of these different diseases, and would be beneficial to take into consideration during experimental design. Lastly, few of these ‘omics studies incorporated histopathological examinations into their analysis thus our understanding of the cellular pathologies is still not connected with the gene expression outcomes. This missing link between the gene expression and the cellular outcomes needs to be further developed so that a better understanding of the disease mechanisms can be achieved.
In this section, we discuss aspects of experimental design and sampling approaches for both field and lab experiments relative to coral immunity and disease. We focus on how to maximize the chance of success in downstream ‘omics analyses, by identifying consistent sampling approaches to enable quantification of molecular processes and to facilitate cross-comparisons across datasets.
The ability to identify and characterize modulation of immune pathways and functions related to disease susceptibility and resistance relies on (1) a focus on hypothesis-driven research in field and lab settings, (2) robust experimental design and consistency in experimental approaches among studies, and (3) communication and collaboration of research efforts to maximize the impacts of individual studies. Oftentimes, we are limited in the ability to sample coral populations affected by disease, particularly when focusing on threatened or endangered species. Sourcing both diseased corals and apparently healthy corals for lab-based experiments can also pose challenges depending on the spatial scale of the endemic zone and potential seasonal variability in disease prevalence. To limit impacts on remaining coral populations affected by disease while still maximizing growth in our understanding of coral immune responses, research efforts could also be directed toward better understanding ‘normal’ metabolic and immune functions through analysis of apparently healthy individuals. Given the variability of diseased corals in the wild, generating additional information about baseline function may provide context to case study observations and experiments, better replicating how we approach rare human diseases by identifying deviations from normality. Additionally, emphasis needs to be placed on consistency of experimental approaches among species, regions, and potentially even diseases, to allow for cross-study comparisons and meta-analyses. Methods used for experimental design and sampling needs to be transparent and well documented. Here, we outline recommendations to improve statistical robustness and enhance comparisons across datasets, in the context of lab- and field-based experiments. These efforts will broadly contribute to the ability to collect robust ‘omics data in both settings and will improve our ability to characterize common immune responses to disease challenges, and subsequently, to identify and validate suitable biomarkers of disease susceptibility/resilience for screening across coral populations. This information can be used for operationalization of ‘omics-based predictions of resilient genotypes to increase success of conservation and restoration initiatives. We acknowledge that this will not be an easy task, as fitness trade-offs exist, however, with the abundance of ‘omics data now available cross analyzation of different types of stressors and diseases are feasible and may help to disentangle potential trade-offs.
There is an inherent difficulty in conducting field sampling efforts and lab-based experiments consistently, in that coral diseases tend to change in terms of prevalence and perhaps even etiology among regions and through time (Aeby et al., 2019; Shore and Caldwell, 2019; Meiling et al., 2021). Few species are readily available in sufficient density for field and lab experiments, and within Florida where the disease was first observed, it is not feasible to source disease-naïve corals from the wild. Furthermore, of the species remaining in sufficient numbers to support sampling and experiments it may be impossible to identify colonies that have not been exposed to prior intervention efforts including antibiotic and probiotic treatments (Paul et al., 2019; Ushijima and Aeby, 2019; Meyer, 2020; Peixoto et al., 2021).
Furthermore, analysis of ‘omics samples from field experiments pose challenges to robustness of statistical designs. For example, environmental variability across microhabitats where coral colonies are sampled, or variation in microbial symbiont communities among coral colonies can introduce unexplained variance into statistical models examining treatment responses (i.e., following antibiotic or probiotic intervention). Most field-based designs also do not allow for replication of coral genotypes across treatments, except in cases such as reciprocal transplant experiments. As a result, the high levels of environmental stochasticity associated with sample collections, in conjunction with relatively low control over genotypic/symbiont variability, can result in data noise that overwhelms any treatment effects. There are several potential ways to mitigate these issues, foremost, increased replication within treatments for better resolution of genotypic and habitat variability. The former may also be improved through data collection of additional environmental parameters, as some ‘omics analysis pipelines (e.g., weighted gene co-expression network analysis, or WGCNA; (Langfelder and Horvath, 2008) can examine links in molecular responses to quantitative traits and parameters. For improved genotypic replication, experimental designs may consider a repeated-measures approach, where colonies are revisited and sampled over time for fate-tracking purposes. This would also facilitate examination of colony survival through time related to disease progression, and responses to intervention treatments. Such designs require extra considerations, however, to minimize overall impacts on colonies related to multiple sampling events, or potential compounding effects on treatments. Repeated-measures approaches should therefore attempt to minimize sample volumes, such as shifting from skeletal fragments taken from a colony margin with hammer/chisel or extraction of 2.2-cm diameter core biopsies, to shallow tissue scrapes or syringe samples of polyps. There are tradeoffs to smaller samples, namely the prevention of conducting other analyses on collected samples (i.e., histology), but the sampling approach can be modified accordingly based on the research questions of the study. More than one sample may be needed from the same colony, for example, core biopsy preserved for histology and core biopsy frozen or preserved for DNA/RNA analyses; these should be taken as close together as possible from each sampling location on a colony with the same visual tissue response. Additionally, efforts to sample from consistent areas on different coral colonies can be prioritized and well documented (e.g. Drake et al., 2021). Finally, field experiments often have unexplained variance due to sampling times, as the amount of time needed to sample a coral underwater and, for example, preserve it on a moving boat, is generally greater than for a lab experiment. Considerations to field operations and personnel should be made to minimize both the time between sampling underwater and topside preservation, as well as the time delay between individual samples collected (such as across sites). Lastly, the environment in which the coral is collected needs to be taken into consideration. Corals are extremely sensitive to their local environment, and measurements such as depth, light, and water flow may have effects on ‘omics outcomes.
While lab-based experiments have some limitations in that they do not often replicate ecologically relevant environmental stochasticity or genotypic variability found in the wild, they are useful for ‘omics analyses that require a consistent treatment effect and controlled genotypic variability to maximize inferential power. For examining disease exposure in particular, ex situ designs allow for greater control over potential co-infections among replicates or environmental effects on disease transmission and progression. As a result, lab experiments could prioritize statistical independence of replicates within treatments using individual micro-aquaria rather than communal tank systems. This eliminates pseudo-replication and minimizes the chance of coinfections occurring from one experimental coral fragment to another. These types of designs often require more coral biomass, especially considering that multiple disease donor fragments are required versus a single fragment in a communal design, but independence of transmission replicates will allow for more robust downstream statistical analysis. Such designs also lend themselves to examination of disease susceptibility/resistance across multiple genotypes, as clonal fragments can be allocated across treatments (Miller et al., 2014; Muller et al., 2018; Young et al., 2020). Additionally, it has become more apparent from a growing number of studies addressing molecular mechanisms related to disease exposure that there may be a sizable impact of genotype and/or symbiont communities on coral responses to disease challenges. Therefore, lab-based experiments accounting for genotypic effects would be beneficial, specifically those that incorporate multiple replicates of genotypes within treatments to allow for statistical comparison of variation in response variables (e.g., gene expression, microbial communities, algal symbiont communities).
Finally, as lab experiments often involve prior fragmentation to achieve a suitable number of coral fragments, future studies could consider a recovery period to allow fragments to heal prior to disease exposure. Recently cut margins may provide a means of entry for pathogens and could artificially inflate transmission rates and time to onset of visible disease signs. Studies have suggested that there is variation in wound healing times among coral species (Work and Aeby, 2010; Rodríguez-Villalobos et al., 2016; Traylor-Knowles, 2016; Edmunds and Yarid, 2017; Counsell et al., 2019), but a period of 2–3 weeks is likely to be sufficient to repair fragmentation damage at cellular levels, provided that water quality, light levels, and food supplementation are optimal. Additionally, fragments should be quarantined and observed for mortality prior to experiments, especially those that are collected from the wild. Conversely, with extended time (months) in laboratory aquaria, there may be changes in holobiont composition, such as microbiomes and/or endosymbiont communities, that can impact the health status or representability of lab-reared corals relative to wild samples (Galand et al. 2018). Such changes through time may influence molecular responses to experimental treatments and may therefore affect downstream ‘omics analyses. To aid in our understanding of the ‘omics outcomes, histological examination of subsets of (1) apparently healthy coral samples collected in the field, (2) samples taken at the end of the acclimation period prior to application of experimental treatments, and (3) control coral samples maintained separately but under the same environmental conditions as experimental corals should be preserved. This would be useful to evaluate health status or rule out other causes of mortality observed in experimental fragments (e.g., husbandry-related, or pathogens other than the disease of interest). Histological markers that indicate stress or disease include areas of necrosis, tissue fragmentation not associated with the sample collection, algal symbiont loss, degeneration, and atrophy of tissue, excessive calicodermal proliferation, granular cell accumulation that can include identifiable etiologies such as algal, fungal, or filamentous bacterial overgrowth, ciliate invasion, or metazoan parasites in a sample (Galloway et al., 2007; Work et al., 2016). These may be found in field sampled corals and are not always associated with visible gross lesions on the coral colony. If this is found in the initially apparently healthy coral samples collected from the field, then it is an indication that the coral is not healthy and the downstream disease and ‘omics analysis may be confounded by a pre-existing disease condition.
Perhaps the second greatest challenge in conducting robust disease exposure experiments comes in the timing of visible disease signs and sampling. There is likely temporal variability at the transcriptomic and holobiont levels related to disease infection and progression, but these processes have not been well-studied in corals. Time series studies are would ideally be performed to identify variation in multiomic responses related to pathogen exposure through time, to better understand the mechanistic processes related to disease infection and progression, and to inform future studies of the ideal time to sample to best capture molecular responses. In the case of SCTLD, several studies have been conducted to examine microbial communities in wild diseased corals and in laboratory transmission experiments (Aeby et al., 2019; Rosales et al., 2020; Meiling et al., 2021; Studivan et al., 2022). It is hypothesized that many of the bacterial taxa found in disease samples represent opportunistic growth of heterotrophs and do not necessarily capture the primary pathogens (Ushijima et al., 2020). It has also been suggested that the white denuded skeleton lesions typically used to characterize SCTLD in the field may represent a late-stage sign of the disease (Aeby et al., 2019). Samples collected earlier in the disease process may facilitate the identification of pathogenic microbes as primary invaders, as well as the ability to quantify the molecular response of the coral to disease, biotic, or abiotic pathogens (Rosales et al., 2020; Traylor-Knowles et al., 2021).
At the analysis stage, increased replication in experimental designs has downstream benefits because outliers are more easily removed from a dataset without sacrificing statistical robustness. Initial data visualization tools can be used to identify potential outliers. These include principal component analysis, and the programs: arrayQualityMetrics (Kauffmann et al., 2008) and ANGSD (Korneliussen et al., 2014). Outputs from these programs can then be compared to metrics captured during library preparation and sequencing (e.g., FASTX-Toolkit (Gordon and Hannon, 2010) or Trimmomatic (Bolger et al., 2014) These metadata can help to remove aberrant samples that may be driving or masking treatment effects in statistical models. Complicated experimental designs, such as those with multiple species or treatments, may also impact robustness through the reduction of replication within groups. An iterative approach may be more useful, where experimental designs are first tested and optimized within a few species, and following success, can be expanded to incorporate other species or treatments. This is especially true for cross-comparisons and meta-analysis of datasets across regions within the Caribbean, where previous experiments have demonstrated substantial variation in disease etiology and susceptibility to SCTLD within Florida and among Caribbean jurisdictions (Aeby et al., 2019; Alvarez-Filip et al., 2019; Estrada-Saldívar et al., 2020; Heres et al., 2021; Meiling et al., 2021). In the case of the ongoing SCTLD outbreak, the adoption of the same designs, approaches, and species for experiments conducted in southeast Florida, the Florida Keys, and the United States Virgin Islands would be extremely beneficial to understand both how SCTLD varies across region. It would also clarify whether corals are responding to disease exposure using similar molecular mechanisms and cellular responses across their ranges.
Here we will discuss different approaches to biomarker development, and how ‘omics resources can be better exploited to develop biomarker systems. We focus on biomarker development in other fields and identify common strategies that are likely to increase success of biomarker development and validation in corals. Case studies are examined for multiple ‘omics approaches, particularly those that are gaining popularity for use in the coral field.
The ultimate goals of biomarker development for diagnosing coral health are twofold: (1) to rapidly identify indications of disease, and (2) to characterize disease resistance and susceptibility for tailored management approaches that protect vulnerable populations and/or restore resilient coral genotypes. Biomarkers, particularly when implemented in relatively inexpensive, rapid diagnostic tests modeled after those used in human medicine (e.g., rapid COVID-19 at home test, ketone test strips) could offer the ability to quickly identify the potential for infection, and to respond to coral disease outbreaks by conserving and propagating disease-resistant genotypes. Biomarker development also facilitates the efficient translation of ‘omics datasets into management initiatives by conservation and restoration practitioners. In theory, this would be expected to greatly reduce the amount of time between initial observations of disease signs in the field and subsequent responses to mitigate the impacts of further disease spread. These same diagnostic and response approaches are likely to be successful in responding to multiple stressors beyond infectious diseases (e.g., bleaching, ocean acidification, sedimentation, nutrification) by allowing the identification of resilient genotypes. Therefore, it is a priority in coral conservation to develop generic biomarkers to broadly assess coral health holistically across a variety of stressors. This will enable high throughput screening of wild and nursery farmed corals to select resilient genotypes that can withstand future climate change scenarios, remain adaptable in response to pollution, and respond to novel disease outbreaks.
Based on the challenges of biomarker development in corals for disease exposure, there is a clear need to develop a consensus of not only what to target (gene, protein, etc.), but also considerations for experimental design and assaying methods as discussed above. And perhaps it would better serve our community to expand outside of what we consider to be a typical medical biomarker. Below, we explore some areas where biomarkers could be expanded from ‘omics data, and when combined with other techniques, may support the monitoring of and predict the requirements for. It is critical to establish markers that can be used as widely as possible and can encompass the inherent genetic variability that defines many corals’ molecular responses. Additionally, expanding our approaches beyond targeting the coral host towards understanding the other holobiont components in tandem with the coral host may provide additional insight towards biomarker development. Below we discuss some of the other techniques from ‘omics that have promise for biomarker application.
A primary focus area of ‘omics applications has been to understand coral disease mechanisms and develop early detection or monitoring systems for disease outbreaks. However, oftentimes late-stage diagnosis is what can be concluded from the collected data. The motivation for using a large-scale ‘omics approach on the coral host is that we can narrow down possible markers for resiliency or tolerance that can then be used in the future for assaying a wider range of corals. This approach appears to be straightforward in theory, but many challenges have been encountered, namely that a universal biomarker for coral stress and/or disease has not been identified because of the inherent variability within coral genetics, the coral holobiome, and the influence of the environment (Traylor-Knowles and Palumbi, 2014; Louis et al., 2017; Parkinson et al., 2020). Results to date have been more successful in identifying biomarkers for pathogenic bacteria, rather than biomarkers in the coral themselves (Ushijima et al., 2020; Rosales et al., 2020). For example, researchers can identify bacteria with strain-level specificity using an Analytical Profile Index (API) strip. The API strip is a common medical diagnostic tool used to quickly identify bacteria of interest. It is stable at room temperature and can easily be modified to identify different bacteria candidates. The API strip has the potential to be used in water monitoring as an early monitoring system for identifying potentially pathogenic bacteria. For example, the VcpA RapidTest was successfully used in coral to identify Vibrio coralliilyticus strains associated with SCTLD (Ushijima et al., 2020). While this approach is applicable for identification of coral-associated bacteria, it does not address the coral host, or other symbiotic partners. Additionally, this approach is not without challenges, most coral diseases are not able to fulfill Koch’s postulates, and the few that have reported doing so are still highly debated (Nagelkerken et al., 1997; Geiser et al., 1998; Richardson et al., 1998; Richie and Smith, 1998; Patterson et al., 2002; Ben-Haim et al., 2003; Pantos et al., 2003; Sussman et al., 2008; Sutherland et al., 2011; Kellogg et al., 2013; Lesser and Jarett, 2014; Pollock et al., 2017). Expanding this approach to also include testing coral lipids, mucous sugars, or coral tissue antioxidant levels could be a valuable tool for assessing coral health to supplement bacterial monitoring (Harriott, 1993; Mydlarz and Palmer, 2011; Seemann et al., 2013; Lee et al., 2016). Additionally, creating approaches to address different partners of the holobiont including Symbiodiniaceae and other microbial partners may generate a more holistic diagnostic tool that can be useful for coral biomarker development, validation, and screening.
The most classical approach to examining cellular aspects of disease has been to use histology. It is a downstream tool that can be compared to changes observed in the transcriptome, metabolome, and proteome. Histopathological studies can encompass both morphological and functional changes of cells and tissue in repsonse to disease. Previous research on coral disease histology have revealed changes in the structural arrangements of cells and tissues, indicating functional impairments of both the coral host and associated endosymbionts (Work and Meteyer, 2014; Aeby et al., 2019). It is a valuable tool because it can reveal the physical location of suspected infectious pathogenic microorganisms responsible for coral diseases, be it the coral host or members of the complex microbiome (Peters, 2015). Knowing the physical location of an infection can help with the disease etiology and future intervention efforts (Work and Meteyer, 2014). Histopathology can also be useful as a screening tool to explain different coral disease pathologies. For example, visual signs of disease on corals can look very similar to the naked eye, especially in the field, but at a tissue level the pathologies can be very different. Using histology, one can differentiate between diseases by examining the cellular aspects to interrogate changes in the tissue integrity, cell death, and presence of bacteria, among many other traits (Work and Meteyer, 2014). In addition to histology, other tissue focused techniques can be used. For example, a tool called microdissection has been used to confirm histological observations; using this technique microorganisms can be dissected and sequenced directly from the preserved coral sample (Ainsworth et al. 2015). Reliance on molecular markers or deep sequencing alone can result in attribution of diseases to microorganisms or viruses identified in diseased tissue that are either secondary invaders or bystanders in the environment (Hewson et al., 2014). These types of challenges are more likely to be avoided if a variety of techniques are used, including cellular observations that can be correlated with ‘omics outcomes (Work and Meteyer, 2014).
Additionally, incorporation of other classical cell biology techniques, such as cell culture and in situ hybridization, as well as contemporary cellular techniques, such as fluorescent activated cell sorting (FACS) and single cell sequencing (SCS) technologies, promise to be informative for our understanding of coral disease. Cell culture techniques can be used to isolate specific coral host cells or specific microbial partners to examine their function and behavior ex vivo. This is a valuable tool because functional assays can be quickly developed and deployed on live cells that are not accessible using the whole coral organism. In situ hybridization is used to examine the spatial gene expression within tissues. This can be done for any stressor but has traditionally been used in coral biology to examine developmental stages (Traylor-Knowles et al., 2017; Traylor-Knowles, 2018). This method is useful because it can show you what cells are expressing a particular gene, and where those cells are located (Traylor-Knowles, 2018). FACS, while not a new technology in human medicine, is relatively new to use in corals (Rosental et al., 2017; Snyder et al., 2020; Snyder et al., 2021). Using this technology, functional cell markers that are non-species specific can be used to identify and quantify the function of different cell populations. It can also be used to collect specific cell populations which can then be sequenced, preserved for microscopy, or grown in cell culture (Rosental et al., 2017; Snyder et al., 2020). SCS encompasses many different types of ‘omics targets and can be useful for isolating the function of different cells. Cell atlases can be developed and using functional markers, specific cell populations can be isolated and sequenced (Sebé-Pedrós et al., 2018; Levy et al., 2021). Using these methods, more targeted biomarkers can be developed that can eventually be species-specific, where in combination with proteomics, peptides for specific markers can be discovered and used as a diagnostic tool that would be useful for understanding disease progression.
Here we discuss different ‘omics approaches that could be incorporated into research on the immune response during disease. Much of the previous work highlighted uses gene expression for biomarker development. While this has been useful, expanding beyond this may give us a more holistic understanding and therefore better equipped to develop informative biomarkers for coral disease.
On its own, proteomics can provide information on the extent of overall peptide expression within a sample, but when combined with other genomic and transcriptomic approaches, proteomics then offers a glimpse into how corals directly respond to disease through changes in protein expression and post-translational modifications. Consequently, while currently limited by the amount of proteomic data available for non-model organisms like corals, its applications within coral disease will continue to grow as the method is used in conjunction with the other technologies. For example, in the most recent reports including that for O. faveolata (Mayfield et al., 2021), Pocillopora. acuta (McRae et al., 2021), P. lobata (Tisthammer et al., 2021), and P. carnosa (Wong et al., 2021), peptide identification and annotation benefited from available or developed transcriptomic data for those associated species. In the future, as whole genomes become more available for those and other species, the process could be further improved as transcriptomic data becomes better annotated (Liew et al., 2016). This would subsequently support the associated proteomic analyses by more confidently identifying each peptide, thereby effectively overcoming a current annotation limitation in some analyses.
While the above example highlights the connectivity within the ‘omics sciences, results can sometimes be obscured by the complex holobiont community within corals (Vega Thurber et al., 2020). Consequently, it is not just coral genomics or proteomics, but rather an entire suite of tools that will be needed to fully understand coral disease etiology. For example, the protein-expression information from proteomics could help identify peptides associated with a specific coral disease (Garcia et al., 2016; Ricci et al., 2019). Additionally, many tools have been previously developed for deconvoluting microbiome data and have the potential to be modified to use on coral holobiont datasets (Carr et al., 2013; Meyer et al., 2018). By developing antibodies for those peptides, further immunohistochemical and microscopic analysis can be performed and potentially offering new insight into the peptide’s cellular expression patterns, timing, and locations (e.g. Mass et al., 2014).
Metabolomics is representative of changes in metabolite function in response to a specific stressor. Measurement of exogenous or endogenous metabolites in a particular physiological or developmental state, termed metabolomics, is an emerging field in coral biology and not yet as established as genomics even in the field of human cancer biomarker discovery (Bailey et al., 2018). Despite the emerging nature of this technique in human medicine, metabolic disorders can easily be measured using metabolite screening and treatment strategies for bacterial infections can be quickly diagnosed using mass spectrometry approaches (Dénes et al., 2012; Sun et al., 2012). Metabolomics has promise for coral disease research because of its direct application for biomarker development.
Relatively little is understood about coral metabolomes (Garg, 2021); however, we do know that the genetic background of the host, the endosymbionts, the microbiome, and the current and historical environment all contribute to variability in metabolomic patterns (Sogin et al., 2016; Lohr et al., 2019; Garg, 2021). If we want to use metabolomics for coral disease biomarker development and to tailor treatment approaches, we first need to be in “data collection” mode. Unique and shared pathways need to be defined among different coral genera while taking into account differences in seasons, bleaching history, coral spawning, genetic background of both the coral host, the endosymbionts, and the differences in the associated microbiome (Garg, 2021). We then will be able to query whether the unique metabolic pathways shift during a pathogenic infection and better define disease status. Lastly, we need to determine the source of these pathways: the host, the endosymbiont, the microbiome, effects from the environment, or a combination of the above (Garg, 2021). Most coral metabolic studies are done on a slurry of tissue that is composed of the coral host and all of its endosymbiotic partners (Garg, 2021). Together, the coral host and endosymbiotic partners represent many sources of confounding factors that can lead to variation in interpretation of the metabolomic profiling. Due to this, data from healthy corals maintained in nurseries, data from controlled laboratory experiments, and data from different species of corals and timepoints provide a valuable resource for the removal of these confounding factors.
Alternatively, multiomics approaches, wherein the genetic background of the host, the microbiome profiles, the variability in endosymbionts paired with metabolome, proteome and transcriptome analysis can enable grouping of variation by its source. We know from previous studies that coral genotypes have unique metabolomes, and that fragments of the same genotype have conserved metabolomes, highlighting the importance of knowing the genotype prior to investigation (Lohr et al., 2019). Visualization of spatial variation in ‘omics data can further enable delineation of source of variation (Little et al., 2021). Together, multi-genotype experiments that use multiomics approaches will enable the development of a knowledge base to stratify corals in terms of disease resilience and holobiont composition to enable biomarker evaluation, and linkage of specific pathways in disease (Muller et al., 2018; Miller et al., 2019).
In abiotic stressors such as thermal stress, Symbiodiniaceae has been used as a marker for stress tolerance. Corals harboring Symbiodiniaceae that are less susceptible to thermal stress will not bleach as easily as corals possessing highly susceptible Symbiodiniaceae. Different Symbiodiniaceae taxa have different thermal tolerances and Symbiodiniaceae switching is known to be a critical strategy for coral survival (Silverstein et al., 2014; Baker, 2004; Cunning et al., 2018). Additionally, the bacterial microbiome of corals has been used as a proxy for health, where the more severe the stress is, the higher beta diversity is observed (Zaneveld et al., 2017). In biotic diseases such as WBD, this same pattern was observed indicating that the bacteria have the potential to be a useful marker for stress (Rosales et al., 2019; Rosales et al., 2020). Within SCTLD, evidence is mounting that the Symbiodiniaceae may harbor or interact with the pathogen(s) that are causing this deadly disease (Work et al., 2021; Veglia et al., 2022), and that the type of Symbiodiniaceae may affect susceptibility to SCTLD (Dennison et al., 2021). While we know that not all diseases are alike, it is important to consider the relationship of the Symbiodiniaceae communities along with the coral host, as the interactions are so tightly linked. By not examining this, we may be missing critical pieces of information including more context to the overall host reaction.
Environmental epigenetics is a burgeoning field that seeks to unveil the mechanisms that drive gene expression changes without changing the DNA sequence (Bollati and Baccarelli, 2010). Given the major pressure of anthropogenic activities such as increasing sea surface temperatures or eutrophication, these tools are starting to be applied to scleractinian corals to understand both phenotypic plasticity as well as role of transgenerational non-mendelian inheritance (Bollati and Baccarelli, 2010; Rodriguez-Casariego et al., 2018; Rodríguez-Casariego et al., 2020). Although the studies on corals are emerging, new genomic approaches that combine high throughput bioinformatics and single cell technology have opened new opportunities to examine a broad range of epigenetic modifications (e.g., DNA methylation or histone modification) (Eirin-Lopez and Putnam, 2019; Zhao et al., 2020; Cowen and Putnam, 2022). With the continual improvement of current coral genome models and the ever-growing availability of new coral ‘omics data, using epigenetic tools such as Assay for Transposase-Accessible Chromatin with high-throughput sequencing (ATAC-seq) is becoming more accessible. ATAC-seq has already been used on Exaptasia pallida, a sea anemone model used to study cnidarian/algal symbiosis but has not yet been applied to the study of coral disease (Weizman and Levy, 2019). In corals, emerging epigenetic tools will be critical for better understanding the diverse phenotypic outcomes in response to disease within and between genotypes.
Ecological approaches including (1) ecological monitoring of the reef biodiversity, (2) environmental monitoring in conjunction with ‘omics sampling, and (3) uses of environmental DNA (eDNA) to monitor DNA in the context of a changing environment can be and have been used as “biomarker” systems. Ecological monitoring to determine the overall biodiversity of a reef system is a proxy for overall reef condition (Kramer, 2003; Rogers, 2013; Díaz-Pérez et al., 2016). This process is usually focused on monitoring the physical state of the reef, environmental monitoring, and coral reef biodiversity but it can incorporate ‘omics approaches including gene expression, protein expression, and recently eDNA monitoring (West et al., 2020) Previous work on environmental monitoring in conjunction with ‘omics sampling has primarily focused on thermal stress (Barshis et al., 2013; Kenkel et al., 2013; Kenkel et al., 2014; Palumbi et al., 2014; Anderson et al., 2016; Davies et al., 2016; Wright et al., 2019; Dixon et al., 2020; Studivan et al., 2021), freshwater discharge (Edge et al., 2013; Mayfield et al., 2013; Aguilar et al., 2017; Aguilar et al., 2019), and synergistic stressors (Wright et al., 2019; Studivan et al., 2021). These environmental stressors can promote the development of disease in reef organisms and reduce reef biodiversity. Several of the previous studies also incorporated genotypic factors into experimental designs using colony fate-tracking or reciprocal transplants, allowing comparisons of genotype x environment impacts on molecular processes. These efforts to monitor wild corals’ responses to environmental variability are extremely useful, especially to characterize ‘normal’ behavior and common responses to multiple stressors but are also time consuming in terms of experimentation and field monitoring, validating the findings, and downstream analysis. Recently, the method of eDNA monitoring has developed into a potentially useful way of monitoring biodiversity (Leray and Knowlton, 2015; Formel et al., 2021). eDNA is a metabarcoding method to detect and quantify multiple types of DNA from environmental samples including water samples (Beng and Corlett, 2020). While this technology still has many challenges including issues with assigning taxa, and incomplete detection, it is currently being developed for use on coral reefs, in conjunction to in-person monitoring practices (Leray and Knowlton, 2015; West et al., 2022). eDNA promises to aid in the discovery of novel species on coral reefs and lead to a more comprehensive picture of reef condition (Leray and Knowlton, 2015; West et al., 2022).
The utilization of specific biomarkers will be directly connected to the development of economically scalable tools that can accurately diagnose or predict coral health. The level of vast complexity in corals calls for an evolution within research institutions that perform these post-genomic investigations to consider the integration of modeling or machine learning tools for enhanced predictive capacity. Research groups performing ‘omics investigations have the computational familiarity that is suitable to evolve into adopting machine learning and modeling tools to better understand ‘omics data. These tools are being developed and innovated in human research and have seen successful application within coral research as well (Hartmann et al., 2017; Mallick et al., 2019; Lima et al., 2020). Human cancer research has integrated gene expression research to identify biomarkers specific to types of cancer, tumor growth patterns, and even the stage and location of cancer (Henry and Hayes, 2012). This research has allowed for predictive modeling in disease and healthy individuals that are at risk among populations (Duan et al., 2005; Kourou et al., 2015; Meyer et al., 2021). Advances in human medical research provide theoretical and technical tools that can be leveraged to achieve overlapping goals in coral research.
The opportunity to forecast bleaching susceptibility from machine learning that utilized proteomic data has already been successfully demonstrated (Mayfield, 2022). Another approach is trait-space modeling, which uses hierarchical clustering and random forest analysis to identify traits that are the most predictive of survival or susceptibility (Darling et al., 2012). This approach is scalable as metrics of microbial ecology, gene expression, immune assays, life history strategies, and other parameters can collectively be compared and ranked for their predictive potential of coral survival. By identifying the metrics that predict survival, these biomarker candidates can be expanded to the broader population. Machine learning and novel computational biology programs can help interpret complex ‘omics datasets to identify a set of genes or microbial targets ideal for biomarker development. The potential of this has been recognized by leading conservation organizations, such as National Oceanic and Atmospheric Administration (NOAA), whose strategic plans are explicitly allocating funds to develop machine learning pipelines (LeBoeuf et al., 2021). The untapped use in machine learning and modeling has the potential to help provide clarity to complex ‘omics datasets and have the potential to aid in the discovery of less variable biomarkers (Marcos-Zambrano et al., 2021).
In this review, we discussed what we have learned about coral immunity using ‘omics during disease outbreaks, how ‘omics experiments can be designed to address these immunological questions, and how to move beyond ‘omics to biomarker development, with an emphasis on broadening our definition of what a biomarker may be. However, in compiling this review, we reflected on how and why we got here as a community and identified the many potential future directions and research needs. Here we will discuss these opportunities and describe the priority needs for furthering our understanding of coral immunity and disease outbreaks.
As a community, we know that there will be another disease outbreak. So we need to be prepared. However, to be ready, this will require infrastructure and coordination. A great example of coordination is the Disease Advisory Committee (DAC), which was started to engage different stakeholders and communities to discuss science quickly and to disburse funding to help push research forward more quickly when the south Florida SCTLD outbreak was recognized (Florida Coral Disease Response Research & Epidemiology Team, 2018). There have been many successes with the development of the DAC including more communication in the coral disease community, engagement, and interaction of scientists in different professional sectors, and funding to support small- and large-scale coral disease studies. This large-scale coordination has helped to promote a lot of important coral disease science that had direct conservation applications. These successes have required a lot of effort, coordination, and funding to be achieved, but despite these big successes, there are still some challenges. For example, with the speed at which coral diseases spread, the ability to get funding dispersed and sample collection permits obtained can lag behind the ecological threat. Coordinating different professional sectors can also be challenging which can inadvertently slow down progress.
As a community, we need the ability to pivot quickly and respond to these new threats and most importantly be prepared before the threat occurs. Ways in which we can be better prepared include developing dedicated facilities and staff that will oversee responding to these threats as well as provide training and facilities for investigation when the disease outbreak occurs. Additionally, a coordinated strategy to establish and maintain a tissue bank for reference samples to be shared and preserved would be beneficial. Lastly, ‘omics research produces a lot of data, and these data would ideally be shared openly and quickly. There have been many previous efforts to share coral ‘omics data through the development of many different databases (Traylor-Knowles et al., 2011; Sun et al., 2013; Liew et al., 2016; Zhang et al., 2019), however these databases require long term support and upkeep. Development of facilities to maintain such resources would benefit the coral research by streamlining resources and creating less redundancy.
In this review we described what is known about the coral immune response during disease and identified areas of overlap between different diseases that include many important signaling pathways and immune receptors. However, translation of these mechanisms into what it means for coral health needs to be would need further development. For example, the range of a normal immune response in many coral species is still not well understood, and so translating these findings into effective biomarkers can be challenging. How the range of a normal immune response interacts with changes in the environment and microbiome disruption are still not well understood but will have big implications for our understanding of coral disease mechanisms. There is also a gap in linking these ‘omics outcomes with cellular pathologies observed during disease outbreaks. We propose that coordinated efforts to correlate immune metrics into existing metrics of coral health will be informative and useful for future conservation applications. Lastly integrating the effects of coral disease treatments on the immune response will also be informative, as many wild corals are currently being treated with antibiotics and probiotics. The long term effects of these treatments is not known and therefore warrants active long-term monitoring.
The use of ‘omics as a tool for studying the coral immune response to disease is powerful and has potential to push forward our understanding of resilience in corals. Quick and coordinated sampling and experimental design efforts, expanding our ideas on how we think of and develop biomarkers, and a better understanding of overall coral immune health will all be essential for us to respond to current and future disease outbreaks. Lastly community efforts to standardize, preserve, and openly share data would be beneficial.
NT-K designed the overall concept for this review and led the discussions. AH designed Figure 1. ECP provided the pictures for Figure 2. All authors contributed to conceptualization, writing, and editing of the manuscript. All authors contributed to the article and approved the submitted version.
NT-K is funded by NSF Award # 2128071, 1951826, and 2013692. NG is supported by NSF CAREER Award # 2047235. JV was supported by the NOAA OAR Omics Program through Award # NA15OAR4320064. MS is supported by the NOAA Coral Reef Conservation Program Award #31252 and the NOAA OAR ‘Omics Program. LM is funded by NSF Award #2109622, MM and JS were supported by NOAA CRCP NA19NOS4820132.
The authors would like to thank Dr. Cheryl Woodley of NOAA for her advice and guidance as this manuscript developed. We also want to thank the members of the Disease Advisory Committee, along with Victoria Barker, Samantha Cook, and Maurizio Martinelli for facilitating collaborations. We thank Jessica M. Deutsch, graduate student in Garg laboratory at Georgia Institute of Technology, for assisting with the metabolomics section of Table 1. We also thank the reviewers for their helpful comments. The authors would also like to thank Janet M. Carter and Dr. Katherine L. D. Richgels for their copy editing. U.S. National Oceanic and Atmospheric Administration Disclaimer: The scientific results and conclusions, as well as any opinions expressed herein, are those of the author(s) and do not necessarily reflect the views of NOAA or the Department of Commerce. The mention of any commercial product is not meant as an endorsement by the Agency or Department. U.S. Geological Survey Disclaimer: This article has been peer reviewed and approved for publication consistent with U.S. Geological Survey Fundamental Science Practices (https://pubs.usgs.gov/circ/1367/). Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aaron C. H., Daniel P., Robert A. Q., Protsyuk I., Frederick I. A., Ransome E., et al. (2017). Meta-mass shift chemical profiling of metabolomes from coral reefs. PNAS 11444), 11685–11690. doi: 10.1073/pnas.1710248114
Abdelhafez O. H., Ali T. F. S., Fahim J. R., Desoukey S. Y., Ahmed S., Behery F. A., et al. (2020). Anti-Inflammatory Potential of Green Synthesized Silver Nanoparticles of the Soft Coral Nephthea Sp. Supported by Metabolomics Analysis and Docking Studies. Int. J. Nanomedicine. 2815, 5345–5360. doi: 10.2147/IJN.S239513
Aeby G. S., Ushijima B., Campbell J. E., Jones S., Williams G. J., Meyer J. L., et al. (2019). Pathogenesis of a tissue loss disease affecting multiple species of corals along the Florida reef tract. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00678
Aeby G. S., Williams G. J., Franklin E. C., Haapkyla J., Harvell C. D., Neale S., et al. (2011). Growth anomalies on the coral genera acropora and porites are strongly associated with host density and human population size across the Indo-Pacific. PloS One 6, e16887. doi: 10.1371/journal.pone.0016887
Aguilar C., Raina J.-B., Fôret S., Hayward D. C., Lapeyre B., Bourne D. G., et al. (2019). Transcriptomic analysis reveals protein homeostasis breakdown in the coral Acropora millepora during hypo-saline stress. BMC Genomics 20, 148. doi: 10.1186/s12864-019-5527-2
Aguilar C., Raina J.-B., Motti C. A., Fôret S., Hayward D. C., Lapeyre B., et al. (2017). Transcriptomic analysis of the response of Acropora millepora to hypo-osmotic stress provides insights into DMSP biosynthesis by corals. BMC Genomics 18, 612. doi: 10.1186/s12864-017-3959-0
Ainsworth T. D., Krause L., Bridge T., et al (2015). The coral core microbiome identifies rare bacterial taxa as ubiquitous endosymbionts. ISME J 9, 2261–2274. doi: 10.1038/ismej.2015.39
Ainsworth T. D., Kvennefors E. C., Blackall L. L., Fine M., Hoegh-Guldberg O. (2007). Disease and cell death in white syndrome of acroporid corals on the great barrier reef. Mar. Biol. 151, 19–29. doi: 10.1007/s00227-006-0449-3
Alvarez-Filip L., Estrada-Saldívar N., Pérez-Cervantes E., Molina-Hernández A., González-Barrios F. J. (2019). A rapid spread of the stony coral tissue loss disease outbreak in the Mexican Caribbean. PeerJ 7, e8069. doi: 10.7717/peerj.8069
Anderson D. A., Walz M. E., Weil E., Tonellato P., Smith M. C. (2016). RNA-Seq of the Caribbean reef-building coral Orbicella faveolata (Scleractinia-merulinidae) under bleaching and disease stress expands models of coral innate immunity. PeerJ 4, e1616. doi: 10.7717/peerj.1616
Andersson E. R., Day R. D., Work T. M., Anderson P. E., Woodley C. M., and Schock T. B, et al (2021). Identifying metabolic alterations associated with coral growth anomalies using 1H NMR metabolomics. Coral Reefs 40, 1195–1209. doi: 10.1007/s00338-021-02125-7
Andersson E.R., Stewart J.A., Work T.M., Woodley C.M., Schock T.B., and Day R.D, et al (2020). Morphological, elemental, and boron isotopic insights into pathophysiology of diseased coral growth anomalies. Sci Rep 10, 8252. doi: 10.1038/s41598-020-65118-6
Andersson E. R., Day R. D., Loewenstein J. M., Woodley C. M., Schock T. B. (2019). Evaluation of sample preparation methods for the analysis of reef-building corals using ¹H-NMR-based metabolomics. Metabolites. 9 (2), 32. doi: 10.3390/metabo9020032
Bailey M. H., Tokheim C., Porta-Pardo E., Sengupta S., Bertrand D., Weerasinghe A., et al. (2018). Comprehensive characterization of cancer driver genes and mutations. Cell 173, 371–385.e18. doi: 10.1016/j.cell.2018.02.060
Baker A. C. (2004). Symbiont diversity on coral reefs and its relationship to bleaching resistance and resilience. In: Rosenberg E., Loya Y. (eds) Coral Health Dis., 177–194. doi: 10.1007/978-3-662-06414-6_8
Barshis D. J., Ladner J. T., Oliver T. A., Seneca F. O., Traylor-Knowles N., Palumbi S. R. (2013). Genomic basis for coral resilience to climate change. Proc. Natl. Acad. Sci. U. S. A. 110, 1387–1392. doi: 10.1073/pnas.1210224110
Bayer F. M. (1954). Anthozoa: alcyonaria. gulf of Mexico: its origin, waters and marine life. Fishery Bull. 89, 279–284.
Beng K. C., Corlett R. T. (2020). Applications of environmental DNA (eDNA) in ecology and conservation: opportunities, challenges and prospects. Biodivers. Conserv. 29, 2089–2121. doi: 10.1007/s10531-020-01980-0
Ben-Haim Y., Zicherman-Keren M., Rosenberg E. (2003). Temperature-regulated bleaching and lysis of the coral Pocillopora damicornis by the novel pathogen Vibrio coralliilyticus. Appl. Environ. Microbiol. 69, 4236 LP–4242. doi: 10.1128/AEM.69.7.4236-4242.2003
Bolger A. M., Lohse M., Usadel B. (2014). Trimmomatic: a flexible trimmer for illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Bollati V., Baccarelli A. (2010). Environmental epigenetics. Heredity 105, 105–112. doi: 10.1038/hdy.2010.2
Bourne D. G., Garren M., Work T. M., Rosenberg E., Smith G. W., Drew Harvell C. (2009). Microbial disease and the coral holobiont. Trends Microbiol. 17, 554–562. doi: 10.1016/j.tim.2009.09.004
Bosh T. V., Long P. Q. A. (2017). Comparison of the composition of wax ester molecular species of different coral groups (subclasses Hexacorallia and Octocorallia). Russ J. Mar Biol. 43, 471–478. doi: 10.1134/S1063074017060049
Brandt M. E., McManus J. W. (2009). Dynamics and impact of the coral disease white plague: insights from a simulation model. Dis. Aquat. Organ. 87, 117–133. doi: 10.3354/dao02137
Brown B. E. (1997). Coral bleaching: causes and consequences. Coral Reefs 16, S129–S138. doi: 10.1007/s003380050249
Brown B. E., Le Tissier M. D. A., Bythell J. C. (1995). Mechanisms of bleaching deduced from histological studies of reef corals sampled during a natural bleaching event. Mar. Biol. 122, 655–663. doi: 10.1007/BF00350687
Brown T., Rodriguez-Lanetty M. (2015). Defending against pathogens - immunological priming and its molecular basis in a sea anemone, cnidarian. Sci. Rep. 5, 17425. doi: 10.1038/srep17425
Bruckner A. W., Bruckner R. J. (2006). Consequences of yellow band disease (YBD) on Montastraea annularis (species complex) populations on remote reefs off Mona island, Puerto Rico. Dis. Aquat. Organ. 69, 67–73. doi: 10.3354/dao069067
Burge C. A., Mouchka M. E., Harvell C. D., Roberts S. (2013). Immune response of the Caribbean sea fan, Gorgonia ventalina, exposed to an aplanochytrium parasite as revealed by transcriptome sequencing. Front. Physiol. 4, 180. doi: 10.3389/fphys.2013.00180
Burns J. H. R., Rozet N. K., Takabayashi M. (2011). Morphology, severity, and distribution of growth anomalies in the coral, Montipora capitata, at Wai’ōpae, Hawai’i. Coral Reefs 30, 819–826. doi: 10.1007/s00338-011-0761-3
Carr R., Shen-Orr S. S., Borenstein E. (2013). Reconstructing the genomic content of microbiome taxa through shotgun metagenomic deconvolution. PloS Comput. Biol. 9, e1003292. doi: 10.1371/journal.pcbi.1003292
Carballeira N. M., Miranda C., Rodríguez A. D. (2002). Phospholipid fatty acid composition of Gorgonia mariae and Gorgonia ventalina. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 131 (1), 83–87. doi: 10.1016/s1096-4959(01)00495-x
Casas V., Kline D. I., Wegley L., Yu Y., Breitbart M., Rohwer F. (2004). Widespread association of a Rickettsiales-like bacterium with reef-building corals. Environ. Microbiol. 6, 1137–1148. doi: 10.1111/j.1462-2920.2004.00647.x
Ceh J., Kilburn M. R., Cliff J. B., Raina J.-B., van Keulen M., Bourne D.G. (2013). Nutrient cycling in early coral life stages: Pocillopora damicornis larvae provide their algal symbiont (Symbiodinium) with nitrogen acquired from bacterial associates. Ecol. Evol. 3 (8), 2393–2400.
Closek C. J., Sunagawa S., DeSalvo M. K., Piceno Y. M., DeSantis T. Z., Brodie E. L., et al. (2014). Coral transcriptome and bacterial community profiles reveal distinct yellow band disease states in Orbicella faveolata. ISME J. 82411–2422. doi: 10.1038/ismej.2014.85
Counsell C. W. W., Johnston E. C., Sale T. L. (2019). Colony size and depth affect wound repair in a branching coral. Mar. Biol. 166. doi: 10.1007/s00227-019-3601-6
Cowen L. J., Putnam H. M. (2022). Bioinformatics of corals: Investigating heterogeneous omics data from coral holobionts for insight into reef health and resilience. Annu. Rev. BioMed. Data Sci 5 (1), 205–231. doi: 10.1146/annurev-biodatasci-122120-030732
Cunning R., Silverstein R. N., Baker A. C. (2018). Symbiont shuffling linked to differential photochemical dynamics of symbiodinium in three Caribbean reef corals. Coral Reefs 37, 145–152. doi: 10.1007/s00338-017-1640-3
Dahlgren C., Pizarro V., Sherman K., Greene W., Oliver J. (2021). Spatial and temporal patterns of stony coral tissue loss disease outbreaks in the Bahamas. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.682114
Daniels C. A., Baumgarten S., Yum L. K., Michell C. T., Bayer T., Arif C., et al. (2015). Metatranscriptome analysis of the reef-building coral Orbicella faveolata indicates holobiont response to coral disease. Front. Mar. Sci. 2. doi: 10.3389/fmars.2015.00062
Darling E. S., Alvarez-Filip L., Oliver T. A., McClanahan T. R., Côté I. M. (2012). Evaluating life-history strategies of reef corals from species traits. Ecol. Lett. 15, 1378–1386. doi: 10.1111/j.1461-0248.2012.01861.x
Davies S. W., Marchetti A., Ries J. B., Castillo K. D. (2016). Thermal and pCO2 stress elicit divergent transcriptomic responses in a resilient coral. Front. Mar. Sci. 3. doi: 10.3389/fmars.2016.00112
Dénes J., Szabó E., Robinette S. L., Szatmári I., Szőnyi L., Kreuder J. G., et al. (2012). Metabonomics of newborn screening dried blood spot samples: a novel approach in the screening and diagnostics of inborn errors of metabolism. Anal. Chem. 84, 10113–10120. doi: 10.1021/ac302527m
Dennison C., Karp R., Weiler B., Goncalves A., del Campo J., Rosales S., et al. (2021). The role of algal symbionts (genus Breviolum) in the susceptibility of corals to stony coral tissue loss disease in south Florida (Miami FL: Florida DEP).
Deutsch J. M., Jaiyesimi O. A., Pitts K. A., Houk J., Ushijima B., Walker B. K., et al. (2021). Metabolomics of healthy and stony coral tissue loss disease affected Montastraea cavernosa corals. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.714778
Díaz-Pérez L., Rodríguez-Zaragoza F. A., Ortiz M., Cupul-Magaña A. L., Carriquiry J. D., Ríos-Jara E., et al. (2016). Correction: Coral reef health indices versus the biological, ecological and functional diversity of fish and coral assemblages in the Caribbean Sea. PloS One 11, e0167252. , 190–3. doi: 10.1371/journal.pone.0161812
Dixon G., Abbott E., Matz M. (2020). Meta-analysis of the coral environmental stress response: Acropora corals show opposing responses depending on stress intensity. Mol. Ecol. 29, 2855–2870. doi: 10.1111/mec.15535
Dobbelaere T., Muller E. M., Gramer L. J., Holstein D. M., Hanert E. (2020). Coupled epidemio-hydrodynamic modeling to understand the spread of a deadly coral disease in Florida. Front. Mar. Sci. 7, 591881 doi: 10.3389/fmars.2020.591881
Domart-Coulon I. J., Traylor-Knowles N., Peters E., Elbert D., Downs C. A., Price K., et al. (2006). Comprehensive characterization of skeletal tissue growth anomalies of the finger coral Porites compressa. Coral Reefs 25, 531–543. doi: 10.1007/s00338-006-0133-6
Downs C.A., Fauth J.E., Robinson C.E., Curry R., Lanzendorf B., Halas J.C., et al. (2005). Cellular diagnostics and coral health: Declining coral health in the Florida Keys. Marine Pollution Bulletin 51, 558–569. doi: 10.1016/j.marpolbul.2005.04.017
Downs C. A., Kramarsky-Winter E., Martinez J., Kushmaro A., Woodley C. M., Loya Y., et al. (2009). Symbiophagy as a cellular mechanism for coral bleaching. Autophagy 5, 211–216. doi: 10.4161/auto.5.2.7405
Downs C. A., Mueller E., Phillips S., Fauth J. E., Woodley C. M. (2000). A molecular biomarker system for assessing the health of coral (Montastraea faveolata) during heat stress. Mar. Biotechnol. 2, 533–544. doi: 10.1007/s101260000038
Drake J. L., Malik A., Popovits Y., Yosef O., Shemesh E., Stolarski J., et al. (2021). Physiological and transcriptomic variability indicative of differences in key functions within a single coral colony. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.685876
Duan K.-B., Rajapakse J. C., Wang H., Azuaje F. (2005). Multiple SVM-RFE for gene selection in cancer classification with expression data. IEEE Trans. Nanobioscience 4, 228–234. doi: 10.1109/tnb.2005.853657
Edge S. E., Shearer T. L., Morgan M. B., Snell T. W. (2013). Sub-lethal coral stress: Detecting molecular responses of coral populations to environmental conditions over space and time. Aquat. Toxicol. 128-129, 135–146. doi: 10.1016/j.aquatox.2012.11.014
Edmunds P. J., Yarid A. (2017). The effects of ocean acidification on wound repair in the coral Porites spp. J. Exp. Mar. Biol. Ecol. 486, 98–104. doi: 10.1016/j.jembe.2016.10.001
Eirin-Lopez J. M., Putnam H. M. (2019). Marine environmental epigenetics. Ann. Rev. Mar. Sci. 11, 335–368. doi: 10.1146/annurev-marine-010318-095114
Emery M. A., Dimos B. A., Mydlarz L. D. (2021). Cnidarian pattern recognition receptor repertoires reflect both phylogeny and life history traits. Front. Immunol. 12, 689463. doi: 10.3389/fimmu.2021.689463
Estrada-Saldívar N., Molina-Hernández A., Pérez-Cervantes E., Medellín-Maldonado F., González-Barrios F. J., Alvarez-Filip L. (2020). Reef-scale impacts of the stony coral tissue loss disease outbreak. Coral Reefs 39, 861–866. doi: 10.1007/s00338-020-01949-z
Farag M. A, Meyer A., Ali S. E., Salem M. A., Giavalisco P., Westphal H., et al (2018). Comparative Metabolomics Approach Detects Stress-Specific Responses during Coral Bleaching in Soft Corals. J Proteome Res 17 (6), 2060–2071. doi: 10.1021/acs.jproteome.7b00929
Ferrier-Pagès C., Houlbrèque F., Wyse E., Richard C., Allemand D., Boisson F. (2005). Bioaccumulation of zinc in the scleractinian coral Stylophora pistillata. Coral Reefs 24, 636–645. doi: 10.1007/s00338-005-0045-x
Florida Coral Disease Response Research & Epidemiology Team (2018) SCTLD case definition. Available at: https://floridadep.gov/sites/default/files/Copy%20of%20StonyCoralTissueLossDisease_CaseDefinition%20final%2010022018.pdf.
Formel N., Enochs I. C., Sinigalliano C., Anderson S. R., Thompson L. R. (2021). Subsurface automated samplers for eDNA (SASe) for biological monitoring and research. HardwareX 10, e00239. doi: 10.1016/j.ohx.2021.e00239
Frazier M., Helmkampf M., Bellinger M. R., Geib S. M., Takabayashi M. (2017). De novo metatranscriptome assembly and coral gene expression profile of Montipora capitata with growth anomaly. BMC Genomics 18, 710. doi: 10.1186/s12864-017-4090-y
Frazier M., Helmkampf M., Bellinger M. R., Geib S. M., Takabayashi M. (2017). De novo metatranscriptome assembly and coral gene expression profile of Montipora capitata with growth anomaly. BMC Genomics 18, 710. doi: 10.1186/s12864-017-4090-y
Fuess L. E., Mann W. T., Jinks L. R., Brinkhuis V., Mydlarz L. D. (2018). Transcriptional analyses provide new insight into the late-stage immune response of a diseased Caribbean coral. R. Soc. Open Sci. 5, 172062. doi: 10.1098/rsos.172062
Fuess L. E., Pinzón C J. H., Weil E., Grinshpon R. D., Mydlarz L. D. (2017). Life or death: disease-tolerant coral species activate autophagy following immune challenge. Proc. Biol. Sci. 284. doi: 10.1098/rspb.2017.0771
Galand PE, Chapron L, Meistertzheim A-L, Peru E, Lartaud F (2018). The effect of captivity on the dynamics of active bacterial communities differs between two deep-sea coral species. Frontiers in Microbiology 9. doi: 10.1126/science.1086050
Galloway S. B., Woodley C. M., McLaughlin S. M., Work T. M., Bochsler V. S., Meteyer C. U., et al. (2007) Coral disease and health workshop: Coral histopathology II. Available at: https://aquadocs.org/handle/1834/20045 (Accessed August 18, 2022).
Garcia G.D., Santos E.O., Sousa G.V., Zingali R.B., Thompson C.C., Thompson F.L, et al. (2016). Metaproteomics reveals metabolic transitions between healthy and diseased stony coral Mussismilia braziliensis. Mol. Ecol. 25, 4632–4644. doi: 10.1111/mec.13775
Gardner T. A., Côté I. M., Gill J. A., Grant A., Watkinson A. R. (2003). Long-term region-wide declines in Caribbean corals. Science 301, 958 LP–958960. doi: 10.1126/science.1086050
Garg N. (2021). Metabolomics in functional interrogation of individual holobiont members. mSystems 6, e0084121. doi: 10.1128/mSystems.00841-21
Geiser D. M., Taylor J. W., Ritchie K. B., Smith G. W. (1998). Cause of sea fan death in the West Indies. Nature 394, 137–138. doi: 10.1038/28079
Glynn P. W., Perez M., Sandra L. G. (1985). “Lipid Decline in Stressed Corals and Their Crustacean Symbionts.” Biol. Bull. 1682, 276–284. doi: 10.2307/1541240
Gochfeld D. J., Ankisetty S., Slattery M. (2015). Proteomic profiling of healthy and diseased hybrid soft corals Sinularia maxima × S. polydactyla. Dis. Aquat. Organisms 116, 133–141. doi: 10.3354/dao02910
Gordon B. R., Leggat W., Motti C. A. (2013). Extraction protocol for nontargeted NMR and LC-MS metabolomics-based analysis of hard coral and their algal symbionts. Methods Mol. Biol. 1055, 129–147. doi: 10.1007/978-1-62703-577-4_10
Gordon, Hannon (2010) Fastx-toolkit. FASTQ/A short-reads preprocessing tools. Available at: http://hannonlab.cshl.edu/fastx_toolkit/
Granados-Cifuentes C., Bellantuono A. J., Ridgway T., Hoegh-Guldberg O., Rodriguez-Lanetty M. (2013). High natural gene expression variation in the reef-building coral Acropora millepora: potential for acclimative and adaptive plasticity. BMC Genomics 14, 228. doi: 10.1186/1471-2164-14-228
Harland A. D., Davies P. S., Fixter L. M. (1992). Lipid content of some Caribbean corals in relation to depth and light. Marine Biol. 113, 357–361. doi: 10.1007/BF00349159
Harland A. D., Navarro J. C., Spencer D. P. (1993). Lipids of some Caribbean and Red Sea corals: total lipid, wax esters, triglycerides and fatty acids. Marine Biol. 117, 113–117. doi: 10.1007/BF00346432
Harriott V. J. (1993). Coral lipids and environmental stress. Environ. Monit Assess. 25 (2), 131–139. doi: 10.1007/BF00549134
Harriott V. J. (1993). Coral lipids and environmental stress. Environ. Monit. Assess. 25, 131–139. doi: 10.1007/BF00549134
Hartmann AC, Petras D, Quinn RA (2017). Meta-mass shift chemical profiling of metabolomes from coral reefs. Proceedings of the National Academy of Sciences. 114, 11685–11690
Harvell C. D., Mitchell C. E., Ward J. R., Altizer S., Dobson A. P., Ostfeld R. S., et al. (2002). Climate warming and disease risks for terrestrial and marine biota. Science 296, 2158–2162. doi: 10.1126/science.1063699
Henry N. L., Hayes D. F. (2012). Cancer biomarkers. Mol. Oncol. 6, 140–146. doi: 10.1016/j.molonc.2012.01.010
Heres M. M., Farmer B. H., Elmer F., Hertler H. (2021). Ecological consequences of stony coral tissue loss disease in the Turks and Caicos islands. Coral Reefs 40, 609–624. doi: 10.1007/s00338-021-02071-4
Hewson I., Button J. B., Gudenkauf B. M., Miner B., Newton A. L., Gaydos J. K., et al. (2014). Densovirus associated with sea-star wasting disease and mass mortality. Proc. Natl. Acad. Sci. U. S. A. 111, 17278–17283. doi: 10.1073/pnas.1416625111
Hillyer K. E., Dias D. A., Lutz A., Wilkinson S. P., Roessner U., Davy S. K. (2017). Metabolite profiling of symbiont and host during thermal stress and bleaching in the coral Acropora aspera. Coral Reefs 36, 105–118. doi: 10.1007/s00338-016-1508-y
Hillyer K. E., Dias D., Lutz A., Roessner U., Davy S. K.. (2018). 13C metabolomics reveals widespread change in carbon fate during coral bleaching. Metabolomics 14, 12. doi: 10.1007/s11306-017-1306-8
Hyman L. H. (1940). Observations and experiments on the physiology of medusae. Biol. Bull. 79, 282–296. doi: 10.2307/1537823
Imbs A. B., Dang L. T. P. (2017). The molecular species of phospholipids of the cold-water soft coral Gersemia rubiformis (Ehrenber) (Alcyonacea, Nephtheidae). Russ J. Mar Biol. 43, 239–244. doi: 10.1134/S1063074017030051
Imbs A. B., Dang L. P. T., Nguyen K. B. (2019). Comparative lipidomic analysis of phospholipids of hydrocorals and corals from tropical and cold-water regions. PloS One 14 (4), e0215759. doi: 10.1371/journal.pone.0215759
Imbs A. B., Dang L. P., Rybin V. G., Svetashev V. I. (2015). Fatty acid, lipid class, and phospholipid molecular species composition of the soft coral Xenia sp. (Nha Trang Bay, the South China Sea, Vietnam). Lipids. 50 (6), 575–589. doi: 10.1007/s11745-015-4021-0
Imbs A. B., Dang L. P. T., Viacheslav R., Nguyen N. T., Quan P. (2015). Distribution of very-long-chain fatty acids between molecular species of different phospholipid classes of two soft corals. Biochem. Analytical Biochem. 4, 205. doi: 10.4172/2161-1009.1000205
Imbs A. B., Demidkova D. A., Dautova T. N., Latyshev N. A. (2009). Fatty acid biomarkers of symbionts and unusual inhibition of tetracosapolyenoic acid biosynthesis in corals (octocorallia). Lipids. 44 (4), 325–335. doi: 10.1007/s11745-008-3266-2
Imbs A. B., Demidkova D. A., Latypov Y. Y., Pham L. Q. (2007). Application of fatty acids for chemotaxonomy of reef-building corals. Lipids. 42 (11), 1035–1046. doi: 10.1007/s11745-007-3109-6
Imbs A. B., Demina O. A., Demidkova D. A. (2006). Lipid class and fatty acid composition of the boreal soft coral Gersemia rubiformis. Lipids. 41 (7), 721–725. doi: 10.1007/s11745-006-5023-8
Imbs A., Latyshev N. (2012). Fatty acid composition as an indicator of possible sources of nutrition for soft corals of the genus Sinularia (Alcyoniidae). J. Marine Biol. Assoc. United Kingdom 92 (6), 1341–1347. doi: 10.1017/S0025315411001226
Imbs A. B., Latyshev N. A., Dautova T. N., Latypov Y. Y. (2010a). Distribution of lipids and fatty acids in corals by their taxonomic position and presence of zooxanthellae. Mar Ecol. Prog. Ser. 409, 65–75. doi: 10.3354/meps08622
Imbs A. B., Latyshev N. A., Zhukova N. V., Dautova T. N. (2007). Comparison of fatty acid compositions of azooxanthellate Dendronephthya and zooxanthellate soft coral species. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 148 (3), 314–321. doi: 10.1016/j.cbpb.2007.06.009
Imbs A. B., Yakovleva I. M. (2012). Dynamics of lipid and fatty acid composition of shallow-water corals under thermal stress: an experimental approach. Coral Reefs 31, 41–53. doi: 10.1007/s00338-011-0817-4
Imbs A. B., Yakovleva I. M., Latyshev N. A., Pham L. Q. (2010b). Biosynthesis of polyunsaturated fatty acids in zooxanthellae and polyps of corals. Russ J. Mar Biol. 36, 452–457. doi: 10.1134/S1063074010060076
Jackson J. B. C., Donovan M. K., Cramer K. L., Lam V. V. (2014). Status and trends of Caribbean coral reefs. Global coral reef Monit. network Switzerland:IUCN Gland, 1970–2012.
Kauffmann A., Gentleman R., Huber W. (2008). arrayQualityMetrics–a bioconductor package for quality assessment of microarray data. Bioinformatics 25, 415–416. doi: 10.1093/bioinformatics/btn647
Kellogg C. A., Piceno Y. M., Tom L. M., DeSantis T. Z., Gray M. A., Zawada D. G., et al. (2013). Comparing bacterial community composition between healthy and white plague-like disease states in Orbicella annularis using PhyloChip™ G3 microarrays. PloS One 8, e79801. doi: 10.1371/journal.pone.0079801
Kelley E.R., Sleith R.S., Matz M.V., Wright R. M. (2021). Gene expression associated with disease resistance and long-term growth in a reef-building coral. R. Soc. Open Sci. 8, 4. doi: 10.1098/rsos.210113
Kenkel C. D., Mocellin V. J. L., Bay L. K. (2020). Global gene expression patterns in Porites white patch syndrome: Disentangling symbiont loss from the thermal stress response in reef-building coral. Mol. Ecol. 29 (20), 3907–3920. doi: 10.1111/mec.15608
Kenkel C. D., Goodbody-Gringley G., Caillaud D., Davies S. W., Bartels E., Matz M. V. (2013). Evidence for a host role in thermotolerance divergence between populations of the mustard hill coral (Porites astreoides) from different reef environments. Mol. Ecol. 22, 4335–4348. doi: 10.1111/mec.12391
Kenkel C. D., Sheridan C., Leal M. C., Bhagooli R., Castillo K. D., Kurata N., et al. (2014). Diagnostic gene expression biomarkers of coral thermal stress. Mol. Ecol. Resour. 14, 667–678. doi: 10.1111/1755-0998.12218
Klinges J. G., Rosales S. M., McMinds R., Shaver E. C., Shantz A. A., Peters E. C., et al. (2019). Phylogenetic, genomic, and biogeographic characterization of a novel and ubiquitous marine invertebrate-associated Rickettsiales parasite, Candidatus Aquarickettsia rohweri, gen. nov., sp. nov. ISME J. 13, 2938–2953. doi: 10.1038/s41396-019-0482-0
Korneliussen T. S., Albrechtsen A., Nielsen R. (2014). ANGSD: Analysis of next generation sequencing data. BMC Bioinf. 15, 356. doi: 10.1186/s12859-014-0356-4
Kourou K., Exarchos T. P., Exarchos K. P., Karamouzis M. V., Fotiadis D. I. (2015). Machine learning applications in cancer prognosis and prediction. Comput. Struct. Biotechnol. J. 13, 8–17. doi: 10.1016/j.csbj.2014.11.005
Kramer P. A. (2003). Synthesis of coral reef health indicators for the Western Atlantic: Results of the AGRRA program, (1997-2000). Atoll Res. Bull. 496, 1–57. doi: 10.5479/si.00775630.496-3.1
Landsberg J. H., Yasunari K., Peters E. C., Wilson P. W., Perry N., Waters Y., et al. (2020). Stony coral tissue loss disease in Florida is associated with disruption of host–zooxanthellae physiology. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.576013
Langfelder P., Horvath S. (2008). WGCNA: an R package for weighted correlation network analysis. BMC Bioinf. 9, 559. doi: 10.1186/1471-2105-9-559
Latyshev N. A., Naumenko N. V., Svetashev V., Latypov ,. Y. Y. (1991). Fatty acids of reef-building corals. Marine Ecology-progress Ser. - MAR ECOL-PROGR Ser. 76, 295–301. doi: 10.3354/meps076295
LeBoeuf N., McLean C., Oliver C., Erickson M., Wohlegemuth M., Hann N. (2021). NOAA Blue economy, strategic plan 2021-2025 (NOAA).
Lee S. T. M., Davy S. K., Tang S.-L., Kench P. S. (2016). Mucus sugar content shapes the bacterial community structure in thermally stressed Acropora muricata. Front. Microbiol. 7, 371. doi: 10.3389/fmicb.2016.00371
Leray M., Knowlton N. (2015). DNA Barcoding and metabarcoding of standardized samples reveal patterns of marine benthic diversity. Proc. Natl. Acad. Sci. U. S. A. 112, 2076–2081. doi: 10.1073/pnas.1424997112
Lesser M. P., Jarett J. K. (2014). Culture-dependent and culture-independent analyses reveal no prokaryotic community shifts or recovery of Serratia marcescens in Acropora palmata with white pox disease. FEMS Microbiol. Ecol. 88, 457–467. doi: 10.1111/1574-6941.12311
Levy S., Elek A., Grau-Bové X., Menéndez-Bravo S., Iglesias M., Tanay A., et al. (2021). A stony coral cell atlas illuminates the molecular and cellular basis of coral symbiosis, calcification, and immunity. Cell 184, 2973–2987.e18. doi: 10.1016/j.cell.2021.04.005
Libro S., Kaluziak S. T., Vollmer S. V. (2013). RNA-Seq profiles of immune related genes in the staghorn coral Acropora cervicornis infected with white band disease. PloS One 8, e81821. doi: 10.1371/journal.pone.0081821
Libro S., Kaluziak S. T., Vollmer S. V. (2013). RNA-seq profiles of immune related genes in the staghorn coral Acropora cervicornis infected with white band disease. PloS One 8 (11), e81821. doi: 10.1371/journal.pone.0081821
Libro S., Vollmer S. V. (2016). Genetic Signature of Resistance to White Band Disease in the Caribbean Staghorn Coral Acropora cervicornis. PloS One 11 (1), e0146636. doi: 10.1371/journal.pone.0146636
Liew Y. J., Aranda M., Voolstra C. R. (2016). Reefgenomics.Org - a repository for marine genomics data. Database 2016, baw152. doi: 10.1093/database/baw152
Lima L.F.O., Weissman M., Reed M, Papudeshi B, Alker A. T., Morris M. M, et al (2020). Modeling of the coral microbiome: the influence of temperature and microbial network. mBio 11, e02691–02619.
Little M., George E. E., Arts M. G. I., Shivak J., Benler S., Huckeba J., et al. (2021). Three-dimensional molecular cartography of the Caribbean reef-building coral Orbicella faveolata. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.627724
Lohr K. E., Khattri R. B., Guingab-Cagmat J., Camp E. F., Merritt M. E., Garrett T. J., et al. (2019). Metabolomic profiles differ among unique genotypes of a threatened Caribbean coral. Sci. Rep. 9, 6067. doi: 10.1038/s41598-019-42434-0
Louis Y. D., Bhagooli R., Kenkel C. D., Baker A. C., Dyall S. D. (2017). Gene expression biomarkers of heat stress in scleractinian corals: Promises and limitations. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 191, 63–77. doi: 10.1016/j.cbpc.2016.08.007
Mallick H., Franzosa E.A., Mclver L.J., Banerjee S., Sirota-Madi A., Kostic A. D., et al. Predictive metabolomic profiling of microbial communities using amplicon or metagenomic sequences. Nat Commun (2019) 10, 3136. doi: 10.1038/s41467-019-10927-1
Marcos-Zambrano L. J., Karaduzovic-Hadziabdic K., Loncar Turukalo T., Przymus P., Trajkovik V., Aasmets O., et al. (2021). Applications of machine learning in human microbiome studies: A review on feature selection, biomarker identification, disease prediction and treatment. Front. Microbiol. 12, 634511. doi: 10.3389/fmicb.2021.634511
Mass T., Drake J. L., Peters E. C., Jiang W., Falkowski P. G. (2014). Immunolocalization of skeletal matrix proteins in tissue and mineral of the coral Stylophora pistillata. Proc. Natl. Acad. Sci. U. S. A. 111, 12728–12733. doi: 10.1073/pnas.1408621111
Matthews J. L., Cunning R., Ritson-Williams R., Oakley C. A., Lutz A., Roessner U., et al. (2020). Metabolite pools of the reef building coral Montipora capitata are unaffected by Symbiodiniaceae community composition. Coral Reefs 39, 1727–1737. doi: 10.1007/s00338-020-01999-3
Mayfield A. B. (2022). Machine-learning-based proteomic predictive modeling with thermally-challenged Caribbean reef corals. Diversity 14, 33. doi: 10.3390/d14010033
Mayfield A. B., Aguilar C., Kolodziej G., Enochs I. C., Manzello D. P. (2021). Shotgun proteomic analysis of thermally challenged reef corals. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.660153
Mayfield A. B., Fan T.-Y., Chen C.-S. (2013). Physiological acclimation to elevated temperature in a reef-building coral from an upwelling environment. Coral Reefs 32, 909–921. doi: 10.1007/s00338-013-1067-4
McRae C. J., Huang W.-B., Fan T.-Y., Côté I. M. (2021). Effects of thermal conditioning on the performance of Pocillopora acuta adult coral colonies and their offspring. Coral Reefs 40, 1491–1503. doi: 10.1007/s00338-021-02123-9
Meiling S. S., Muller E. M., Lasseigne D., Rossin A., Veglia A. J., MacKnight N., et al. (2021). Variable species responses to experimental stony coral tissue loss disease (SCTLD) exposure. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.670829
Menzel L. P., Tondo C., Stein B., Bigger C. H. (2015). Histology and ultrastructure of the coenenchyme of the octocoral Swiftia exserta, a model organism for innate immunity/graft rejection. Zoology 118, 115–124. doi: 10.1016/j.zool.2014.09.002
Meszaros A., Bigger C. (1999). Qualitative and quantitative study of wound healing processes in the Coelenterate, Plexaurella fusifera: Spatial, temporal, and environmental (light attenuation) influences. J. Invertebr. Pathol. 73, 321–331. doi: 10.1006/jipa.1999.4851
Meyer J. (2020) Development of probiotics and alternative treatments for stony coral tissue loss disease. Available at: https://floridadep.gov/sites/default/files/DEP%202020-2021_FinalReport_Meyer_508.pdf (Accessed August 2, 2022).
Meyer D., Dimitriadou E., Hornik K., Weingessel A., Leisch F. (2021) e1071: Misc functions of the department of statistics, probability theory group (Formerly: E1071). Available at: https://CRAN.R-project.org/package=e1071.
Meyer F., Hofmann P., Belmann P., Garrido-Oter R., Fritz A., Sczyrba A., et al. (2018). AMBER: Assessment of metagenome BinnERs. Gigascience 7 (6), giy069. doi: 10.1093/gigascience/giy069
Miller M. W., Colburn P. J., Pontes E., Williams D. E., Bright A. J., Serrano X. M., et al. (2019). Genotypic variation in disease susceptibility among cultured stocks of elkhorn and staghorn corals. PeerJ 7, e6751. doi: 10.7717/peerj.6751
Miller M. W., Lohr K. E., Cameron C. M., Williams D. E., Peters E. C. (2014). Disease dynamics and potential mitigation among restored and wild staghorn coral, Acropora cervicornis. PeerJ 2, e541. doi: 10.7717/peerj.541
Mohamed A. F., Achim M., Sara E. A., Mohamed A. S., Patrick G., Hildegard W., et al. (2018). Comparative metabolomics approach detects stress-specific responses during coral bleaching in soft corals. Wessjohann J. Proteome Res. 176), 2060–2071. doi: 10.1021/acs.jproteome.7b00929
Montilla L. M., Ascanio A., Verde A., Croquer A. (2019). Systematic review and meta-analysis of 50 years of coral disease research visualized through the scope of network theory. PeerJ 7, e7041. doi: 10.7717/peerj.7041
Morgan M., Goodner K., Ross J., Poole A. Z., Stepp E., Stuart C. H., et al. (2015). Development and application of molecular biomarkers for characterizing Caribbean Yellow Band Disease in Orbicella faveolata. PeerJ. 3, e1371. doi: 10.7717/peerj.1371
Mullen K. M., Peters E. C., Harvell C. D. (2004). “Coral resistance to disease,” in Coral health and disease. Eds. Rosenberg E., Loya Y. (Berlin, Heidelberg: Springer Berlin Heidelberg), 377–399.
Muller E. M., Bartels E., Baums I. B. (2018). Bleaching causes loss of disease resistance within the threatened coral species Acropora cervicornis. Elife 7, e35066. doi: 10.7554/eLife.35066.028
Muller E. M., Sartor C., Alcaraz N. I., Van Woesik R. (2020). Spatial epidemiology of the stony-coral-tissue-loss disease in Florida. Front. Mar. Sci 7:163. doi: 10.3389/fmars.2020.00163
Mydlarz L. D., Couch C. S., Weil E., Smith G. (2009). Immune defenses of healthy, bleached and diseased Montastraea faveolata during a natural bleaching event. Dis. Aquat. Organ. 87, 67–78. doi: 10.3354/dao02088
Mydlarz L. D., Harvell C. D. (2007). Peroxidase activity and inducibility in the sea fan coral exposed to a fungal pathogen. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 146, 54–62. doi: 10.1016/j.cbpa.2006.09.005
Mydlarz L. D., Holthouse S. F., Peters E. C., Harvell C. D. (2008). Cellular responses in sea fan corals: Granular amoebocytes react to pathogen and climate stressors. PloS One 3, e1811. doi: 10.1371/journal.pone.0001811
Mydlarz L. D., Jacobs R. S. (2006). An inducible release of reactive oxygen radicals in four species of gorgonian corals. Mar. Freshw. Behav. Physiol. 39, 143–152. doi: 10.1080/10236240600708512
Mydlarz L. D., Palmer C. V. (2011). The presence of multiple phenoloxidases in Caribbean reef-building corals. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 159, 372–378. doi: 10.1016/j.cbpa.2011.03.029
Nagelkerken I., Buchan K., Smith G. W., Bonair K., Bush P., Garzón-Ferreira J., et al. (1997). Widespread disease in Caribbean sea fans: II. patterns of infection and tissue loss. Mar. Ecol. Prog. Ser. 160, 255–263. doi: 10.3354/meps160255
Neely K. L., Macaulay K. A., Hower E. K., Dobler M. A. (2020). Effectiveness of topical antibiotics in treating corals affected by stony coral tissue loss disease. PeerJ 8, e9289. doi: 10.7717/peerj.9289
Ocampo I. D., Zárate-Potes A., Pizarro V., Rojas C. A., Vera N. E., Cadavid L. F. (2015). The immunotranscriptome of the Caribbean reef-building coral Pseudodiploria strigosa. Immunogenetics. 67 (9), 515–530. doi: 10.1007/s00251-015-0854-1
Ochsenkühn M. A., Schmitt-Kopplin P., Harir M., Amin S. A. (2018). Coral metabolite gradients affect microbial community structures and act as a disease cue. Commun. Biol. 1, 184. doi: 10.1038/s42003-018-0189-1
Oku H., Yamashiro H., Onaga K., et al. (2003). Seasonal changes in the content and composition of lipids in the coral Goniastrea aspera. Coral Reefs 22, 83–85. doi: 10.1007/s00338-003-0279-4
Palmer C. V., McGinty E. S., Cummings D. J., Smith S. M., Bartels E., Mydlarz L. D. (2011a). Patterns of coral ecological immunology: variation in the responses of Caribbean corals to elevated temperature and a pathogen elicitor. J. Exp. Biol. 214, 4240–4249. doi: 10.1242/jeb.061267
Palmer C. V., Modi C. K., Mydlarz L. D. (2009). Coral fluorescent proteins as antioxidants. PloS One 4, e7298. doi: 10.1371/journal.pone.0007298
Palmer C. V., Traylor-Knowles N. (2012). Towards an integrated network of coral immune mechanisms. Proc. R. Soc. B: Biol. Sci. 279, 4106–4114. doi: 10.1098/rspb.2012.1477
Palmer C. V., Traylor-Knowles N. G. (2018). “Cnidaria: Anthozoans in the hot seat,” in Advances in Comparative Immunology (Cham: Springer), 51–93.
Palmer C. V., Traylor-Knowles N. G., Willis B. L., Bythell J. C. (2011b). Corals use similar immune cells and wound-healing processes as those of higher organisms. PloS One 6, e23992. doi: 10.1371/journal.pone.0023992
Palumbi S. R., Barshis D. J., Traylor-Knowles N., Bay R. A. (2014). Mechanisms of reef coral resistance to future climate change. Science 344, 895–898. doi: 10.1126/science.1251336
Pantos O., Cooney R. P., Le Tissier M. D. A., Barer M. R., O’Donnell A. G., Bythell J. C. (2003). The bacterial ecology of a plague-like disease affecting the Caribbean coral Montastrea annularis. Environ. Microbiol. 5, 370–382. doi: 10.1046/j.1462-2920.2003.00427.x
Parkinson J. E., Baker A. C., Baums I. B., Davies S. W., Grottoli A., Kitchen S. A., et al. (2020). Molecular tools for coral reef restoration: beyond biomarker discovery. Conserv. Lett, 13:e12687. doi: 10.1111/conl.12687
Papina M., Meziane T., van Woesik R. (2007). Acclimation effect on fatty acids of the coral Montipora digitata and its symbiotic algae. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 147 (4), 583–589. doi: 10.1016/j.cbpb.2007.02.011
Patterson K. L., Porter J. W., Ritchie K. B., Polson S. W., Mueller E., Peters E. C., et al. (2002). The etiology of white pox, a lethal disease of the Caribbean elkhorn coral, Acropora palmata. Proc. Natl. Acad. Sci. 99, 8725 LP–8730. doi: 10.1073/pnas.092260099
Paul VJ, Ushijima B, Aeby G. (2019). Studies of the Ecology and Microbiology of Florida’s Coral Tissue Loss Diseases Miami, FL:Florida DEP, Pp 1–33.
Peixoto R. S., Sweet M., Villela H. D. M., Cardoso P., Thomas T., Voolstra C. R., et al. (2021). Coral probiotics: Premise, promise, prospects. Annu. Rev. Anim. Biosci. 9, 265–288. doi: 10.1146/annurev-animal-090120-115444
Pernice M., Meibom A., Van Den H. A., Kopp C., Domart-Coulon I., Hoegh-Guldberg O., et al. (2012). A single-cell view of ammonium assimilation in coral–dinoflagellate symbiosis. ISME J. 6, 1314–1324. doi: 10.1038/ismej.2011.196
Peters E. C. (2015). “Diseases of coral reef organisms,” in Coral reefs in the anthropocene. Ed. Birkeland C. (Dordrecht: Springer Netherlands), 147–178.
Petes L. E., Harvell C. D., Peters E. C., Webb M. A. H., Mullen K. M. (2003). Pathogens compromise reproduction and induce melanization in Caribbean sea fans. Mar. Ecol. Prog. Ser. 264, 167–171. doi: 10.3354/meps264167
Petrou K., Nielsen D. A., Heraud P. (2018). Single-cell biomolecular analysis of coral algal symbionts reveals opposing metabolic responses to heat stress and expulsion. Front. Mar. Sci. 5. doi: 10.3389/fmars.2018.00110
Pinzón C. J. H., Beach-Letendre J., Weil E., Mydlarz L. D. (2014). Relationship between phylogeny and immunity suggests older Caribbean coral lineages are more resistant to disease. PloS One 9, e104787. doi: 10.1371/journal.pone.0104787
Pollock F. J., Joseph Pollock F., Lamb J. B., Field S. N., Heron S. F., Schaffelke B., et al. (2014). Sediment and turbidity associated with offshore dredging increase coral disease prevalence on nearby reefs. PloS One 9, e102498. doi: 10.1371/journal.pone.0102498
Pollock F. J., Wada N., Torda G., Willis B. L., Bourne D. G. (2017). White syndrome-affected corals have a distinct microbiome at disease lesion fronts. Appl. Environ. Microbiol. 83, e02799-16. doi: 10.1128/AEM.02799-16
Precht W. F., Gintert B. E., Robbart M. L., Fura R., van Woesik R. (2016). Unprecedented disease-related coral mortality in southeastern Florida. Sci. Rep. 6, 31374. doi: 10.1038/srep31374
Quinn Robert A., Vermeij Mark J. A., Hartmann Aaron C., Galtier d'Auriac I., Benler S., Haas A., et al. (2016). Metabolomics of reef benthic interactions reveals a bioactive lipid involved in coral defence. Proc. R. Soc B., 2832016046920160469. doi: 10.1098/rspb.2016.0469
Reeves L. (1994). Newly discovered: yellow band disease strikes keys reefs. Underwater U.S.A. 11, 16.
Ricci C. A., Kamal A. H. M., Chakrabarty J. K., Fuess L. E., Mann W. T., Jinks L. R., et al. (2019). Proteomic investigation of a diseased gorgonian coral indicates disruption of essential cell function and investment in inflammatory and other immune processes. Integr. Comp. Biol. 59, 830–844. doi: 10.1093/icb/icz107
Richardson L., Goldberg W. M., Carlton R. G., Halas J. C. (1998). Coral disease outbreak in. tbe Florida keys: Plague type II. Rev. Biol. Trop. 46, 187–198.
Roach T. N. F., Dilworth J., Jones A. D., Quinn R. A., Drury C. (2021). Metabolomic signatures of coral bleaching history. Nat. Ecol. Evol. 5, 495–503. H, C. M. doi: 10.1038/s41559-020-01388-7
Roach T. N. F., Little M., Arts M. G. I., Huckeba J., Haas A. F., George E. E., et al. (2020). A multiomic analysis of in situ coral–turf algal interactions. PNAS 117 (24), 13588–13595. doi: 10.1073/pnas.1915455117
Rodriguez-Casariego J. A., Ladd M. C., Shantz A. A., Lopes C., Cheema M. S., Kim B., et al. (2018). Coral epigenetic responses to nutrient stress: Histone H2A.X phosphorylation dynamics and DNA methylation in the staghorn coral Acropora cervicornis. Ecol. Evol. 8, 12193–12207. doi: 10.1002/ece3.4678
Rodríguez-Casariego J. A., Mercado-Molina A. E., Garcia-Souto D., Ortiz-Rivera I. M., Lopes C., Baums I. B., et al. (2020). Genome-wide DNA methylation analysis reveals a conserved epigenetic response to seasonal environmental variation in the staghorn coral Acropora cervicornis. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.560424
Rodríguez-Villalobos J. C., Work T. M., Calderon-Aguilera L. E. (2016). Wound repair in Pocillopora. J. Invertebr. Pathol. 139, 1–5. doi: 10.1016/j.jip.2016.07.002
Rogers C. S. (2013). Coral reef resilience through biodiversity. ISRN Oceanogr 2013, 1–18. doi: 10.5402/2013/739034
Rohwer F., Seguritan V., Azam F., Knowlton N. (2002). Diversity and distribution of coral-associated bacteria. Mar Ecol Prog Ser 243, 1–10. doi: 10.3354/meps243001
Rosales S. M., Miller M. W., Williams D. E., Traylor-Knowles N., Young B., Serrano X. M. (2019). Microbiome differences in disease-resistant vs. susceptible acropora corals subjected to disease challenge assays. Sci. Rep. 9, 18279. doi: 10.1038/s41598-019-54855-y
Rosales S. M., Clark A. S., Huebner L. K., Ruzicka R. R., Muller E. M. (2020). Rhodobacterales and rhizobiales are associated with stony coral tissue loss disease and its suspected sources of transmission. Front. Microbiol. 11, 681. doi: 10.3389/fmicb.2020.00681
Rosenau N. A., Gignoux-Wolfsohn S., Everett R. A., Miller A. W., Minton M. S., Ruiz G. M. (2021). Considering commercial vessels as potential vectors of stony coral tissue loss disease. Front. Mar. Science 8, 709764 doi: 10.3389/fmars.2021.709764
Rosental B., Kozhekbaeva Z., Fernhoff N., Tsai J. M., Traylor-Knowles N. (2017). Coral cell separation and isolation by fluorescence-activated cell sorting (FACS). BMC Cell Biol. 18, 1–30. doi: 10.1186/s12860-017-0146-8
Rosset S., Koster G., Brandsma J., Hunt A. N., Postle A. D., D’Angelo C. (2019). Lipidome analysis of symbiodiniaceae reveals possible mechanisms of heat stress tolerance in reef coral symbionts. Coral Reefs 38, 1241–1253. doi: 10.1007/s00338-019-01865-x
Roth L., Kramer P., Doyle E., O’Sullivan C. (2020) Caribbean SCTLD dashboard. Available at: www.agrra.org.
Rowan R. (2004). Coral bleaching: thermal adaptation in reef coral symbionts. Nature 430, 742. doi: 10.1038/430742a
Schutte V. G. W., Selig E. R., Bruno J. F. (2010). Regional spatio-temporal trends in Caribbean coral reef benthic communities. Mar. Ecol. Prog. Ser. 402, 115–122. doi: 10.3354/meps08438
Sebé-Pedrós A., Saudemont B., Chomsky E., Plessier F., Mailhé M.-P., Renno J., et al. (2018). Cnidarian cell type diversity and regulation revealed by whole-organism single-cell RNA-seq. Cell 173, 1520–1534.e20. doi: 10.1016/j.cell.2018.05.019
Seemann J., Sawall Y., Auel H., Richter C. (2013). The use of lipids and fatty acids to measure the trophic plasticity of the coral Stylophora subseriata. Lipids 48, 275–286. doi: 10.1007/s11745-012-3747-1
Sharp W. C., Shea C. P., Maxwell K. E., Muller E. M., Hunt J. H. (2020). Evaluating the small-scale epidemiology of the stony-coral -tissue-loss-disease in the middle Florida keys. PloS One 15, e0241871. doi: 10.1371/journal.pone.0241871
Shore A., Caldwell J. M. (2019). Modes of coral disease transmission: how do diseases spread between individuals and among populations? Mar. Biol. 166. doi: 10.1007/s00227-019-3490-8
Sikorskaya T. V., Efimova K. V., Imbs A. B. (2021). Lipidomes of phylogenetically different symbiotic dinoflagellates of corals. Phytochemistry 181, 112579. doi: 10.1016/j.phytochem.2020.112579
Santacruz L., Hurtado D. X., Doohan R., et al. (2020). Metabolomic study of soft corals from the Colombian Caribbean: PSYCHE and 1H-NMR comparative analysis. Sci. Rep. 10, 5417. doi: 10.1038/s41598-020-62413-0
Seveso D., Montano S., Reggente M. A., Maggioni D., Orlandi I., Galli P., et al. (2017). The cellular stress response of the scleractinian coral Goniopora columna during the progression of the black band disease. Cell Stress Chaperones. 22 (2), 225–236. doi: 10.1007/s12192-016-0756-7
Shinzato C., Shoguchi E., Kawashima T., Hamada M., Hisata K., Tanaka M., et al. (2011). Using the Acropora digitifera genome to understand coral responses to environmental change. Nature 476, 320–323. doi: 10.1038/nature10249
Sikorskaya T. V., Ermolenko E. V., Imbs A. B. (2020). Effect of experimental thermal stress on lipidomes of the soft coral Sinularia sp. and its symbiotic dinoflagellates. J. Exp. Mar. Bio. Ecol. 524, 151295. doi: 10.1016/j.jembe.2019.151295
Sikorskaya T. V., . Ermolenko E.V., Imbs A. B. (2020). Effect of experimental thermal stress on lipidomes of the soft coral Sinularia sp. and its symbiotic dinoflagellates. J. Exp. Marine Biol. Ecol. 524, 51295. doi: 10.1016/j.jembe.2019.151295
Sikorskaya T. V., Imbs A. B. (2018). Study of total lipidome of the Sinularia siaesensis soft coral. Russ J. Bioorg Chem. 44, 712–723. doi: 10.1134/S1068162019010151
Silverstein R. N., Cunning R., Baker A. C. (2014). Change in algal symbiont communities after bleaching, not prior heat exposure, increases heat tolerance of reef corals. Glob. Change Biol. 21, 236–249. doi: 10.1111/gcb.12706
Silva-Lima A. W., Froes A. M., Garcia G. D., Tonon L. A. C., Swings J., Cosenza C. A. N., et al. (2021). Mussismilia braziliensis white plague disease is characterized by an affected coral immune system and dysbiosis. Microb. Ecol. 81, 795–806. doi: 10.1007/s00248-020-01588-5
Slattery M., Renegar D. A., Gochfeld D. J. (2013). Direct and indirect effects of a new disease of alcyonacean soft corals. Coral Reefs 32, 879–889. doi: 10.1007/s00338-013-1035-z
Snyder G. A., Browne W. E., Traylor-Knowles N., Rosental B. (2020). Fluorescence-activated cell sorting for the isolation of scleractinian cell populations. J. Vis. Exp. 159, e60446. doi: 10.3791/60446
Snyder G. A., Eliachar S., Connelly M. T., Talice S., Hadad U., Gershoni-Yahalom O., et al. (2021). Functional characterization of hexacorallia phagocytic cells. Front. Immunol. 12, 2402. doi: 10.3389/fimmu.2021.662803
Sogin E. M., Putnam H. M., Anderson P. E., Gates R. D. (2016). Metabolomic signatures of increases in temperature and ocean acidification from the reef-building coral, Pocillopora damicornis. Metabolomics 12. doi: 10.1007/s11306-016-0987-8
Sogin E. M., Anderson P., Williams P., Chen C. S., Gates R. D. (2014). Application of 1H-NMR metabolomic profiling for reef-building corals. PloS One 9 (10), e111274. doi: 10.1371/journal.pone.0111274
Spies NP, Takabayashi M (2013). (2013) Expression of galaxin and oncogene homologs in growth anomaly in the coral Montipora capitata. Dis Aquat Org 104, 249–256. doi: 10.3354/dao02603
Studivan M. S., Rossin A. M., Rubin E., Soderberg N., Holstein D. M., Enochs I. C. (2022). Reef sediments can act as a stony coral tissue loss disease vector. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.815698
Studivan M. S., Shatters A., Dodge D. L., Beal J. L., Voss J. D. (2021). Synergistic effects of thermal stress and estuarine discharge on transcriptomic variation of Montastraea cavernosa corals in southeast Florida. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.662220
Sun J., Chen Q., Lun J. C. Y., Xu J., Qiu J.-W. (2013). PcarnBase: development of a transcriptomic database for the brain coral Platygyra carnosus. Mar. Biotechnol. 15, 244–251. doi: 10.1007/s10126-012-9482-z
Sun A., Lam C., Wong D. A. (2012). Expanded newborn screening for inborn errors of metabolism: overview and outcomes. Adv. Pediatr. 59, 209–245. doi: 10.1016/j.yapd.2012.04.013
Sussman M., Willis B. L., Victor S., Bourne D. G. (2008). Coral pathogens identified for white syndrome (WS) epizootics in the Indo-Pacific. PloS One 3, e2393. doi: 10.1371/journal.pone.0002393
Sutherland K. P., Porter J. W., Torres C. (2004). Disease and immunity in Caribbean and Indo-Pacific zooxanthellate corals. Mar. Ecol. Prog. Ser. 266, 273–302. doi: 10.3354/meps266273
Sutherland K. P., Shaban S., Joyner J. L., Porter J. W., Lipp E. K. (2011). Human pathogen shown to cause disease in the threatened eklhorn coral Acropora palmata. PloS One 6, e23468. doi: 10.1371/journal.pone.0023468
Sweet M., Bulling M., Varshavi D., Lloyd G. R., Jankevics A., Najdekr L., et al. (2021). Species-specific variations in the metabolomic profiles of Acropora hyacinthus and Acropora millepora mask acute temperature stress effects in adult coral colonies. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.574292
Takabayashi M., Gregg T. M., Farah E., Burns J., Teves K., Cody N. H. (2010). The prevalence of skeletal growth anomaly and other afflictions in scleractinian corals at Wai ‘opae, Hawai ‘i. Proc. 11th Int. Coral Reef Symp, 820–824.
Takagi T., Yoshioka Y., Zayasu Y., Satoh N., Shinzato C. (2020). Transcriptome analyses of immune system behaviors in primary polyp of coral Acropora digitifera exposed to the bacterial pathogen Vibrio coralliilyticus under thermal loading. Mar Biotechnol. (NY). 22 (6), 748–759. doi: 10.1007/s10126-020-09984-1
Tang C. H., Ku P. C., Lin C. Y., Chen T. H., Lee K. H., Lee S. H., et al. (2015). Intra-colonial functional differentiation-related modulation of the cellular membrane in a pocilloporid coral Seriatopora caliendrum. Mar Biotechnol. (NY). 17 (5), 633–643. doi: 10.1007/s10126-015-9645-9
Tang C. H., Lin C. Y., Lee S. H., Wang W. H. (2017). Membrane lipid profiles of coral responded to zinc oxide nanoparticle-induced perturbations on the cellular membrane. Aquat Toxicol. 187, 72–81. doi: 10.1016/j.aquatox.2017.03.021
Tang C. H., Lin C. Y., Sun P. P., Lee S. H., Wang W. H. (2018). Modeling the effects of Irgarol 1051 on coral using lipidomic methodology for environmental monitoring and assessment. Sci. Total Environ. 627, 571–578. doi: 10.1016/j.scitotenv.2018.01.276
Tang C.-H., Shi S.-H., Lin C.-Y., LI H.-H., Wang W.-H. (2019). Using lipidomic methodology to characterize coral response to herbicide contamination and develop an early biomonitoring model. Sci. Total Environ. 648, 1275–1283. doi: 10.1016/j.scitotenv.2018.08.296
Tchernov D., Gorbunov M. Y., de Vargas C., Narayan Yadav S., Milligan A. J., Häggblom M., et al. (2004). Membrane lipids of symbiotic algae are diagnostic of sensitivity to thermal bleaching in corals. Proc. Natl. Acad. Sci. U. S. A. 101, 13531–13535. doi: 10.1073/pnas.0402907101
Tisthammer K. H., Timmins-Schiffman E., Seneca F. O., Nunn B. L., Richmond R. H. (2021). Physiological and molecular responses of lobe coral indicate nearshore adaptations to anthropogenic stressors. Sci. Rep. 11. doi: 10.1038/s41598-021-82569-7
Tracy A. M., Weil E., Burge C. A. (2020). Ecological factors mediate immunity and parasitic co-infection in sea fan octocorals. Front. Immunol. 11, 608066. doi: 10.3389/fimmu.2020.608066
Tracy A.M., Weil E., Burge C. A (2021). Ecological factors mediate Immunity and parasitic co-infection in sea fan octocorals. Front Immunol, 11–824. doi: 10.3389/fimmu.2020.608066
Traylor-Knowles N., Granger B. R., Lubinski T. J., Parikh J. R., Garamszegi S., Xia Y., et al. (2011). Production of a reference transcriptome and transcriptomic database (Pocillopora Base) for the cauliflower coral, Pocillopora damicornis. BMC Genomics 12, 585. doi: 10.1186/1471-2164-12-585
Traylor-Knowles N., Palumbi S. R. (2014). Translational environmental biology: Cell biology informing conservation. Trends Cell Biol. 24, 265–267. doi: 10.1016/j.tcb.2014.03.001
Traylor-Knowles N. (2016). Distinctive wound-healing characteristics in the corals Pocillopora damicornis and Acropora hyacinthus found in two different temperature regimes. Mar. Biol. 163. doi: 10.1007/s00227-016-3011-y
Traylor-Knowles N., Rose N.H., Sheets E.A., Palumbi S.R (2017). Early transcriptional responses during heat stress in the coral Acropora hyacinthus. The Biological Bulletin 2017 232(2), 91–100
Traylor-Knowles N., Rose N. H., Palumbi S. R. (2017). The cell specificity of gene expression in the response to heat stress in corals. J. Exp. Biol. 220, 1837–1845. doi: 10.1242/jeb.155275
Traylor-Knowles N. (2018). In situ hybridization techniques for paraffin-embedded adult coral samples. J. Vis. Exp. 138, e57853. doi: 10.3791/57853
Traylor-Knowles N., Connelly M. T., Young B. D., Eaton K., Muller E. M., Paul V. J., et al. (2021). Gene expression response to stony coral tissue loss disease transmission in M. cavernosa and O. faveolata from Florida. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.681563
Treignier C., Grover R., Ferrier-Pagés C., Tolosa I. (2008). Effect of light and feeding on the fatty acid and sterol composition of zooxanthellae and host tissue isolated from the scleractinian coral Turbinaria reniformis. Limnology Oceanography 53. doi: 10.4319/lo.2008.53.6.2702
Ushijima P., Aeby (2019). Studies of the ecology and microbiology of Florida’s coral tissue loss diseases (Miami, FL: Florida DEP). Available at: https://floridadep.gov/sites/default/files/Paul%20Ushijima%20Aeby_Final%20Report_508_2019.June:0.pdf.
Ushijima B., Meyer J. L., Thompson S., Pitts K., Marusich M. F., Tittl J., et al. (2020). Disease diagnostics and potential coinfections by Vibrio coralliilyticus during an ongoing coral disease outbreak in Florida. Front. Microbiol 11, 569354. doi: 10.3389/fmicb.2020.569354
Ushijima B., Videau P., Burger A. H., Shore-Maggio A., Runyon C. M., Sudek M., et al. (2014). Vibrio coralliilyticus strain OCN008 is an etiological agent of acute Montipora white syndrome. Appl. Environ. Microbiol. 80. doi: 10.1128/AEM.03463-13
van de Water J. A. J. M., Chaib De Mares M., Dixon G. B., Raina J.-B., Willis B. L., Bourne D. G., et al. (2018). Antimicrobial and stress responses to increased temperature and bacterial pathogen challenge in the holobiont of a reef-building coral. Mol. Ecol. 27, 1065–1080. doi: 10.1111/mec.14489
Vega Thurber R., Mydlarz L. D., Brandt M., Harvell D., Weil E., Raymundo L., et al. (2020). Deciphering coral disease dynamics: Integrating host, microbiome, and the changing environment. Front. Ecol. Evol. 8. doi: 10.3389/fevo.2020.575927
Veglia A. J., Beavers K., Van Buren E. W., Meiling S. S., Muller E. M., Smith T. B., et al. (2022). Alphaflexivirus genomes in stony coral tissue loss disease-affected, disease-exposed, and disease-unexposed coral colonies in the U.S. Virgin Islands. Microbiol. Resour. Announc. 11, e0119921. doi: 10.1128/mra.01199-21
Vidal-Dupiol J., Dheilly N. M., Rondon R., Grunau C., Cosseau C., Smith K. M, et al. (2014). Thermal stress triggers broad Pocillopora damicornis transcriptomic remodeling, while Vibrio coralliilyticus infection induces a more targeted immuno-suppression response. PloS One 9 (9), e107672. doi: 10.1371/journal.pone.0107672
Vohsen S. A., Fisher C. R., Baums I. B. (2019). Metabolomic richness and fingerprints of deep-sea coral species and populations. Metabolomics 15, 34. doi: 10.1007/s11306-019-1500-y
Wall C. B., Ricci C. A., Foulds G. E., Mydlarz L. D., Gates R. D., Putnam H. M. (2018). The effects of environmental history and thermal stress on coral physiology and immunity. Mar. Biol. 165, 56. doi: 10.1007/s00227-018-3317-z
Weiss Y., Forêt S., Hayward D. C., Ainsworth T., King R., Ball E. E., et al. (2013). The acute transcriptional response of the coral Acropora millepora to immune challenge: expression of GiMAP/IAN genes links the innate immune responses of corals with those of mammals and plants. BMC Genomics 14, 400. doi: 10.1186/1471-2164-14-400
Wiedenmann J., D’Angelo C., Smith E., Hunt A. N., Legiret F. E., Postle A. D., et al. (2013). Nutrient enrichment can increase the susceptibility of reef corals to bleaching. Nat. Clim Change 3, 160–164. doi: 10.1038/nclimate1661
Wright R. M., Aglyamova G. V., Meyer E., et al. (2015). Gene expression associated with white syndromes in a reef building coral, Acropora hyacinthus. BMC Genomics 16, 371. doi: 10.1186/s12864-015-1540-2
Weis V. M. (2008). Cellular mechanisms of cnidarian bleaching: stress causes the collapse of symbiosis. J. Exp. Biol. 211, 3059–3066. doi: 10.1242/jeb.009597
Weizman E., Levy O. (2019). The role of chromatin dynamics under global warming response in the symbiotic coral model Aiptasia. Commun. Biol. 2, 282. doi: 10.1038/s42003-019-0543-y
West K. M., Adam A. A. S., White N., Robbins W. D., Barrow D., Lane A., et al. (2022). The applicability of eDNA metabarcoding approaches for sessile benthic surveying in the Kimberley region, north-western Australia. Environ. DNA 4, 34–49. doi: 10.1002/edn3.184
West K. M., Stat M., Harvey E. S., Skepper C. L., DiBattista J. D., Richards Z. T., et al. (2020). eDNA metabarcoding survey reveals fine-scale coral reef community variation across a remote, tropical island ecosystem. Mol. Ecol. 29, 1069–1086. doi: 10.1111/mec.15382
Wong Y.-H., Zhang Y., Lun J. C. Y., Qiu J.-W. (2021). A proteomic analysis of skeletal tissue anomaly in the brain coral Platygyra carnosa. Mar. pollut. Bull. 164, 111982. doi: 10.1016/j.marpolbul.2021.111982
Work T. M., Aeby G. S. (2010). Wound repair in Montipora capitata. J. Invertebrate Pathol. 105, 116–119. doi: 10.1016/j.jip.2010.05.009
Work T. M., Aeby G. S., Hughen K. A. (2016). Gross and microscopic lesions in corals from Micronesia. Vet. Pathol. 53, 153–162. doi: 10.1177/0300985815571669
Work T. M., Kaczmarsky L. T., Peters E. C. (2015). Skeletal growth anomalies in corals. Dis. Coral, 291–299. doi: 10.1002/9781118828502.ch20
Work T., Meteyer C. (2014). To understand coral disease, look at coral cells. Ecohealth 11, 610–618. doi: 10.1007/s10393-014-0931-1
Work T. M., Weatherby T. M., Landsberg J. H., Kiryu Y., Cook S. M., Peters E. C. (2021). Viral-like particles are associated with endosymbiont pathology in Florida corals affected by stony coral tissue loss disease. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.750658
Wright R. M., Aglyamova G. V., Meyer E., Matz M. V. (2015). Gene expression associated with white syndromes in a reef building coral, Acropora hyacinthus. BMC Genomics 16, 371. doi: 10.1186/s12864-015-1540-2
Wright R. M., Correa A. M. S., Quigley L. A., Santiago-Vázquez L. Z., Shamberger K. E. F., Davies S. W. (2019). Gene expression of endangered coral (Orbicella spp.) in flower garden banks national marine sanctuary after hurricane Harvey. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00672
Wright R. M., Kenkel C. D., Dunn C. E., Shilling E. N., Bay L. K., Matz M. V. (2017). Intraspecific differences in molecular stress responses and coral pathobiome contribute to mortality under bacterial challenge in Acropora millepora. Sci. Rep. 7, 2609. doi: 10.1038/s41598-017-02685-1
Yamashiro H., Oku H., Higa H., Chinen I., Sakai K. (2019). Composition of lipids, fatty acids and sterols in Okinawan corals, Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology 1224, 397–407. doi: 10.1016/S0305-0491(99)00014-0
Young B. D., Serrano X. M., Rosales S. M., Miller M. W., Williams D., Traylor-Knowles N. (2020). Innate immune gene expression in Acropora palmata is consistent despite variance in yearly disease events. PloS One 15, e0228514. doi: 10.1371/journal.pone.0228514
Yuan C., Zhou Z., Zhang Y., Chen G., Yu X., Ni X., et al (2017). Effects of elevated ammonium on the transcriptome of the stony coral Pocillopora damicornis. Mar. Pollut. Bull. 114 (1), 46–52. doi: 10.1016/j.marpolbul.2016.08.036
Zaneveld J. R., McMinds R., Vega Thurber R. (2017). Stress and stability: applying the Anna Karenina principle to animal microbiomes. Nat. Microbiol. 2, 17121. doi: 10.1038/nmicrobiol.2017.121
Zhang Y., Chen Q., Xie J. Y., Yeung Y. H., Xiao B., Liao B., et al. (2019). Development of a transcriptomic database for 14 species of scleractinian corals. BMC Genomics 20, 387. doi: 10.1186/s12864-019-5744-8
Zhang Y., Sun J., Mu H., Lun J. C. Y., Qiu J.-W. (2017). Molecular pathology of skeletal growth anomalies in the brain coral Platygyra carnosa: A meta-transcriptomic analysis. Mar. pollut. Bull. 124, 660–667. doi: 10.1016/j.marpolbul.2017.03.047
Zhao L.-Y., Song J., Liu Y., Song C.-X., Yi C. (2020). Mapping the epigenetic modifications of DNA and RNA. Protein Cell 11, 792–808. doi: 10.1007/s13238-020-00733-7
Keywords: genomics, transcriptomics, coral disease, immunity, ‘omics, coral immunity, biomarker
Citation: Traylor-Knowles N, Baker AC, Beavers KM, Garg N, Guyon JR, Hawthorn A, MacKnight NJ, Medina M, Mydlarz LD, Peters EC, Stewart JM, Studivan MS and Voss JD (2022) Advances in coral immunity ‘omics in response to disease outbreaks. Front. Mar. Sci. 9:952199. doi: 10.3389/fmars.2022.952199
Received: 24 May 2022; Accepted: 26 August 2022;
Published: 07 October 2022.
Edited by:
Sarah Annalise Gignoux-Wolfsohn, Smithsonian Environmental Research Center (SI), United StatesReviewed by:
David Anderson, Washington University in St. Louis, United StatesCopyright © 2022 Traylor-Knowles, Baker, Beavers, Garg, Guyon, Hawthorn, MacKnight, Medina, Mydlarz, Peters, Stewart, Studivan and Voss. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nikki Traylor-Knowles, bmd0MTRAbWlhbWkuZWR1
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.