 Zhiping Feng
Zhiping Feng Wei Yu
Wei Yu Yang Zhang
Yang Zhang Yunkai Li1,2,3,4
Yunkai Li1,2,3,4- 1College of Marine Sciences, Shanghai Ocean University, Shanghai, China
- 2National Engineering Research Center for Oceanic Fisheries, Shanghai Ocean University, Shanghai, China
- 3Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Ministry of Education, Shanghai Ocean University, Shanghai, China
- 4Key Laboratory of Oceanic Fisheries Exploration, Ministry of Agriculture and Rural Affairs, Shanghai, China
- 5State Key Laboratory of Satellite Ocean Environment Dynamics, Second Institute of Oceanography, Ministry of Natural Resources, Hangzhou, China
The synchronous response of the spatial distribution pattern of Dosidicus gigas and Trachurus murphyi habitat along the Chilean waters to different-intensity El Niño events was examined based on the habitat suitability index (HSI) models inclusive with the key environmental factors of sea surface height anomaly (SSHA), sea surface salinity (SSS), water temperature at 400-m depth (Temp_400 m) for D. gigas, sea surface temperature (SST), mixed layer depth (MLD), Temp_400 m for T. murphyi, and Niño 1 + 2 index. Results showed that all the environmental factors except MLD were significantly correlated with the Niño 1 + 2 index. Compared with weak (WE) and moderate (ME) El Niño events, the suitable range of SSHA and SSS for D. gigas decreased, resulting in a decreased suitable habitat area and increased poor habitat area during the strong (SE) El Niño years. In space, the suitable habitat of D. gigas was mainly distributed in north-central Chile with an east–west strip pattern; it decreased significantly in the northwestern waters off Chile and moved southwestward under SE El Niño events. For T. murphyi, the ME El Niño events yielded a contracted suitable range of water temperature and MLD decreased, consequently leading to a decreased suitable habitat area and increased poor habitat area of T. murphyi compared to WE and SE El Niño events. The suitable habitat of T. murphyi was mainly distributed in the 42°–47°S area and varied non-significantly during the WE and SE El Niño events; however, it clearly decreased in the southwest area of Chile and shifted northeastward under ME El Niño events. Our results indicated that El Niño events with different intensities strongly affected the habitat pattern of D. gigas and T. murphyi off Chile by changing the regional marine environment.
Introduction
Climate variability drives the synchronous response of marine ecosystems at a global scale and their internal species resources. The synchronous variation of marine species in a marine ecosystem is largely explained by the driving force of climate variability on marine environmental elements in different time scales, which produces a synchronous impact on the abundance of marine species or the distribution of their habitat in marine areas with various spatial scales (Goberville et al., 2014; Oozeki et al., 2019). In the Southeast Pacific Ocean, the Humboldt Current System (HCS) is the eastern boundary upwelling system with the highest fish abundance in the world. Due to its geographical proximity to El Niño events, the HCS is vulnerable to El Niño-Southern Oscillation (ENSO) phenomena and the replacement of its steady state was always promoted by the combined impacts of a seasonal or permanent upwelling in different latitudes, polar undercurrents near the coast, and many other complexly various ocean currents (Penven et al., 2005; Chavez et al., 2008; Freon et al., 2009; Brochier et al., 2013; Gutiérrez et al., 2016; Mogollón and Calil, 2017). Driven by the change in the marine environment or climate events at different scales, changes in species interaction within the HCS were often accompanied by synchronous changes in their abundance level, habitat quality, and spatial distribution pattern. For example, Engraulis ringens and Sardinops sagax presented an interdecadal regime shift in the HCS ecosystem during the warm and cold periods in a synchronized way (Zhang and Xu, 2000; Chavez et al., 2003) . In addition, with the influences of various climatic and environmental conditions, the synchronicity of key species in HCS contributed to the annual variation of the total output in the ecosystem as a whole (Zhang and Xu, 2000; Li et al., 2016; Yang et al., 2019). Therefore, exploring the synchronous change of marine species is helpful to understanding and mastering the variations of ecosystem structure and function under the changing environments, which provide help on a scientific basis for the monitoring and evaluation of ecosystem quality.
The Chilean fishing ground on the western coast of South America is one of the vital fishing grounds in the world, with many economically important species inhabiting in this area such as jumbo flying squid Dosidicus gigas and Chilean jack mackerel Trachurus murphyi (Zhang and Xu, 2000). D. gigas stocks grow fast and are highly abundant in this area. They mainly feed on zooplankton, crustaceans, fish, and cephalopods. D. gigas is an important target species for distant-water fishery countries such as Japan, South Korea, and China (Hu et al., 2018; Yu et al., 2019). For T. murphyi off Chile, its annual catch follows Peruvian anchovy which is the highest in the world, so its fishery occupies an important position in the global marine fishery. Expect Peru, Chile, and other coastal countries, the main fishing countries also involve Japan, South Korea, the European Union, and China (Zhang and Xu, 2000; Gerlotto and Dioses, 2013; Li et al., 2016). Small T. murphyi are mainly clustered in the northern part of Chile, while large individuals mainly inhabit in south-central Chile, supporting fisheries in south-central Chile (Zhang and Xu, 2000; Xie et al., 2021; Feng et al., 2021b). D. gigas and T. murphyi are ecologically important pelagic species in the HCS, occupying the middle position of the trophic niche in this ecosystem; therefore, the climate-related habitat changes of the two species tend to directly affect other trophic-level species through ecological effects (Cury et al., 2000; Hu et al., 2018).
At present, however, some studies stated that the abundance and habitat suitability of D. gigas and T. murphyi in the Southeast Pacific Ocean were regulated by the El Niño event (Yang et al., 2019; Yu et al., 2019; Wen et al., 2020). However, the duration, intensity, and zonal variation characteristics of the El Niño event are particularly changeable, which have impacts on fish species to varying degrees (Liu and Xue, 2010; Yu et al., 2018). For example, Guo et al. (2018) found that compared with the medium-intensity El Niño events, the strong El Niño events decreased the temperature and increased the sea surface height on the main fishing ground of Scomber japonicus in the East China Sea, and suitable habitats of S. japonicus significantly were reduced, resulting in a sharp decrease in catch per unit effort (CPUE).
The Eighth Scientific Committee Report of the South Pacific Regional Fisheries Management Organization (SPRFMO) pointed out that, on the one hand, it is necessary to set the total allowable catch (TAC) and the investment in regulating the fishing efforts when managing the resources of D. gigas and T. murphyi in the southeast Pacific Ocean. On the other hand, it is also important to strengthen the monitoring of their habitat changes driven by climatic and environmental conditions in order to ensure a more reasonable and comprehensive regulation of the fishing quota of the two species (Ianelli and Hintzen, 2020). Therefore, the habitat patterns of D. gigas and T. murphyi were examined and the synergistic response of their habitat suitability to El Niño events with different intensities was further explored. The primary objectives of this study are (1) to assess the impacts of different-intensity El Niño events on environmental conditions; (2) to examine the synchronous response of the suitable habitat patterns of D. gigas and T. murphyi under El Niño events with different intensities; and (3) to evaluate the relationship between environment and species habitat. This study can provide scientific supports for SPRFMO to manage the two species under a changing climate.
Materials and Methods
Fisheries, Environmental, and Climate Index Data
The autumnal fisheries data of D. gigas from 2011 to 2017 and T. murphyi from 2013 to 2017 with a spatiotemporal resolution of season and 0.5° × 0.5° were supplied by the National Data Center for Distance-Water Fisheries of China (NDCDF), Shanghai Ocean University, China. The fisheries data information includes fishing location, fishing time, catch (unit: t), and fishing effort. The study region in the southeast Pacific Ocean covers both central fishing grounds of D. gigas and T. murphyi and is bounded by 70°–97°W and 20°–47°S (Figure 1).
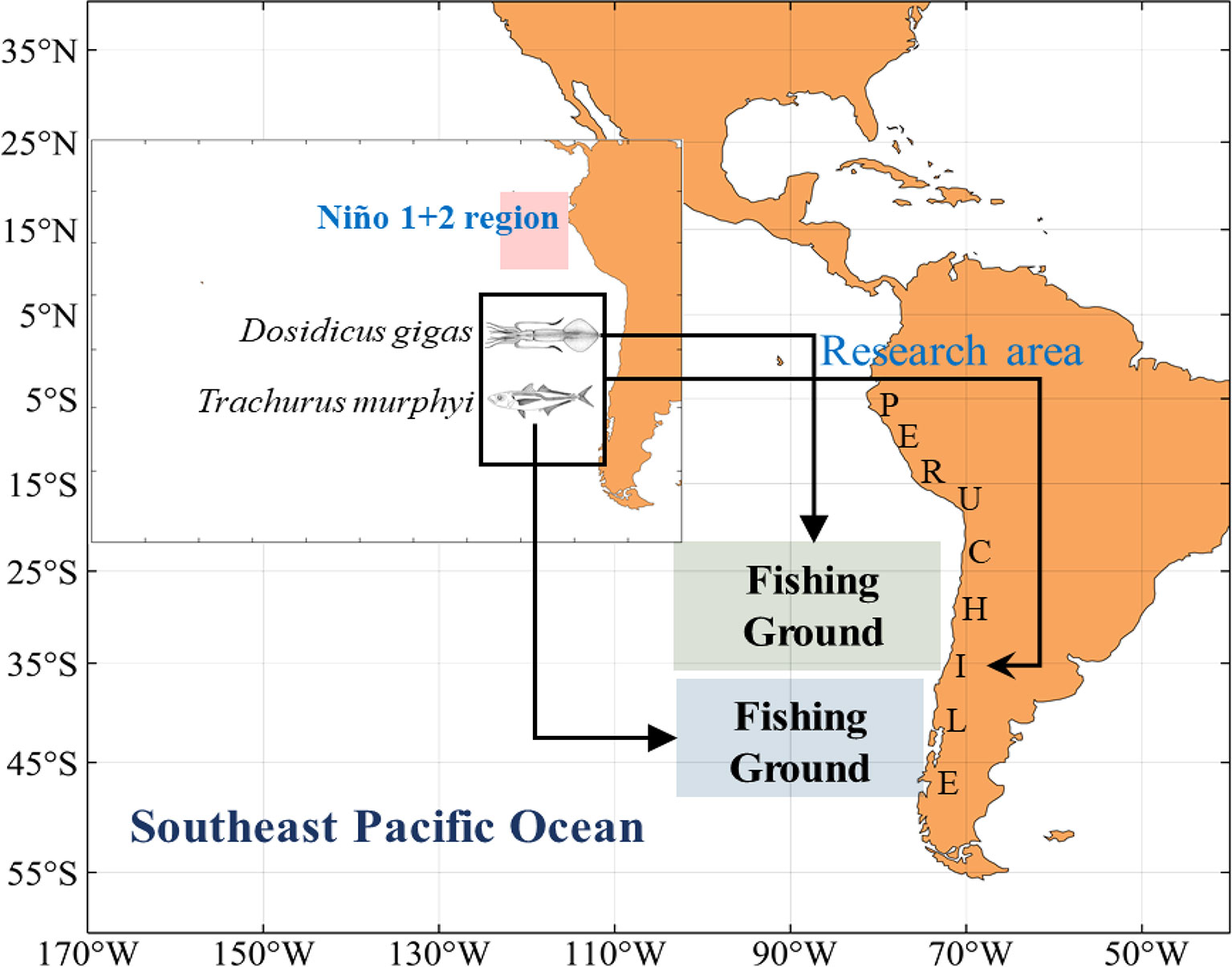
Figure 1 Geographical distribution of fishing ground of Dosidicus gigas and Trachurus murphyi off Chile and the Niño 1 + 2 region in the Eastern Pacific Ocean.
According to previous studies in assessing the impacts of environmental factors affecting the habitat distribution of D. gigas and T. murphyi, sea surface height anomaly (SSHA), sea surface salinity (SSS), and water temperature at 400-m depth (Temp_400 m) were selected as key variables for D. gigas, while sea surface temperature (SST), mixed layer depth (MLD), and Temp_400 m were selected for T. murphyi in this study (Feng et al., 2021a; Feng et al., 2021b). All environmental variables covering the study region from 1950 to 2017 were obtained from the Asia Pacific data research center (http://apdrc.soest.hawaii.edu/las_ofes/v6/dataset?catitem=71); the data period and its spatiotemporal resolution were matched with fisheries data.
In order to explore the synchronous response of the suitable habitat patterns of D. gigas and T. murphyi off Chile in the southeast Pacific Ocean under the long-term series of El Niño events with different intensities, the monthly SSTA of the Niño 1 + 2 region (between 80°–90°W and 0°–10°S, close to the research area) was considered as an indicator of climate variability, which is sourced from the following website: http://iridl.ldeo.columbia.edu/SOURCES/.Indices/.
Classification of El Niño Events With Different Intensities From 1950 to 2017
According to the definition of different ENSO events by the National Oceanic and Atmospheric Administration (NOAA), El Niño events are divided into four intensities based on the Oceanic Niño index (ONI): weak El Niño events (WE) with 0.5 ≤ ONI ≤ 0.9, moderate El Niño events (ME) with 1.0 ≤ ONI ≤ 1.4, strong El Niño events (SE) with 1.5 ≤ ONI ≤ 1.9, and very strong El Niño events (VSE) with ONI ≥2.0 (http://ggweather.com/enso/oni.htm). According to the above delimitation method, the classification results of El Niño events with different intensities from 1950 to 2017 were as follows: years with WE events included 1952, 1953, 1958, 1969, 1976, 1977, 1979, 2004, 2006, and 2014; years with ME events included 1951, 1963, 1968, 1986, 1994, 2002, and 2009; and years with SE and VSE events included 1957, 1965, 1972, 1982, 1987, 1991, 1997, and 2015.
Construction of HSI Models for D. gigas and T. murphyi off Chile
For marine species vulnerable to environmental change, the response of the spatial distribution pattern of their habitat to climate stress is significant. Based on the long-term impacts of climate variability on marine species in a large spatial domain, the response of species habitat pattern in the same-area or cross-area is often concurrent. At the spatial level, the synchronous impacts sourced from climate variability on species may be positive or negative. At the time level, the changes in species habitat may occur in the same period, in spite of the wide geographical locations across species. In this study, we assumed that the different-intensity El Niño events yielded significant impacts on the habitat patterns of the two species through changing the environments on the fishing ground. Thus, we developed the habitat model for them inclusive with the key environmental factors.
The optimal weighting-based habitat suitability index (HSI) models were developed to evaluate the synchronous response of the spatial distribution patterns of D. gigas and T. murphyi habitat off Chile under different-intensity El Niño events, respectively. The first step in the construction of the HSI model was to fit the suitability index (SI) model for each environmental variable (SSHA, SSS, and Temp_400 m for D. gigas, and SST, MLD, and Temp_400 m for T. murphyi). The second step was to combine all SI models into an integrated HSI model for D. gigas and T. murphyi based on the empirical arithmetic weighting method (AMM) and then calculate the HSI from 1950 to 2017 for the fishing grounds of D. gigas and T. murphyi, respectively.
The range of SI for each environmental variable and the HSI was in the range of 0–1, where 0 indicated the poor habitat with the most unfavorable environmental condition and the least fishing effort, while 1 indicated the suitable habitat with the most favorable habitat and the highest fishing effort (Tian et al., 2009; Feng et al., 2020). According to the values of SI and HSI, SI ≥0.6, HSI ≤0.2, HSI ≥0.6, and HSI ≥0.8 were regarded as the suitable environmental ranges, poor habitat, suitable habitat, and optimal habitat for D. gigas and T. murphyi, respectively (Fang et al., 2014; Li et al., 2014; Feng et al., 2021b). In this study, the SI models and HSI models were sourced from Feng et al. (2021b) whose studies showed that each SI model of D. gigas and T. murphyi passed the significant test (P < 0.05) with a low root mean squared error (RMSE) and a high correlation coefficient (R2), and the HSI model of these two species also showed good model prediction performance.
Exploring the Relationship Between Climate Change and Environment
To analyze the connection between the climate variability in the southeast Pacific Ocean with the environmental variables of D. gigas and T. murphyi, the relationship between SSTA in the Niño 1 + 2 regions and all environmental variables was evaluated using cross-correlation functions (CCFs). Furthermore, in order to evaluate the variation of the marine environment under El Niño events with different intensities, the anomalies of SSH, SSS, and Temp_400 m for D. gigas and SST, MLD, and Temp_400 m for T. murphyi were calculated, respectively.
Based on the established SI model of each environmental variable, the average of the SI value and the proportion of the suitable environmental range (SI ≥0.6) of each environmental factor were determined to evaluate the variation difference in the suitable environment of D. gigas and T. murphyi under El Niño events with different intensities, respectively.
Exploring the Relationship Between Climate Variability and the Two Species
To explore the synchronous variation of suitable habitat patterns, the established HSI model of D. gigas and T. murphyi was used to calculate the HSI anomaly and the proportion of poor, suitable, and optimal habitat areas of the two species. Spatial distributions of suitable habitat were also plotted under El Niño events with different intensities. Moreover, the longitudinal and latitudinal gravity centers of suitable habitat of D. gigas and T. murphyi under El Niño events with different intensities were examined to reflect the changing characteristics of the spatiotemporal position of the suitable habitat of these two species. The gravity centers of suitable habitat were determined as follows (Li et al., 2014):
where LONGHSI and LATGHSI represent the longitudinal and latitudinal gravity centers of suitable habitat, respectively; i and j denote longitude and latitude, respectively.
Exploring the Relationship Between Environmental Change and the Two Species
A spatial correlation analysis method was applied to assess the spatial correlation between key environmental factors and habitat suitability for D. gigas and T. murphyi, respectively, and spatial correlations were further plotted. In addition, the CCF was used to analyze the relationship between the environmental variables and the latitudinal gravity centers of suitable habitat of D. gigas and T. murphyi, and then the most preferred environmental factors of the two species were selected. On this basis, the variation difference of the average latitudinal gravity center of the most preferred environmental factor was analyzed under El Niño events with different intensities. Three El Niño years with different intensities were selected as the case study, and the isolines of the most preferred environmental factor of D. gigas and T. murphyi were plotted in order to explore the correlation between key environmental factors and habitat suitability of the two species.
Results
Variations in Environmental Conditions Under Different-Intensity El Niño Events
Cross-correlation analysis showed that the Niño 1 + 2 index was significantly positive with SSHA and Temp_400 m at a time lag of -8–14 months (P < 0.001), and the highest correlation occurred at 1 month with correlation coefficient values of 0.2686 and 0.2643, respectively (Figure 2). The Niño 1 + 2 index was significantly positive with SST and SSS at time lags of -1–1 and -2–1 months, respectively (P < 0.05). The highest correlation occurred at -1 and 0 months with correlation coefficient values of 0.1443 and 0.1090, respectively (Figure 2). The Niño 1 + 2 index was negatively correlated with MLD but not significantly (P > 0.05) (Figure 2). From the perspective of the variable-driven processes, the above results of CCF indicated that the increase in Niño 1 + 2 SSTA would yield an increase in SSHA, SSS, and Temp_400 m and a decrease in MLD. At the same time, the short leading or lagging time of 0 or 1 month implied that the response of the marine environmental variables affecting D. gigas and T. murphyi to El Niño events was rapid and even concurrent with climate variability. Under El Niño events with different intensities, SSHA increased with the strengthening of the intensity, while SSSA and MLDA gradually decreased. SSTA and Temp_400A were higher during WE and SE El Niño events compared to ME El Niño events (Figure 2).
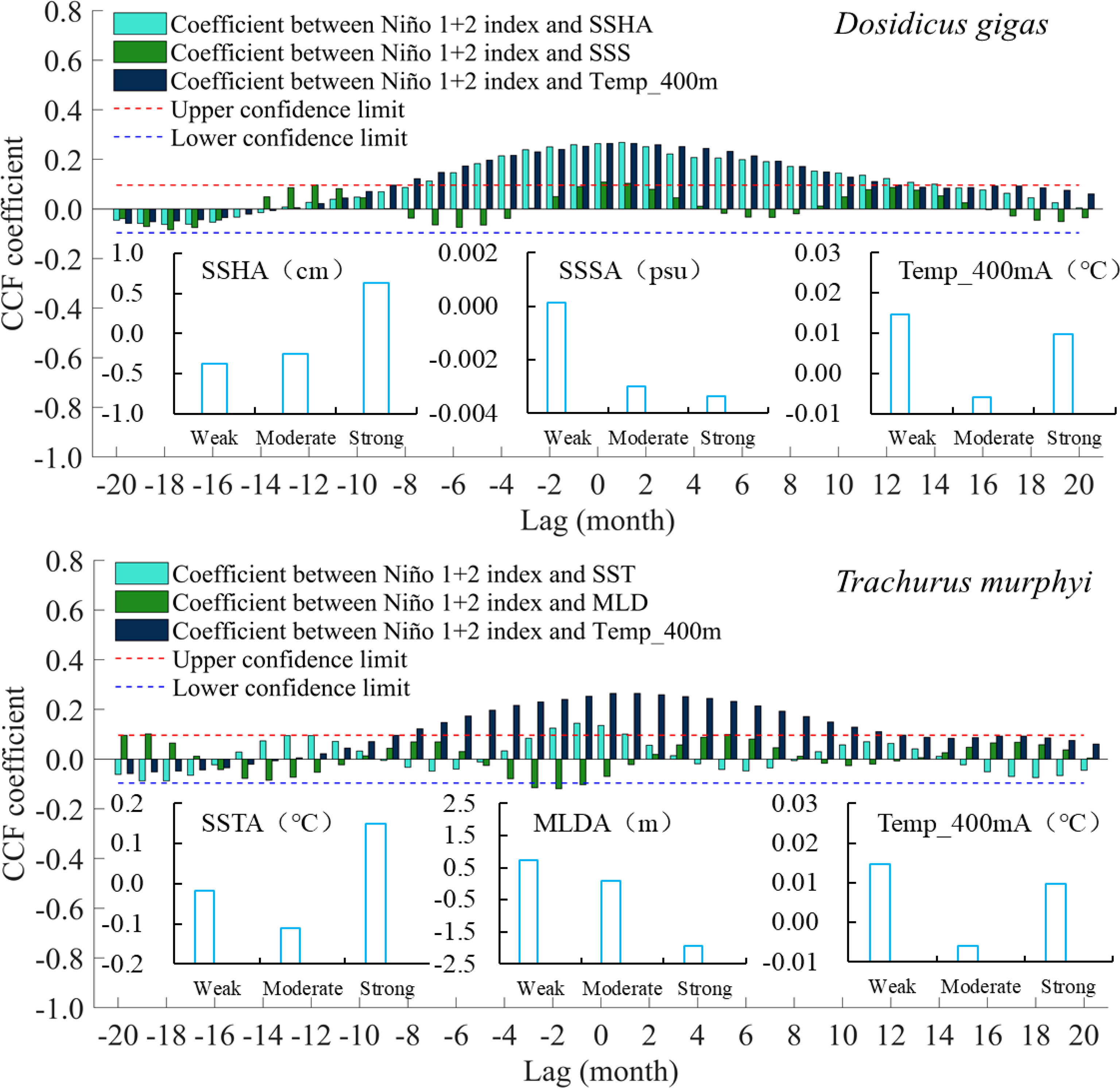
Figure 2 Cross-correlation coefficient between the Niño 1 + 2 index and environmental factors and variation of anomaly value of environmental factors under El Niño events with different intensities.
The suitability indexes of SI-SSHA, SI-SSS, and SI-Temp_400 m for D. gigas gradually decreased with the strengthening of the intensity (Figure 3). However, the suitability indexes of SI-SST, SI-MLD, and SI-Temp_400 m for T. murphyi were higher in WE and SE El Niño years than those in ME El Niño years (Figure 3). The proportion of suitable environmental ranges (SI ≥0.6) of each environmental variable for D. gigas and T. murphyi varied significantly (Figure 4). Among them, the proportion of suitable SI-SSHA and SI-Temp_400 m for D. gigas gradually decreased with the El Niño event intensities, while SI-SSS for D. gigas was highest under ME El Niño events compared to that under WE and SE El Niño events. The proportion of suitable SI-SST for T. murphyi was gradually decreased, while the suitable SI-Temp_400 m for T. murphyi showed the opposite variation. The proportion of suitable SI-MLD for T. murphyi varied contrarily to that of SI-SSS for D. gigas.
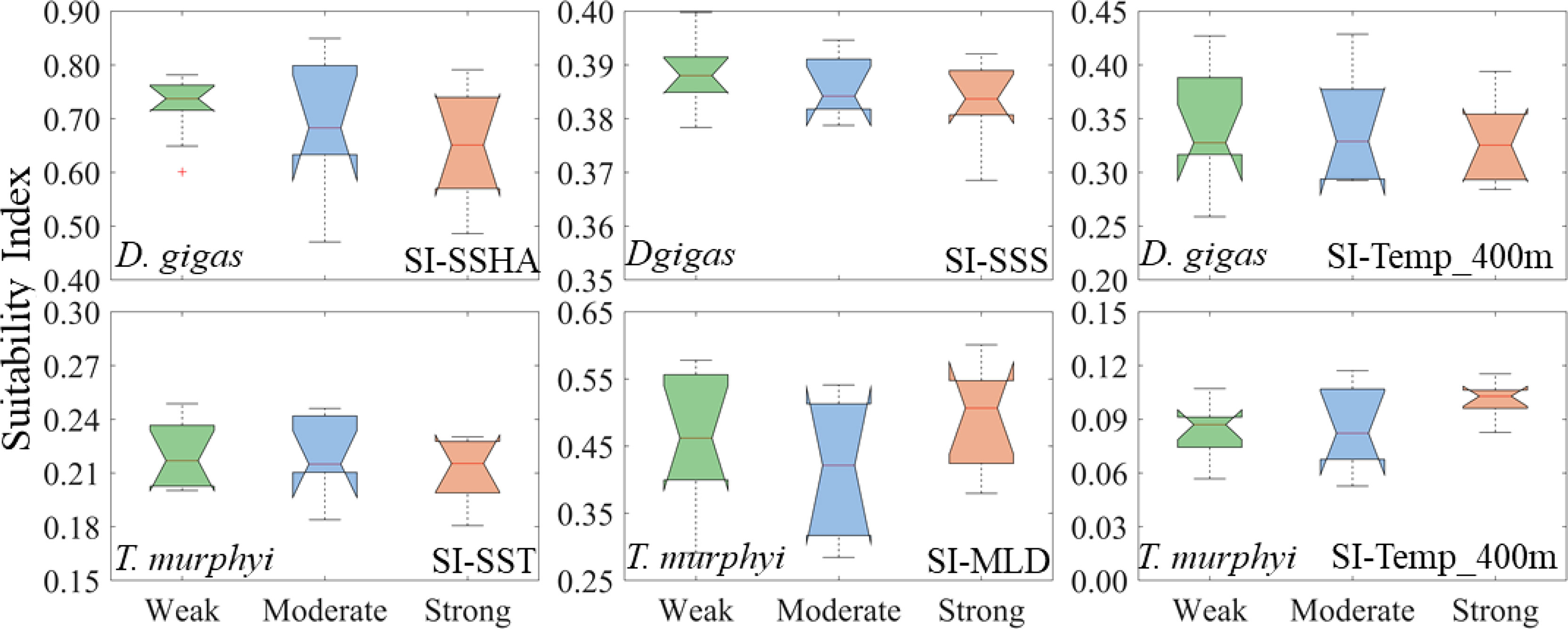
Figure 3 Changes of suitability index of key environmental factors of Dosidicus gigas and Trachurus murphyi under El Niño events with different intensities. (The nodes of the boxplot from top to bottom are the upper edge, upper quartile, median, lower quartile, and lower edge, with “+” representing outliers).
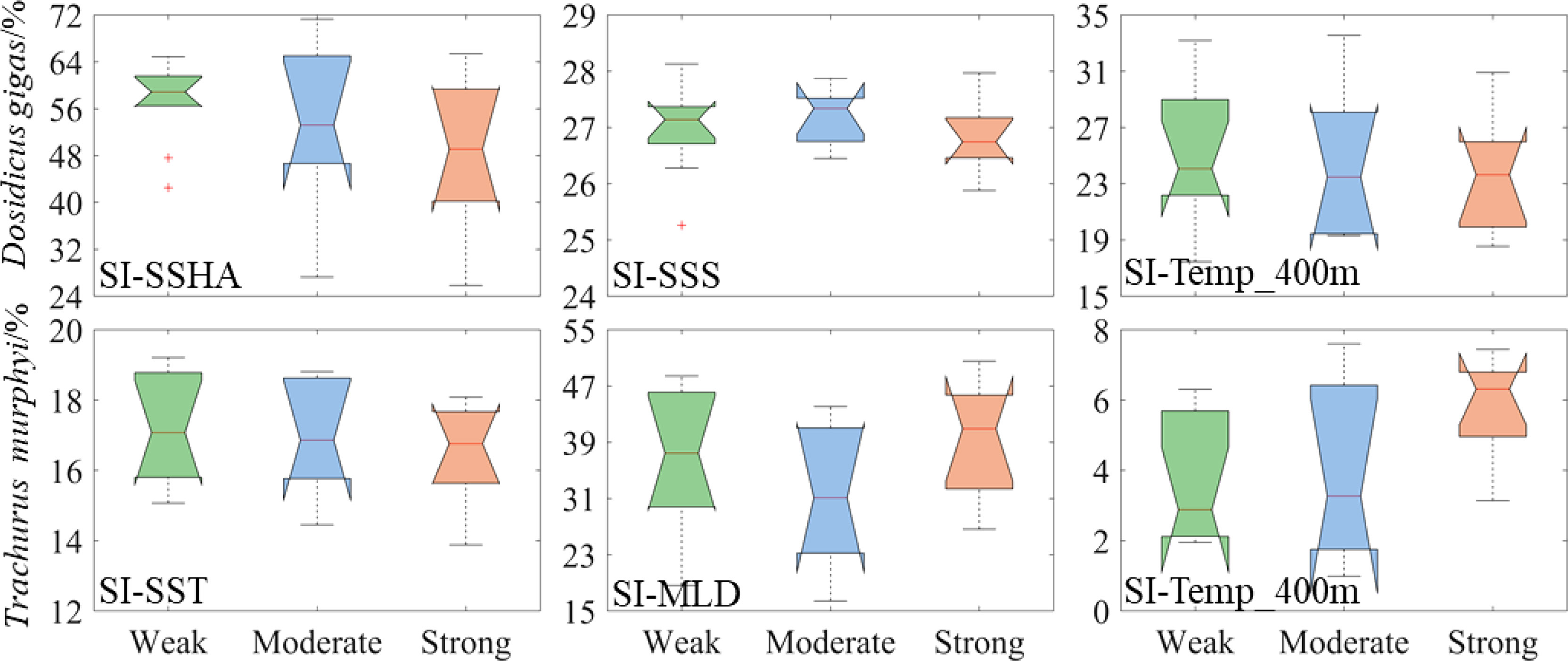
Figure 4 Changes in the proportion of suitability index (SI >0.6) of key environmental factors of Dosidicus gigas and Trachurus murphyi under El Niño events with different intensity. (The nodes of the boxplot from top to bottom are the upper edge, upper quartile, median, lower quartile, and lower edge, with “+” representing outliers).
Different-Intensity El Niño-Related Habitat Changes of D. gigas and T. murphyi
The HSI anomaly of D. gigas gradually decreased with the El Niño intensities; at the same time, the proportion of its poor habitat area gradually increased, while the proportion of suitable and optimal habitat ranges during SE events was lowest (Figure 5). For T. murphyi, the HSI anomaly and suitable and optimal habitats were higher under WE and SE El Niño events than those under ME El Niño events. Compared to the WE and SE El Niño events, the ME El Niño events yielded expanded poor habitats (Figure 5).
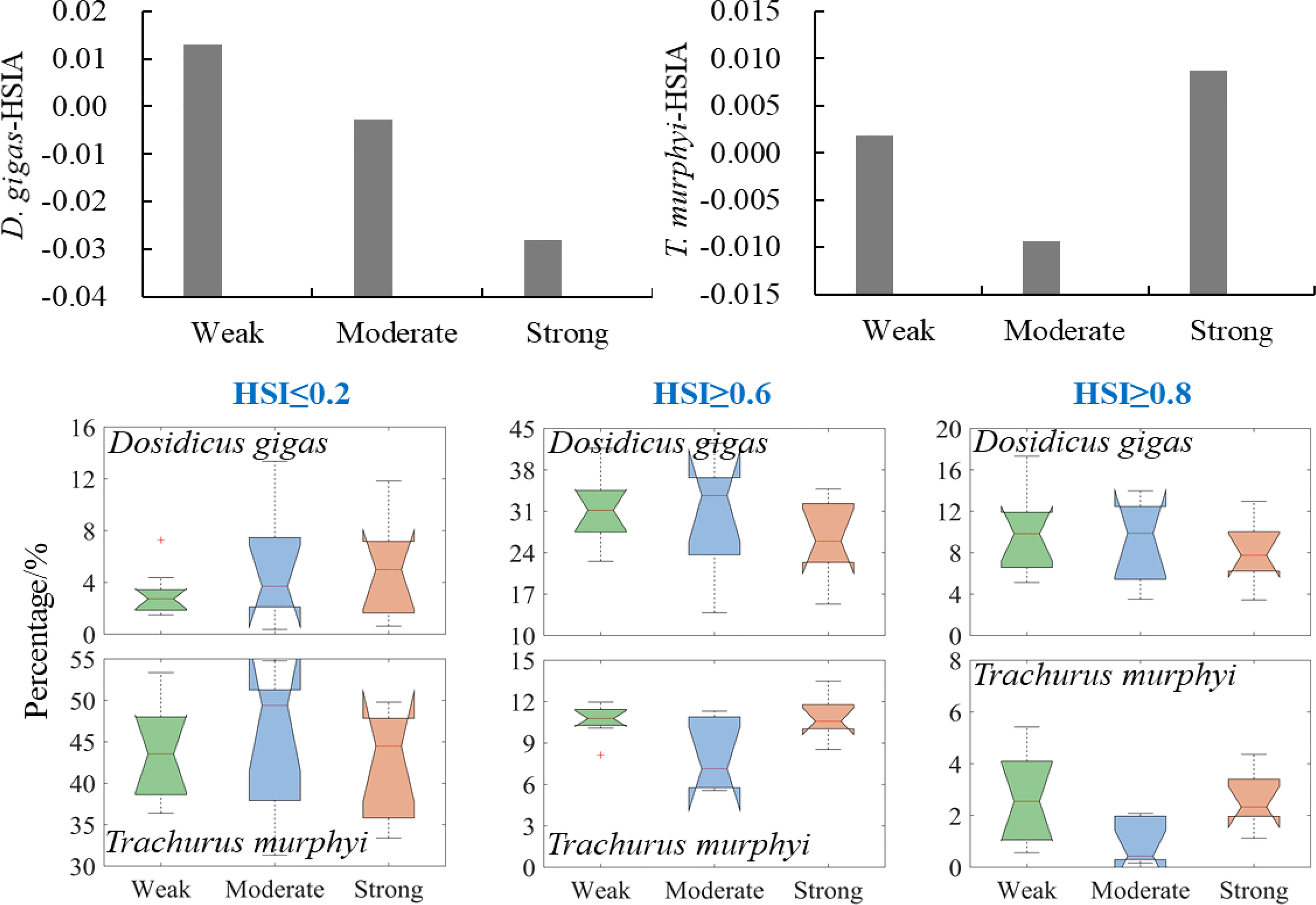
Figure 5 Changes in proportion of poor, suitable, and optimal habitat areas of Dosidicus gigas and Trachurus murphyi under El Niño events with different intensities. (The nodes of the boxplot from top to bottom are the upper edge, upper quartile, median, lower quartile, and lower edge, with “+” representing outliers).
Regarding spatial distribution, the suitable habitat of D. gigas was distributed in the areas between 75°–97°W and 22°–38°S in the central and northern regions of Chile, which significantly reduced with the strengthening of El Niño intensities. Under the SE El Niño events, the suitable habitats of D. gigas largely reduced in the west of 85°W between 30° and 35°S (Figure 6). A minor difference was found in the suitable habitat range and spatial distribution during the WE and SE El Niño years for T. murphyi. Suitable habitats mainly concentrated in the waters between 42° and 47°S, which significantly decreased in the southwest region of Chile under ME El Niño events (Figure 6).

Figure 6 Spatial distribution of suitable habitats of Dosidicus gigas and Trachurus murphyi under El Niño events with different intensities.
The spatial location of D. gigas suitable habitat gradually moved southwestward with the El Niño event intensity (Figure 7). For T. murphyi, the longitudinal and latitudinal gravity centers of suitable habitat moved southwestward under WE and SE El Niño events compared with ME El Niño events (Figure 7). In the longitudinal direction, the gravity center of the suitable habitat of D. gigas during SE events was significantly different from that under WE and SE events, with the longitudinal difference of about 1° (Figure 7). The gravity center of the suitable habitat of T. murphyi during WE and SE years was clearly different from that in ME years, with the longitudinal difference of about 1.5° (Figure 7). The changing trends of the gravity center of the suitable habitats of D. gigas and T. murphyi in the latitudinal direction were both similar to those of the longitudinal direction, with a small variation difference of the gravity center in the latitude under El Niño events with different intensities (Figure 7).
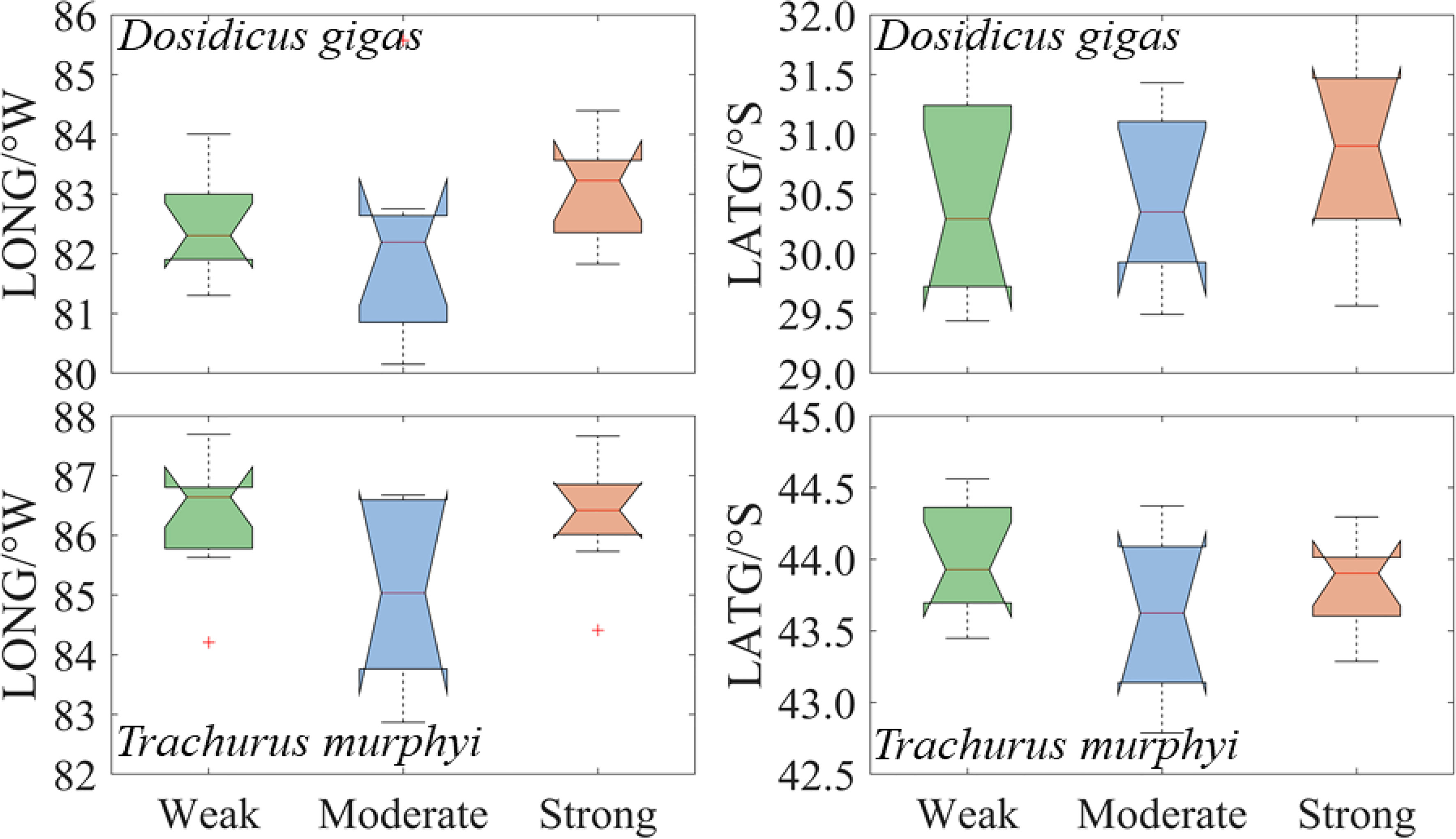
Figure 7 Changes in the longitudinal and latitudinal gravity center of the suitable habitat of Dosidicus gigas and Trachurus murphyi under El Niño events with different intensities. (The nodes of the boxplot from top to bottom are the upper edge, upper quartile, median, lower quartile, and lower edge).
Impact of Environmental Changes on D. gigas and T. murphyi
Spatial correlation results showed that SSHA, SSS, and Temp_400 m were negatively correlated with HSI on the fishing ground of D. gigas, SST and Temp_400 m were positively correlated, and MLD was negatively correlated with HSI on the fishing ground of T. murphyi (Figure 8).
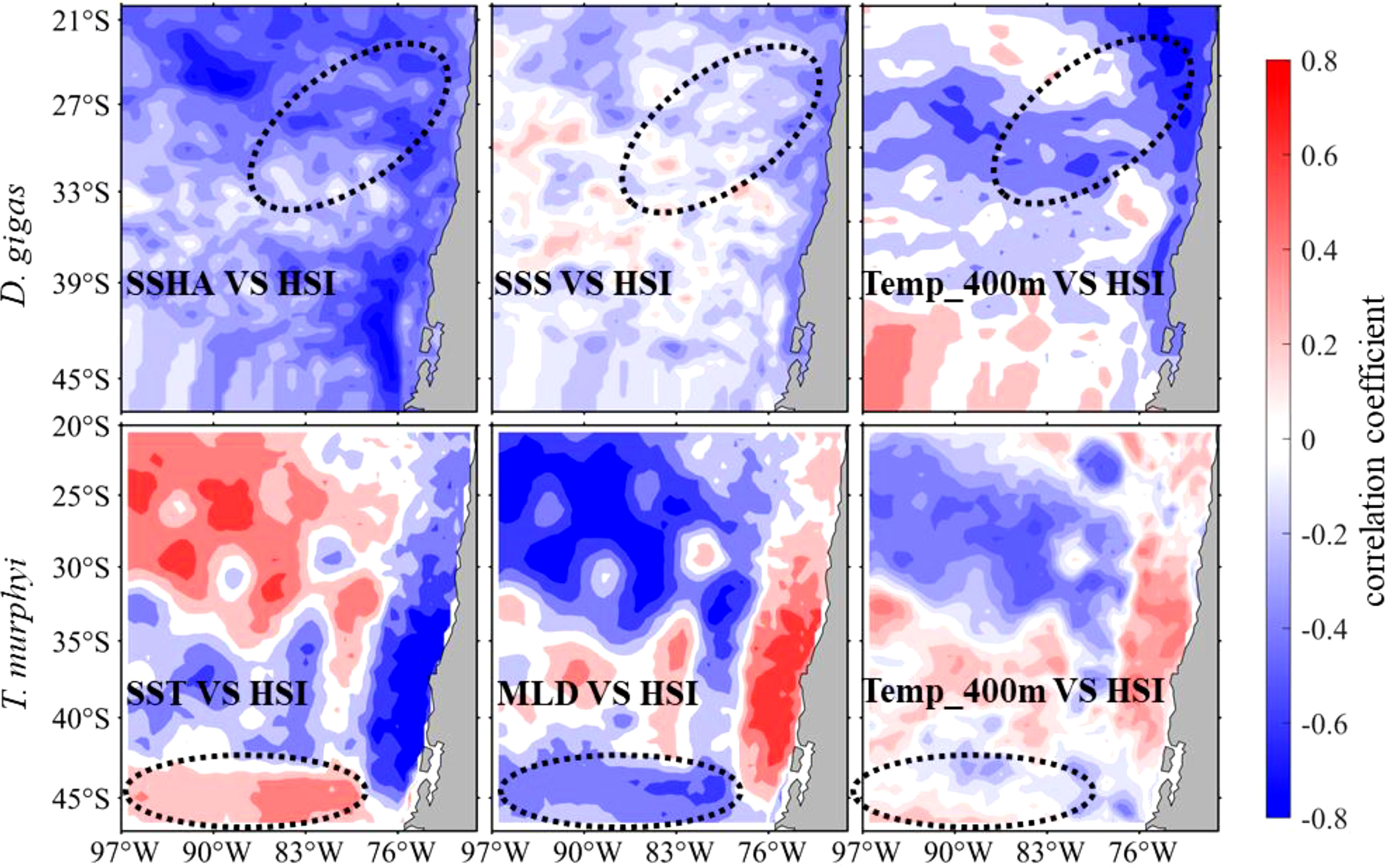
Figure 8 Spatial correlation between key environmental factors of Dosidicus gigas (top panel) and Trachurus murphyi (bottom panel) overlaid with a fishing ground range.
Cross-correlation analysis showed that the latitudinal gravity center of the suitable habitat of D. gigas was significantly positive with SSHA (P < 0.001), SSS (P < 0.05), and Temp_400 m (P < 0.001), respectively (Figure 9). The environmental variable with the highest correlation coefficient value of 0.5219 was Temp_400 m (Figure 9). The latitudinal gravity center of the suitable habitat of T. murphyi was significantly positive with SST (P < 0.001) and MLD (P < 0.05) and was positively related to Temp_400 m (P > 0.05) but not significantly (Figure 9). The environmental variable with the highest correlation coefficient value of 0.5094 was SST (Figure 9). The above results of CCF also showed that the changes in the key environmental factors of Dosidicus gigas and Trachurus murphyi drove the changes in the latitudinal gravity center of the suitable habitats of the two species, and the influence of seawater temperature on the latitudinal changes of their habitats was more important.
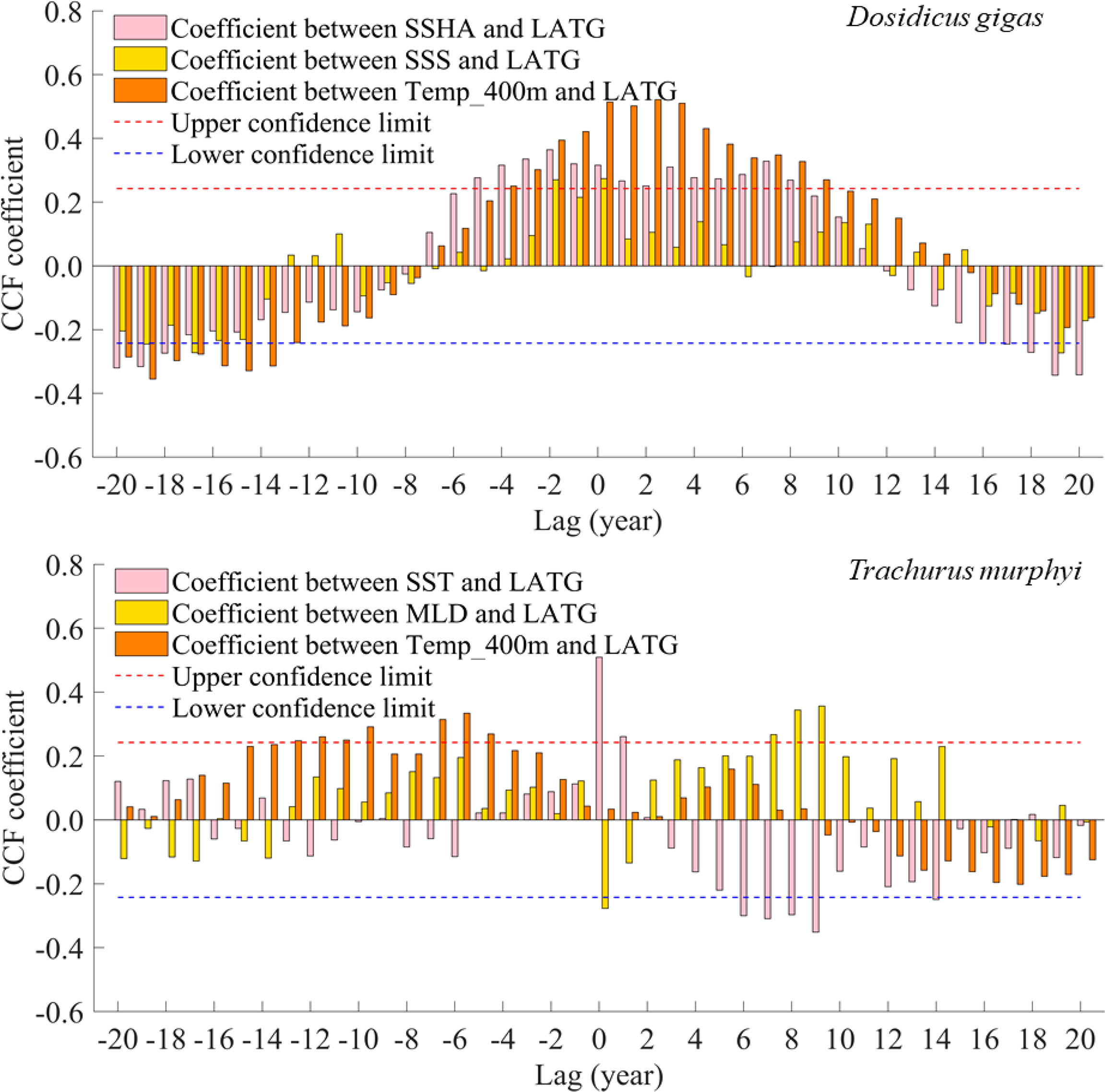
Figure 9 Cross-correlation coefficients of key environmental factors of Dosidicus gigas (top panel) and Trachurus murphyi (bottom panel) and latitudinal gravity center of habitat suitability index.
The contour lines of Temp_400 m = 7.25°C for D. gigas and SST = 12°C for T. murphyi under El Niño events with different intensities showed that the average latitude of the most preferred Temp_400 m of D. gigas gradually moved southward under SE El Niño events, while the average latitude of the most preferred SST of T. murphyi moved southward under WE and SE El Niño events than that under ME El Niño events (Figure 10). Furthermore, 3 years was selected as the case to show the movement of the most preferred Temp_400 m for D. gigas and SST for T. murphyi (Figure 11); the result was consistent with the findings shown in Figure 10.
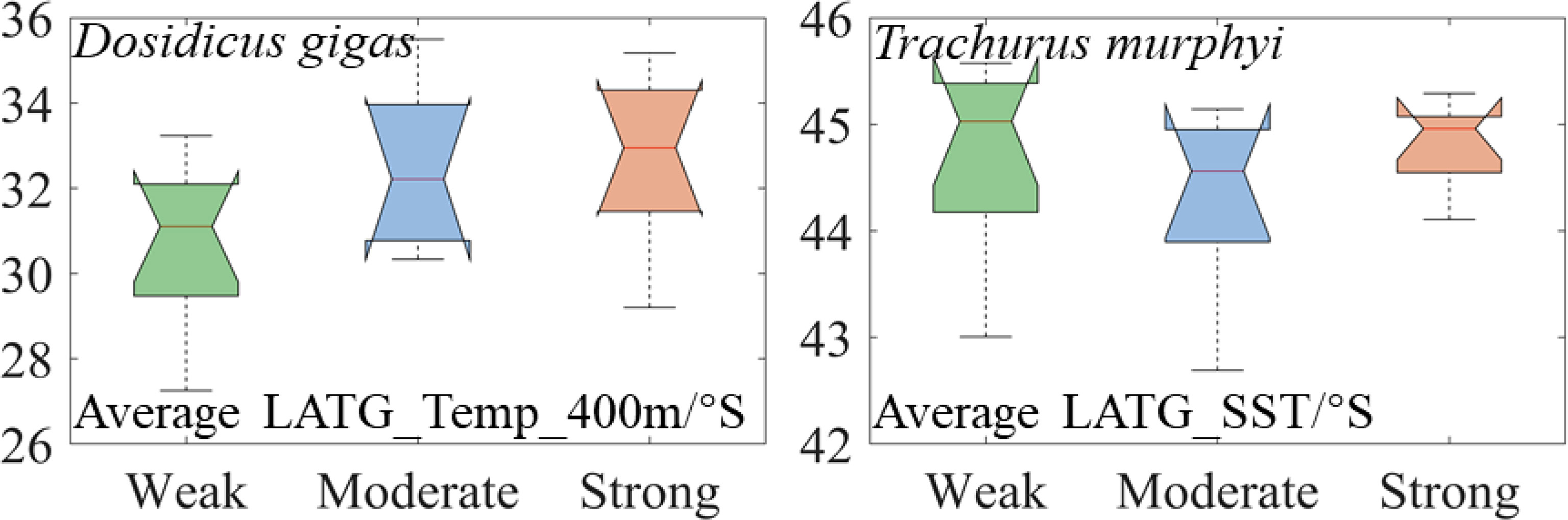
Figure 10 Average latitude changes of the most preferred Temp_400 m of Dosidicus gigas and SST of Trachurus murphyi under El Niño events with different intensities. (The nodes of the boxplot from top to bottom are the upper edge, upper quartile, median, lower quartile, and lower edge, with “+” representing outliers).
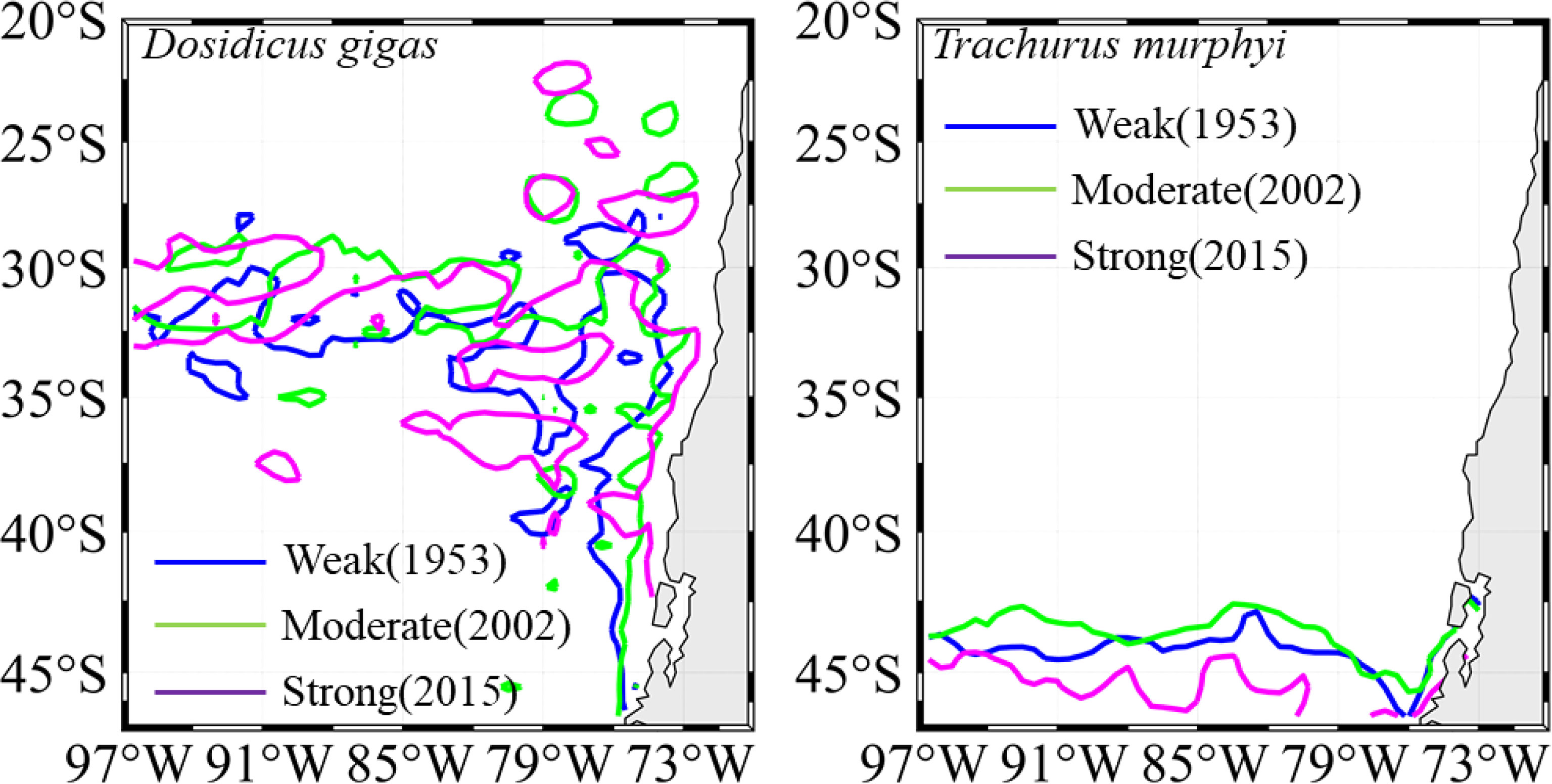
Figure 11 Spatial distribution of the isolines of the most preferred Temp_400 m for Dosidicus gigas and the most preferred SST for Trachurus murphyi in the years of 1953 (weak El Niño), 2002 (moderate El Niño), and 2015 (strong El Niño).
Discussion
Climate variability plays important roles in the variation of environmental conditions such as ocean temperature, chemical composition, and primary productivity and then drives the changes in marine species abundance and distribution. In this study, in order to better understand the synchronous response of the spatial distribution patterns of D. gigas and T. murphyi habitats off Chile in the Southeast Pacific Ocean to climate variability, the environmental variables included in the HSI model mainly referred to Feng et al. (2021b) on the selection of key environmental factors of D. gigas and T. murphyi off Chile based on the maximum entropy model (MaxEnt). Then, the factors with a high correlation among the environmental factors were excluded. Meanwhile, according to the biological characteristics of the vertical movement and multifaceted ecological demand for D. gigas and T. murphyi off Chile, SSHA, SSS, Temp_400 m, and SST, MLD, and Temp_400 m were selected as the key environmental factors for D. gigas and T. murphyi, respectively, in order to explore the synergistic response of their suitable habitats to different-intensity El Niño events.
The El Niño event is the warm regime of the ENSO phenomenon with the strongest interannual variation signal in the sea–air coupling system of large-scale surface seawater and tropical atmospheric circulation anomalies in the equatorial Pacific Ocean every 3–7 years, resulting from the strong air–sea interaction (Montecinos and Aceituno, 2005; Behera et al., 2021). The changes in equatorial currents and trade winds caused by the El Niño events have a strong impact on regional oceanic conditions, and the changing environments further significantly affect the life cycle of marine species, such as gonad maturation time, migration path, spawning location, and habitat distribution range (Chen and Cane, 2008; Pecl and Jackson, 2008; Brander, 2010). However, previous studies stated that the El Niño events showed obvious irregularity in the tropical Pacific, specifically reflecting the irregularity of the duration and intensity of SST anomalies, which has evolved different-intensity El Niño events and yielded various impacts on the abundance and distribution of marine species (Liu and Xue, 2010; Zhou and Fang, 2018). In addition, the redistribution of the species population depends not only on the climate variability but also on the synchronous response of species related to them (Davis et al., 1998; Hastings et al., 2020). Therefore, exploring the coordinated response of habitat distribution of key pelagic species such as D. gigas and T. murphyi off Chile to climatic and environmental changes driven by the El Niño events with different intensities at various spatial and temporal scales can provide a scientific basis for the rational utilization and management of their resources.
Our results presented that the suitable habitats of D. gigas gradually decreased with the strengthening of the El Niño event intensity, and its key environmental factor SSHA increased with the strengthening of the intensity, while variations of SSS were opposite to those of SSHA. It indicated that a lower SSHA and a more suitable SSS were favorable for yielding a high-quality habitat of D. gigas (Figures 2 and 5). SSH represents the convergence and surge of seawater and also reflects the cumulative results of the thermodynamics and dynamics between ocean and atmosphere. Its impact on marine species often reflects the impact of different ENSO events (Li et al., 2016; Chen and Chen, 2020). During the ME and SE El Niño events, the weakened trade wind in the equatorial Pacific broke the balance between the surface wind stress and the upper boundary pressure gradient of the thermocline. Consequently, the sea surface wind stress anomaly in the tropical western Pacific expanded from west to east along the equator with the oscillation of the water body in the tropical Pacific (Blanco et al., 2002; Strub and James, 2002). Moreover, the generated abnormal wind stress and divergence fields were conducive to water body oscillation, which in turn further strengthened the eastward expansion of water body oscillation in the mixed layer, resulting in the decline of the sea level in the tropical western Pacific due to water body eastward movement, while the sea level in the equatorial eastern Pacific rose and thermocline deepened due to water body accumulation (Zhao et al., 2007; Neelin, 2010; Alizadeh-Choobari, 2017). The increasing of SSH in the eastern equatorial Pacific and South America also indirectly reflected that the accumulation of warm water weakened the upwelling along the west coast of South America and the upwelling intensity of inorganic salts and other nutrients in the deep waters during the El Niño years, leading to the reduction in plankton richness and further weakening the food availability of D. gigas (Halpern, 2002; Rahn, 2012). Inevitably, suitable habitats of D. gigas largely contracted.
As an indicator of the global water cycle, SSS plays an important role in the dynamic process of tropical climate; it often reflects the changes in ocean fresh water flux, ocean advection, and MLD caused by ENSO atmospheric circulation anomaly (Huang et al., 2008; Cravatte et al., 2009; Nguyen et al., 2017; Fang et al., 2019; Qi et al., 2019). Surface advection and precipitation change are the main mechanisms of SSS response to the El Niño events (Huang et al., 2008; Fang et al., 2019). During the El Niño years, the warm fresh water in the equatorial western Pacific moved eastward, resulting in the reduction of precipitation in the South Pacific Composite Belt (SPCZ) region and indirectly leading to the SSS decline (Cravatte et al., 2009; Zhi et al., 2020). The decreased SSS also reflected the weakening of the intersection intensity between the cold eddy in the north and the warm eddy in the south equator, yielding unfavorable impacts on the development of fishery productivity and the expansion of suitable habitat for D. gigas (Niu et al., 2009).
The suitable habitat range of T. murphyi during WE and SE El Niño events enlarged significantly compared to ME El Niño events, and key environmental factors SST, Temp_400 m, and MLD played an important role in regulating the spatial distribution of the T. murphyi habitat (Figures 2 and 5). Zheng et al. (2012) clarified that under the central and Western El Niño events, the negative salinity anomaly and the MLD shifted westward, showing consistency in the variation. Therefore, MLD decreased with the strengthening of the El Niño intensity and the decrease of SSS. MLD can reflect the change of sea–air flux, and its thickness determines the vertical distribution volume of ocean net surface heat flux (Chang et al., 2013; Xu, 2014). During the El Niño years, with the combined influences of ocean surface heat flux and the large-scale wind field, the warming of the ocean surface in the eastern equatorial Pacific was much stronger than that in the subsurface, inducing a shallow upper ocean stratification and a mixed layer depth. This further accumulated the solar radiation absorbed by the ocean surface and heated up and shallowed the mixed layer under the strengthening El Niño intensity (Sutton et al., 2007; Xu, 2014). In addition, the North–South movement of the SST isoline is consistent with the shift of the T. murphyi suitable habitat. It showed that with the rise of SST in Chilean waters, the preferred SST isoline of T. murphyi moved southward, and suitable habitat also migrated southward. On the contrary, they both shifted northward (Li et al., 2016; Feng et al., 2021b). However, the key environmental factors SST and Temp_400 m of T. murphyi were higher during WE and SE El Niño events than those under ME events, which may be related to the occurrence of multiple strong sporadic westerly events and different types of El Niño events in the central and eastern Pacific (Figure 2).
There are two types of El Niño events: eastern El Niño and central El Niño (Philander, 1983). The impacts of central and eastern El Niño events on the West Coast of America may be opposite. For example, with the occurrence of the central El Niño event, the western United States was mainly dry in the north and wet in the south in winter, while the eastern El Niño event produced a humid climate (Weng et al., 2007; Yu and Zou, 2013). According to the meteorological industry standard “El Niño/La Niña events discrimination method” issued by the China Meteorological Administration in 2017, the moderate-intensity El Niño years 1968, 1994, 2002, and 2009 in this study belong to central El Niño events, while weak and strong El Nino event years belong to eastern El Niño events. Compared with the SST anomaly under the eastern El Niño events, the maximum warming phenomenon under the central El Niño events is concentrated in the central equatorial Pacific, and the warming effect on the eastern equatorial Pacific and South America is relatively short, limited, and weak (McPhaden, 2004; Alizadeh-Choobari, 2017; Zhou and Fang, 2018). In addition, during the eastern El Niño years, the easterly stress anomaly along the equator of the eastern Pacific and the central Pacific usually remains in the east of 100°–120°W (McPhaden, 2004). However, it increases in the zonal direction during the moderate-intensity El Niño years, which may weaken the deepening trend of the thermocline in the eastern equatorial Pacific to a certain extent, resulting in the weakening variation of the SST anomaly in eastern Pacific and South America (McPhaden, 2004). Therefore, the SST and Temp_400 m of T. murphyi and its suitable habitat range were higher during WE and SE El Niño events than those in ME El Niño years. Compared with those of T. murphyi, D. gigas abundance and recruitment are more vulnerable to climatic and environmental change, and its distribution area is closer to the equatorial region, so it is more significantly affected by the El Niño events (Yu et al., 2019; Fang et al., 2021).
Climate-induced variations in ocean current intensity, circulation axis, and inter-tropical convergence zone (ITCZ) affected the sea substrate conditions and forced marine species with strong migration ability to abandon their original habitat and move to a new suitable and stable living space (Tian et al., 2004; Lee, 2009). In this study, the latitudinal gravity center of the suitable habitat for D. gigas off Chile gradually moved southward with the strengthening of El Niño intensities, which was mainly associated with seawater temperature at a depth of 400 m (Figures 7, 9–11). For T. murphyi, the latitudinal gravity center of the suitable habitat shifted southward under WE and SE El Niño events compared with ME El Niño events, which could be explained by the movement of the preferred SST (Figures 7, 9–11). A stable ocean current system is the fundamental reason for the formation of the marine species fishing ground. The fishing ground of D. gigas distributed in the northern region of Chile is mainly formed by the coastal upwelling driven by wind (Blanco et al., 2002). During normal climate years, the equatorial wind promotes the coastal upwelling on the west coast of South America, uplifts the thermocline, strengthens the upwelling intensity, promotes phytoplankton growth, and provides rich primary productivity for the growth and reproduction of pelagic species (Blanco et al., 2002). However, during the El Niño years, the south Pacific highly weakened and ITCZ moved southward, which was unfavorable to upwelling in southern Peru and northern Chile. The cold eddy force in the north of southern Chile was weaker than the South Equatorial Countercurrent in the north; the isoline of the preferred Temp_400 m moved southward. At the same time, the accumulation of warm water in the equatorial eastern Pacific and south American enhanced the surface and polar undercurrent in northern Chile. The transport of equatorial warm water to the polar direction depressed the thermocline and reduced the productivity of plankton, promoting a large southward shift of the suitable habitat of D. gigas (Halpern, 2002; Strub and James, 2002; Penven et al., 2005). Arcos et al. (2001) proposed that the habitat of T. murphyi moved southwest under the strong El Niño events. This study showed that the suitable habitat of T. murphyi was mainly distributed in the southwest of Chile during the WE and SE El Niño events, which was consistent with previous studies. In addition, under the ME El Niño events, the gravity center of the suitable habitat of T. murphyi moved northeastward, which may be related to the latitudinal change of the most preferred SST under different-intensity El Niño events (Figure 11).
This study focuses on the coordinated response of the suitable habitats of D. gigas and T. murphyi off Chile in the Southeast Pacific Ocean under the long-time series of El Niño events with different intensities, but there are some uncertainties in this study. Firstly, the research focus of this study is limited to the habitat distribution changes of the two species. There is limited fisheries/survey data to validate the synchronous variation between species and habitat in a long term. Second, the mechanism leading to the difference in SST change under El Niño events with different intensities and the synchronous change mechanism of the suitable habitat of D. gigas and T. murphyi are still unclear. In the future, interdisciplinary studies including but not limited to fishery biology, physiology, and marine science should be conducted to establish an appropriate physical biological coupling model or marine dynamics model in order to explore or clarify the mechanism of the coordinated response of D. gigas and T. murphyi to climate variability.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Ethics Statement
Ethical review and approval were not required for the animal study because this study just uses fisheries and environmental data and did not do any animal experiment.
Author Contributions
WY, ZF, and XC conceptualized the study. WY, YZ, and ZF designed the methodology, provided the software, and analyzed the data for the study. WY and ZF wrote the original draft. WY, YL, and XC involved in the funding acquisition. The manuscript was written through contributions of all authors. All authors contributed to the article and approved the submitted version.
Funding
This study was financially supported by the National Natural Science Foundation of China (41906073), Shanghai talent development funding for the project (2021078), the National Key R&D Program of China (2019YFD0901405), and the open fund of State Key Laboratory of Satellite Ocean Environment Dynamics, Second Institute of Oceanography (QNHX2232).
Conflict of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alizadeh-Choobari O. (2017). Contrasting Global Teleconnection Features of the Eastern Pacific and Central Pacific El Niño Events. Dynam. Atmosph. Oceans. 80, 139–154. doi: 10.1016/j.dynatmoce.2017.10.004
Arcos D. F., Cubillos L. A., Núñez S. P. (2001). The Jack Mackerel Fishery and El Niño 1997-98 Effects Off Chile. Prog. Oceanog. 49 (1-4), 597–617. doi: 10.1016/S0079-6611(01)00043-X
Behera S. K., Doi T., Luo J. J. (2021). Air–Sea Interaction in Tropical Pacific: The Dynamics of El Niño/southern Oscillation. Trop. Extratrop. Air-Sea Interact. 61-92. doi: 10.1016/B978-0-12-818156-0.00005-8
Blanco J. L., Carr M. E., Thomas A. C., Strub P. T. (2002). Hydrographic Conditions Off Northern Chile During the 1996–1998 La Niña and El Niño Events. J. Geophys. Res. 107 (C3), 3017. doi; 10.1029/2001JC001002
Brander K. (2010). Impacts of Climate Change on Fisheries. J. Mar. Syst. 79, 389–402. doi: 10.1016/j.jmarsys.2008.12.015
Brochier Timothée, Echevin V., Tam J., Chaigneau A., Goubanova K., Bertrand A. (2013). Climate Change Scenarios Experiments Predict a Future Reduction in Small Pelagic Fish Recruitment in the Humboldt Current System. Global Change Biol. 19 (6), 1841–1853. doi: 10.1111/gcb.12184
Chang Y. J., Sun C. L., Chen Y., Yeh S. Z., DiNardo G., Su N. (2013). Modelling the Impacts of Environmental Variation on the Habitat Suitability of Swordfish, Xiphias Gladius, in the Equatorial Atlantic Ocean. J. ICES J. Mar. Sci. 70, 1000–1012. doi: 10.1093/icesjms/fss190
Chavez F. P., Bertrand A., Guevara-Carrasco R., Soler P., Csirke J. (2008). The Northern Humboldt Current System: Brief History, Present Status and a View Towards the Future. Prog. Oceanog. 79 (2-4), 95–105. doi: 10.1016/j.pocean.2008.10.012
Chavez F. P., Ryan J., Lluch-Cota S. E., Ñiquen C. M. (2003). From Anchovies to Sardines and Back: Multidecadal Change in the Pacific Ocean. Science 299, 217–221. doi: 10.1126/science.1075880
Chen D., Cane M. A. (2008). El Niño Prediction and Predictability. J. Comput. Phys. 227 (7), 3625–3640. doi: 10.1016/j.jcp.2007.05.014
Chen P., Chen X. J. (2020). Relationship Between the Variation of Peru Coastal Engraulis Ringens Fishing Ground and Oceanic Environmental Factors. J. Shanghai Ocean. Univ. 29 (4), 611–621. doi: 10.12024/jsou.20190702736
Cravatte S., Delcroix T., Zhang D., Mcphaden M., Leloup J. (2009). Observed Freshening and Warming of the Western Pacific Warm Pool. Clim. Dynam. 33 (4), 565–589. doi: 10.1007/s00382-009-0526-7
Cury P., Bakun A., Crawford R. J. M., Jarre A., Quiñones R. A., Shannon L. J., et al. (2000). Small Pelagics in Upwelling Systems: Patterns of Interaction and Structural Changes in "Wasp-Waist" Ecosystems. ICES J Mar. Sci. 57 (3), 603–618. doi: 10.1006/jmsc.2000.0712
Davis A. J., Jenkinson L. S., Lawton J. H., Shorrocks B., Wood S. (1998). Making Mistakes When Predicting Shifts in Species Range in Response to Global Warming. Nature 391 (6669), 783–786. doi: 10.1038/35842
Fang X. Y., Chen X. J., Ding Q. (2014). Optimization Fishing Ground Prediction Models of Dosidicus Gigas in the High Sea Off Chile Based on Habitat Suitability Index. J. Guangdong Ocean. Univ. 34 (4), 67–73. doi: 10.3969/j.issn.1673-9159.2014.04.012
Fang X. N., Yu W., Chen X. J., Zhang Y. (2021). Response of Abundance and Distribution of Humboldt Squid (Dosidicus Gigas) to Short-Lived Eddies in the Eastern Equatorial Pacific Ocean From April to June 2017. Front. Mar. Sci. 8, 721291. doi: 10.3389/fmars.2021.721291
Fang Z. J., Zhi H., Lin P. F., Wei X. (2019). Distinguishing Two Types of El Niño in the Tropical Pacific Using Key Region Sea Surface Salinity Index. J. Trop. Oceanog. 38 (2), 32–42. doi: 10.11978/2018061
Feng Z. P., Yu W., Chen X. J., Liu B. L., Zhang Z. (2020). Analysis of Fishing Ground of Jumbo Flying Squid Dosidicus Gigas in the Southeast Pacific Ocean Off Peru Based on Weighting-Based Habitat Suitability Index Model. J. Shanghai Ocean. Univ. 29 (6), 878–888. doi: 10.12024/jsou.20191102862
Feng Z. P., Yu W., Chen X. J., Zou X. R. (2021b). Distribution of Chilean Jack Mackerel (Trachurus Murphyi) Habitats Off Chile Based on a Maximum Entropy Model. J. Fish. Sci. China 28 (4), 431–441. doi: 10.12264/JFSC2020-0258
Feng Z. P., Zhang Y. J., Yu W., Chen X. J. (2021a). Differences in Habitat Pattern Response to Various ENSO Events in Trachurus Murphyi and Dosidicus Gigas Located Outside the Exclusive Economic Zones of Chile. J. Fish. Sci. China 28 (9), 1195–1207. doi: 10.12264/JFSC2020-0533
Freon P., Barange M., Aristegui J. (2009). Eastern Boundary Upwelling Ecosystems: Integrative and Comparative Approaches. Prog. Oceanog. 83 (1-4), 1–14. doi: 10.1016/j.pocean.2009.08.001
Gerlotto F., Dioses T. (2013). “Bibliographical Synopsis on the Main Traits of Life of Trachurus Murphyi in the South Pacific Ocean,” in The First Meeting of the Scientific Committee of SPRFMO. 21–7. (California, USA).
Goberville E., Beaugrand G., Edwards M. (2014). Synchronous Response of Marine Plankton Ecosystems to Climate in the Northeast Atlantic and the North Sea. J. Mar. Syst. 129, 189–202. doi: 10.1016/j.jmarsys.2013.05.008
Guo A., Zhang Y., Yu W., Chen X. J., Qian W. G., Li Y. S. (2018). Influence of El Niño and La Niña With Different Intensity on Habitat Variation of Chub Mackerel Scomber Japonicas in the Coastal Waters of China. Haiyang Xuebao. 40 (12), 58–67. doi: 10.3969/ji.ssn.0253-4193.2018.12.007
Gutiérrez D., Akester M., Naranjo L. (2016). Productivity and Sustainable Management of the Humboldt Current Large Marine Ecosystem Under Climate Change. Environ. Dev. 17, 126–144. doi: 10.1029/2001GL014097
Halpern D. (2002). Offshore Ekman Transport and Ekman Pumping Off Peru During the 1997–1998 EL Niño. Geophys. Res. Lett. 29 (5), 19–11. doi: 10.1029/2001GL014097
Hastings R. A., Rutterford L. A., Freer J. J., Collins R. A., Simpson S. D., Genner M. J. (2020). Climate Change Drives Poleward Increases and Equatorward Declines in Marine Species. Curr. Biol. 30 (8), 1572–1577. doi: 10.1016/j.cub.2020.02.043
Huang B. Y., Xue Y., Behringer D. W. (2008). Impacts of Argo Salinity in NCEP Global Ocean Data Assimilation System: The Tropical Indian Ocean. J. Geophys. Res.: Oceans. 113 (C8), C08002. doi: 10.1029/2007JC004388
Hu G. Y., Fang Z., Chen X. J. (2018). Review on the Life History of Jumbo Squid (Dosidicus Gigas) in the Eastern Pacific Ocean. J. Fish. China 42 (8), 1315–1328. doi; 10.11964/jfc.20170610869
Ianelli J., Hintzen N. 8th SPRFMO Scientific Committee Meeting Report (Wellington, New Zealand), 3–8, 76 p.
Lee H. C. (2009). Impact of Atmospheric CO2 Doubling on the North Pacific Subtropical Mode Water. Geophys. Res. Lett. 36 (6), 295–311. doi: 10.1029/2008GL037075
Li G., Cao J., Zou X. R., Chen X. J., Runnebaum J. (2016). Modeling Habitat Suitability Index for Chilean Jack Mackerel (Trachurus Murphyi) in the South East Pacific. Fish. Res. 178, 47–60. doi: 10.1016/j.fishres.2015.11.012
Li G., Chen X. J., Lei L., Guan W. J. (2014). Distribution of Hotspots of Chub Mackerel Based on Remote-Sensing Data in Coastal Waters of China. Int. J. Remote Sens. 35 (11-12), 4399–4421. doi: 10.1080/01431161.2014.916057
Liu C. Z., Xue F. (2010). The Decay of El Niño With Different Intensity. The Decay of the Strong El Niño. Chin. J. Geophys. 53 (1), 39–48. doi: 10.3969/j.issn.0001-5733.2010.01.005
McPhaden M. J. (2004). Evolution of the 2002/03 El Niño. Bull. Am. Meteorolog. Soc. 85 (5), 677–696. doi: 10.1175/BAMS-85-5-677
Mogollón R., Calil P. (2017). On the Effects of ENSO on Ocean Biogeochemistry in the Northern Humboldt Current System (NHCS): A Modeling Study. J. Mar. Syst. 172, 137–159. doi: 10.1016/j.jmarsys.2017.03.011
Montecinos A., Aceituno P. (2005). Seasonality of the ENSO-Related Rainfall Variability in Central Chile and Associated Circulation Anomalies. J. Clim. 16 (2), 281–296. doi: 10.1175/1520-0442(2003)016<0281:SOTERR>2.0.CO;2
Neelin J. D. (2010). Climate Change and Climate Modeling. Cambridge Univ. Press Cambridge 92 (92), 198–199. doi: 10.1017/CBO9780511780363
Nguyen H., Hendon H., Lim E., Boschat G., Timbal B. (2017). Variability of the Extent of the Hadley Circulation in the Southern Hemisphere: A Regional Perspective. Clim. Dynam. 21, 1–14. doi: 10.1007/s00382-017-3592-2
Niu M. X., Li X. S., Xu Y. C. (2009). Preliminary Study on Spatio-Temporal Change of Central Fishing Ground of Chilean Jack Mackerel (Trachurus Murphyi) in the Offshore Waters of Chile. Mar. Sci. 33 (11), 105–109. doi: CNKI:SUN:HYKX.0.2009-11-020
Oozeki Y., Carranza M.Ñ., Takasuka A., Dejo P. A., Kuroda H., Malagas J. T., et al. (2019). Synchronous Multi-Species Alternations Between the Northern Humboldt and Kuroshio Current Systems. Deep Sea Res. Part II: Topic. Stud. Oceanog. 159, 11–21. doi: 10.1016/j.dsr2.2018.11.018
Pecl G. T., Jackson G. D. (2008). The Potential Impacts of Cli- Mate Change on Inshore Squid: Biology, Ecology and Fisheries. Rev. Fish Biol. Fish. 18 (4), 373–385. doi: 10.1007/s11160-007-9077-3
Penven P., Echevin V., Pasapera J., Colas F., Tam J. (2005). Average Circulation, Seasonal Cycle, and Mesoscale Dynamics of the Peru Current System: A Modeling Approach. J. Geophys. Res. Oceans. 110 (c10), C10021. doi: 10.1029/2005JC002945
Philander S. G. H. (1983). El Niño Southern Oscillation Phenomena. Nature 302 (5906), 295–301. doi: 10.1038/302295a0
Qi J., Zhang L., Qu T., Yin B., Xu Z., Yang D., et al. (2019). Salinity Variability in the Tropical Pacific During the Central-Pacific and Eastern-Pacific El Niño Events. J. Mar. Syst. 199, 103225. doi: 10.1016/j.jmarsys.2019.103225
Rahn D. A. (2012). Influence of Large-Scale Oscillations on Upwelling-Favorable Coastal Wind Off Central Chile. J. Geophys. Res. Atmos 117, D19114. doi: 10.1029/2012JD018016
Strub P. T., James C. (2002). The 1997–1998 Oceanic El Niño Signal Along the Southeast and Northeast Pacific Boundaries—An Altimetric View. Prog. Oceanog. 54 (1-4), 439–458. doi: 10.1016/S0079-6611(02)00063-0
Sutton R. T., Dong B., Gregory J. M. (2007). Land/sea Warming Ratio in Response to Climate Change: IPCC AR4 Model Results and Comparison With Observations. Geophys. Res. Lett. 34 (2), L02701. doi: 10.1029/2006GL028164
Tian S. Q., Chen X. J., Chen Y., Xu L. X., Dai X. J. (2009). Evaluating Habitat Suitability Indices Derived From CPUE and Fishing Effort Data for Ommatrephes Bartramii in the Northwestern Pacific Ocean. Fish. Res. 95 (2), 181–188. doi: 10.1016/j.fishres.2008.08.012
Tian Y. J., Ueno Y., Suda K., Akmine T. (2004). Decadal Variability in the Abundance of Pacific Saury and Its Response to Climatic/ Oceanic Regi me Shifts in the Northwestern Subtropical Pacific During the Last Half Century. J. Mar. Syst. 52 (1–4), 235–257. doi: 10.1016/j.jmarsys.2004.04.004
Weng H. Y., Ashok K., Behera S. K., Rao S. A., Yamagata T. (2007). Impacts of Recent El Niño Modoki on Dry/Wet Conditions in the Pacific Rim During Boreal Summer. Clim. Dynam. 29, 113–129. doi: 10.1007/s00382-007-0234-0
Wen J., Gong J. W., Li T., Zhao S. L., Tian Y. Y., Chen X. J., et al. (2020). Spatiotemporal Variations in the Habitat of Jumbo Squid Dosidicus Gigas in the Southeast Pacific Ocean Off Peru Under Anomalous Climate Conditions. Haiyang Xuebao. 42 (10), 92–99.doi: 10.3969/j.issn.0253-4193.2020.10.009
Xie F., Zhang M., Chen X. J. (2021). Current Situation and Suggestions on the Exploitation of Jack Mackerel Resources in the Word. J. Fish. Res. 43 (3), 307–315. doi: 10.14012/j.cnki.fjsc.2021.03.010
Xu L. X. (2014). Mode Waters and the North Pacific Subtropical Countercurrent: Its’ Decadal Variability and the Response to Global Warming (Ocean University of China, Qingdao).
Yang X. S., Zhou X. R., Xu X. X., Wang Z. A. (2019). Effects of ENSO on Abundance Index and Spatial-Temporal Change of Chilean Jack Mackerel in the Southeast Pacific Ocean. J. Shanghai Ocean. Univ. 28 (2), 290–297. doi: 0.12024/jsou.20180902406
Yu W., Chen X. ,. J., Zhang Y. (2019). Seasonal Habitat Patterns of Jumbo Flying Squid Dosidicus Gigas Off Peruvian Waters. J. Mar. Syst. 194, 41–51. doi: 10.1016/j.jmarsys.2019.02.011
Yu W., Zhang Y., Chen X. J., Yi Q., Qian W. G. (2018). Response of Winter Cohort Abundance of Japanese Common Squid Todarodes Pacificus to the ENSO Events. Acta Oceanol. Sinica. 37 (6), 61–71. doi: 10.1007/s13131-018-1186-4
Yu J. Y., Zou Y. H. (2013). The Enhanced Drying Effect of Central-Pacific El Niño on US Winter. Environ. Res. Lett. 8 (1), 3865–3879. doi: 10.1088/1748-9326/8/1/014019
Zhang M., Xu L. X. (2000). Analysis and Discussion on the Exploitation and Utilization of the Resources of Trachurus Murphyi in the Southeast Pacific Ocean. Mar. Fish. 22 (3), 137–140. doi: 10.3969/j.issn.1004-2490.2000.03.015
Zhao Y. P., Chen Y. L., Wang F., Wu A. M. (2007). Water Oscillation and ENSO Cycle in the Tropical Pacific Ocean Mixed Layer. Sci. China (D: Earth Sci.). 37 (8), 1120–1133. doi: 10.3321/j.issn:1006-9267.2007.08.016
Zheng F., Wan L. Y., Wang H. (2012). Distinguished Effects of Interannual Salinity Variability on the Development of the Central-Pacific El Niño Events. Atmos. Ocean. Sci. Lett. 5 (2), 123–127. doi: 10.1080/16742834.2012.11446977
Zhi H., Zhang R. H., Lin P., Yu P., Zhou G., Shi S. (2020). Interannual Salinity Variability Associated With the Central Pacific and Eastern Pacific El Niños in the Tropical Pacific. J. Geophys. Res. Oceans. 125, e16090. doi: 10.1029/2020JC016090
Keywords: Dosidicus gigas, Trachurus murphyi, off Chile, spatial distribution, habitat change, El Niño, different intensities
Citation: Feng Z, Yu W, Zhang Y, Li Y and Chen X (2022) Habitat Variations of Two Commercially Valuable Species Along the Chilean Waters Under Different-Intensity El Niño Events. Front. Mar. Sci. 9:919620. doi: 10.3389/fmars.2022.919620
Received: 13 April 2022; Accepted: 16 May 2022;
Published: 13 June 2022.
Edited by:
Jose Luis Iriarte, Austral University of Chile, ChileReviewed by:
Luis A. Cubillos, University of Concepcion, ChileEleuterio Yáñez, Pontificia Universidad Católica de Valparaíso, Chile
Copyright © 2022 Feng, Yu, Zhang, Li and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wei Yu, d3l1QHNob3UuZWR1LmNu