 Juan José Alvarado1,2,3*†
Juan José Alvarado1,2,3*† José Leonardo Chacón-Monge1,2†
José Leonardo Chacón-Monge1,2† Juan Carlos Azofeifa-Solano1,3†
Juan Carlos Azofeifa-Solano1,3† Jorge Cortés1,2,3†
Jorge Cortés1,2,3†- 1Centro de Investigación en Ciencias del Mar y Limología (CIMAR), Universidad de Costa Rica, San José, Costa Rica
- 2Escuela de Biología, Universidad de Costa Rica, San José, Costa Rica
- 3Centro de Investigación en Biodiversidad y Ecología Tropical (CIBET), Universidad de Costa Rica, San José, Costa Rica
Echinoderms are a highly diverse group and one of the most conspicuous in the deep sea, playing ecological key roles. We present a review about the history of expeditions and studies on deep-sea echinoderms in Costa Rica, including an updated list of species. We used literature and information gathered from the databases of the California Academy of Sciences, the Benthic Invertebrate Collection of the Scripps Institution of Oceanography, the National Museum of Natural History, the Museum of Comparative Zoology and the Museo de Zoología from the Universidad de Costa Rica. A total of 124 taxa (75 confirmed species) have been collected from the Costa Rican deep sea, 112 found in the Pacific Ocean, 13 in the Caribbean Sea, and one species shared between the two basins. We report 22 new records for the Eastern Tropical Pacific, 46 for Central American waters, and 58 for Costa Rica. The most specious group was Ophiuroidea with 37 taxa, followed by Holothuroidea (34 taxa), Asteroidea (23 taxa), Echinoidea (17 taxa), and Crinoidea (11 taxa). The highest number of species (64) was found between 800 m and 1200 m depth. Only two species were found deeper than 3200 m. Further efforts on identification will be required for a better comprehension of the diversity of deep-sea echinoderms. Limited research has been done regarding the biology and ecology of deep-sea echinoderms in Costa Rica, so additional approaches will be necessary to understand their ecological functions.
Introduction
Costa Rica is located in the Central American Isthmus facing the Caribbean Sea and the Pacific Ocean, it has an oceanic island 500 km offshore (Isla del Coco), and a marine extension covering 572,877 km2, that is more than 11-fold its land area, making it the largest country on Central America (Cortés, 2008; Cortés, 2016a; Cortés, 2016b; Cortés, 2016c). Recent studies estimated that over 60% of the total area of the country is below 2,000 m depth, and over one-third below 3,000 m (Cortés and Benavides-Varela, in prep.). Eastern Tropical Pacific (ETP) deep waters (below 200 m) constitute a particularly important ecosystem in Costa Rica, since they represent about 90% of the whole territory and the last frontier in the international scientific agendas (Cortés, 2019; Rojas-Jiménez et al., 2020; Azofeifa-Solano and Cortés, 2021).
Few explorations have been done on the Costa Rican Caribbean (Alvarado et al., 2013; Cortés, 2016c; Cambronero-Solano et al., 2019). The continental shelf is relatively narrow, with a total area of 2,310 km2, and an Exclusive Economic Zone (EEZ) of 28,064 km2 (Cortés, 2016c; Cortés and Benavides-Varela, in prep.). In the nearshore area, there are mangrove forests, seagrass beds, coral reefs, rocky shores, and sandy beaches, further offshore the bottom is overall sandy, but muddy near river mouths, also some shallow calcareous mounds have been observed (Cortés, 2016c).
The Costa Rican Pacific area is much larger than the Caribbean, covering 544,813 km2, hosting a greater variety of deep-sea environments (Cortés, 2016a; Cortés, 2016b). There are mesophotic ecosystems (Cortés, 2019), methane seeps (Sahling et al., 2008; Levin et al., 2012; Levin et al., 2015), seamounts (Auscavitch, 2020), deep areas of fluid discharge (Wheat et al., 2019), extensive abyssal plains (Agassiz, 1898; Townsend, 1901) and a large deep pelagic region. The extensive Coco Submarine Volcanic Range (CSVR, also called Cocos Ridge) has been explored too (Neuhaus, 2004; Alvarado-Induni, 2021).
For Costa Rica, there have been registered ~306 echinoderm species (Alvarado et al., 2017; Cambronero-Solano et al., 2019; Chacón-Monge et al., 2021), of the 420 reported species for Central America (Alvarado and Fabregat-Malé, 2021). Most of them inhabit shallow water or intertidal environments, mainly related to coral and rocky reefs, sand, and mudflats (Solís-Marín et al., 2013). Most collecting efforts have been carried out on Coco, Caño, and Murciélago islands in the Pacific Ocean, while on the Caribbean it has been between Punta Cahuita and Punta Mona (Alvarado et al., 2017; Chacón-Monge et al., 2021). However, the real status of the deep-sea echinoderm’s fauna has not been evaluated. Deep-water biodiversity assessments have always been a challenge due to the complexity and cost in logistic operations (Costello and Chaudhary, 2017). In this paper, we undertook the task of compiling the available information in the literature and museum collections about deep-sea echinoderms collected in Costa Rica. In this way, we are aiming to have a better understanding of the deep-sea fauna contribution to the total echinoderm diversity in Costa Rican waters. Likewise, to be able to identify areas or groups that require a greater sampling effort for future collections and research.
Material and Methods
To compile the list of deep-sea echinoderms collected in Costa Rica, a literature search was carried out to corroborate records, ecological investigations, and further aspects of deep-sea Costa Rican echinoderms. In addition, the following databases of biological collections were visited: 1) California Academy of Sciences (CAS: http://researcharchive.calacademy.org/research/izg/); 2) Benthic Invertebrate Collection, Scripps Institution of Oceanography (SIO: https://scripps.ucsd.edu/benthic-invertebrate-collection); 3) National Museum of Natural History, Smithsonian Institution (NMNH: http://collections.nmnh.si.edu/); 4) Museum of Comparative Zoology, Harvard University (MCZ: https://mcz.harvard.edu/database; and 5) Museo de Zoología, Universidad de Costa Rica (MZUCR).
A list of deep-water echinoderms collected in Costa Rican waters was elaborated (Table 1), following the phylogenetic order for the five classes of Echinodermata according to the World Register of Marine Species (WoRMS, WoRMS Editorial Board, 2022), including the taxonomy, location, depth, status as new record for the country or Central America, the reported depth range (if available), and the references. A map of the Costa Rican Exclusive Economic Zones was constructed using ArcGIS Desktop 10.8, plotting the location, species richness, and abundance of specimens (Figure 1). We followed the definition for the deep sea as oceanic waters and seabed below 200 m depth (UNESCO, 2009).
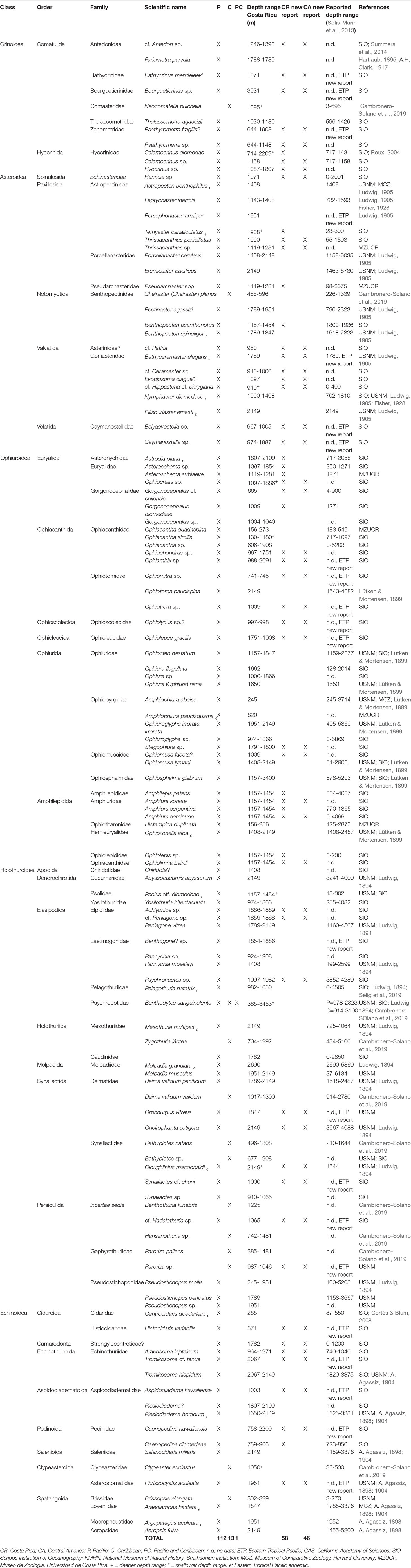
Table 1 Taxonomic list of deep-sea echinoderms from Costa Rican waters. CR: Costa Rica; CA: Central America; P: Pacific; C: Caribbean; PC: Pacific and Caribbean; n.d: no data; ETP: Eastern Tropical Pacific; CAS: California Academy of Sciences; SIO: Scripps Institution of Oceanography; NMHN: National Museum of Natural History, Smithsonian Institution; MCZ: Museum of Comparative Zoology, Harvard University; MZUCR: Museo de Zoología, Universidad de Costa Rica. + = deeper depth range; * = shallower depth range. ϵ: Eastern Tropical Pacific endemic.
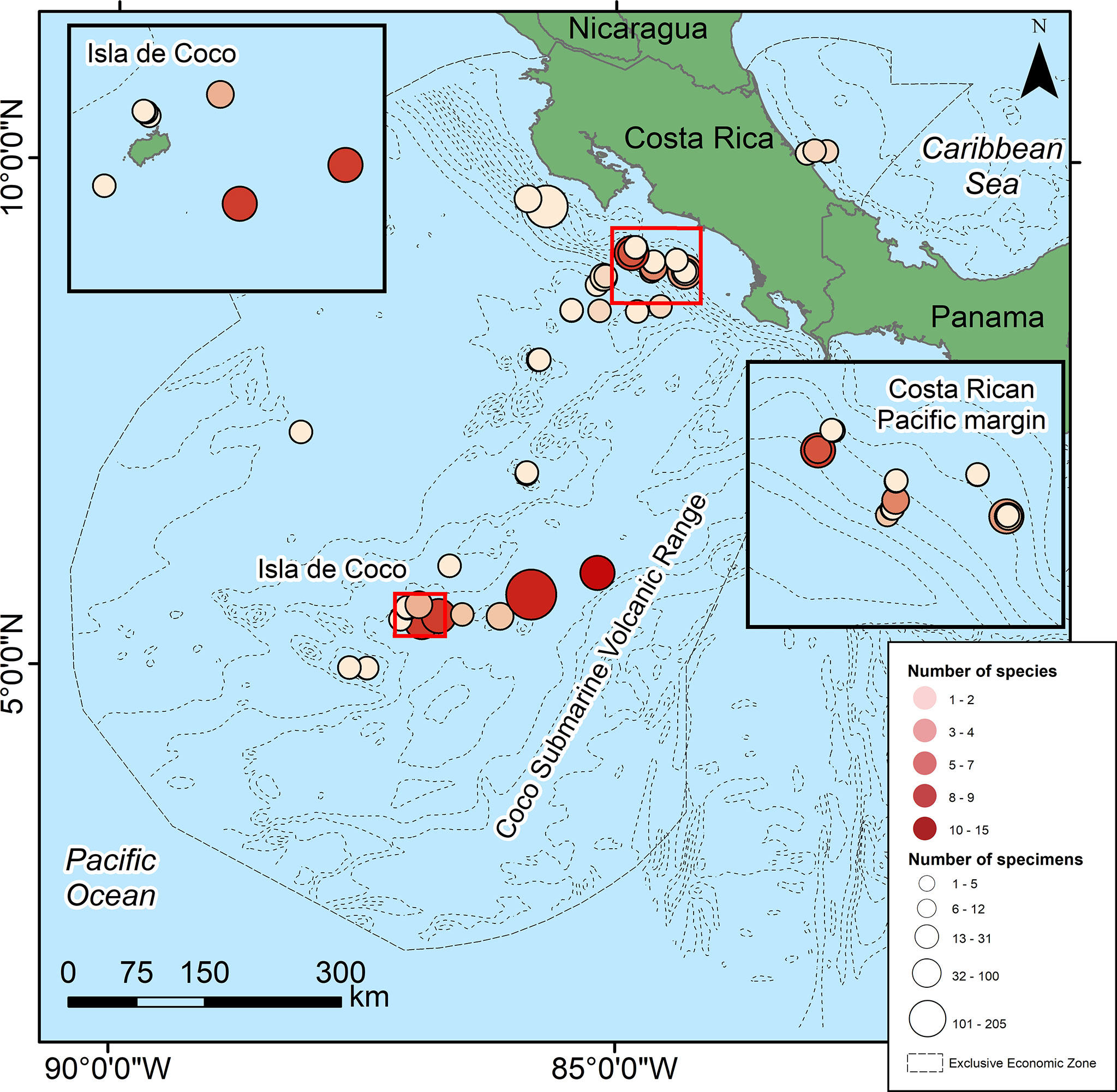
Figure 1 Sampling localities of deep-sea echinoderms recorded on museum collections (Scripps Institution of Oceanography, National Museum of Natural History of Smithsonian Institution, Museum of Comparative Zoology of Harvard University, Museo de Zoología Universidad de Costa Rica) from the Costa Rican Economic Exclusive Zone.
The biogeographic affinity of the echinoderm species was estimated based on the Ocean Biodiversity Information System (www.obis.org) and WoRMS (WoRMS Editorial Board, 2022). We used the classification of marine biogeographic realms proposed by Costello et al. (2017). We found biogeographic affinities for only 120 species. Based on this affinity, a presence/absence matrix was developed, and a Bray-Curtis similarity matrix was made. A Non-metric Multi-Dimensional Scaling was elaborated, and we overlay vectors using a Pearson correlation based on the realms indicated by the Similarity Percentages - species contributions analysis (SIMPER). These analyzes were performed in PRIMER 7.0.
Results
A total of 124 taxa of deep-sea echinoderms have been collected in Costa Rica, 75 identified to species level with certainty (Tables 1, 2), the remaining 49 representing morphospecies (i.e. morphologically similar individuals). There are 112 taxa for the Pacific Ocean and 13 for the Caribbean Sea, with only one shared species between the two basins (the holothuroid Benthodytes sanguinolenta). We found 22 new reports for the Eastern Tropical Pacific, 46 for Central America, and 58 for Costa Rica (57 on the Pacific Ocean and one in the Caribbean Sea) (Table 1). With this, we reached a total of 364 species of echinoderms for Costa Rica, 78% of the total species registered in Central America (466). 34% of the echinoderm species of Costa Rica are deep-sea species.
In the Costa Rican deep sea, the most diverse class is Ophiuroidea with 37 taxa and 22 confirmed species, all from the Pacific (Table 2), with the families Ophiacanthidae and Ophiopyrgidae as the most specious (five taxa each). The Holothuroidea is the second in richness, with 35 taxa in 19 species, the Synallactidae had most of the taxa (five), followed by Laetmogonidae and Deimatidae (four each). Asteroidea and Echinoidea have 24 and 18 taxa, respectively (14 and 15 confirmed species, respectively). For the asteroids, the families Goniasteridae and Astropectinidae were the richest (six taxa each). While Echinothutiridae and Aspidodiadematidae were the most specious families in Echinoidea (three taxa each). Finally, Crinoidea have 10 taxa and five confirmed species, where Hyocrinidae is the richest family (three genera) (Table 1, 2).
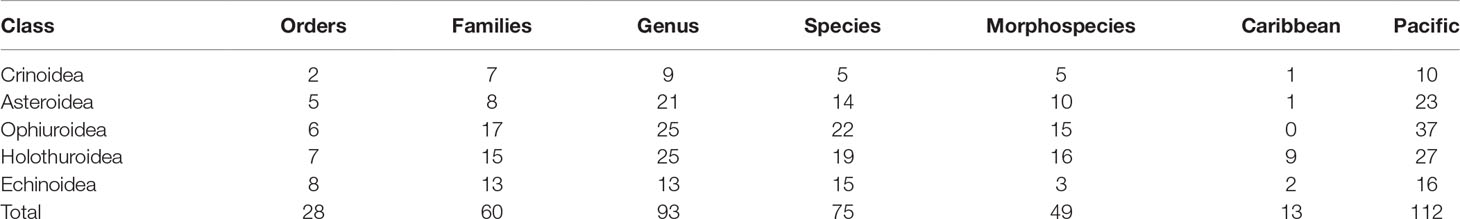
Table 2 Number of deep-sea echinoderms taxa at different taxonomic levels from Costa Rican waters.
According to their distribution and diversity (Figure 1), the greatest collecting effort has been around the CSVR, Isla del Coco, the seamount subduction zone, and along the continental margin (9°4’26.09’’-8°36’37.88’’N and 85°14’11.77’’-84°17’050’’W) on the Pacific side, and in front of the Port of Limón (10°4’35.77’’N-82°54’9.38’’W) on the Caribbean coast. Other areas of the country have been scarcely sampled, like the extensive abyssal plains of the Pacific and most of the EEZ in the Caribbean (Figure 1).
The greatest number of taxa by depth range (Figure 2) was found between 800 and 1200 m with 64 taxa, followed by 52 and 51 taxa between 1600-2000 m and 1200-1600 m, respectively. The holothuroid Benthodytes sanguinolenta and the ophiuroid Ophiosphalma glabrum were the species with the largest bathymetric distribution range (385-3453 m and 1157-3400 m, respectively), and the only two species found deeper than 3200 m. One species of ophiuroid, Ophiacantha similis, had a shallower deep range than previously reported, while the holothuroids, Psolus aff. diomedeae, Benthodytes sanguinolenta, Oloughlinius macdonaldi, and the echinoid, Clypeaster euclastus, had deeper depth ranges than previously reported (Table 1).
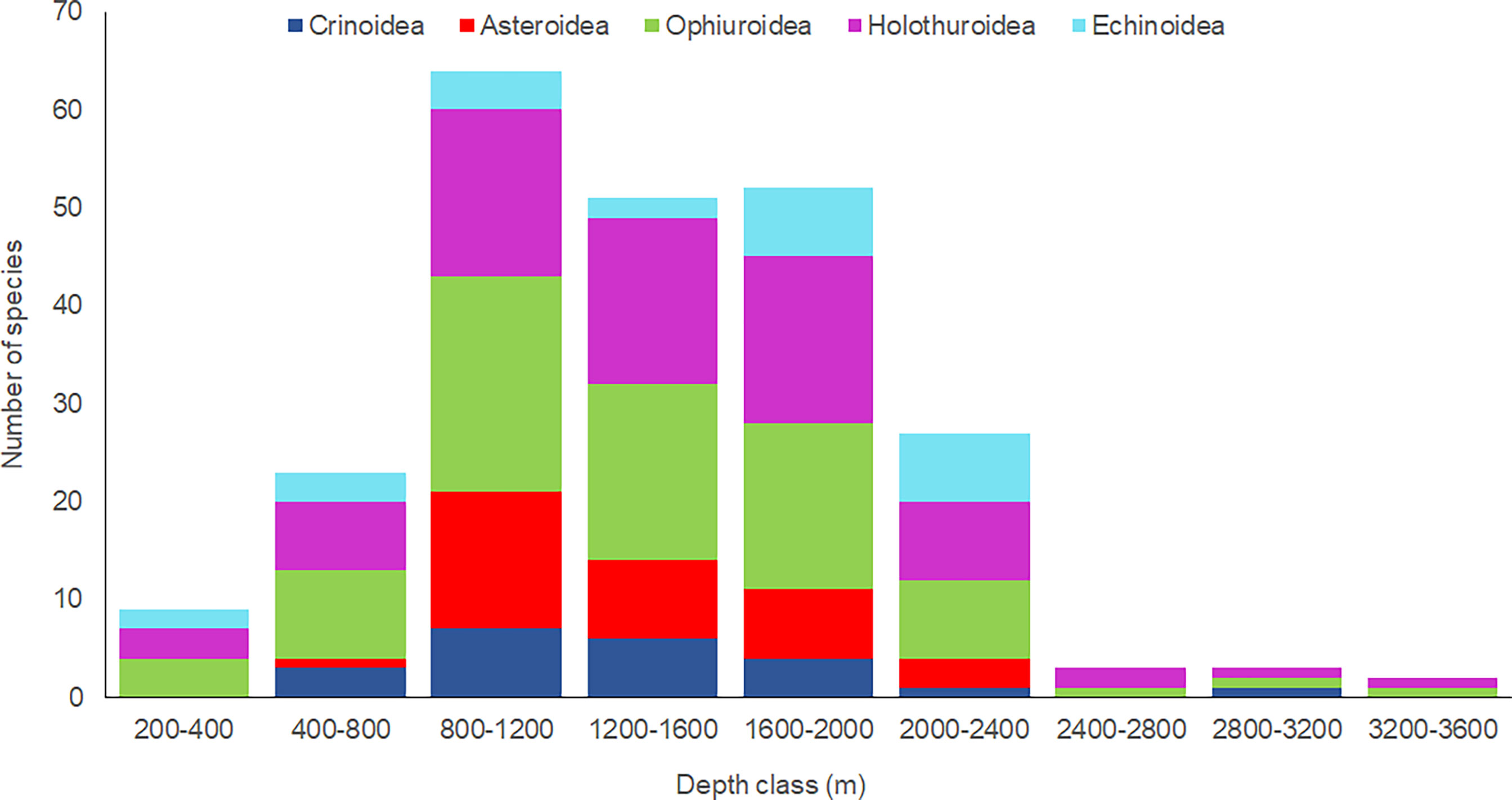
Figure 2 Bathymetric variation in deep sea echinoderm species richness by class.
Most taxa of deep-sea echinoderms collected in Costa Rica are vouchered in the collections at SIO (73 species) and the NMNH (35 species) (Figure 3). Otherwise, the MZUCR and MCZ have six taxa and three species respectively. No reports were found in the CAS database. Additionally, we found 49 records in the literature (Table 1), mostly all deposited in museum collections, except for the echinoderms reported by Cambronero-Solano et al. (2019) for the Caribbean Sea.
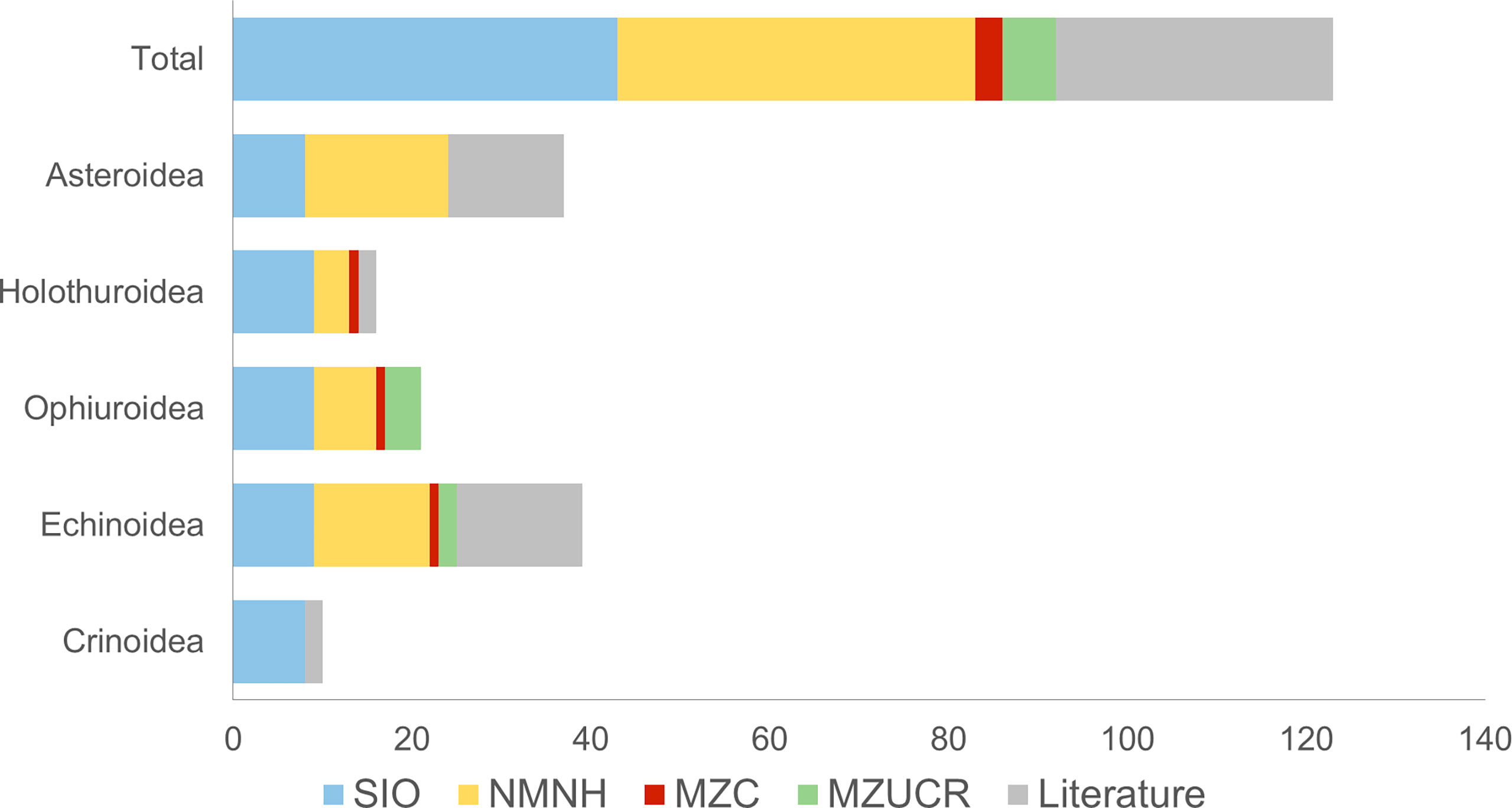
Figure 3 Number of echinoderms species registered by museum collection or literature for Costa Rica deep waters. Scripps Oceanographic Institution (SIO); National Museum of Natural History, Smithsonian Institution (NMNH); Museum of Comparative Zoology, Harvard University (MCZ); Museo de Zoología, Universidad de Costa Rica (MZUCR).
About 78% of the species present in the Costa Rican deep sea have a biogeographic affinity with the Eastern Tropical Pacific realm (Table 3), 23 species are endemic to this region (Table 1). Other regions with higher biogeographic affinity are the North Pacific (40%), followed by Tropical Indo-Pacific & coastal Indian Ocean (36%), Western Tropical Atlantic (35%), offshore Western Pacific and South Australia with 33% both. By class, most of the species have an ETP affinity, there are also several species that can be found in other regions (Table 3; Figure 4).

Table 3 Biogeographic distribution of Costa Rican deep sea echinoderms species based on the Ocean Biodiversity Information System (www.obis.org) and the World Register of Marine Species (www.marinespecies.org), and according to the marine biogeographic realms proposed by Costello et al. (2017).
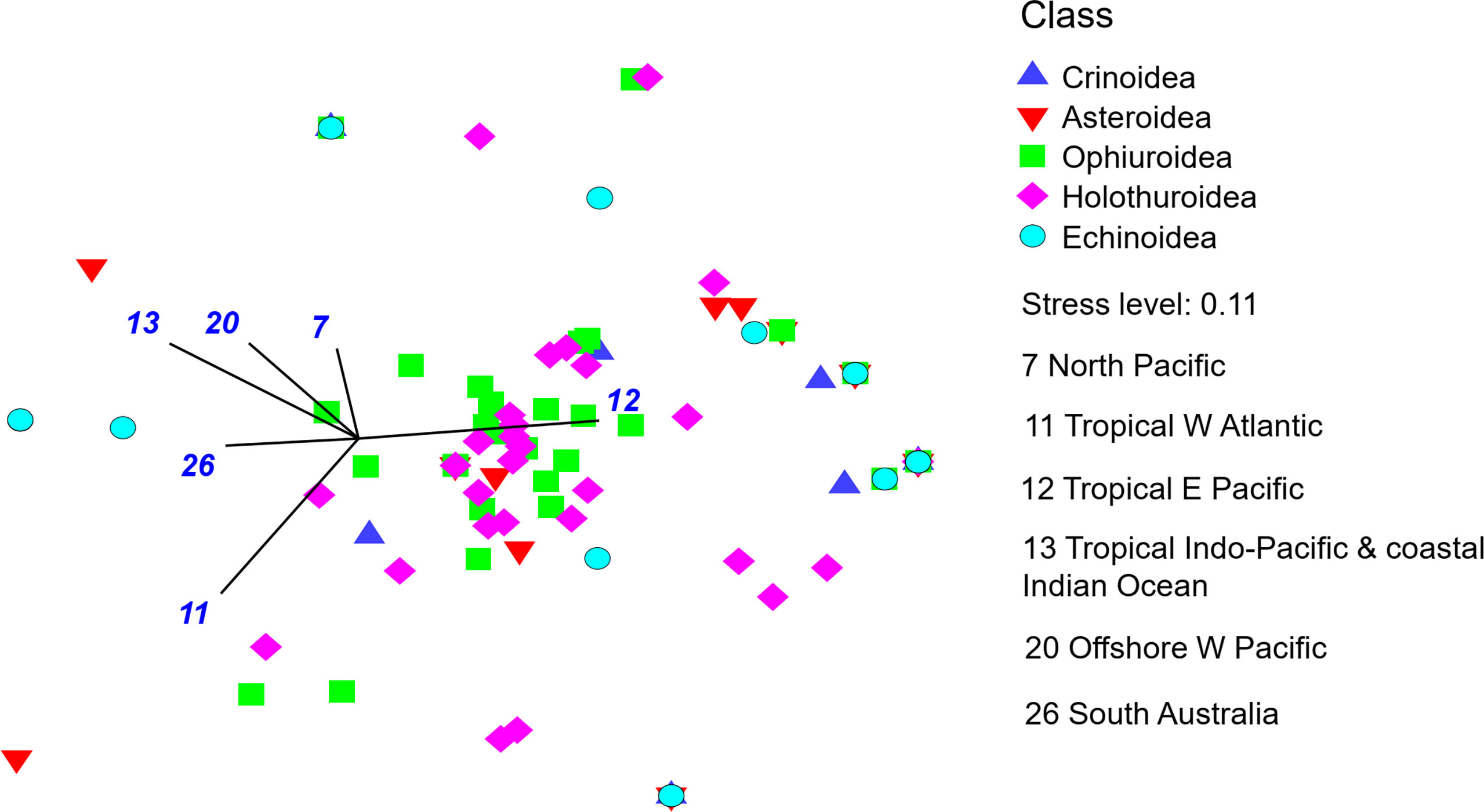
Figure 4 A Non-metric Multi-Dimensional Scaling based on a Bray-Curtis similarity matrix of the presence/absence of the biogeographic affinities of Costa Rica deep sea echinoderms. Vectors indicated the greatest contributions of the realms based on the SIMPER analysis. Number is blue refer to the marine biogeographic realms proposed by Costello et al. (2017) in Table 3.
Discussion
Historical Perspectives
The Costa Rican coastal areas have been relatively well studied, but not so the deep regions (Cortés, 2009; Cortés, 2016a; Cortés, 2016b; Cortés, 2016c). In the Caribbean, few studies have been carried out, and only two publications mentioned echinoderms collected below 200 m depth (Voss, 1971; Cambronero-Solano et al., 2019). In 1971, the R/V John Elliot Pillsbury, cruised to Central America, sampling off the Costa Rican Caribbean coast at two deep stations (Voss, 1971). Voss (1971) reported “small ophiuroids” (Ophiuroidea) and three “large sea biscuits” (Echinoidea: Spatangoida) from a “gray muddy bottom”.
In 2011, the R/V Miguel Oliver, under the Central America Fisheries and Aquaculture Organization (OSPESCA), trawled into two areas of the Costa Rican Caribbean. Trawls ranged from 385 to 1,481 m and found six new echinoderm reports for Costa Rica (Cambronero-Solano et al., 2019). Holothuroids made up 99% of the total biomass, mainly comprised of Benthodytes sanguinolenta (60%) (Cambronero-Solano et al., 2019).
The deep waters of the Costa Rican Pacific were first studied in the late 19th century, when the United States Fisheries Committee visited the area with the steamer USS Albatross, with Alexander Agassiz as Chief Scientist (Agassiz, 1898; Townsend, 1901; Agassiz, 1904). They sampled near Isla del Coco between February and March 1891 and carried dredging and trawling on abyssal plains at 12 stations (#3362-3373), ranging from 95 m to 3433 m (Azofeifa-Solano and Cortés, 2021). From the material collected during the Albatross expeditions, Ludwig (1894; 1905), Hartlaub (1895); Agassiz (1898; 1904), Lütken and Mortensen (1899), and Clark (1917) described many Central American deep-sea echinoderms (Alvarado et al., 2013). Ludwig (1894) worked on Holothuroidea and recorded 11 species from Costa Rican deep waters. Hartlaub (1895) reported one comatulid Crinoidean species. In the preliminary report on the Echini, Agassiz (1898) registered five Costa Rican deep-sea urchin species, later he added one species (Agassiz, 1904). Lütken and Mortensen (1899) worked on Ophiuroidea and reported eight species from Costa Rican deep sea. Finally, Ludwig (1905) reported 14 deep-sea species of Asteroidea for Costa Rican deep sea.In 1925, during the oceanographic expedition of the Zoological Society of New York, with the yacht Arcturus led by William Beebe, deep-sea stars were collected from one station close to Isla del Coco (Fisher, 1928).In 1973, the Scripps Institution of Oceanography realized the first expedition to the Eastern Pacific (EP) with the R/V Agassiz, in which some sample stations included deep-sea echinoderms (Luke, 1982; Cortés, 2009). In 1986, a submersible was used at Isla del Coco, the Johnson Sea-Link I of Harbor Branch Oceanographic Institute (HBOI), as part of the SeaPharm Project (a pharmaceutical prospecting expedition). Six dives were done ranging from 105 to 785 m (Cortés, 2008). The crinoid Calamocrinus diomedae was collected southeast Isla del Coco at 714 m deep and cataloged in the collections of the HBIO (Roux, 2004). Starting in 2007 a three-person submarine, DeepSee, able to reach 450 m has been used regularly at Isla del Coco. Echinoderms and other taxonomic groups have been video recorded and/or collected (Cortés, 2008; Cortés and Blum, 2008).
Since the 1990’s the methane seeps along the Pacific margin have drawn attention, mainly from a geological point of view, but also there are some images of deep-water echinoderms available (Sahling et al., 2008). The methane seeps off the Central Pacific coast of Costa Rica have been studied using the Human Operated Vehicle (HOV) Alvin in 2009-2010, led by Lisa A. Levin, and again in 2017-2018, led by Erik E. Cordes, this last one also included the exploration of several seamounts farther offshore. Echinoderms were collected between 974-1866 m depth, most of the specimens were deposited at SIO.Based on the 2009-2010 expeditions, Levin et al. (2012) described “hydrothermal seep ecosystems” hosting high densities of ophiuroids (Ophiuridae). Levin et al. (2015) demonstrated the role of authigenic carbonate rocks, providing a unique habitat and food resources for macrofaunal assemblages at seep sites on the Costa Rican margin (400-1850 m). The presence of high densities of ophiuroids is strongly related to overlying water’s hydrography. Based on those collections, Summers et al. (2014) described species of Myzostomida (Annelida) which are obligate associates, mostly of echinoderms. One of them, Pulvinomyzostomum inaki Summers & Rouse, 2014, was collected on the crinoid Antedon sp., at Jaco Scarp in 2009.In 2019 an expedition led by Erik E. Cordes visited the methane seeps and more seamounts using the ROV SuBastian onboard the R/V Falkor. Echinoderms were video recorded and collected. Selig et al. (2019), reported aggregations of Pelagothuria sp. in deep sea regions of minimum oxygen concentration, based on data from R/V Falkor that includes observations from Costa Rica.
Deep-Sea Echinoderm Fauna
According to the compiled information, the main component of the known deep sea echinoderm fauna in Costa Rica inhabits from the bathyal to the abyssal zone (200-3000 m depth). This view could be biased by the exploration approaches, the scientific historic background and the relative representativeness of the deep sea total area covered (particularly scarce in the Caribbean) and their proportional size, but depicts the biogeographic predicted pattern (Mah and Blake, 2012; Stöhr et al., 2012; Pérez-Ruzafa et al., 2013). It is important to recognize that the taxa list reported in this study was obtained from literature and zoological museum databases, in which taxa are classified at the genus level or identified with affinities towards a species. Therefore, further taxonomic efforts are required.
For Crinoids, the genus Antendon is reported in the Costa Rican deep sea, but the only species listed near the ETP is A. bifida from Chile (Solís-Marín et al., 2013). Similarly, the only Psathyrometra reported before was P. bigradata, from Panama, Galapagos and Chile (Solís-Marín et al., 2013), but now there is also P. fragilis from Costa Rica. For Calamocrinus, the only species reported is Calamocrinus diomedae, from Panama and Galapagos, while Hyocrinus foelli is also the only species reported for the genus, in Mexico (Solís-Marín et al., 2013).
In Asteroidea, at museum collections we found Thrissacanthias sp., being T. penicillatus the only species reported for Mexico and Peru (Solís-Marín et al., 2013). In other genera, the situation is more complex, as there are different possible species. Pseudarchaster has five species in the ETP: P. discus, P. pectinifer, P. pulcher, P. pusilus, and P. verrilli, scattered from Mexico to Peru. For Patiria two species have been identified, P. chilensis (Chile-Pascua) and P. miniata (Mexico-Revillagigedo), but with bathymetric distributions below 40 m and 300 m respectively, while our specimen was found at 950 m. Ceramaster (Figure 5) has three species (C. grenadensis, C. patagonicus and C. leptoceramus) while Henricia has nine, all with overlapping bathymetric ranges but with geographically isolated records (USA, Mexico, Galapagos, Peru and Chile) (Solís-Marín et al., 2013). Evoplosoma claguei is the only reported species for this genus in Mexico, while Hippasteria has two, H. phrygiana and H. falklandica, both from deep waters, but with different geographic distribution, one is present in the Mexican Pacific waters while the other is from Chile (Solís-Marín et al., 2013). Finally, the family Caymanostellidae has been registered for the first time in the ETP. The genus Belyaevostella has two recognized species, while the genus Caymanostella has five. Belyaevostella hyugaensis was described from samples collected on sunken wood from the Southern of Japan (Fujita et al., 1994), while Belyaevostella hispida have been described from deep sea Indo-Malaysian region (Aziz and Jangoux, 1984), otherwise Caymanostella spp. are widely distributed (Table 3).

Figure 5 Costa Rican deep-sea echinoderms. (A) Antedonidae sp.; (B) Antedonidae sp.; (C) Ceramaster sp.; (D) Asteroschema sp.; (E) Ophiacantha sp. and the fish Lophiodes caulinaris (Lophiidae); (F) Synallactida sp.; (G) Psychropotidae sp.; (H) Peniagone sp.; (I) Caenopedina sp.; (J) Aspidodiadema sp. Images were taken by the HOV Alvin and ROV SuBastian onboard of the Costa Rica expeditions.
For Ophiuroidea, Ophiocreas is a new report for Central America and has at least 16 species described. Asteroschema (Figure 5) and Gorgonocephalus have two species reported for the ETP (Solís-Marín et al., 2013); A. rubrum from Pascua (below 731 m) and A. sblaeve from the USA (1235 m), Guatemala, Panama (1271 m), and Costa Rica (1119-1281 m); G. chilensis was registered from Chile (4-900 m) while G. diomedeae was found in Panama (1271 m). The worldwide distributed Ophiacantha has 25 species present in the ETP, but only O. phragma is registered for Costa Rica, at Isla del Coco (Solís-Marín et al., 2013). For Ophiochondrus, only O. stelliger is reported for Chile, between 73-439 m, while our specimen was collected between 1005-1008 m, and for Ophiambix there are about six species and is a genus widely distributed (Table 3). Ophiomitra has 11 valid species and Ophiotreta 17 (Stöhr et al., 2022), both genera are widely distributed (Table 3). Ophioscolecidae is a new family for the ETP, where the widely distributed genus Ophiolycus has three species (Stöhr et al., 2022). Ophiura has 16 species from shallow to deep sea on the ETP, two of them have been identified from Isla del Coco (Solís-Marín et al., 2013). In the family Ophiopyrgidae, both Ophiuroglypha and Stegophiura are widely distributed genera (Table 3), which have about 19 recognized species each (Stöhr et al., 2022), only one registered at Isla del Coco for Ophiuroglypha (Solís-Marín et al., 2013) while Stegophiura has been found in USA and Mexico. Ophiomusa is the unique genus for the family Ophiomusaidae, but it has about 50 valid and widely distributed species (Table 3). Finally, Ophiolepis has five species reported from the ETP nevertheless, only O. crassa, reach deeps beyond 200 m.
In holothuroids, Chiridota pisanii has been the unique species found in the deep sea for the genus in the ETP, but Psolus is represented by eight species at wide depths ranges. P. digitatus has been listed for Panama, P. diomedae is reported for Mexico, Isla del Coco, Panama, and Galapagos, while P. squamatus for Mexico and Chile (Solís-Marín et al., 2013). For the genus Achlyonice, the only species reported for the ETP is A. ecalcarea, while Peniagone has seven deep sea species at the ETP. P. vitrea has the largest extension range including Isla del Coco, P. papillata is also presented in Panama, but others are from Galapagos, Ecuador and Peru (Solís-Marín et al., 2013). The family Laetmogonidae is represented with three genera, Benthogone is widely distributed and has three accepted species (Table 3). The genus Pannychia has two species; P. taylorae was described from the Indian Ocean (O’Loughlin et al., 2013), while P. moseleyi has been found in Mexico, Panama, Galapagos and Peru. Psychronaetes hanseni is monospecific for its genus and was described from the Eastern Central Pacific Ocean (Pawson, 1983). Synallactidae is represented in two genera, Bathyplotes natans has been found in the Caribbean deep sea from Costa Rica but also in other countries in Latin America at both Pacific and Atlantic basins and also there have been another three species registered for the ETP (Solís-Marín et al., 2013). Synallactes has three species reported for Panama, Colombia, Malpelo, Galapagos, Ecuador and Peru, S. chuni has been reported from West Pacific (Table 3). Hadalothuria and Hansenothuria are monospecific genera. Hadalothuria wolffi was described from the hadal zone (>6000 m; Hansen, 1956) however, our specimen was found on the mesopelagic zone (~1000 m), whereas Hansenothuria benti was described from the Tropical Western Atlantic (Miller and Pawson, 1989). Gephyrothuriidae is a new family in the ETP represented by the genus Paroriza, which has four recognized species, only P. prouhoi has been reported from Chile. Finally, the genus Pseudostichopus registered three species at deep sea in ETP, P. mcdonaldi and P. mollis were found at Isla del Coco, but the second is more widely distributed (Solís-Marín et al., 2013).
In the case of echinoids, the echinothuriids Tromikosoma and Plesiodiadema have two species in the region each, and for both they have been previously listed from ETP at similar depths (Solís-Marín et al., 2013). Tromikosoma hispidum and P. horridum were reported for Isla del Coco and Costa Rica (Solís-Marín et al., 2013).
Regional Comparison
In Latin America, most echinoderm species were found only in one or two bathymetric intervals, Ophiuroidea was dominant from 200 to 2000 m at Pacific and West-Atlantic basins, but Asteroidea and Holothuroidea (respectively) where dominant from 2000 to 6000 m (Pérez-Ruzafa et al., 2013). For Asteroidea as also as Ophiuroidea, it has been suggested a highly conservative morphology in deep sea species and recent molecular analysis are suggesting cryptic and species complexes, thus deep sea and especially abyssal echinoderm diversity is considered underestimated (Mah and Blake, 2012; Stöhr et al., 2012).
Diversity information of deep-sea echinoderm fauna through recent research is scarce (Pawson, 1982; Stöhr and Segonzac, 2005; Mecho et al., 2014; Moles et al., 2015; Calero et al., 2017; Mironov et al., 2018; Setyastuti and Wirawati, 2018; Stöhr and O’Hara, 2021). Strong progress has been made, especially for Colombia, Chile, Brazil, Argentina and Mexico (González et al., 2002; Borrero-Pérez et al., 2003; Benavides-Serrato and Borrero-Pérez, 2010; Campos et al., 2010; Manso, 2010; Massin and Hendrickx, 2011; Borrero-Pérez et al., 2012; Hendrickx et al., 2014; Solís-Marín et al., 2014; Martínez et al., 2014; Martínez et al., 2015; Martínez, 2016; Conejeros-Vargas et al., 2017; Martinez and Penchaszadeh, 2017;Martínez et al., 2017, Rivadeneira et al., 2017; Luna-Cruz and Hendrickx, 2018; Borrero-Pérez et al., 2019; Flores et al., 2019; Pertossi et al., 2019; Martínez et al., 2019; Borrero-Pérez et al., 2020; Martinez et al., 2020; Rivadeneira et al., 2020; Luna-Cruz and Hendrickx, 2020; Catalán et al., 2020; Luna-Cruz and Hendrickx, 2021; Flores et al., 2021).The deep sea of the Colombian Caribbean has been extensively studied (González et al., 2002; Borrero-Pérez et al., 2003; Benavides-Serrato and Borrero-Pérez, 2010; Borrero-Pérez et al., 2012; Borrero-Pérez et al., 2019; Dueñas et al., 2021). For the Southern Colombian Caribbean region, 16 species of sea cucumbers are found between 596 and 2566 m (Borrero-Pérez et al., 2020), of which only four have been reported for the Costa Rican Caribbean (B. sanguinolenta, Benthothuria funebris, Deima validum validum, and Paroriza pallens). For the SeaFlower Biosphere Reserve area (Borrero-Pérez et al., 2019), a region immediately adjacent to the exclusive economic zone of Costa Rica, they found 111 deep-sea echinoderm species, that represents 10 times more species than those reported for the Costa Rican Caribbean (Cambronero-Solano et al., 2019). Dueñas et al. (2021) reported the presence at cold-seep communities in the Colombian Caribbean between 2300 and 3300 m including the sea star family, Solasteridae, and two sea cucumbers, Chiridota cf. heheva and Pseudostichopus sp. This indicates a high potential for research and discovery of new reports for our waters.
In the ETP, Stöhr and O’Hara (2021) report 17 species of ophiuoids from waters deeper than 400 m as part of the Danish Galathea II Expedition, at stations in Nicaragua and Panama. Of these 17 species, only three are reported in Costa Rican waters in our review (Astrodia plana, Ophiosphalma glabrum, and Ophiura flagellata), which indicates a high potential for an increase in the number of ophiuroids in our region. Manso (2010) reports the presence of 15 species of brittle stars in Chile, of which only three are present in our list (Gorgonocephalus chilensis, Ophiolimna bairdi and Ophiomusium lymani). According to Stöhr et al. (2012), globally, the greatest diversity of ophiuroids occurs in shallow waters between 0 and 200 m deep, with 1313 species. Between 200 and 3500 m they indicate that there are 1297 species. For the ETP, they mention that the greatest diversity occurs between 200 and 3500 m with 111 species, while for shallow waters only 92 species. For the abyssal and hadal zone they indicate 28 and one species, respectively. For the Western Atlantic, the species richness of brittle stars is quite similar between the platform (217) and the bathyal zone (229), and only 16 species for the abyssal zone.
Perhaps the country in the region that has the most complete evaluation of its deep-sea fauna is Mexico (Solís-Marín et al., 2014). This country has 348 species of echinoderms that inhabit deep waters, which corresponds to 54.4% of the total species reported for the country. At the Caribbean and Gulf of Mexico they have been listed 111 and 103 deep sea species respectively (Crinoidea 25, Asteroidea 39, Ophiuroidea 100, Holothuroidea 9, Echinoidea 41) but sharing some species, while for the Pacific coast a total of 188 species are included (Crinoidea 3, Asteroidea 63, Ophiuroidea 61, Holothuroidea 34, Echinoidea 26) (Solís-Marín et al., 2014; Conejeros-Vargas et al., 2017). For the west coast of the Baja California Peninsula, Luna-Cruz and Hendrickx (2021) indicate the presence of 18 species of sea cucumbers between 554 m and 2082 m depth. Probably many of the widespread West-Atlantic or Pacific deep sea Asteroidea and Ophiuroidea present in Mexico are also found in Costa Rica (Pawson et al., 2015; Conejeros-Vargas et al., 2017; Luna-Cruz and Hendrickx, 2021). In Costa Rica, the deep fauna corresponds to 33% of the total diversity of echinoderms, while the Mexican coast and its economic exclusive zone are much larger than those of Costa Rica, and the sampling effort has been greater due to the presence of several oceanographic vessels such as the R/V Puma or R/V Justo Sierra of the National Autonomous University of Mexico, among others.
Threats for Conservation
The deep sea is under increasing pressure from exploration and extraction activities during the last decades, fueled by modern technological advances, depletion of terrestrial and shallow-water resources, growing global population with rising demands for food, energy, and raw materials (Ramírez-Llodra et al., 2011; Norse et al., 2012; Hefferman, 2019), climate change (Levin and Le Bris, 2015; Sweetman et al., 2017), and ocean acidification (Solís-Marín et al., 2014). Deep-sea species and ecosystems are more vulnerable than its shallow-water counterparts, due to the life-history traits of most deep-sea species such as slow growth, delayed maturity, extended longevities, and slow colonization (Cheung et al., 2007). In addition, recovery of deep-sea habitats is slow, studies have found that extraction impacts (scars on the sea bottom) are still visible after 26 years (1989-2015), while some organisms found prior to the extraction have not returned (e.g., sponges, soft corals, or sea anemones) (Hefferman, 2019).
Costa Rica has prohibited oil, gas and mineral exploration and exploitation in the sea until 2050 through the government moratorium decree 36693-MINAET (MINAET, 2010). Nonetheless, the debate on whether Costa Rica should or should not exploit the deep-sea mineral resources has been increasing in media articles and social media recently. The only known fishing activity that exploited resources deeper than 200 m in Costa Rica was shrimp trawling. The target species were the kolibri shrimp (Solenocera agassizii), the northern nylon shrimp (Heterocarpus vicarius), and to a less extent the three-spined nylon shrimp (Heterocarpus affinis) (Wehrtmann and Nielsen-Muñoz, 2009). Data from scientific surveys indicated a decreasing deep-sea shrimp catch while bycatch was increasing (Wehrtmann and Nielsen-Muñoz, 2009; Wehrtmann et al., 2012). These surveys did not record any echinoderm (I. Wehrtmann per. comm.). The shrimp trawling is currently prohibited in Costa Rica (Sala Constitucional, 2013), however, there has been recent and continuous efforts to re-activate this activity. Finally, litter has also been observed in Costa Rican deep-sea waters around Isla del Coco, composed mainly of plastics and lost fishing gear (Naranjo-Elizondo and Cortés, 2018), and in much deeper areas in several locations (J. Cortés per. obs.).
Future Perspectives
The current knowledge on deep-sea echinoderms is limited and more research is needed. Publishing diversity records with taxonomic precision contributes to improve future efforts in research and management and contributes to assess future impacts on the marine ecosystems (Worm et al., 2006; Costello et al., 2013).
Deep-sea research is costly and requires highly specialized vessels, equipment, and trained scientists. These issues preclude research of deep-sea habitats in most developing and undeveloped countries. Companies will continue to be important allies in the exploration of the deep sea, including in Costa Rica (Brewin et al., 2007; Wehrtmann and Nielsen-Muñoz, 2009). Further collaborations with international institutions will be necessary to advance our knowledge on deep-sea echinoderms.
There is no doubt that the deep sea plays a unique and outstanding role on sustaining the health and functioning of the oceans (Sweetman et al., 2017). Deep-sea ecosystems provide habitat provision for commercial species (e.g., tuna, large bill fishes), nutrient cycling, heat absorption, trophic and diversity support services, and carbon sequestration, all of which are vital ecological processes to maintain diversity and humanity (Thurber et al., 2014). Van Dover (2011) suggest that having a coherent conservation, management, and mitigation framework for the deep sea is necessary before undergoing deep-sea resources exploitation. We call for further active action and advocacy for working towards science-based management conservation of the deep sea in Costa Rica, following the precautionary principles since the impacts on the deep sea could be irreversible at human timescale (Roberts, 2002; Waller et al., 2007; Hefferman, 2019).
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
The authors contribute in equal proportions to the elaboration of the work, both in the search for information, analysis, elaboration of figures, and writing. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank the University of Costa Rica for supporting basic research in all topics, and especially the Vicerrectoría de Investigación and the logistic support of the Center for Research in Marine Science and Limnology (CIMAR). We are grateful to the Undersea Hunter Group for their support and access to their submarine DeepSee, and all the videos. We greatly appreciate the invitation by Erik Cordes, Lisa Levin, Victoria Orphan, Greg Rouse, and Shana Goffredi, to participate in deep-sea expeditions off the Pacific coast of Costa Rica. We thank the crew and staff of the R/V Atlantis and the HOV Alvin, and the crew and staff of the R/V Falkor and the ROV SuBastian. We are thankful to Odalisca Breedy, Beatriz Naranjo and Charlotte Seid for their collaboration in deep-sea research. We appreciate the help with picture identifications by Rebeca Granja-Fernández, Francisco A. Solis-Marin, Tania Pineda-Enríquez, Carlos A. Conejeros-Vargas and Arturo Angulo.
References
Agassiz A. (1898). Reports on the Dredging Operations Off the West Coast of Central America to the Galápagos, to the West Coast of Mexico, and in the Gulf of California, in Charge of Alexander Agassiz, Carried on by the U.S. Fish Commission Steamer “Albatross”, Lieut. Commander Z.L. Tanner U.S.N., Commanding. XXIII. Preliminary Report on the Echini. Bull. Mus. Comp. Zoöl. Harvard Coll. 32, 71–86.
Agassiz A. (1904). Reports on an Exploration Off the West Coast of Mexico, Central and South America, and Off Galapagos Islands, in Charge of Alexander Agassiz, by the U.S. Fish Commission Steamer “Albatross”, During 1891, Lieut. Commander Z.L. Tanner, U.S. Commanding. XXXII. Panamic. deep sea. echini. Mem. Mus. Comp. Zoöl. Harvard Coll. 31, 1–246.
Alvarado J. J., Barraza E., Sancho-Mejía T. (2013). “Chapter 3: Central America Echinoderms: Diversity, Ecology and Future Perspectives,” in Echinoderm Research and Diversity in Latin America. Eds. Alvarado J. J., Solís-Marín F. A. (Berlin: Springer), 67–106. doi: 10.1007/s12526-009-0032-5
Alvarado J. J., Chacón-Monge J. L., Solís-Marín F. A., Pineda-Enríquez T., Caballero-Ochoa A. A., Solano-Rivera S., et al. (2017). Equinodermos Del Museo De Zoología De La Universidad De Costa Rica. Rev. Biol. Trop. 65 (Suppl. 1), S272–S287. doi: 10.15517/RBT.V65I1-1.31695
Alvarado J. J., Fabregat-Malé S. (2021). Echinoderm Research Perspectives: A Central American Bibliometric Review. UNED Res. Jour. 13, e3535. doi: 10.22458/urj.v13i2.3535
Alvarado-Induni G. E. (2021). Costa Rica Y Sus Volcanes (San José, Costa Rica: Universidad de Costa Rica), ISBN: 978-9968-46-776-6.
Auscavitch S. R. (2020). Deep-Sea Coral Biogeography and Community Structure in Tropical Seamount Environments. PhD Dissertation (Philadelphia, Pennsylvania, USA: Temple University).
Aziz A., Jangoux M. (1984). Description De Quatre Nouvelles Especes D’asterides Profonds (Echinodermata) De La Region Indo-Malaise. Indo-Malayan Zool. 2, 187–194.
Azofeifa-Solano J. C., Cortés J. (2021). “Chapter 24: Diversity and Biology of Deep-Water Crustaceans in Costa Rica,” in Deep-Sea Pycnogonids and Crustaceans of the Americas. Ed. Hendrickx M. E. (Switzerland: Springer Nature), 633–658. doi: 10.1007/978-3-030-58410-8_24
Benavides-Serrato M., Borrero-Pérez G. H. (2010). “Equinodermos De La Plataforma Y La Franja Superior Del Talud Continental Del Caribe Colombiano,” in Biodiversidad Del Margen Continental Del Caribe Colombiano, vol. 20 . Ed. Invemar, (INVEMAR: Serie de Publicaciones Especiales), 255–266.
Borrero-Pérez G. H., Benavides-Serrato M., Campos N. H., Galeano-Galeano E., Gavio B., Medina J., et al. (2019). Echinoderms of the Seaflower Biosphere Reserve: State of Knowledge and New Findings. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00188
Borrero-Pérez G. H., Benavides-Serrato M., Diaz-Sánchez C. M. (2012). Equinodermos Del Caribe Colombiano II: Echinoidea Y Holothuroidea. Ser. Publ. Esp. INVEMAR 30, 1–250.
Borrero-Pérez G. H., Benavides-Serrato M., Solano O. D., Navas G. R. (2003). Equinoideos (Echinodermata: Echinoidea) Colectados En La Franja Superior Del Talud Continental Del Caribe Colombiano. Bol. Invest. Cost. Mar. 30, 133–166. doi: 10.25268/bimc.invemar.2002.31.0.289
Borrero-Pérez G. H., Dueñas L. F., León J., Puentes V. (2020). Deep-Sea Holothurians (Echinodermata, Holothuroidea) From the Colombian Southern Caribbean Sea. Check. List 16, 535–551. doi: 10.15560/16.3.535
Brewin P. E., Stocks K. I., Menezes G. (2007). “A History of Seamount Research,” in Seamounts: Ecology, Conservation and Management. Eds. Pitcher T. J., Morato T., Hart P. J. B., Clark M. R., Haggan N., Santos R. S. (Oxford: Blackwell), 41–61. doi: 10.1002/9780470691953.ch3
Calero B., Ramil F., Ramos A. (2017). “Echinoderms of the Mauritanian Deep-Sea Waters,” in Deep-Sea Ecosystems Off Mauritania. Eds. Ramos A., Ramil F., Sanz J. L. (Dordrecht: Springer). doi: 10.1007/978-94-024-1023-5_12
Cambronero-Solano S., Benavides R., Solís-Marín F. A., Alvarado J. J. (2019). New Reports of Echinoderms on the Caribbean Continental Slope of Central America. Zoosymposia 15, 5–12. doi: 10.11646/zoosymposia.15.1.3
Campos L. S., Moura R. B., Veríssimo I., Curbelo-Fernandez M. P., Cavalcanti G. H., Brasil A. C. S. (2010). “ROV Imaging of Deep-Sea Echinoderms From the Brazilian Continental Margin, Southwest Atlantic,” in Echinoderms: Durham. Proc. Of the 12th Int. Echinoderm Conf. Eds. Harris L. G., Böttger S. A., Walker C. W., Lesser M. P. ( Balkema, Leiden), 147–152.
Catalán J., Merino-Yunnissi C., Martínez A., Sellanes J., Ibáñez C. M. (2020). Nuevos Registros De Crinoideos (Echinodermata: Crinoidea) En El Talud Continental De Chile. Rev. Biol. Mar. Oceanogr. 55, 68–72. doi: 10.22370/rbmo.2020.55.1.2395
Chacón-Monge J. L., Azofeifa-Solano J. C., Alvarado J. J., Cortés J. (2021). Área De Conservación Guanacaste Echinoderms, North Pacific of Costa Rica. Rev. Biol. Trop. 69 (Suppl. 1), S487–S500. doi: 10.15517/rbt.v69iSuppl.1.46391
Cheung W. W. L., Watson R., Morato T., Pitcher T. J., Pauly D. (2007). Intrinsic Vulnerability in the Global Fish Catch. Mar. Ecol. Prog. Ser. 333, 1–12. doi: 10.3354/meps333001
Clark H. L. (1917). Reports on the Scientific Results of the Expedition to the Tropical Pacific in Charge of Alexander Agassiz, on the U.S. Fish Commission Steamer “Albatross” From August 1899 to March 1900, Commander Jefferson F. Mooser, U.S.N Commanding. XVIII. Reports on the Scientific Results of the Expedition to the Tropical Pacific in Charge of Alexander Agassiz, on the U.S. Fish Commission Steamer “Albatross” From October 1904 to March 1905, Lieut. Commander L.M. Garret, U.S.N. Commanding. Bull. Mus. Comp. Zoöl. Harvard Coll. 61, 429–466.
Conejeros-Vargas C. A., Solís-Marín F. A., Laguarda-Figueras A. (2017). Equinoideos De Mar Profundo (Echinodermata: Echinoidea) Del Pacífico Mexicano. Rev. Biol. Trop. 65 (Suppl. 1), S244–S252. doi: 10.15517/RBT.V65I1-1.31692
Cortés J. (2008). Historia De La Investigación Marina De La Isla Del Coco, Costa Rica. Rev. Biol. Trop. 56 (Suppl 2), 1–18. doi: 10.15517/RBT.V56I2.26934
Cortés J. (2009). “A History of Marine Biodiversity Scientific Research in Costa Rica,” in Marine Biodiversity of Costa Rica, Central America. Eds. Wehrtmann I. S., Cortés J. (Dordrecht: Springer), 47–80. doi: 10.1007/978-1-4020-8278-8_2
Cortés J. (2016a). “The Pacific Coastal and Marine Ecosystems,” in Costa Rican Ecosystems. Ed. Kappelle M. (Chicago: University of Chicago Press), 97–138. doi: 10.7208/chicago/9780226121642.003.0005
Cortés J. (2016b). “Isla Del Coco: Coastal and Marine Ecosystems,” in Costa Rican Ecosystems. Ed. Kappelle M. (Chicago: University of Chicago Press), 162–191. doi: 10.7208/chicago/9780226121642.003.0007
Cortés J. (2016c). “The Caribbean Coastal and Marine Ecosystems,” in Costa Rican Ecosystems. Ed. Kappelle M. (Chicago: University of Chicago Press), 591–617. doi: 10.7208/chicago/9780226121642.003.0017
Cortés J. (2019). “Isla Del Coco, Costa Rica, Eastern Tropical Pacific,” in Mesophotic Coral Ecosystems. Coral Reefs of the World 12. Eds. Loya Y., Puglise K. A., Bridge T. C. L. (Switzerland: Springer Nature), 465–474. doi: 10.1007/978-3-319-92735-0_26
Cortés J., Blum S. (2008). Life to 450 M Depth at Isla Del Coco, Costa Rica. Rev. Biol. Trop. 56 (Suppl. 2), 189–206. doi: 10.15517/RBT.V56I2.27018
Costello M. J., Chaudhary C. (2017). Marine Biodiversity, Biogeography, Deep-Sea Gradients, and Conservation. Curr. Biol. 27, R511–R527. doi: 10.1016/j.cub.2017.04.060
Costello M. J., Michener W. K., Gahegan M., Zhang Z. Q., Bourne P. E. (2013). Biodiversity Data Should be Published, Cited, and Peer Reviewed. Trends Ecol. Evol. 28, 454–461. doi: 10.1016/j.tree.2013.05.002
Costello M. J., Tsai P., Wong P. S., Cheung A. K. L., Basher Z., Chaudhary C. (2017). Marine Biogeograohic Realms and Species Endemicity. Nat. Commun. 8, 1057. doi: 10.1038/s41467-017-01121-2
Dueñas L. F., Puentes V., León J., Herrera S. (2021). Fauna Associated With Cold Seeps in the Deep Colombian Caribbean. Deep–Sea Res. I 173, 103552. doi: 10.1016/j.dsr.2021.103552
Fisher W. K. (1928). Sea Stars From the Arcturus Oceanographic Expedition. Zool. 8, 487–493. doi: 10.5962/p.203759
Flores J. N., Brogger M. I., Penchaszadeh P. E. (2019). Reproduction and Development of the Brooding Sea Urchin Austrocidaris Canaliculata From Deep-Sea Off Argentina. Deep. Sea. Res. I: Oceanogr. Res. Pap. 143, 35–42. doi: 10.1016/j.dsr.2018.11.012
Flores J. N., Penchaszadeh P. E., Brogger M. I. (2021). Heart Urchins From the Depths: Corparva Lyrida Gen. Et Sp. Nov. (Palaeotropidae), and New Records for the Southwestern Atlantic Ocean. Rev. Biol. Trop. 69 (Suppl. 1), S14–S34. doi: 10.15517/rbt.v69iSuppl.1.46320
Fujita T., Stampanato S., Jangoux M. (1994). Belyaevostella Hyugaensis, a New Species of Deep-Sea Asteroid (Asteroidea, Caymanostellidae) Found on a Sunken Wood From Off Southern Japan. Bull. Nat. Sci. Mus. Tokyo A 20, 183–188.
González G., Dunia N., Solano P., Oscar D., Navas S. (2002). Equinodermos Colectados Por La Expedición CIOH-INVEMAR-Smithsonian Desde Cartagena Hasta El Golfo De Urabá, Caribe Colombiano. Bol. Invest. Mar. Cost. 31, 85–132.
Hartlaub C. (1895). Reports on the Dredging Operations Off the West Coasts of Central America to the Galapagos, to the West Coast of Mexico, and in the Gulf of California, in Charge of Alexander Agassiz, Carried on by the U.S. Fish Commission Steamer “Albatross”, During 1891, Lieut. Commander Z.L. Tanner, U.S.N., Commanding. XVIII. Die Comatuliden. Bull. Mus. Comp. Zoöl. Harvard Coll. 27 (4), 127–152.
Hendrickx M. E., Pawson D., Mejía-Mercado B. E., Ayón-Parente M., Hinojosa-Corona A. (2014). “Registros De Especies De Holothuroidea (Echinodermata),” in Explorando El Mar Profundo Del Golfo De California 2008-2014. Eds. Mejía-Mercado B. E., Hinojosa-Corona A., Hendrickx M. E. ( CICESE), 177–191.
Levin L. A., Le Bris N. (2015). The Deep Ocean Under Climate Change. Science 350, 766–768. doi: 10.1126/science.aad0126
Levin L. A., Mendoza G. F., Grupe B. M., Gonzalez J. P., Jellison B., Rouse G. W., et al. (2015). Biodiversity on the Rocks: Macrofauna Inhabiting Authigenic Carbonate at Costa Rica Methane Seeps. PLoS One 10, e0136129. doi: 10.1371/journal.pone.0131080
Levin L. A., Orphan V. J., Rouse G. W., Rathburn A. E., Ussler W., Cook G. S., et al. (2012). A Hydrothermal Seep on the Costa Rica Margin: Middle Ground in a Continuum of Reducing Ecosystems. Proc. R. Soc B 279, 2580–2588. doi: 10.1098/rspb.2012.0205
Ludwig H. (1894). Reports on an Exploration Off the West Coast of Mexico, Central America and South America, and Off the Galápagos Islands, in Charge of Alexander Agassiz, by the U.S. Fish Commission Steamer “Albatross”, During 1891. XII. The Holothurioidea. Mem. Mus. Comp. Zoöl., Harvard Coll.17 (3), 1–183.
Ludwig H. (1905). Reports on the Exploration Off the West Coasts of Mexico, Central and South America, and Off the Galapagos Islands, in Charge of Alexander Agassiz by the U.S. Fish Commission Steamer “Albatross,” During 1891, Lieut.-Commander Z.L. Tanner, U.S.N., Commanding. V. Reports on the Scientific Results of the Expedition to the Tropical Pacific in Charge of Alexander Agassiz, by the U.S. Fish Commission Steamer “Albatross”, From August 1899, to March 1900, Commander Jefferson F. Moser, U.S. N., Commanding. VII. Asteroidea. Mem. Mus. Comp. Zoöl. Harvard Coll. 32, 1–291.
Luke R. (1982). Catalog of the Benthic Invertebrate Collections of the Scripps Institution of Oceanography. Echinodermata. Scripps Institution Oceanogr. Ref. Ser., 82–85.
Luna-Cruz A. K., Hendrickx M. E. (2018). New Records of Synallactes Virgulasolida Massin & Hendrickx 2010 (Echinodermata: Holothuroidea) From the Eastern Pacific. Zootaxa 4444, 491–500. doi: 10.11646/zootaxa.4444.4.9
Luna-Cruz A. K., Hendrickx M. E. (2020). Rediscovery of the Deep-Water Species Ypsilocucumis Californiae Massin & Hendrickx 2011 (Echinodermata; Holothuroidea; Ypsilothuriidae) in Western Mexico. Zootaxa 4742, 139–148. doi: 10.11646/zootaxa.4742.1.9
Luna-Cruz A. K., Hendrickx M. E. (2021). Deep-Water Holothuroids Echinodermata, Holothuroidea) Collected Off the West Coast of Baja California Península, Mexico During the TALUD Cruises. Geomare. Zool. 3, 31–40.
Lütken C. F., Mortensen T. (1899). Reports on an Exploration Off the West Coast of Mexico, Central America and South America, and Off theGalápagos Islands, in Charge of Alexander Agassiz byU.S Fish Commission Steamer “Albatross”, During 1891, Lieut.-Commander Z.L. Tanner, U.S.N., Commanding. XXV. Ophiuridae. Mem. Mus. Comp. Zoöl. Harvard Coll. 23 (2), 93–208.
Mah C. L., Blake D. B. (2012). Global Diversity and Phylogeny of the Asteroidea (Echinodermata). PLoS One 7, e35644. doi: 10.1371/journal.pone.0035644
Manso C. L. C. (2010). Deep-Water Ophiuroidea (Echinodermata) From Off Chile in the Eastern South Pacific. Biota. Neotrop. 10 (2), 185–199. doi: 10.1590/S1676-06032010000200023
Martínez M. I. (2016). The Sea Cucumber Psolus Patagonicus Ekman 1925 (Echinodermata: Holothuroidea) From the Southwestern Atlantic, Redescription of the Holotype and New Synonym. Rev. Mus. Arg. Cien. Nat. 18, 237–241. doi: 10.22179/REVMACN.18.474
Martinez M. I., Alba−Posse E. J., Lauretta D., Penchaszadeh P. E. (2020). Reproductive Features in the Sea Cucumber Pentactella Perrieri (Ekman 1927) (Holothuroidea: Cucumariidae): A Brooding Hermaphrodite Species From the Southwestern Atlantic Ocean. Pol. Biol. 43, 1383–1389. doi: 10.1007/s00300-020-02715-1
Martínez M. I., Arribas L. P., Berecoechea J. J., Brogger M. I., Penchaszadeh P. E. (2015). Echinoderm Diversity in the Southwestern Atlantic. Rev. Biol. Trop. 63 (Suppl. 2), 115–120. doi: 10.15517/RBT.V63I2.23132
Martinez M. I., Penchaszadeh P. E. (2017). A New Species of Brooding Psolidae (Echinodermata: Holothuroidea) From Deep-Sea Off Argentina, Southwestern Atlantic Ocean. Deep-Sea Res. Part II 146, 13–17. doi: 10.1016/j.dsr2.2017.05.007
Martínez M. I., Solís-Marín F. A., Penchaszadeh P. E. (2014). Benthodytes Violeta, a New Species of a Deep-Sea Holothuroid (Elasipodida: Psychropotidae) From Mar Del Plata Canyon (South-Western Atlantic Ocean). Zootaxa 3760, 89–95. doi: 10.11646/zootaxa.3760.1.6
Martínez M. I., Solís-Marín F. A., Penchaszadeh P. E. (2019). First Report of Paelopatides (Synallactida, Synallactidae) for the SW Atlantic, With Description of a New Species From the Deep-Sea Off Argentina. Zool. Anzeiger 278, 21e27. doi: 10.1016/j.jcz.2018.10.010
Massin C., Hendrickx M. E. (2011). Deep-Water Holothuroidea (Echinodermata) Collected During the TALUD Cruises Off the Pacific Coast of Mexico, With the Description of Two New Species. Rev. Mex. Biodiv. 82, 413–443. doi: 10.22201/ib.20078706e.2011.2.476
Mecho A., Billett D. S. M., Ramírez-Llodra E., Aguzzi J., Tyler P. A., Company J. B. (2014). First Records, Rediscovery and Compilation of Deep-Sea Echinoderms in the Middle and Lower Continental Slope of the Mediterranean Sea. Sci. Mar. 78, 281–302. doi: 10.3989/scimar.03983.30C
Miller J. E., Pawson D. L. (1989). Hansenothuria Benti, New Genus, New Species (Echinodermata: Holothuroidea) From the Tropical Western Atlantic: A Bathyal, Epibenthic Holothurian With Swimming Abilities. Proc. Biol. Soc Wash. 102, 977–986.
MINAET (2010). “Se Decreta Moratoria En Minería,” in Decreto N°35982-MINAET. Gaceta N° 90(San José, Costa Rica). Available at: http://www.pgrweb.go.cr/scij/Busqueda/Normativa/Normas/nrm_texto_completo.aspx?param1=NRTC&nValor1=1&nValor2=67911&nValor3=80690&strTipM=TC#:~:text=Art%C3%ADculo%201%C2%BA%2D%20Se%20declara%20la,extra%C3%ADdos%20utilizando%20cianuro%20o%20mercurio
Mironov A. N., Minina K. V., Dilmana A. B., Smirnovb I. S. (2018). Deep-Sea Echinoderms of the Sea of Okhotsk. Deep-Sea Res. Part II 154, 342–357. doi: 10.1016/j.dsr2.2017.10.003
Moles J., Figuerola B., Campanya-Llovet N., Monleón-Getino T., Taboada S., Avila C. (2015). Distribution Patterns in Antarctic and Subantarctic Echinoderms. Pol. Biol. 38, 799–813. doi: 10.1007/s00300-014-1640-5
Naranjo-Elizondo B., Cortés J. (2018). Observations of Litter Deposited in the Deep Waters of Isla Del Coco National Park, Eastern Tropical Pacific. Front. Mar. Sci. 5. doi: 10.3389/fmars.2018.00091
Neuhaus B. (2004). Description of Campyloderes Cf. Vanhoeffeni (Kinorhyncha, Cyclorhagida) From the Central American East Pacific Deep With a Review of the Genus. Meiofauna Mar. 13, 3–20.
Norse E. A., Brooke S., Cheung W. W. L., Clark M. R., Ekeland I., Froese R., et al. (2012). Sustainability of Deep-Sea Fisheries. Mar. Pol. 36, 307–320. doi: 10.1016/j.marpol.2011.06.008
O’Loughlin P. M., Mackenzie M., VandenSpiegel D. (2013). New Sea Cucumber Species From the Seamounts on the Southwest Indian Ocean Ridge (Echinodermata: Holothuroidea: Aspidochirotida, Elasipodida, Dendrochirotida). Mem. Mus. Victoria 70, 37–50. doi: 10.24199/j.mmv.2013.70.04
Pawson D. L. (1982). Deep-Sea Echinoderms in the Tongue of the Ocean, Bahama Islands: A Survey, Using the Research Submersible Alvin. Aust. Mus. Mem. 16, 129–145. doi: 10.3853/j.0067-1967.16.1982.362
Pawson D. L. (1983). Psychronaetes Hanseni, a New Genus and Species of Elasipodan Sea Cucumber From the Eastern Central Pacific (Echinodermata: Holothurioidea). Proc. Biol. Soc Wash. 96, 154–159.
Pawson D. L., Nizinski M., Lewis C., Pawson D. (2015). Deep-Sea Echinoids and Holothurians (Echinodermata) Near Cold Seeps and Coral Communities in the Northern Gulf of Mexico. Bull. Mar. Sci. 91, 167–204. doi: 10.5343/bms.2014.1064
Pérez-Ruzafa A., Alvarado J. J., Solís-Marín F. A., Hernández J. C., Morata A., Marcos C., et al. (2013). “Appendix,” in Echinoderm Research and Diversity in Latin America. Eds. Alvarado J. J., Solís-Marín F. A. (Berlin: Springer), 543–654.
Pertossi R. M., Brogger M. I., Penchaszadeh P. E., Martinez M. I. (2019). Reproduction and Developmental Stages in the Crinoid Isometra Vivipara Mortensen 1917 From the Southwestern Atlantic. Pol. Biol. 42, 807–816. doi: 10.1007/s00300-019-02477-5
Ramírez-Llodra E. R., Tyler P. A., Baker M. C., Bergstad O. A., Clark M. R., Escobar E., et al. (2011). Man and the Last Great Wilderness: Human Impact on the Deep Sea. PLoS One 6, e22588. doi: 10.1371/journal.pone.0022588
Rivadeneira P. R., Brogger M. I., Penchaszadeh P. E. (2017). Aboral Brooding in the Deep Water Sea Star Ctenodiscus Australis Lütken 1871 (Asteroidea) From the Southwestern Atlantic. Deep-Sea Res. Part I: Oceanogr. Res. Pap. 123, 105–109. doi: 10.1016/j.dsr.2017.03.011
Rivadeneira P. R., Martinez M. I., Penchaszadeh P. E., Brogger M. I. (2020). Reproduction and Description of a New Genus and Species of Deep-Sea Asteriid Sea Star (Echinodermata; Asteroidea) From the Southwestern Atlantic. Deep. Sea. Res. Part I: Oceanogr. Res. Pap. 163,0967–0637. doi: 10.1016/j.dsr.2020.103348
Roberts C. M. (2002). Deep Impact: The Rising Toll of Fishing in the Deep Sea. Trends Ecol. Evol. 17, 242–245. doi: 10.1016/S0169-5347(02)02492-8
Rojas-Jiménez K., Grossart H. P., Cordes E., Cortés J. (2020). Fungal Communities in Sediments Along a Depth Gradient in the Eastern Tropical Pacific. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.575207
Roux M. (2004). New Hyocrinid Crinoids (Echinodermata) From Submersible Investigations in the Pacific Ocean. Pac. Sci. 58, 597–613. doi: 10.1353/psc.2004.0042
Sahling H., Masson D. G., Ranero C. R., Hühnerbach V., Weinrebe W., Klaucke I., et al. (2008). Fluid Seepage at the Continental Margin Offshore Costa Rica and Southern Nicaragua. Geochem. Geophys. Geosyst. 9 (5), 1–22. doi: 10.1029/2008GC001978
Sala Constitucional (2013). Resolución N° 10540-2013 Del 7 De Agosto De 2013 (San José, Costa Rica). Available at: http://www.pgrweb.go.cr/scij/Busqueda/Normativa/Normas/nrm_articulo.aspx?param1=NRA&nValor1=1&nValor2=86448&nValor3=112177&nValor5=2
Selig G. M., Netburn A. N., Malik M. (2019). Distributions of the Pelagic Holothurian Pelagothuria in the Central Pacific Ocean as Observed by Remotely-Operated Vehicle Surveys. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00684
Setyastuti A., Wirawati I. (2018). The Banda Sea: A Hotspot of Deep-Sea Echinoderms Diversity in Indonesia. IOP Conf. Series: Earth Environ. Sci. 184, 12003. doi: 10.1088/1755-1315/184/1/012003
Solís-Marín F. A., Alvarado J. J., Abreu-Pérez M., Aguilera O., Alió J., Bacallado-Aránega J. J., et al. (2013). “Appendix,” in Echinoderm Research and Diversity in Latin America. Eds. Alvarado J. J., Solís-Marín F. A. (Berlin: Springer), 543–654.
Solís-Marín F. A., Laguarda-Figueras A., Durán-González A., Vázquez-Bader A. R., Gracia-Gasca A. (2014). “Biodiversidad De Los Equinodermos (Echinodermata) Del Mar Profundo Mexicano,” in La Frontera Final: El Océano Profundo. Eds. Low-Pfeng A., Peters E. (México: INECC), 207–246.
Stöhr S., O’Hara T. D. (2021). Deep-Sea Ophiuroidea (Echinodermata) From the Danish Galathea II Expedition 1950–52, With Taxonomic Revisions. Zootaxa 4963, 505–529. doi: 10.11646/zootaxa.4963.3.6
Stöhr S., O’Hara T. D., Thuy B. (2012). Global Diversity of Brittle Stars (Echinodermata: Ophiuroidea). PLoS One 7, e31940. doi: 10.1371/journal.pone.0031940
Stöhr S., O’Hara T., Thuy B. (2022) World Ophiuroidea Database. Available at: https://www.marinespecies.org/aphia.php?p=taxdetails&id=123598on2022-04-08.
Stöhr S., Segonzac M. (2005). Deep-Sea Ophiuroids (Echinodermata) From Reducing and non-Reducing Environments in the North Atlantic Ocean. J. Mar. Biol. Assoc. U.K. 85, 383–402. doi: 10.1017/S0025315405011318h
Summers M. M., Al-Hakim I. I., Rouse G. W. (2014). Turbo-Taxonomy: 21 New Species of Myzostomida (Annelida). Zootaxa 3873, 301–344. doi: 10.11646/zootaxa.3873.4.1
Sweetman A. K., Thurber A. R., Smith C. R., Levin L. A., Mora C., Wei C. L., et al. (2017). Major Impacts of Climate Change on Deep-Sea Benthic Ecosystems. Elementa Sci. Anthrop. 5, 4. doi: 10.1525/elementa.203
Thurber A. R., Sweetman A. K., Narayanaswamy B. E., Jones D. O. B., Ingels J., Hansman R. L. (2014). Ecosystem Functions and Services in the Deep Sea. Biogeosciences 11, 3941–3963. doi: 10.5194/bg-11-3941-2014
Townsend C. H. (1901). Dredging and Other Records of the United States Fish Commission Steamer “Albatross”, With Bibliography Relative to the Work of the Vessel. U.S. Fish. Commun. Rep. 1900, 387–562.
UNESCO (2009). “Global Open Oceans and Deep Seabed (GOODS) – Biogeographic Classification,” in IOC Technical Series, vol. 84. (Paris: UNESCO-IOC).
Van Dover C. (2011). Tighten Regulations on Deep-Sea Mining. Nature 470, 31–33. doi: 10.1038/470031a
Voss G. L. (1971). Narrative of RV John Elliot Pillsbury Cruise P-7101: Central America, January 20-February 5, 1971 (Miami, Florida: School of Marine and Atmospheric Science, University of Miami).
Waller R., Watling L., Auster P., Shank T. (2007). Anthropogenic Impacts on the Corner Rise Seamounts, North-West Atlantic Ocean. J. Mar. Biol. Assoc. UK 87, 1075–1107. doi: 10.1017/S0025315407057785
Wehrtmann I. S., Arana P. M., Barriga E., Gracia A., Pezzuto P. R. (2012). Deep-Water Shrimp Fisheries in Latin America: A Review. Lat. Amer. J. Aquat. Res. 40, 497–535. doi: 10.3856/vol40-issue3-fulltext-2
Wehrtmann I. S., Nielsen-Muñoz V. (2009). The Deepwater Fishery Along the Pacific Coast of Costa Rica, Central America. Lat. Amer. J. Aquat. Res. 37, 543–554. doi: 10.3856/vol37-issue3-fulltext-19
Wheat C. G., Hartwell A. M., McManus J., Fisher A. T., Orcutt B. N., Schlicht L. E. M., et al. (2019). Geology and Fluid Discharge at Dorado Outcrop, a Low Temperature Ridge-Flank Hydrothermal System. Geochem. Geophys. Geosyst. 20, 487–504. doi: 10.1029/2018GC007933
Worm B., Barbier E. B., Beaumont N., Duffy J. E., Folke C., Halpern B. S., et al. (2006). Impacts of Biodiversity Loss on Ocean Ecosystem Services. Science 314, 787–790. doi: 10.1126/science.1132294
WoRMS Editorial Board (2022) World Register of Marine Species. Available at: https://www.marinespecies.org.
Keywords: Central America, Eastern Tropical Pacific, Caribbean, last frontier, new records, Ophiroidea, Holothuroidea
Citation: Alvarado JJ, Chacón-Monge JL, Azofeifa-Solano JC and Cortés J (2022) Diversity of Deep-Sea Echinoderms From Costa Rica. Front. Mar. Sci. 9:918878. doi: 10.3389/fmars.2022.918878
Received: 12 April 2022; Accepted: 13 June 2022;
Published: 15 July 2022.
Edited by:
Paris Vasileios Stefanoudis, University of Oxford, United KingdomReviewed by:
Edgardo Londono-Cruz, University of Valle, ColombiaIlana Rosental Zalmon, State University of the North Fluminense Darcy Ribeiro, Brazil
Copyright © 2022 Alvarado, Chacón-Monge, Azofeifa-Solano and Cortés. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juan José Alvarado, anVhbmFsdmE3NkB5YWhvby5jb20=
†ORCID: Juan José Alvarado, orcid.org/0000-0002-2620-9115
José Leonardo Chacón-Monge, orcid.org/0000-0002-9754-1254
Juan Carlos Azofeifa-Solano, orcid.org/0000-0002-2105-1056
Jorge Cortés, orcid.org/0000-0001-7004-8649