 Ye Yuan1,2,3,4
Ye Yuan1,2,3,4 Min Jin1,5*
Min Jin1,5* Fang Fang1,5
Fang Fang1,5 Douglas R. Tocher2Mónica B. Betancor6
Douglas R. Tocher2Mónica B. Betancor6 Lefei Jiao1,5
Lefei Jiao1,5 Yucong Hong7
Yucong Hong7 Qicun Zhou1,5*
Qicun Zhou1,5*- 1Laboratory of Fish and Shellfish Nutrition, School of Marine Sciences, Ningbo University, Ningbo, China
- 2Guangdong Provincial Key Laboratory of Marine Biotechnology, Institute of Marine Sciences, Shantou University, Shantou, China
- 3Guangdong Provincial Key Laboratory of Marine Disaster Prediction and Prevention, Shantou University, Shantou, China
- 4STU-UMT Joint Shellfish Research Laboratory, Shantou University, Shantou, China
- 5Key Laboratory of Aquacultral Biotechnology Ministry of Education, Ningbo University, Ningbo, China
- 6Institute of Aquaculture, Faculty of Natural Sciences, University of Stirling, Stirling, United Kingdom
- 7Guangdong Yuequn Biotechnology Co., Ltd, Guangdong Provincial Key Laboratory of Aquatic Larvae Feed, Jieyang, China
Organisms can regulate mitochondrial metabolic adaptation, further ameliorate the energy homeostasis to control the ATP production for the energy expenditure during the growth process under different physiological and nutritional conditions. Based on this understanding, the objective of the present study was to investigate how different dietary n-3 PUFA (polyunsaturated fatty acid) lipid sources modify the mitochondrial metabolic adaptation, and further affect the energy homeostasis and the growth of swimming crab (Portunus trituberculatus). A total of 120 swimming crab juveniles with an average initial weight of 10.17 ± 0.12 g were fed one of three diets (4 replicates/treatment) containing either fish oil (control), krill oil or linseed oil as dietary lipid sources for 8 weeks, and the effects of dietary lipid sources on the growth and energy homeostasis via the regulation of mitochondrial metabolic adaptation were evaluated. The study revealed that, compared with linseed oil rich in 18:3n-3, fish oil and krill oil rich in 20:5n-3 and 22:6n-3 significantly promoted the molting and growth of juvenile swimming crab, increased the ATP level, mitochondrial membrane potential, NAD+ substrate level, NAD+/NADH ratio and the mitochondrial DNA copy number. Furthermore, crabs fed the diet supplemented with krill oil can up-regulate the expression levels of genes related to energy metabolism. In addition, dietary krill oil also specifically improved the ability for scavenging free radicals produced in the process of physiological metabolism, reduced the level of lipid peroxidation and the degree of DNA oxidative damage, and improved the health status of swimming crab. The present study revealed the adaptation of mitochondrial metabolism and the regulation of the energy homeostasis of swimming crab to different dietary n-3 PUFA lipid sources, and provided a new insight into the relationship between the growth as well as molting and the energy homeostasis, which provided a novel insight into the lipid nutrition and energy metabolism of crustacean species.
Introduction
Molting, a critical biological process in crustaceans that requires a large amount of energy to absorb water and swell the exoskeleton, is highly related to growth, gonad development and reproduction (Panganiban et al., 1995; Jung et al., 2013; Huang et al., 2015). Hepatopancreas is the key organ responsible for not only the metabolism of nutrients (particularly lipids and fatty acids) in crustaceans, but also the production of energy for the process of molting which maintain energy homeostasis and survival after molting (Wang et al., 2014; Huang et al., 2015; Fang et al., 2021). Hence, the hepatopancreas is not only the critical organ in which to study the metabolic response to exogenous nutrients in crustaceans, but also the bridge between the molting process and energy homeostasis.
Endogenous factors such as physiological status, growth stage and nutritional condition, as well as exogenous factors such as temperature, toxins, exercise and food availability, can alter the balance of energy expenditure and supply in organisms (Bremer and Moyes, 2011; Dos Santos et al., 2012; Sokolova et al., 2012; Conde-Sieira and Soengas, 2017). Organisms can regulate mitochondrial metabolic adaptation and affect the processes of energy metabolism to control ATP (adenosine triphosphate) production under different physiological and environmental conditions, which further alters the balance between energy expenditure and energy supply (Bremer and Moyes, 2011). It was reported that mitochondrial metabolic adaptation was affected by many factors including the fatty acid profile of membranes (Kraffe et al., 2007; Yuan et al., 2019) and membrane potential (Zorova et al., 2018), volume density (Urschel and O'Brien, 2008; Morris et al., 2021), ETC (electron transport chain) complex activity (Duggan et al., 2011; Cogliati et al., 2018) and NAD+ (nicotinamide adenine dinucleotide) homeostasis (Cantó et al., 2015; Jokinen et al., 2017), mtDNA (mitochondrial DNA) copy number (Hartmann et al., 2011; Liao et al., 2015), mitochondrial transcription factors (Bremer et al., 2012; Yuan et al., 2019) and sirtuins (Lombard et al., 2011; Rahman et al., 2014). As the metabolic adaptability of mitochondria changes (especially inefficient mitochondrial metabolism), ROS (reactive oxygen species) are produced accordingly, and overproduction of ROS can lead to a series of metabolic diseases including diabetes mellitus, insulin resistance and obesity (Alfadda and Sallam, 2012). In Arthropoda such as crustacean species, molting can strongly influence the energy homeostasis owing to the recovery process requiring additional energy. Furthermore, mitochondria produce ROS in response to the production of energy, which may cause a series of changes in the nucleus at the transcriptional level (Yun and Finkel, 2014). Many studies have focused on the relationship between mitochondrial metabolic adaptation and exogenous factors including temperature, toxins, photoperiod and exercise in organisms (LeMoine et al., 2010; Bremer and Moyes, 2011; Sokolova et al., 2012), but few studies have investigated the effect and mechanism of dietary stimuli (especially dietary fatty acids) on the mitochondrial metabolic adaptation and energy homeostasis in marine crustaceans.
Swimming crab (Portunus trituberculatus) is widely distributed in the coastal waters of China, Japan, Korea and Malaysia, and is a well-studied marine crustacean in China due to its high nutritional quality and economic value (Yuan et al., 2020). In the past decade, research on lipid and fatty acid nutrition as well as the metabolism of swimming crab has advanced greatly (Yuan et al., 2019; Yuan et al., 2019; Yuan et al., 2020; Fang et al., 2021; Yuan et al., 2021; Xie et al., 2022). However, until now, mitochondrial metabolic adaptability has not been studied in crustaceans with few studies designed to specifically investigate the mechanisms of how dietary lipid sources and fatty acids impact energy homeostasis. In the present study, we hypothesized that different dietary n-3 PUFA (polyunsaturated fatty acid) lipid sources would differentially affect mitochondrial metabolic adaptation of hepatopancreas of swimming crab, and further influence the energy homeostasis after molting, and finally alter the growth performance of swimming crab. The overarching aim of the research was to provide reliable evidence and novel insights into the mechanisms of mitochondrial metabolic responses and energy homeostasis of crustaceans in response to different dietary lipid sources.
Materials and Methods
Experimental Design and Diet Preparation
Three isoproteic and isolipidic experimental diets varying in lipid source (fish oil, krill oil and linseed oil) were formulated and manufactured to contain approximately 45 g kg-1 crude protein and 9 g kg-1 crude lipid (Table 1). The three diets were: 1) Diet FO with fish oil as a source of n-3 LC-PUFA (long-chain polyunsaturated fatty acids) (the control diet), 2) Diet KO with krill oil as a source of n-3 LC-PUFA, and 3) Diet LO with linseed oil as a source of 18:3n-3. The lipid sources were added at 45 g kg-1 (FO), 44.3 g kg-1 (KO) and 26.4 g kg-1 (LO), respectively. And the addition of palmitic acid were 0 g kg-1, 0.7 g kg-1 and 18.6 g kg-1, respectively, to equalize the total n-3 PUFA and lipid contents in three diets. The fatty acid compositions of lipid sources and diets are shown in Table S1 and Table S2, respectively. The contents of n-3 LC-PUFA (EPA and DHA) in the FO and KO diets were 9.13 and 9.23 mg g-1 (dry matter), respectively, and the content of 18:3n-3 in the LO diet was 9.22 mg g-1 (Table S2). The diets were prepared following the procedure described in detail previously (Yuan et al., 2019). Briefly, all ingredients were firstly ground into fine powder by ultra-fine pulverizer, then passed through a screen cloth (178 μm). The ingredients were thoroughly mixed with lipid sources and water in a Hobart type mixer (F-26, Machine factory of South China University of Technology, Guangzhou, China), and the mixture was processed into pellets in two specifications (3.0 mm in diameter and 10.0 mm in length; 5.0 mm in diameter and 15.0 mm in length) using a twin screw extruder (G-250, Machine factory of South China University of Technology, Guangzhou, China), then pellets were steamed for 30 min at 90°C in a constant temperature oven. After air-drying to approximately 10% moisture, the feeding stuff were sealed in vacuum-packed bags and stored at −20°C until used. Proximate compositions of experimental diets were determined following the methods of the Association of Official Analytical Chemists (AOAC, 2006). Briefly, moisture was determined by drying the samples to a constant weight at 105°C. Crude protein content was measured by using the Dumas combustion method directly with a protein analyzer (FP-528, Leco, USA). Crude lipid content was determined via petroleum ether extraction using the Soxtec method (Soxtec System HT6, Tecator, Sweden). Ash content was determined using a muffle furnace at 550°C for 8 h.
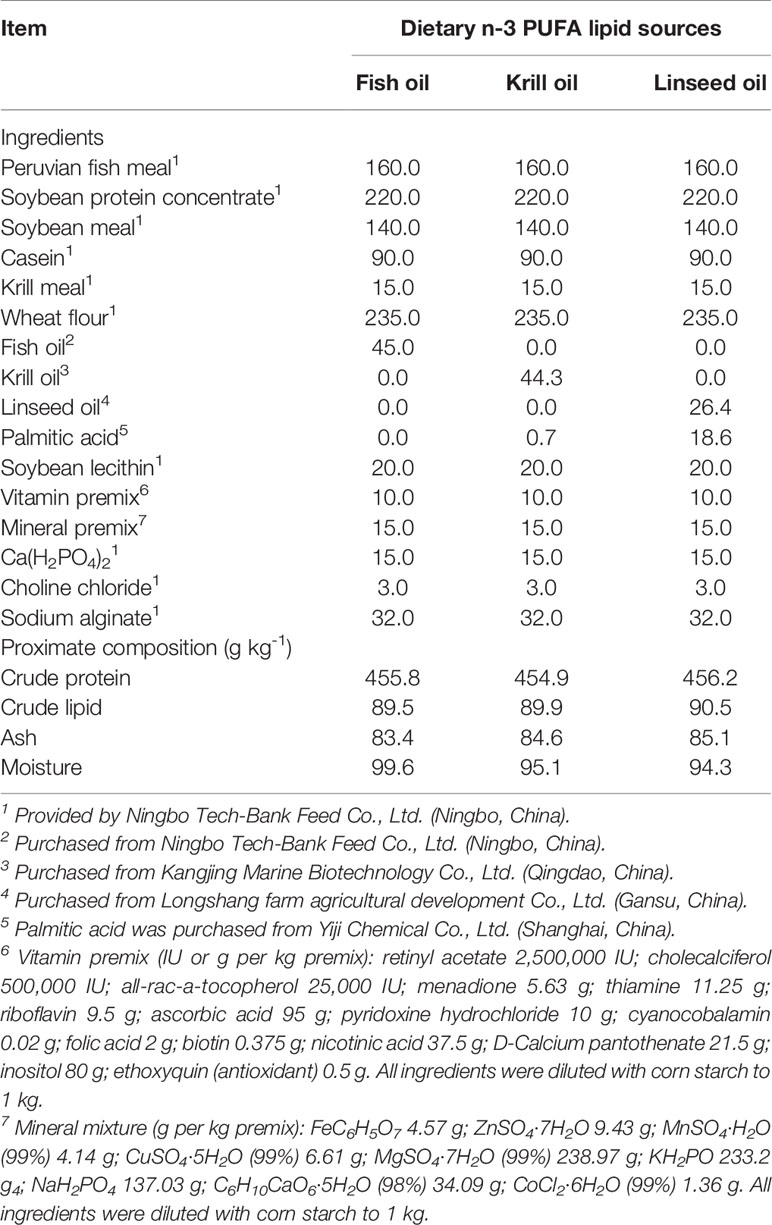
Table 1 Formulation and analyzed proximate composition of experimental diets (g kg-1, DM basis).
Crab Rearing and Experimental Conditions
Swimming crab juveniles were sourced from a commercial crab farm in Xiangshan, Ningbo, China. Prior to the start of the feeding trial, the crabs were maintained in an indoor rectangular cement pool (8.5 m × 3.0 m × 1.5 m) for acclimation over 2 weeks during which time they were fed a commercial feed containing 45 g kg-1 crude protein and 8 g kg-1 crude lipid (Ningbo Tech-Bank Feed Co. Ltd., Ningbo, China). The feeding trial was conducted at the Ningbo Marine and Fishery Science and Technology Innovation Base (Ningbo, China), and the procedures for crab culture and daily management were conducted according to the methods described in detail previously (Yuan et al., 2020). Briefly, 120 uniformly sized and healthy swimming crab juveniles (mean weight ± SE: 10.17 ± 0.12 g) were weighed and randomly allocated into 120 individual plastic baskets (35 cm × 30 cm × 35 cm) in a new cement pool (6.8 m × 3.8 m × 1.7 m). Each treatment had four replicates, each consisting of 10 crabs (10 plastic baskets), which placing in a line next to each other in the cement pool. All of crabs were hand-fed once per day at 17:00 with a ration of 6–8% of wet body weight. Each crab was weighed every 2 weeks and the daily ration of the feed was adjusted accordingly. The uneaten feed and feces were removed with a nylon screen, and 60% of seawater in the cement pool was exchanged daily to maintain water quality. During the feeding trial, the seawater temperature ranged from 28.5 to 32.1°C; the salinity was 24.5–28.1 g L-1; the dissolved oxygen concentration was not less than 6.0 mg L-1; and NH4-N concentration was lower than 0.05 mg L-1.
Sample Collection
At the termination of the feeding trial, crab in each plastic basket was weighed and counted to calculate the survival, percent weight gain (PWG), molting ratio (MR), specific growth rate (SGR) and feed efficiency (FE). Before sample collection, all crabs were fasted for 12 h. The crabs were placed on ice and anesthetized, and hepatopancreas samples were rapidly dissected out. A small portion of the hepatopancreas (approximately 20 mg each crab) from 3 crabs per biological replicate of each treatment were pooled into 1.5-mL RNAase-free tubes (Axygen, USA) as one sample, immediately frozen in liquid nitrogen, and stored at −80°C prior to analysis of gene expression (n = 4 per treatment). The remainder of the hepatopancreas (approximately 1.5 g each crab) from another three crabs (1 g per crab) per biological replicate of each group were dissected, collected and mixed as one sample (n = 4 per dietary treatment) in a 5-mL microfuge tube, then stored immediately at −80°C for further analysis. All the determination and tests were repeated twice, and the average of the two measured values were taken as the final result.
Quantification of Fatty Acids
The fatty acid profiles of lipid sources and diets were determined according to our previous method (Yuan et al., 2020). Briefly, the samples were first treated by methyl esterification, then the fatty acid methyl esters (FAMEs) were resuspended in 500 μL n-hexane, finally separated and measured on a gas chromatograph mass spectrometer (GC-MS, Agilent 7890B-5977A, Agilent Technologies, CA, USA) fitted with a fused-silica ultra-inert capillary column (DB-WAX, 30 m × 250 μm internal diameter, film thickness 0.25 μm, Agilent J & W Scientific, CA, USA), with the following temperature program and column conditions: the oven temperature 100°C, increasing at a rate of 10°C/min up to 200°C, held at 200°C for 5 min, then increasing at 2°C/min up to 230°C and held at 230°C for 10 min, with a final ramp from 230 to 240 at 10°C/min. The injection, interface and ion source temperatures were adjusted to 250, 240 and 230°C, respectively. High purity helium (99.999%) was used as the carrier gas with a constant flow rate of 1.0 mL/min. 0.5 μL of sample was injected in a 1: 20 split ratio by autosampler. The acquisition of mass spectra data was carried out in fullscan mode (mass range m/z 40–500). The absolute quantification (mg g-1) of each fatty acid was quantified by calculating the peak area ratio of each fatty acid with that of the internal standard (methyl tricosanoate) (ANPEL, Shanghai, China). The values were represented as absolute concentration (mg g-1, dry matter).
Oxidation and Antioxidant Parameters Assays
Hepatopancreas samples were homogenized in 9 volumes of ice-cold physiological saline, then centrifuged at 4000 × g for 10 min at 4°C (Eppendorf centrifuge 5418R, Germany) and protein concentrations of homogenates were determined according to the method of Bradford (1976) with a diagnostic reagent kit (Coomassie protein assay dye; Nanjing Jiancheng Bioengineering Institute, Nanjing, China). The activity of total superoxide dismutase (T-SOD) and the concentration of malondialdehyde (MDA) were measured with reagent kits (Nanjing Jiancheng Bioengineering Institute) based on the previous methods (Placer et al., 1966; Zuo et al., 2013). Catalase (CAT) activity was measured by the diagnostic reagent kit (Nanjing Jiancheng Bioengineering Institute) following the method described by Goth (1991). Superoxide radical scavenging activity was detected by the nitrite method following the instruction of reagent kit (Nanjing Jiancheng Bioengineering Institute) (Oyanagui, 1984). Hydroxyl radical scavenging activity was assayed by the Fenton reaction by monitoring the decrease in H2O2 level using a reagent kit (Nanjing Jiancheng Bioengineering Institute) (Moore et al., 2006). The concentration of 8-hydroxydeoxyguanosine (8-OHdG) was assessed by the enzyme-linked immunosorbent assay following the manufacturer’s instructions of kit (Nanjing Jiancheng Bioengineering Institute) (Wu et al., 2017).
Hepatopancreas Mitochondria Isolation and Purification
Mitochondria were isolated from hepatopancreas samples by the differential centrifugation method with some modification according to the manufacturer’s instructions for the mitochondria isolation kit (G006-1-1; Nanjing Jiancheng Bioengineering Institute), and used for the determination of mitochondrial complex activity, ATP levels and mitochondrial membrane potential (Bustamante et al., 1977; Yuan et al., 2019). Briefly, hepatopancreas samples (approximately 200 mg) were rinsed with saline, minced in 1.5-mL pre-cooled lysis buffer, homogenized in a glass homogenizer, and centrifuged at 800 × g for 5 min at 4°C. The supernatant (0.5 mL) was transferred into a new pre-cooled microfuge tube and centrifuged at 15,000 × g for 10 min at 4°C. The sedimented mitochondria were resuspended in 0.2 mL rinse solution and centrifuged at 15,000 × g for 10 min at 4°C. Finally, the resultant mitochondrial pellet was resuspended in the appropriate buffer and stored at −80°C. All operations were carried out on ice. The concentration of mitochondrial protein was determined using a BCA protein assay kit according to the manufacturer’s protocol (P0012; Beyotime, Shanghai, China), and the data expressed as μg mL-1.
Mitochondrial Electron Transport Chain Complex Activity Assays
The mitochondrial supernatants were used to determine the activities of mitochondrial complexes I-V using mitochondrial enzyme activity assay kits (Solarbio, Beijing, China) based on the previous method (Kirby et al., 2007). In brief, the activity of mitochondrial complex I was analysed using a Complex I Activity Assay Kit (BC0515, Solarbio, Beijing, China) through determining the NADH oxidation rate at 340 nm. The activity of mitochondrial complex II was determined by detecting the 2,6-dichloroindophenol reduction rate using a Complex II Activity Assay Kit (BC3230, Solarbio, Beijing, China). The activities of mitochondrial complex III and complex IV were measured by detecting light absorption at 550 nm using Complex III Activity Assay Kit (BC3240, Solarbio, Beijing, China) and Complex IV Activity Assay Kit (BC0945, Solarbio, Beijing, China), respectively. We use the Complex V Activity Assay Kit (BC1445, Solarbio, Beijing, China) to measure the increasing rate of phosphate radical, which reflected mitochondrial complex V activity. The activities of mitochondrial complexes I-V were presented as U mg-1 protein.
ATP Levels Determination
The ATP levels in hepatopancreas of swimming crab were determined using ATP assay kit (A095-1-1; Nanjing Jiancheng Bioengineering Institute) according to the protocol used previously with some modification (Xu et al., 2017). Briefly, hepatopancreas samples were homogenized in boiling double distilled water (10%, w/v), boiled in a water bath for 10 min, and then centrifuged at 4000 × g for 10 min at room temperature, and the supernatant collected for later measurement. The concentration of ATP was determined based on the phosphomolybdic acid colorimetry method which quantified phosphocreatine production at 636 nm (Spectramax M2, Molecular Devices, USA) with data expressed as μmol mg-1 protein.
Mitochondrial Membrane Potential Measurement
Mitochondrial membrane potential of purified mitochondria of hepatopancreas was assessed using JC-1 mitochondrial membrane potential assay kit (C2006; Beyotime, Shanghai, China) based on the method described in detail previously (Reers et al., 1995; Lauer et al., 2012). Briefly, the mitochondria suspension solution (10 μL) and JC-1 working solution (90 μL) were mixed in the 96-well microplate. After incubation for 1 hr at 20°C in the dark, fluorescence generated in the reaction mixture was read (excitation/emission wavelength = 485/590 nm) using a fluorescence microplate reader (Spectramax M2, Molecular Devices, USA) and mitochondrial membrane potential (ΔΨm) expressed as × 103 U mg-1 mitochondrial protein.
NAD+ and NADH Quantification Assays
The NAD+ and NADH (reduced nicotinamide adenine dinucleotide) concentrations in hepatopancreas of crabs were determined using NAD+/NADH assay kit (S0175; Beyotime) following the manufacturer’s protocol by measuring absorbance at 450 nm and calculating according to the NADH standard curve (Sun et al., 2019). Briefly, hepatopancreas samples (approximately 40 mg) were homogenized in NAD+/NADH extracting solution (400 μL), and then centrifuged at 12,000 × g for 10 min at 4°C, and the supernatant collected. The supernatant samples were divided to measure NAD+total (the total of NAD+ and NADH) and NADH separately. A portion of the supernatant was incubated for 30 min at 60°C to decompose NAD+ and translate to NADH. Data were normalized to total protein content as determined by the Bradford protein assay kit (W042-1-1; Nanjing Jiancheng Bioengineering Institute).
MtDNA Copy Number Analysis
Hepatopancreas mitochondrial DNA (mtDNA) copy number was determined as described previously (Hartmann et al., 2011; Liao et al., 2016). The relative mtDNA copy number was measured by quantitative real time PCR with primers for the mitochondrial 16S rRNA gene and the nuclear β-actin gene. The primer sequences of 16S rRNA were CGCCTGTTTATCAAAAACAT (forward primer) and CCGGTCTGAACTCAGATCACG (reverse primer) (Dai et al., 2010). The primer sequences of β-actin were TCACACACTGTCCCCATCTACG (forward primer) and ACCACGCTCGGTCAGGATTTTC (reverse primer) (Cui et al., 2010). Total DNA of hepatopancreas was extracted using the FastPure Cell/Tissue DNA Isolation Mini Kit (DC102; Vazyme, Nanjing, China) according to the manufacturer’s instruction. Quantitative real time PCR was performed as described below.
Assay of Total RNA Extraction, and Reverse Transcription and Quantitative Real-Time PCR
Total RNA isolation, cDNA synthesis and quantitative real-time PCR (qRT-PCR) were conducted based on the procedures as described previously (Yuan et al., 2021). Total RNA of hepatopancreas was extracted using RNA Isolater Total RNA Extraction Reagent (R401-01; Vazyme), and cDNA was synthesized by HiScript II Q RT SuperMix (R223-01; Vazyme). Quantitative real-time PCR was conducted using ChamQ Universal SYBR qPCR Master Mix (Q711-02; Vazyme) and run on a Lightcycler 96 Real Time PCR system (Roche, Switzerland). The specific primers were designed using the online Primer3Plus program (http://www.bioinformatics.nl/primer3plus), and synthesized by TSINGKE Biological Technology (Beijing, China) (Table S3). The expression levels of genes were calculated using the 2−ΔΔt method (Livak and Schmittgen, 2001). Gene expression was normalized to β-actin (housekeeping gene) expression with the FO sample as calibration control.
Statistical Analysis
Results are presented as means ± SE (n = 4). Before statistical analysis, the data were first checked for normality of distribution and homogeneity of variance by using the Kolmogorov-Smirnoff and Levene’s tests, respectively. One-factor analysis of variance (ANOVA) with Tukey’s multiple range post-hoc test were used to analyze the data. The statistical analysis was conducted by using SPSS software version 22 for Windows (IBM, Chicago, USA), with P < 0.05 considered as being significant.
Results
Growth Performance
The results of growth performance have shown that crabs fed the diet supplemented with fish oil and krill oil had a significantly higher PWG, survival, MR, SGR and FE than those fed the diet contained with linseed oil (P < 0.05). Furthermore, it was interesting to note that krill oil was significantly better than fish oil in promoting the growth of swimming crab, which was reflected in each growth performance index (P < 0.05) (Figure 1).
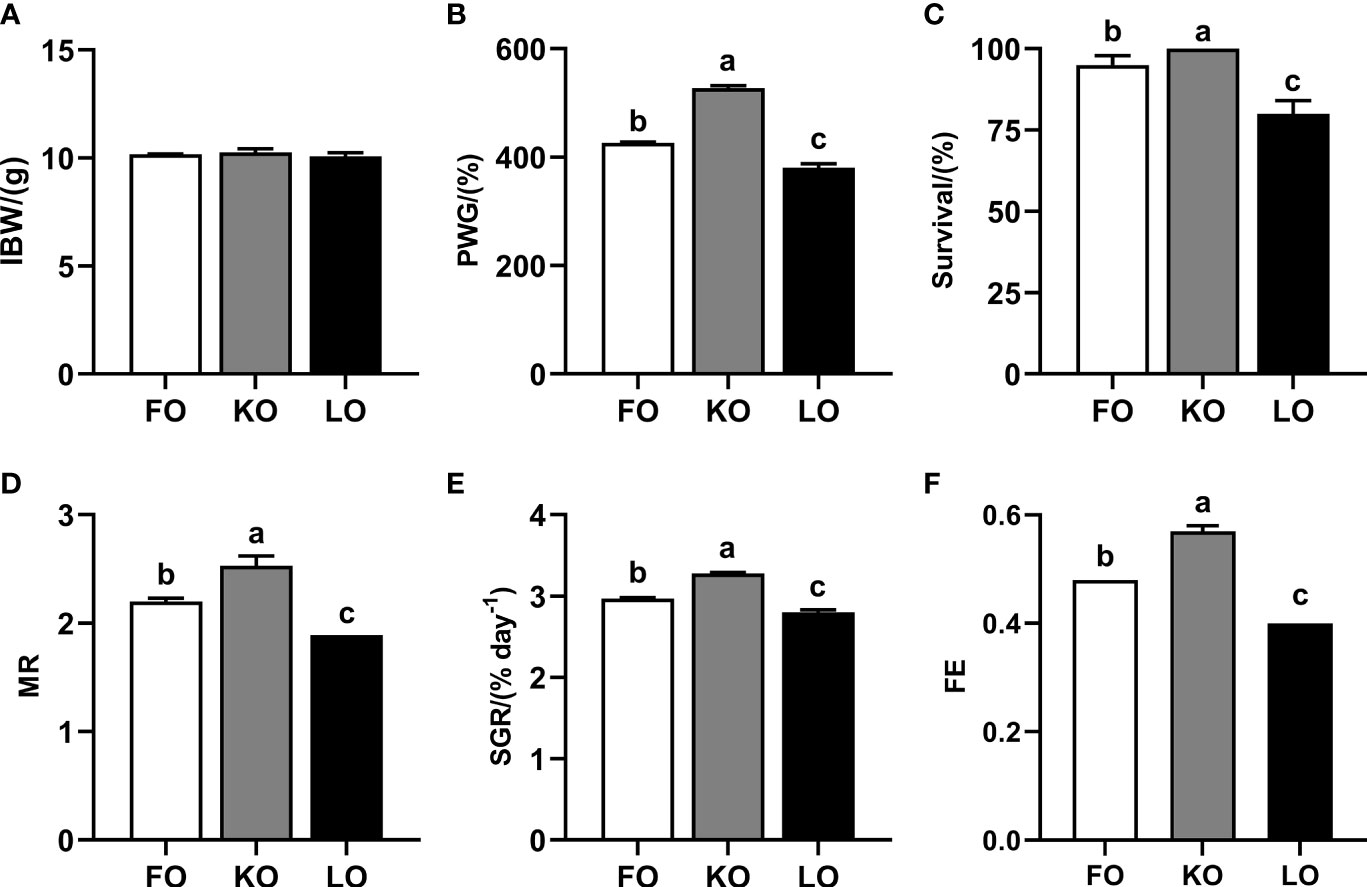
Figure 1 Growth performance and feed utilization of swimming crab fed with different dietary lipid sources. Data are presented as the mean ± SE of four replicates (n = 4). Values in the same row with different superscripts are significantly different (P < 0.05). (A) IBW, initial body weight; (B) PWG (percent weight gain, %) = 100 × [final body weight (g) − initial body weight (g)]/initial body weight (g); (C) Survival (%) = 100 × (final number of crab) / (initial number of crab); (D) MR (molting ratio) = 2 × the number of molting/(final number of crab + initial number of crab); (E) SGR (specific growth rate, % d-1) = 100 × [Ln (final body weight) − Ln(initial body weight)]/days; (F) FE (feed efficiency) = weight gain (g, wet weight)/feed consumed (g, dry weight).
Oxidative Stress in the Hepatopancreas
Total superoxide dismutase (T-SOD) activity in hepatopancreas was significantly higher in crabs fed the KO and LO diets than that in crabs fed the FO diet (P < 0.05) (Figure 2A). The concentration of malondialdehyde (MDA) was significantly lower in crabs fed KO diet than crabs fed the other diets (P < 0.05), with the FO group highest (Figure 2B). Crabs fed the diet supplemented with krill oil had significantly higher activities of catalase (CAT), superoxide radical scavenging and hydroxyl radical scavenging than those fed the diets containing fish oil and linseed oil (P < 0.05) (Figures 2C–E). The levels of 8-hydroxydeoxyguanosine (8-OhdG) in hepatopancreas were significantly lower in crabs fed the KO diet than in crabs fed the FO diet (P < 0.05) (Figure 2F).
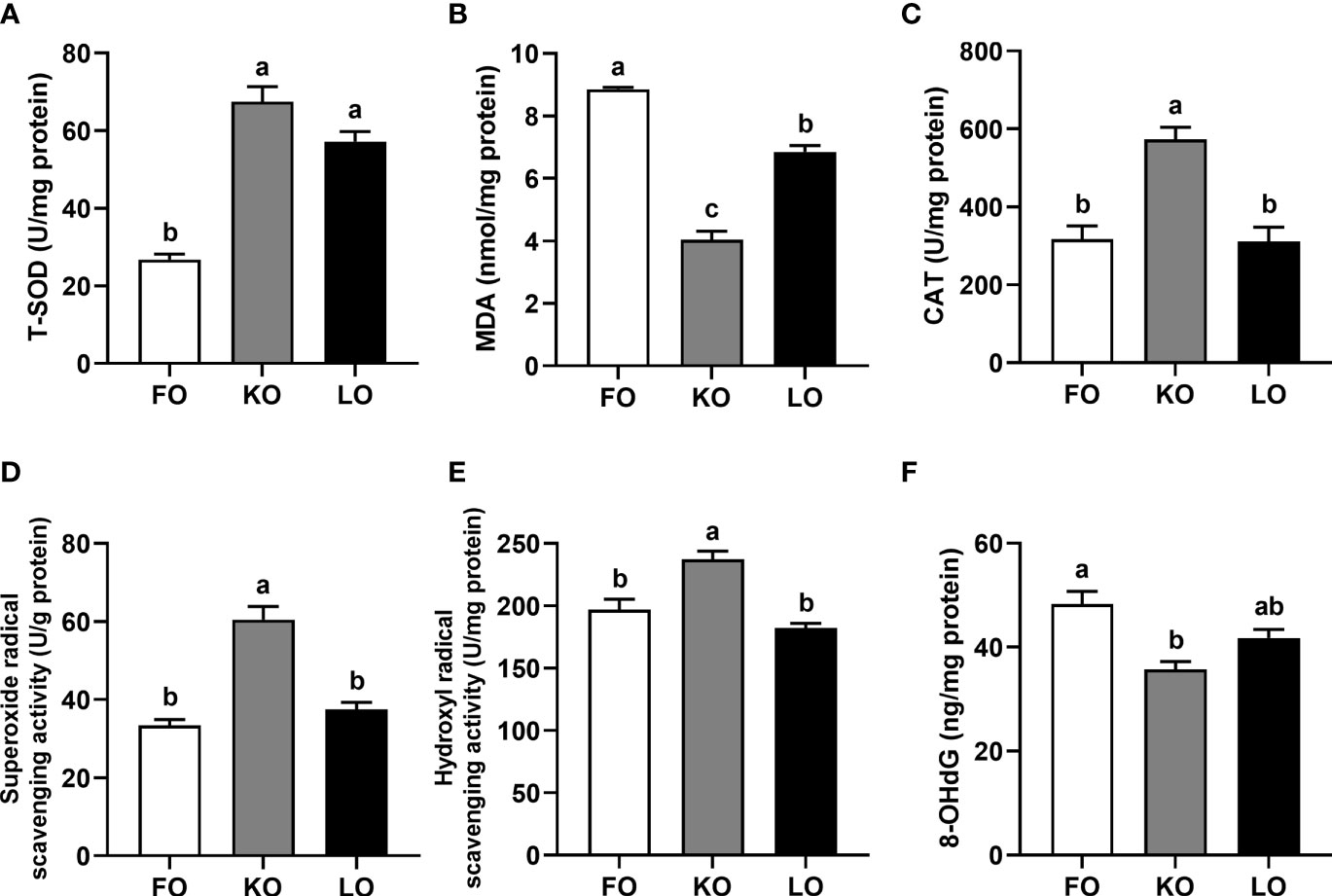
Figure 2 Effects of different dietary lipid sources on indicators of oxidative stress in the hepatopancreas of swimming crab. Values are mean ± SE (n = 4). Bars with different superscripts represent significant difference by one-factor ANOVA followed by a Tukey’s multiple range post-hoc test (P < 0.05). (A) T-SOD, total superoxide dismutase; (B) MDA, malondialdehyde; (C) CAT, catalase; (D) Superoxide radical scavenging activity; (E) Hydroxyl radical scavenging activity; (F) 8-OhdG, 8-hydroxydeoxyguanosine.
Mitochondrial Electron Transport Chain Complex Activity
The activities of mitochondrial complex I and complex III were significantly higher in crabs fed the FO and KO diets than in crabs fed the LO diet (P < 0.05) (Figures 3A, C), and crabs fed diet containing krill oil had a significantly higher activity of mitochondrial complex II than crabs fed the other diets (P < 0.05) (Figure 3B). Although the activity of mitochondrial complex IV was not affected by dietary lipid sources (P > 0.05), it showed a trend with FO > KO > LO (Figure 3D). A similar pattern was observed with mitochondrial complex V activity, which was significantly higher in crabs fed the FO diet than in crabs fed LO (P < 0.05) with crabs fed KO showing an intermediate level not significantly different to either FO or LO (P > 0.05) (Figure 3E).
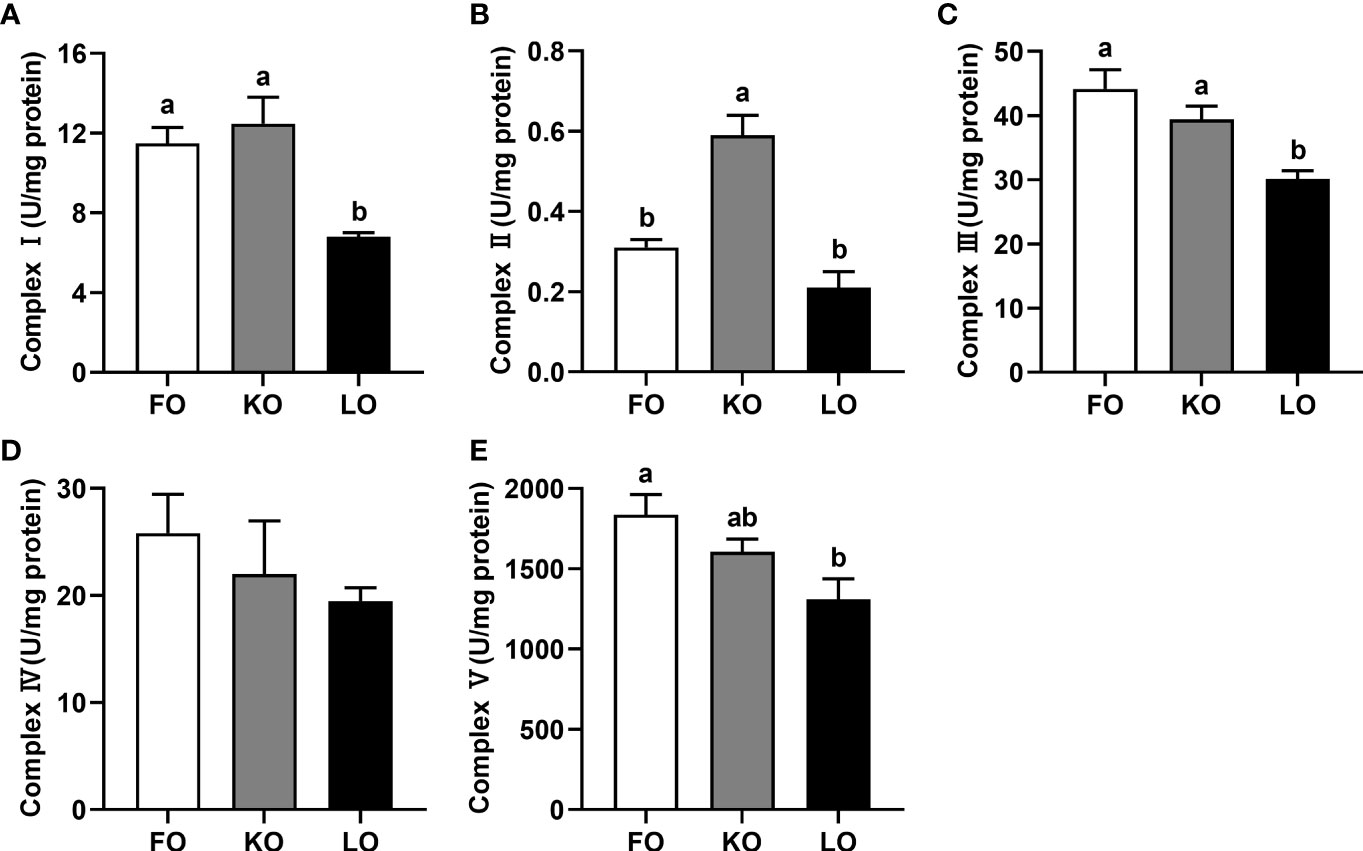
Figure 3 Effects of dietary n-3 PUFA lipid sources on activities of mitochondrial electron transport chain complexes in the hepatopancreas of swimming crab. Values are mean ± SE (n = 4). Bars with different superscripts represent significant difference by one-factor ANOVA followed by a Tukey’s multiple range post-hoc test (P < 0.05). (A) Complex I; (B) Complex II; (C) Complex III; (D) Complex IV; (E) Complex V.
ATP Level
The ATP level of the hepatopancreas in the FO group was significantly higher than other groups (P < 0.05), with the ATP level in crabs fed the KO diet being intermediate, and lowest in the LO group (Figure 4A).
Mitochondrial Membrane Potential
Crabs fed the diets formulated with fish and krill oils had significantly higher mitochondrial membrane potentials in the hepatopancreas than crabs fed the diet containing linseed oil (P < 0.05), while no statistic differences between the FO and KO groups (Figure 4B).
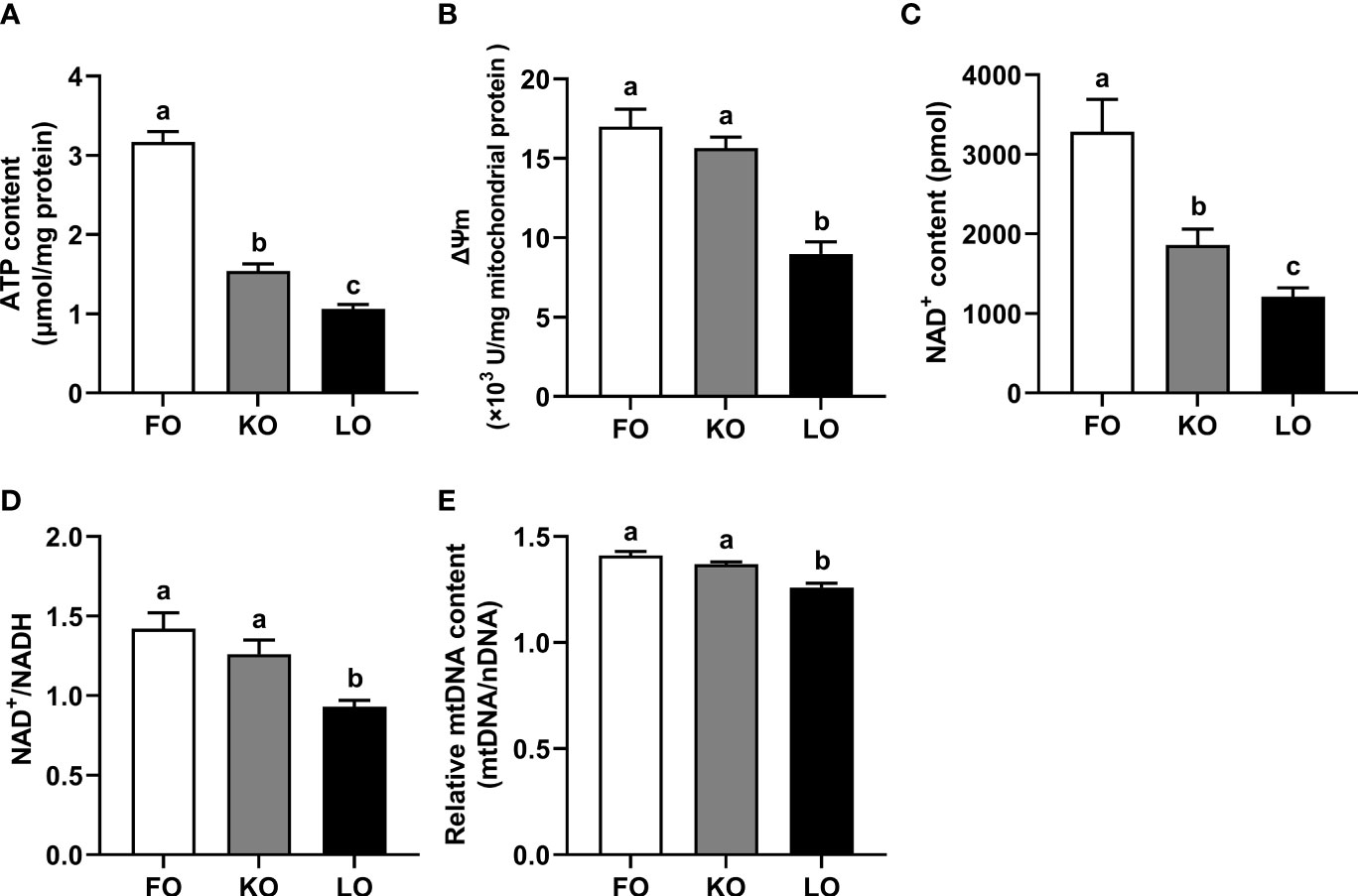
Figure 4 Effects of different dietary lipid sources on the mitochondrial metabolic adaptation of swimming crab. Values are mean ± SE (n = 4). Bars with different superscripts represent significant difference by one-factor ANOVA followed by a Tukey’s multiple range post-hoc test (P < 0.05). (A) ATP, adenosine triphosphate; (B) ΔΨm, mitochondrial membrane potential; (C) NAD+, nicotinamide adenine dinucleotide; (D) NAD+/NADH, the ratio of NAD+ (nicotinamide adenine dinucleotide) and NADH (reduced nicotinamide adenine dinucleotide); (E) Relative mtDNA content, relative mtDNA copy number (mtDNA/nDNA).
NAD+ and NAD+/NADH Ratio
The NAD+ level of the hepatopancreas in crabs fed the FO diet were significantly higher than that in crabs fed other diets (P < 0.05) (Figure 4C), followed by KO group, and lowest in the LO group. The NAD+/NADH ratio of the hepatopancreas in FO and KO treatments were significantly higher than those in LO group (P < 0.05) (Figure 4D), while no statistic differences between the FO and KO groups (P > 0.05).
MtDNA Copy Number
The relative mtDNA copy number was significantly higher in the FO and KO groups compared to the LO group (P < 0.05), whereas no significant difference was found between the FO and KO treatments (P > 0.05) (Figure 4E).
Relative mRNA Expression of Genes Related to Lipolysis, Electron Transport Chain and Energy Metabolism
Dietary lipid sources significantly influenced the expression levels of genes related to lipolysis, electron transport chain and energy metabolism of hepatopancreas (P < 0.05). It was found that the expression levels of lipoprotein lipase (lpl), carnitine palmitoyltransferase 1 (cpt1) and carnitine palmitoyltransferase 2 (cpt2) mRNA were significantly higher in crabs fed KO than in crabs fed FO (P < 0.05) (Figure 5A). Furthermore, the mRNA abundances of genes related to the electron transport chain complexes [NADH dehydrogenase subunit 1 (nd1), succinate dehydrogenase subunit c (sdhc)] were higher in crabs fed KO than in crabs fed the other diets (P < 0.05). The expression levels of cytb, cox I and cox III were significantly higher in crabs fed KO compared to crabs fed LO (P < 0.05), while no significant differences were found between crabs fed KO and FO (P > 0.05) (Figure 5B). Furthermore, the mRNA abundances of ATP synthase 6 (Atpase6), silent information regulator 1(sirt1) and silent information regulator 3 (sirt3) were significantly higher in crabs fed KO than in crabs fed the other diets (P < 0.05). The expression levels of nuclear respiratory factor 1 (nrf1) in crabs fed the FO and KO diets were significantly higher than in crabs fed the LO diet (P < 0.05) (Figure 5C).
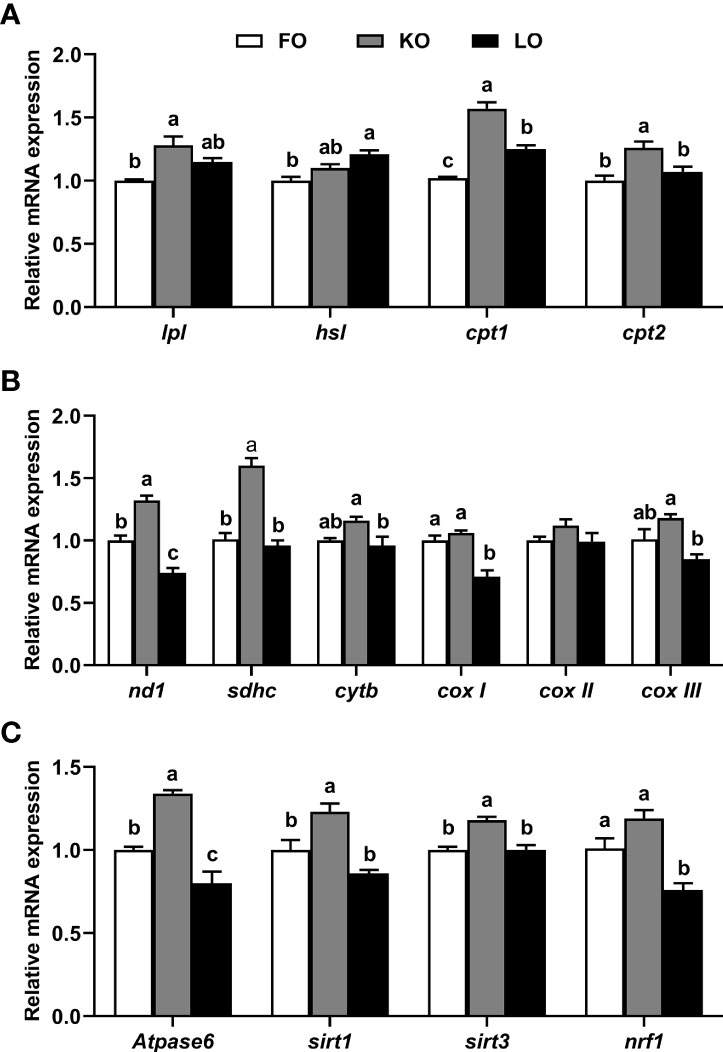
Figure 5 Effects of dietary n-3 PUFA lipid sources on the expression levels of genes related to lipolysis (A), electron transport chain complexes (B) and energy metabolism (C) in the hepatopancreas of swimming crab (Portunus trituberculatus). Values are mean ± SE (n = 4). Bars with different superscripts represent significant difference by one-factor ANOVA followed by a Tukey’s multiple range post-hoc test (P < 0.05). Atpase6, ATP synthase 6; cox I, cytochrome c oxidase subunit I; cox II, cytochrome c oxidase subunit II; cox III, cytochrome c oxidase subunit III; cpt, carnitine palmitoyltransferase; cytb, cytochrome b; hsl, hormone-sensitive lipase; lpl, lipoprotein lipase; nd1, NADH dehydrogenase subunit 1; nrf1, nuclear respiratory factor 1; sdhc, succinate dehydrogenase subunit c; sirt, silent information regulator.
Discussion
In brief, crabs fed the diets containing with krill oil had a significantly higher growth performance and molting ratio than those of crabs fed diets supplemented with linseed oil and even the fish oil. Actually, a recent lipidomic study of hepatopancreas of swimming crab from our research team has evaluated and tested the hypothesis that different n-3 PUFA lipid sources cause different growth performance of swimming crab from the perspective of the lipid molecular species and the stereospecific position of individual fatty acid on the glycerol backbone (Yuan et al., 2021). The previous study has demonstrated that dietary lipid and fatty acid sources could significantly affect the stereospecific position of individual fatty acids on the glycerol backbone of triglyceride (TG), phosphatidylcholine (PC) and phosphatidylethanolamine (PE) and further alter the fluidity of cell membranes and impact metabolic reactions (Yuan et al., 2021). Furthermore, the transcriptomics based on high-throughput sequencing was used to investigate the molecular mechanism by which krill oil promotes the growth and molting of the swimming crab (Fang et al., 2021). That research has showed that swimming crab fed the diet supplemented with krill oil can promote the growth performance and molting through enhancing the transport of glucose, lipids and fatty acids, and meanwhile up-regulating the expression levels of genes related to molting, immunity and energy metabolism (Fang et al., 2021). Here, the present study attempted to explain why dietary krill oil promoted the growth and molting of swimming crab from the perspective of mitochondrial metabolic adaptation and energy metabolism. The possible mechanism may be through improving the mitochondrial metabolic responses and maintaining the energy homeostasis after molting of swimming crab, and further increasing the molting ratio.
Hepatopancreas, as the main metabolic organ of crustaceans, carries out complex and varied metabolic activities, with one of the most crucial being lipid catabolism and mitochondrial fatty acid β-oxidation, which are key for producing the energy required to drive growth, molting, reproduction and other energy-consuming activities (Wang et al., 2014). In this way, hepatopancreas is arguably the tissue most sensitive to dietary lipid sources or fatty acids, just as the liver is in fish and mammals (Xu et al., 2020). Therefore, in the present study, hepatopancreas was considered to be the ideal organ to explore the effects of dietary lipid sources and fatty acids on mitochondrial metabolic adaptation in swimming crab.
The continuous energy supply for metabolic activities is based on the ability of cells to sense, metabolize and convert nutrients into chemical energy via mitochondria, the primary organelle for energy generation (Seungyoon and Pekkurnaz, 2018). Furthermore, mitochondria, as the metabolic center of cellular pathways including the citric acid cycle (Martínez-Reyes et al., 2016), ROS generation (Sena and Chandel, 2012) and calcium signaling (Raffaello et al., 2016), play a vital role in energy homeostasis in the organism (Cogliati et al., 2018). Much of the mitochondrial energy conversion is dependent upon the activity of electron transport chain (ETC), which is composed of 5 protein complexes including NADH dehydrogenase (complex I), succinate-ubiquinone oxidoreductase (complex II), ubiquinol-cytochrome c oxidoreductase (complex III), cytochrome c oxidase (complex IV) and ATP synthase (complex V), the basis of ATP production during oxidative phosphorylation (OXPHOS) (Duchen, 2004). Complex I, as the initiation complex and the major rate-limiting enzyme of the respiratory chain electron transport, is primarily responsible for transfer electrons from matrix NADH to ubiquinone (Zhao et al., 2019). Complex II, as a component of the ETC and Krebs cycle, plays an important role in serving as a bridge between the physiological metabolism and OXPHOS (Cecchini, 2003). Complex III is considered to be required for the hypoxic stabilization of hypoxia-inducible factors (HIF) (Guzy et al., 2005). Complex IV, as the terminal enzyme of the ETC, can transfer electrons from cytochrome c to O2 and reduce the oxygen to H2O (Soro-Arnaiz et al., 2016). Complex V can phosphorylate ADP to ATP through the energy generated by the gradient of the proton electrochemical (Jonckheere et al., 2012). According to the data of present study, crabs fed the KO diet showed a high activity of ETC complexes and a coordinated upregulation of nuclear-encoded genes regulating mitochondrial ETC. It was similar to the results of a previous study in mice which the genes involved in the synthesis of mitochondrial OXPHOS proteins were up-regulated in mice fed the diet supplemented with krill oil compared to dietary fish oil (Burri et al., 2011). Furthermore, a research has also reported that dietary krill oil supplementation could improve the complex activities of rat when it treated with high-fat diet (Ferramosca et al., 2015). The possible mechanism may be the high levels of EPA and DHA structured phospholipids in krill oil, on the one hand, phospholipids, the important components of the mitochondria system, can directly impacts the mitochondrial function (Mejia and Hatch, 2016); on the other hand, high contents of EPA and DHA structured phospholipids supplementation can increase the levels of EPA and DHA in the sn-2 positions of phosphatidylcholine (PC) and phosphatidylethanolamine (PE) molecules and may further promote the fluidity of mitochondrial membranes and its function (Yuan et al., 2021).
ATP, as the energy currency, is mainly generated by mitochondrial ATP synthase, with the energy produced by the hydrolysis of ATP to ADP and phosphate used to fulfill the requirement of cells (Hardie et al., 2012). In addition, the mitochondrial membrane potential (ΔΨm) is generated by proton pumps (i.e. complexes I, III and IV) and can also serve as an intermediate form of energy storage (Zorova et al., 2018). Indeed, the mitochondrial membrane potential is a prerequisite to drive mitochondrial oxidative phosphorylation and ATP synthesis, and stability of the mitochondrial membrane potential is conducive to maintaining normal physiological functions of cells (Izyumov et al., 2004). In the present study, the mitochondrial membrane potential was significantly higher in crabs fed FO and KO compared to crabs fed LO, which may indicate a higher ability of ATP synthesis and viability of mitochondria in swimming crab of FO and KO groups. In addition to the above, nicotinamide adenine dinucleotide (NAD+), a central and universal metabolic cofactor, not only participates in the production of ATP and the maintenance of mitochondrial membrane potential, but also activate sirtuins, and maintains mitochondrial fitness (Stein and Imai, 2012; Cantó et al., 2015). Furthermore, an optimal NAD+/NADH ratio is also crucial for mitochondrial function (Ying, 2008; Houtkooper et al., 2010). The present study showed that crabs fed the FO and KO diets displayed a significantly higher level of NAD+ and NAD+/NADH ratio in comparison with crabs fed diet LO, which indicated greater mitochondrial function.
The oxidative phosphorylation capacity of mitochondria is regulated by the concordant expression of mtDNA and nDNA genes and, therefore, mitochondrial DNA copy number and expression are essential to maintain mitochondrial function (Jeng et al., 2008). While the copy number reflects the state of energy metabolism of mitochondria, mitochondria are also the main intracellular source and immediate target of ROS (Lee and Wei, 2005). Due to the lack of histone protection and the limited capacity for repair of DNA damage, mtDNA is vulnerable to ROS and this can affect ETC function (Lee and Wei, 2005). However, attenuated mitochondrial ETC function can be compensate and normal energy homeostasis maintained by increased mitochondria copy number, which mitigates the mitochondrial dysfunction resulting from decreased mitochondrial content and lower mRNA expression of mtDNA (Bohr, 2002; Lee and Wei, 2005). Consistent with the above statement, crabs fed diets FO and KO displayed a significant increase in the relative content of mtDNA compared to crabs fed LO. Meanwhile, organisms have developed antioxidant defence mechanisms (such as T-SOD, CAT and GSH) to cope with the ROS produced by aerobic metabolism and thus reduce oxidative stress (Meng et al., 2014; Ren et al., 2017). High dietary n-3 LC-PUFA may induce increased lipid peroxidation and MDA (an important indicator for oxidative damage induced by ROS) owing to the high degree of unsaturation in the aliphatic tail (Liao et al., 2015). In the present study, crabs fed the FO diet had a significantly lower activity of T-SOD and a significantly higher content of MDA than those fed the other diets, which indicated that high levels of dietary n-3 LC-PUFA (especially EPA and DHA) could impose a peroxidation burden on swimming crab. However, crabs fed KO had the highest T-SOD activity, superoxide radical scavenging activity and hydroxyl radical scavenging activity as well as the lowest levels of MDA and 8-OHdG in hepatopancreas. This may be related to the high level of astaxanthin in krill oil, which is regarded as an effective antioxidant against lipid peroxidation and oxidative stress (Ambati et al., 2014). Many studies have shown that astaxanthin can protect mitochondrial redox state and functional integrity from oxidative stress (Wolf et al., 2010; Lee et al., 2011; Kim and Kim, 2018). Thus, dietary krill oil could reduce the negative effects of high level of n-3 LC-PUFA, promote the capacity against the lipid peroxidation and prevent a condition of oxidative damage, and further ameliorate the mitochondrial function.
Mitochondria fatty acid β-oxidation is the major pathway of fatty acid catabolism in the cells (Tocher, 2003). The carnitine palmitoyltransferase system, as the most important system in the process of fatty acid β-oxidation, plays an irreplaceable role in the β-oxidation of long chain fatty acids (LCFA) (Tocher, 2003; Lopes-Marques et al., 2015). CPT1, a rate-limiting regulatory enzyme in mitochondrial fatty acid β-oxidation, can participate in catalyzing the synthesis of long-chain fatty acylcarnitine (Kerner and Hoppel, 2000). Then CPT2 can break down the long-chain fatty acylcarnitine into the long-chain fatty acids with the liberation of carnitine (Glatz et al., 2010). In our previous comparative transcriptomic study of hepatopancreas of swimming crab, the expression level of cpt1 of hepatopancreas was significantly up-regulated by dietary krill oil compared to fish and linseed oils, which was consistent with the result of present study (Fang et al., 2021). One study on tiger puffer (Takifugu rubripes) has reported that dietary astaxanthin supplementation could significantly up-regulate the expression level of cpt1, and further enhance the fatty acid β-oxidation in fish (Liao et al., 2018). Furthermore, it has been shown that dietary astaxanthin can enhance the function of CPT1 through the accumulation of astaxanthin on the mitochondrial membrane (Takahashi et al., 2004). However, more researches are needed to elucidate the specific mechanism.
Sirtuins, as the conserved protein NAD+-dependent deacylases, plays an important role in the regulation of cellular processes including cell proliferation, gene expression and apoptosis (Poulose and Raju, 2015). In addition, sirtuins have been proved to be not only the protector of cells against metabolic stresses but also the important energy status sensors which were sensitive to the diet and themselves regulated by diet and environmental stress (Chang and Guarente, 2014). Over past decades, accumulating evidence has indicated that SIRT1 is an essential regulator of systemic energy homeostasis (Li, 2013). The result of the gene expression has showed that crabs fed the diet supplemented with krill oil can significantly up-regulate the expression level of sirt1, and may further improve the level of energy homeostasis. Furthermore, it was also found that significantly up-regulation of sirt3 in KO group compared to other groups. SIRT3, as a regulator of mitochondrial energy metabolism, can deacetylate and activate various target substrates in the oxidation of fatty acids and further maintain the ATP levels (Ahn et al., 2008). Nuclear respiratory factor 1, also known as nrf1, encodes a protein that homodimerizes and directly regulates the expression of nuclear genes encoding subunits of the respiratory complexes, indirectly regulating the three mitochondrial-encoded COX subunit genes for respiration, mitochondrial DNA transcription and replication (Gleyzer et al., 2005; Wang et al., 2006). High expression of nrf1 in crabs fed the KO diet demonstrated a positive effect of krill oil on the up-regulation of genes and pathways involved in hepatopancreas mitochondrial ETC and energy metabolism.
Conclusion
In conclusion, the results showed that compared with linseed and fish oils, krill oil significantly promoted the growth and molting of juvenile swimming crab, increased activities of ETC complexes, the membrane potential, NAD+ level, NAD+/NADH ratio and the copy number of mitochondrial DNA, as well as up-regulated the expression levels of genes related to lipolysis, ETC and energy metabolism. Furthermore, dietary krill oil also specifically improved the ability for scavenging free radicals produced in the process of physiological metabolism, reduced the level of lipid peroxides and the degree of DNA oxidative damage, mitigated the damage to mitochondrial ETC function caused by lipid peroxidation, and improved the health status of swimming crab. Overall, results of the present study clearly suggested that dietary krill oil promoted the molting and growth via enhancing the mitochondrial metabolic adaptation and ameliorating the energy homeostasis (Figure 6).
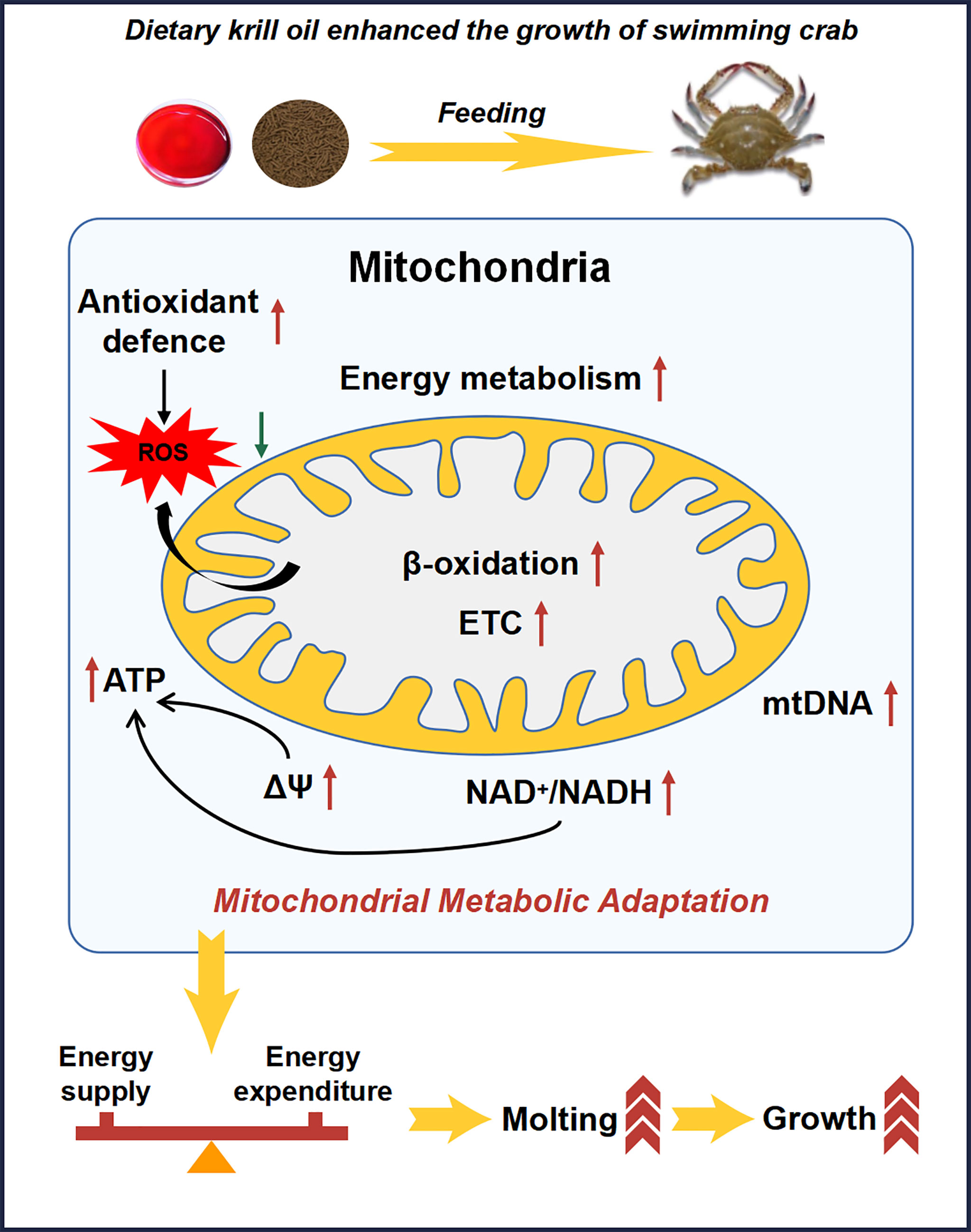
Figure 6 Schematic of dietary krill oil greatly enhances the growth of swimming crab via promoting the mitochondrial metabolic adaptation and energy homeostasis.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics Statement
The animal study was reviewed and approved by Ethics Scientific Committee for Experiments on Animals of Ningbo University.
Author Contributions
YY: Conceptualization, methodology, formal analysis, investigation, visualization, writing-original draft, writing-review & editing. MJ: Conceptualization, investigation, supervision, writing-review & editing, project administration, funding acquisition. FF: Methodology, software, writing-review & editing. DT: Conceptualization, methodology, writing-review & editing. MB: Conceptualization, methodology, writing-review & editing. LJ: Software, writing-review & editing, visualization. YH: Writing-review & editing. QZ: Conceptualization, methodology, supervision, data curation, writing-review & editing, project administration and funding acquisition. All authors contributed to the article and approved the submitted version.
Funding
This research was supported by National Natural Science Foundation of China (32072987), China Agriculture Research System of MOF and MARA (CARS-48), National Key R & D Program of China (2018YFD0900400), Nature Science Foundation of Zhejiang Province (LY21C190006) and the K. C. Wong Magna Fund in Ningbo University.
Conflict of Interest
Author YH is employed by Guangdong Yuequn Biotechnology Co., Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgements
The authors are grateful to Man Liu (Ningbo Institute of Materials Technology and Engineering, Ningbo, China) for her technical support and valuable help during the analysis of fatty acids.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.914590/full#supplementary-material
Glossary

References
Ahn B. H., Kim H. S., Song S., Lee I. H., Liu J., Vassilopoulos A., et al. (2008). A Role for the Mitochondrial Deacetylase Sirt3 in Regulating Energy Homeostasis. P Natl. Acad. Sci. U.S.A. 105, 14447–14452. doi: 10.1073/pnas.0803790105
Alfadda A. A., Sallam R. M. (2012). Reactive Oxygen Species in Health and Disease. J. BioMed. Biotechnol. 2012, 936486. doi: 10.1155/2012/936486
Ambati R. R., Phang S. M., Ravi S., Aswathanarayana R. G. (2014). Astaxanthin: Sources, Extraction, Stability, Biological Activities and its Commercial Applications–a Review. Mar. Drugs 12, 128–152. doi: 10.3390/md12010128
AOAC. (2006). Official Methods of Analysis, 18th ed. Arlington: VA: Association of Official Analytical Chemists.
Bohr V. A. (2002). Repair of Oxidative DNA Damage in Nuclear and Mitochondrial DNA, and Some Changes With Aging in Mammalian Cells. Free Radical Bio Med. 32, 804–812. doi: 10.1016/S0891-5849(02)00787-6
Bradford M. (1976). A Rapid Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 72, 248–254. doi: 10.1016/0003-2697(76)90527-3
Bremer K., Monk C. T., Gurd B. J., Moyes C. D. (2012). Transcriptional Regulation of Temperature-Induced Remodeling of Muscle Bioenergetics in Goldfish. Am. J. Physiol. Regul. Integr. Comp. Physiol. 303, R150–R158. doi: 10.1152/ajpregu.00603.2011
Bremer K., Moyes C. D. (2011). Origins of Variation in Muscle Cytochrome C Oxidase Activity Within and Between Fish Species. J. Exp. Biol. 214, 1888–1895. doi: 10.1242/jeb.053330
Burri L., Berge K., Wibrand K., Berge R. K., Barger J. L. (2011). Differential Effects of Krill Oil and Fish Oil on the Hepatic Transcriptome in Mice. Front. Genet. 2. doi: 10.3389/fgene.2011.00045
Bustamante E., Soper J. W., Pedersen P. L. (1977). A High-Yield Preparative Method for Isolation of Rat Liver Mitochondria. Anal. Biochem. 80, 401–408. doi: 10.1016/0003-2697(77)90661-3
Cantó C., Menzies K. J., Auwerx J. (2015). NAD+ Metabolism and the Control of Energy Homeostasis: A Balancing Act Between Mitochondria and the Nucleus. Cell Metab. 22, 31–53. doi: 10.1016/j.cmet.2015.05.023
Cecchini G. (2003). Function and Structure of Complex II of the Respiratory Chain. Annu. Rev. Biochem. 72, 77–109. doi: 10.1146/annurev.biochem.72.121801.161700
Chang H. C., Guarente L. (2014). SIRT1 and Other Sirtuins in Metabolism. Trends Endocrin Met. 25, 138–145. doi: 10.1016/j.tem.2013.12.001
Cogliati S., Lorenzi I., Rigoni G., Caicci F., Soriano M. E. (2018). Regulation of Mitochondrial Electron Transport Chain Assembly. J. Mol. Biol. 430 (24), 4849–4873. doi: 10.1016/j.jmb.2018.09.016
Conde-Sieira M., Soengas J. L. (2017). Nutrient Sensing Systems in Fish: Impact on Food Intake Regulation and Energy Homeostasis. Front. Neurosci-Switz. 10. doi: 10.3389/fnins.2016.00603
Cui Z. X., Liu Y., Luan W. S., Li Q. Q., Wu D. H., Wang S. Y. (2010). Molecular Cloning and Characterization of a Heat Shock Protein 70 Gene in Swimming Crab (Portunus trituberculatus). Fish Shellfish Immun. 28, 56–64. doi: 10.1016/j.fsi.2009.09.018
Dai Y. J., Liu P., Gao B. Q., Li J., Wang Q. Y. (2010). Sequence Analysis of Mitochondrial 16S rRNA and COI Gene Fragments of Four Wild Populations of Portunus trituberculatus. Periodical Ocean Univ. China. 40(3), 054–60 doi: 10.3969/j.issn.1672-5174.2010.03.009
Dos Santos R. S., Galina A., Da-Silva W. S. (2012). Cold Acclimation Increases Mitochondrial Oxidative Capacity Without Inducing Mitochondrial Uncoupling in Goldfish White Skeletal Muscle. Biol. Open 2, 82–87. doi: 10.1242/bio.20122295
Duchen M. R. (2004). Mitochondria in Health and Disease: Perspectives on a New Mitochondrial Biology. Mol. Aspects Med. 25, 365–451. doi: 10.1016/j.mam.2004.03.001
Duggan A. T., Kocha K. M., Monk C. T., Bremer K., Moyes C. D. (2011). Coordination of Cytochrome C Oxidase Gene Expression in the Remodelling of Skeletal Muscle. J. Exp. Biol. 214, 1880–1887. doi: 10.1242/jeb.053322
Fang F., Yuan Y., Jin M., Shi B., Zhu T. T., Luo J. X., et al. (2021). Hepatopancreas Transcriptome Analysis Reveals the Molecular Responses to Different Dietary N-3 PUFA Lipid Sources in the Swimming Crab Portunus trituberculatus. Aquaculture 543, 737016. doi: 10.1016/j.aquaculture.2021.737016
Ferramosca A., Conte A., Zara V. (2015). Krill Oil Ameliorates Mitochondrial Dysfunctions in Rats Treated With High-Fat Diet. BioMed. Res. Int. 2015, 645984. doi: 10.1155/2015/645984
Glatz J. F., Luiken J. J., Bonen A. (2010). Membrane Fatty Acid Transporters as Regulators of Lipid Metabolism: Implications for Metabolic Disease. Physiol. Rev. 90, 367–417. doi: 10.1152/physrev.00003.2009
Gleyzer N., Vercauteren K., Scarpulla R. C. (2005). Control of Mitochondrial Transcription Specificity Factors (TFB1M and TFB2M) by Nuclear Respiratory Factors (NRF-1 and NRF-2) and PGC-1 Family Coactivators. Mol. Cell Biol. 25, 1354–1366. doi: 10.1128/MCB.25.4.1354-1366.2005
Goth L. (1991). A Simple Method for Determination of Serum Catalase Activity and Revision of Reference Range. Clinica Chimica Acta 196, 143–151. doi: 10.1016/0009-8981(91)90067-M
Guzy R. D., Hoyos B., Robin E., Chen H., Liu L., Mansfield K. D., et al. (2005). Mitochondrial Complex III is Required for Hypoxia-Induced ROS Production and Cellular Oxygen Sensing. Cell Metab. 1, 401–408. doi: 10.1016/j.cmet.2005.05.001
Hardie D. G., Ross F. A., Hawley S. A. (2012). AMPK: A Nutrient and Energy Sensor That Maintains Energy Homeostasis. Nat. Rev. Mol. Cell Bio. 13, 251–262. doi: 10.1038/nrm3311
Hartmann N., Reichwald K., Wittig I., Dröse S., Schmeisser S., Lück C., et al. (2011). Mitochondrial DNA Copy Number and Function Decrease With Age in the Short-Lived Fish Nothobranchius Furzeri. Aging Cell 10, 824–831. doi: 10.1111/j.1474-9726.2011.00723.x
Houtkooper R. H., Cantó C., Wanders R. J., Auwerx J. (2010). The Secret Life of NAD+: An Old Metabolite Controlling New Metabolic Signaling Pathways. Endocr. Rev. 31, 194–223. doi: 10.1210/er.2009-0026
Huang S., Wang J., Yue W. C., Chen J., Gaughan S., Lu W. Q., et al. (2015). Transcriptomic Variation of Hepatopancreas Reveals the Energy Metabolism and Biological Processes Associated With Molting in Chinese Mitten Crab, Eriocheir sinensis. Sci. Rep-UK 5, 1–11. doi: 10.1038/srep14015
Izyumov D. S., Avetisyan A. V., Pletjushkina O. Y., Sakharov D. V., Wirtz K. W., Chernyak B. V., et al. (2004). “Wages of Fear”: Transient Threefold Decrease in Intracellular ATP Level Imposes Apoptosis. BBA-Bioenergetics 1658, 141–147. doi: 10.1016/j.bbabio.2004.05.007
Jeng J. Y., Yeh T. S., Lee J. W., Lin S. H., Fong T. H., Hsieh R. H. (2008). Maintenance of Mitochondrial DNA Copy Number and Expression are Essential for Preservation of Mitochondrial Function and Cell Growth. J. Cell Biochem. 103, 347–357. doi: 10.1002/jcb.21625
Jokinen R., Pirnes-Karhu S., Pietiläinen K. H., Pirinen E. (2017). Adipose Tissue NAD+-Homeostasis, Sirtuins and Poly (ADP-Ribose) Polymerases-Important Players in Mitochondrial Metabolism and Metabolic Health. Redox Biol. 12, 246–263. doi: 10.1016/j.redox.2017.02.011
Jonckheere A. I., Smeitink J. A. M., Rodenburg R. J. T. (2012). Mitochondrial ATP Synthase: Architecture, Function and Pathology. J. Inherit Metab. Dis. 35, 211–225. doi: 10.1007/s10545-011-9382-9
Jung H., Lyons R. E., Hurwood D. A., Mather P. B. (2013). Genes and Growth Performance in Crustacean Species: A Review of Relevant Genomic Studies in Crustaceans and Other Taxa. Rev. Aquacult. 5, 77–110. doi: 10.1111/raq.12005
Kerner J., Hoppel C. (2000). Fatty Acid Import Into Mitochondria. BBA-Mol Cell Biol. L. 1486, 1–17. doi: 10.1016/S1388-1981(00)00044-5
Kim S. H., Kim H. (2018). Inhibitory Effect of Astaxanthin on Oxidative Stress-Induced Mitochondrial Dysfunction-a Mini-Review. Nutrients 10, 1137. doi: 10.3390/nu10091137
Kirby D. M., Thorburn D. R., Turnbull D. M., Taylor R. W. (2007). Biochemical Assays of Respiratory Chain Complex Activity. Method Cell Biol. 80, 93–119. doi: 10.1016/S0091-679X(06)80004-X
Kraffe E., Marty Y., Guderley H. (2007). Changes in Mitochondrial Oxidative Capacities During Thermal Acclimation of Rainbow Trout Oncorhynchus Mykiss: Roles of Membrane Proteins, Phospholipids and Their Fatty Acid Compositions. J. Exp. Biol. 210, 149–165. doi: 10.1242/jeb.02628
Lauer M. M., de Oliveira C. B., Yano N. L. I., Bianchini A. (2012). Copper Effects on Key Metabolic Enzymes and Mitochondrial Membrane Potential in Gills of the Estuarine Crab Neohelice Granulata at Different Salinities. Comp. Biochem. Phys. C 156, 140–147. doi: 10.1016/j.cbpc.2012.08.001
Lee D. H., Kim C. S., Lee Y. J. (2011). Astaxanthin Protects Against MPTP/MPP+-Induced Mitochondrial Dysfunction and ROS Production In Vivo and In Vitro. Food Chem. Toxicol. 49, 271–280. doi: 10.1016/j.fct.2010.10.029
Lee H. C., Wei Y. H. (2005). Mitochondrial Biogenesis and Mitochondrial DNA Maintenance of Mammalian Cells Under Oxidative Stress. Int. J. Biochem. Cell B. 37, 822–834. doi: 10.1016/j.biocel.2004.09.010
LeMoine C. M., Craig P. M., Dhekney K., Kim J. J., McClelland G. B. (2010). Temporal and Spatial Patterns of Gene Expression in Skeletal Muscles in Response to Swim Training in Adult Zebrafish (Danio Rerio). Comp. Physiol. B. 180, 151–160. doi: 10.1007/s00360-009-0398-5
Li X. (2013). SIRT1 and Energy Metabolism. Acta Biochim. Biophys. Sin. 45, 51–60. doi: 10.1093/abbs/gms108
Liao Z., Xu H., Wei Y., Zhang Q., Liang M. (2018). Dietary Astaxanthin Differentially Affected the Lipid Accumulation in the Liver and Muscle of the Marine Teleost, Tiger Puffer Takifugu Rubripes. Aquac Res. 49, 3421–3433. doi: 10.1111/are.13806
Liao K., Yan J., Mai K. S., Ai Q. H. (2015). Dietary Olive and Perilla Oils Affect Liver Mitochondrial DNA Methylation in Large Yellow Croakers. J. Nutr. 145, 2479–2485. doi: 10.3945/jn.115.216481
Liao K., Yan J., Mai K. S., Ai Q. H. (2016). Dietary Lipid Concentration Affects Liver Mitochondrial DNA Copy Number, Gene Expression and DNA Methylation in Large Yellow Croaker (Larimichthys Crocea). Comp. Biochem. Phys. B. 193, 25–32. doi: 10.1016/j.cbpb.2015.11.012
Livak K. J., Schmittgen T. D. (2001). Analysis of Relative Gene Expression Data Using Realtime Quantitative PCR and the 2–ΔΔCT Method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Lombard D. B., Tishkoff D. X., Bao J. (2011). Mitochondrial Sirtuins in the Regulation of Mitochondrial Activity and Metabolic Adaptation. Handb. Exp. Pharmacol. 206, 163–188. doi: 10.1007/978-3-642-21631-2_8
Lopes-Marques M., Delgado I. L., Ruivo R., Torres Y., Sainath S. B., Rocha E., et al. (2015). The Origin and Diversity of Cpt1 Genes in Vertebrate Species. PloS One 10, e0138447. doi: 10.1371/journal.pone.0138447
Martínez-Reyes I., Diebold L. P., Kong H., Schieber M., Huang H., Hensley C. T. (2016). TCA Cycle and Mitochondrial Membrane Potential are Necessary for Diverse Biological Functions. Mol. Cell. 61, 199–209. doi: 10.1016/j.molcel.2015.12.002
Mejia E. M., Hatch G. M. (2016). Mitochondrial Phospholipids: Role in Mitochondrial Function. J. Bioenerg Biomembr. 48 (2), 99–112. doi: 10.1007/s10863-015-9601-4
Meng X. L., Liu P., Li J., Gao B. Q., Chen P. (2014). Physiological Responses of Swimming Crab Portunus trituberculatus Under Cold Acclimation: Antioxidant Defense and Heat Shock Proteins. Aquaculture 434, 11–17. doi: 10.1016/j.aquaculture.2014.07.021
Moore J., Yin J. J., Yu L. (2006). Novel Fluorometric Assay for Hydroxyl Radical Scavenging Capacity (HOSC) Estimation. J. Agr Food Chem. 54, 617–626. doi: 10.1021/jf052555p
Morris M. R., Wuitchik S. J., Rosebush J., Rogers S. M. (2021). Mitochondrial Volume Density and Evidence for its Role in Adaptive Divergence in Response to Thermal Tolerance in Threespine Stickleback. J. Comp. Physiol. B 191, 657–668. doi: 10.1007/s00360-021-01366-w
Oyanagui Y. (1984). Reevaluation of Assay Methods and Establishment of Kit for Superoxide Dismutase Activity. Anal. Biochem. 142, 290–296. doi: 10.1016/0003-2697(84)90467-6
Panganiban G., Sebring A., Nagy L., Carroll S. (1995). The Development of Crustacean Limbs and the Evolution of Arthropods. Science 270, 1363–1366. doi: 10.1126/science.270.5240.1363
Placer Z. A., Cushman L. L., Johnson B. C. (1966). Estimation of Product of Lipid Peroxidation (Malonyl Dialdehyde) in Biochemical Systems. Anal. Biochem. 16, 359–364. doi: 10.1016/0003-2697(66)90167-9
Poulose N., Raju R. (2015). Sirtuin Regulation in Aging and Injury. BBA-Mol Basis Dis. 1852 (11), 2442–2455. doi: 10.1016/j.bbadis.2015.08.017
Raffaello A., Mammucari C., Gherardi G., Rizzuto R. (2016). Calcium at the Center of Cell Signaling: Interplay Between Endoplasmic Reticulum, Mitochondria, and Lysosomes. Trends Biochem. Sci. 41, 1035–1049. doi: 10.1016/j.tibs.2016.09.001
Rahman M., Nirala N. K., Singh A., Zhu L. J., Taguchi K., Bamba T., et al. (2014). Drosophila Sirt2/mammalian SIRT3 Deacetylates ATP Synthase β and Regulates Complex V Activity. J. Cell Biol. 206, 289–305. doi: 10.1083/jcb.201404118
Reers M., Smiley S. T., Mottola-Hartshorn C., Chen A., Lin M., Chen L. B. (1995). Mitochondrial Membrane Potential Monitored by JC-1 Dye. Method Enzymol. 260, 406–417. doi: 10.1016/0076-6879(95)60154-6
Ren X., Wang Z., Gao B., Liu P., Li J. (2017). Toxic Responses of Swimming Crab (Portunus trituberculatus) Larvae Exposed to Environmentally Realistic Concentrations of Oxytetracycline. Chemosphere 173, 563–571. doi: 10.1016/j.chemosphere.2017.01.078
Sena L. A., Chandel N. S. (2012). Physiological Roles of Mitochondrial Reactive Oxygen Species. Mol. Cell. 48, 158–167. doi: 10.1016/j.molcel.2012.09.025
Seungyoon B. Y., Pekkurnaz G. (2018). Mechanisms Orchestrating Mitochondrial Dynamics for Energy Homeostasis. J. Mol. Biol. 430, 3922–3941. doi: 10.1016/j.jmb.2018.07.027
Sokolova I. M., Frederich M., Bagwe R., Lannig G., Sukhotin A. A. (2012). Energy Homeostasis as an Integrative Tool for Assessing Limits of Environmental Stress Tolerance in Aquatic Invertebrates. Mar. Environ. Res. 79, 1–15. doi: 10.1016/j.marenvres.2012.04.003
Soro-Arnaiz I., Li Q. O. Y., Torres-Capelli M., Meléndez-Rodríguez F., Veiga S., Veys K., et al. (2016). Role of Mitochondrial Complex IV in Age-Dependent Obesity. Cell Rep. 16, 2991–3002. doi: 10.1016/j.celrep.2016.08.041
Stein L. R., Imai S. I. (2012). The Dynamic Regulation of NAD Metabolism in Mitochondria. Trends Endocrin Met. 23, 420–428. doi: 10.1016/j.tem.2012.06.005
Sun Y., Teng Z. Y., Sun X. J., Zhang L. X., Chen J., Wang B. Z., et al. (2019). Exogenous H2S Reduces the Acetylation Levels of Mitochondrial Respiratory Enzymes via Regulating the NAD+-SIRT3 Pathway in Cardiac Tissues of Db/Db Mice. Am. J. Physiol-Endoc M. 317, E284–E297. doi: 10.1152/ajpendo.00326.2018
Takahashi K., Watanabe M., Takimoto T., Akiba Y. (2004). Uptake and Distribution of Astaxanthin in Several Tissues and Plasma Lipoproteins in Male Broiler Chickens Fed a Yeast (Phaffia rhodozyma) With a High Concentration of Astaxanthin. Brit Poultry Sci. 45, 133–138. doi: 10.1080/00071660410001668950a
Tocher D. R. (2003). Metabolism and Functions of Lipids and Fatty Acids in Teleost Fish. Rev. Fisheries Sci. 11, 107–184. doi: 10.1080/713610925
Urschel M. R., O'Brien K. M. (2008). High Mitochondrial Densities in the Hearts of Antarctic Icefishes are Maintained by an Increase in Mitochondrial Size Rather Than Mitochondrial Biogenesis. J. Exp. Biol. 211, 2638–2646. doi: 10.1242/jeb.018598
Wang C. G., Li Z. P., Lu Y. N., Du R. L., Katiyar S. J., Yang J. G., et al. (2006). Cyclin D1 Repression of Nuclear Respiratory Factor 1 Integrates Nuclear DNA Synthesis and Mitochondrial Function. P Natl. Acad. Sci. U.S.A. 103, 11567–11572. doi: 10.1073/pnas.0603363103
Wang W., Wu X. G., Liu Z. J., Zheng H. J., Cheng Y. X. (2014). Insights Into Hepatopancreatic Functions for Nutrition Metabolism and Ovarian Development in the Crab Portunus trituberculatus: Gene Discovery in the Comparative Transcriptome of Different Hepatopancreas Stages. PloS One 9, e84921. doi: 10.1371/journal.pone.0084921
Wolf A. M., Asoh S., Hiranuma H., Ohsawa I., Iio K., Satou A., et al. (2010). Astaxanthin Protects Mitochondrial Redox State and Functional Integrity Against Oxidative Stress. J. Nutr. Biochem. 21, 381–389. doi: 10.1016/j.jnutbio.2009.01.011
Wu D. N., Liu B. D., Yin J. F., Xu T., Zhao S. L., Xu Q., et al. (2017). Detection of 8-Hydroxydeoxyguanosine (8-OHdG) as a Biomarker of Oxidative Damage in Peripheral Leukocyte DNA by UHPLC–Ms/MS. J. Chromatogr B. 1064, 1–6. doi: 10.1016/j.jchromb.2017.08.033
Xie S. C., Zhou Q. C., Zhang X. S., Zhu T. T., Guo C., Yang Z., et al. (2022). Effect of Dietary Replacement of Fish Meal With Low-Gossypol Cottonseed Protein Concentrate on Growth Performance and Expressions of Genes Related to Protein Metabolism for Swimming Crab (Portunus trituberculatus). Aquaculture 549, 737820. doi: 10.1016/j.aquaculture.2021.737820
Xu F., Liu Y., Zhao H., Yu K., Song M., Zhu Y., et al. (2017). Aluminum Chloride Caused Liver Dysfunction and Mitochondrial Energy Metabolism Disorder in Rat. J. Inorg Biochem. 174, 55–62. doi: 10.1016/j.jinorgbio.2017.04.016
Xu H. G., Turchini G. M., Francis D. S., Liang M. Q., Mock T. S., Rombenso A., et al. (2020). Are Fish What They Eat? A Fatty Acid’s Perspective. Prog. Lipid Res. 80, 101064. doi: 10.1016/j.plipres.2020.101064
Ying W. (2008). NAD+/NADH and NADP+/NADPH in Cellular Functions and Cell Death: Regulation and Biological Consequences. Antioxid Redox Sign. 10, 179–206. doi: 10.1089/ars.2007.1672
Yuan Y., Sun P., Jin M., Wang X. X., Zhou Q. C. (2019). Regulation of Dietary Lipid Sources on Tissue Lipid Classes and Mitochondrial Energy Metabolism of Juvenile Swimming Crab, Portunus Trituberculatus. Front. Physiol. 10. doi: 10.3389/fphys.2019.00454
Yuan Y., Wang X. X., Jin M., Jiao L. F., Sun P., Betancor M. B., et al. (2020). Modification of Nutritional Values and Flavor Qualities of Muscle of Swimming Crab (Portunus Trituberculatus): Application of a Dietary Lipid Nutrition Strategy. Food Chem. 308, 125607. doi: 10.1016/j.foodchem.2019.125607
Yuan Y., Wang X. X., Jin M., Sun P., Zhou Q. C. (2019). Influence of Different Lipid Sources on Growth Performance, Oxidation Resistance and Fatty Acid Profiles of Juvenile Swimming Crab, Portunus Trituberculatus. Aquaculture 508, 147–158. doi: 10.1016/j.aquaculture.2019.04.068
Yuan Y., Xu F. M., Jin M., Wang X. X., Hu X. Y., Zhao M. M., et al. (2021). Untargeted Lipidomics Reveals Metabolic Responses to Different Dietary N-3 PUFA in Juvenile Swimming Crab (Portunus Trituberculatus). Food Chem. 354, 129570. doi: 10.1016/j.foodchem.2021.129570
Zhao R. Z., Jiang S., Zhang L., Yu Z. B. (2019). Mitochondrial Electron Transport Chain, ROS Generation and Uncoupling. Int. J. Mol. Med. 44, 3–15. doi: 10.3892/ijmm.2019.4188
Zorova L. D., Popkov V. A., Plotnikov E. Y., Silachev D. N., Pevzner I. B., Jankauskas S. S., et al. (2018). Mitochondrial Membrane Potential. Anal. Biochem. 552, 50–59. doi: 10.1016/j.ab.2017.07.009
Zuo R. T., Ai Q. H., Mai K. S., Xu W. (2013). Effects of Conjugated Linoleic Acid on Growth, non-Specific Immunity, Antioxidant Capacity, Lipid Deposition and Related Gene Expression in Juvenile Large Yellow Croaker (Larmichthys Crocea) Fed Soyabean Oil-Based Diets. Br. J. Nutr. 110, 1220–1232. doi: 10.1017/S0007114513000378
Keywords: krill oil, n-3 PUFA, mitochondrial metabolic adaptation, oxidative stress, Portunus trituberculatus
Citation: Yuan Y, Jin M, Fang F, Tocher DR, Betancor MB, Jiao L, Hong Y and Zhou Q (2022) New Insight Into the Molting and Growth in Crustaceans: Regulation of Energy Homeostasis Through the Lipid Nutrition. Front. Mar. Sci. 9:914590. doi: 10.3389/fmars.2022.914590
Received: 07 April 2022; Accepted: 27 May 2022;
Published: 30 June 2022.
Edited by:
Ivan Viegas, University of Coimbra, PortugalReviewed by:
Houguo Xu, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences (CAFS), ChinaSonglin Li, Shanghai Ocean University, China
Copyright © 2022 Yuan, Jin, Fang, Tocher, Betancor, Jiao, Hong and Zhou. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Min Jin, amlubWluQG5idS5lZHUuY24=; Qicun Zhou, emhvdXFpY3VuQG5idS5lZHUuY24=