 Ainara Ballesteros1*
Ainara Ballesteros1* Diego Páez2
Diego Páez2 Andreu Santín1Alba García2Yeray Martín2Ester Alonso2Eric Jourdan3Josep-Maria Gili1
Andreu Santín1Alba García2Yeray Martín2Ester Alonso2Eric Jourdan3Josep-Maria Gili1- 1Department of Marine Biology and Oceanography, Institute of Marine Sciences, Spanish National Research Council (ICM-CSIC), Barcelona, Spain
- 2Loro Parque, Santa Cruz de Tenerife, Spain
- 3ISDIN, Innovation and Development, Barcelona, Spain
The scyphozoan Pelagia noctiluca has potential value for research and a wide variety of industrial applications. However, its holoplanktonic life cycle makes it difficult to maintain a permanent stock in aquaculture, unlike the continuous supply of newly-released ephyrae from jellyfish species with a benthic life stage. We investigated optimal feeding conditions for the captive breeding of P. noctiluca, monitoring total body diameter, morphological development, survival rate, and instantaneous growth rate. Diets of different items encapsulated in gelatin or non-motile prey were not suitable for successful growth of the early stages (ephyra and metaephyra), whereas rotifers were a sufficient food source for healthy growth. Fertilized eggs from sexually-mature jellyfish were obtained at 84.5 ± 2.1 and 87.0 ± 9.9 days post-fecundation in the presence and absence of gelatinous prey (Aurelia sp.), respectively. In contrast with the general consensus, we demonstrate that the incorporation of medusivorous prey was not essential for proper growth and closure of P. noctiluca life cycle. Instead, we recommend the inclusion of white shrimps (Acetes sp.) for the formation and maturation of gonads and production of eggs. Instantaneous growth rates (57.0% d-1), the highest reported to date for this species in captive conditions, were higher during the early stages. A third generation of cultured P. noctiluca was obtained for the first time (169 – 179 experimental days), demonstrating the durability of the culture, providing permanent stock and halving the production time of mature individuals compared to previous studies. Finally, we validated the use of captive-bred specimens through quantification of the hemolytic area after the sting process using the tentacle skin blood agarose assay methodology.
1 Introduction
Since marine bioprospecting began, the world’s oceans and their ecosystems have been considered valuable sources to explore for new natural products with a variety of biotechnological applications (Rocha et al., 2011; Merquiol et al., 2019). Although sponges are the foremost providers of marine bioactive compounds, animals belonging to the phylum Cnidaria (such as jellyfish, sea anemones and corals) are also promising reservoirs of compounds with potential drug activity (Oliveira et al., 2018; Merquiol et al., 2019).
Most of the bioactive compounds are extracted from cnidarians derived from benthic species, while a limited number come from pelagic cnidarians such as scyphozoan jellyfish (Rocha et al., 2011). Although the full exploitation of jellyfish is still in its early stages, recent efforts have investigated the use of scyphozoan jellyfish as a natural source of collagen, fatty acids and bioactive compounds mainly extracted from the venom stored in their cnidocytes (Frazão et al., 2017; Merquiol et al., 2019; Duarte et al., 2021).
Jellyfish blooms may have been increasing in frequency and magnitude in some areas of the world (Brotz et al., 2012) but the scarcity of historical datasets covering large temporal and spatial scales makes it difficult to determine the biomass increase on a global scale (Marambio et al., 2021). Moreover, the variability of environmental conditions and the seasonality of most species do not guarantee their availability when needed (Purcell et al., 2013). Under this premise, wild jellyfish can be a useful resource, but their production by means of aquaculture techniques is key to guarantee a regular supply for research, aquarium exhibitions, stock enhancement for fisheries and successful commercial exploitation (Dong et al., 2009; Purcell et al., 2013; Duarte et al., 2021).
The mauve stinger Pelagia noctiluca is an oceanic jellyfish, widely distributed in the open waters of the Mediterranean Sea (Mariottini et al., 2008; Canepa et al., 2014; Marambio et al., 2021). The lack of a polyp phase, a peculiarity within its life cycle (Sandrini and Avian, 1983; Helm, 2018; Ballesteros et al., 2021b), allows it to inhabit oceanic as well as coastal ecosystems (Canepa et al., 2014). This scyphozoan is considered the most important jellyfish in the Mediterranean basin; its massive outbreaks and the high toxicity of its venom (Mariottini et al., 2008) lead to serious negative effects on tourism, fisheries, aquaculture, and the energy industry (Canepa et al., 2014; Bosch-Belmar et al., 2020).
Pelagia noctiluca is an ideal subject for research in biomedical, cosmetic, and pharmacological industries due to the potential value of the bioactive compounds from toxins identified in its venom (Ayed et al., 2012; Ayed et al., 2016; Frazão et al., 2017). The crude venom, stored in 4 nematocyst types (a-isorhiza, A-isorhiza, O-isorhiza and eurytele) (Ballesteros et al., 2021b), has hemolytic, cytotoxic, dermonecrotic and local tissue damage activity (D’Ambra and Lauritano, 2020). Its proteome, including not only the venom from its nematocysts, but also tissue proteins, revealed 68 different proteins (Frazão et al., 2017), with a wide range of biomedical implications in fields such as oncology, diabetes, HIV, immunology, as well as cardiovascular, musculoskeletal, neurological and hematological disorders (Ayed et al., 2012; Ayed et al., 2016; Frazão et al., 2017). Red fluorescent protein, a protein with high biotechnological importance in molecular biology and previously reported in anthozoans (Verkhusha and Lukyanov, 2004), was first discovered in scyphozoans in P. noctiluca, as was peroxiredoxin, a potential natural source of antioxidants and anti-UV radiation agents (Frazão et al., 2017).
The sting of P. noctiluca is painful and inconvenient for beach users and represents a high proportion of cases of those seeking help for jellyfish stings on the beaches of the Mediterranean Sea (Mariottini et al., 2008; De Donno et al., 2014). Although it is not considered a life-threatening species, its severe sting generates edema, burning sensation, vesicles and/or scabs immediately after inoculation of the venom (Montgomery et al., 2016; Hall et al., 2018). Hence, the cosmetic and pharmaceutical industries research and sell products to prevent and ameliorate its sting (Hall et al., 2018; Morabito et al., 2020).
Pelagia noctiluca is present all year in the Mediterranean Sea in diverse stages of its life cycle, and reproductive medusae appear in different periods in the basin (Rosa et al., 2013; Canepa et al., 2014; Milisenda et al., 2018a). Its oceanic distribution, vertical migrations, and spatiotemporal variability along the coast (Marambio et al., 2021; Pastor-Prieto et al., 2021) make sampling a difficult task. Moreover, many individuals stranded on the beaches are not in optimal conditions to be used for research or commercial purposes. Under these premises, and expanding its use to a larger and more sustainable scale, it is essential to establish P. noctiluca cultures. The holoplanktonic life cycle of P. noctiluca, which lacks a benthic stage (Sandrini and Avian, 1983; Ballesteros et al., 2021b), has limited scope for research in captive conditions. Maintaining its culture presents a host of challenges (Ramondenc et al., 2019) not only for academic and industrial research but also for aquarium exhibitions. Ephyrae released from polyp colonies through the strobilation process (Helm, 2018) represent a continuous source of individuals, an advantage that has allowed improvements in husbandry techniques, for example with the well-known moon jellyfish Aurelia spp. (Schaadt et al., 2017) and the upside-down jellyfish Cassiopea spp. (Pierce, 2005). To our knowledge, only two studies have shown long-term culture of P. noctiluca, but none of them managed to reproduce the second generation to ensure durability of the culture (Lilley et al., 2014; Ramondenc et al., 2019).
Considering this approach, and the great value of P. noctiluca in aquarium exhibitions, research and industrial applications, the objectives of this research were: (I) to determine the optimal food regimen for culture, (II) to obtain a third generation of P. noctiluca to guarantee durability of the culture, and (III) to validate the use of captive-bred P. noctiluca specimens by evaluating the hemolytic area after a sting event. In this study, we halved the production time of mature P. noctiluca individuals, acquiring a third generation in the presence and absence of gelatinous prey. The use of captive-bred P. noctiluca specimens for future venom experiments using the tentacle skin blood agarose assay was validated. The results represent an important contribution to the improvement of nutrition, production time and breeding techniques in aquaculture of the most important jellyfish in the Mediterranean Sea.
2 Material and Methods
2.1 Breeding of Pelagia noctiluca in Captivity
2.1.1 Culture Establishment
Mucus strips with fertilized eggs were collected at ∼ 11:00 AM from a mature jellyfish aquarium in Loro Parque Aquarium (Tenerife, Spain), 3 hours after the photoperiod was switched on (post-fecundation day 0). The fertilized eggs were transferred to a plastic container with 8 L of natural filtered seawater (34–36 salinity) and abundant aeration (Figure 1A) placed in a water bath (22°C) and using a wide plastic pipette. 22°C of temperature was chosen for being the water inlet temperature from the ocean to the Loro Parque Aquarium during the period of the experiments. Within 3 days, the eggs metamorphosed into the ephyra stage and the experiments could be started (post-fecundation day 3).
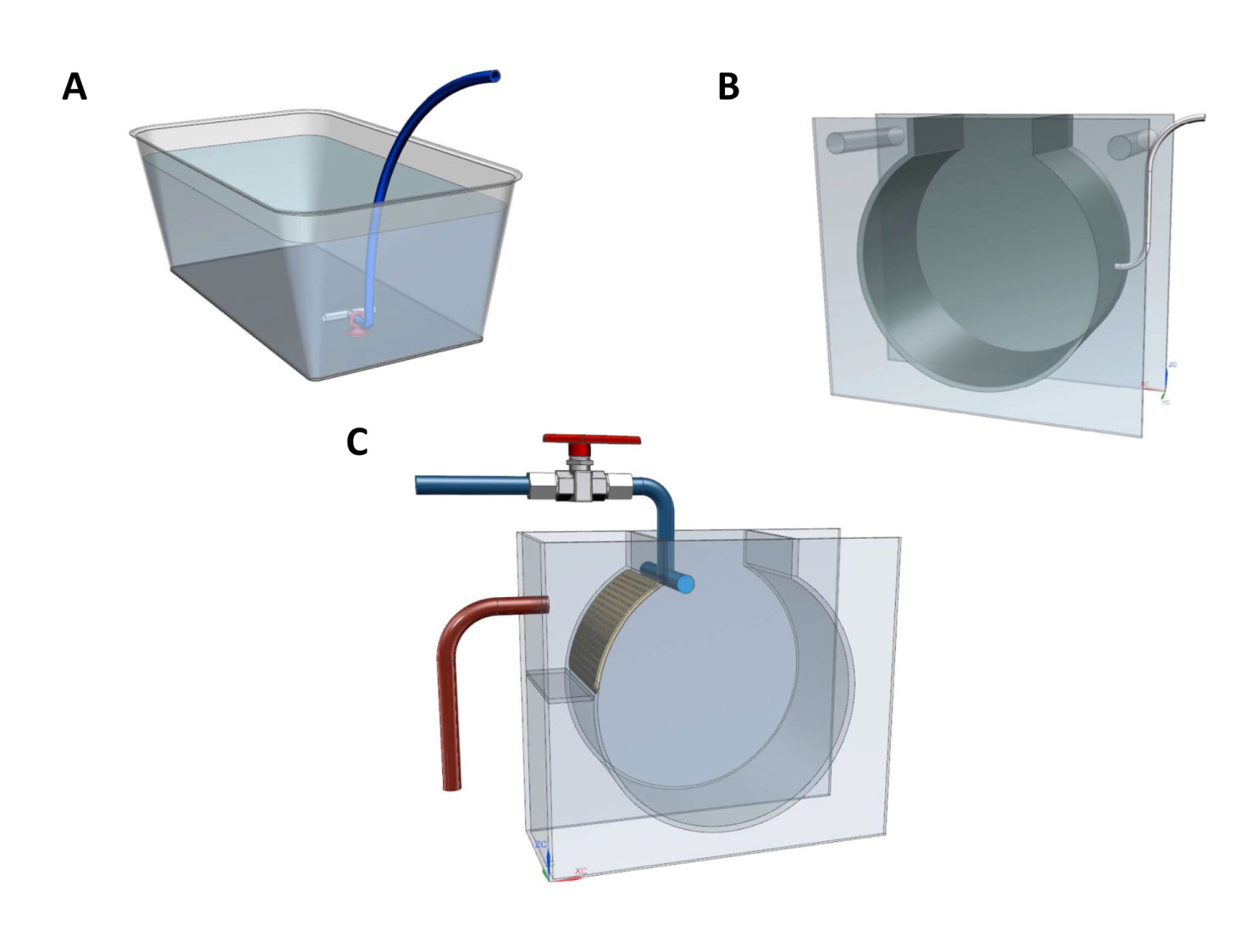
Figure 1 Types of rearing vessels and tanks used for Pelagia noctiluca during the experiments. (A) Culture establishment from fertilized eggs to first ephyrae. (B) Air-kreisel tank for the farming of early stages. (C) Water-kreisel tank for the breeding of advanced stages. Illustrations by Carlos Mengod.
2.1.2 Maintenance and Feeding
Early stages. Four groups of 150 randomly-selected ephyrae, all from the same fecundation event, were transferred to four air-kreisel tanks (Figure 1B) with 12 L of natural filtered seawater (34–36 salinity), which were placed in a water bath to reach 22°C. The kreisel aquarium is a well-known tank designed to keep gelatinous zooplankton (Purcell et al., 2013). The air inlet, helped by its circular shape, creates a continuous circular water flow, allowing animals to remain in the water column. The light cycle was 12h light/12h dark. The early stages were cultivated in a closed water system. Every day, 15% of the seawater was renewed. Every three days, a complete deep cleaning was performed coinciding with the renewal of 100% of the seawater to ensure optimal culture conditions (e.g., pH and nitrogen and oxygen concentrations).
Dispensing with additional cultures such as Brachionus spp. (rotifers) or Artemia spp., helps optimize time in captive husbandry. Under this premise, we used two diets that did not involve additional cultures: Culture 1 (gelatin) was encapsulated food, made by us, inside a neutral gelatin (Promolac). It was composed of razor shell (Ensis directus), shrimp (Neomysis sp.), sardine (Sardina pilchardus), mussel (Mytilus galloprovincialis), hake (Merluccius hubbsi), pacific krill (Euphausia pacifica, Ocean Nutrition™) and red plankton (Calanus finmarchicus, Ocean Nutrition™). Culture 2 (non-motile prey) was formed of frozen Artemia sp. nauplii and codfish eggs (Gadus morhua). Amounts of food and feeding times used for each experimental week can be consulted in Supplementary Material Tables 1, 2.
Gelatinous zooplankton, such as the scyphozoan Aurelia spp. and Salpa democratica, seem to be important in the farming of medusivorous jellyfish such as P. noctiluca (Purcell et al., 2013). Two more types of food regimens were added. Culture 3 (medusivorous) was based on the presence of mucus and small pieces of Aurelia sp. as an addition to rotifers; both came from Loro Parque Aquarium cultures. Culture 4 (non-medusivorous) contained only rotifers. Amounts of food and feeding times used for each experimental week can be found in Supplementary Material Tables 1, 2.
Advanced stages. The optimal cultures, at day 31 of the experiments, were considered juvenile medusae and they were easily recognized for their typical brown coloration – detailed morphological descriptions can be consulted in Ballesteros et al. (2021b) –. Juvenile medusae were transferred to a water-kreisel tank (190 L) with a direct inlet of filtered natural seawater (34–36 salinity and 22°C temperature) (Figure 1C). The light cycle was 12h light/12h dark. The cultures were deep cleaned on a weekly basis.
Culture 3 (medusivorous) was separated into two cultures. The first, culture 3 (medusivorous), continued the research line in the presence of pieces of Aurelia sp. along with Artemia sp. nauplii, codfish eggs, encapsulated food in gelatin (described above) and frozen white shrimp (Acetes sp.). The other, culture 3.1 (gelatin), was fed only with encapsulated food in gelatin. Finally, culture 4 (non-medusivorous), with no Aurelia sp., was fed with Artemia sp. nauplii, codfish eggs, encapsulated food in gelatin and white shrimp. Amounts of food, feeding and times for each experimental week can be consulted in Supplementary Material Tables 1, 2.
2.1.3 Morphological Descriptions, Measurements, Statistical Analysis and Graphical Representation
To describe and monitor the growth of the jellyfish on each food regimen, each week, 10 living individuals were randomly collected from each culture. Ephyrae and metaephyrae were relaxed with a menthol solution (4% w/v), transferred into a ventral position on a glass slide and photographed using a camera (Imaging Source) attached to a binocular loupe (ZUZI 220). Afterwards, the individuals were carefully returned to their respective cultures. The morphology and the total body diameter (TBD) were described and measured (from lappet to lappet) as in Ballesteros et al. (2021b) by photo-analysis with the Fiji version of ImageJ software (Schindelin et al., 2012). For advanced stages, jellyfish were transferred to a plastic tray with a small amount of seawater. TBD was measured directly with a digital caliper with the animals in a ventral position. As in the early stages, individuals were carefully returned to each culture. A total of 602 individuals were measured and analyzed during the research.
The growth of P. noctiluca (TBD; mm) over time (days post-fecundation) was represented graphically by means of boxplots using the R software platform (R Core Team, 2017). Instantaneous growth rate (µ; % d-1) was estimated as in Lilley et al. (2014). TBD measures were converted into carbon weight (CW) according to the formula (Lilley et al., 2014):
and µ was calculated as:
The differences between µ for successful food regimens were calculated using dynamic time warping (DTW). DTW provides a similarity score or distance between two sequences by means of the following formula (Giorgino, 2009):
where M is a matrix of X and Y series, i is the iterator for series X and j is the iterator for series Y.
A DTW distance of zero implies that the compared series are almost identical, whereas high distance values imply dissimilarities between the compared series (Giorgino, 2009). This was performed using the dtw package available in the R software platform (R Core Team, 2017).
Finally, during the deep cleaning of the systems, every three days for the early stages and once a week for the advanced stages, individuals were counted to calculate the survival rate (%):
where Nt is the individual number at that time point and N0 is the number of individuals at the beginning of the culture.
2.2 Validation of the Hemolytic Activity of the Venom
In order to be able to use P. noctiluca bred in captivity in future toxicity tests, the hemolytic property of wild and captivity jellyfish venoms was compared using the tentacle skin blood agarose assay (TSBAA) (Yanagihara et al., 2016).
2.2.1 Origin of Jellyfish
Mature jellyfish from the open sea were collected in 2019 in Menorca (Spain) using hand nets and plastic jars. The individuals were then transferred to plastic bags full of seawater, avoiding air bubbles to ensure good conditions during transport. For the captive jellyfish, we used mature jellyfish from culture 3 (medusivorous), once the third generation was achieved. The culture 3 (medusivorous) was chosen because it represented a traditional diet in presence of gelatinous prey (Aurelia sp.).
2.2.2 Tentacle Skin Blood Agarose Assay
The venom load was evaluated by the hemolytic area in a modified protocol adapted from the TSBAA method (Yanagihara et al., 2016). Briefly, an agarose gel preparation incorporating sheep red blood cells (SRBCs) (Thermo Fisher Scientific) was used, covered by a thin tissue of pig small intestine to simulate the effect of the human skin barrier (Ballesteros et al., 2021a).
Cutting tentacles to the same size is a complex process due to their contraction and expansion after manipulation. With the aim of improving the methodology and standardizing the sting area, we designed a plastic mold (Supplementary Material Figure 1). These plastic molds were placed on top of the agarose rectangles with pig intestine. Consecutively, the sting process was carried out with tentacles from wild and captive jellyfish for 10 min. The sting diameter was 57.4 mm2. A total of 8 replicates for wild jellyfish and 10 for captive jellyfish were performed. After this natural-method sting event, the plastic molds and the intestine sections were removed and the SRBC agarose rectangles were stored in the humidification chamber at room temperature.
2.2.3 Hemolytic Areas, Statistical Analysis and Graphical Representation
After a period of 22 hours from the sting process, images of the hemolytic areas were obtained with a camera. The data was tested for normality and homogeneity using the stats package available for the R software platform (R Core Team, 2017). As data did not display homogenous variances, a Mann-Whitney test was applied by means of the wilcox.test function to test if significant differences existed between hemolytic areas. Finally, a graphical representation was performed using the ggplot2 package (Wickham, 2016) from the R software platform (R Core Team, 2017).
3 Results
3.1 Morphology, Total Body Diameter and Survival Rate
3.1.1 First Generation
Culture 1 (gelatin). Early stages. Feeding with gelatin was not satisfactory for early stages. The culture was maintained only up to 18 days post-fecundation with a 0% survival rate (Figures 2, 3A and Table 1). No gelatin pieces were observed in the ephyrae gastric system. Throughout the culture there was an absence of gastric filaments, velar canals, and nematocyst batteries. Additional information is presented in Supplementary Material Table 1.
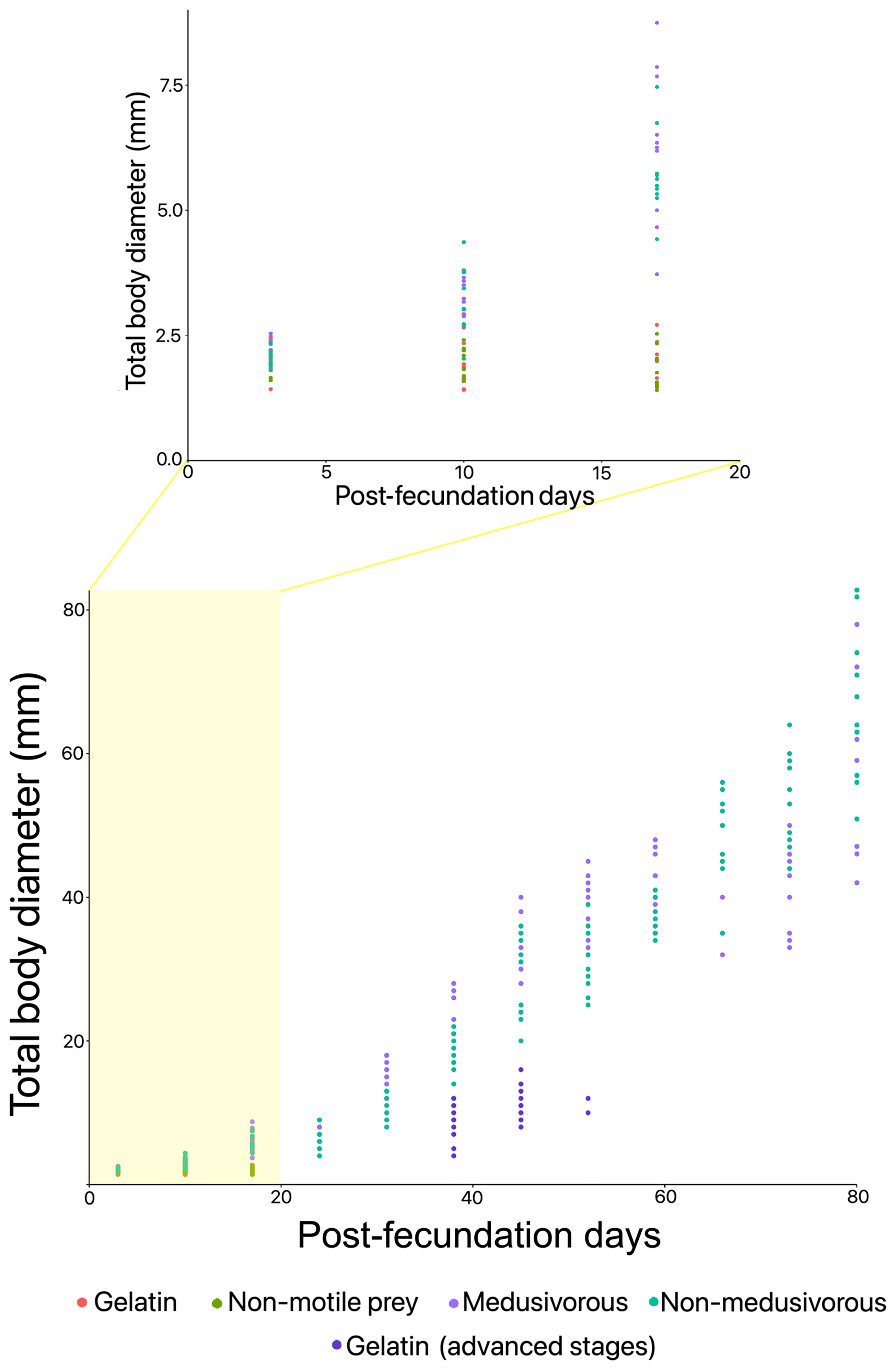
Figure 2 Total body diameter over time in the presence of different food regimens for the first generation of Pelagia noctiluca. Food regimens: Culture 1, encapsulated food in gelatin; Culture 2, non-motile prey; Culture 3, medusivorous diet (presence of Aurelia sp.); Culture 3.1, medusivorous diet during early stages and gelatin from advanced stages; and Culture 4, non-medusivorous diet (absence of Aurelia sp.).
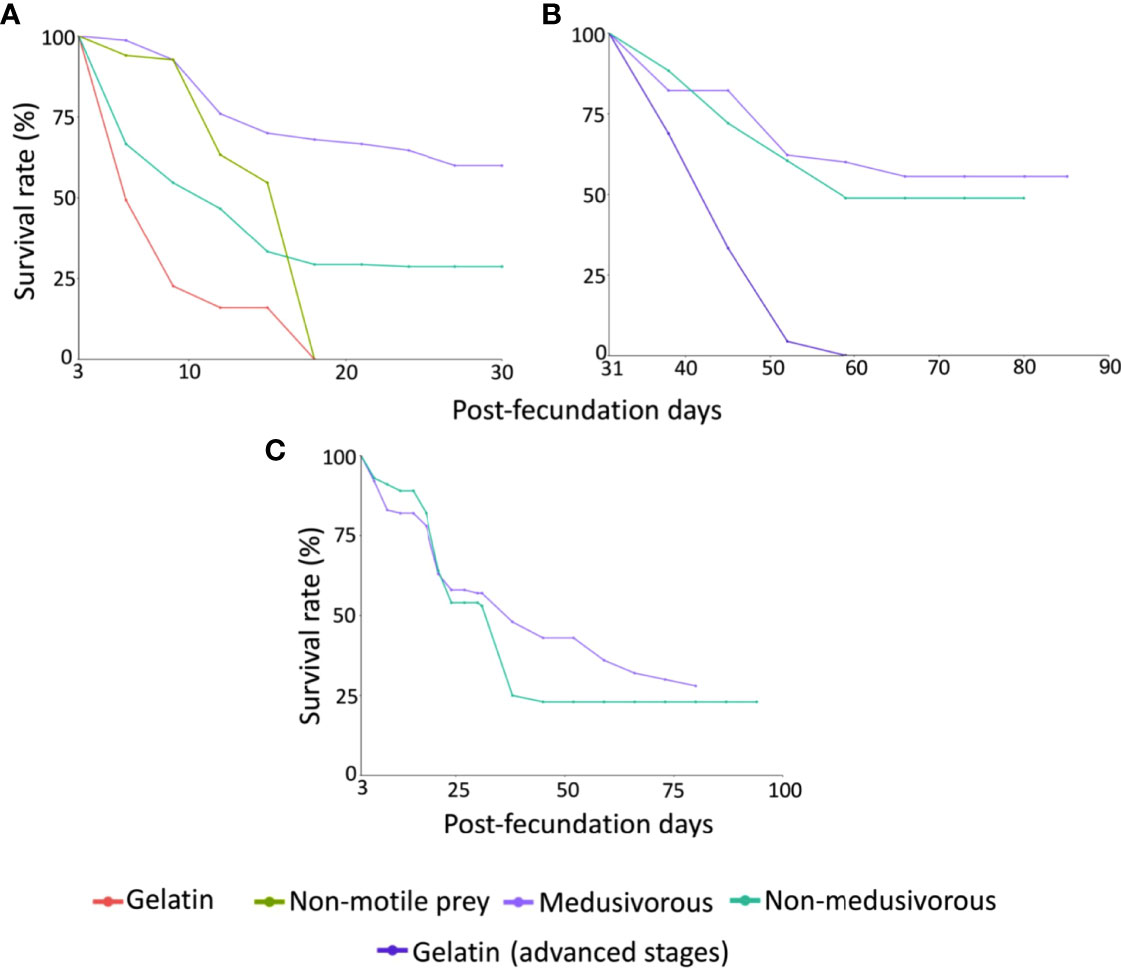
Figure 3 Survival rates (%) of Pelagia noctiluca for: (A) Early stages of the first generation (ephyra and metaephyra stages), (B) Advanced stages of the first generation (juvenile and adult stages), (C) Early and advanced stages of the second generation. Food regimens: Culture 1, encapsulated food in gelatin; Culture 2, non-motile prey; Culture 3, medusivorous diet (presence of Aurelia sp.); Culture 3.1, medusivorous diet during early stages and gelatin from advanced stages; and Culture 4, non-medusivorous diet (absence of Aurelia sp.).
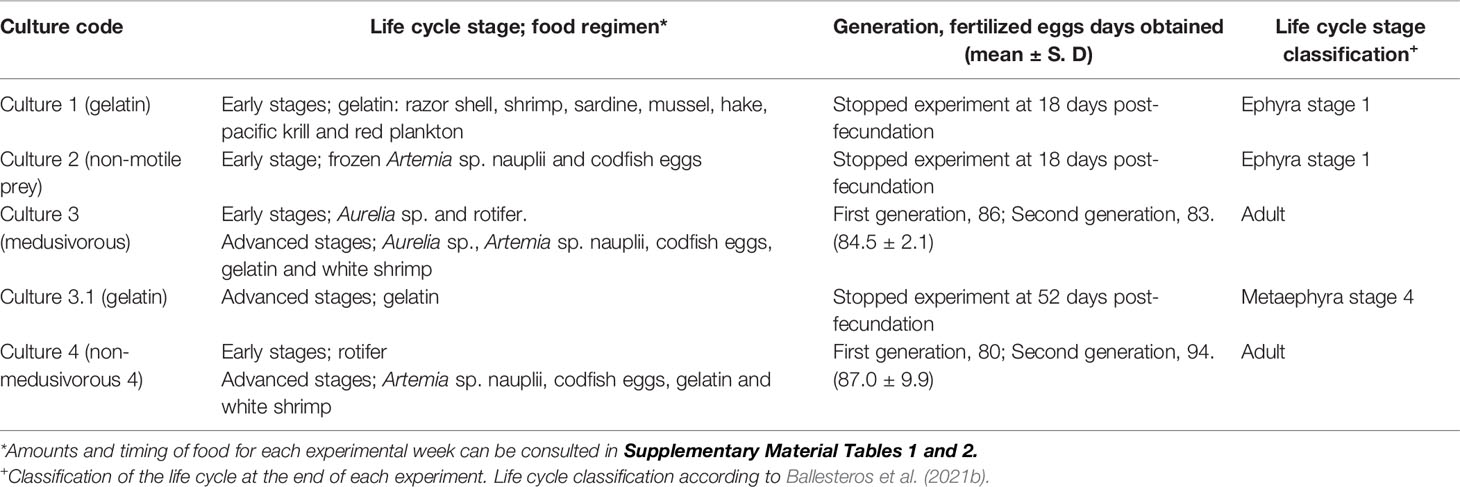
Table 1 Food regimens according to the life cycle stages, post-fecundation days and life cycle classification at the end of each experiment.
Culture 2 (non-motile prey). Early stages. A food regimen based on non-motile prey was insufficient for growth during the early stages. As with culture 1 (gelatin), this culture was stopped on the 18th day post-fecundation with a survival rate of 0% due to the absence of gastric filaments, velar canals, and nematocyst batteries (Figures 2, 3A and Table 1). Additional information can be consulted in Supplementary Material Table 1.
Culture 3 (medusivorous). Early and advanced stages. First gastric filaments and velar canals were observed on the 10th day post-fecundation (mean TBD for this culture was 3.3 ± 0.3 mm). Tentacle buds or incipient tentacles and nematocyst batteries appeared on the 17th day after fecundation (mean TBD 6.3 ± 1.5 mm). Marginal tentacles fully developed in some individuals at the 24th day post-fecundation (mean TBD of 6.4 ± 1.5 mm) with a 64.7% survival rate (Figures 2, 3A). Culture 3 was split at 31 days post-fecundation into two cultures, named culture 3 (medusivorous) and culture 3.1 (gelatin) (Figure 2), each with 45 individuals. Culture 3 (medusivorous), which included culture 3 early stages, was maintained for a period of 86 days post-fecundation. Gonad formation began 52 days after the fecundation event (mean TBD for this culture, 38.6 ± 3.9 mm), and incorporation of white shrimp as an additional protein was essential (Supplementary Material Table 1). Fertilized eggs from 25 mature jellyfish were observed at ~ 11:00 AM, closing the P. noctiluca life cycle in laboratory conditions at 86 days post-fecundation (survival rate of 55.6%) (Figure 3B and Table 1). Eggs were observed daily. Ephyrae of the second generation were obtained 3 days after egg production. Additional information is presented in Supplementary Material Table 1.
Culture 3.1 (gelatin). Advanced stages. 45 juvenile medusae (survival rate of 100%) (Figure 3B) from culture 3 (medusivorous) early stages were fed only with pieces of gelatin from the 31st day post-fecundation. Unfortunately, gelatin alone was also not a suitable food for the advanced stages. Young medusae did not grow. They were deformed and lost their typical juvenile brownish coloration. The culture was stopped on the 52nd day post-fecundation (survival rate of 4.4%) with a mean TBD of 11.4 ± 1.4 mm, compared with culture 3 (medusivorous) which had a mean TBD of 38.6 ± 3.9 (survival rate of 62.2%) and was formed of healthy juvenile medusae that produced gonads (Figures 2, 3B and Table 1). Additional information can be consulted in Supplementary Material Table 1.
Culture 4 (non-medusivorous). Early and advanced stages. The culture was maintained for a period of 83 days post-fecundation. As in culture 3 (medusivorous), the first gastric filaments and velar canals were observed at the 10th day post-fecundation (mean TBD of 3.2 ± 0.7 mm). Tentacle buds or incipient marginal tentacles and nematocyst batteries appeared at 17 days post-fecundation (mean TBD of 5.7 ± 0.8 mm). Marginal tentacles fully developed in some metaephyrae at 24 days after fecundation with a survival rate of 28.7% (mean TBD of 6.3 ± 1.3 mm) (Figures 2, 3B). Rotifers as a single food source were sufficient for normal development in each early stage. Gonad formation started 52 days after fecundation (mean TBD of 31.2 ± 4.5 mm). As in culture 3 (medusivorous), white shrimps were added to the diet as a rich source of protein. Fertilized eggs were observed from 21 mature jellyfish, on the 83rd day post-fecundation, at ~ 11:00 AM (Table 1). The life cycle was closed in laboratory conditions without the presence of medusae in the food regimen. Release of eggs and sperm occurred daily. Eggs metamorphosed into ephyrae (second generation) 3 days after egg production. Additional information is presented in Supplementary Material Table 1.
3.1.2 Second Generation
Culture 3 (medusivorous). Early and advanced stages. The culture was maintained for a period of 80 days (Table 1). First gastric filaments and velar canals were observed on the 10th day post-fecundation (mean TBD of 4.0 ± 0.6 mm). As in the first generation, tentacle buds or incipient marginal tentacles and nematocyst batteries were observed at 17 days after fecundation (mean TBD of 9.4 ± 1.6 mm). Marginal tentacles were fully developed at the 24th day post-fecundation (mean TBD of 12.3 ± 1.2 mm). Gonad formation occurred at 52 days after fecundation (mean TBD of 39.6 ± 6.3 mm). Fertilized eggs were collected on the 80th day post-fecundation with 28 jellyfish (survival rate of 28%) (Figure 3C and Table 1). Eggs metamorphosed into ephyrae (third generation) 3 days after fecundation, closing the second generation. Additional information is presented in Supplementary Material Table 2.
Culture 4 (non-medusivorous). Early and advanced stages. The culture was maintained until 94 days post-fecundation. First gastric filaments and velar canals appeared on the 10th day post-fecundation (mean TBD of 5.1 ± 0.4 mm). Tentacle buds or incipient marginal tentacles and nematocyst batteries were observed at 17 days after fecundation (mean TBD of 9.2 ± 0.8 mm). Marginal tentacles were fully developed at the 24th day post-fecundation (mean TBD of 10.1 ± 1.3 mm). Gonad formation occurred at 66 days after fecundation (mean TBD of 43.0 ± 8.9 mm), and again, white shrimps were incorporated in the diet. Fertilized eggs were collected on the 94th day post-fecundation from 23 jellyfish (survival rate of 23%) (Figure 3C and Table 1). Eggs metamorphosed into ephyrae (third generation) 3 days after fecundation, closing the second generation. Additional information can be consulted in Supplementary Material Table 2.
3.1.3 Third Generation
Culture 3 (medusivorous). The experiment was stopped after the observation of a healthy third generation of ephyrae (stage 1) with a mean TBD of 3.1 ± 0.2 mm, demonstrating the durability of the culture. The ephyrae were obtained after a total of 169 experimental days (Figures 4, 5). Additional information can be consulted in Supplementary Material Table 2.
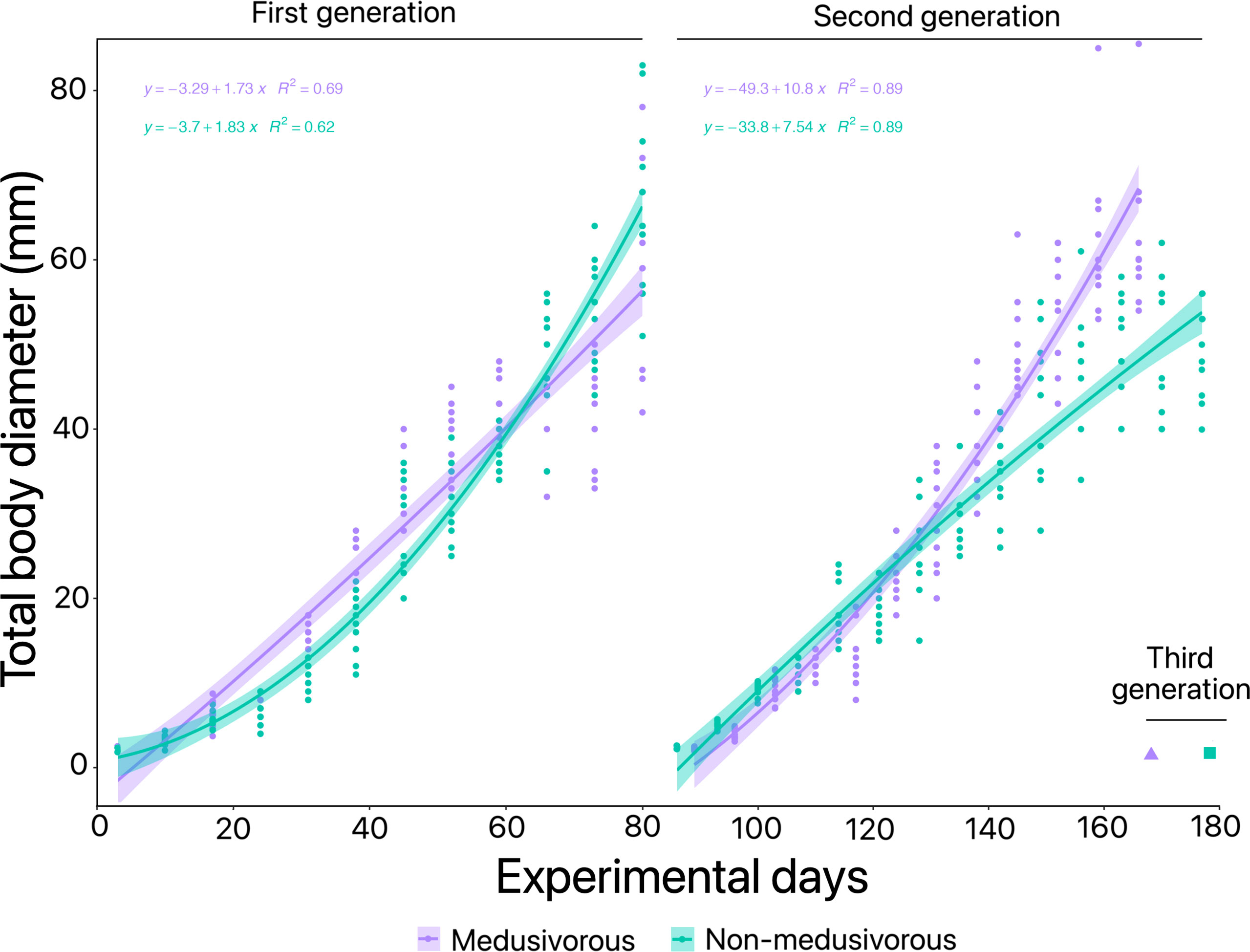
Figure 4 Optimization of Pelagia noctiluca farming in the presence and absence of medusivorous prey. The three generations obtained from the beginning of the experiments are included. Culture 3, medusivorous diet (presence of Aurelia sp.) and Culture 4, non-medusivorous diet (absence of Aurelia sp.).
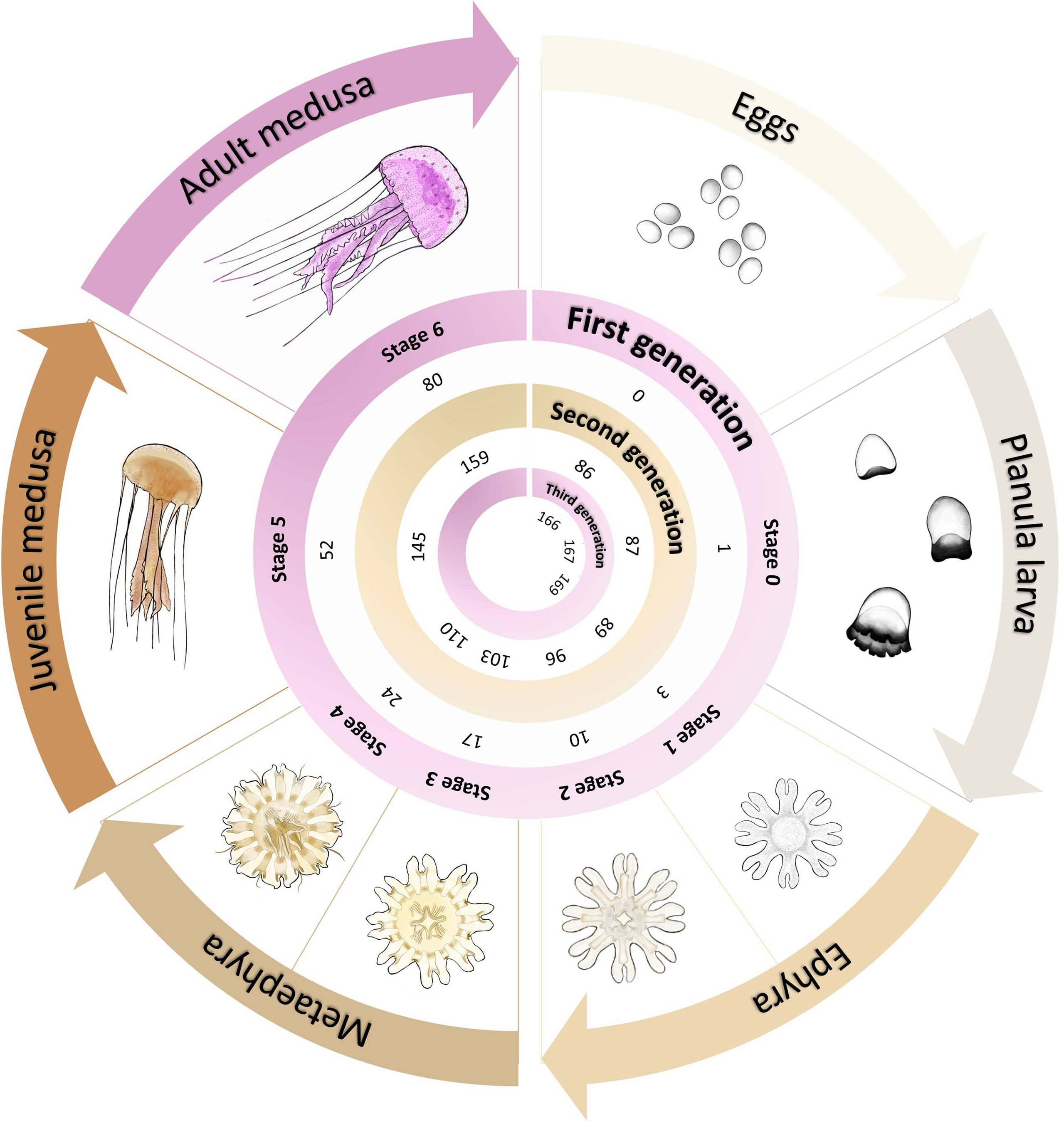
Figure 5 Graphical representation of the Pelagia noctiluca life cycle optimization at 22°C. It includes the post-fecundation days until each stage of its life cycle is reached, when fed with the medusivorous diet (culture 3). Detailed morphological and cnidome descriptions for each life cycle stage can be consulted in Ballesteros et al. (2021b). Illustrations of the life cycle by Lau López.
Culture 4 (non-medusivorous). The third generation of healthy ephyrae (stage 1), with a mean TBD of 2.7 ± 0.3 mm, was obtained after a total of 179 experimental days (Figure 4). As in culture 3 (medusivorous), the experiment was then stopped. Additional information is presented in Supplementary Material Table 2.
3.2 Instantaneous Growth Rate
For the first generation, growth rates of P. noctiluca varied depending on the feeding regime (Supplementary Material Table 1). Changes in carbon weight in culture 1 (gelatin), culture 2 (non-motile prey) and advanced stages of culture 3.1 (gelatin) reflected the lack of growth and development of the gastric cavity (Figure 2 and Supplementary Material Table 1). The growth rates with the successful feeding regimens in culture 3 (medusivorous) and culture 4 (non-medusivorous) were predominantly positive, with greater rates during the early stages of the life cycle (Figure 6A). Considering only positive changes in carbon weight, early stages grew up to 57.0% d-1 for culture 3 (medusivorous) and 48.9% d-1 for culture 4 (non-medusivorous). The instantaneous growth rates for advanced stages reached 33.0% d-1 for culture 3 (medusivorous) and 51.5% d-1 for culture 4 (Figure 6A). Additional information is presented in Supplementary Material Table 1.
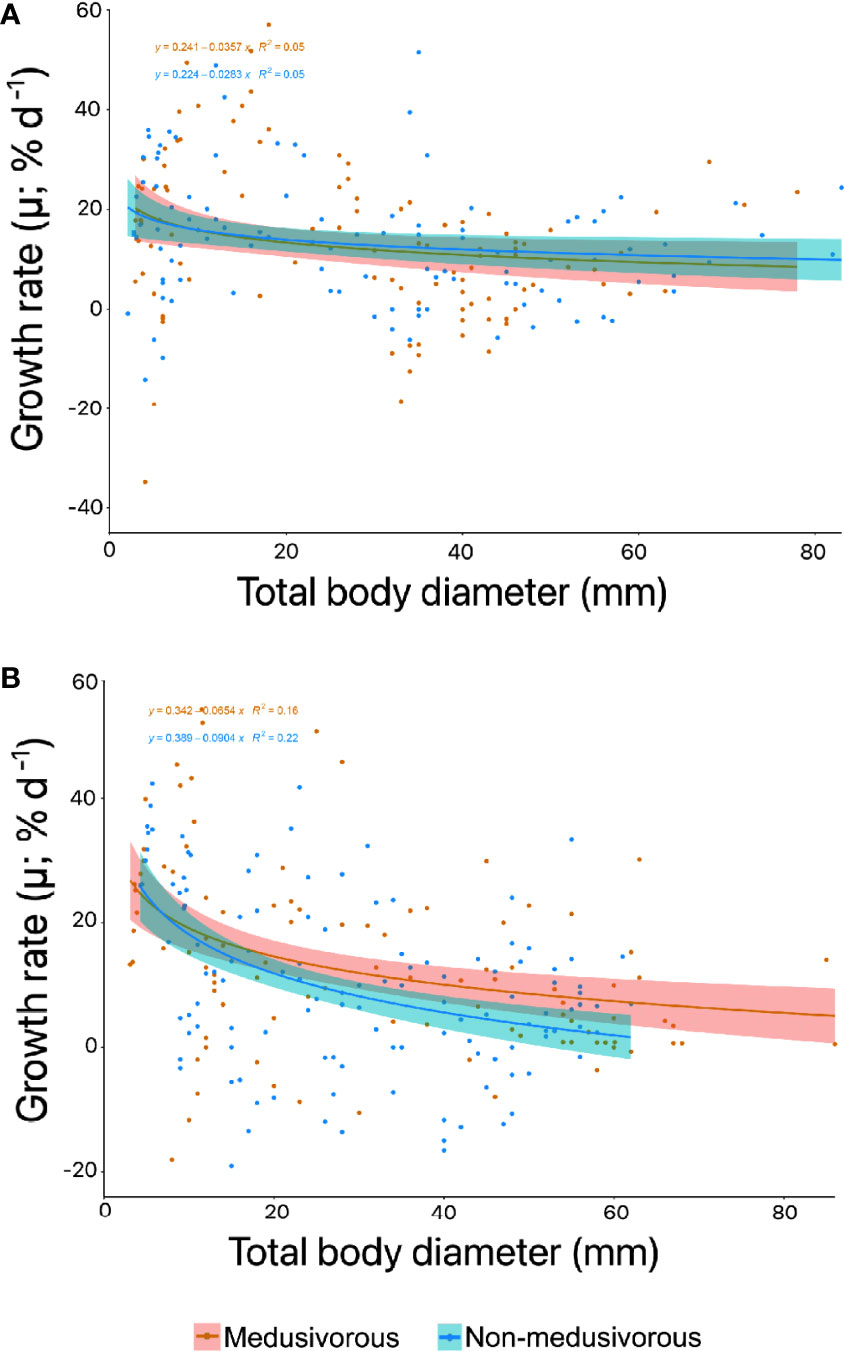
Figure 6 Instantaneous growth rate (% d-1) for the successful food regimens for Pelagia noctiluca in (A) first generation and (B) second generation. The decrease in growth rate with total body diameter (TBD) (solid line) represents a power relationship according to positive instantaneous growth rates. The shaded area indicates 95% confidence limits. Food regimens: Culture 3, medusivorous diet (presence of Aurelia sp.) and Culture 4, non-medusivorous diet (absence of Aurelia sp.).
The trends in instantaneous growth in the second generation were very similar to the first generation (Figure 6). Positive instantaneous growth rates predominated, and greater rates corresponded to the early stages of the life cycle (Figure 6B). Early stages grew up to 54.2% d-1 for culture 3 (medusivorous) and 42.4% d-1 for culture 4 (non-medusivorous). Advanced stages reached a maximum growth rate of 50.7% d-1 for culture 3 (medusivorous) and 33.3% d-1 for culture 4 (non-medusivorous) (Figure 6B). Additional information can be consulted in Supplementary Material Table 2.
The comparison of growth rate data between non-medusivorous and medusivorous by means of DTW yielded a score of 0.2.
3.3 Validation of the Use of Captive Individuals in Venom Experiments
In order to validate the use of captive jellyfish in venom-related experiments, the hemolytic areas were evaluated after the sting process from captive-bred and wild jellyfish. The hemolytic area obtained a value of 44.3 ± 3.4% for wild jellyfish and 39.3 ± 10.2% for captive jellyfish (Figure 7). No significant difference (p value 0.2) was observed between wild and captive jellyfish.
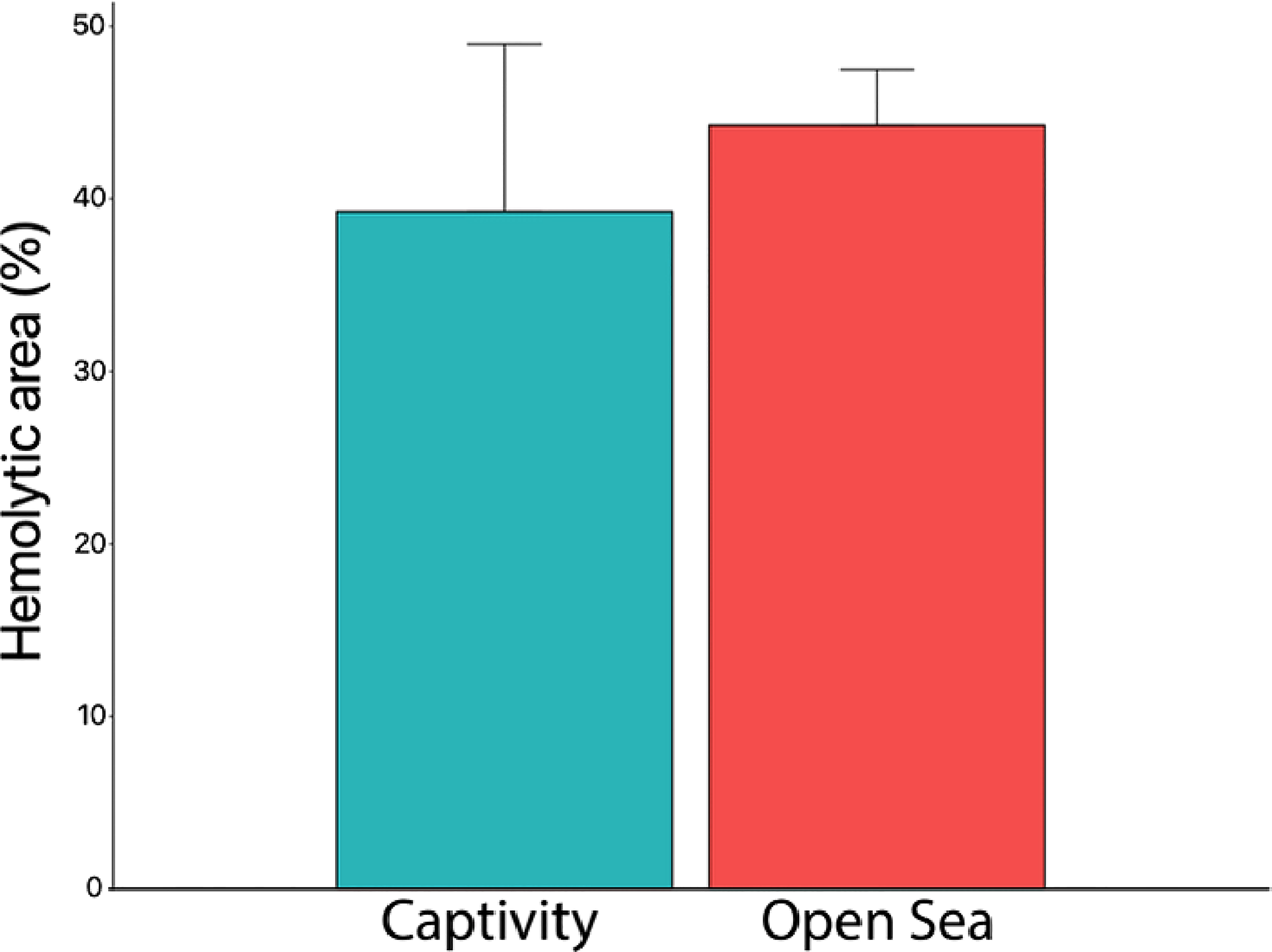
Figure 7 Hemolytic area (%) produced by the venom from captive and open-sea jellyfish. No significant differences were observed between jellyfish from the open sea and those from captivity (p value 0.2) (Mann–Whitney test). Captivity replicates, n = 10; open sea replicates, n = 8.
4 Discussion
The present study improves the culture of P. noctiluca. Here, we halved the production time of mature individuals and doubled the instantaneous growth rates, in the presence and absence of gelatinous prey (Aurelia sp.). This is the first time that a third generation of P. noctiluca has been obtained, guaranteeing its continuous availability, and the first time that the use of captive-bred individuals for venom-related experiments has been validated.
The culture of P. noctiluca is a challenge for most research institutions, aquariums and companies who wish to keep this species in captivity (Purcell et al., 2013; Ramondenc et al., 2019). To date, only two studies have focused on the culture of P. noctiluca over time (Lilley et al., 2014; Ramondenc et al., 2019). Lilley et al. (2014) cultured P. noctiluca eggs through to reproductive adults up to a maximum of ∼ 500 days post-fecundation. Although gamete release was observed, second generation was not achieved. Despite the use of a closed system with artificial seawater, a limitation for the cultivation of jellyfish, Ramondenc et al. (2019) improved on the results by Lilley et al. (2014), obtaining egg production in jellyfish with a mean TBD of ~ 90 mm at 140 days post-fecundation. Comparing previous research (Lilley et al., 2014; Ramondenc et al., 2019) and optimizing the growth time of individuals, our study obtained fertilized eggs in 84.5 ± 2.1 and 87.0 ± 9.9 days post-fecundation in the presence and absence of gelatinous prey, respectively (Figure 4 and Table 1). We halved the time of production of mature P. noctiluca individuals, growing two generations and obtaining a third generation in a period of 169 (medusivorous prey) and 179 (non-medusivorous prey) days (Figures 4, 5). The TBDs we observed in mature individuals were comparable to wild P. noctiluca individuals (Avian, 1986; Milisenda et al., 2018a; Shtewi et al., 2022), and egg production was observed ∼ 3 hours after the light was switched on, as in previous studies (Lilley et al., 2014; Ramondenc et al., 2019).
Exponential somatic growth during the first weeks of development is a typical trend for jellyfish species (Widmer, 2005; Lilley et al., 2014; Ramondenc et al., 2019; Schäfer et al., 2021). Instantaneous growth rates were fastest during early stages and gradually slowed thereafter, when jellyfish invested more energy in the formation and maturation of gonads and egg production than in growth (Figure 6). Lilley et al. (2014) and Ramondenc et al. (2019) obtained growth rates up to 30% d-1 for the first weeks of development. In this study, we observed a maximum of 57.0% d-1 (Figure 6A), the highest values for captive-bred P. noctiluca early stages. Therefore, the satisfactory feeding regimens (Figure 4 and Table 1) accompanied by the maintenance techniques used in the present research (see materials and methods section) represent an improvement in the aquaculture techniques of P. noctiluca.
The development from eggs to ephyrae of P. noctiluca is temperature-dependent, warmer waters favoring its growth (Avian, 1986; Purcell et al., 2013; Rosa et al., 2013). The temperature range for culture is from 9–24°C (Purcell et al., 2013; Duarte et al., 2021), although temperatures around 14°C slowed down the development of planulae (Avian, 1986; Rosa et al., 2013), and previous captivity studies considered 18°C the optimal temperature for experiments (Lilley et al., 2014; Ramondenc et al., 2019). In the present study we observed an improvement in the development time from planulae to ephyrae at 3 days post-fecundation (22°C) vs. the previously-reported 4 days post-fecundation at 18°C (Ramondenc et al., 2019; Ballesteros et al., 2021b). Regarding morphological descriptions, Ramondenc et al. (2019) identified first gastric filaments and marginal tentacles at 18 and 38 post-fecundation days, respectively, at 18°C. In our study, gastric filaments and marginal tentacles appeared at 10 and 24 post-fecundation days in the presence and absence of gelatinous prey, respectively. Similar morphological and cnidome descriptions were obtained in Ballesteros et al. (2021b) during the early stages at 18°C. Considering our results, we recommended a temperature of 22°C to encourage healthy growth (without any visible signs of stress) of P. noctiluca in captivity.
Unlike mixotrophic scyphozoa such as Cotylorhiza tubercula, Phyllorhiza punctata and Cassiopea spp. that obtain part of their nutrition from zooxanthellae (Raskoff et al., 2003; Pierce, 2005; Straehler-Pohl and Jarms, 2010; Crow et al., 2013), P. noctiluca is a heterotroph and opportunistic jellyfish that uses nematocysts to penetrate and entangle prey as an exclusive way of obtaining food (Larson, 1987; Milisenda et al., 2018b; Ballesteros et al., 2021b). The gut content analysis from wild P. noctiluca revealed a wide variety of zooplankton prey (e.g., Copepoda, Cladocera and Siphonophore) (Tilves et al., 2016), which has been provided in captive conditions through its daily collection to reproduce the natural diet (Lilley et al., 2014; Ramondenc et al., 2019). However, rotifers and Artemia spp. nauplii are the most widely used food for jellyfish farming (Raskoff et al., 2003; Crow et al., 2013; Purcell et al., 2013; Schaadt et al., 2017; Duarte et al., 2021). Previous studies identified good survival and ephyrae growth of P. noctiluca with Artemia spp. nauplii and small sea urchin eggs in the diet (Lilley et al., 2014; Ramondenc et al., 2019) while our results suggested rotifers or rotifers plus Aurelia sp. mucus was the best food regimen during early stages (Figures 2–4). The improvements observed in this study in terms of TBD, growth rates, development time of the gastric cavity and survival rates are linked with the ideal size and longevity of the rotifers offered during the early stages. Pelagia noctiluca does not have nematocyst batteries during ephyra stage 1 and 2 (Ballesteros et al., 2021b), thus it is preferable to feed them with smaller prey than Artemia spp. nauplii. The nematocyst batteries are a strategy to capture larger prey, as groups of nematocysts discharge simultaneously and operate more efficiently than a single nematocyst (Östman and Hydman, 1996; Ballesteros et al., 2021b). To avoid Artemia spp. nauplii escaping from young ephyrae of P. noctiluca as previously reported by Lilley et al. (2014), rotifers are fundamental for feeding in early stages, especially during the 3–17 days post-fecundation when nematocyst batteries are not yet formed and isolated a-isorhiza nematocysts dominate (Ballesteros et al., 2021b). Improved growth rate in the presence of rotifers or mix of rotifers and microalgae has been reported previously for some early stages of scyphozoans (Widmer et al., 2005; Miranda et al., 2016). Artemia spp. nauplii can be added as a nutritional complement from metaephyra stage 3, when the first nematocyst batteries appear, the gastric cavity is more developed and feeding structures such as the manubrium or marginal tentacles start to grow longitudinally (Ballesteros et al., 2021b).
Non-motile prey (Table 1) sunk rapidly to the bottom of the tank according to Crow et al. (2013). This short-lived food item explains the 0% survival rate at 18 days post-fecundation for gelatin and non-motile prey food regimens (Figures 2, 3A). Contrary to Lilley et al. (2014), we do not recommend the use of immobile prey to feed early stages of P. noctiluca. According to the guidelines for jellyfish aquaculture techniques, a variety of frozen and immobile items (e.g., mysids, krill, bloodworms, mixed fish, fish eggs, and/or fish larvae) is suitable for advanced stages when feeding structures are fully developed, to provide additional nutritional value (Raskoff et al., 2003; Crow et al., 2013; Purcell et al., 2013; Duarte et al., 2021). Although the use of gelatin as the only food source was not satisfactory for P. noctiluca advanced stage (Figures 2, 3B), encapsulating a variety of items in gelatin helped keep the water cleaner and offered a wide range of elements in a single feeding time; therefore, its use is suggested as a nutritional supplement in the food regimens for jellyfish husbandry.
Moon jellyfish Aurelia spp. and the upside-down jellyfish Cassiopea spp. are the most popular cultured gelatinous prey offered for the husbandry of medusivorous jellyfish such as P. noctiluca, Cyanea spp., Chrysaora spp., and Phacellophora camtschatica (Pierce, 2005; Purcell et al., 2013; Duarte et al., 2021). Until now, their use was necessary for proper growth of medusivorous jellyfish (Crow et al., 2013; Purcell et al., 2013; Duarte et al., 2021). In particular, previous studies highlighted high values of growth, gonadal maturity and release of eggs in the presence of gelatinous prey for captive-bred P. noctiluca (Lilley et al., 2014; Ramondenc et al., 2019). Here, we achieved a production of individuals that lasted over time without the presence of gelatinous prey (Aurelia sp.) (Figure 4). Besides, the comparison of growth rate between medusivorous and non-medusivorous cultures obtained a value of DTW = 0.2, so, it can be concluded that there were no differences in growth rate regardless of the presence of gelatinous prey (Aurelia sp.). This result is an important finding for jellyfish aquaculture techniques, since up until now all the growth manuals and guidelines for medusivorous jellyfish underlined the use of other jellyfish for good growth (Raskoff et al., 2003; Crow et al., 2013; Purcell et al., 2013; Duarte et al., 2021). Aurelia spp. pieces or mucus can be added as a nutritional supplement but are not an essential item to maintain a successful culture of P. noctiluca. The key is a varied diet in advanced stages (Raskoff et al., 2003; Crow et al., 2013; Purcell et al., 2013; Duarte et al., 2021), where the jellyfish invest energy in the production and maturation of gonads for reproduction. Here, the addition of white shrimp, when gonads began to be visible, was a sufficient energy source to ensure gonadal maturation and egg production and close the life cycle in captivity (Figure 4 and Supplementary Material Tables 1, 2). Our research supports the use of white shrimp instead of gelatinous prey to avoid investing time in the culture of additional jellyfish destined solely for food (Pierce, 2005) or daily gelatinous plankton collection from the wild.
As with feeding, each stage of the life cycle requires different rearing vessels and aquarium tanks (Raskoff et al., 2003; Crow et al., 2013; Purcell et al., 2013; Duarte et al., 2021). Tank design, flow patterns and good regulation of suction velocity are critical factors for captive husbandry (Purcell et al., 2013). The ephyrae of most scyphozoans are successfully farmed in a closed system using glass dishes, beakers, plastic jugs and round glass tanks with or without aeration (Widmer et al., 2005; Purcell et al., 2013; Duarte et al., 2021). The early stages of P. noctiluca can be maintained in 5 L plastic jugs, round glass tanks or air-kreisel aquariums in a closed system with a flow of air (Figure 1B) or water that keeps them in suspension (Purcell et al., 2013; Ramondenc et al., 2019; Ballesteros et al., 2021b). Newly-developed ephyrae of P. noctiluca are transparent, in contrast to the colored newly-released ephyrae from polyps in some scyphozoan species (Straehler-Pohl and Jarms, 2010; Ballesteros et al., 2021b), a difference that causes even more time to be consumed during continuous water changes in a closed system due to the lack of visibility. Purcell et al. (2013) recommend the use of a kreisel aquarium with a gentle current for ephyrae of P. noctiluca; however, due to their small size, young ephyrae can get sucked in by the drain and trapped in the mesh screen (personal observation). From our experience, we do not recommend the use of conventional kreisel tanks for early stages of P. noctiluca. With the aim of reducing the time required for maintenance routines, novel system designs and safe techniques are required to grow the young ephyrae of P. noctiluca in an open system. Advanced stages of P. noctiluca can be maintained in conventional kreisel, pseudo-kreisel or rectangular tanks without any danger (Purcell et al., 2013; Duarte et al., 2021).
Jellyfish cultures allow availability of specimens when required and are a unique opportunity to study different areas of knowledge: from each stage of the life cycle and evolution of the cnidome (Holst, 2012; Heins et al., 2015; Ballesteros et al., 2021b) to the impact of climate change or ocean acidification on jellyfish populations (Olariaga et al., 2014). Without doubt, P. noctiluca stands out for its stinging capacity and the high number of incidents with beach-goers (Mariottini et al., 2008; De Donno et al., 2014), making it an ideal model for venom-toxicity experiments (Morabito et al., 2020; Ballesteros et al., 2021a). No significant differences were observed between the hemolytic areas caused by wild and captive specimens (p value 0.2) (Figure 7). Therefore, the optimization of its life cycle and the validation of the use of captive individuals in the TSBAA methodology may help clarify discrepancies among the scientific community about the best first-aid protocol to quickly and effectively treat its sting (Remigante et al., 2018; Ballesteros et al., 2021a), without depending on the presence of P. noctiluca in the marine environment. However, to definitively conclude the similarity in the properties of the venom, new lines of research are required, such as the study of the venom proteome of wild and captive jellyfish.
5 Conclusion
For the proper development of P. noctiluca in captive conditions, a diet that includes rotifers is recommended during the early life stages. Mucus from the jellyfish Aurelia spp. can be added but is not essential. For advanced stages, Artemia spp., codfish eggs, a variety of different food items encapsulated inside a neutral gelatin (razor shell, shrimp, sardine, mussel, hake, pacific krill and red plankton) and white shrimp provide the food sources essential for proper development. The use of gelatin-encapsulated food is a suitable technique to supply additional nutritional value without compromising water quality. As with the early stages, the presence of gelatinous prey (pieces of Aurelia spp.) is not a necessary condition for the growth of P. noctiluca juvenile and adult stages, yet it can be used as a food supplement. When the gonads begin to be visible, inclusion of white shrimp is suggested to help ensure suitable development, maturation and egg production. Warmer waters reaching 22°C are beneficial for the growth of P. noctiluca without any sign of stress. Captive-bred individuals can be used in the TSBAA methodology, since the hemolytic areas caused by their sting do not show differences from the hemolytic areas caused by wild jellyfish.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author Contributions
AB and EA conceived the study and supervised the jellyfish cultures. DP, AG, YM, and EA cared for and fed the jellyfish cultures. AB and DP carried out the research and the data curation. AS performed the statistical analyses, and drew up the graphs. AB wrote and edited the original manuscript. DP, AS, AG, YM, EA, EJ, and J-MG revised the manuscript and contributed to its improvement. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by ISDIN (Barcelona, Spain).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
Many thanks to the late Verónica Fuentes for all the knowledge that she provided on jellyfish aquaculture techniques during her life. The authors would like to thank all the staff from Loro Parque who assisted with the experiments, as well as the Loro Parque Fundación. Authors AB, AS and J-MG acknowledge the institutional support of the ‘Severo Ochoa Centre of Excellence’ accreditation (CEX2019-000928-S). Finally, we want to thank to the reviewers for their comments and suggestions to improve the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.911383/full#supplementary-material
Abbreviations
DTW, dynamic time warping; TBD, total body diameter; TSBAA, tentacle skin blood agarose assay.
References
Avian M. (1986). Temperature Influence on In Vitro Reproduction and Development of Pelagia Noctiluca (Forskål). Boll. Di. Zoolog. 53, 385–391. doi: 10.1080/11250008609355528
Ayed Y., Dellai A., Mansour H. B., Bacha H., Abid S. (2012). Analgesic and Antibutyrylcholinestrasic Activities of the Venom Prepared From the Mediterranean Jellyfish Pelagia Noctiluca (Forsskal 1775). Ann. Clin. Microbiol. Antimicrob. 11, 1–8. doi: 10.1186/1476-0711-11-15
Ayed Y., Sghaier R. M., Laouini D., Bacha H. (2016). Evaluation of Anti-Proliferative and Anti-Inflammatory Activities of Pelagia Noctiluca Venom in Lipopolysaccharide/Interferon-γ Stimulated RAW264.7 Macrophages. Biomed. Pharmacoth. 84, 1986–1991. doi: 10.1016/j.biopha.2016.11.010
Ballesteros A., Marambio M., Fuentes V., Narda M., Santín A., Gili J. M. (2021a). Differing Effects of Vinegar on Pelagia Noctiluca (Cnidaria: Scyphozoa) and Carybdea Marsupialis (Cnidaria: Cubozoa) Stings—Implications for First Aid Protocols. Toxins 13(8), 509. doi: 10.3390/toxins13080509
Ballesteros A., Östman C., Santín A., Marambio M., Narda M., Gili J.-M. (2021b). Cnidome and Morphological Features of Pelagia Noctiluca (Cnidaria: Scyphozoa) Throughout the Different Life Cycle Stages. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.714503
Bosch-Belmar M., Milisenda G., Basso L., Doyle T. K., Leone A., Piraino S. (2020). Jellyfish Impacts on Marine Aquaculture and Fisheries. Rev. Fish. Sci. Aquacult. 29, 242–259. doi: 10.1080/23308249.2020.1806201
Brotz L., Cheung W. W. L., Kleisner K., Pakhomov E., Pauly D. (2012). Increasing Jellyfish Populations: Trends in Large Marine Ecosystems. Hydrobiologia 690, 3–20. doi: 10.1007/s10750-012-1039-7
Canepa A., Fuentes V., Sabateís A., Piraino S., Boero F., Gili J. M. (2014). “Pelagia Noctiluca in the Mediterranean Sea” in Jellyfish Blooms. Eds. Pitt K. A., Lucas C. H. (Dordrecht: Springer), 237–266. doi: 10.1007/978-94-007-7015-7_11
Crow G., Howard M., Levesque V., Matsushige L., Spina S., Schaadt M., et al. (2013). “Association of Zoos and Aquariums Aquatic Invertebrate TAG.” In Jellyfish (Cnidaria/ Ctenophora) Care Manual Ed. M. Schaadt (Silver Spring), 5–79. doi: 10.13140/RG.2.1.4344.3045
D’Ambra I., Lauritano C. (2020). A Review of Toxins From Cnidaria. Mar. Drugs 18, 507. doi: 10.3390/md18100507
De Donno A., Idolo A., Bagordo F., Grassi T., Leomanni A., Serio F., et al. (2014). Impact of Stinging Jellyfish Proliferations Along South Italian Coasts: Human Health Hazards, Treatment and Social Costs. Int. J. Environ. Res. Public Health 11, 2488–2503. doi: 10.3390/ijerph110302488
Dong J., Jiang L., Tan K., Liu H., Purcell J. E., Li P., et al. (2009). Stock Enhancement of the Edible Jellyfish (Rhopilema Esculentum Kishinouye) in Liaodong Bay, China: A Review. Hydrobiologia 616, 113–118. doi: 10.1007/s10750-008-9592-9
Duarte I. M., Marques S. C., Leandro S. M., Calado R. (2021). An Overview of Jellyfish Aquaculture: For Food, Feed, Pharma and Fun. Rev. Aquacult. 14, 265–287. doi: 10.1111/raq.12597
Frazão B., Campos A., Osório H., Thomas B., Leandro S., Teixeira A., et al. (2017). Analysis of Pelagia Noctiluca Proteome Reveals a Red Fluorescent Protein, a Zinc Metalloproteinase and a Peroxiredoxin. Protein J. 36, 77–97. doi: 10.1007/s10930-017-9695-0
Giorgino T. (2009). Computing and Visualizing Dynamic Time Warping Alignments in R: The Dtw Package. J. Stat. Softw. 31, 1–24. doi: 10.18637/jss.v031.i07
Hall A., Mathieu L., Blomet J. (2018). Pelagia Noctiluca Jellyfish: Can Lesions and Symptoms be Prevented or Ameliorated? J. Mar. Biol. Aquacult. 4, 53–57. doi: 10.15436/2381-0750.18.1971
Heins A., Glatzel T., Holst S. (2015). Revised Descriptions of the Nematocysts and the Asexual Reproduction Modes of the Scyphozoan Jellyfish Cassiopea Andromeda (Forskål 1775). Zoomorphology 134, 351–366. doi: 10.1007/s00435-015-0263-x
Helm R. R. (2018). Evolution and Development of Scyphozoan Jellyfish. Biol. Rev. 93, 1228–1250. doi: 10.1111/brv.12393
Holst S. (2012). Morphology and Development of Benthic and Pelagic Life Stages of North Sea Jellyfish (Scyphozoa, Cnidaria) With Special Emphasis on the Identification of Ephyra Stages. Mar. Biol. 159, 2707–2722. doi: 10.1007/s00227-012-2028-0
Larson R. J. (1987). A Note on the Feeding, Growth, and Reproduction of the Epipelagic Scyphomedusa Pelagia Noctiluca (Forskål). Biol. Oceanogr. 4, 447–454. doi: 10.1080/01965581.1987.10749501
Lilley M. K. S., Ferraris M., Elineau A., Berline L., Cuvilliers P., Gilletta L., et al. (2014). Culture and Growth of the Jellyfish Pelagia Octiluca in the Laboratory. Mar. Ecol. Prog. Ser. 510, 265–273. doi: 10.3354/meps10854
Marambio M., Canepa A., Lòpez L., Gauci A. A., Gueroun S. K. M., Zampardi S., et al. (2021). Unfolding Jellyfish Bloom Dynamics Along the Mediterranean Basin by Transnational Citizen Science Initiatives. Diversity 13, 274. doi: 10.3390/d13060274
Mariottini G. L., Elisabetta G., Luigi P. (2008). The Mauve Stinger Pelagia Noctiluca (Forsskål 1775). Distribution, Ecology, Toxicity and Epidemiology of Stings. A Review. Mar. Drugs 6, 496–513. doi: 10.3390/md20080025
Merquiol L., Romano G., Ianora A., D’Ambra I. (2019). Biotechnological Applications of Scyphomedusae. Mar. Drugs 17, 604. doi: 10.3390/md17110604
Milisenda G., Martinez-Quintana A., Fuentes V. L., Bosch-Belmar M., Aglieri G., Boero F., et al. (2018a). Reproductive and Bloom Patterns of Pelagia Noctiluca in the Strait of Messina, Italy. Estuar. Coast. Shelf. Sci. 201, 29–39. doi: 10.1016/j.ecss.2016.01.002
Milisenda G., Rossi S., Vizzini S., Fuentes V. L., Purcell J. E., Tilves U., et al. (2018b). Seasonal Variability of Diet and Trophic Level of the Gelatinous Predator Pelagia Noctiluca (Scyphozoa). Sci. Rep. 8 (1), 1–13. doi: 10.1038/s41598-018-30474-x
Miranda F. S., Chambel J., Almeida C., Pires D., Duarte I., Esteves L., et al. (2016). Effect of Different Diets on Growth and Survival of the White-Spotted Jellyfish, Phyllorhiza Punctata. Front. Mar. Sci. doi: 10.3389/conf.FMARS.2016.04.00042
Montgomery L., Seys J., Mees J. (2016). To Pee, or Not to Pee: A Review on Envenomation and Treatment in European Jellyfish Species. Mar. Drugs 14, 127. doi: 10.3390/md14070127
Morabito R., Cornara L., la Spada G., Marino A., Mariottini G. L., Remigante A., et al. (2020). Inhibitory Effect of Plant Extracts on the Cytotoxicity of Eurytele Nematocysts From Pelagia Noctiluca. J. Biol. Res. (Italy) 93, 96–101. doi: 10.4081/jbr.2020.9136
Olariaga A., Guallart E. F., Fuentes V., López-Sanz À., Canepa A., Movilla J., et al. (2014). Polyp Flats, a New System for Experimenting With Jellyfish Polyps, With Insights Into the Effects of Ocean Acidification. Limnol. Oceanogr.: Methods 12, 212–222. doi: 10.4319/LOM.2014.12.212/FORMAT/PDF
Oliveira C. S., Caldeira C. A. S., Diniz-Sousa R., Romero D. L., Marcussi S., Moura L. A., et al. (2018). Pharmacological Characterization of Cnidarian Extracts From the Caribbean Sea: Evaluation of Anti-Snake Venom and Antitumor Properties. J. Venom. Anim. Toxin. Includ. Trop. Dis. 24 (1), 22. doi: 10.1186/s40409-018-0161-z
Östman C., Hydman J. (1996). Nematocyst Analysis of Cyanea Capillata and Cyanea Lamarckii (Scyphozoa, Cnidaria)*. Scient. Mar. 61, 313–344.
Pastor-Prieto M., Bahamon N., Sabatés A., Canepa A., Gili J. M., Carreton M., et al. (2021). Spatial Heterogeneity of Pelagia Noctiluca Ephyrae Linked to Water Masses in the Western Mediterranean. PloS One 16, e0249756. doi: 10.1371/journal.pone.0249756
Pierce J. (2005). A System for Mass Culture of Upside-Down Jellyfish Cassiopea Spp as a Potential Food Item for Medusivores in Captivity. Int. Zoo Yearbook 39, 62–69. doi: 10.1111/j.1748-1090.2005.tb00005.x
Purcell J. E., Baxter E. J., Fuentes V. L. (2013). “Jellyfish as Products and Problems of Aquaculture,” in Advances in Aquaculture Hatchery Technology. Eds.Allan G., Burnell G. (Cambridge, United Kingdom: Elsevier), 404–430. doi: 10.1533/9780857097460.2.404
Ramondenc S., Ferrieux M., Collet S., Benedetti F., Guidi L., Lombard F. (2019). From Egg to Maturity: A Closed System for Complete Life Cycle Studies of the Holopelagic Jellyfish Pelagia Noctiluca. J. Plankt. Res. 41, 207–217. doi: 10.1093/plankt/fbz013
Raskoff K. A., Sommer F. A., Hamner W. M., Cross K. M. (2003). Collection and Culture Techniques for Gelatinous Zooplankton. Biol. Bull. 204, 68–80. doi: 10.2307/1543497.z
R Core Team (2017). A Language and Environment for Statistical Computing (Vienna: R Foundation for Statistical Computing).
Remigante A., Costa R., Morabito R., la Spada G., Marino A., Dossena S. (2018). Impact of Scyphozoan Venoms on Human Health and Current First Aid Options for Stings. Toxins. 10, 133. doi: 10.3390/toxins10040133
Rocha J., Peixe L., Gomes N. C. M., Calado R. (2011). Cnidarians as a Source of New Marine Bioactive Compounds—An Overview of the Last Decade and Future Steps for Bioprospecting. Mar. Drugs 9, 1860–1886. doi: 10.3390/md9101860
Rosa S., Pansera M., Granata A., Guglielmo L. (2013). Interannual Variability, Growth, Reproduction and Feeding of Pelagia Noctiluca (Cnidaria: Scyphozoa) in the Straits of Messina (Central Mediterranean Sea): Linkages With Temperature and Diet. J. Mar. Syst. 111–112, 97–107. doi: 10.1016/j.jmarsys.2012.10.001
Sandrini L. R., Avian M. (1983). Biological Cycle of Pelagia Noctiluca: Morphological Aspects of the Development From Planula to Ephyra. Mar. Biol. 74, 169–174. doi: 10.1007/BF00413920
Schaadt M., Widmer C. L., Sowinski N. (2017). “Chapter Jellyfish” in Marine Ornamental Species Aquaculture. Eds. Calado R., Olivotto I., Oliver M., Holt J. (New Jersey: John Wiley & Sons Ltd), 457–473. doi: 10.1002/9781119169147.ch2
Schäfer S., Gueroun S. K. M., Andrade C., Canning-Clode J. (2021). Combined Effects of Temperature and Salinityon Polyps and Ephyrae of Aurelia Solida (Cnidaria: Scyphozoa). Diversity 13, 573. doi: 10.3390/d13110573
Schindelin J., Arganda-Carreras I., Frise E., Kaynig V., Longair M., Pietzsch T., et al. (2012). Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 9, 676–682. doi: 10.1038/nmeth.2019
Shtewi H., Abushaala N., Elfituri A., Abdullah A. B. (2022). Morphological Feature of Pelagia Noctiluca (Forskål 1775) (Cnidaria: Scyphozoa) in Western Libyan Coast, Tripoli. J. Pure. Appl. Sci. 21, 24–27. doi: 10.51984/jopas
Straehler-Pohl I., Jarms G. (2010). Identification Key for Young Ephyrae: A First Step for Early Detection of Jellyfish Blooms. Hydrobiologia 645, 3–21. doi: 10.1007/s10750-010-0226-7
Tilves U., Purcell J. E., Fuentes V. L., Torrents A., Pascual M., Raya V., et al. (2016). Natural Diet and Predation Impacts of Pelagia Noctiluca on Fish Eggs and Larvae in the NW Mediterranean. J. Plankt. Res. 38, 1243–1254. doi: 10.1093/plankt/fbw059
Verkhusha V. V., Lukyanov K. A. (2004). The Molecular Properties and Applications of Anthozoa Fluorescent Proteins and Chromoproteins. Nat. Biotechnol. 22, 289–296. doi: 10.1038/nbt943
Wickham H. (2016). Ggplot2 Elegant Graphics for Data Analysis (New York, NY: Springer). doi: 10.1007/978-3-319-24277-4
Widmer C. L. (2005). Effects of Temperature on Growth of North-East Pacific Moon Jellyfish Ephyrae, Aurelia Labiata (Cnidaria: Scyphozoa). J. Mar. Biol. Assoc. Unite. Kingd. 85, 569–573. doi: 10.1017/S0025315405011495
Widmer C. L., Voorhees J. P., Badger M. A., Lambert J. W., Block N. M. (2005). The Effects of Rearing Vessels and Laboratory Diets on Growth of Northeast Pacific Jellyfish Ephyrae (Cnidaria: Scyphozoa). Drum. Croaker. 36, 29–36.
Keywords: captivity, early stage, generation, growth rate, kreisel, life cycle, rotifer, venom
Citation: Ballesteros A, Páez D, Santín A, García A, Martín Y, Alonso E, Jourdan E and Gili J-M (2022) Successful Culture of Pelagia noctiluca (Cnidaria: Scyphozoa) Over Time: A Continuous Supply of the Holoplanktonic Jellyfish for Research and Industrial Applications. Front. Mar. Sci. 9:911383. doi: 10.3389/fmars.2022.911383
Received: 02 April 2022; Accepted: 20 April 2022;
Published: 13 May 2022.
Edited by:
Sílvia Lourenço, Instituto Politécnico de Leiria, PortugalReviewed by:
Ricardo Calado, University of Aveiro, Santiago University Campus, PortugalMar Bosch Belmar, University of Palermo, Italy
Copyright © 2022 Ballesteros, Páez, Santín, García, Martín, Alonso, Jourdan and Gili. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ainara Ballesteros, YmFsbGVzdGVyb3NAaWNtLmNzaWMuZXM=