 Md Hadiuzzaman1,2
Md Hadiuzzaman1,2 Mohammad Moniruzzaman
Mohammad Moniruzzaman Md Shahjahan
Md Shahjahan Sungchul C. Bai
Sungchul C. Bai Taesun Min
Taesun Min Zakir Hossain
Zakir Hossain
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Mar. Sci., 15 July 2022
Sec. Marine Fisheries, Aquaculture and Living Resources
Volume 9 - 2022 | https://doi.org/10.3389/fmars.2022.905986
This article is part of the Research TopicNew Progresses and Effects of Functional Feed Additives on Marine Aquatic AnimalsView all 13 articles
β-glucan is considered as an effective immunostimulant because of its binding capacity to different receptors on leukocytes leading to the stimulation of immune responses including bactericidal activity, cytokine productivity, and survival fit ability at cellular levels. In response to immune cell surface receptors, β-glucan stimulates to release cytokines and chemokines. It has been found that these signaling proteins eventually stimulate the immunocompetent cells in fish such as monocytes, macrophages, and neutrophils for killing pathogens by phagocytosis, oxidative burst, and cytotoxic killing activities. They also procreate immunological memories and specific antibodies through activation of T and B lymphocytes. Researchers have proved that β-glucan can modulate some important biochemical (serum hemoglobin, serum protein, and total hemocyte count) and immunological (lysozyme activity, phagocytic activity, oxidative burst activity, and phenoloxidase activity) properties providing more competent immune profile for treating fish and aquatic organisms. β-glucan-supplemented fish showed limited sensitivity of genes involved in acute inflammatory reactions. Findings have shown that β-glucan exerts a positive impact on fish and aquatic organisms’ immunity, enhancing their disease resistance by increasing functional and decreasing deleterious responses. This review focuses on the basic bump of β-glucan on fish and shellfish immunity and recent information on the uses of β-glucan in progressive aquaculture.
Different studies in fishes have proven β-glucan as a potent immunostimulant for improving the immunity of cultured fish against disease and stress (Figueras et al., 1998; Kawakami et al., 1998; Meena et al., 2013; Yamamoto et al., 2018a; Yamamoto et al., 2018b; Yamamoto et al., 2020). The immunostimulatory properties of β-glucans were first identified in mammals resulting in an increased resistance to infectious pathogens (Di Luzio, 1985). However, in recent years, special attention has been given on the use of β-glucan in stimulating the recovery potential of immunocompromised fish and aquatic organisms. Many studies have proven the significant effect of β-glucan on fish growth and survival (Cook et al., 2003; Misra et al., 2006), protective resistance to specific pathogen (Welker et al., 2007; Sealey et al., 2008), and adjuvant effect on increased antibody production (Selvaraj et al., 2005; Kamilya et al., 2006). The introductory section of this article aims to provide an elaborate discussion on β-glucan sources, their distinguishing properties, and the scope of using it in aquaculture.
Many fungi possess β-glucans in their cell wall, which has important immunomodulatory properties (Brown et al., 2003). The cell walls of many cereal plants (barley, oat, etc.) are accordingly flourished with β-glucans (Volman et al., 2008). The yeast species Saccharomyces cerevisiae cell wall comprises 50%–55% β (1/3) glucan and 10%–15% β-(1/6) glucan of the total polysaccharides (Figure 1) (Lesage and Bussey, 2006). Mushroom species Lentinus edodes, Grifola frondosa, and Ganoderma lucidum are reliable sources of good-quality β-glucans having therapeutic significance (Wasser and Weis, 1999). Laminaria sp., the members of brown seaweed, may also be better sources of β-glucan. They are extensively cultivated for commercial extraction of laminaran, fucoidan, alginate, etc. (Rioux et al., 2007). The pathogenic fungus Pneumocystis carinii holds cell wall β-glucan responsible for the secretion of pro-inflammatory signaling proteins (cytokines and chemokines) (Lebron et al., 2003). Cell wall composition of Cryptococcus neoformans demonstrates β-glucan as an essential structural element of this morbific mold (Reese et al., 2007). The ascomycetes member Sclerotinia sclerotiorum retains reactive β-glucan in their cell wall components (Borchers et al., 2004). The cell walls of some non-pathogenic bacteria of the Rhizobiaceae family are also composed of β-glucan (McIntosh et al., 2005).

Figure 1 Molecular structure of a typical yeast β-glucan.
β-glucan is an indigestible constituent (fiber substance) that makes up the cell wall of cereal plants (Figure 2) (e.g., oat, barley, etc.) DeVries (2003). β-glucan is a starch-less polysaccharide with recurrent units of glucose monomers united by (1/3)-β-D-glycosidic linkages and very often arranged with β (1/6)-linked side chains of non-identical presentation (Buckeridge et al., 2004). The most convenient forms of β-glucans are those composed of glucose monomers linked by β (1/3) glycosidic bonds (Goodridge et al., 2009; Meena et al., 2013). Glucans may be branched in several ways depending on their sources. The yeast and fungal glucans share β (1/3, 1/6) glycosidic bonds that are usually highly branched (Figure 1). Yeast’s β-glucans consist of β (1/3)-linked backbones and β (1/6)-linked side chains (Zlatkovic et al., 2003). Algal β-glucan laminaran has a skeleton of β (1/3) glucopyranosyl units with β (1/6) branching (Zvyagintseva et al., 1999). In nature, cereals’ (oat, barley, and rye) β-glucans contain β (1/3, 1/4)-joined links (Figure 2), whereas mushrooms contain higher amounts of β (1/3, 1/6)-joined links (Volman et al., 2008). Some oat β-glucans are linear, unbranched polysaccharides, furnished with 70% 1/4-O-linked bondages and 30% 1/3-O-linked bondages between their β-D-glucopyranosyl units (Butt et al., 2008). On the contrary, bacterial β-glucan (curdlan) is unbranched with only (1/3)-β-D-linkages between its glycopyranosyl molecules (Johansson et al., 2008).
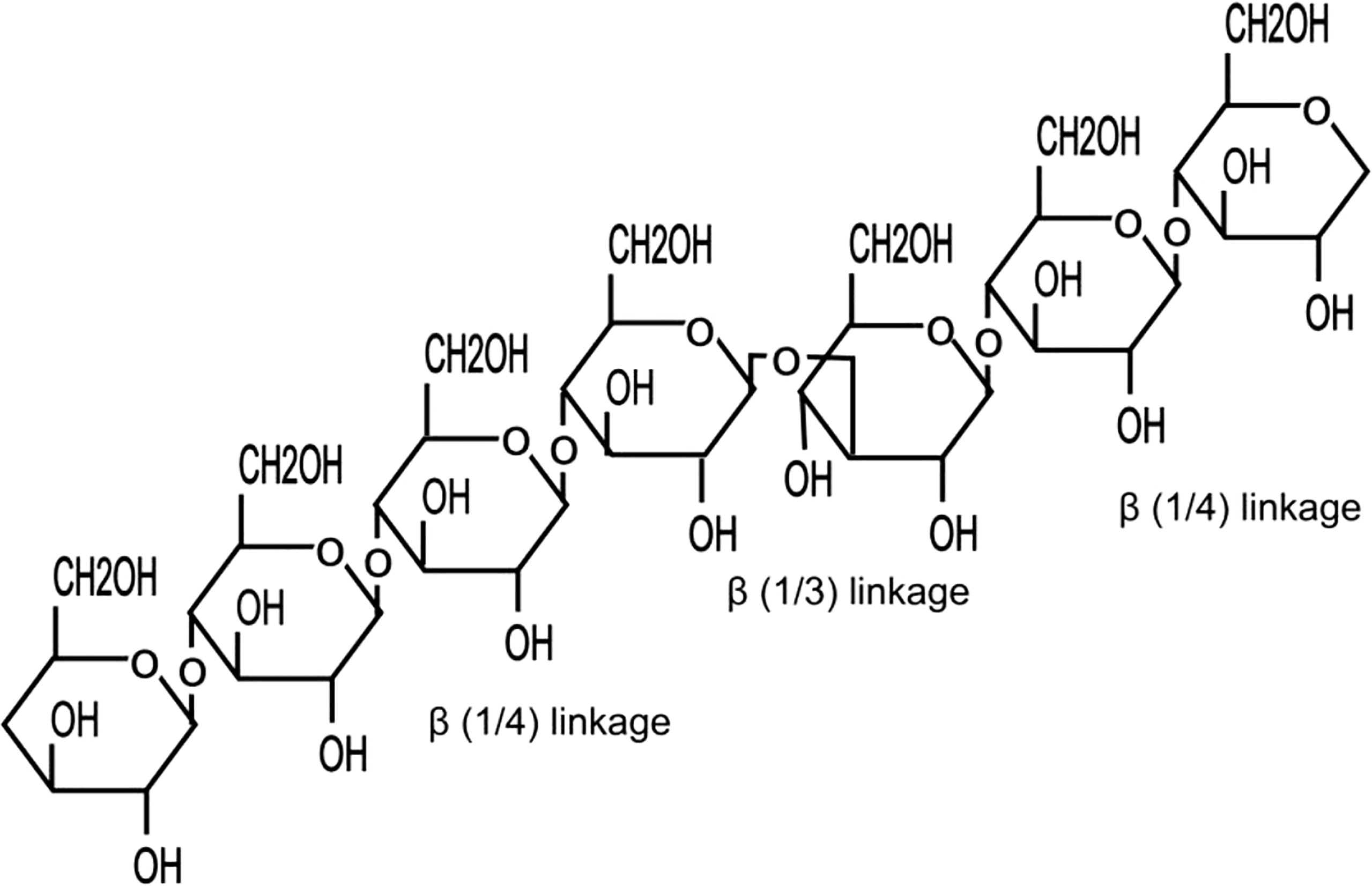
Figure 2 Molecular structure of a typical cereal β-glucan.
Larger (molecular weight > 5 kDa and < 200 kDa) sizes of β-glucans have regulatory effects on the host immune system (Bagni et al., 2005). They boost up the host’s non-specific defense mechanism and instigate leukocytes triggering their phagocytic and anti-pathogenic reactions through production of pro-active oxygen species (superoxide anions, hydrogen peroxide, hydroxyl radical, hypochlorous acid, etc.) and nitrogen intermediates (nitrite, amides, and nitrogen dioxide) (Lee et al., 2002). The variations in molecular weight, shape, and structure of β-glucans have an impact on their immune performances (Akramiene et al., 2007; Meena et al., 2013; Chu, 2014). Lines of evidence demonstrate that specific physicochemical properties, for example, molecular structure, solution strength, molecular weight, and net charge of β-glucan, play a vital role in determining the magnitude of β-glucan binding to macrophage receptor(s) and how it modulates the immune responses (Mueller et al., 2000). Researchers have found that insoluble (1/3, 1/6) β-glucans have higher biological involvement than that of its soluble β (1/3, 1/4) counterparts (Ooi and Liu, 2000). Indigestible β-glucans may lead to an alteration in the population of gut microbiota (Swennen et al., 2006). β-glucans are capable of modulating biological immune responses (Miura et al., 1996). β-glucan modulates immune cells but never overstimulates, which is key to the safety of this product (Gatlin and Li, 2004).
The provision of β-glucans, either dietary or supplementary injection, stimulates the recovery potential of immunosuppressed cells in teleost fishes (e.g., Atlantic salmon and rainbow trout) against infectious diseases under ordinary habitat condition (Petit and Wiegertjes, 2016). Most of the traditional antibiotics are banned due to the potential exposure of antibiotic-resistant bacteria, their residual effects on aquatic habitat, and suppressing repercussion on aquatic organisms’ immune system (FAO, 2002). Moreover, antibiotics may disrupt growth and feed efficiency by declining gut microbes and reducing amino acid utilization by the host animal (Rawles et al., 1997). These adverse effects can successfully be overcome by exploring the prebiotic nature of β-glucan (Meena et al., 2013). Thus, the immunomodulating effects of β-glucan can be characterized into (1) prebiotic effects of β-glucan as indirect immunomodulation in terms of fermentation of β-glucan by naïve bacteria and changes in microbial composition as well as the shift in the production of short chain fatty acid (SCFA) metabolites in the gastrointestinal tract (GIT) of fish, and (2) immunity enhancement of host by β-glucan as the direct immunomodulation in terms of receptor-mediated recognition in GIT of fish (Petit et al., 2022).
The production of fish fry is often hampered up to 10% in the aquaculture sector by high mortality rates due to infectious diseases. The delivery of β-glucan as a dietary supplement to larval fish can have a considerable effect in improving the animals’ innate defenses (Bricknell and Dalmo, 2005). A considerable amount of protective resistance has been achieved with the β-glucan adjuvanted vaccine in rainbow trout (Onchorynchus mykiss) (Siwicki et al., 2004), catla (Catla catla) (Kamilya et al., 2006), and sea bass (Dicentrarchus labrax) (Bonaldo et al., 2007). Thus, the potential of adjuvanticity of β-glucan can also be explored for vaccination of fish in commercial aquaculture.
The innate immune system comprises different integrants including physical barriers (skin, epithelial cell surfaces, and mucus itself), phagocytic cells (monocytes, macrophages, and neutrophils), antibacterial enzymes and peptides [lysozymes, phospholipase (A2), defensins, cathelicidins, transferrin, etc.], inflammation responsive serum proteins (complement, C-reactive protein, lectins, and ficolins), cells that produce cytokines and inflammatory mediators (macrophages, mast cells, and natural-cytotoxic cells), and their surface receptors [toll-like receptors (TLRs) and other pattern recognition receptors (PRRs)]. These components initiate the enzyme cascade system to establish the first-line defense system by eliminating or destroying pathogens (Kumagai and Akira, 2010). The first line of defense identifies pathogens or immunostimulants by decoding the generic properties of their macromolecules (carbohydrates, lipids, nucleic acids, and proteins), the pathogen-associated molecular patterns (PAMPs) (Akira and Hemmi, 2003). This first line of defense has evolved receptors, PRRs, capable of pointing out and encountering pathogens via their PAMPs (Brown and Gordon, 2003). Microorganisms or their toxins or bioactive ingredients are identified by PRRs when first encountering the immune cells and mechanisms of the baseline defense (Medzhitov, 2007). The innate immune defenses are non-specific, predominantly respond to pathogens in a customary way, and destroy them by an inclusive manner (Alberts et al., 2002). Thus, these non-specific defenses can also be triggered by damage-associated (injured or abnormal cells’ mediated) molecular patterns (DAMPs) (Matzinger, 2002). Following the initial responses to a specific pathogenic infection, the invading organism acquires an immunological memory (pathogen-specific receptor) and leads to a more specific reaction (specific antibody production through genetic recombination) to succeeding the infection by the previous one (Figure 3). This secondary response is known as adaptive immunity. Like the inherent immune defense, the acquired immunity comprises both humoral immunity complements and cell-mediated immunity complements (Alberts et al., 2002). Humoral components are soluble proteins of the plasma and body fluids. The molecules include transferrin, interferons, lytic enzymes, macroglobulin, natural antibodies, proteins of the classical and alternative pathways, and proteins under the group cytokines and chemokines (Rodrigues et al., 2020). A study in seabream (Sparus aurata) fed a diet supplemented with yeast cell showed humoral immune response (antibody production and specific defense) (Cuesta et al., 2004). Teleostean B cells are reported to produce immunoglobulin M (IgM), immunoglobulin D (IgD), and immunoglobulin T (IgT) (Danilova et al., 2005; Solem and Stenvik, 2006). Glucans are generally believed to be associated with a first-line defense mechanism by binding to specific receptors on major phagocytic cells and other components of the inherent immune system (Mueller et al., 2000). Different cell surface receptors contain lectins, scavenger receptors (SRs), transmembrane proteins on monocytes/macrophages, natural cytotoxic cells (NCC), and other lymphocyte subcomponents binding various types of β-glucan (Brown and Gordon, 2003). A variety of glucan binding sites on macrophages have been identified, but their mechanism of modulating an innate immune system is not fully clear.

Figure 3 Mechanism of modulating the fish immune system by β-glucan.
The first step of β-glucan–macrophage complex formation is binding to specific receptors present on the immune cell surface (Figure 3). Complement receptor type 3 (CR3) (Vetvicka et al., 1996), lactosylceramide moiety (Zimmerman et al., 1998), dectin-1 (C-type lectin) (Rice et al., 2002), and carboxymethylated (CM) SRs (Vereschagin et al., 1998) have been considered to be major glucan binding sites on macrophages and other immunocompetent cells. Receptor-bound β-glucan may mediate the production of inflammatory cytokines (interleukins, interferons, lymphokines, and tumor necrosis factor) and chemokines (CC, CXC, C, and CX3C). These signaling proteins are believed to aggravate phagocytic activity and microbial killing efficiency of immune cells through oxidative burst and natural cytotoxic liquidation (Brown et al., 2003; Misra et al., 2006). Presently, instead of dectin-1 and CR3, attention has been given to toll-like receptor 2 (TLR-2) inducing important roles in innate immunity. Glucan instigates different cell types (macrophages, neutrophils, and T lymphocytes) to mediate its stimulatory signals by forming bondage to TLR-2 associating other cell membrane receptors (Aizawa et al., 2018). The process may first induce membrane receptors and then form subsequent complexation with TLR homologues. TLR homologues have been identified in Atlantic salmon and puffer fish. The homologues have also been described in zebrafish, flounder, and goldfish (Rodrigues et al., 2020).
CR3 (type ι membrane protein, an integrin dimer) consists of αMβ2 CD11b⁄CD18 (Cluster of differentiation 18) and is manifested exclusively by myeloid cells (evolved from progenitor cells and target oriented stem cells) comprising monocytes/macrophages, neutrophils, and NCCs (Blystone and Brown, 1999). They are frequently characterized by dendritic cells (DCs) and antigen-presenting cells having a central role in adaptive immune response (Banchereau and Steinman, 1998), present on the cell surfaces of skin, nose, lungs, stomach, and intestines. CR3 has multiple types of ligands (a molecule that binds to another molecule) inclined to bind with different fungal, parasitic, and microbial cells, and also encrypted with some complementary sites for inducing glycoprotein complex (fibrinogen), coagulation factors, and intercellular adhesion molecule-1 (ICAM-1). CR3, in association with complement component iC3b, phagocytizes complement-opsonized foreign particles (Blystone and Brown, 1999). Some studies suggest that CR3 may persuade the assemblage of low-affinity receptors and their binding to the interlinking protein filaments (cytoskeleton) of various microorganisms. Particular propensity to co-receptors proves that CR3 has distinguished binding capacity (5 × 10-8 M) to β-glucan and the domain is distinct from the iC3b binding site. Lectin domain is also involved in CR3 assemblage with urokinase plasminogen activator receptor (uPAR), an autogenous surface receptor, for binding to certain microbial carbohydrates that do not contain β-glucan (Xia and Ross, 1999). A recent study reveals that antibodies responsive to CR3 or lactosylceramide hamper β-glucan particle fixing to human neutrophils. Thus, CR3 generated signaling requires β-glucan-mediated interaction of CR3 with lactosylceramide-linked lyn-kinase. The finding suggests that CR3 may act as a co-receptor with lactosylceramide in binding β-glucan (Nakayama et al., 2008). A study on zebrafish genome revealed that two genes related to CR3 proved the indirect existence of the receptor in fish (Petit et al., 2019).Lactosylceramide (a glycosphingolipid), which consists of an aquaphobic ceramide (waxy lipid) and an aquaphile sugar moiety (a portion of a complex molecule), is found in microdomains (lipid rafts) on the cytomembranes of immune cells. Lactosylceramide (LacCer)’s ability to interact with (1/3)-β-D-glucans is first recognized through biochemical reaction of distinct human leukocyte membrane constituents (Zimmerman et al., 1998). The reaction between glycosphingolipid and β-glucans promotes cellular responses, the release of cytokines, and the release of MIP-2 (macrophage inflammatory protein-2) and TNF (tumor necrosis factor) (Evans et al., 2005). These signaling proteins initiate the respiratory oxidative burst and anti-microbial activities of leukocytes (Wakshull et al., 1999).
Dectin-1 (type ιι membrane receptor) predominantly binds protein ligands (Brown et al., 2003) and is considered to be the prominent β-glucan binding receptor in mammals (Dennehy and Brown, 2007; Petit et al., 2019). Dectin-1 is assessed as a β-glucan receptor (βGR) for its particular (opsonin-independent) β-glucan binding affinity (Brown et al., 2003). It consists of a recognition domain (lectin-like carbohydrate) with a short stalk [C-terminal C-type lectin or C-type lectin receptor (CLR)] and a cytoplasmic tail (consists of 40 amino acids, with an amino terminal intracellular tail), and therefore has the ability to recognize carbohydrate having β (1/3) and/or β (1/6) glycosidic bondages (Dennehy and Brown, 2007). Dendritic cells and a subpopulation of T lymphocytes are reported to lower the expression level of C-type lectin (Taylor et al., 2002). This membrane protein can be detected by flow cytometry on a subcomponent of B and T lymphocytes, though its function on these cells is not well-defined (Willment et al., 2001). However, other lines of evidence reveal that Dectin-1, in association with toll-like receptor 2 (TLR-2), results in pro-inflammatory reaction to mycobacterial contamination (Yadav and Schorey, 2006). For structural resemblance, C-type lectin has higher affinity to β-glucans having (1/3)-β-D-glucopyranosyl backbones (Adam et al., 2008). Dectin-1, as a response to antigens, recruits a variety of macrophages, DCs, and neutrophils responsible for the production of inflammatory mediators and signaling proteins (cytokines, chemokines, etc.). These antigen-presenting substances are believed to accelerate phagocytosis, oxidative burst, and neutrophil degranulation into the phagosomes of immune cells. An investigation on Candida albicans, P. carinii, and Aspergillus fumigatus has proven that βGR is capable of recognizing β-glucan resembling compounds (e.g., zymosan and curdlan) and defying fungal infections in mouse (Taylor et al., 2007; Werner et al., 2009). In fish genomes, there is no clear homologue of Dectin-1 identified so far (Petit et al., 2019). However, Dectin-1 homologues have yet been identified as β-glucan pattern recognition receptors on salmon macrophages and catfish neutrophils (Magnadottir, 2006). A receptor associated with dectin-1 activation through the CLR signaling pathway is reported to mediate the immunostimulatory functions of β-glucan in common carp macrophages (Petit et al., 2019). In a recent study, Petit et al. (2022) reported that β-glucans such as curdlan and MacroGard can be fermented and produced three dominant SCFAs such as acetate, butyrate, and propionate through the production of nitric oxide and expressions of several cytokines (interleukin-1b, -6, -10, and TNF-α) in vitro using head kidney leukocytes of common carp. Moreover, the researchers confirmed the fermentation of MacroGard (β-glucan) by specific bacteria and immunomodulation by β-glucan through the production of SCFAs in the GIT of common carp in vivo.
Scavenger receptors are a superfamily of cell surface receptors having the properties of recognizing and binding to diverse ligands (Patten and Shetty, 2018). These ligands usually include low-density lipoproteins (LDL), apoptotic cells (pyknotic cells due to apoptosis), phospholipids (lecithin and phosphatidylcholine), proteoglycans (testican and versican), and carbohydrates (mainly polysaccharides) (Murphy and Weaver, 2017). This wide recognition range allows scavenger receptors to play a vital role in homeostasis (equilibrium among biological functions), apoptosis, inflammatory disorders, and pathogen removal (PrabhuDas et al., 2017).
Purified β (1/3, 1/6) glucan has been increasingly used as an immunostimulant (biologically active compounds) and/or an adjuvant (substances enhancing immune response) to improve the fish immune system (Petit and Wiegertjes, 2016; Filho et al., 2019). The most useful bioactive ingredient used in aquaculture is β-glucan extracted from yeasts, mushrooms, seaweeds, and cereal plants (Bagni et al., 2005). Researchers have proven that β-glucans increase fish resistance to infectious diseases primarily by boosting their non-specific defense mechanisms. Some studies in fish have demonstrated evidence of developing the specific defense as well (Siwicki et al., 2004). The biological extracts (Lentinan, Laminaran, and Schizophyllan) and/or therapeutic chemicals (Levamisole, MacroGard, EcoActiva, Ergosan, and VitaStim) trigger the immune cells or non-specific cytotoxic cells for microbial killing through phagocytosis and/or oxidative burst (Sakai, 1999). Among the frequently trialed immunostimulants, only a few are considered suitable for preventive measure in aquaculture (Siwicki et al., 1998). Major reports that have been made on the immunomodulatory effects of β-glucans in aquaculture are related to yeast, fungi, and macro algal extracts having molecular weights ranging from 5 kDa (kelp, Laminaria digitata) to 200 kDa (yeast, Saccharomyces cerevisae) (Bagni et al., 2005). Many experiments have been conducted on different fish species to determine the effects of purified β (1/3, 1/6) glucan on their immune responses (Table 1). Researchers have found that β-glucan from different sources, despite their similarity in structures, molecular weights, and solution strength, can differ significantly. Even the same β-glucan administered through different routes responds in varying magnitude (Chen and Seviour, 2007).
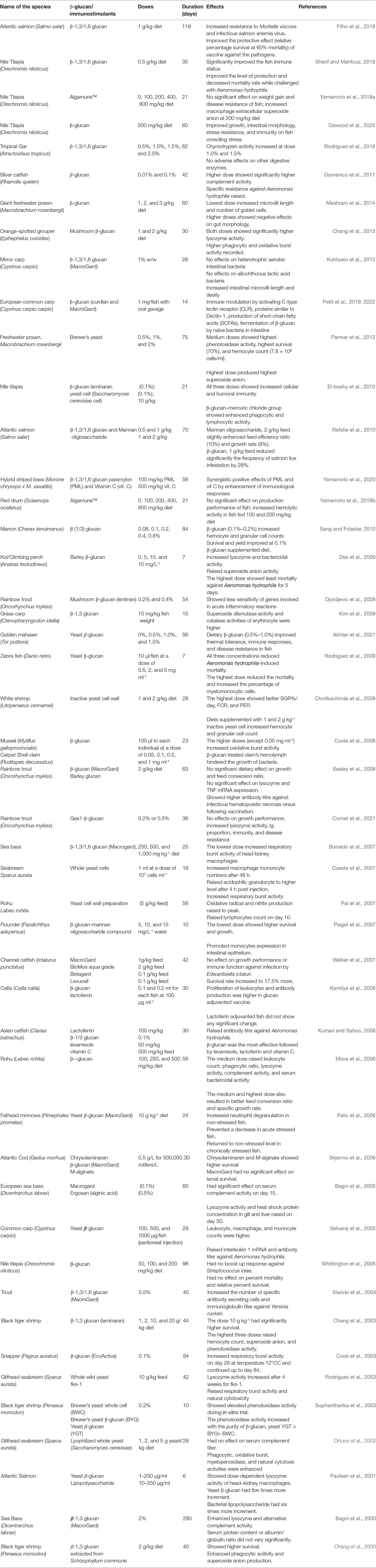
Table 1 Some research findings on the uses of β-glucan in aquaculture.
Misra et al. (2006) found that rohu fish (Labeo rohita) fed a diet containing 250 mg of β-glucan/kg showed maximum level of immune activities (serum bactericidal activity, lysozyme activity, complement activity, and phagocytic activity) after 42 days of feeding while the indices started increasing from the 28th day of the experiment. Superoxide anion and lymphokine production made a remarkable change in their volume. The magnitude of the activities started to decrease after 42 days, maintaining a peak at a dose of 250 mg of β-glucan kg-1 diet. In another investigation, Pal et al. (2007) reported similar results in rohu (L. rohita) fish. They evidenced that the ingestion of pelleted feed containing the yeast (S. cerevisiae) cell wall preparation (5 g/kg feed) increased the volume of reactive oxidative radicals and nitrogen intermediates (nitrite). Lymphocyte proliferation and phagocytic activity were also enhanced during the experimental feeding. These four parameters reached a peak in treated fish on day 10 and continued until day 20 with a significant difference on day 15 compared to the control group. Paulsen et al. (2001) found that head kidney macrophages of Atlantic salmon (Salmo salar), supplemented with yeast β-glucan, increased extracellular lysozyme production five times higher than control after 6 days of cell incubation. The optimum concentration for linear dose-dependent curve was between 1 and 250 µg/ml. The stimulated cells showed enhanced lysozyme gene (lysozyme mRNA) transcription responsible for escalated lysozyme secretion. Chang et al. (2013) observed that the orange-spotted grouper (Epinephelus coioides) fed mushroom β-glucan at 1 g and 2 g per kg diet had significantly higher lysozyme and complement activities against Vibrio alginolyticus. The activities started to increase from day 6 and continued up to day 30. On the contrary, phagocytic activity started to increase from day 9 and respiratory burst activity started to increase from day 12. In comparison to the control fish, the minimum level of incorporation (0.5 g/kg diet) of β-glucan also brought a significant change in these parameters after feeding for 15 days. El-boshy et al. (2010) observed similar responses in Nile tilapia when fed a diet supplemented with whole yeast (S. cerevisiae, 10 g/kg diet), β-glucan (0.1%), and laminaran (0.1%). β-glucan-treated Nile tilapia showed elevated phagocytic activity, phagocytic index, bactericidal activity, and total lymphocyte count when challenged with Aeromonas hydrophila for 21 days.
Bagni et al. (2000) conducted a similar research to assess the effect of an immunostimulant on sea bass (D. labrax) fed a diet supplemented with β-glucan (2%), ascorbic acid (500 ppm), and α-tocopherol (500 ppm). After 40 weeks of feeding (diets fed at 2% of body weight for 14 days with 3 months interval), the treated fish showed increased lysozyme activity (672 mg/ml) compared to the control group (455 mg/ml). Plasma complement activity was also found to be significantly high (868 ± 157 units/ml) in the treated group. Bagni et al. (2005) also conducted another research on sea bass where the fish were fed with alginic acid (0.5%) and MacroGard (0.1%) for 30 days. The experiment showed elevated serum lysozyme and complement activity, which returned to the control level after 45 days from the start of the trial. Some immune parameters (lysozyme activities and bactericidal activities) of the treated fish were found to be significantly higher at week 4 compared to week 6. In a separate study, Cuesta et al. (2007) injected whole yeast (S. cerevisiae, a single peritoneal injection of 107 cells ml-1) preparation to gilthead seabream (S. aurata) to assess the effect. The study showed increased respiratory burst and cytotoxic activity in peritoneal exudate leukocytes. The competent cells (monocytes and macrophages) started to increase their number after 24-h post-injection and reached the significant level after 48 to 72 h post-injection. A similar experiment was conducted by Cook et al. (2003) on snapper fish (Pagrus auratus) fed a diet containing EcoActiva (β-glucan-based immunostimulant) and showed increased macrophage respiratory burst activity through in vitro superoxide anion production on day 28 at 12°CC. Up to day 84, there was a higher burst activity but with no significant difference compared to the control. Castro et al. (1999) conducted a relevant experiment to assess the in vitro effect of MacroGard and Fibosel on respiratory burst activity of turbot (Psetta maxima) and gilthead seabream (S. aurata) where head kidney phagocytes were pre-incubated with β-glucan for 1, 3, and 6 h. MacroGard and Fibosel induced maximum responses when the cells were pre-incubated for 2 h with β-glucan (50–100 µg ml-1). However, the activity started to decrease with higher β-glucan concentration becoming significantly low at 500 µg ml-1. In contrast, Sealey et al. (2008) found no significant dietary effect on lysozyme and TNF-α mRNA expression when rainbow trout was fed a high amount barley β-glucan and commercial β-glucan (MacroGard, 2 g/kg diet).
Researchers have proven that β-glucan can be used as a vaccine adjuvant. Siwicki et al. (2004) conducted an investigation on rainbow trout (Oncorhynchus mykiss) fed diets containing β (1/3, 1/6) glucan (MacroGard) at a dose of 0.5% before immunization with anti-Yersinia ruckeri vaccine. The fish produced higher specific antibody secreting cells (ASC) and specific immunoglobulin levels in blood serum compared to control (provided vaccine only). The highest levels of specific antibody titers and highest number of ASCs were observed between 21 and 28 days after immunization (Table 1). Similar research was conducted by Sahoo and Mukherjee (2001) feeding 0.1% β (1/3) glucan to rohu fish (L. rohita). The fish raised some non-specific and specific immunity indices and resistance against Aeromonas hydrophila challenge compared to control fish. A remarkable rise (4.25 times higher than control group) in bacterial agglutination titer was shown by the glucan-fed fish group while the Aflatoxin B1-injected fish group showed a 10 times lower titer than the control group. The titer of the Aflatoxin-treated fish group was restored to the control level when they were supplemented with β (1/3) glucan in their diet. The serum bactericidal activity, the phagocytic ratio, and the leukocyte numbers were also significantly higher in glucan-fed fish. Selvaraj et al. (2005) obtained similar results in common carp (Cyprinus carpio) injected with β-glucan (100, 500, and 1,000 µg/fish). The treated fish showed a significant increase in total leukocyte counts and enhanced the proportion of neutrophils and monocytes. The fish pre-injected with β-glucan showed adjuvanted effect and ensured higher amount of antibody titers against vaccination with Aeromonas hydrophila. On the other hand, Parmar et al. (2012) conducted an experiment to assess the effect of brewer’s yeast on immune response and resistance of giant freshwater prawn, Macrobrachium rosenbergii, to white muscle disease. The prawns supplemented with 1% brewer’s yeast in their diet showed significantly higher (p < 0.05) phenoloxidase activity compared to the control. Significantly higher total hemocyte count (7.81 × 106 cells/ml) was also observed by this group. However, superoxide anion production differed significantly (p < 0.05) among the treatment groups, highest in prawns provided with 2% brewer’s yeast. Dawood et al. (2015) found a significant combined effect on immunity (lysozyme activity, serum bactericidal activity, and complement activity) when red sea bream (Pagrus major) was treated with heat-killed Lactobacillus plantarum (0.025%) and β-glucan (0.1%). Bonaldo et al. (2007) vaccinated sea bass (D. labrax) with Vibrio anguillarum and the medium dose (500 ppm β-glucan) group showed the highest antibody titers at the second week. Sealey et al. (2008) found that rainbow trout fed with a higher amount of barley β-glucan displayed higher antibody titers following vaccination with infectious hematopoietic necrosis virus (IHNV) than the fish fed commercial β-glucan, MacroGard. In contrast, some researchers found no adjuvanted effect of β-glucan parallel to fish vaccination. Welker et al. (2007) did not observe any rise in antibody titers in channel catfish when challenged with Edwardsiella ictaluri for 21 days.
Skjermo et al. (2006) conducted an experiment on Atlantic cod (Gadus morhua) fed algal (Chaetoceros mulleri) glucan chrysolaminaran (0.5 g/L rotifer culture), commercial β-glucan (MacroGard), and M-alginate to test the larval response. The stimulants were fed through rotifers and weaning from the supplemented diet was started on day 17 or 18 after hatching. The larvae fed C. mulleri glucan had higher survival and higher body weight at day 30. A similar experiment was also conducted by Misra et al. (2006) on rohu (L. rohita) fish to evaluate the effect of different doses of glucan on immunity, growth, and survival against Aeromonas hydrophila and Edwardsiella tarda. Doses between 250-500 mg β-glucan kg-1 diets showed better specific growth rate and food conversion ratio. Another study was conducted by Piaget et al. (2007) for six days post-hatch larvae of flounder (Paralicthys adspersus) to assess the effect of β-glucan and mannan oligosaccharide (applied first five days of the experiment at 5, 10, & 15 mg L-1 culture water) on larval growth and survival. A histological study of intestinal epithelium suggested that combined effect of β-glucan and mannan oligosaccharide enhanced monocytes expression. The immunostimulants had positive impact on the growth and survival of the larvae when provided with 5 mg L-l culture water. Dawood et al. (2015) tested red sea bream (Pagrus major) treated with heat-killed Lactobacillus plantarum (0.025%) and β-glucan (0.1%). The stimulants had significant combined effects on growth (feed intake, digestibility, growth rate, protein efficiency ratio, and plasma protein level). Kumari and Sahoo (2006) carried out a 30 day long investigation to assess the effect of lactoferrin (100 mg/kg feed), β (1/3) glucan (0.1%), levamisole (50 mg/kg feed) and vitamin C (500 mg/kg feed) on the immune function of Asian catfish (Clarias batrachus) vaccinated with formalin killed A. hydrophila. The study proved the stimulants efficient for inducing immunity against the bacteria and the immunosuppressant (the cyclophosphamide) injected peritoneally at a dose of 200 mg kg-1 body weight. These four substances significantly increased survival rates in both immunosuppressed and healthy vaccinated and non-vaccinated fish compared to their corresponding controls. On the contrary, channel catfish (Ictalurus punctatus) fed yeast subcomponents did not show a significant increase in survival percentage, though 5%–17.5% elevated survival was ensured in treated fish compared to the control ones (Welker et al., 2007). Sealey et al. (2008) conducted an investigation for 9 weeks on rainbow trout (O. mykiss) against three barley genotypes (with different amounts of β-glucan) regarding its growth and disease resistance. The high β-glucan barley diets (52 g/kg, 82 g/kg diet) had no significant effect on growth and feed conversion ratio compared to the lower amount of barley diet (38 g/kg diet) and the control diet (wheat supplemented).
β-glucan alone and/or coupled with other bioactive compounds (alginic acid, lactoferrin, mannan oligosaccharide, etc.) can be an effective immunostimulant. It may also be effective in improving specific immunity in fish for pursuing adjuvanticity. More than 3,000 papers have reported the effectiveness of β-glucan in improving fish immunity, but the detailed knowledge of the receptors involved in recognizing β-glucans and their downstream signaling mechanism is yet to be clarified in teleosts (Rodrigues et al., 2020). The β-glucans often differ in their activities. From thousands of published papers, it has been evidenced that β-glucans from different sources having similarities in structures, molecular weights, and solution strength can differ remarkably. Even the same β-glucan administering through immersion, dietary inclusion, or supplementary injection can respond in different ways (Chen and Seviour, 2007). For this, it is challenging to make a consensus on β-glucan sources, doses, and duration for an individual fish species.
Conceptualization, ZH and MH. Supervision, ZH. Writing—reviewing and editing, MH, MM, MS, SB, TM, and ZH. All authors have read and agreed to the published version of the manuscript.
This work was supported by the Krishi Gobeshona Foundation (KGF project ID: TF 40-F/17) under Bangladesh Agricultural Research Council (BARC). The research work was also supported by the Basic Science Research Program (2019R1A6A1A110 52070) to TM through the National Research Foundation of Korea (NRF) funded by the Ministry of Education.
The authors declare that the review work was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The authors wish to extend their immense gratitude to the authority of the Krishi Gobeshona Foundation (KGF project ID: TF 40-F/17), Bangladesh Agricultural Research Council, Farmgate, Dhaka, Bangladesh for their financial support and cordial cooperation to carry out the present study. The authors also offer thanks to the Department of Animal Biotechnology, Jeju International Animal Research Center (JIA) and Sustainable Agriculture Research Institute (SARI), Jeju National University for financial support to publish the review article. MM would like to acknowledge the National Research Foundation of Korea (NRF) for providing the postdoctoral research fellowship under the Brain Pool Program (Grant No. 2019H1D3A1A01101555) funded by the Ministry of Science, ICT, and Future Planning.
Adam E. L., Rice P. J., Graves B., Ensley H. E., Yu H., Brown G. D. (2008). Differential High Affinity Interaction of Dectin-1 With Natural or Synthetic Glucans Is Dependent Upon Primary Structure and is Influenced by Polymer Chain Length and Side Chain Branching. J. Pharmacol. Exp. Ther. 325 (1), 115–123. doi: 10.1124/jpet.107.133124
Aizawa M., Watanabe K., Tominari T., Matsumoto C., Hirata M., Grundler F. M. W., et al. (2018). Low Molecular-Weight Curdlan, (1→3)-Beta-GlucanSuppresses TLR2-Induced RANKL-Dependent Bone Resorption. Biol. Pharm. Bull. 41, 1282–1285. doi: 10.1248/bpb.b18-00057
Akhtar M. S., Tripathi P. H., Pandey A., Ciji A. (2021). β-Glucan Modulates non-Specific Immune Gene Expression, Thermal Tolerance and Elicits Disease Resistance in Endangered Tor Putitora Fry Challenged With Aeromonas Salmonicida. Fish Shell. Immunol. 119, 154–162. doi: 10.1016/j.fsi.2021.09.038
Akira S., Hemmi H. (2003). Recognition of Pathogen Associated Molecular Patterns by TLR Family. Immunol.Lett. 85, 85–95. doi: 10.1016/S0165-2478(02)00228-6
Akramiene D., Kondrotas A., Didziapatriene J., Kevelaitis E. (2007). Effects of Beta-Glucans on the Immune System. Med. Lith. 43, 597–606. doi.org/10.3390/medicina43080076
Alberts B., Johnson A., Lewis J., Raff M., Roberts K., Walters P. (2002). Molecular Biology of the Cell. 4th ed. (New York and London: Garland Science), ISBN: ISBN-10: 0-8153-3218-1.
Bagni M., Archetti L., Amadori M., Marino G. (2000). Effect of Long-Term Oral Administration of an Immunostimulant Diet on Innate Immunity in Sea Bass (Dicentrarchus Labrax). J. Vet. Med. B. 47, 745–751. doi: 10.1046/j.1439-0450.2000.00412.x
Bagni M., Romano N., Finoia M. G., Abelli L., Scapigliati G., Tiscar P. G. (2005). Short- and Long-Term Effects of a Dietary Yeast β-Glucan (Macrogard) and Alginic Acid (Ergosan) Preparation on Immune Response in Sea Bass (Dicentrarchus Labrax). Fish Shell. Immunol 18, 311–325. doi: 10.1016/j.fsi.2004.08.003
Banchereau J., Steinman R. M. (1998). Dendritic Cells and Control of Immunity. Nature 392 (6673), 245–252. doi.org/10.1038/32588
Blystone S. D., Brown E. J. (1999). Integrin Receptors of Phagocytes. Adv. Cell Mol. Biol. Membranes Organdies 5, 103–147. doi: 10.1016/S1874-5172(99)80030-3
Bonaldo A., Thompson K. D., Manfrin A., Adams A., Murano E., Mordenti A. L. (2007). The Influence of Dietary β-Glucans on the Adaptive and Innate Immune Responses of European Sea Bass (Dicentrarchus Labrax) Vaccinated Against Vibriosis. Ital. J. Anim. Sci. 6, 151–164. doi: 10.4081/ijas.2007.151
Borchers A. T., Keen C. L., Gershwin M. E. (2004). Mushrooms, Tumors, and Immunity: An Update. Exp. Biol. Med. 229, 393–406. doi: 10.1177/153537020422900507
Bricknell I, Dalmo R. A. and . (2005). The Use of Immunostimulants in Fish Larval Aquaculture. Fish. Shellfish Immunol. 19, 457–472. doi: 10.1016/j.fsi. 2005.03.008
Brown G. D., Gordon S. (2003). Fungal β-Glucans and Mammalian Immunity. Immunity 19, 311–315. doi: 10.1016/S1074-7613(03)00233-4
Brown G. D., Herre J., Williams D. L., Willment J. A., Marshall A. S. J., Gordon S. (2003). Dectin-1 Mediates the Biological Effects of Beta-Glucans. J. Exp. Med. 197 (9), 1119–1124. doi: 10.1084/jem.20021890
Buckeridge M. S., Rayon C., Urbanowicz B., Tine M. A. S., Carpita N. C. (2004). Mixed Linkage (1→3) (1→4) - β-D-Glucans of Grasses. Cereal Chem. 81 (1), 115–127. doi: 10.1094/CCHEM.2004.81.1.115
Butt M. S., Tahir-Nadeem M., Khan M. K. I., Shabir R. (2008). Oat: Unique Among the Cereals. Eur. J. Nutr. 47, 68–79. doi: 10.1007/s00394-008-0698-7
Castro R., Couso N., Obach A., Lamas J. (1999). Effect of Different β-Glucans on the Respiratory Burst of Turbot (Psetta Maxima) and Gilthead Seabream (Sparusaurata) Phagocytes. Fish Shellfish Immunol. 9, 529–541. doi: 10.1006/fsim.1999.0210
Chang C. F., Chen H. Y., Su M. S., Liao I. C. (2000). Immunomodulation by Dietary β-1, 3-Glucan in the Brooders of the Black Tiger Shrimp Penaeus Monodon. Fish Shellfish Immunol. 10, 505–514. doi: 10.1006/fsim.2000.0266
Chang C. S., Huang S. L., Chen S. (2013). Innate Immune Responses and Efficacy of Using Mushroom Beta-Glucan Mixer (MBG) on Orange-Spotted Grouper, Epinephelus Coioides, Aquaculture. Fish Shellfish Immunol. 35, 115–125. doi: 10.1016/j.fsi.2013.04.004
Chang C. F., Sub M. S., Chenc H. Y., Liao I. (2003). Dietary β-1,3-Glucan Effectively Improves Immunity and Survival of Penaeusmonodon Challenged With White Spot Syndrome Virus. Fish Shellfish Immunol. 15, 297–310. doi: 10.1016/S1050-4648(02)00167-5
Chen J., Seviour R. (2007). Medicinal Importance of Fungal Beta-(1→3), (1→6)-Glucans. Mycol. Res. 111, 635–652. doi: 10.1016/j.mycres.2007.02.011
Chotikachinda R., Lapjatupon W., Chaisilapasung S., Sangsue D., Tantikitt C. (2008). Effect of Inactive Yeast Cell Wall on Growth Performance, Survival Rate and Immune Parameters in Pacific White Shrimp (Litopenaeus Vannamei). Songklanakarin J. Sci. Technol. 30 (6), 687–692.
Chu Y. (2014). Oats Nutrition and Technology (Barrington, Illinois: Wiley Blackwell), 35411–35417, ISBN: ISBN 978-1-118.
Cook M. T., Hayball P. J., Hutchinson W., Nowak B. F., Hayball J. D. (2003). Administration of a Commercial Immunostimulant Preparation, EcoActivaTM as a Feed Supplement Enhances Macrophage Respiratory Burst and Growth Rate of Snapper (Pagrus Auratus, Sparidae (Bloch and Schneider)) in Winter. Fish Shellfish Immunol. 14, 333–345. doi: 10.1006/fsim.2002.0441
Cornet V., Khuyen T. D., Mandiki S. N. M., Betoulle S., Bossier P., Reyes-Lopez F. E., et al. (2021). GAS1: A New B-Glucan Immunostimulant Candidate to Increase Rainbow Trout (Oncorhynchus Mykiss) Resistance to Bacterial Infections With Aeromonas Salmonicida Achromogenes. Front. Immunol. 12, 693613. doi: 10.3389/fimmu.2021.693613
Costa M. M., Novoa B., Figueras A. (2008). Influence of B-Glucans on the Immune Responses of Carpet Shell Clam (Ruditapes Decussatus) and Mediterranean Mussel (Mytilusgallo Provincialis). Fish Shellfish Immunol. 24, 498–505. doi: 10.1016/j.fsi.2007.10.003
Cuesta A., Meseguer J., Esteban M. A. (2004). Total Serum Immunoglobulin M Levels are Affected by Immunomodulators in Seabream (Sparus Aurata L.). Veterinary Immunol. Immunopathol. 101, 203–210. doi: 10.1016/j.vetimm.2004.04.021
Cuesta A., Rodrıguez A., Salinas I., Meseguer J., Esteban M. A. (2007). Early Local and Systemic Innate Immune Responses in the Teleost Gilthead Seabream After Intraperitoneal Injection of Whole Yeast Cells. Fish Shellfish Immunol. 22, 242–251. doi: 10.1016/j.fsi.2006.05.005
Danilova N., Bussmann J., Jekosch K., Steiner L. A. (2005). The Immunoglobulin Heavy-Chain Locus in Zebrafish: Identification and Expression of a Previously Unknown Isotype, Immunoglobulin Z. Nat. Immunol. 6, 295–302. doi: 10.1038/ni1166
Das B. K., Debnath C., Patnaik P., Swain D. K., Bibudhendra K. K., Misrhra K. (2009). Effect of β-Glucan on Immunity and Survival of Early Stage of Anabas Testudineus (Bloch). Fish Shellfish Immunol. 27, 1–6. doi: 10.1016/j.fsi.2009.08.002
Dawood M. A., Koshio S., Ishikawa M., Yokoyama S. (2015). Interaction Effects of Dietary Supplementation of Heat-Killed Lactobacillus Plantarum and Beta-Glucan on Growth Performance, Digestibility and Immune Response of Juvenile Red Sea Bream, Pagrus Major. Fish Shellfish Immunol. 45, 33–42. doi: 10.1016/j.fsi.2015.01.033
Dawood M. A., Metwally A. E. S., El-Sharawy M. E., Atta A. M., Elbialy Z. I., Abdel-Latif H. M. R., et al. (2020). The Role of β-Glucan in the Growth, Intestinal Morphometry, and Immune Related Gene and Heat Shock Protein Expressions of Nile Tilapia (Oreochromis Niloticus) Under Different Stocking Densities. Aquaculture 523, 735205. doi: 10.1016/j.aquaculture.2020.735205
Dennehy K. M., Brown G. D. (2007). The Role of the β-Glucan Receptor Dectin-1 in Control of Fungal Infection. J. Leukoc. Biol. 82, 253–258. doi: 10.1189/jlb.1206753
DeVries J. W. (2003). On Defining Dietary Fiber. Proc. Nutr. Society. 62, 37–43. doi: 10.1079/PNS2002234
Di Luzio N. (1985). Update on the Immunomodulating Activities of Glucans. Springer Semin. Immunopathol. 8, 387–400. doi: 10.1007/BF01857392
Djordjevic B., Skugor S., Jorgensen S. M., Overland M., Myland L. T., Krasnov A. (2009). Modulation of Splenic Immune Response to Bacterial Lipopolysaccharide in Rainbow Trout (Oncorhynchus Mykiss) Fed Lentinin, a β-Glucan From Mushroom Lentinula Edodes. Fish Shell.Immunol. 26 (2), 201–209. doi: 10.1016/j.fsi.2008.10.012
Domenico J. D., Canova R., Soveral L. D. F., Nied C. O., Costa M. M., Frandoloso R., et al. (2017). Immunomodulatory Effects of Dietary β-Glucan in Silver Catfish (Rhamdia Quelen). Pesq. Vet. Bras. 37 (1), 73–78. doi: 10.1590/s0100-736x2017000100012
El-boshy M. E., El-Ashram A. M., AbdelHamd F. M., Gadalla H. A. (2010). Immunomodulatory Effect of Dietary Saccharomyces Cerevisiae, Beta-Glucan and Laminaran in Mercuric Chloride Treated Nile Tilapia (Oreochromis Niloticus) and Experimentally Infected With Aeromonas Hydrophila. Fish Shellfish Immunol. 28, 802–808. doi: 10.1016/j.fsi.2010.01.017
Evans S. E., Hahn P. Y., McCann F., Kottom T. J., Pavlovic Z. V., Limper A. H. (2005). Pneumocystis Cell Wall β-Glucans Stimulate Alveolar Epithelial Cell Chemokine Generation Through Nuclear Factor-B–Dependent Mechanisms. Am. J. Respir. Cell Mol. Biol. 32, 490–497. doi: 10.1165/rcmb.2004-0300OC
FAO (2002). “Antibiotics Residue in Aquaculture Products,” in The State of World Fisheries and Aquaculture(Rome, Italy), Food and Agriculture Organization of the United Nations, pp 74–pp 82.
Figueras A., Santarem M. M., Nov B. (1998). Influence of the Sequence of Administration of B-Glucans and a Vibrio Damsela Vaccine on the Immune Response of Turbot (Scophthalmus Maximus L.). Vet. Immunol. Immunopathol. 64 (1), 59–68. doi: 10.1016/S0165-2427(98)00114-7
Filho F. D. O. R., Koch J. F. A., Wallace C., Leal M. C. (2019). Dietary β-1,3/1,6-Glucans Improve the Effect of a Multivalent Vaccine in Atlantic Salmon Infected With Moritella Viscosa or Infectious Salmon Anemia Virus. Aquaculture Int. 27, 1825–1834. doi: 10.1007/s10499-019-00436-9
Gatlin D. M., Li P. (2004). Dietary Supplementation of Prebiotics for Haelth Management of Hybrid Striped Bass Morone Chrysops X M. Saxatilis. Aqua Feeds Formul Beyond. 1 (4), 19–21.
Goodridge H. S., Wolf A. J., Underhill D. M. (2009). Beta-Glucan Recognition by the Innate Immune System. Immunol. Rev. 230, 38–50. doi: 10.1111/j.1600-065X.2009.00793.x
Johansson L., Karesoja M., Ekholm P. (2008). Comparison of the Solution Properties of (1→3),(1→4)-β-D-Glucans Extracted From Oats and Barley. LWT. 41, 180–184. doi: 10.1016/j.lwt.2007.01.012
Kamilya D., Maiti T. K., Joardar S. N., Mal B. C. (2006). Adjuvant Effect of Mushroom Glucan and Bovine Lactoferrin Upon Aeromonas Hydrophila Vaccination in Catla, Catla (Hamilton). J. Fish Dis. 29, 331–337. doi: 10.1111/j.1365-2761.2006.00722.x
Kawakami H., Shinohara N., Sakai M. (1998). The non-Specific Immunostimulation and Adjuvant Effects of Vibrio Anguillarum Bacterin, M-Glucan, Chitin and Freund’s Complete Adjuvant Against Pasteurella Piscicida Infection in Yellowtail. Fish Pathol. 33, 287–292. doi: 10.3147/jsfp.33.287
Kim Y., Ke F., Zhang Q. (2009). Effect of B-Glucan on Activity of Antioxidant Enzymes and Mx Gene Expression in Virus Infected Grass Carp. Fish Shellfish Immunol. 27, 336–340. doi: 10.1016/j.fsi.2009.06.006
Kuhlwein H., Emery M. J., Rawling M. D., Harper G. M., Merrifield D. L., Davies S. J. (2013). Effects of a Dietary Beta-(1,3)(1,6)-D-Glucan Supplementation on Intestinal Microbial Communities and Intestinal Ultrastructure of Mirror Carp (Cyprinus Carpio L.). J. Appl. Microbiol. 115, 1091–1106. doi: 10.1111/jam.12313
Kumagai Y., Akira S. (2010). Identification and Functions of Pattern-Recognition Receptors. J. Aller. Clin.Immunol. 125, 985–992. doi: 10.1016/j.jaci.2010.01.058
Kumari J., Sahoo P. K. (2006). Non-Specific Immune Response of Healthy and Immunocompromised Asian Catfish (Clarias Batrachus) to Several Immunostimulants. Aquaculture 255, 133–141. doi: 10.1016/j.aquaculture.2005.12.012
Lebron F., Vassallo R., Puri V., Limper A. H. (2003). Pneumocystis Carinii Cell Wall β-Glucans Initiate Macrophage Inflammatory Responses Through NF-Kappa B Activation. J. Biol. Chem. 278 (4), 25001–25008. doi: 10.1074/jbc.M301426200
Lee D. Y., Ji I. H., Chang H. I., Kim C. W. (2002). High-Level TNF-A Secretion and Macrophage Activity With Soluble β-Glucans From Saccharomyces Cerevisiae. Biosci. Biotechnol. Biochem. 66, 233–238. doi: 10.1271/bbb.66.233
Lesage G., Bussey H. (2006). Cell Wall Assembly in Saccharomyces Cerevisiae.Microbiol. Mol. Biol. Rev. 70, 317–343. doi: 10.1128/MMBR.00038-05
Magnadottir B. (2006). Innate Immunity of Fish (Overview). Fish Shellfish Immunol. 20, 137–151. doi: 10.1016/j.fsi.2004.09.006
Matzinger P. (2002). The Danger Model: A Renewed Sense of Self (PDF). Science 296 (5566), 301–305. doi.org/10.1126/science.1071059
McIntosh M., Stone B. A., Stanisich V. A. (2005). Curdlan and Other Bacterial (1/3)-Beta-D-Glucans. Appl. Microbiol. Biotechnol. 68 (2), 163–173. doi: 10.1007/s00253-005-1959-5
Medzhitov R. (2007). Recognition of Microorganisms and Activation of the Immune Response. Nature 449 (7164), 819–826. doi: 10.1038/nature06246
Meena D. K., Das P., Kumar S., Mandal S. C., Prusty A. K., Singh S. K., et al. (2013). Beta Glucan: An Ideal Immunostimulant in Aquaculture (a Review). Fish Physiol. Biochem. 39, 431–457. doi: 10.1007/s10695-012-9710-5
Meshram S. J., Murthy S., Swain H. S., Ali H., Jagadesh T. D., Dhamgaye H. B. (2014). Preliminary Study of the Effect of Dietary Beta-Glucan on the Gut Morphology of Giant Freshwater Prawn, Macrobrachium Rosenbergii. Int. J. Life Sci. 9 (2), 543–546.
Misra C. K., Das B. K., Mukherjee S. C., Pattnaik P. (2006). Effect of Long Term Administration of Dietary β-Glucan on Immunity, Growth and Survival of Labeo Rohita Fingerlings. Aquaculture 255, 82–94. doi: 10.1016/j.aquaculture.2005.12.009
Miura N. N., Ohno N., Aketagawa J., Tamura H., Tanaka S., Yadomae T. (1996). Blood Clearance of (1–3)-Beta-D-Glucan in MRL Lpr/Lpr Mice. FEMS Immunol. Med. Microbiol. 13 (1), 51–57. doi.org/10.1111/j.1574-695X.1996.tb00215.x
Mueller A., Raptis J., Rice P. J., Kalbfleisch J. H., Stout R. D., Ensley H. E. (2000). The Influence of Glucan Polymer Structure and Solution Conformation on Binding to (1/3)-β-D-Glucan Receptors in a Human Monocyte- Like Cell Line. Glycobiology 10, 339–346. doi: 10.1093/glycob/10.4.339
Murphy K., Weaver C. (2017). Janeway’s Immunobiology. 9th edition, Garland science, Taylor and Francis group, New York, USA), ISBN: ISBN 978-0-8153-4505-3.
Nakayama H., Yoshizaki F., Prinetti A., Sonnino S., Mouri L., Takamori K. (2008). Lyn-Coupled LacCer-Enriched Lipid Rafts are Required for CD11b⁄CD18-Mediated Neutrophil Phagocytosis of Nonopsonized Microorganisms. J. Leukoc. Biol. 83, 728–741. doi: 10.1189/jlb.0707478
Ooi V. E. C., Liu F. (2000). Immunomodulation and Anti-Cancer Activity of Polysaccharide-Protein Complexes. Curr. Med. Chem. 7, 715–729. doi: 10.2174/0929867003374705
Ortuno J, Cuesta A., Rodriguez A., Esteban M. A., Meseguer J., (2002). Oral Administration of Yeast, Saccharomyces Cerevisiae, Enhances the Cellular Innate Immune Response of Gilthead Seabream (Sparus Aurata L.). Vet. Immunol. Immunopathology 85, 41–50. doi: 10.1016/S0165-2427(01)00406-8
Palic D., Andreasen C. B., Dawn M. H., Menzeld B. W., Roth J. A. (2006). Immunomodulatory Effects of B-Glucan on Neutrophil Function in Fathead Minnows (Pimephales Promelas Rafinesque 1820). Dev. Comp. Immunol. 30, 817–830. doi: 10.1016/j.dci.2005.11.004
Pal D., Siddhartha N., Joardar S. N., Roy B. (2007). Immunostimulatory Effects of a Yeast (Saccharomyces Cerevisiae) Cell Wall Feed Supplement on Rohu (Labeo Rohita), an Indian Major Carp. Israeli J. Aquacult. –Bamidgeh 59 (3), 175–181. doi: 10.46989/001c.20525
Parmar P. V., Murthy H. S., Tejpal C. S., Kumar B. T. N. (2012). Effect of Brewer’s Yeast on Immune Response of Giant Freshwater Prawn, Macrobrachium Rosenbergii, and its Resistance to White Muscle Disease. Aquacult. Int. 20, 951–964. doi: 10.1007/s10499-012-9520-x
Patten D. A., Shetty S. (2018). More Than Just a Removal Service: Scavenger Receptors in Leukocyte Trafficking. Front. Immunol. 9, 2904. doi: 10.3389/fimmu.2018.02904
Paulsen S. M., Engstad R. E., Robertsen B. (2001). Enhanced Lysozyme Production in Atlantic Salmon (Salmo Salar L.) Macrophages Treated With Yeast β-Glucan and Bacterial Lipopolysaccharide. Fish Shellfish Immunol. 11, 23–37. doi: 10.1006/fsim.2000.0291
Petit J., Bailey E. C., Wheeler R. T., de Oliveira C. A. F., Forlenza M., Wiegertjes G. F. (2019). Studies Into Beta-Glucan Recognition in Fish Suggests a Key Role for the C-Type Lectin Pathway. Front. Immunol. 10, 280. doi: 10.3389/fimmu.2019.00280
Petit J., de Bruijn I., Goldman M. R. G., van den Brink E., Pellikaan W. F., Forlenza M., et al. (2022). β-Glucan-Induced Immuno Modulation: A Role for the Intestinal Microbiota and Short-Chain Fatty Acids in Common Carp. Front. Immunol. 12, 761820. doi: 10.3389/fimmu.2021.761820
Petit J., Wiegertjes G. F. (2016). Long-Lived Effects of Administering β-Glucans: Indications for Trained Immunity in Fish. J. Dev. Comp. Immunol. 64, 93–102. doi: 10.1016/j.dci.2016.03.003
Piaget N., Vegal A., Silva A., Toledo P. (2007). Effect of the Application of β-Glucans and Mannan-Oligosaccharides (βg MOS) in an Intensive Larval Rearing System of Paralichthys Adspersus (Paralichthydae). Invest. Mar. Valparaiso. 35 (2), 35–43. doi: 10.4067/S0717-71782007000200004
PrabhuDas D. M. R., Baldwin C. L., Bollyky P. L., Bowdish D. M. E., Drickamer K., Febbraio M. (2017). A Consensus Definitive Classification of Scavenger Receptors and Their Roles in Health and Disease. J. Immunol. 198 (10), 3775–3789. doi: 10.4049/jimmunol.1700373
Rawles S. D., Kocabas A., Gatlin D. M., Du W. X., Wei C. I. (1997). Dietary Supplementation of Terramycin and Romet-30 Does Not Enhance Growth of Channel Catfish But Does Influence Tissue Residues. J. World Aquac. Soc 28, 392–401. doi: 10.1111/j.1749-7345.1997.tb00286.x
Reese A. J., Yoneda A., Breger J. A., Beauvais A., Liu H., Griffith C. L. (2007). Loss of Cell Wall Alpha(1-3) Glucan Affects Cryptococcus Neoformansfrom Ultrastructure to Virulence. Mol. Microb. 63 (5), 1385–1398. doi: 10.1111/j.1365-2958.2006.05551.x
Refstie S., Baeverfjord G., Seim R. R., Elvebo O. (2010). Effects of Dietary Yeast Cell Walls Beta-Glucans and MOS on Performance, Gut Health, and Salmon Lice Resistance in Atlantic Salmon (Salmo Salar) Fed Sunflower and Soybean Meal. Aquaculture 305, 109–116. doi: 10.1016/j.aquaculture.2010.04.005
Rice P. J., Kelley J. L., Kogan G., Ensley H. E., Kalbfleisch H. J., Browder I. W. (2002). Human Monocyte Scavenger Receptors are Pattern Recognition Receptors for (1/3)-Beta-D-Glucans. J. Leukoc. Biol. 72, 140–146. doi.org/10.1189/jlb.72.1.140
Rioux L. E., Turgeon S. L., Beaulieu M. (2007). Characterization of Polysaccharides Extracted From Brown Seaweeds. Carbohydr. Pol. 69, 530–537. doi: 10.1016/j.carbpol.2007.01.009
Rodrigues M. V., Zanuzzo F. S., Koch J. F. A., de Oliveira C. A. F., Sima P., Vetvicka V. (2020). Development of Fish Immunity and the Role of β-Glucan in Immune Responses. Molecules 25, 5378. doi: 10.3390/molecules25225378
Rodriguez K. N. N., Alvarez-González A. C., Pena-Marin E. S., Vega-Villasante F., Martinez-Garcia R., Camarillo-Coop S., et al. (2018). Effect of β-Glucans in Diets on Growth, Survival, Digestive Enzyme Activity, and Immune System and Intestinal Barrier Gene Expression for Tropical Gar (Atractosteus Tropicus) Juveniles. Fishes 3 (27), 1–13. doi:10.3390/fishes3030027
Rodriguez I., Chamorro R., Novoa B., Figueras A. (2009). β-Glucan Administration Enhances Disease Resistance and Some Innate Immune Responses in Zebrafish (Danio Rerio). Fish Shell.Immunol. 27, 369–373. doi: 10.1016/j.fsi.2009.02.007
Rodriguez A., Cuesta A., Ortuno J., Esteban M. A., Meseguer J. (2003). Immunostimulant Properties of a Cell Wall-Modified Whole Saccharomyces Cerevisiae Strain Administered by Diet to Seabream (Sparus Aurata L.). Vet. Immunol. Immunopathol. 96, 183–192. doi: 10.1016/j.vetimm.2003.07.001
Sahoo P. K., Mukherjee S. C. (2001). Effect of Dietary β-1,3glucan on Immune Responses and Disease Resistance of Healthy and Aflatoxin B1-Induced Immuno- Compromisedrohu (Labeo Rohita, Hamilton). Fish Shellfish Immunol. 11, 683–695. doi: 10.1006/fsim.2001.0345
Sakai M. (1999). Current Research Status of Fish Immunostimulants. Aquaculture 172, 63–92. doi: 10.1016/S0044-8486(98)00436-0
Sang H. M., Fotedar R. (2010). Effects of Dietary β (1,3) Glucan on the Growth, Survival, Physiological and Immune Response of Marron, Cherax Tenuimanus (Smith 1912). Fish Shellfish Immunol. 28, 957–960. doi: 10.1016/j.fsi.2010.01.020
Sealey W. M., Barrows F. T., Hang A., Johansen K. A., Overturf K., LaPatra S. E. (2008). Evaluation of the Ability of Barley Genotypes Containing Different Amount of B-Glucan to Alter Growth and Disease Resistance of Rainbow Trout (Oncorhynchus Mykiss). Anim. Feed Sci. Tech. 141 (1–2), 115–128. doi: 10.1016/j.anifeedsci.2007.05.022
Selvaraj V., Sampath K., Sekar V. (2005). Administration of Yeast Glucan Enhances Survival and Some non-Specific and Specific Immune Parameters in Carp (Cyprinus Carpio) Infected With Aeromonas Hydrophila. Fish Shell.Immunol. 19 (4), 293–306. doi: 10.1016/j.fsi.2005.01.001
Sherif A. H., Mahfouz M. E. (2019). Immune Status of Oreochromis Niloticus Experimentally Infected With Aeromonas Hydrophila Following Feeding With 1, 3 β-Glucan and Levamisole Immunostimulants. Aquaculture 509, 40–46. doi: 10.1016/j.aquaculture.2019.05.016
Siwicki A. K., Kazun K., Edward G., Terech-Majewska E., Baranowski P., Trapkowska S. (2004). The Effect of Beta-1.3/1.6–Glucan in Diets on the Effectiveness of Anti-Yersinia Ruckeri Vaccine–an Experimental Study in Rainbow Trout (Oncorhynchus Mykiss). Pol. J. Food Nutr. Sci. 2, 59–61. 13/54, SI.
Siwicki A. K., Morand M., Terech-Majevska E., Niemczuk W., Kazun K., Glabsky E. (1998). Influence of Immunostimulant on the Effectiveness of Vaccines in Fish: In Vitro and In Vivo Study. J. Appl. Ichthyol. 14, 225–227. doi: 10.1111/j.1439-0426.1998.tb00646.x
Skjermo J., Trond R., Hansen S. K., Handa A., Oie G. (2006). Evaluation of β-(1→3, 1→6)-Glucans and High-M Alginate Used as Immunostimulatory Dietary Supplement During First Feeding and Weaning of Atlantic Cod (Gadus Morhua L.). Aquaculture 261, 1088–1101. doi: 10.1016/j.aquaculture.2006.07.035
Solem S. T., Stenvik J. (2006). Antibody Repertoire Development in Teleosts—A Review With Emphasis on Salmonids and Gadus Morhua L. Dev. Comp. Immunol. 30, 57–76. doi: 10.1016/j.dci.2005.06.007
Suphantharika M., Khunrae P., Thanardkit P., Verduyn C. (2003). Preparation of Spent Brewer’s Yeast β-Glucans With a Potential Application as an Immunostimulant for Black Tiger Shrimp, Penaeus Monodon. Biores. Technol. 88, 55–60. doi: 10.1016/S0960-8524(02)00257-2
Swennen K., Courtin C. A., Delcour J. A. (2006). Non-Digestible Oligosaccharides With Prebiotic Properties. Crit. Rev. Food. Sci. Nutr. 46, 459–471. doi: 10.1080/10408390500215746
Taylor P. R., Brown G. D., Reid D. M., Willment J. A., Martinez-Pomares L., Gordon S. (2002). The β-Glucan Receptor, Dectin-1, is Predominantly Expressed on the Surface of Cells of the Monocyte/Macrophage and Neutrophil Lineages. J. Immunol. 169, 3876–3882. doi: 10.4049/jimmunol.169.7.3876
Taylor P. R., Tsoni S. V., Willment J. A., Dennehy K. M., Rosas M., Findon H. (2007). Dectin-1 is Required for Beta-Glucan Recognition and Control of Fungal Infection. Nat. Immunol. 8, 31–38. doi: 10.1038/ni1408
Vereschagin E. I., Van Lambalgen A. A., Dushkin M. I., Schwartz Y. S., Polyakov L., Heemskerk A. (1998). Soluble Glucan Protects Against Endotoxin Shock in the Rat: The Role of the Scavenger Receptor. Shock 9 (3), 193–198. doi: 10.1097/00024382-199803000-00006
Vetvicka V., Thornton B. P., Ross G. D. (1996). Soluble Beta-Glucan Polysaccharide Binding to the Lectin Site of Neutrophils or Natural Killer Cell Complement Receptor Type 3 (CD11b/ CD18) Generates a Primed State of the Receptor Capable of Mediating Cytotoxicity of Ic3b-Opsonized Target Cells. J. Clin. Investig. 98, 50–61. doi: 10.1172/JCI118777
Volman J. J., Ramakers J. D., Plat J. (2008). Dietary Modulation of Immune Function by β-Glucans. Physiol. Behav. 94 (2), 276–284. doi: 10.1016/j.physbeh.2007.11.045
Wakshull E., Brunke-Reese D., Lindermuth J., Fisette L., Nathans R. S., Crowley J. J. (1999). PGG-Glucan, a Soluble β-1,3 -Glucan, Enhances the Oxidative Burst Response, Microbicidal Activity, and Activates an NF-kB-Like Factor in Human PMN: Evidence for a Glycosphingolipid β-1,3-Glucan Receptor. Immunopharmacology 41, 89–107. doi: 10.1016/S0162-3109(98)00059-9
Wasser S. P., Weis A. L. (1999). Therapeutic Effects of Substances Occurring in Higher Basidiomycetes Mushrooms: A Modern Perspective. Crit. Rev. Immunol. 19 (1), 65–96. doi: 10.1615/CritRevImmunol.v19.i1.30
Welker T. L., Lim C., Yildrim-Aksoy M., Shelby R., Klesius P. H. (2007). Immune Response and Resistance to Stress and Edwardsiella Ictaluri Challenge in Channel Catfish, Ictalurus Punctatus, Fed Diets Containing Commercial Whole-Cell or Yeast Subcomponents. J. World Aquac. Soc 38, 24–35. doi: 10.1111/j.1749-7345.2006.00070.x
Werner J. L., Metz A. E., Horn D., Schoeb T. R., Hewitt M. M., Schwiebert L. M. (2009). Requisite Role for the Dectin-1 Beta-Glucan Receptor in Pulmonary Defense Against Aspergillus Fumigatus. J. Immunol. 182, 4938–4946. doi: 10.4049/jimmunol.0804250
Whittington R., Lim C., Klesius P. H. (2005). Effect of Dietary β-Glucan Levels on the Growth Response and Efficacy of Streptococcus Iniae Vaccine in Nile Tilapia, Oreochromis Niloticus. Aquaculture 248, 217–225. doi: 10.1016/j.aquaculture.2005.04.013
Willment J. A., Gordon S., Brown G. D. (2001). Characterization of the Human Beta-Glucan Receptor and its Alternatively Spliced Isoforms. J. Biol. Chem. 76, 43818–43823. doi: 10.1074/jbc.M107715200
Xia Y., Ross G. D. (1999). Generation of Recombinant Fragments of CD11b Expressing the Functional Beta-Glucan-Binding Lectin Site of CR3 (CD11b/ Cd18). J. Immunol. 162, 7285–7293.
Yadav M., Schorey J. S. (2006). The Beta-Glucan Receptor Dectin-1 Functions ogether With TLR2 to Mediate Macrophage Activation by Mycobacteria. Blood 108, 3168–3175. doi: 10.1182/blood-2006-05-024406
Yamamoto F. Y., Castillo S., de Cruz C. R., Chen K., Hume M. E., Gatlin III, D.M. (2020). Synergistic Effects of the β-1,3 Glucan Paramylon and Vitamin C on Immunological Responses of Hybrid Striped Bass (Morone Chrysops × M. Saxatilis) Were Pronounced In Vitro But More Moderate In Vivo. Aquaculture 526, 735394. doi.org/10.1016/j.aquaculture.2020.735394
Yamamoto F. Y., Sutili F. J., Hume M., Gatlin III, D.M. (2018a). The Effect of β-1,3-Glucan Derived From Euglena Gracilis (Algamune™) on the Innate Immunological Responses of Nile Tilapia (Oreochromis Niloticus L.). J. Fish Dis. 41, 1579–1588. doi.org/10.1111/jfd.12871
Yamamoto F. Y., Yin F., Rossi W. Jr., Hume M., Gatlin III, D.M. (2018b). β-1,3 Glucan Derived From Euglena Gracilis and Algamune™ Enhances Innate Immune Responses of Red Drum (Sciaenops Ocellatus L.). Fish Shell. Immunol. 77, 273–279. doi.org/10.1016/j.fsi.2018.04.003
Zimmerman J. W., Lindermuth J., Fish P. A., Palace G. P., Stevenson T. T., DeMong D. E. (1998). A Novel Carbohydrate Glycosphingolipid Interaction Between a Beta-(1–3)-Glucanimmunomodulator, PGG-Glucan, and Lactosylceramide of Human Leukocytes. J. Biol. Chem. 273 (34), 22014–22020.
Zlatkovic D., Jakovljevic D., Zekovic D., Vrvic M. M. (2003). A Glucan From Active Dry Baker’s Yeast (Sachharomyces Cerevisiae): A Chemical and Enzymatic Investigation of the Structure. J. Serb. Chem. Soc 68 (11), 805–809. doi: 10.2298/JSC0311805Z
Keywords: beta-glucan, mechanism of action, fish growth, fish health, immunomodulation, aquaculture
Citation: Hadiuzzaman M, Moniruzzaman M, Shahjahan M, Bai SC, Min T and Hossain Z (2022) β-Glucan: Mode of Action and Its Uses in Fish Immunomodulation. Front. Mar. Sci. 9:905986. doi: 10.3389/fmars.2022.905986
Received: 28 March 2022; Accepted: 09 June 2022;
Published: 15 July 2022.
Edited by:
Min Jin, Ningbo University, ChinaReviewed by:
Miriam Angulo, Centro de Investigación Biológica del Noroeste (CIBNOR), MexicoCopyright © 2022 Hadiuzzaman, Moniruzzaman, Shahjahan, Bai, Min and Hossain. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Taesun Min, dHNtaW5AamVqdW51LmFjLmty; Zakir Hossain, emFraXIuZmJnQGJhdS5lZHUuYmQ=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.