 Susannah J. Buchan1,2,3,4*Laura Gutiérrez1,5Mark F. Baumgartner4Kathleen M. Stafford6Nadin Ramirez7Oscar Pizarro7,8Jose Cifuentes1
Susannah J. Buchan1,2,3,4*Laura Gutiérrez1,5Mark F. Baumgartner4Kathleen M. Stafford6Nadin Ramirez7Oscar Pizarro7,8Jose Cifuentes1- 1Center for Oceanographic Research COPAS Sur-Austral and COPAS COASTAL, Universidad de Concepción, Concepción, Chile
- 2Departamento de Oceanografía, Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción, Concepción, Chile
- 3Centro de Estudios Avanzados en Zonas Áridas (CEAZA), La Serena, Chile
- 4Biology Department, Woods Hole Oceanographic Institution, Woods Hole, MA, United States
- 5Graduate Program in Oceanography, Departamento de Oceanografía, Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción, Concepción, Chile
- 6Marine Mammal Institute, Oregon State University, Newport, OR, United States
- 7Instituto Milenio de Oceanografía, Universidad de Concepción, Concepción, Chile
- 8Departamento de Geofísica, Facultad de Ciencias Físicas y Matemáticas, Universidad de Concepción, Concepción, Chile
Northern Chilean Patagonia is a mega-estuarine system where oceanic waters mix with freshwater inputs in the coastal fjords, channels and gulfs. The aim of this study was to examine the distribution of blue and sei whales with respect to oceanographic conditions of the study area from the estuarine inner sea to the outer ocean. Ocean gliders were used, mounted with a hydrophone to determine acoustic presence of whales (Southeast Pacific and Antarctic blue whale song calls, and blue whales D-calls; sei whale downsweeps and upsweeps), and a temperature and salinity instrument. Four glider deployments were carried out in April 2018 and April-June 2019 navigating a total of 2817 kilometers during 2110 hours. To examine interannual variation, the average percentage of day with presence of calls was compared between years using the adjusted p-values for one-way ANOVA and descriptive statistics. To examine spatial variation between the hourly acoustic presence of blue whales and sei whales and temperature and salinity conditions, Generalized Linear Models (GLMs) were used. Salinities were higher in 2019 compared to 2018. Southeast Pacific blue whales produced song calls throughout the study area in both years, across estuarine and oceanic areas, but percentage of day with presence was higher in 2019 vs 2018. Percentage of day with presence of D-calls was similar between years, but higher in oceanic areas during both study periods. In contrast, the spatial pattern of sei whale acoustic presence was ambiguous and interannual variability was high, suggesting that sei whales preferred estuarine areas in 2018 and oceanic areas in 2019. We discuss possible explanations for observed acoustic presence in relation to foraging behavior and prey distribution.
Introduction
Understanding the distribution of baleen whales on their coastal feeding grounds and the oceanographic processes that drive distribution is central to developing effective conservation measures for the recovery of populations and for a better understanding of pelagic ecology. Off the coast of southern Chile baleen whales, predominantly blue (Balaenoptera musculus) and sei (B. borealis) whales, forage on a seasonal feeding ground in Northern Chilean Patagonia (NCP, Figure 1) (e.g. (Hucke-Gaete et al., 2004; Buchan and Quiñones, 2016; Vernazzani et al., 2017; Hucke-Gaete et al., 2018; Bedriñana-Romano et al., 2021; Buchan et al., 2021a). In this area, blue whales are known to forage on the euphausiid Euphausia vallentini (Buchan and Quiñones, 2016) while sei whales forage on lobster krill Munida gregaria (Buchan et al., 2021b).
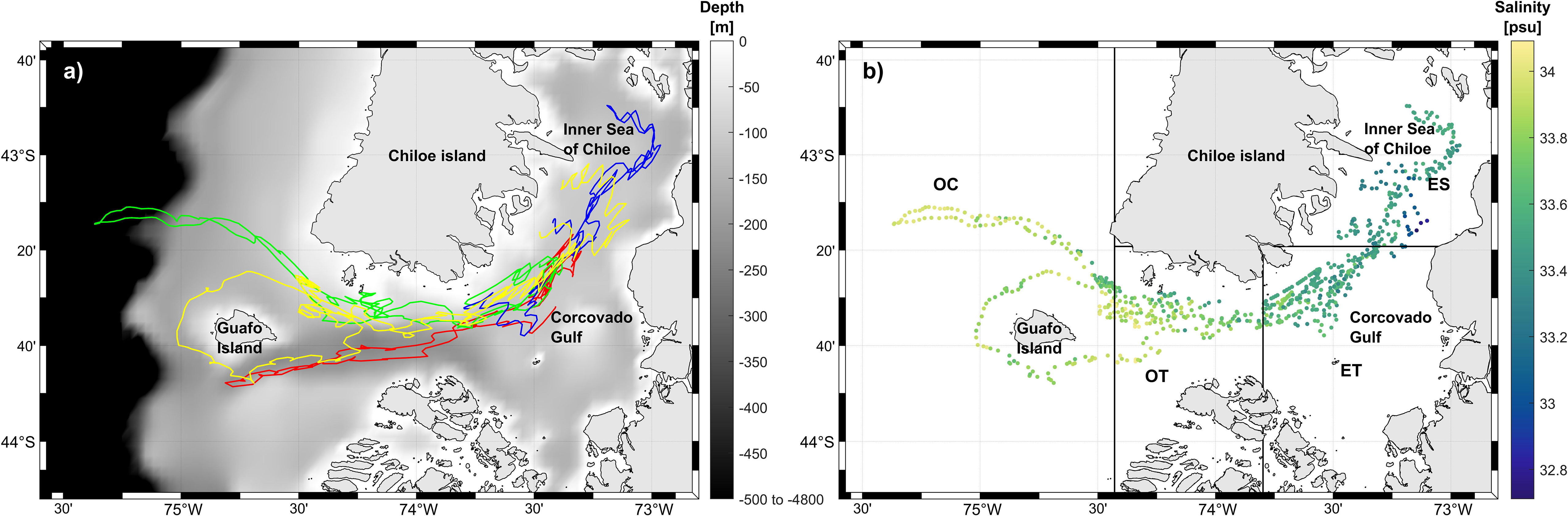
Figure 1 (A) Glider deployment tracks in 2018 and 2019 between the inner sea of Chiloe and Guafo Island. Red track: deployment 1, April 2018; green track: deployment 2, April 2019; blue track: deployment 3, April 2019; yellow track: deployment 4, May 2019. Grey scale: bathymetry from Instituto de Fomento Pesquero (IFOP; (Pinilla et al., 2020)), Chile with depths ranging from 0 to -4800 m (B) Map of deep layer salinity for 2019 deployments (d2, d3 and d4) throughout the study area. ES, Estuarine zone; ET, Estuarine Transition zone; OT, Oceanic Transition zone; OC, Oceanic zone.
The NCP is a mega-estuarine system where oceanic water masses from the Pacific Ocean enter through open gulfs and restricted channels and mix with freshwater inputs from high levels of rainfall running off the mainland into coastal fjords and channels. This mix of water masses contains nutrients necessary for the high observed levels of primary and secondary production (e.g. (Sievers and Silva, 2008; Iriarte et al., 2010; Buchan and Quiñones, 2016). High primary production has been particularly associated with the Modified Subantarctic Water Mass (MSAAW) (Silva and Guzman, 2006; Torres et al., 2014). Based on a study in the inner Corcovado Gulf over two summer seasons, (Buchan and Quiñones, 2016) found higher blue whale sightings in the summer where MSAAW was the predominant water mass over the higher salinity Subantarctic Water (SAAW). However, this study was restricted to a very small area near the mainland and whales are known to forage further out in oceanic areas as well, including the northwest coast of Chiloe Island (Vernazzani et al., 2012; Bedriñana-Romano et al., 2018; Bedriñana-Romano et al., 2021).
Blue whales produce loud song (sounds in repetitive patterned phrases) and non-song calls throughout their migratory range; therefore, acoustic detections can be used as an indicator of their presence (e.g. (Stafford et al., 1999; Stafford et al., 2009; Buchan et al., 2014; Romagosa et al., 2020). Silent animals are not detected by this method. The Southeast Pacific (SEP) blue whales present in NCP can be monitored acoustically using their highly stereotyped regional song dialect, known as “Southeast Pacific 2” or “SEP2” (Buchan et al., 2014; Buchan et al., 2015). Antarctic blue whales are also known to be present in this area (Buchan et al., 2018) and are known to produce stereotyped “Z-call” songs (Širović et al., 2004). In addition to regional song calls, blue whales also produce non-regionally distinct highly variable downswept calls or “D-calls” (Thompson et al., 1996) (see Figure 2). In blue whales, long-range songs are produced by males and are thought to serve some reproductive function, while D-calls are produced by both males and females and are thought to be short-range social calls between animals that are in close proximity (Lewis et al., 2018). D-calls have been reported during a range of behaviors including feeding (Oleson et al., 2007; Lewis et al., 2018) and potential reproductive behaviors (Schall et al., 2019).

Figure 2 Spectrograms of (A) blue whale D-calls; (B) sei whale downsweep calls; (C) sei whale upsweep calls. Spectrogram parameters: FFT: 512 samples, 50% overlap, Hann Window.
Less is known of sei whale acoustic behavior. Sei whales can produce both downsweep (Baumgartner et al., 2008) and upsweep calls (Calderan et al., 2014) (Figure 2). The downswept calls are distinct from blue whale D-calls as they are less variable in bandwidth and are more stereotyped (Figure 2). The function and behavioral context of sei whale calls is unknown. As with blue whales, detections of these calls in passive acoustic datasets collected by hydrophones indicate the acoustic presence of each of this species.
Ocean gliders are autonomous vehicles now widely used to increase the spatial and temporal coverage of ocean observation (e.g. Webb et al., 2001; Testor et al., 2019). Gliders can be equipped with oceanographic sensors, such as conductivity-temperature-depth (CTD) sensors, but also hydrophones to collect passive acoustic data for the duration of their deployment e.g. (Klinck et al., 2012; Baumgartner et al., 2014; Baumgartner et al., 2020; Cauchy et al., 2020; Johnson et al., 2022). In this study, we examine the distribution of blue and sei whales with respect to oceanographic conditions from the estuarine pole (inner sea of Chiloe) to the oceanic pole (Guafo Island) of the NCP mega-estuarine system (Figure 1). We used ocean gliders mounted with a hydrophone to determine whale acoustic presence, and a CTD instrument to determine temperature and salinity along the glider route.
Materials and methods
Data collection
Two Slocum G1 gliders, one owned by the University of Concepción (gladis) and one owned by Woods Hole Oceanographic Institution (we04), surveyed between the inner sea of Chiloe and Guafo Island through the Corcovado Gulf to cover the range of estuarine to oceanic oceanographic conditions (Figure 1). One deployment was conducted in 2018 and three were conducted in 2019 of which two were simultaneous (Table 1). Both gliders were deployed with a digital acoustic monitoring instrument (DMON; (Johnson and Hurst, 2007) (Baumgartner et al., 2013) which is a hydrophone and recording device that recorded audio continuously at 2 kHz (for an effective bandwidth of 5-1000 Hz) to.wav files of 1 hour duration. The DMON has a total system sensitivity of -169.8 dB re V/µPa rms. The sample rate and system sensitivity was the same across all deployments. A CTD instrument sampling at 1 Hz was deployed on both gliders. A Neil Brown Ocean Sensors, Inc. CTD was deployed on the we04 glider and a Sea Bird Electronics model 41CP was deployed on the gladis glider.
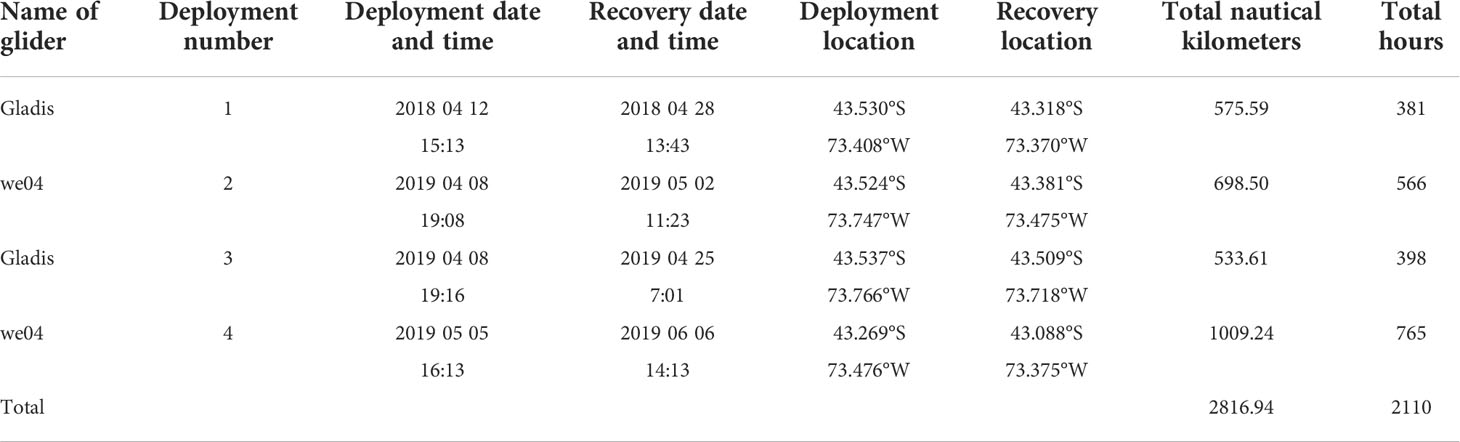
Table 1 Glider deployment metadata. Deployment date, time, location, total km and hours of survey effort.
Data analysis
Passive acoustic data from the DMON were analyzed via manual annotation in Raven Pro 1.6 (K. Lisa Yang Center for Conservation Bioacoustics at the Cornell Lab of Ornithology., 2022) for the following target signals: unit C of the four-unit “SEP2” song calls of Chilean blue whales (as per Buchan et al., 2021a), “Z” song calls of Antarctic blue whales (as per Buchan et al., 2018), blue whale D-calls (e.g. Buchan et al., 2021a; Figure 2), and upsweep and downsweep calls of sei whales (e.g. Calderan et al., 2014; Figure 2). The date and time of each manual annotation of a call was compiled as a time series, and from the time stamp of each annotation a latitude and longitude was allocated to each annotation from the glider track information based on interpolation between known surface positions using MATLAB, 2020b (The Math Works, 2020).
Temperature (°C) and salinity (psu) data from the CTD instrument sampling on the downward portion of dives were processed in MATLAB (The Math Works, 2020) using custom routines and compiled as time series. Each data point was allocated a latitude and longitude based on interpolation between known surface positions along the glider track. Temperature and salinity data were examined for surface and deep layers given the stratified nature of estuarine environments. From density profiles, the mixed layer depth (MLD) was determined for each profile by determining the first depth (starting from the surface) at which a gradient of 0.03 kg/m4 was found (Holte and Talley, 2009). All MLD values less than 2 m and greater than 50 m were eliminated as unrealistic. Based on the MLD values, surface (above MLD) and deep layer (below MLD) average temperature and salinity were calculated for each profile to describe the spatial variation of these variables; where no MLD was found (either because the water column was well-mixed or because the method used did not detect MLD), the average of MLD over the entire dataset was applied, i.e. 24.09 m for the purposes of obtaining surface and deep layer temperature and salinity values.
For a comparison of temperature and salinity conditions between years, data from April 2018 and only April 2019 were compared. All temperature and salinity data (April 2018 and April-June 2019) were used in the statistical analyses described below.
Statistical analyses
To examine spatial variation in the presence of whale calls and temperature and salinity conditions, the study area was divided into 4 large areas, which we refer to as oceanographic zones, based on visual inspection of the deep layer salinity values for the 2019 data (Figure 1B), which was the larger dataset with greater spatial coverage compared to 2018. This resulted in the following oceanographic zones which range from generally low to high salinity conditions (Figure 1B): Estuarine zone (ES); Estuarine Transition zone (ET); Oceanic Transition zone (OT); and Oceanic zone (OC). Temperature and salinity trends per oceanographic zone were overall similar between years (see Results) so we are confident that this division of zones is reasonably applied to the 2018 data.
Based on the whale call data, hourly presence/absence and percentage of day with presence were calculated. Hourly presence/absence of a call was determined where any hour with 1 or more calls was assigned a score of 1; and hours with no calls were assigned a score of 0. Percentage of day with presence of a call type was determined by the number of hours with 1+ calls/number of hours of recording per day. Percentage of day with presence of blue whale song calls (SEP2 and Antarctic), and D-calls were examined separately; for sei whales, upsweep and downsweep calls were pooled together.
To examine interannual variation, the average percentage of day with presence of calls was compared between years using the adjusted p-values for one-way ANOVA with post-hoc Tukey Honest Significant Differences test (Table 2). Boxplots of temperature and salinity values and percentage of day with calls present were used to examine temporal and spatial variation between years (2018 vs 2019), study periods (April 2018, deployment 1; April 2019, deployments 2 and 3; and May 2019, deployment 4), and oceanographic zones (ES, ET, OT, OC).

Table 2 Summary statistics for percentage of day with presence (Pd) (+/- s.d.) for blue whale song and D-calls, and sei whale calls (downsweeps and upsweeps) per year.
To examine the relationship between the hourly acoustic presence of blue whales and sei whales and oceanographic conditions, Generalized Linear Models (GLMs) with a binomial link function were fitted in R (R Core Team, 2019) with presence/absence (0 or 1) as the response variable and the following explanatory variables (one per GLM): surface temperature, deep layer temperature, surface salinity and deep layer salinity. For blue whales, only D-calls were used, based on the findings by (Buchan et al., 2021a) and references therein, that suggest that the production of D-calls is related to environmental changes and/or may indicate whale aggregation, whereas SEP2 song calls follow seasonal cycles likely linked to reproductive behavior rather than environmental drivers. Statistical significance was corrected with the Bonferroni adjustment.
Results
Four glider deployments were completed between 12 – 28 April 2018, and 8 April – 7 June 2019. Deployment tracks are shown in Figure 1 and deployment details are summarized in Table 1. A total of 2,110 hours of passive acoustic monitoring (PAM) data were collected: 381 h in 2018 and 1,729 h in 2019 (Table 1). A total of 43,566 SEP2 song calls (unit C), 1,239 Antarctic blue whale Z calls, 5,656 blue whale D-calls, and 8,238 sei whale calls (6,500 upsweep calls, and 1,738 downsweep calls) were annotated. Supplementary Material Figure 1 shows maps of the spatial distribution of detections for the four calls types and the three study periods. A total of 993 CTD vertical profiles were collected.
Interannual variability
Between the 2018 and 2019 deployments, there was no significant difference in the percentage of day with presence of blue whales D-calls, but there was a significant difference between higher percentage of day with presence of SEP2 song calls in 2019 as compared to 2018. Percentage of day with presence of Antarctic blue whales was also significantly higher in 2019. Percentage of day with presence of sei whales was significantly higher (p < 0.001) in 2018 compared with 2019 (Table 2).
Salinity conditions displayed differences between years, with 2018 being a lower salinity year across all oceanographic zones compared with 2019 (Figure 3). Based on the water mass definitions for this area, there was greater influence of Modified Subantarctic Water (MSAAW) during 2018 compared with 2019 in surface layers (Figures 3A, B), and greater influence of Equatorial Subantarctic Water (ESSW), especially in the oceanic (OT and OC) zones, in 2019 compared with 2018 (Figures 3E, F). No outstanding difference between temperature conditions were apparent between 2018 and 2019 (Figures 3C, D, G, H).
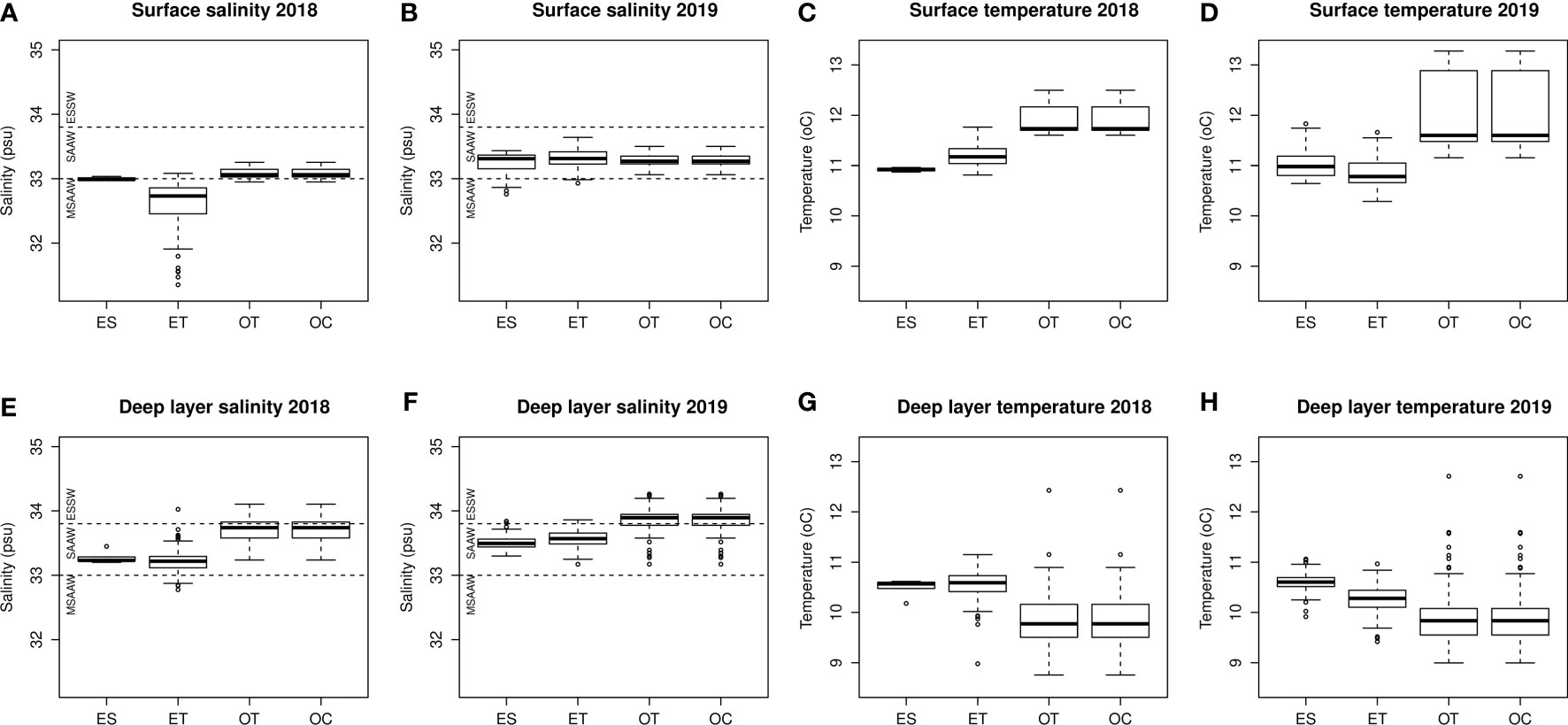
Figure 3 Boxplots of surface salinity (A, B), surface temperature (C, D), deep layer salinity (E, F), and deep layer temperature (G, H) for April 2018 and April 2019, and per oceanographic zone. ES, Estuarine zone; ET, Estuarine Transition zone; OT, Oceanic Transition zone; OC, Oceanic zone. Dashed lines show salinity-based water mass definitions according to (Pérez-Santos et al., 2014) for this area. ESSW, Equatorial Subsurface Water; SAAW, Subantarctic Water; MSAAW, Modified Subantarctic Water.
Spatial variability
The percentage of day with presence of blue whale SEP2 song calls was high over all study periods and zones, except for the ES in 2018 (Figure 4, first row). Antarctic blue whale songs were only present in 2019, with highest percentage of day with presence in estuarine zones (ES and ET) especially in May 2019 (Figure 4, second row). For blue whale D-calls, the percentage of day with presence was progressively higher from the estuarine zone (ES) to the oceanic zone (OC) during all study periods (Figure 4, third row). For sei whales the percentage of day with presence was higher in estuarine zone (ES) in April 2018. In contrast, in April 2019 percentage of day with presence was very low throughout the study area, with slightly higher values in the oceanic transition zone (OT). In May 2019, percentages of day with call presence were variable although slightly higher in the oceanic zone (OC) (Figure 4, fourth row).
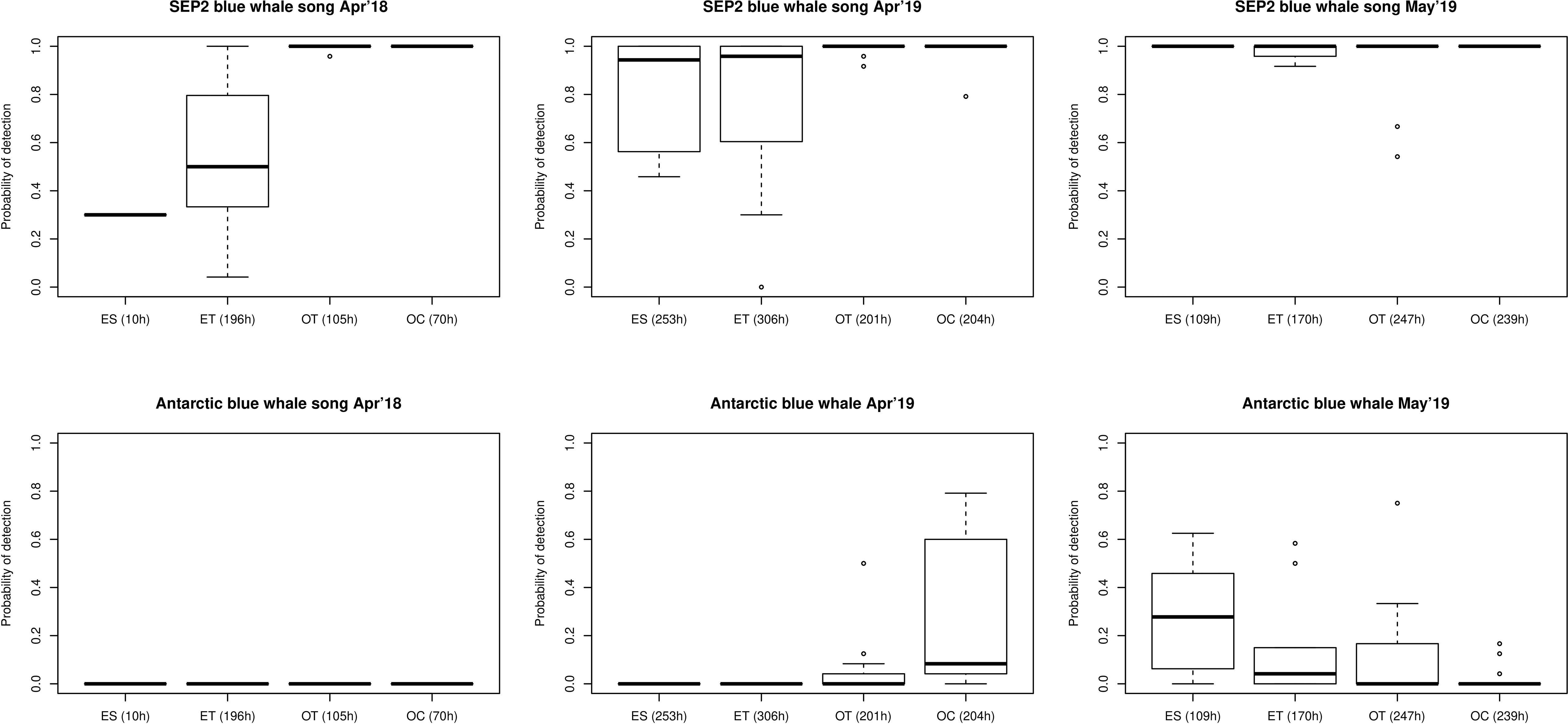
Figure 4 Boxplots of percentage of day with presence of SEP2 and Antarctic blue whale songs, blue whales D-calls and sei whale calls, per study period and per oceanographic zone. First column: April 2018 (deployment 1); Second column: April 2019 (deployment 2 and 3); Third column: May 2019 (deployment 4). ES, Estuarine zone; ET, Estuarine Transition zone; OT, Oceanic Transition zone; OC, Oceanic zone. For each zone, the number of hours that the glider was in the zone in indicated in the x-axis labels.
Progressively, from the ES to the OC zones, deep layer salinities increased in 2018 and 2019 and salinities were higher in the deep layer compared to the surface layer in all sectors; surface layer salinities only increased very slightly from ES to OC in 2018 but appeared the same in all oceanographic zones in 2019 (Figures 3A, B, E, F). Overall, surface and deep layer temperatures were not coupled: surface temperatures increased between the ES and the OC, and deep layer temperatures decreased between the ES and the OC (Figures 3C, D, G, H). Spatial differences between the estuarine pole (ES and ET) and the oceanic pole (OT and OC) were more marked in 2018 compared with 2019 in both the surface and deep layers.
Generalized Linear Models
Generalized Linear Models for blue whales D-calls performed better compared with sei whale calls, with higher percentages of deviance explained (maximum 26.1%), lower AIC values and statistically significant adjusted p-values. All GLMs for sei whale presence had percentages of deviance explained below 10% (Supplementary Image 1).
For blue whales, GLMs indicated that D-call presence was significantly associated with higher salinities in both the surface and deep layers, except for surface layer salinity during deployments 2 and 3, (Figure 5). GLM performance was better for deep layer salinity, which showed an increase in presence with increasing salinity levels (models 1 and 3, Table 3). During the study period, call presence increased significantly with decreasing temperature in the deep layer, but results were more ambiguous for surface temperature with the only significant relationship between call presence and increasing surface temperature found during April 2018, but not April or May 2019. Overall, GLMs for deep layer conditions performed generally better than surface layer conditions.
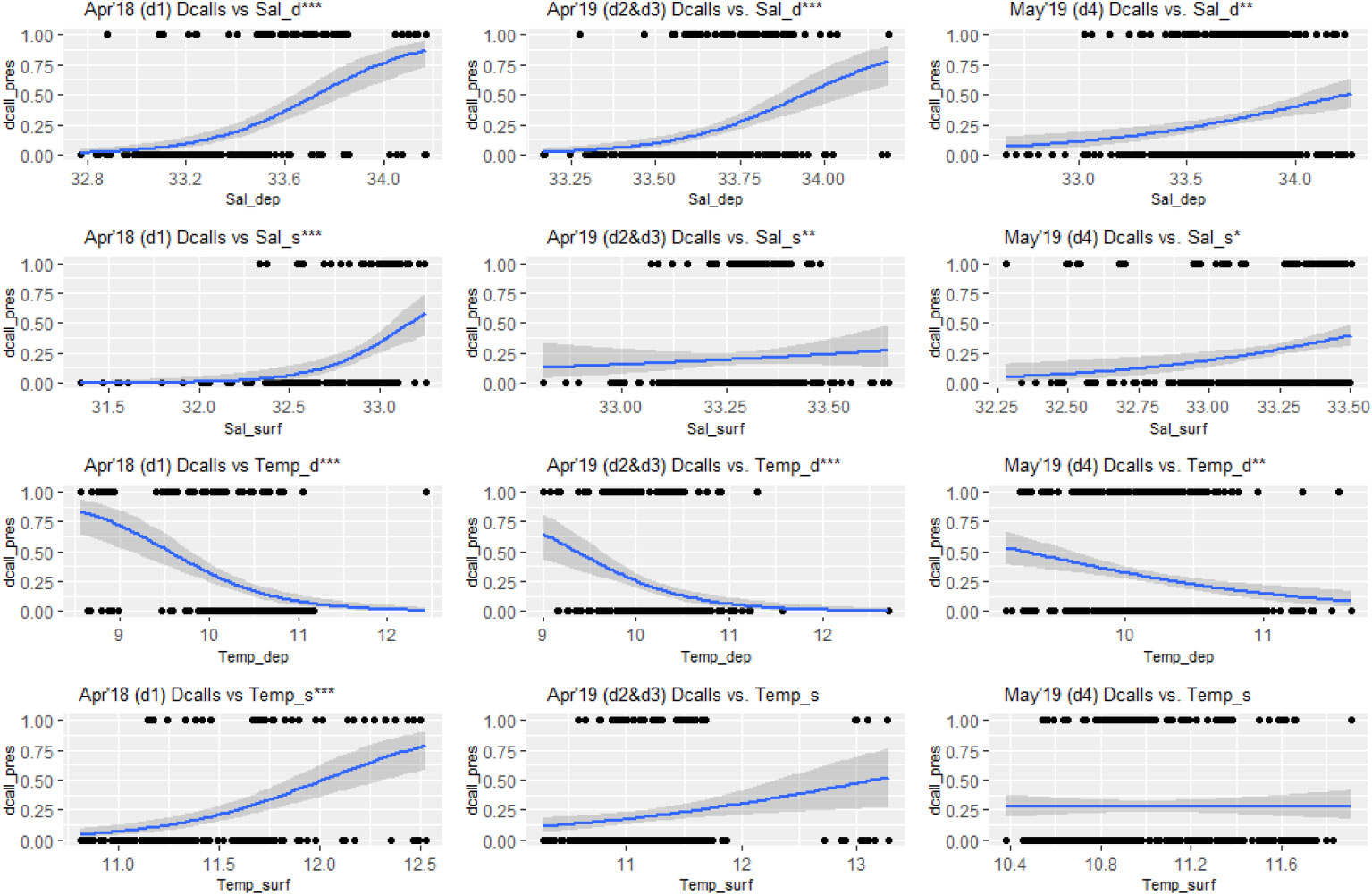
Figure 5 Generalized Linear Model response curves for blue whale D-call presence and deep layer salinity (SalD), surface layer salinity (SalS), deep layer temperature (TempD) and surface layer temperature (TempS). Salinity in psu and temperature in °C. Study periods: April 2019 (deployment 1), April 2019 (deployments 2 and 3), May 2019 (deployment 4). Bonferroni adjusted significance codes: *0.00416 > p > 0.00083; **0.00083 > p > 0.000083; *** p < 0.000083.
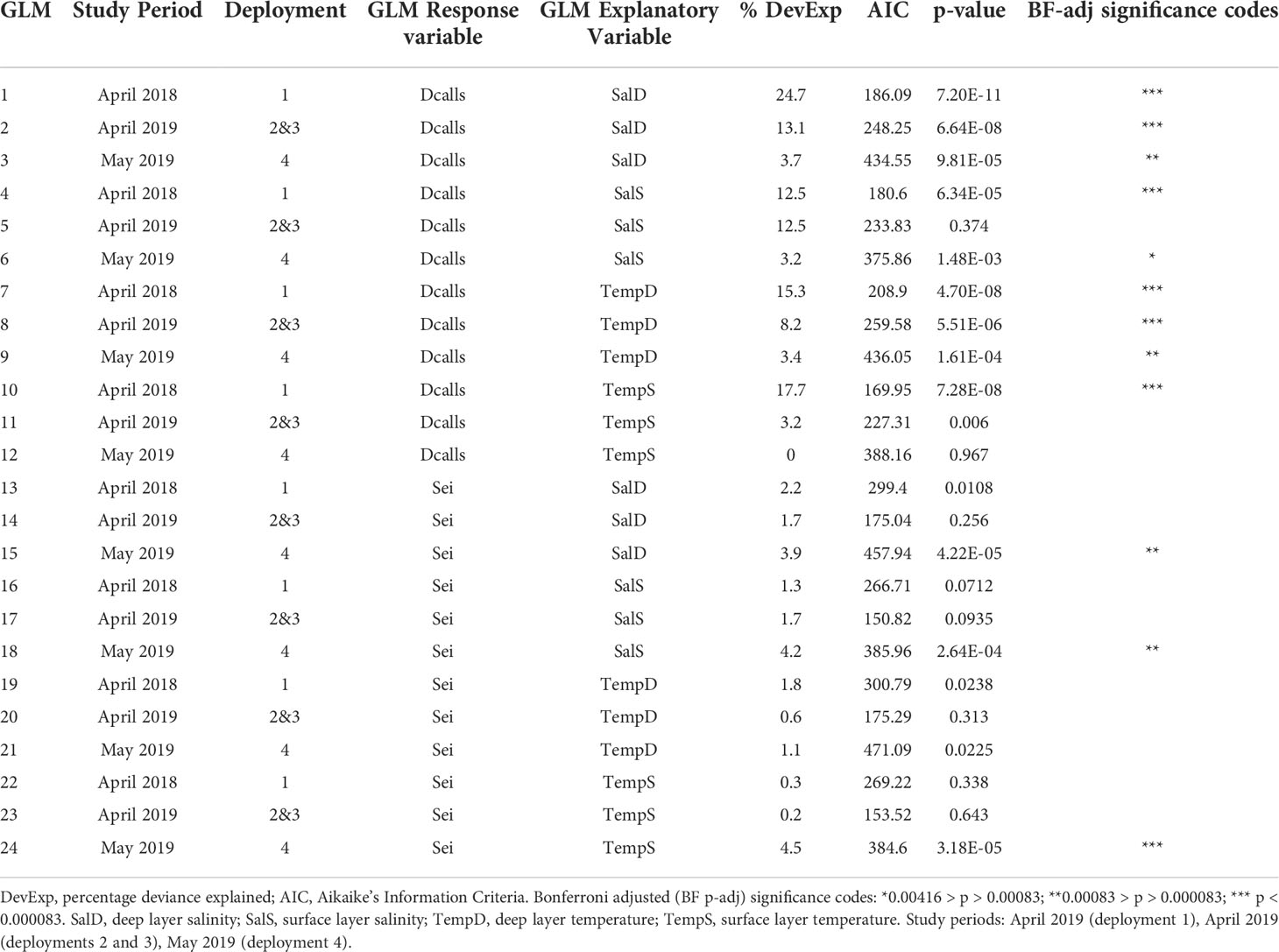
Table 3 Generalized Linear Models for percentage of day with presence of blue whale D-calls or sei whale calls (downsweeps and upsweeps) with temperature and salinity as explanatory variables, sorted by species and ranked by highest to lowest percentage deviance explained.
GLMs performed poorly for sei whales (Table 3) and statistically significant relationships only occurred for three GLMs (Table 3; Figure 6) during the May 2019 study period, revealing increased call presence with increasing surface and deep layer salinity, and increasing surface temperature. These conditions are characteristic of oceanic areas. This is interesting given that during the low-salinity year 2018, presence of sei whale calls was overall higher in the study area compared with 2019, but none of the GLMs for the 2018 study period showed statistically significant relationships.
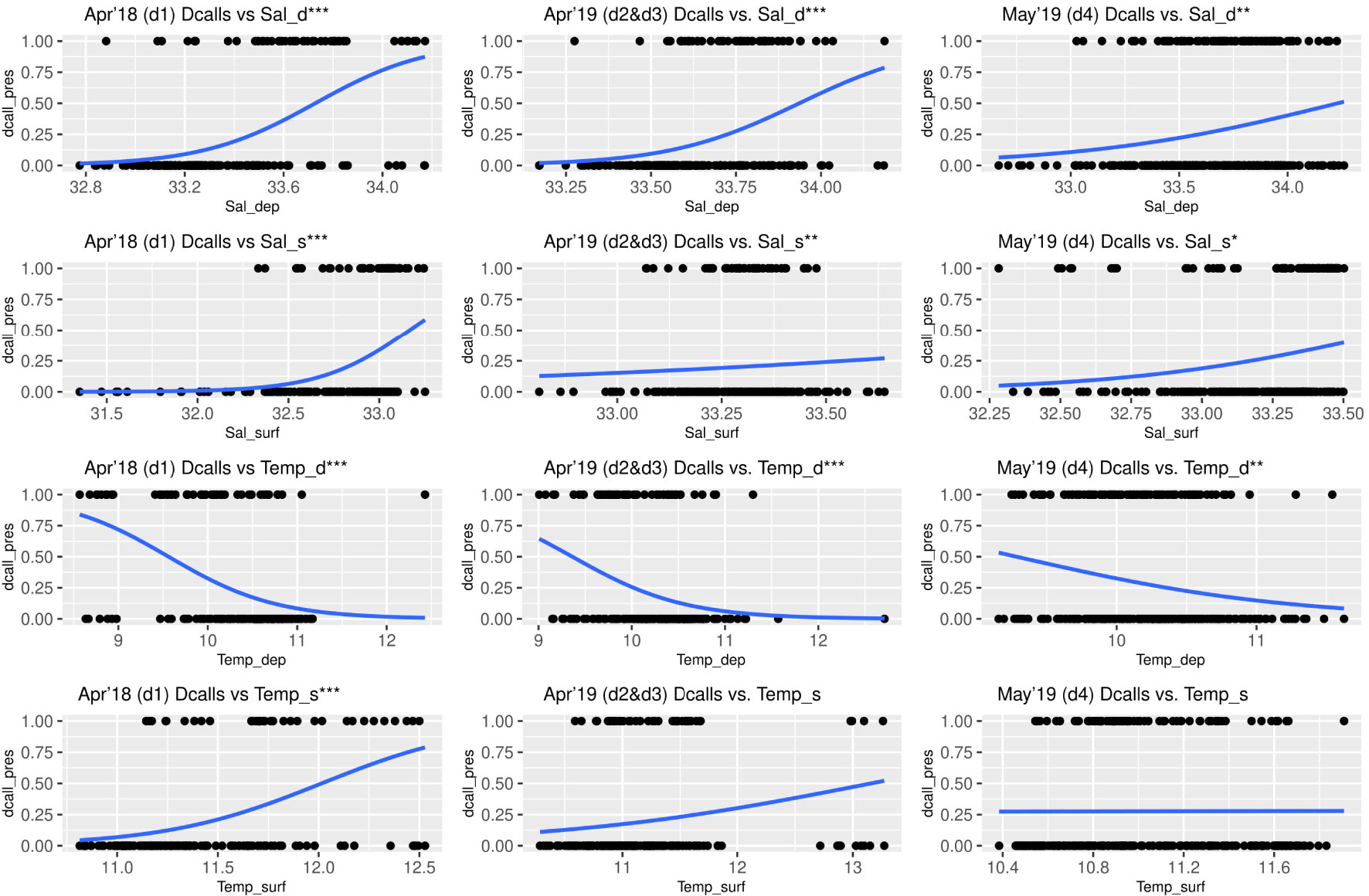
Figure 6 Generalized Linear Model response curves for probability of sei whale detection (downsweeps and upsweeps) and deep layer salinity (SalD), surface layer salinity (SalS), deep layer temperature (TempD) and surface layer temperature (TempS). Salinity in psu and temperature in °C. Study periods: April 2019 (deployment 1), April 2019 (deployments 2 and 3), May 2019 (deployment 4). Bonferroni adjusted significance codes: *0.00416 > p > 0.00083; **0.00083 > p > 0.000083; *** p < 0.000083.
Discussion
Blue whales were detected throughout the study area, with a preference for oceanic areas and little interannual variation, based on D-calls detections. Sei whale detections were highly variable from one year to another, with an ambiguous spatial pattern. Below we discuss possible causes of interannual and spatial variability for both species, based on deployments during approximately two weeks in April 2018 and approximately two months April-June 2019.
Interannual variation
No interannual variation was detected for blue whale D-call detections, but the percentage of day with presence of Antarctic and SEP2 blue whale song calls was higher in 2019. This suggests that the interannual variation in the presence of Antarctic blue whales is high, and in effect this species has only been previously found to occasionally visit the NCP feeding ground (Buchan et al., 2018). For Chilean blue whales, the explanation for this is less evident. Possibly, during the study periods, blue whales exploited the area for feeding with similar intensity (assuming D-calls are indicative of whale aggregation around feeding patches), but song might have varied due to interannual changes in monthly song production as reported by (Oestreich et al., 2020). This may also be due to a much shorter study period in 2018 that introduced bias in percentage of day with presence of song but not for D-calls.
Percentage of day with presence of sei whale calls showed interannual variation. Low salinities in 2018 in the study area may explain significantly higher sei whale acoustic presence in 2018 compared to 2019. From sei whale fecal plume sampling conducted at the same time as the glider surveys in this study, sei whales were found to feed on Munida gregaria (Buchan et al., 2021a), also known as squat lobster or lobster krill. This crustacean has been linked to freshwater inputs in this study area (Meerhoff et al., 2013) and has only be studied in the estuarine areas, i.e. in the NCP inner sea and fjords (Castro et al., 2011; Meerhoff et al., 2014a; Meerhoff et al., 2014b; Betti et al., 2020). A low-salinity year may favor increased abundance and spatial distribution of lobster krill in the study area and therefore attract foraging sei whales. This is not the case for the known prey item of blue whales in this area, E. vallentini (Buchan and Quiñones, 2016), which is the dominant species of meso-zooplankton throughout the study area (González et al., 2011) which may explain why no interannual variation was found in blue whale percentage of day with presence.
Spatial variation
SEP2 and Antarctic blue whale song calls
Percentage of day with presence was high for blue whale SEP2 song calls throughout all study periods and oceanographic zones (except for ES during April 2018). D-calls were more strongly associated with oceanic areas. The prevalence of SEP2 song calls vs. D-calls may be due to the large detection range of SEP2 song calls. In effect, (Buchan et al., 2021a) found that the maximum detection range of SEP2 song calls (up to 15.3 km) from a bottom-mounted hydrophone in this area was over twice the maximum detection range of D-calls.
Detection ranges on the glider have not been assessed in this study for any call types, and will be the focus of future work. Detection ranges are likely to change as the glider moves through the water column, i.e. changes in receiver depth; and through the study area, i.e. changes in temperature and salinity conditions which lead to changes in sound speed profile. Sound speed profiles for each oceanographic zone can be found in Supplementary Material Figure 2 and show that profiles are more stratified in oceanic zones compared with estuarine zones, which could lead to difference in detection ranges between areas. Changes in ambient noise between years and areas could also modify detection range (e.g. Fregosi et al., 2020). However, if for arguments sake, we were to assume that detection ranges for song calls and D-calls were similar to the bottom-mounted hydrophone in (Buchan et al., 2021a), these remain much smaller than the total distance covered by the glider in the study area, i.e. 150 km between the estuarine pole and oceanographic pole, with Chiloe Island largely intervening (Figure 1). Thus, although detection ranges must be assessed for the glider, we have confidence that the spatial variations in call presence observed in this study are due to differences in whale acoustic presence and not simply due to differences in detection range.
In contrast to SEP2 song calls, the percentage of day with presence of Antarctic blue whale songs was low throughout the study area and periods. This is not surprising given that NCP is not their primary feeding ground and Antarctic blue whales have only been heard occasionally in this area (Buchan et al., 2018). Interestingly however, Antarctic blue whales were detected in both oceanic and estuarine areas. Although the detection range of Antarctic blue whale song calls in this area was not assessed, these findings suggest that Antarctic blue whales that visit NCP, possibly to forage, do not exclusively remain in oceanic area but do enter the Patagonian inner sea.
Blue whale D-calls
D-calls are thought to be short-range social calls produced by males and females when individuals are in close proximity (Lewis et al., 2018) and have been reported during a range of behaviors including feeding (Oleson et al., 2007; Lewis et al., 2018) and possible reproductive behaviors (Schall et al., 2019). D-calls appear to respond to short-term (days to weeks) environmental changes much more than song calls (Szesciorka et al., 2020; Buchan et al., 2021a). Based on the results of this study, although blue whales were producing song calls throughout the study area, D-calls were more abundant in oceanic areas.
Presence of blue whale D-calls was consistently higher in oceanic areas across all study periods, with statistically significant relationships found with increasing surface and deep layer salinity, and decreased deep layer temperature. The apparent preference of blue whales for higher salinity, lower deep layer temperature oceanic areas may be because their only prey, Euphausia vallentini (Buchan and Quiñones, 2016), is a subantarctic oceanic species that has colonized the NCP inner sea system (Hamame and Antezana, 2010). This preference does not mean that blue whales do not enter the inner sea, as SEP2 song calls were detected in estuarine areas, but it may mean that if D-calls are associated with foraging activity, oceanic areas may be preferred foraging areas. This spatial pattern is no doubt overlaid by temporal variation at submonthly scales possibly linked to patch depletion and/or aggregation effects, driven by tidal amplitude or wind stress, as suggested by findings by (Buchan et al., 2021a). Conclusions regarding spatial distribution and preference for oceanic areas should be drawn with caution given the short time periods in this study; more extensive temporal and spatial coverage is certainly needed to confirm this pattern.
Sei whale calls
For sei whales, the overall NCP spatial pattern is harder to understand. If we assume that lobster krill prefers low-salinity areas, which is supported by results in (Meerhoff et al., 2013) but has not been studied over the entire study area including oceanic zones, and sei whales were feeding exclusively on lobster krill during the entire study period, we would have expected sei whales to prefer estuarine areas. However, sei whales did not only prefer estuarine areas. In April 2018, when the probability of sei whale detection was highest, sei whales preferred estuarine zones (Figure 4), however the GLM analysis revealed no statistically significant relationship between sei whale presence and temperature or salinity variables (Figure 6). Sei whale acoustic presence was very low overall during April 2019, but in May 2019 sei whales appeared to prefer oceanic areas with higher deep and surface layer salinities and higher surface temperatures, similar to blue whales during the same time period.
The spatial pattern in sei whale acoustic presence may be confounded because sei whales are known to feed on a range of prey types, including copepods, euphausiids, lobster krill and small schooling fish (Matthews, 1932; Kawamura, 1974) (Burkhardt-Holm and N’Guyen, 2019) (Buchan et al., 2021b). Although sei whales were definitely feeding on lobster krill in the inner sea area (ES and ET zones) at the time of the 2018 glider survey (Buchan et al., 2021b), no fecal samples were collected from the other study periods or oceanographic regions, so it is possible that sei whales were feeding on other prey type(s), e.g. euphausiids, similar to the blue whales foraging in oceanic areas during May 2019. It is also possible that lobster krill aggregations may have extended out to the OT and OC zones in May 2019. Lobster krill has only been studied in inner sea estuarine areas so little is known about its distribution in oceanic areas and its tolerance for higher salinity conditions.
In polar latitudes (>60°) sei whales have been associated with warmer waters (Kapel, 1985; Heide-Jorgensen and Simon, 2007), but a more recent study in the Fram Strait found no relationship between sei whale acoustic presence and water temperature time series (Nieukirk et al., 2020). Given that the site in this study is in mid-latitudes, the association with warmer water may not necessarily apply here and/or this association may apply over a broader spatial scale.
Sei whales have been called the “forgotten whale” and there is little information about their diet and distribution, the latter of which appears to be highly variable between the two years. There have been documented “sei whale invasions” or “sei whale years” and other years where sei whales are largely absent (Prieto et al., 2012). Clearly, more information is needed on the dietary plasticity of sei whales and the distribution of lobster krill in NCP to better understand the drivers of sei whale distribution in this area.
Conclusion
Oceanographic conditions were different between 2018 and 2019 in the NCP area making broad conclusions about the physical drivers of baleen whales in the area difficult to encapsulate. Salinity conditions were highly variable between the two years in the NCP estuarine system, with 2018 being a lower-salinity year and 2019 being a higher-salinity year.
Blue whales were producing song calls throughout the study area in both years, across estuarine and oceanic areas, and overall presence was higher in 2019 compared with 2018. Presence of D-calls was similar between years, but higher in oceanic areas during both study periods, possibly indicating aggregation of animals and/or foraging behavior in oceanic areas. Higher D-call presence during all study periods in higher-salinity oceanic areas may have been driven by higher prey density in these areas, given that E. vallentini is an oceanic species. In contrast, the spatial pattern of sei whale acoustic presence was ambiguous and interannual variability was high, suggesting that sei whales preferred estuarine areas in 2018 and oceanic areas in 2019. In 2018, sei whales were certainly feeding on lobster krill, M. gregaria, which is known to be associated with freshwater input in NCP, explaining the high acoustic presence in estuarine areas. In 2019, we have no dietary information for sei whales, so we do not know if they were feeding on lobster krill aggregations that may have extended further offshore or if they switched prey type to feed on euphausiids in oceanic areas. Future studies in this area should focus on (1) acquiring glider-based zooplankton backscatter data during glider-based acoustic surveys, (2) increasing the spatial and temporal coverage of glider surveys, (3) understanding lobster krill and E. vallentini distributions, and other zooplankton distribution between the estuarine and oceanic poles of NCP, and (4) improving knowledge of sei whale diet plasticity.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
SB contributed to study conception, design, data collection and analysis, manuscript preparation, including writing the first draft of this manuscript, and securing funding. LG contributed to data collection and analysis, and manuscript preparation. MB and KS contributed to study conception, design, data collection and analysis, manuscript preparation, and securing funding. NR contributed to data collection and analysis, and manuscript preparation. OP contributed to study design, data collection, and securing funding. JC contributed to data analysis. All authors commented on all previous versions of the manuscript and read and approved the final manuscript.
Funding
Financial support was provided by the Office of Naval Research grant N00014-17-1-2606. Partial funding was provided by Centro COPAS Sur-Austral PFB31 and ANID APOYO CCTE AFB170006, and Centro COPAS Coastal 210021, funded by the Chilean Agencia Nacional de Investigación y Desarrollo (ANID) (https://www.anid.cl/). All deployments were carried out under SHOA resolutions 13270_24_136 and 13270_24_155. A MATLAB, 2020b Academic License to the University of Concepción was used in this research. A free student license of Raven Pro 1.6 was kindly provided to LG by the K. Lisa Yang Center for Conservation Bioacoustics.
Acknowledgments
Our sincere thanks to the captain and crew of the Don Felipe, and Matias Pizarro for their valuable support during the fieldwork. Our thanks to the Instituto de Fomento Pesquero for bathymetric data shown in Figure 1.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.903964/full#supplementary-material
Supplementary Figure 1 | Maps of spatial distribution of detections of all four call types (Southeast Pacific blue whales Sep2 song, blue whale D-calls, sei whale upcalls and downsweep calls, and Antarctic blue whale song) per study period. Study periods: April 2019 (d1, deployment 1), April 2019 (d2 & d3, deployments 2 and 3), May 2019 (d4 = deployment 4). ES, Estuarine zone; ET, Estuarine Transition zone; OT, Oceanic Transition zone; OC, Oceanic zone.
Supplementary Figure 2 | Sound speed profiles per year (2018: top row; 2019: bottom row) per oceanographic zone, ES=Estuarine zone; ET= Estuarine Transition zone; OT=Oceanic Transition zone; OC=Oceanic zone. Black line indicates average and green line indicates the standard deviation.
References
Baumgartner M. F., Bonnell J., Corkeron P. J., Van Parijs S. M., Hotchkin C., Hodges B. A., et al. (2020). Slocum Gliders provide accurate near real-time estimates of baleen whale presence from human-reviewed passive acoustic detection information. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00100
Baumgartner M. F., Fratantoni D. M., Hurst T. P., Brown M. W., Cole T. V. N., Van Parijs S. M., et al. (2013). Real-time reporting of baleen whale passive acoustic detections from ocean gliders. J. Acoust. Soc Am. 134, 1814–1823. doi: 10.1121/1.4816406
Baumgartner M. F., Stafford K. M., Winsor P., Statscewich H., Fratantoni D. M. (2014). Glider-based passive acoustic monitoring in the Arctic. Mar. Technol. Soc J. 48, 40–51. doi: 10.4031/MTSJ.48.5.2
Baumgartner M. F., Van Parijs S. M., Wenzel F. W., Tremblay C. J., Carter Esch H., Warde A. M. (2008). Low frequency vocalizations attributed to sei whales ( balaenoptera borealis ). J. Acoust. Soc Am. 124, 1339–1349. doi: 10.1121/1.2945155
Bedriñana-Romano L., Hucke-Gaete R., Viddi F. A., Johnson D., Zerbini A. N., Morales J., et al. (2021). Defining priority areas for blue whale conservation and investigating overlap with vessel traffic in Chilean Patagonia, using a fast-fitting movement model. Sci. Rep. 11, 1–16. doi: 10.1038/s41598-021-82220-5
Bedriñana-Romano L., Hucke-Gaete R., Viddi F. A., Morales J., Williams R., Ashe E., et al. (2018). Integrating multiple data sources for assessing blue whale abundance and distribution in Chilean northern Patagonia. Divers. Distrib. 24, 991–1004. doi: 10.1111/ddi.12739
Betti F., Castro L. R., Bavestrello G., Enrichetti F., Daneri G. (2020). Distribution, abundance and ecological requirements of the benthic phase of munida gregaria (Anomura; munididae) in the puyuhuapi fjord (Chilean Patagonia). Reg. Stud. Mar. Sci. 40, 101534. doi: 10.1016/j.rsma.2020.101534
Buchan S. J., Hucke-Gaete R., Rendell L., Stafford K. (2014). A new song recorded from blue whales in the corcovado gulf, southern Chile, and an acoustic link to the Eastern tropical pacific. Endanger. Species Res. 23, 241–252. doi: 10.3354/esr00566
Buchan S. J., Hucke-Gaete R., Stafford K. M., Clark C. W. (2018). Occasional acoustic presence of Antarctic blue whales on a feeding ground in southern Chile. Mar. Mammal Sci. 34, 220–228. doi: 10.1111/mms.12441
Buchan S. J., Pérez-Santos I., Narváez D., Castro L., Stafford K. M., Baumgartner M. F., et al. (2021a). Intraseasonal variation in southeast pacific blue whale acoustic presence, zooplankton backscatter, and oceanographic variables on a feeding ground in northern Chilean Patagonia. Prog. Oceanogr. 199, 1–14. doi: 10.1016/j.pocean.2021.102709
Buchan S. J., Quiñones R. A. (2016). First insights into the oceanographic characteristics of a blue whale feeding ground in northern Patagonia, Chile. Mar. Ecol. Prog. Ser. 554, 183–199. doi: 10.3354/meps11762
Buchan S. J., Stafford K. M., Hucke-Gaete R. (2015). Seasonal occurrence of southeast pacific blue whale songs in southern chile and the eastern tropical pacific. Mar. Mammal Sci. 31, 440–458. doi: 10.1111/mms.12173
Buchan S. J., Vásquez P., Olavarría C., Castro L. R. (2021b). Prey items of baleen whale species off the coast of Chile from fecal plume analysis. Mar. Mammal Sci. 37, 1116–1127. doi: 10.1111/mms.12782
Burkhardt-Holm P., N’Guyen A. (2019). Ingestion of microplastics by fish and other prey organisms of cetaceans, exemplified for two large baleen whale species. Mar. pollut. Bull. 144, 224–234. doi: 10.1016/j.marpolbul.2019.04.068
Calderan S., Miller B., Collins K., Ensor P., Double M., Leaper R., et al. (2014). Low-frequency vocalizations of sei whales ( balaenoptera borealis ) in the southern ocean. J. Acoust. Soc Am. 136, EL418–EL423. doi: 10.1121/1.4902422
Castro L. R., Cáceres M. A., Silva N., Muñoz M. I., León R., Landaeta M. F., et al. (2011). Short-term variations in mesozooplankton, ichthyoplankton, and nutrients associated with semi-diurnal tides in a patagonian gulf. Cont. Shelf Res. 31, 282–292. doi: 10.1016/j.csr.2010.09.005
Cauchy P., Heywood K. J., Risch D., Merchant N. D., Queste B. Y., Testor P. (2020). Sperm whale presence observed using passive acoustic monitoring from gliders of opportunity. Endanger. Species Res. 42, 133–149. doi: 10.3354/ESR01044
Fregosi S., Harris D. V., Matsumoto H., Mellinger D. K., Negretti C., Martin S. W., et al. (2020). Comparison of fin whale 20 Hz call detections by deep-water mobile autonomous and stationary recorders. J. Acoust. Soc Am. 147, 961–977. doi: 10.1121/10.0000617
González H. E., Castro L., Daneri G., Iriarte J. L., Silva N., Vargas C. A., et al. (2011). Seasonal plankton variability in Chilean Patagonia fjords: Carbon flow through the pelagic food web of aysen fjord and plankton dynamics in the moraleda channel basin. Cont. Shelf Res. 31, 225–243. doi: 10.1016/j.csr.2010.08.010
Hamame M., Antezana T. (2010). Vertical diel migration and feeding of euphausia vallentini within southern Chilean fjords. Deep. Res. Part II Top. Stud. Oceanogr. 57, 642–651. doi: 10.1016/j.dsr2.2009.10.013
Heide-Jorgensen M. P., Simon M. J. (2007). Estimates of large whale abundance in greenlandic waters from a ship-based survey in 2005. J. Cetacean Res. Manage. 9, 95–104.
Holte J., Talley L. (2009). A new algorithm for finding mixed layer depths with applications to argo data and subantarctic mode water formation. J. Atmos. Ocean. Technol. 26, 1920–1939. doi: 10.1175/2009JTECHO543.1
Hucke-Gaete R., Bedriñana-Romano L., Viddi F. A., Ruiz J. E., Torres-Florez J. P., Zerbini A. N. (2018). From Chilean Patagonia to Galapagos, Ecuador: Novel insights on blue whale migratory pathways along the Eastern south pacific. PeerJ 2018, 1–22. doi: 10.7717/peerj.4695
Hucke-Gaete R., Osman L. P., Moreno C. A., Findlay K. P., Ljungblad D. K. (2004). Discovery of a blue whale feeding and nursing ground in southern Chile. Proc. R. Soc B Biol. Sci. 271, S170–S173. doi: 10.1098/rsbl.2003.0132
Iriarte J. L., Gonzalez H. E., Nahuelhual L. (2010). Patagonian Fjord ecosystems in southern Chile as a highly vulnerable region: Problems and needs. Ambio 39, 463–466. doi: 10.1007/s13280-010-0049-9
Johnson M., Hurst T. (2007). “The DMON: An open-hardware/open software passive acoustic detector,” in 3rd International Workshop on the Detection and Classification of Marine Mammals using Passive Acoustics(Boston).
Johnson H. D., Taggart C. T., Newhall A. E., Lin Y.-T., Baumgartner M. F. (2022). Acoustic detection range of right whale upcalls identified in near-real time from a moored buoy and a Slocum glider. J. Acoust. Soc Am. 151, 2558–2575. doi: 10.1121/10.0010124
Kapel F. O. (1985). On the occurence of sei whales (Balaenoptera borealis) in the southern hemisphere (the soviet marking programme). Rep. Int. Whal. Commun. 35, 349–352.
Kawamura A. (1974). Food and feeding ecology in the southern sei whale*. Sci. Rep. Whales Res. Inst 1974, 25–144.
Klinck H., Mellinger D. K., Klinck K., Bogue N. M., Luby J. C., Jump W. A., et al. (2012). Near-real-time acoustic monitoring of beaked whales and other cetaceans using a SeagliderTM. PLoS One 7, 1–8. doi: 10.1371/journal.pone.0036128
K. Lisa Yang Center for Conservation Bioacoustics at the Cornell Lab of Ornithology (2022). Raven pro: Interactive sound analysis software (Version 1.6.3). (Ithaca, USA).
Lewis L. A., Calambokidis J., Stimpert A. K., Fahlbusch J., Friedlaender A. S., McKenna M. F., et al. (2018). Context-dependent variability in blue whale acoustic behaviour. R. Soc Open Sci. 5, 1–20. doi: 10.1098/rsos.180241
Matthews L. H. (1932). Lobster-krill. anomuran Crustacea that are the food of whales. Discovery Rep. 5, 467–484. 1 fig., 1 pl.
Meerhoff E., Castro L., Tapia F. (2013). Influence of freshwater discharges and tides on the abundance and distribution of larval and juvenile munida gregaria in the baker river estuary, chilean patagonia. Cont. Shelf Res. 61–62, 1–11. doi: 10.1016/j.csr.2013.04.025
Meerhoff E., Tapia F. J., Castro L. R. (2014a). Spatial structure of the meroplankton community along a Patagonian fjord - the role of changing freshwater inputs. Prog. Oceanogr. 129, 125–135. doi: 10.1016/j.pocean.2014.05.015
Meerhoff E., Tapia F. J., Sobarzo M., Castro L. (2014b). Influence of estuarine and secondary circulation on crustacean larval fluxes: A case study from a Patagonian fjord. J. Plankton Res. 37, 168–182. doi: 10.1093/plankt/fbu106
Nieukirk S. L., Mellinger D. K., Dziak R. P., Matsumoto H., Klinck H. (2020). Multi-year occurrence of sei whale calls in north Atlantic polar waters. J. Acoust. Soc Am. 147, 1842–1850. doi: 10.1121/10.0000931
Oestreich W. K., Fahlbusch J. A., Cade D. E., Calambokidis J., Margolina T., Joseph J., et al. (2020). Animal-borne metrics enable acoustic detection of blue whale migration. Curr. Biol. 30, 4773–4779.e3. doi: 10.1016/j.cub.2020.08.105
Oleson E. M., Calambokidis J., Burgess W. C., McDonald M. A., LeDuc C. A., Hildebrand J. A. (2007). Behavioral context of call production by eastern north pacific blue whales. Mar. Ecol. Prog. Ser. 330, 269–284. doi: 10.3354/meps330269
Pinilla E., Soto P., Soto-Riquelme C., Venegas O., Salas P., Cortés J. (2020). Determinación de las escalas de intercambio de agua en fiordos y canales de la región de Los Lagos y región de aysén del general carlos ibáñez del campo. 65.
Prieto R., Janiger D., Silva M. A., Waring G. T., Goncalves J. M. (2012). The forgotten whale: a bibliometric analysis and literature review of the north Atlantic sei whale balaenoptera borealis. Mamm. Rev. 42, 235–272. doi: 10.1111/j.1365-2907.2011.00195.x
Pérez-Santos I., Garcés-Vargas J., Schneider W., Ross L., Parra S., Valle-Levinson A. (2014). Double-diffusive layering and mixing in Patagonian fjords. Progress in Oceanography 129, 35–49. doi: 10.1016/j.pocean.2014.03.012
R Core Team. (2019). R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. Available at: https://www.R-project.org/
Romagosa M., Baumgartner M., Cascão I., Lammers M. O., Marques T. A., Santos R. S., et al. (2020). Baleen whale acoustic presence and behaviour at a mid-Atlantic migratory habitat, the Azores archipelago. Sci. Rep. 10, 1–11. doi: 10.1038/s41598-020-61849-8
Schall E., Di Iorio L., Berchok C., Filún D., Bedriñana-Romano L., Buchan S. J., et al. (2019). Visual and passive acoustic observations of blue whale trios from two distinct populations. Mar. Mammal Sci 36, 1–10. doi: 10.1111/mms.12643
Sievers H. A., Silva N. (2008). 4 . 1 . Water masses and circulation in austral Chilean channels and fjords. Water, 53–58.
Silva N., Guzman D. (2006). Condiciones oceanograficas fisicas y quimicas, entre boca del guafo y fiordo aysen CIMAR 7. Cienc. y Tecnol. del Mar. 29, 25–44.
Širović A., Hildebrand J. A., Wiggins S. M., McDonald M. A., Moore S. E., Thiele D. (2004). Seasonality of blue and fin whale calls and the influence of sea ice in the Western Antarctic peninsula. Deep. Res. Part II Top. Stud. Oceanogr. 51, 2327–2344. doi: 10.1016/j.dsr2.2004.08.005
Stafford K. M., Citta J. J., Moore S. E., Daher M. A., George J. E. (2009). Environmental correlates of blue and fin whale call detections in the north pacific ocean from 1997 to 2002. Mar. Ecol. Prog. Ser. 395, 37–53. doi: 10.3354/meps08362
Stafford K. M., Nieukirk S. L., Fox C. G. (1999). An acoustic link between blue whales in the eastern tropical pacific and the northeast pacific. Mar. Mammal Sci. 15, 1258–1268. doi: 10.1111/j.1748-7692.1999.tb00889.x
Szesciorka A. R., Ballance L. T., Širović A., Rice A., Ohman M. D., Hildebrand J. A., et al. (2020). Timing is everything: Drivers of interannual variability in blue whale migration. Sci. Rep. 10, 1–9. doi: 10.1038/s41598-020-64855-y
Testor P., DeYoung B., Rudnick D. L., Glenn S., Hayes D., Lee C., et al. (2019). OceanGliders: A component of the integrated GOOS. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00422
The Math Works (2020). MATLAB Release 2020b, The MathWorks, Inc., Natick, Massachusetts, United States.
Thompson P. O., Findley L. T., Vidal O., Cummings W. C. (1996). Underwater sounds of blue whales, balaenoptera musculus, in the gulf of California, Mexico. Mar. Mammal Sci. 12, 288–293. doi: 10.1111/j.1748-7692.1996.tb00578.x
Torres R., Silva N., Reid B., Frangopulos M. (2014). Silicic acid enrichment of subantarctic surface water from continental inputs along the Patagonian archipelago interior sea (41-56°S). Prog. Oceanogr. 129, 50–61. doi: 10.1016/j.pocean.2014.09.008
Vernazzani B. G., Carlson C. A., Cabrera E., Brownell R. L. (2012). Chilean Blue whales off isla grande de chiloe 2004-2010: Distribution, site-fidelity and behaviour. J. Cetacean Res. Manage. 12, 353–360.
Vernazzani B. G., Jackson J. A., Cabrera E., Carlson C. A., Brownell R. L. (2017). Estimates of abundance and trend of Chilean blue whales off isla de chiloe, Chile. PLoS One 12, 1–16. doi: 10.1371/journal.pone.0168646
Keywords: pelagic ecology, biological oceanography, baleen whales, passive acoustic monitoring, Slocum glider
Citation: Buchan SJ, Gutiérrez L, Baumgartner MF, Stafford KM, Ramirez N, Pizarro O and Cifuentes J (2022) Distribution of blue and sei whale vocalizations, and temperature - salinity characteristics from glider surveys in the Northern Chilean Patagonia mega-estuarine system. Front. Mar. Sci. 9:903964. doi: 10.3389/fmars.2022.903964
Received: 24 March 2022; Accepted: 24 August 2022;
Published: 07 October 2022.
Edited by:
Mónica A. Silva, University of the Azores, PortugalReviewed by:
Sofie Van Parijs, Northeast Fisheries Science Center, (NOAA), United StatesSelene Fregosi, Pacific Islands Fisheries Science Center, (NOAA), United States
Copyright © 2022 Buchan, Gutiérrez, Baumgartner, Stafford, Ramirez, Pizarro and Cifuentes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Susannah J. Buchan, sbuchan@udec.cl