
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 01 June 2022
Sec. Marine Megafauna
Volume 9 - 2022 | https://doi.org/10.3389/fmars.2022.863602
 Leire Lopetegui-Eguren1,2*
Leire Lopetegui-Eguren1,2* Jan Jaap Poos2,3
Jan Jaap Poos2,3 Haritz Arrizabalaga1
Haritz Arrizabalaga1 Gency L. Guirhem1,4
Gency L. Guirhem1,4 Hilario Murua5
Hilario Murua5 Nerea Lezama-Ochoa6,7
Nerea Lezama-Ochoa6,7 Shane P. Griffiths8
Shane P. Griffiths8 Jon Ruiz Gondra1
Jon Ruiz Gondra1 Philippe S. Sabarros9,10
Philippe S. Sabarros9,10 José Carlos Báez11,12Maria José Juan-Jordá1
José Carlos Báez11,12Maria José Juan-Jordá1Oceanic whitetip shark (Carcharhinus longimanus) is an important top predator in pelagic ecosystems currently classified as globally Critically Endangered by the International Union for the Conservation of Nature. This species is incidentally caught by fisheries targeting highly migratory tunas and billfishes throughout the Indian Ocean. Understanding the temporal, spatial and environmental factors influencing the capture of this species is essential to reduce incidental catches. In this study, we used generalized additive models to analyze the spatio-temporal distributions of the juvenile oceanic whitetip shark catches and the environmental conditions in the western Indian Ocean using observer data from 2010 to 2020 of the European Union and associated flags purse seine fishery. We found sea surface temperature and nitrate concentration to be the most important environmental variables predicting the probability of catching an oceanic whitetip shark. A higher probability of capture was predicted in areas where sea surface temperature was below 24°C and with low nitrate concentrations close to zero and intermediate values (1.5-2.5 mmol.m-3). We also found a higher probability of capture in sets on fish aggregating devices than in sets on free schools of tuna. The Kenya and Somalia basin was identified to have higher probabilities of capture during the summer monsoon (June to September) when upwelling of deep cold waters occurs. We provide the first prediction maps of capture probabilities and insights into the environmental preferences of oceanic whitetip shark in the western Indian Ocean. However, the causal mechanisms behind these insights should be explored in future studies before they can be used to design spatial management and conservation strategies, such as time-area closures, for bycatch avoidance.
The incidental catch continues to be a key management problem globally for sustainable fisheries. Fisheries targeting large predatory tuna, billfish and some shark species can have particularly large impacts on a wide variety of non-target species including sharks, rays, sea turtles, seabirds, and marine mammals. In this study, we define bycatch as all non-targeted species plus the small or damaged target tuna species that are not marketed through canneries (Amandè et al., 2010). Bycatch species often have biological traits that make them more vulnerable to overfishing than most teleosts (Hall and Roman, 2013; Clarke et al., 2014). Tuna Regional Fisheries Management Organizations (tRFMOs)—responsible for conserving and managing highly migratory species of tunas and billfishes—have adopted various management measures to mitigate or reduce impacts on vulnerable bycatch taxa. However, most tRFMOs have made only modest progress in managing bycatch (Gilman et al., 2014; Juan-Jordá et al., 2018). The lack or insufficient data collection and the low level of observer coverage in some fishery programs, together with the frequent lack of compliance with conservation and management measures have all contributed to insufficient bycatch governance and management in tRFMOs (Maury et al., 2013; Clarke et al., 2014; Gilman et al., 2014). Furthermore, existing management measures have mostly focused on modifying fishing gear design and practices to reduce bycatch of vulnerable taxa, retention bans for some species, and adopting best-practices for handling and releasing-at-sea to increase their probability of post-release survival (Poisson et al., 2014; Goñi et al., 2015; Grande et al., 2020). Other measures such as bycatch limits or time-area closures to reduce interactions of fisheries with vulnerable species in tRFMOs have been limited to dolphins in the eastern Pacific Ocean (IATTC, 1999) and closures to reduce the catch of juvenile tunas (e.g. bigeye tuna in the Atlantic) and swordfish (IATTC, 2009; Hall et al., 2017; Boerder et al., 2019). The efficacy of spatial management strategies including static and dynamic time-area closures to reduce interactions with vulnerable bycatch species while keeping similar yields for target species remains a priority research area in tRFMOs (Kaplan et al., 2014; Tolotti et al., 2015b; Hilborn et al., 2022).
Understanding the temporal, spatial, and environmental factors influencing species distributions is essential to minimize the interactions of fisheries with bycatch species and identify areas of high bycatch rates (Deakos et al., 2011). Species distribution models (SDMs), also known as ecological niche models, are widely used to predict species distributions and environmental niche using species occurrence or abundance and environmental information (Elith and Leathwick, 2009). These models are increasingly being used to support the development of conservation planning and spatial management tools such as static and dynamic time-area closures (Hazen et al., 2018; Passadore et al., 2018; Welch et al., 2020). SDMs have been developed for highly migratory pelagic fish species of commercial importance such as tunas (Setiawati et al., 2015; Erauskin-Extramiana et al., 2019), and billfishes (Su et al., 2011; Rooker et al., 2012; Brodie et al., 2018). However, applying SDMs to bycatch species caught in tuna and billfish fisheries is challenging due to the difficulty of collecting quality bycatch data across vast oceanic environments (McKinney et al., 2012), including the low observer coverage in some fisheries, and the difficulties of accessing data from national observer datasets (Ewell et al., 2020). To date, SDM studies of vulnerable bycatch species have been mostly conducted in the Eastern Pacific Ocean using Inter-American Tropical Tuna Commission (IATTC) observer data from the purse seine fishery—where there is 100% observer coverage. These studies include dolphinfish (Coryphaena spp.) (Marín-Enríquez et al., 2018), sailfish (Istiophorus platypterus) (Martinez-Rincon et al., 2015), wahoo (Acanthocybium solandri) (Martínez-Rincón et al., 2012), olive ridley sea turtle (Lepidochelys olivacea) (Montero et al., 2016), and spinetail devil ray (Mobula mobular) (Lezama-Ochoa et al., 2019). Since 2012, the European Union (EU) and associated flags tropical tuna purse seine fishery have been voluntarily increasing its observer coverage over time, which is currently 100% in the Atlantic and Indian Oceans (Escalle et al., 2016; Ruiz et al., 2018). These fishery observer datasets were recently used to develop SDMs for vulnerable bycatch species, such as whale shark (Rhincodon typus) (Báez et al., 2020), spinetail devil ray (Lezama-Ochoa et al., 2020), and silky shark (Carcharhinus falciformis) (Lopez et al., 2020) in the Atlantic Ocean.
The EU and associated flags tropical tuna purse seine fishery targets skipjack (Katsuwonus pelamis), yellowfin (Thunnus albacares), and bigeye tunas (Thunnus obesus), but also incidentally catches non-target species (Amandè et al., 2012; Ruiz et al., 2018). This fishery operates using two types of fishing operations (or set types): sets associated with drifting floating objects such as Fish Aggregating Devices (FAD) designed to attract and aggregate tunas, and sets associated with free-swimming tuna schools (FSC) (Marsac, 2017). Currently, around 90% of the tuna catches by the purse seine fishery in the Indian Ocean are derived from FAD sets, while the remaining 10% comes from FSC sets (IOTC, 2021). While FSC sets are generally made on monospecific schools of tuna, FAD sets attract tuna species and other pelagic fish and non-fish species, and have 2.8–6.7 times higher catches of non-target species compared to FSC sets (Murua et al., 2021a). Incidentally captured species in both FAD and FSC sets primarily include sharks, rays, and a range of bony fishes, while the interaction with sea turtles and marine mammals is less frequent (Gray and Kennelly, 2018). Due to their specific life-history traits (generally slow growth and low reproductive potential), many shark and ray species are inherently susceptible to overfishing (Dulvy et al., 2021).
The oceanic whitetip shark (Carcharhinus longimanus) is a highly migratory top predator in pelagic ecosystems, and is the second most caught shark bycatch species in the tropical tuna purse seine fishery in the western Indian Ocean (Clavareau et al., 2020). This species was once described as one of the most abundant shark species in tropical waters worldwide (Compagno, 1984). However, in recent decades, the species has experienced marked population declines throughout the majority of its global range due to overfishing (Tremblay-Boyer et al., 2019; Pacoureau et al., 2021). In 2019, oceanic whitetip shark was classified as globally Critically Endangered by the International Union for the Conservation of Nature (IUCN) (Rigby et al., 2019), and was included in Appendix II of the Convention on International Trade in Endangered Species (CITES; www.cites.org), which imposes strict regulation of its international trade. It is also listed in Annex I, Highly Migratory Species, of the United Nations Convention on the Law of the Sea (UNCLOS; www.un.org). All tRFMOs have adopted conservation and management measures to prohibit the retention on board and trade of oceanic whitetip sharks caught in tuna and billfish fisheries (ICCAT, 2010; IATTC, 2011; IOTC, 2013; WCPFC, 2019). Despite these measures there is no evidence that the oceanic whitetip shark decline has stopped (Pacoureau et al., 2021).
Oceanic whitetip shark is epipelagic and spends most of its time within the upper 200 m of the water column, in oceanic waters between 20-28°C (Tolotti et al., 2017; Andrzejaczek et al., 2018). However, the species has been recorded diving to > 200 m and temperatures down to 7.8°C for short periods (Howey et al., 2016; Tolotti et al., 2017). These dives may be linked to foraging or navigation behavior (Howey et al., 2016). Long distance migrations have been recorded for this species as well as presumed philopatry (i.e., site fidelity) (Musyl et al., 2011; Howey-Jordan et al., 2013; Tolotti et al., 2015a). For example, in the Indian Ocean, the species has exhibited trans-equatorial movements of up to 6,500 km (Filmalter et al., 2012). Despite relevant ecological research on this species there is little understanding of the relationship between their spatial distribution and prevailing environmental conditions across large spatial oceanic scales, such as the western Indian Ocean.
The main objective of this study was to investigate the spatio-temporal distribution of the oceanic whitetip shark incidental catch related to environmental conditions in the western Indian Ocean. We used the EU and associated flags tropical tuna purse seine fishery observer data between 2010 and 2020 to develop yearly and seasonal SDMs using generalized additive models to identify areas with high incidental bycatch probability. We provide the first prediction maps of capture probabilities and insights into the environmental preferences of oceanic whitetip shark in the western Indian Ocean. Yet our model results should be further tested and validated with additional sampling and modeling approaches, before it can be used to inform spatial management and conservation strategies, such as time-area closures, to reduce the bycatch of this shark species while maintaining the yield of target species.
The EU and associated flags purse seine fishery targeting tropical tuna species in the Indian Ocean is primarily concentrated in the western Indian Ocean (Figure 1 and Supplementary Figure 1). In the western Indian Ocean, the ocean surface circulation is influenced by seasonal monsoon winds that affect biological productivity (Schott and McCreary, 2001). Two major monsoon regimes occur in the region: the winter monsoon from December to March (Northeast monsoon), and the summer monsoon from June to September (Southwest monsoon). These are separated by the spring intermonsoon (April and May) and autumn intermonsoon (October and November). During the summer monsoon, a strong upwelling occurs in the western Indian Ocean where cold and highly saline waters come to the surface, increasing primary production, particularly along the coast of Somalia to 500 km offshore (Hitchcock et al., 2000; Wiggert et al., 2006). Other mesoscale processes such as eddies, filaments, fronts, and whirls also affect biological productivity, affecting the distribution of oceanodromous species such as tunas (Orue et al., 2019). For example, the Seychelles-Chagos thermocline ridge (55°E-65°E; 5°S-12°S) features a productive open-ocean upwelling area during the winter monsoon (Hermes and Reason, 2008), while the Mozambique Channel has a complex circulation influenced by mesoscale eddies (Schott et al., 2009). Such oceanic features affect the biophysical characteristic of the water column (i.e., chlorophyll-a, temperature, salinity, dissolved oxygen), which in turn affect the abundance of pelagic species in the area (Kai et al., 2009; Escalle et al., 2015; Orue et al., 2019) and the spatial dynamics of the fisheries targeting them (Marsac, 2017).
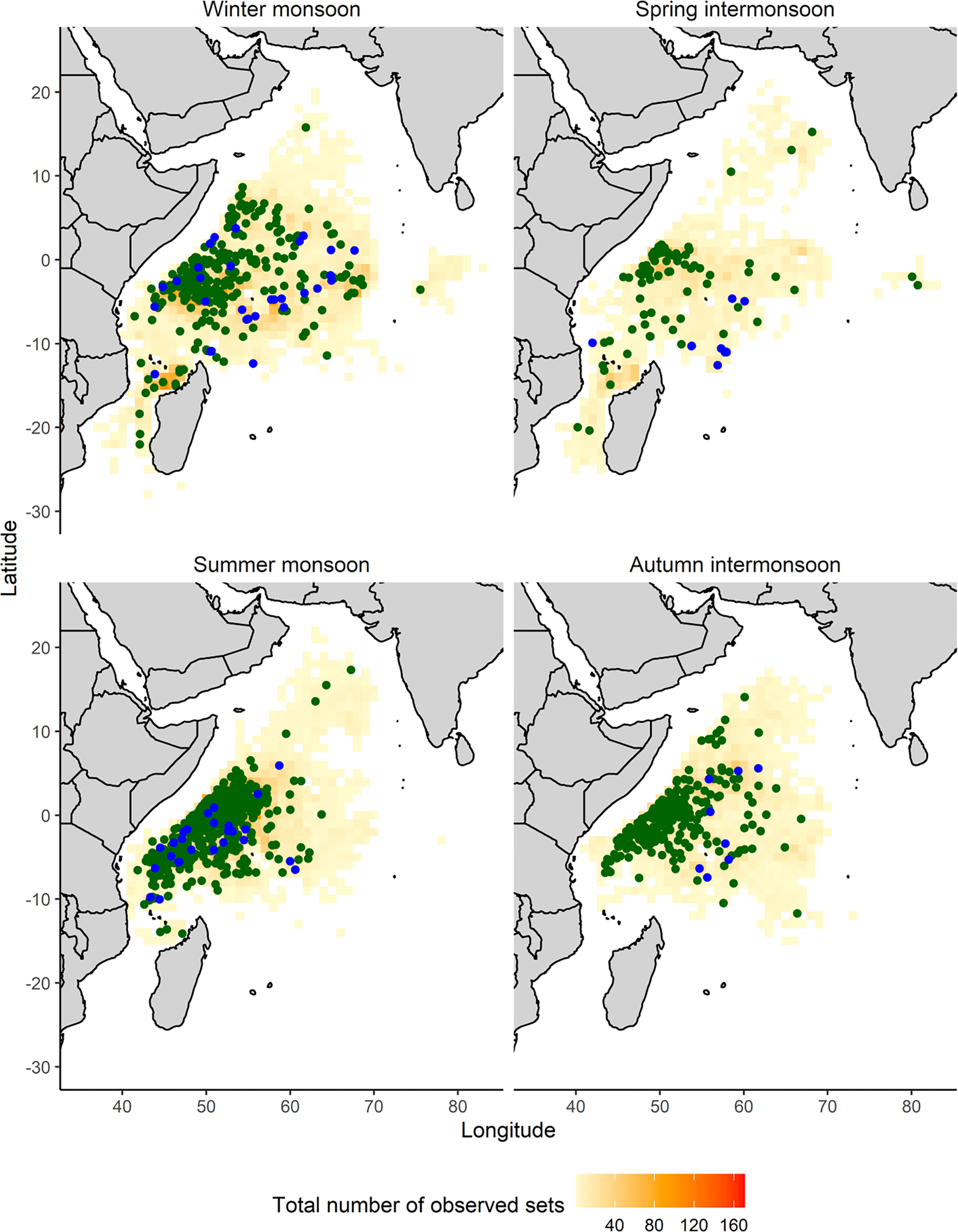
Figure 1 Spatial distribution of cumulative effort (observed sets) in the EU and associated flags purse seine fishery from 2010-2020 and observed presences of oceanic whitetip shark catch by set type (FAD in green dots and FSC in blue dots) by monsoon regimes: winter monsoon (December-March), spring intermonsoon (April and May), summer monsoon (June-September) and autumn intermonsoon (October and November). Check Supplementary Figure 1 for a better appreciation of the cumulative number of observed sets and oceanic whitetip shark catches by set type.
We used observer data collected by human observers and Electronic Monitoring System (EMS) on board the EU and associated flags (Seychelles) tropical tuna purse seine fishery between 2010 and 2020 (Supplementary Figure 2A). We analyzed a total of 26,523 observed sets, covering around 27% of all sets within this fishery between 2010-2020. About 87.4% and 12.6% of the observed sets analyzed were FADs and FSC sets, respectively (Supplementary Figure 2B). We considered all drifting floating objects as FADs irrespective whether they were artificial or natural. While the observer program started in 2003 under the EU Data Collection Framework (Regulation (EU) 2017/1004) with a 10% observer coverage, sampling effort dramatically decreased during 2009-2011 owing to the high incidences of piracy in the area. Sampling resumed in 2012 and the fleet voluntarily agreed to monitor 100% of sets. Since 2014, the observed spatial coverage has progressively increased through private contracts, presently reaching almost 100% coverage of the fleet (Goujon et al., 2017; Ruiz et al., 2018) (Supplementary Figure 2C). In 2017 EMS started to complement human observers in the task of collecting fishery data (Supplementary Figure 2A).
The observer dataset contained operational set data including set positions (longitude, latitude), dates, GMT hours, fishing set type (FAD or FSC), and vessel and observer identifiers. For each set, the total catch (in tons) and target tuna composition of the catch (skipjack, yellowfin, and bigeye tunas) and non-target species catch was recorded. The non-target catch (here referred as bycatch) was either retained for its commercial value (e.g., small tunas Auxis sp.) or discarded at sea (dead or alive) because of its low commercial value or prohibition of retention as is the case with the oceanic whitetip shark (Amandè et al., 2012; Ruiz et al., 2018).
For bycatch shark species, the number and mean size of individuals by set were recorded. The mean length of oceanic whitetip sharks caught in each set ranged from 48 to 350 cm of total length (TL), while 90.2% of the sets had average lengths < 186 cm. Given this species reach sexual maturity at 186 cm (Young and Carlson, 2020), the vast majority of individuals caught were juveniles, with individuals caught in FAD sets being slightly smaller than in FSC sets (Supplementary Figure 3). Therefore, our habitat modeling is reflective of the spatio-temporal distribution of juvenile oceanic whitetip sharks across the fishing grounds of the EU and associated flags purse seine tropical tuna fishery in the western Indian Ocean.
Environmental data were obtained at 0.25° spatial and daily temporal resolution from the EU Copernicus Marine Environment Monitoring Service (CMEMS) (https://marine.copernicus.eu/). We selected biological and physical variables based on previous research, for the position and date of each fishing set: chlorophyll-a (Chl), primary production of phytoplankton (NPPV), oxygen (O2), nitrate (NO3), phosphate (PO4) and silicate (Si) concentrations, sea surface temperature (SST), sea surface height (SSH), mixed layer depth (MLD) and salinity (Sal) (Table 1). We also extracted the eastward (Uo) and northward (Vo) velocity vectors from CMEMS and used them to calculate the eddy kinetic energy (Ke), velocity (Vel), and heading of the current (Heading). Last, we calculated chlorophyll-a and sea surface temperature fronts based on previously extracted sea surface temperature and chlorophyll-a data using a front detection algorithm (Belkin and O'Reilly, 2009).
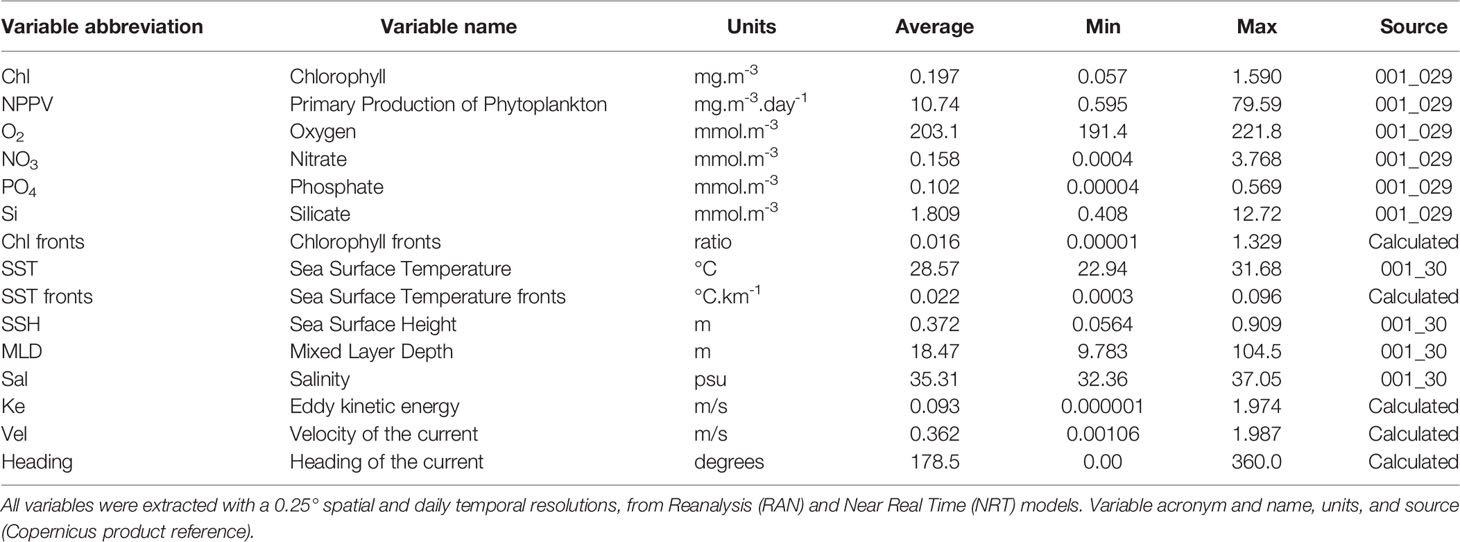
Table 1 Summary of the predictor environmental variables used in the analysis.
We analyzed the relationships between oceanic whitetip shark capture (presence/absence in the set) and environmental, fishery and spatio-temporal variables using Generalized Additive Models (GAMs). GAMs are a widely used statistical modeling tool to analyze relationships between the distribution of marine species and their environment, as they are capable of capturing non-linear relationships by fitting smoothing functions to predictor variables (Guisan et al., 2002). This method is based on the use of non-parametric smoothing functions that allows a flexible description of complex species relationships with different predictor variables (Zuur et al., 2007). The general structure of the GAM used was:
where g is the link function (logit for binomial family), μi is the expected response variable (presence/absence in our case), α is the intercept, f1 to fn are smooth functions (thin plate or cyclic cubic regression splines), and X1i to Xni are the covariates (Guisan et al., 2002).
We modelled the probability of catching an oceanic whitetip shark in an individual set as a function of the predictor variables. We modelled the catch data as present-absent and not the total catch of sharks as numbers of fish in each specific set because one single oceanic whitetip shark was caught in most of the sets (Supplementary Figure 4). Therefore, the catch of oceanic whitetip sharks was transformed to the unit of presence/absence (1/0) and considered as the dependent variable in the model.
The predictor variables considered in the modeling were environmental (Table 1), spatial (latitude and longitude), temporal (year, week, hours from sunrise) and operational information including set type (FSC or FAD) and the total catch of target and non-target species in each set. The target tuna catch (TargetTunaC, in tons) and total fish bycatch (TotalBC, in tons) of each fishing set, excluding oceanic whitetip sharks, accounted for the potential effect of the fish community size, as sharks may be found in association with other species when they are juveniles (Jacoby et al., 2012).
The degrees of freedom of the smooth functions for each predictor variable in the models were restricted to avoid over-fitting and to simplify interpretation of the results (Wood, 2006). We limited the maximum degrees of freedom (measured as number of knots, k) in the smoothing functions to k = 6 for main effects and, k = 20 for interaction effects (Lezama-Ochoa et al., 2019; Lopez et al., 2020). Each GAM was fitted using thin plate regression splines for non-linear covariates, except for week and heading variation, where a cyclic cubic regression spline was used to account for a cyclical effect, and a two-dimensional thin plate regression spline surface to account for spatial effects (latitude, longitude) of each fishing set (Wood, 2006). The gam function of the mgcv package was used to fit the model (Wood, 2014).
We used two measures to determine the correlation and multicollinearity between predictor variables before fitting the GAMs. First, all predictor variables were explored using Pearson’s rank correlation (Wood, 2006). Pairs of variables with high correlation values (|r| > 0.6) were detected (Supplementary Figure 5). The pairs of (1) sea surface temperature - oxygen, (2) sea surface temperature - primary production of phytoplankton, (3) primary production of phytoplankton - nitrate, (4) primary production of phytoplankton - oxygen and (5) salinity - latitude were highly correlated and thus we did not include them in the model at the same time (Supplementary Figure 5). Furthermore, multicollinearity between variables was examined conducting a Variance Factor Analysis (VIF) with a cut-off value of 5 (Zuur et al., 2009) using the vifstep function of the usdm package in R (Naimi et al., 2014). This function deals with multicollinearity problems by excluding highly collinear variables from a set through a stepwise procedure. Based on the VIF test, the variables total catch, chlorophyll-a and velocity of the current were excluded due to high collinearity with target tuna catch, primary production of phytoplankton and kinetic energy (Supplementary Table 1).
We applied a forward stepwise variable selection procedure to build models using the full dataset. This consisted of building the null model (intercept only model) and then adding one new covariate at a time to check its contribution to the model (Venables and Dichmont, 2004). Covariate contributions were evaluated using model Akaike Information Criterion (AIC) and studying their significance (based on p-value). We only included significant covariates (p < 0.05) and those with large relative contributions to AIC (ΔAIC > 2) in each step of the selection procedure. At the end, the final model was selected based on the lowest AIC value and the highest explained deviance (Akaike, 1974). We assessed the relative contribution of each predictor variable on the oceanic whitetip shark catch using partial effect plots. These plots show the effect of each predictor variable on the dependent variable (presence/absence of oceanic whitetip shark catch) after accounting for the average effect of all other variables in the model. Therefore, they provide an indication of how the oceanic whitetip shark catch depends on each predictor variable (Wood, 2006). We also run univariate binomial GAMs for each predictor variable, which provided information both on their raw likely contribution to the deviance explained and the potential functional shape of each predictor variable on the response variable.
We validated the final model using a cross-validation procedure (Elith and Leathwick, 2009). The dataset was randomly split into two sets: a training dataset to calibrate the model, and a testing dataset to evaluate the predictions. A k-fold cross-validation (k = 5) method was applied to split the training (80%) and testing (20%) data (Elith and Leathwick, 2009) using the kfold function from the dismo package (Freeman and Moisen, 2008) in R software (Hijmans et al., 2020). We repeated this procedure 5 times following the 5-fold cross-validation, and the performance scores’ obtained were averaged over the different random sets to evaluate the predictive performance of the distribution model (Pearson et al., 2006). Model performance was evaluated by computing a confusion matrix of the predicted and observed values using the PresenceAbsence R package (Freeman and Moisen, 2008). From the confusion matrix, we calculated the Area Under the receiver operating Curve (AUC), sensitivity (proportion of presences correctly predicted), specificity (proportion of absences correctly predicted), and the mean True Skill Statistic (TSS) validation indices (Pearson, 2010). The AUC is a threshold independent index that ranges from 0 to 1, and measures the ability of the model to correctly predict species presence or absence (Elith et al., 2006). An AUC value of 0.5 indicates that the prediction is as good as random, whereas 1 indicates perfect prediction (Fielding and Bell, 1997). The TSS index, which is calculated as sensitivity plus specificity minus 1, ranges from -1 to +1, where 0 indicates no predictive skill, +1 indicates perfect agreement, and values of zero or less indicate a performance no better than random (Brodie et al., 2015). The sensitivity, specificity and TSS indices are threshold dependent and thus, for these indices a selection of a threshold is necessary to transform the probabilities into binary predictions (presence or absence) (Jiménez-Valverde and Lobo, 2007). Different methods can be used in order to select this threshold probability value (Pearson, 2010). Based on the low prevalence (number of presences) in our dataset, we used the Maximized Sum Threshold (MST) method to establish the threshold for the accuracy indices (Liu et al., 2005; Jiménez-Valverde and Lobo, 2007). The MST method gives the most accurate predictions with low prevalence data while avoiding omission (false negative) errors.
We used the final model to predict the probability of catching an oceanic whitetip shark weekly between 2010-2020 and with a spatial resolution of 1° latitude x 1° longitude, using the predict.gam function of the mgcv package (Wood, 2014). For the model predictions, we used the environmental conditions of sea surface temperature and nitrate (the selected variables in the final model) present in each time period (each week for 11 years [2010-2020]) with the spatial resolution of 1° latitude x 1° longitude grid cell. We also used the variable FAD set type as the baseline for the predictions and set the variable of target tuna catch to mean levels. Then, we averaged the predicted weekly probabilities (and calculated the standard deviation) to obtain an overall mean prediction map (considering 11 years of weekly predictions). We also calculated seasonal prediction maps by averaging weekly probabilities for each monsoon regime: winter monsoon, spring intermonsoon, summer monsoon, and autumn intermonsoon.
In addition, we also produced yearly predictions of oceanic whitetip shark catch probabilities to assess interannual patterns using two approaches. First, we allowed the year effect (selected as significant in the final model) to vary in the predictions to explore potential interannual changes. Thus, by considering the estimated year specific coefficients, we predicted the yearly probability of oceanic whitetip capture, reflecting changes due to the environment as well as overall resource abundance and catchability. Second, we fixed the year effect (using the year 2010 as the baseline for the predictions) to examine interannual changes in the probability of oceanic whitetip shark capture solely due to the environment (Arrizabalaga et al., 2015). The ranges for the environmental variables in the environmental prediction dataset extended only 0.7% beyond the ranges observed in the environmental variables recorded in the observed sets.
The fishing grounds of this fishery varied by season (Figure 1 and Supplementary Figure 1). During the winter monsoon, the observed fishing effort spread across the equatorial region (southeast Seychelles and Chagos region) (Supplementary Figure 1). In the spring intermonsoon the fleet operated mainly in the Mozambique Channel and northwest Seychelles region using both fishing techniques (Supplementary Figures 1A, B). During the summer monsoon the fleet operated predominantly using FADs in the northwest Indian Ocean where the coastal upwelling takes place (Supplementary Figure 1A). In the autumn intermonsoon, as primary productivity levels fall and the catch rate on FADs decreases, the fleet moved into the equatorial Indian Ocean to set on free-swimming schools of tunas, which are aggregated for spawning (Supplementary Figure 1B).
The number of observed sets was lower between 2010 and 2014, compared to the later years. After 2015, the number of observed sets progressively increased (Figure 2A). Over the whole period (2010-2020), observed fishing sets were distributed across all months of the year, with slightly larger numbers during the winter monsoon (Figure 2A). Oceanic whitetip sharks were present in 4.5% of observed sets. The catch of this shark was recorded throughout the year with some apparent differences between monsoon regimes (Figures 1, 2B). The largest number of presences of oceanic whitetip sharks were recorded during summer monsoon months (Figure 2B). Moreover, of the total number of sets where oceanic whitetip sharks were present, 93.7% were FAD sets (presence in FAD sets by regime: 84% winter, 82.3% spring, 88.7% summer and 95% autumn, Supplementary Figures 1C, D).
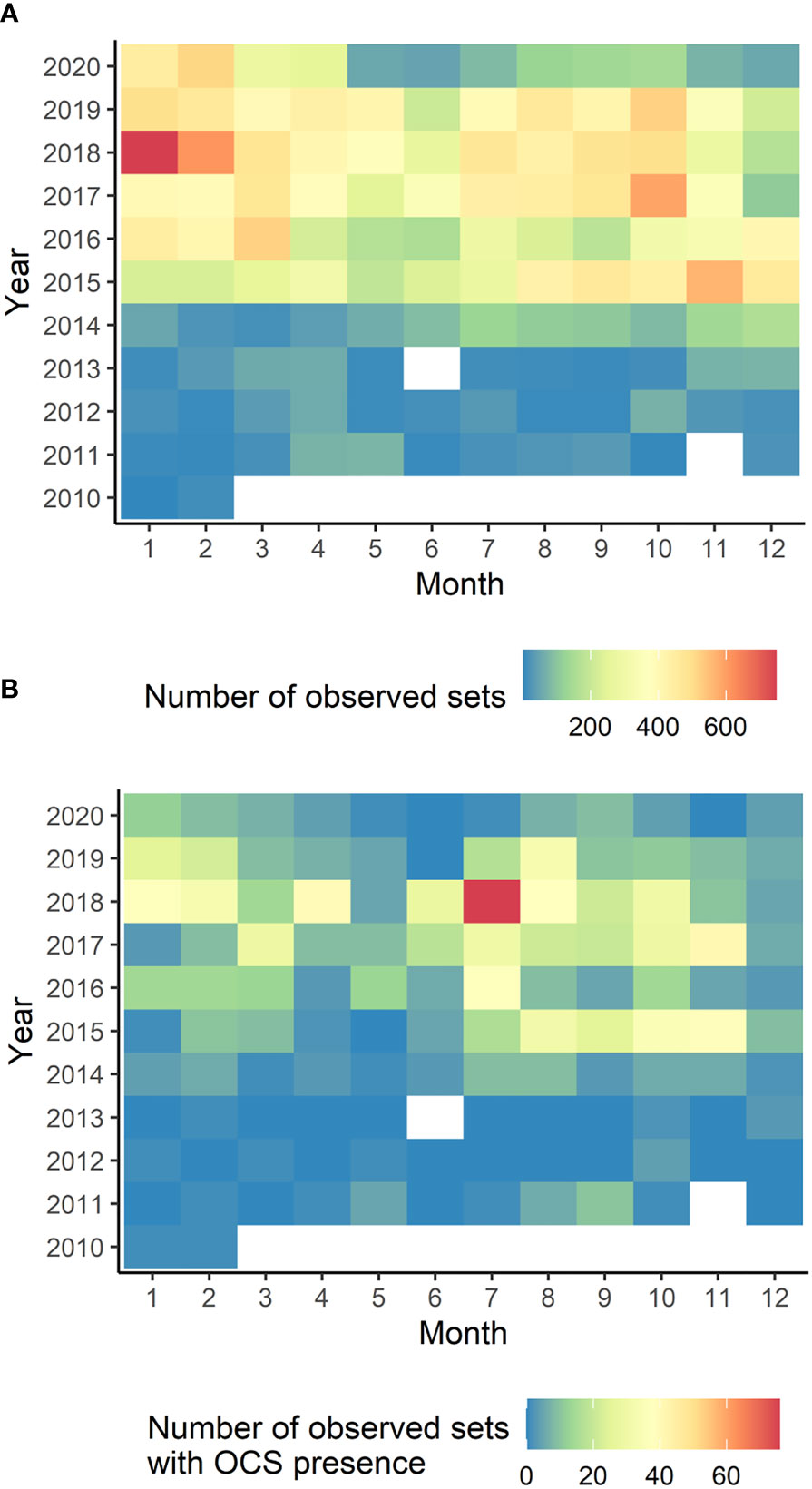
Figure 2 Temporal patterns of the EU and associated flags purse seine fishing activity (observed sets) and observed presences of oceanic whitetip shark catch. (A) Total number of observed sets in the fishery represented per month between 2010-2020. (B) Total number of sets with presence of oceanic whitetip shark catch per month between 2010-2020.
Considering the correlation and multicollinearity of covariates, we considered ten possible combinations as candidate models in the variable selection procedure (Table 2). The final model (GAM 1, Table 2) explained 8.96% of the total deviance (the most parsimonious model based on the lowest AIC value) with an adjusted r2 of 0.043 (Table 3).
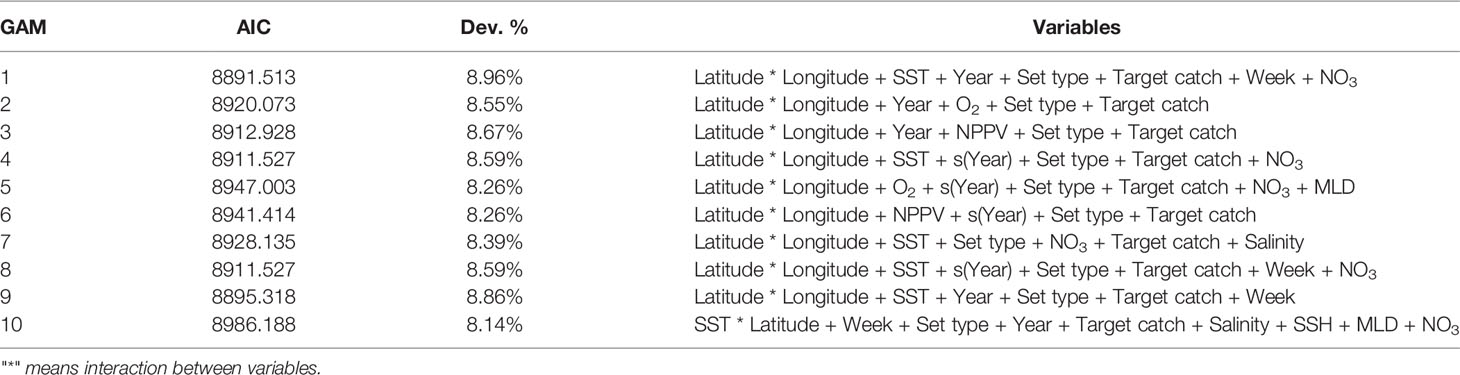
Table 2 Explored GAMs candidates with corresponding Akaike Information Criteria (AIC), explained deviance (Dev. %) values and the variables selected for each model.
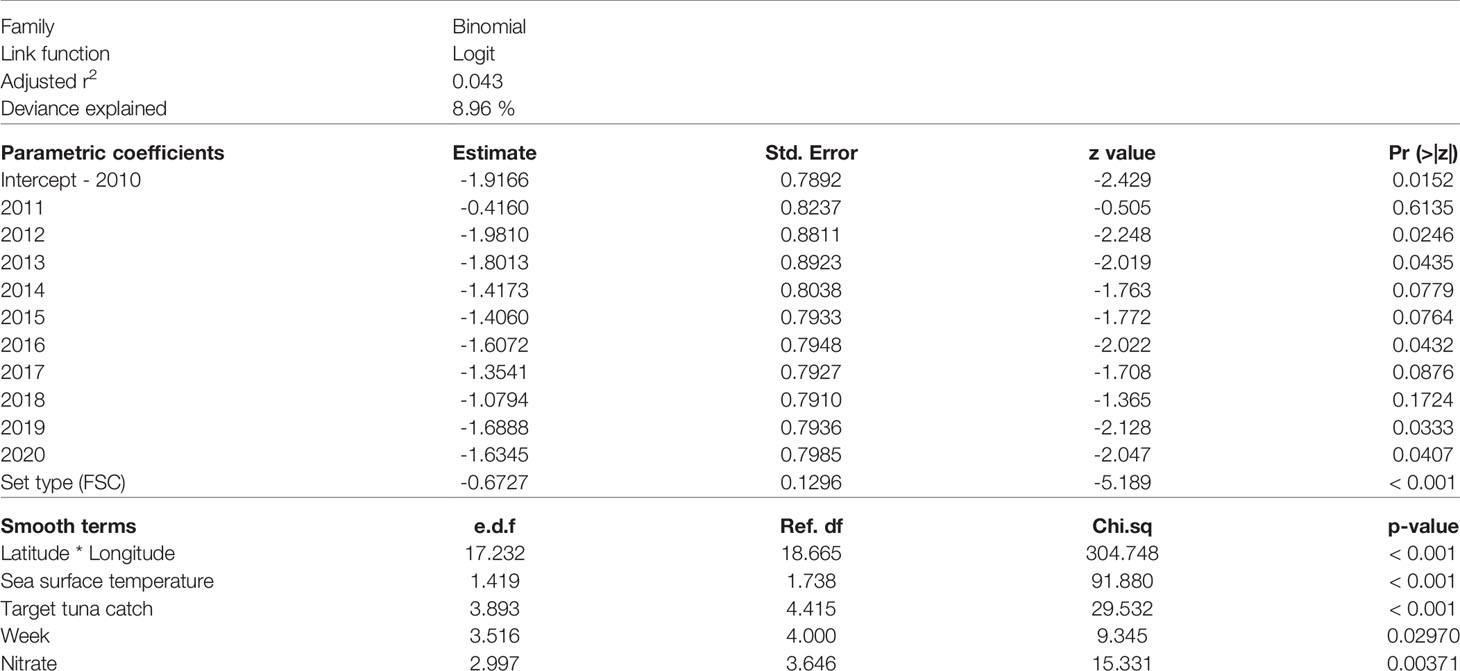
Table 3 Summary results for the parametric coefficients and smooth terms of the final GAM selected to model the probability of catching an oceanic whitetip shark in the western Indian Ocean over 2010-2020. Estimated degrees of freedom (e.d.f.).
The final model included (1) as predictors environmental variables sea surface temperature and nitrate, (2) as spatial and temporal variables the latitude and longitude interaction, year, week, and (3) as fishery related variables the set type and the total amount of target tuna caught per set (Supplementary Table 2). The individual contribution of each variable to the model revealed the interaction between latitude and longitude (5.30%), sea surface temperature (3.55%) and nitrate (1.39%) were the most significant variables (Supplementary Table 3).
The final model indicated a significant interaction between latitude and longitude and highlighted the area off the Kenya-Somalia coast as an area with a higher probability of catching an oceanic whitetip shark (Figure 3). The model also indicated a linear increase in the catch probability with decreasing temperatures, with higher probabilities in areas with lower sea surface temperature (< 24°C) relative to the range encountered by the fishery (23-32°C, Figure 4A). Higher catch probabilities were also predicted in areas with low values of nitrate (NO3) close to zero and intermediate values of nitrate (1.5-2.5 mmol.m-3), relative to the range encountered by the fishery (0-3.77 mmol.m-3, Figure 4B). There was also a relationship between the probability of catching an oceanic whitetip shark and the total target tuna catch in the set, with higher catch probabilities observed in sets with relatively large target tuna catch (between 200 and 300 tons, Figure 4C). The time of the year also affected the catch probability, with higher probabilities peaking around week 15 (April, spring intermonsoon) and week 43 (October), right after summer monsoon (Figure 4D).
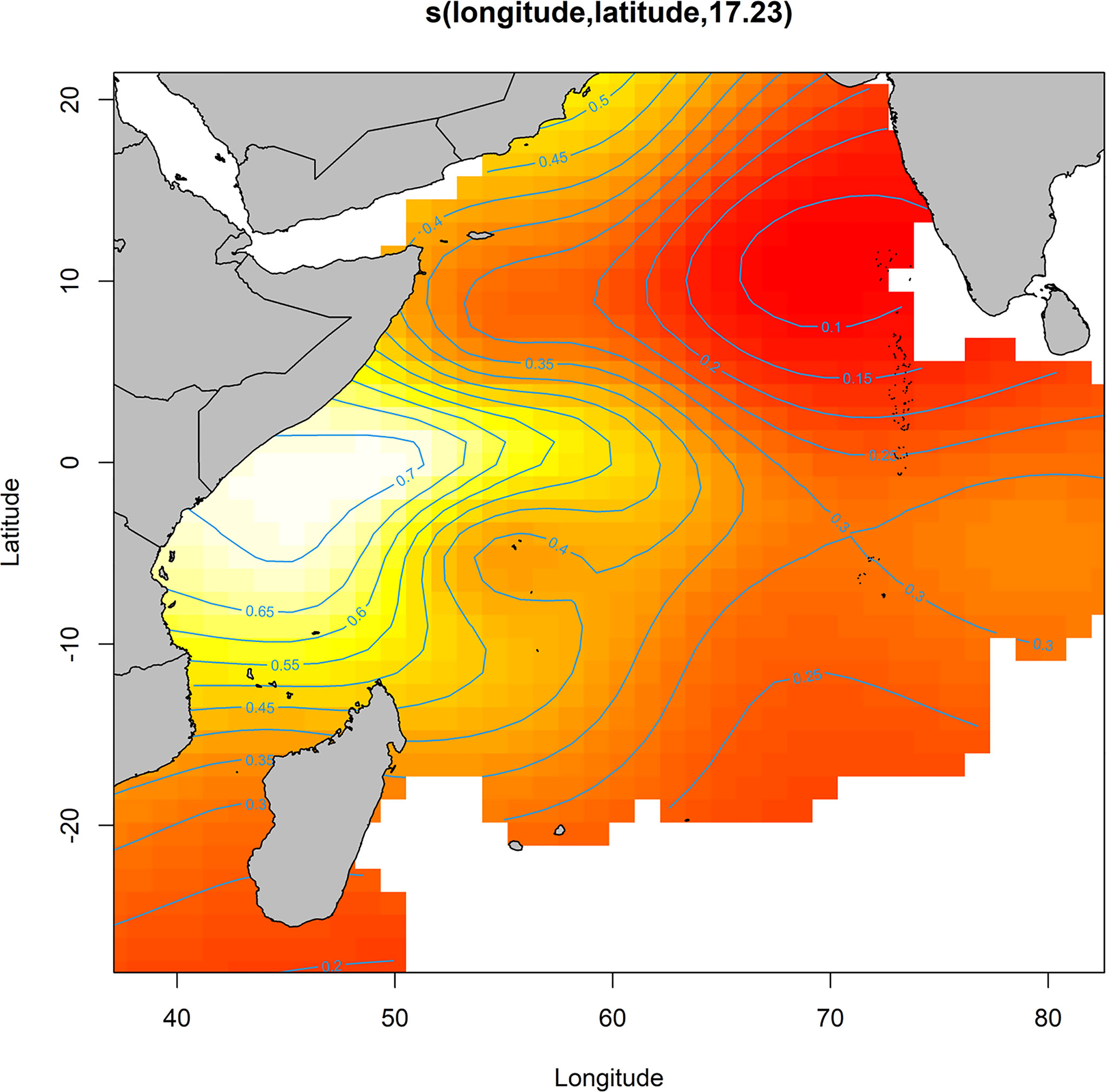
Figure 3 Partial effect of the interaction between latitude and longitude in the final model for predicting the catch probability of an oceanic whitetip shark. Isoclines indicate the catch probability.
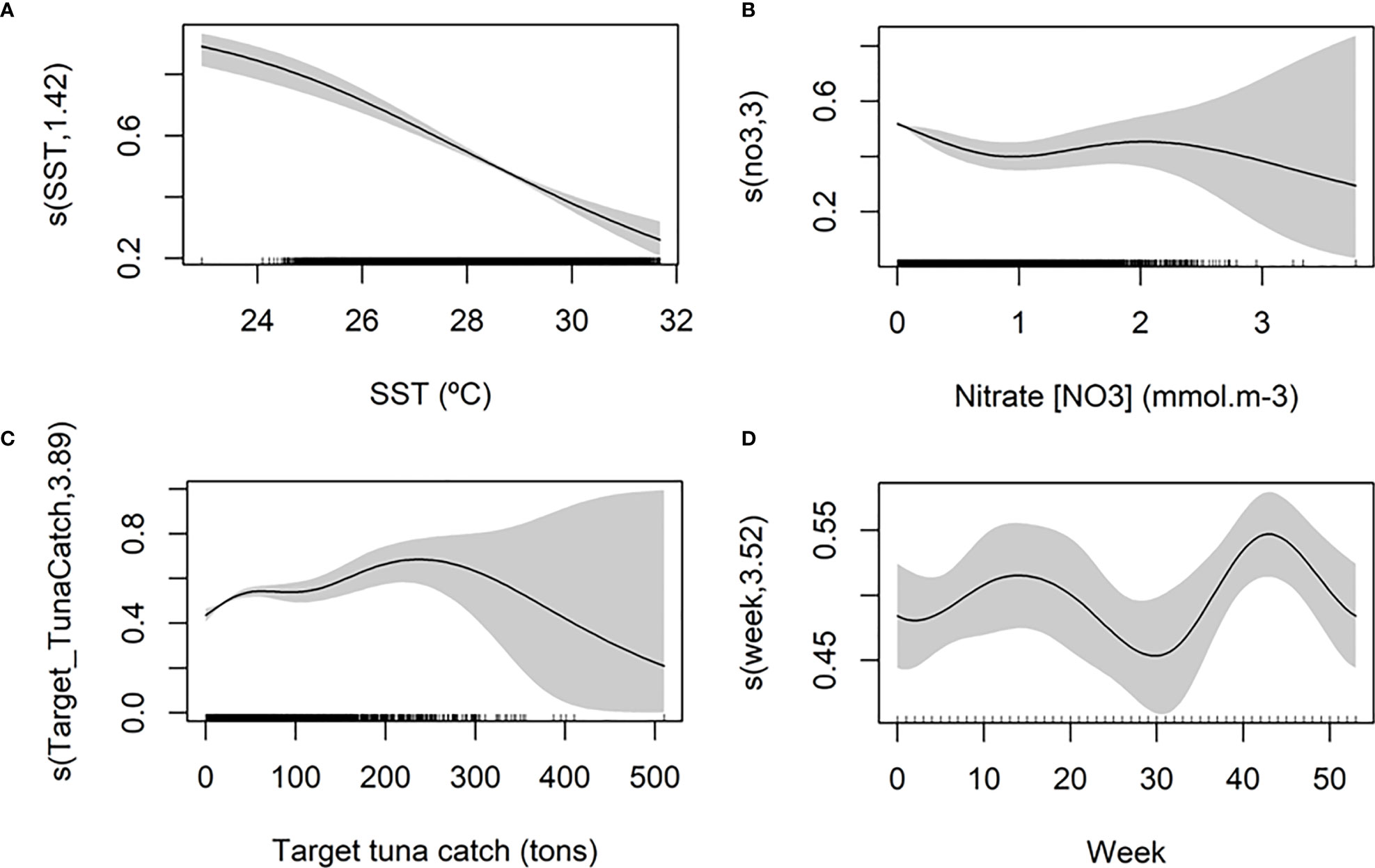
Figure 4 Partial effects of four predictor variables in the final model for predicting the catch probability of an oceanic whitetip shark. (A) Sea surface temperature (SST) (°C), (B) nitrate (mmol.m-3), (C) target tuna catch (tons) and (D) week. Effect size for each variable is expressed in probability units (0-1) and indicates the individual contribution of each variable to total catch probability. Shaded polygons indicate approximate 95% confidence intervals for smooth terms, and the black vertical lines in the x-axis the distribution of the data.
The year and set type modelled as categorical variables also contributed to explain the catch of an oceanic whitetip shark (Figure 5). Higher probabilities of capture were predicted at the beginning of the study period (year 2010 and 2011), followed by a decrease in 2012 and stabilization for the rest of the study period (Figure 5A). Finally, higher probabilities of oceanic whitetip shark capture were observed in FAD sets compared to FSC sets (Figure 5B).
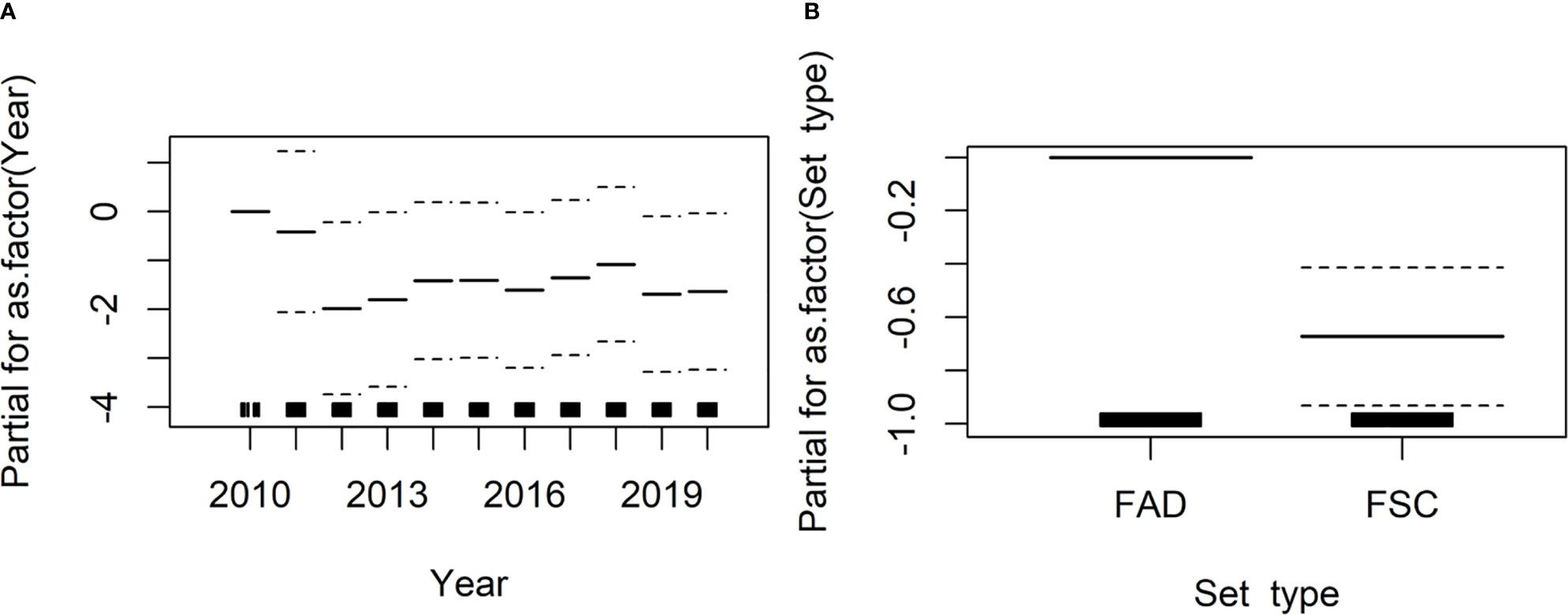
Figure 5 Partial effects of two categorial predictor variables in the final model for predicting the catch probability of an oceanic whitetip shark. (A) Year, and (B) Set type (FAD or FSC). The dashed lines indicate the 95% confidence interval for the parametric terms, and black boxes in the x-axis the distribution of the data.
The accuracy indices used to evaluate the model performance showed moderate values (AUC: 0.72, Sensitivity: 0.68, Specificity: 0.68, TSS: 0.36, Table 4) suggesting a fair model accuracy. Despite the low prevalence of the species (4.5%) in the total observed sets, the model was able to predict the probability of catching an oceanic whitetip shark and identify areas with higher catch probabilities for this species. Yet the moderate values of the accuracy indices imply that the model may have over or under predicted catch probabilities.
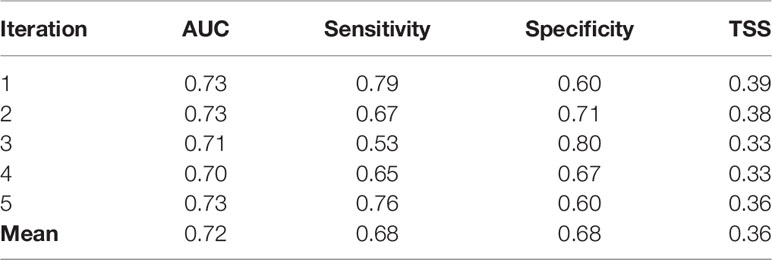
Table 4 Accuracy indices to evaluate the predictive performance of the model: Area Under the receiver operating Curve (AUC), Sensitivity, Specificity and True Skill Statistic (TSS).
The overall mean predictions of the probability of catching an oceanic whitetip shark over 2010-2020 suggest higher catch probabilities in the area offshore of Kenya-Somalia coast (Figure 6). The seasonal mean predictions for each monsoon period indicated that the high probability of capturing an oceanic whitetip shark in the Kenya-Somali basin remained throughout all the seasons but its extent varied seasonally (Figure 7). The probability of capture was higher and extended to a larger area during the summer monsoon (June-September), reaching down to the southern Mozambique Channel. During the winter monsoon (December-March) the capture of this species was also predicted off the coast of Oman (Figure 7).
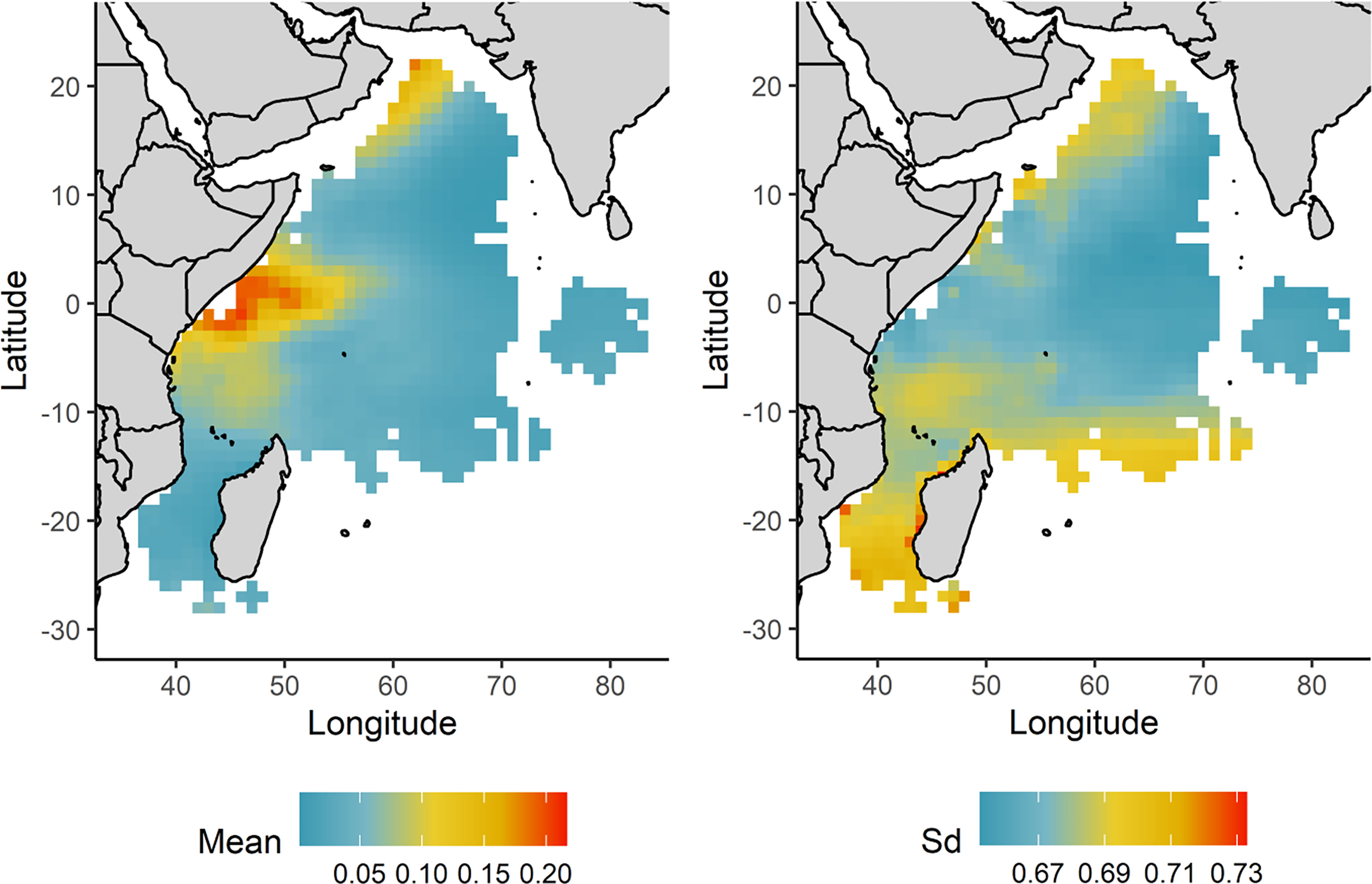
Figure 6 Spatial distribution of the mean and standard deviation (Sd) predicted catch probability of an oceanic whitetip shark over 2010-2020 and across the fishing grounds of the tropical tuna purse seine fishery in the western Indian Ocean. For predictions the FAD fishing technique was used as baseline.
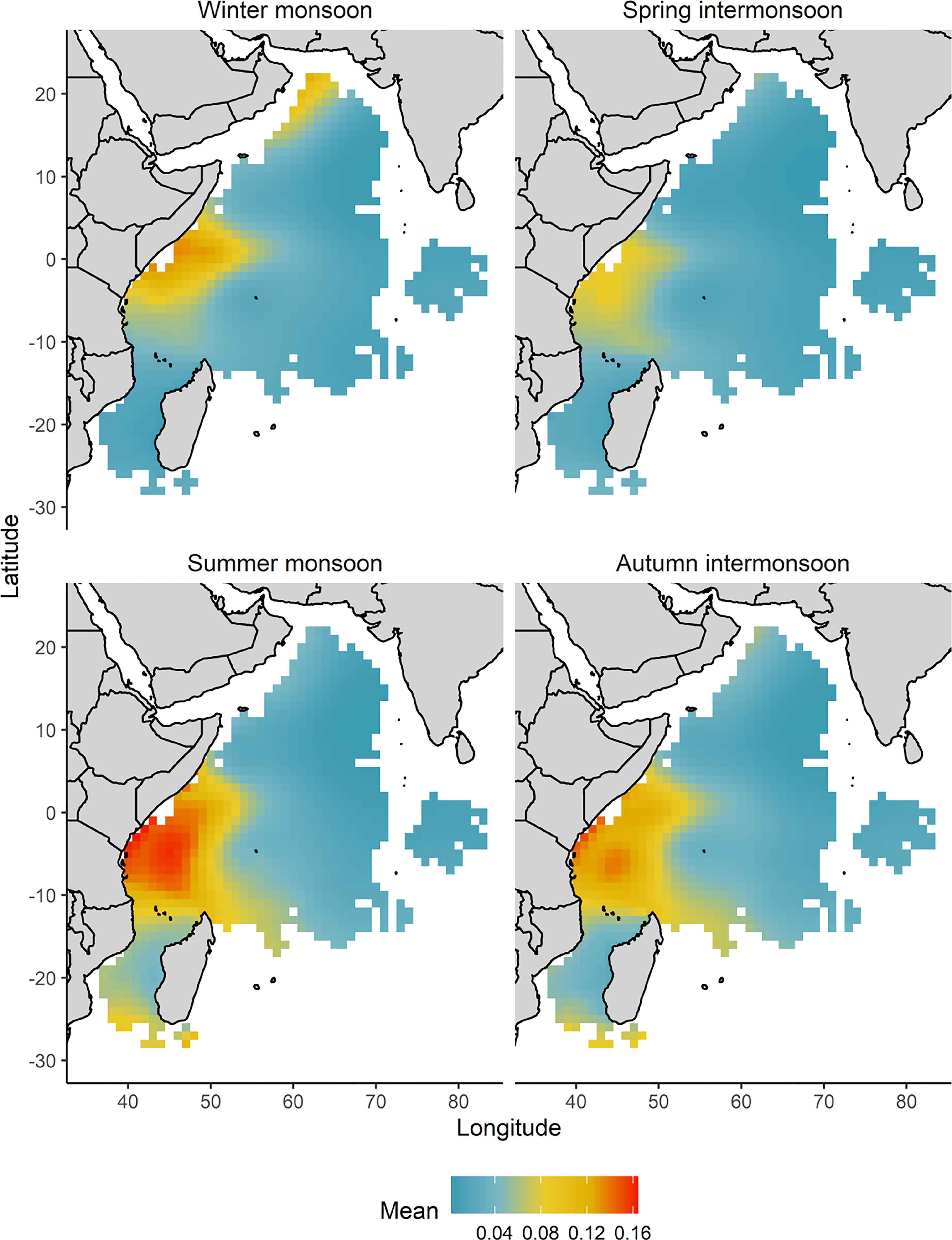
Figure 7 Spatial distribution of the mean predicted catch probability of an oceanic whitetip shark by monsoon regimes and across the fishing grounds of the tropical tuna purse seine fishery in the western Indian Ocean. Winter monsoon (December-March), spring intermonsoon (April and May), summer monsoon (June-September) and autumn intermonsoon (October and November) in the western Indian Ocean. For predictions the FAD fishing technique was used as baseline.
Yearly predictions of the probability of catching an oceanic whitetip shark showed the highest catch probability in the Kenya-Somali basin at the beginning of the study period (year 2010 and 2011) and intermediate catch probability in the year 2018 relative to the rest of the period (Figure 8A).
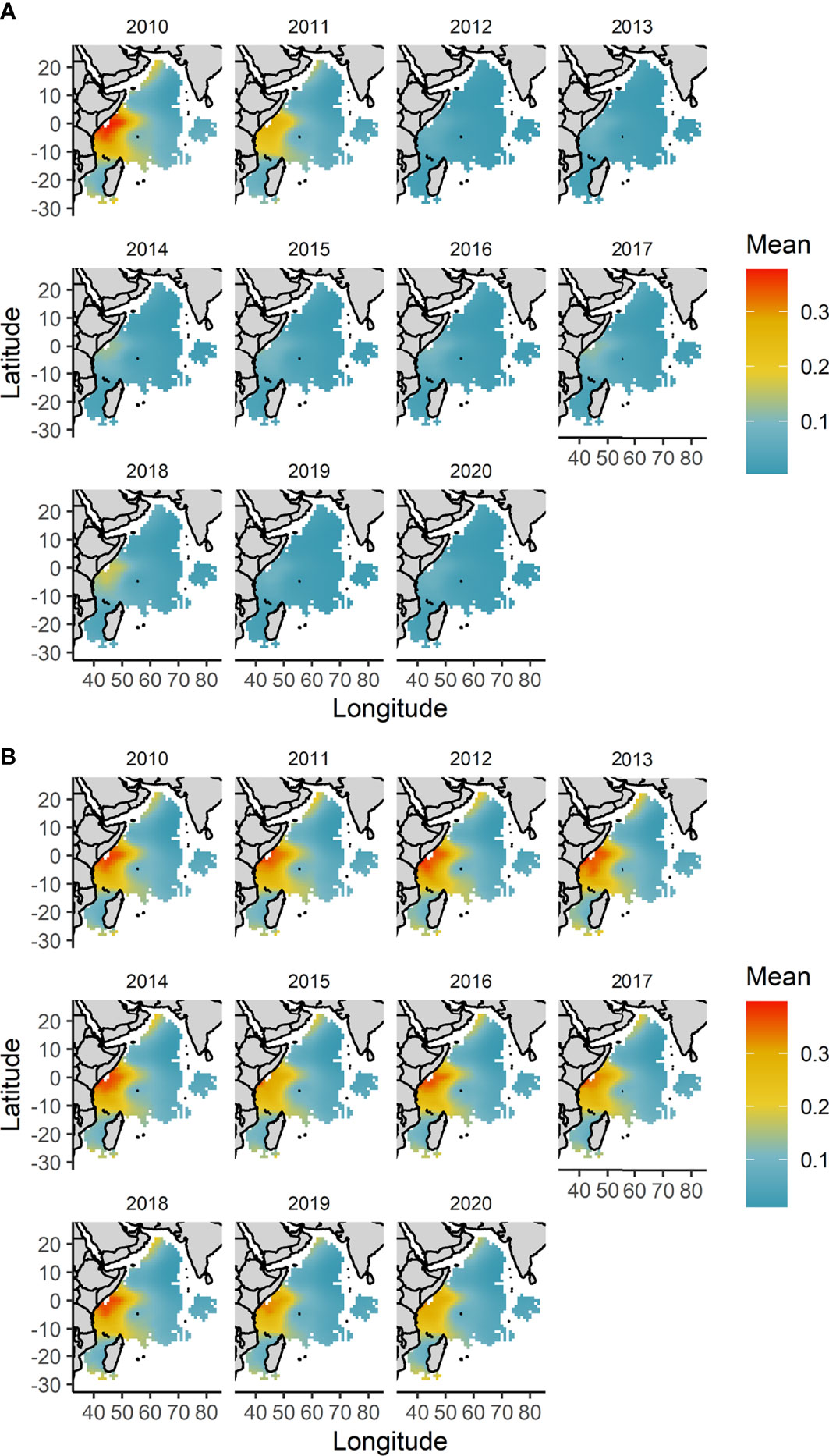
Figure 8 Spatial distribution of the yearly mean predicted catch probability of an oceanic whitetip shark between 2010-2020 and across the fishing grounds of the tropical tuna purse seine fishery in the western Indian Ocean. (A) Model predictions account for both the year effect and changing environmental conditions. (B) Model predictions are made fixing the year 2010 as baseline and only allowing the environmental variables of sea surface temperature and nitrogen concentration to change interannually. For predictions the FAD fishing technique was used as baseline.
When only the environmental predictor variables were allowed to vary and the year 2010 was used as baseline, the interannual variation in the catch probability was substantially reduced. However, some interannual variability was also observed (Figure 8B). Predictions were consistent over time, showing a higher probability of catching an oceanic whitetip shark in the Kenya-Somali basin, though this probability was lower in the years 2015, 2019 and 2020 relative to the other years (Figure 8B).
Modeling and predicting the distribution of oceanic species is challenging due to the difficulties in obtaining extensive spatial and temporal sampling coverage of oceanic environments. The approach taken in the current study is a practical and cost-effective approach to glean important aspects of the distribution of species for which data are scant, but which require management because of their vulnerability to fishing. While in this research we identified the main factors analyzed in this study driving the probability of catching a juvenile oceanic whitetip shark in the western Indian Ocean, the results should be interpreted with caution. Our species distribution model explained a small proportion of total deviance (9%). However, such low explained deviance is common in studies modeling the spatio-temporal distributions of data poor bycatch species (10.2-19%) (Lopez et al., 2017; Lezama-Ochoa et al., 2020). Studies modeling the spatial distributions of commercially important target species of tunas usually explain a higher percentage of total deviance in the models (33.7-62.4%) (Su et al., 2011; Erauskin-Extramiana et al., 2019), in part because these studies include data from fisheries that cover a large (or even whole) area of the species distribution.
Sea surface temperature was the most important environmental predictor explaining the catch of a juvenile oceanic whitetip shark in the purse seine fishery, suggesting higher catch probabilities decreasing linearly with increasing temperature. Due to the small thermal range encountered by the fishery in the study area (23-32°C), the model was not able to find a unimodal distribution of preferred temperatures as expected based on the ecological niche theory (Hutchinson, 1957). However, our results are consistent with existing tagging studies that found out that the thermal range of preference of this shark is between 20-28°C (Musyl et al., 2011; Tolotti et al., 2017; Andrzejaczek et al., 2018). Sea surface temperature of 28°C marked a distinct change in vertical movements of tagged oceanic whitetip sharks affected by thermocline depth (Tolotti et al., 2017; Andrzejaczek et al., 2018) and thus, we expected this variable to influence the spatial distribution of this species. Studies on distribution of pelagic shark species, such as blue shark (Prionace glauca), whale shark and silky shark, also suggest that sea surface temperature strongly influences the distribution of these species (Carvalho et al., 2011; Afonso et al., 2014; Hacohen-Domené et al., 2015; Lezama-Ochoa et al., 2016; Lopez et al., 2020). Other studies in the Indian Ocean have also found out that sea surface temperature to be the main environmental predictor explaining the distribution of pelagic fish species such as albacore (Thunnus alalunga) (Chen et al., 2005), bigeye (Lee et al., 2005), yellowfin and skipjack tunas (Arrizabalaga et al., 2015).
The other environmental variable selected in the final model was nitrate concentration, yet its contribution to the model was small. We expected to find increasing probabilities of capturing oceanic whitetip shark with increasing concentrations of nitrate, as nitrate increases phytoplankton growth (Dugdale, 1967), which is indicative of high primary production, and therefore, it may be associated with areas known to attract pelagic species for feeding (Young et al., 2015; Williamson et al., 2019). Instead, we found higher probabilities of catching oceanic whitetip shark in areas with low values of nitrate (close to zero) and intermediate values of nitrate. It may be that there is a spatial and temporal lag between the coastal upwelling of colder and nutrients-rich waters and the occurrence of oceanic whitetip sharks, as it has been observed in other pelagic species (Barlow et al., 2021). In addition, low concentrations of nitrate can be an indicative of oligotrophic waters where distribution of zooplankton and schools of foraging preys are more patchy (Priyadarshi et al., 2019). We suggest that oceanic whitetip shark could be associated with low-nitrogen oligotrophic waters as it might facilitate, as a visual predator, its foraging behavior on patchy prey distributions (Collin, 2018). This is supported by other studies with higher catches of other visual predators, such as tunas, along the offshore side of an upwelling frontal boundary, which is characterized with high frontal occurrence supporting schools of forage fish, yet clear, low nutrient and low-chlorophyll waters facilitating foraging for visual predators (Hahlbeck et al., 2017). Its association with low-nitrogen oligotrophic waters could also be a behavioral mechanism to avoid competition and predator avoidance (Lima and Dill, 1990; Brown and Kotler, 2004), yet this explanation for juvenile oceanic whitetip sharks needs to be further examined. Moreover, the range of nitrate values found in our study area (0-3.8 mmol.m-3) was very low compared to other research studies modeling other migratory species, such as the spinetail devil ray (Mobula mobular) distribution in upwelling areas in the eastern Pacific Ocean where the nitrate concentrations were around 145.2 mmol.m-3 (Lezama-Ochoa et al., 2019). The oceanic regions in the Indian Ocean are characterized by low concentrations of nitrate, in comparison to other oceanic regions in the Atlantic and Pacific Oceans (Pennington et al., 2006). These low concentrations might also confound the interpretation of the effect of nitrate in the oceanic whitetip shark catch probability.
We also found higher probabilities of juvenile oceanic whitetip shark catch when sets were made under FADs compared to FSC of tunas. This is consistent with the association behavior and attraction of juvenile oceanic whitetip sharks to FADs (Filmalter et al., 2013). The affinity of pelagic species with FADs is not fully understood but a widely accepted theory relies on their social behavior (Jacoby et al., 2012) and suggests that FADs may act as a “meeting point” (Dagorn and Fréon, 1999). While it has been hypothesized that FADs may be acting as an “ecological trap” for pelagic species aggregating underneath them impacting their behavior, biology, and migration (Marsac et al., 2000), these factors are little known for oceanic whitetip shark and other sharks. Furthermore, the majority of the observed sets in our study were made in FADs, and our models did not account for fishing effort and FAD densities. We recommend future studies to explore the effect of the densities, drift and distribution of FADs on the spatial distribution and behavior of oceanic whitetip shark and other pelagic species with aggregation behaviors to FADs.
Higher catch probabilities were also predicted at the beginning of the study period (year 2010 and 2011), followed by a decrease in 2012 and stabilization for the rest of the study period. While this temporal trend in the year effect may suggest a population decline in oceanic whitetip shark and then a stabilization of this species in the study area, we need to interpret this result with caution as the number of observed sets in the first two years (2010 and 2011) were considerably lower resulting in high uncertainty in these early years. However, this explanation cannot be completely ruled out as over the last decades large declines in the abundance of oceanic whitetip shark have been reported throughout its global range (Rigby et al., 2019; Tremblay-Boyer et al., 2019; Pacoureau et al., 2021), including the Indian Ocean (IOTC, 2015; Young and Carlson, 2020). Population declines in oceanic whitetip shark are believed to have been primarily driven by the increasing susceptibility of this species to different fishing gears (primarily longline and gillnets, followed by purse seine fisheries) in the Indian Ocean (Garcia and Herrera, 2019), together with its sensitive life history traits (low fecundity, slow-moderate growth rate, and late sexual maturity), making this shark species to be highly vulnerable to overfishing (Young and Carlson, 2020).
The model predicted a higher catch probability of an oceanic whitetip shark offshore Kenya-Somalia coast during summer monsoon suggesting this may be an important area for this species. Considering that most of the catches of oceanic whitetip shark through all the study area were juveniles, we suggest that this upwelling area may be used as a feeding ground by the young individuals of this species. In line with our results, previous studies on the distribution and mortality of shark species caught as bycatch also found catches of juvenile oceanic whitetip shark to be localized near the Somalian coast (Clavareau et al., 2020). Other studies also found this area to be highly suitable for pelagic species such as silky sharks (Lezama-Ochoa et al., 2016) and adult individuals of blue sharks (Coelho et al., 2018). The seasonality of the higher catch probabilities in this important area appears to be associated to the upwelling seasonality in Kenya-Somali basin (Schott and McCreary, 2001). We found that the highest catch probabilities occurred during the summer monsoon regime when the upwelling decreases sea surface temperature and increases biological productivity (Supplementary Animations 1, 2), thus attracting schools of prey fishes to top predators (Young et al., 2015). In line with this, other studies have also suggested this area to be a feeding area for migratory pelagic species such as skipjack tuna (Druon et al., 2017), and blue whale (Balaenoptera musculus) (Charles et al., 2012) during the summer monsoon.
Interannual predictions in response to only environmental changes in sea surface temperature and nitrate concentrations showed high consistency in the higher catch probabilities of this species in the Kenya-Somali basin, yet with some interannual variation in this area. Interannual climatic processes are well known to affect the spatio-temporal distributions of pelagic migratory fish species (Bacha et al., 2017; Kai et al., 2017; Marsac, 2017). We observed the catch probability of this species decreased slightly in the Kenya-Somali basin in the years 2015, 2019 and 2020 driven by higher sea surface temperature values observed during these years explained by marine heatwaves associated to Indian Ocean basin wide warming and positive Indian Ocean Dipole events (Zhang et al., 2018; Holbrook et al., 2019; Saranya et al., 2021; Shi and Wang, 2021). This ocean-atmosphere phenomenon resulted in a higher sea surface temperature in the western side of the Indian Ocean as well as a less intense Somali upwelling with a reduction in the nitrate concentrations and the primary production in the western Indian Ocean (Yang et al., 2020). The frequency, duration and intensity of heatwaves are also increasingly affecting the distribution of coastal and oceanic pelagic species leading to shifts in their distributions (Lonhart et al., 2019; Cheung and Frölicher, 2020). Furthermore, as studied in other shark species, projected increases in sea surface temperature induced by climate change could also redistribute and shift oceanic whitetip distributions to more southern waters in the Indian Ocean (Sequeira et al., 2014; Lezama-Ochoa et al., 2016) or drive them to deeper waters (Dulvy et al., 2008; Tolotti et al., 2017; Andrzejaczek et al., 2018) with unknown consequences to their populations. Climate-driven changes on shark distributions and the timing of their migrations can also impact their risks to fisheries by decreasing their spatial protection, as it has been observed in the apex predator tiger shark (Galeocerdo cuvier) in the northwest Atlantic Ocean (Hammerschlag et al., 2022). Considering the sea surface temperature effect on the distribution of the oceanic whitetip shark and that climate change is increasing the positive Indian Ocean Dipole events and thus marine heatwaves (Cai et al., 2009; Frölicher et al., 2018; Oliver et al., 2019), understanding the effect of multiple climate change scenarios in combination with its interactions with multiple fisheries in the Indian Ocean is necessary to ensure that this globally Critically Endangered species is recovered to sustainable levels.
It is important to emphasize that the species distribution model for juvenile oceanic whitetip shark derived in the present study is based on data collected by only the EU and associated flags tropical tuna purse seine fishery, which mostly captures juveniles and operates in the western part of the Indian Ocean. This obviously limits the applicability of our model and interpretations to the region of the Indian Ocean where the fishery operates, to the years where the fishery collected observer data, and to the limited size range of oceanic whitetip sharks that are susceptible to capture by the fishery. Future modeling efforts could improve on our efforts by collecting and analyzing datasets from multiple fisheries including longline and gillnet fisheries that are also known to interact with this species throughout the Indian Ocean (Garcia and Herrera, 2019). Not only will this approach increase the spatial coverage of the model to the entire Indian Ocean but also provide greater coverage of the population’s length distribution as longline gear tends to capture larger individuals than purse seine. This will ultimately improve the predictive abilities of the model, as well as its validation, and provide an improved representation of the overall distribution of this species.
Using exclusively fisheries-dependent data to model the distribution of species can also result in biased models because catch probabilities are biased by fisheries behavior that is limited to the fishing grounds (Montero et al., 2016; Pennino et al., 2016). However, fishery-dependent data may be less biased for bycatch species as these are non-targeted and fishing locations are not selected to maximize the catch of these species (Pennino et al., 2016). To account for these potential biases, future modeling studies could also attempt to include additional fisheries independent data such as acoustic or satellite tagging data and if possible and use them to validate the models to identify if the predictions accurately represent the observed species distributions. Tagging studies not only become critical for model validation purposes, but to also understand the habitat utilization in three dimensions. Long distance migrations (up to 6,500 Km) have been recorded by tagging studies in oceanic whitetip sharks in the Indian and central Pacific Oceans (Musyl et al., 2011; Filmalter et al., 2012). Despite the growing number of tagging studies focusing on oceanic whitetip shark movements (Mejuto et al., 2005; Musyl et al., 2011; Filmalter et al., 2012; Howey-Jordan et al., 2013; Howey et al., 2016; Tolotti et al., 2017; Andrzejaczek et al., 2018), the migrations patterns and habitat utilization in two and three dimensions of the oceanic whitetip shark in the western Indian Ocean are still poorly known.
Species distribution models are increasingly being used to inform and guide spatially based fisheries management measures to minimize the interactions of bycatch species with fisheries (Hobday et al., 2010; Urbisci et al., 2016; Hazen et al., 2018), as well as in marine spatial planning and biodiversity conservation efforts (Marshall et al., 2014; Passadore et al., 2018). This study examines the spatio-temporal distributions of the juvenile oceanic whitetip shark using generalized additive models and provides the first prediction maps of capture probabilities and insights into its environmental preferences in the western Indian Ocean. While our model results suggest that the area off Kenya-Somalia during the summer monsoon (June-September) could be a potential area to be protected to reduce shark interactions, we think the model results are preliminary. Therefore, they should be treated as a working hypothesis to be further tested and validated (Jarnevich et al., 2015) before they are used to inform spatial management measures to reduce shark interactions in the purse seine fishery operating in the western Indian Ocean. To design and apply effective spatial management measures to reduce shark interactions in this fishery, we think it is necessary (1) to improve the species distribution model for the oceanic whitetip shark with additional sampling beyond the spatial coverage of our study area (e.g. including observer data collected by longline and other fisheries), additional modeling (e.g. comparing with alternative algorithms such as Boosted Regression Trees, Bayesian Additive Regression Trees and ensemble methods), and as well as model validation with alternative data sources (e.g., using tracking data), (2) understand better the co-occurrence of multiple bycatch shark species caught by the purse seine fishery (e.g., silky shark) together with the occurrence of the targeted tropical tuna species, in order to evaluate the effectiveness of potential spatial management measures across multiple species of interest and avoid piecemeal bycatch management measures (Gilman et al., 2019; Pons et al., 2022), and (3) closely evaluate the cost to the fisheries and a consequence of fishing reallocation of the time-area closures (Watson et al., 2009). In the short term, we also think that spatially explicit ecological risk assessment approaches, which have been developed as an alternative to traditional fishery stock assessment models for bycatch in data-limited settings (Murua et al., 2021b), can also assist in informing and evaluating the complementarity of management measures to reduce shark interactions. The most recent development is EASI-Fish (Ecological Assessment of the Sustainable Impacts of Fisheries) (Griffiths et al., 2019), which is capable of estimating the cumulative impact of multiple fisheries on vulnerable data-limited bycatch species. The species distribution model for oceanic whitetip shark derived in this paper (as well as its future refinements) together with this risk assessment approach would allow the evaluation of the potential impact of current and other hypothetical management measures including time-area closures on the vulnerability status of oceanic whitetip shark in the Indian Ocean. This information is crucial to reduce fishery interactions and mortality of the oceanic whitetip shark in the Indian Ocean and ensure the conservation of this species.
The data analyzed in this study is subject to the following licenses/restrictions: The datasets for this article are not publicly available because they reflect individual fishing strategies of private fishing vessel companies. Requests to access these datasets should be directed to Jon Ruiz Gondra,anJ1aXpAYXp0aS5lcw==; Jose Carlos Báez, am9zZWNhcmxvcy5iYWV6QGllby5jc2ljLmVz; Philippe Sabarros,cGhpbGlwcGUuc2FiYXJyb3NAaXJkLmZy.
LL-E, MJ-J, JP, and HA conceived the study and designed the methodology. JG, PS, and JB provided the data. LL-E, MJ-J, JP, HA, and GL-G performed the analysis. LL-E and MJ-J wrote the first draft of manuscript. JP, HA, GL-G, HM, NL-O, SG, JR, PS, and JB provided feedback on the analysis and manuscript. All authors contributed to manuscript revision, read and approved the submitted version.
MJ-J and LL-E were supported by “la Caixa” Foundation Postdoctoral Junior Leader Fellowship under agreement N° 847648.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The data analyzed in this study were collected by AZTI, Institut de Recherche pour le Développement (IRD), and Instituto Español de Oceanografía (IEO) through the EU-funded Data Collection Framework (DCF, Reg (EC) 1543/2000, 1639/2001, and 665/2008) and through private contracts funded by the purse seiner associations ANABAC, OPAGAC and ORTHONGEL. We thank all the observers from the three institutions that collected the data. This is contribution number 1095 from AZTI.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.863602/full#supplementary-material
Afonso P., McGinty N., Machete M. (2014). Dynamics of Whale Shark Occurrence at Their Fringe Oceanic Habitat. PloS One 9 (7), e102060. doi: 10.1371/journal.pone.0102060
Akaike H. (1974). A New Look at the Statistical Model Identification. IEEE Trans. Auto Contr 19 (6), 716–723. doi: 10.1109/tac.1974.1100705
Amandè M. J., Ariz J., Chassot E., de Molina A. D., Gaertner D., Murua H., et al. (2010). Bycatch of the European Purse Seine Tuna Fishery in the Atlantic Ocean for the 2003–2007 Period. Aquat. Living Resour. 23 (4), 353–362. doi: 10.1051/alr/2011003
Amandè M. J., Chassot E., Chavance P., Murua H., de Molina A. D., Bez N. (2012). Precision in Bycatch Estimates: The Case of Tuna Purse-Seine Fisheries in the Indian Ocean. ICES J. Mar Sci. 69 (8), 1501–1510. doi: 10.1093/icesjms/fss106
Andrzejaczek S., Gleiss A. C., Jordan L. K. B., Pattiaratchi C. B., Howey L. A., Brooks E. J., et al. (2018). Temperature and the Vertical Movements of Oceanic Whitetip Sharks, Carcharhinus Longimanus. Sci. Rep. 8 (1), 8351. doi: 10.1038/s41598-018-26485-3
Arrizabalaga H., Dufour F., Kell L., Merino G., Ibaibarriaga L., Chust G., et al. (2015). Global Habitat Preferences of Commercially Valuable Tuna. Deep Sea Res. Part II: Top Stud. Oceanog. 113, 102–112. doi: 10.1016/j.dsr2.2014.07.001
Bacha M., Jeyid M. A., Vantrepotte V., Dessailly D., Amara R. (2017). Environmental Effects on the Spatio-Temporal Patterns of Abundance and Distribution of Sardina Pilchardus and Sardinella Off the Mauritanian Coast (North-West Africa). Fisher. Oceanog. 26 (3), 282–298. doi: 10.1111/fog.12192
Báez J. C., Barbosa A. M., Pascual P., Ramos M. L., Abascal F. (2020). Ensemble Modeling of the Potential Distribution of the Whale Shark in the Atlantic Ocean. Ecol. Evol. 10 (1), 175–184. doi: 10.1002/ece3.5884
Barlow D. R., Klinck H., Ponirakis D., Garvey C., Torres L. G. (2021). Temporal and Spatial Lags Between Wind, Coastal Upwelling, and Blue Whale Occurrence. Sci. Rep. 11 (1), 1–10. doi: 10.1038/s41598-021-86403-y
Belkin I. M., O'Reilly J. E. (2009). An Algorithm for Oceanic Front Detection in Chlorophyll and SST Satellite Imagery. J. Mar. Syst. 78 (3), 319–326. doi: 10.1016/j.jmarsys.2008.11.018
Boerder K., Schiller L., Worm B. (2019). Not All Who Wander are Lost: Improving Spatial Protection for Large Pelagic Fishes. Mar. Policy 105, 80–90. doi: 10.1016/j.marpol.2019.04.013
Brodie S., Hobday A. J., Smith J. A., Everett J. D., Taylor M. D., Gray C. A., et al. (2015). Modelling the Oceanic Habitats of Two Pelagic Species Using Recreational Fisheries Data. Fisher. Oceanog. 24 (5), 463–477. doi: 10.1111/fog.12122
Brodie S., Jacox M. G., Bograd S. J., Welch H., Dewar H., Scales K. L., et al. (2018). Integrating Dynamic Subsurface Habitat Metrics Into Species Distribution Models. Front. Mar. Sci. 5, 219. doi: 10.3389/fmars.2018.00219
Brown J. S., Kotler B. P. (2004). Hazardous Duty Pay and the Foraging Cost of Predation. Ecol. Lett. 7 (10), 999–1014. doi: 10.1111/j.1461-0248.2004.00661.x
Cai W., Sullivan A., Cowan T. (2009). Climate Change Contributes to More Frequent Consecutive Positive Indian Ocean Dipole Events. Geophys. Res. Lett. 36 (23). doi: 10.1029/2009GL040163
Carvalho F. C., Murie D. J., Hazin F. H., Hazin H. G., Leite-Mourato B., Burgess G. H. (2011). Spatial Predictions of Blue Shark (Prionace Glauca) Catch Rate and Catch Probability of Juveniles in the Southwest Atlantic. ICES J. Mar. Sci. 68 (5), 890–900. doi: 10.1093/icesjms/fsr047
Charles A., Branch T. A., Alagiyawadu A., Baldwin R., Marsac F. (2012). Seasonal Distribution, Movements and Taxonomic Status of Blue Whales (Balaenoptera Musculus) in the Northern Indian Ocean. J. Cetacean. Resour. Manage. 12 (2), 203–218.
Chen I.-C., Lee P.-F., Tzeng W.-N. (2005). Distribution of Albacore (Thunnus Alalunga) in the Indian Ocean and Its Relation to Environmental Factors. Fisher. Oceanog. 14 (1), 71–80. doi: 10.1111/j.1365-2419.2004.00322.x
Cheung W. W., Frölicher T. L. (2020). Marine Heatwaves Exacerbate Climate Change Impacts for Fisheries in the Northeast Pacific. Sci. Rep. 10 (1), 1–10. doi: 10.1038/s41598-020-63650-z
Clarke S., Sato M., Small C., Sullivan B., Inoue Y., Ochi D. (2014). Bycatch in Longline Fisheries for Tuna and Tuna-Like Species: A Global Review of Status and Mitigation Measures. FAO Fisher. Aquacul. Tech. Paper No 588. Food and Agriculture Organization of the United Nations, Rome
Clavareau L., Sabarros P. S., Escalle L., Bach P., Abascal F. J., Lopez J., et al. (2020). Elasmobranch Bycatch Distributions and Mortality: Insights From the European Tropical Tuna Purse-Seine Fishery. Global Ecol. Conserv. 24, e01211. doi: 10.1016/j.gecco.2020.e01211
Coelho R., Mejuto J., Domingo A., Yokawa K., Liu K.-M., Cortés E., et al. (2018). Distribution Patterns and Population Structure of the Blue Shark (Prionace Glauca) in the Atlantic and Indian Oceans. Fish Fisher. 19 (1), 90–106. doi: 10.1111/faf.12238
Collin S. P. (2018). Scene Through the Eyes of an Apex Predator: A Comparative Analysis of the Shark Visual System. Clin. Exp. Optom. 101 (5), 624–640. doi: 10.1111/cxo.12823
Compagno L. J. V. (1984). “FAO Species Catalogue,” in Sharks of the World: An Annotated and Illustrated Catalogue of Shark Species Known to Date. Parts 1 and 2 FAO Fisheries Synopsis No. 125. Food and Agriculture Organization of the United Nations, Rome.
Dagorn L., Fréon P. (1999). Tropical Tuna Associated With Floating Objects: A Simulation Study of the Meeting Point Hypothesis. Can. J. Fisher Aquat. Sci. 56 (6), 984–993. doi: 10.1139/cjfas-56-6-984
Deakos M. H., Baker J. D., Bejder L. (2011). Characteristics of a Manta Ray Manta Alfredi Population Off Maui, Hawaii, and Implications for Management. Mar. Ecol. Prog. Ser. 429, 245–260. doi: 10.3354/meps09085
Druon J.-N., Chassot E., Murua H., Lopez J. (2017). Skipjack Tuna Availability for Purse Seine Fisheries Is Driven by Suitable Feeding Habitat Dynamics in the Atlantic and Indian Oceans. Front. Mar. Sci. 4 (315). doi: 10.3389/fmars.2017.00315
Dugdale R. C. (1967). Nutrient Limitation in the Sea: Dynamics, Identification, and Significance. Limnol. Oceanog. 12 (4), 685–695. doi: 10.4319/lo.1967.12.4.0685
Dulvy N. K., Pacoureau N., Rigby C. L., Pollom R. A., Jabado R. W., Ebert D. A., et al. (2021). Overfishing Drives Over One-Third of All Sharks and Rays Toward a Global Extinction Crisis. Curr. Biol. 31 (21), 4773–4787. e4778. doi: 10.1016/j.cub.2021.08.062
Dulvy N. K., Rogers S. I., Jennings S., Stelzenmüller V., Dye S. R., Skjoldal H. R. (2008). Climate Change and Deepening of the North Sea Fish Assemblage: A Biotic Indicator of Warming Seas. J. Appl. Ecol. 45 (4), 1029–1039. doi: 10.1111/j.1365-2664.2008.01488.x
Elith J., Graham C. H., Anderson R. P., Dudík M., Ferrier S., Guisan A., et al. (2006). Novel Methods Improve Prediction of Species' Distributions From Occurrence Data. Ecography, 129—151. doi: 10.1111/j.2006.0906-7590.04596.x
Elith J., Leathwick J. (2009). Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol Evol. Syst. 40, 677–697. doi: 10.1146/annurev.ecolsys.110308.120159
Erauskin-Extramiana M., Arrizabalaga H., Hobday A. J., Cabré A., Ibaibarriaga L., Arregui I., et al. (2019). Large-Scale Distribution of Tuna Species in a Warming Ocean. Global Change Biol. 25 (6), 2043–2060. doi: 10.1111/gcb.14630
Escalle L., Capietto A., Chavance P., Dubroca L., De Molina A. D., Murua H., et al. (2015). Cetaceans and Tuna Purse Seine Fisheries in the Atlantic and Indian Oceans: Interactions But Few Mortalities. Mar. Ecol. Prog. Ser. 522, 255–268. doi: 10.3354/meps11149
Escalle L., Gaertner D., Chavance P., Delgado de Molina A., Ariz J., Mérigot B. (2016). Forecasted Consequences of Simulated FAD Moratoria in the Atlantic and Indian Oceans on Catches and Bycatches. ICES J. Mar. Sci. 74 (3), 780–792. doi: 10.1093/icesjms/fsw187
Ewell C., Hocevar J., Mitchell E., Snowden S., Jacquet J. (2020). An Evaluation of Regional Fisheries Management Organization at-Sea Compliance Monitoring and Observer Programs. Mar. Policy 115, 103842. doi: 10.1016/j.marpol.2020.103842
Fielding A. H., Bell J. F. (1997). A Review of Methods for the Assessment of Prediction Errors in Conservation Presence/Absence Models. Environ. Conserv. 24 (01), 38—49. doi: 10.1017/S0376892997000088
Filmalter J. D., Capello M., Deneubourg J.-L., Cowley P. D., Dagorn L. (2013). Looking Behind the Curtain: Quantifying Massive Shark Mortality in Fish Aggregating Devices. Front. Ecol. Environ. 11 (6), 291–296. doi: 10.1890/130045
Filmalter J., Forget F., Poisson F., Vernet A. L., Bach P., Dagorn L. (2012). Vertical and Horizontal Behavior of Silky, Oceanic White Tip and Blue Sharks in the Western Indian Ocean. IOTC-2012-WPEB08-23. Indian Ocean Tuna Commission, Mahé, 1–8
Freeman E. A., Moisen G. (2008). PresenceAbsence: An R Package for Presence Absence Analysis. J. Stat. Softw. 23 (11), 1–31. doi: 10.18637/jss.v023.i11
Frölicher T. L., Fischer E. M., Gruber N. (2018). Marine Heatwaves Under Global Warming. Nature 560 (7718), 360–364.
Garcia A., Herrera M. (2019). Assessing the Contribution of Purse Seine Fisheries to Overall Levels of Bycatch in the Indian Ocean. IOTCY2108YWPDCS14-26_Rev1. Indian Ocean Tuna Commission, Mahé, 1–95.
Gilman E., Chaloupka M., Dagorn L., Hall M., Hobday A., Musyl M., et al. (2019). Robbing Peter to Pay Paul: Replacing Unintended Cross-Taxa Conflicts With Intentional Tradeoffs by Moving From Piecemeal to Integrated Fisheries Bycatch Management. Rev. Fish Biol. Fisher 29 (1), 93–123. doi: 10.1007/s11160-019-09547-1
Gilman E., Passfield K., Nakamura K. (2014). Performance of Regional Fisheries Management Organizations: Ecosystem-Based Governance of Bycatch and Discards. Fish Fisher 15 (2), 327–351. doi: 10.1111/faf.12021
Goñi N., Ruiz J., Murua H., Santiago J., Krug I., de Olano B. S., et al. (2015). System of Verification of the Code of Good Practices on Board ANABAC and OPAGAC Tuna Purse Seiners and Preliminary Results for the Atlantic Ocean. System 5, 13.
Goujon M., Maufroy A., Relot-Stirnemann A., Moëc E., Bach P., Cauquil P., et al. (2017) Collecting Data on Board French and Italian Tropical Tuna Purse Seiners With Common Observers: Results of Orthongel's Voluntary Observer Program OCUP, (2013-2017) in the Indian Ocean. IOTC-2017-WPDCS13-22_Rev1, 22. Indian Ocean Tuna Comission, Mahé, 1–23.
Grande M., Ruiz J., Murua H., Murua J., Goñi N., Arregi I. K. I., et al. (2020). Progress on the Code of Good Practices on the Tropical Tuna Purse Seine Fishery in the Atlantic Ocean. Collect Vol Sci. Pa. ICCAT 76 (9), 193—234.
Gray C. A., Kennelly S. J. (2018). Bycatches of Endangered, Threatened and Protected Species in Marine Fisheries. Rev. Fish Biol. Fisher 28 (3), 521–541. doi: 10.1007/s11160-018-9520-7
Griffiths S. P., Kesner-Reyes K., Garilao C., Duffy L. M., Román M. H. (2019). Ecological Assessment of the Sustainable Impacts of Fisheries (EASI-Fish): A Flexible Vulnerability Assessment Approach to Quantify the Vumulative Impacts of Fishing in Data-limited SettingsMar. Ecol. Prog. Series. 625, 89–113.
Guisan A., Edwards T. C., Hastie T. (2002). Generalized Linear and Generalized Additive Models in Studies of Species Distributions: Setting the Scene. Ecol. Model 157 (2), 89–100. doi: 10.1016/S0304-3800(02)00204-1
Hacohen-Domené A., Martínez-Rincón R. O., Galván-Magaña F., Cárdenas-Palomo N., de la Parra-Venegas R., Galván-Pastoriza B., et al. (2015). Habitat Suitability and Environmental Factors Affecting Whale Shark (Rhincodon Typus) Aggregations in the Mexican Caribbean. Environ. Biol. Fish 98 (8), 1953–1964. doi: 10.1007/s10641-015-0413-5
Hahlbeck N., Scales K. L., Dewar H., Maxwell S. M., Bograd S. J., Hazen E. L. (2017). Oceanographic Determinants of Ocean Sunfish (Mola Mola) and Bluefin Tuna (Thunnus Orientalis) Bycatch Patterns in the California Large Mesh Drift Gillnet Fishery. Fisher Res. 191, 154–163. doi: 10.1016/j.fishres.2017.03.011
Hall M., Gilman E., Minami H., Mituhasi T., Carruthers E. (2017). Mitigating Bycatch in Tuna Fisheries. Rev. Fish Biol. Fisher 27, 1–28. doi: 10.1007/s11160-017-9478-x
Hall M., Roman M. (2013). Bycatch and Non-Tuna Catch in the Tropical Tuna Purse Seine Fisheries of the World. FAO Fisher Aquacul. Tech. Paper 568). Food and Agriculture Organization of the United Nations, Rome
Hammerschlag N., McDonnell L. H., Rider M. J., Street G. M., Hazen E. L., Natanson L. J., et al. (2022). Ocean Warming Alters The Distributional Range, Migratory Timing, and Spatial Protections of an Apex Predator, the Tiger Shark (Galeocerdo Cuvier). Global Change Biol. doi: 10.1111/gcb.16045
Hazen E. L., Scales K. L., Maxwell S. M., Briscoe D. K., Welch H., Bograd S. J., et al. (2018). A Dynamic Ocean Management Tool to Reduce Bycatch and Support Sustainable Fisheries. Sci. Adv. 4 (5), eaar3001. doi: 10.1126/sciadv.aar3001
Hermes J. C., Reason C. J. C. (2008). Annual Cycle of the South Indian Ocean (Seychelles-Chagos) Thermocline Ridge in a Regional Ocean Model. J. Geophys. Res.: Ocean 113 (C4), 148–227. doi: 10.1029/2007JC004363
Hijmans R. J., Phillips S., Leathwick J., Elith J. (2020). Dismo: Species Distribution Modeling. R Package Version 1. 3–3 .
Hilborn R., Agostini V. N., Chaloupka M., Garcia S. M., Gerber L. R., Gilman E., et al. (2022). Area-Based Management of Blue Water Fisheries: Current Knowledge and Research Needs. Fish Fisher. 23 (2), 492–518. doi: 10.1111/faf.12629
Hitchcock G. L., Key E. L., Masters J. (2000). The Fate of Upwelled Waters in the Great Whirl, August 1995. Deep Sea Res. Part II: Top Stud. Oceanog 47 (7), 1605–1621. doi: 10.1016/S0967-0645(99)00156-3
Hobday A. J., Hartog J. R., Timmiss T., Fielding J. (2010). Dynamic Spatial Zoning to Manage Southern Bluefin Tuna (Thunnus Maccoyii) Capture in a Multi-Species Longline Fishery. Fisher Oceanog 19 (3), 243–253. doi: 10.1111/j.1365-2419.2010.00540.x
Holbrook N., Scannell H., Gupta A., Benthuysen J., Feng M., Oliver E., et al. (2019). A Global Assessment of Marine Heatwaves and Their Drivers. Nat. Commun. 10, 2624. doi: 10.1038/s41467-019-10206-z
Howey-Jordan L. A., Brooks E. J., Abercrombie D. L., Jordan L. K. B., Brooks A., Williams S., et al. (2013). Complex Movements, Philopatry and Expanded Depth Range of a Severely Threatened Pelagic Shark, the Oceanic Whitetip (Carcharhinus Longimanus) in the Western North Atlantic. PloS One 8 (2), e56588. doi: 10.1371/journal.pone.0056588
Howey L. A., Tolentino E. R., Papastamatiou Y. P., Brooks E. J., Abercrombie D. L., Watanabe Y. Y., et al. (2016). Into the Deep: The Functionality of Mesopelagic Excursions by an Oceanic Apex Predator. Ecol. Evol. 6 (15), 5290–5304. doi: 10.1002/ece3.2260
Hutchinson G. (1957). Concluding Remarks Cold Spring Harbor Symposia on Quantitative Biology. GS Search 22, 415–427.
IATTC. (1999). Agreement on the International Dolphin Conservation Program. Inter-American Tropical Tuna Commission, La Jolla
IATTC. (2009). Resolution C-09-01 on a Multiannual Program for the Conservation of Tuna in the Eastern Pacific Ocean in 2009-2011. Inter-American Tropical Tuna Commission, La Jolla
IATTC. (2011). Resolution C-11-10 on the Conservation of Oceanic Whitetip Sharks Caught in Association With Fisheries in the Antigua Convention Area. Inter-American Tropical Tuna Commission, La Jolla
ICCAT. (2010). Recommendation 10-07 by ICCAT on the Conservation of Oceanic Whitetip Shark Caught in Association With Fisheries in the ICCAT Convention Area. International Commission for the Conservation of Atlantic Tunas, Madrid
IOTC. (2013). Resolution 13/06 on a Scientific and Management Framework on the Conservation of Sharks Species Caught in Association With IOTC Managed Fisheries. Indian Ocean Tuna Commission, Mahé
IOTC. (2015). Status of the Indian Ocean Oceanic Whitetip Shark (OCS: Carcharhinus Longimanus). Indian Ocean Tuna Commission, Mahé
IOTC. (2021). Review of Data on Drifting Fish Aggregating Devices. Indian Ocean Tuna Commission, Mahé
Jacoby D. M. P., Croft D. P., Sims D. W. (2012). Social Behaviour in Sharks and Rays: Analysis, Patterns and Implications for Conservation. Fish Fisher 13 (4), 399–417. doi: 10.1111/j.1467-2979.2011.00436.x
Jarnevich C. S., Stohlgren T. J., Kumar S., Morisette J. T., Holcombe T. R. (2015). Caveats for Correlative Species Distribution Modeling. Ecol. Inf. 29, 6–15. doi: 10.1016/j.ecoinf.2015.06.007
Jiménez-Valverde A., Lobo J. M. (2007). Threshold Criteria for Conversion of Probability of Species Presence to Either–or Presence–Absence. Acta Oecologica 31 (3), 361–369. doi: 10.1016/j.actao.2007.02.001
Juan-Jordá M. J., Murua H., Arrizabalaga H., Dulvy N. K., Restrepo V. (2018). Report Card on Ecosystem-Based Fisheries Management in Tuna Regional Fisheries Management Organizations. Fish Fisher 19 (2), 321–339. doi: 10.1111/faf.12256
Kai E. T., Rossi V., Sudre J., Weimerskirch H., Lopez C., Hernandez-Garcia E., et al. (2009). Top Marine Predators Track Lagrangian Coherent Structures. Proc. Natl. Acad. Sci. 106 (20), 8245–8250.
Kai M., Thorson J. T., Piner K. R., Maunder M. N. (2017). Predicting the Spatio-Temporal Distributions of Pelagic Sharks in the Western and Central North Pacific. Fisher Oceanog 26 (5), 569–582. doi: 10.1111/fog.12217
Kaplan D. M., Chassot E., Amandé J. M., Dueri S., Demarcq H., Dagorn L., et al. (2014). Spatial Management of Indian Ocean Tropical Tuna Fisheries: Potential and Perspectives. ICES J. Mar Sci. 71 (7), 1728–1749. doi: 10.1093/icesjms/fst233
Lee P.-F., Chen I., Tzeng W. (2005). Spatial and Temporal Distribution Patterns of Bigeye Tuna (Thunnus Obesus) in the Indian Ocean. Zoological Stud. 44, 260–270.
Lezama-Ochoa N., Hall M. A., Pennino M. G., Stewart J. D., López J., Murua H. (2019). Environmental Characteristics Associated With the Presence of the Spinetail Devil Ray (Mobula Mobular) in the Eastern Tropical Pacific. PloS One 14 (8), e0220854. doi: 10.1371/journal.pone.0220854
Lezama-Ochoa N., Lopez J., Hall M., Bach P., Abascal F., Murua H. (2020). Spatio-Temporal Distribution of the Spinetail Devil Ray Mobula Mobular in the Eastern Tropical Atlantic Ocean. Endanger Species Res. 43, 447–460. doi: 10.3354/esr01082
Lezama-Ochoa N., Murua H., Chust G., Van Loon E., Ruiz J., Hall M., et al. (2016). Present and Future Potential Habitat Distribution of Carcharhinus Falciformis and Canthidermis Maculata By-Catch Species in the Tropical Tuna Purse-Seine Fishery Under Climate Change. Front. Mar Sci. 3 (34). doi: 10.3389/fmars.2016.00034
Lima S. L., Dill L. M. (1990). Behavioral Decisions Made Under the Risk of Predation: A Review and Prospectus. Can. J. Zoology 68 (4), 619–640. doi: 10.1139/z90-092
Liu C., Berry P. M., Dawson T. P., Pearson R. G. (2005). Selecting Thresholds of Occurrence in the Prediction of Species Distributions. Ecography 28 (3), 385–393. doi: 10.1111/j.0906-7590.2005.03957.x
Lonhart S. I., Jeppesen R., Beas-Luna R., Crooks J. A., Lorda J. (2019). Shifts in the Distribution and Abundance of Coastal Marine Species Along the Eastern Pacific Ocean During Marine Heatwaves From 2013 to 2018. Mar Biodiver Record 12 (1), 1–15. doi: 10.1186/s41200-019-0171-8
Lopez J., Alvarez-Berastegui D., Soto M., Murua H. (2020). Using Fisheries Data to Model the Oceanic Habitats of Juvenile Silky Shark (Carcharhinus Falciformis) in the Tropical Eastern Atlantic Ocean. Biodiver Conserv. 29 (7), 2377–2397. doi: 10.1007/s10531-020-01979-7
Lopez J., Moreno G., Lennert-Cody C., Maunder M., Sancristobal I., Caballero A., et al. (2017). Environmental Preferences of Tuna and non-Tuna Species Associated With Drifting Fish Aggregating Devices (DFADs) in the Atlantic Ocean, Ascertained Through Fishers’ Echo-Sounder Buoys. Deep Sea Res. Part II Top. Stud. Oceanogr. 140, 127–138. doi: 10.1016/j.dsr2.2017.02.007
Marín-Enríquez E., Seoane J., Muhlia-Melo A. (2018). Environmental Modeling of Occurrence of Dolphinfish (Coryphaena Spp.) in the Pacific Ocean Off Mexico Reveals Seasonality in Abundance, Hot Spots and Migration Patterns. Fisher Oceanog 27 (1), 28–40. doi: 10.1111/fog.12231
Marsac F. (2017). The Seychelles Tuna Fishery and Climate Change. Climate Change Impacts on Fisheries and Aquaculture: A global Analysis 2, 523–568.
Marsac F., Fonteneau A., Ménard F. (2000). Drifting FADs Used in Tuna Fisheries: an Ecological Trap? Proceedings of the 1st Symposium on Tuna fisheries and FADs, Martinique October 1999: 537–552.
Marshall C. E., Glegg G. A., Howell K. L. (2014). Species Distribution Modelling to Support Marine Conservation Planning: The Next Steps. Mar Policy 45, 330–332. doi: 10.1016/j.marpol.2013.09.003
Martínez-Rincón R. O., Ortega-García S., Vaca-Rodríguez J. G. (2012). Comparative Performance of Generalized Additive Models and Boosted Regression Trees for Statistical Modeling of Incidental Catch of Wahoo (Acanthocybium Solandri) in the Mexican Tuna Purse-Seine Fishery. Ecol. Model 233, 20–25. doi: 10.1016/j.ecolmodel.2012.03.006
Martinez-Rincon R. O., Ortega-Garcia S., Vaca-Rodriguez J. G., Griffiths S. P. (2015). Development of Habitat Prediction Models to Reduce by-Catch of Sailfish (Istiophorus Platypterus) Within the Purse-Seine Fishery in the Eastern Pacific Ocean. Mar Freshwater Res. 66 (7), 644–653. doi: 10.1071/MF14062
Maury O., Miller K., Campling L., Arrizabalaga H., Aumont O., Bodin Ö., et al. (2013). A Global Science–Policy Partnership for Progress Toward Sustainability of Oceanic Ecosystems and Fisheries. Curr. Opin. Environ. Sustain 5 (3), 314–319. doi: 10.1016/j.cosust.2013.05.008
McKinney J. A., Hoffmayer E. R., Wu W., Fulford R., Hendon J. (2012). Feeding Habitat of the Whale Shark Rhincodon Typus in the Northern Gulf of Mexico Determined Using Species Distribution Modelling. Mar Ecol. Prog. Ser. 458, 199–211. doi: 10.3354/meps09777
Mejuto J., García-Cortés B., Ramos-Cartelle A. (2005). Tagging-Recapture Activities of Large Pelagic Sharks Carried Out by Spain or in Collaboration With the Tagging Programs of Other Countries. Coll. Vol Sci. Pap. ICCAT 58, 974–1000.
Montero J. T., Martinez-Rincon R. O., Heppell S. S., Hall M., Ewal M. (2016). Characterizing Environmental and Spatial Variables Associated With the Incidental Catch of Olive Ridley (Lepidochelys Olivacea) in the Eastern Tropical Pacific Purse-Seine Fishery. Fisher. Oceanog. 25 (1), 1–14. doi: 10.1111/fog.12130
Murua H., Dagorn L., Justel-Rubio A., Moreno G., Restrepo V. (2021a). Questions and Answers About FADs and Bycatch. ISSF Technical Report 2021-11. International Seafood Sustainability Foundation, Washington, D.C.
Murua H., Griffiths S. P., Hobday A. J., Clarke S. C., Cortés E., Gilman E. L., et al. (2021b). Shark Mortality Cannot be Assessed by Fishery Overlap Alone. Nature 595 (7866), E4–E7. doi: 10.1038/s41586-021-03396-4
Musyl M. K., Brill R. W., Curran D. S., Fragoso N. M., McNaughton L. M., Nielsen A., et al. (2011). Postrelease Survival, Vertical and Horizontal Movements, and Thermal Habitats of Five Species of Pelagic Sharks in the Central Pacific Ocean. Fisher Bull. 109 (4), 341–368.
Naimi B., Hamm N., Groen T., Skidmore A., Toxopeus A. (2014). Where is Positional Uncertainty a Problem for Species Distribution Modelling. Ecography 37, 191–203. doi: 10.1111/j.1600-0587.2013.00205.x
Oliver E. C., Burrows M. T., Donat M. G., Sen Gupta A., Alexander L. V., Perkins-Kirkpatrick S. E., et al. (2019). Projected Marine Heatwaves in the 21st Century and the Potential for Ecological Impact. Front. Mar Sci. 6, 734. doi: 10.3389/fmars.2019.00734
Orue B., Lopez J., Moreno G., Santiago J., Soto M., Murua H. (2019). Aggregation Process of Drifting Fish Aggregating Devices (DFADs) in the Western Indian Ocean: Who Arrives First, Tuna or non-Tuna Species? PloS One 14 (1), e0210435. doi: 10.1371/journal.pone.0210435
Pacoureau N., Rigby C. L., Kyne P. M., Sherley R. B., Winker H., Carlson J. K., et al. (2021). Half a Century of Global Decline in Oceanic Sharks and Rays. Nature 589 (7843), 567–571. doi: 10.1038/s41586-020-03173-9
Passadore C., Möller L. M., Diaz-Aguirre F., Parra G. J. (2018). Modelling Dolphin Distribution to Inform Future Spatial Conservation Decisions in a Marine Protected Area. Sci. Rep. 8 (1), 1–14. doi: 10.1038/s41598-018-34095-2
Pearson R. G. (2010) Species Distribution Modeling for Conservation Educators and Practitioners. Lesson Conserv. 3, 54–89
Pearson R. G., Thuiller W., Araújo M. B., Martinez-Meyer E., Brotons L., McClean C., et al. (2006). Model-Based Uncertainty in Species Range Prediction. J. Biogeograph 33 (10), 1704–1711. doi: 10.1111/j.1365-2699.2006.01460.x
Pennington J. T., Mahoney K. L., Kuwahara V. S., Kolber D. D., Calienes R., Chavez F. P. (2006). Primary Production in the Eastern Tropical Pacific: A Review. Prog. Oceanog 69 (2), 285–317. doi: 10.1016/j.pocean.2006.03.012
Pennino M. G., Conesa D., López-Quílez A., Muñoz F., Fernández A., Bellido J. M. (2016). Fishery-Dependent and -Independent Data Lead to Consistent Estimations of Essential Habitats. ICES J. Mar Sci. 73 (9), 2302–2310. doi: 10.1093/icesjms/fsw062
Poisson F., Séret B., Vernet A.-L., Goujon M., Dagorn L. (2014). Collaborative Research: Development of a Manual on Elasmobranch Handling and Release Best Practices in Tropical Tuna Purse-Seine Fisheries. Mar Policy 44, 312—320. doi: 10.1016/j.marpol.2013.09.025
Pons M., Watson J. T., Ovando D., Andraka S., Brodie S., Domingo A., et al. (2022). Trade-Offs Between Bycatch and Target Catches in Static Versus Dynamic Fishery Closures. Proc. Natl. Acad. Sci. 119 (4). doi: 10.1073/pnas.2114508119
Priyadarshi A., Smith S. L., Mandal S., Tanaka M., Yamazaki H. (2019). Micro-Scale Patchiness Enhances Trophic Transfer Efficiency and Potential Plankton Biodiversity. Sci. Rep. 9 (1), 1–9. doi: 10.1038/s41598-019-53592-6
Rigby C., Barreto R., Carlson J., Fernando D., Fordham S., Francis M., et al. (2019). Carcharhinus Longimanus. The IUCN Red List of Threatened Species 2019-3. doi: 10.2305/IUCN.UK.2019-3.RLTS.T39374A2911619.en
Rooker J. R., Simms J. R., Wells R. J. D., Holt S. A., Holt G. J., Graves J. E., et al. (2012). Distribution and Habitat Associations of Billfish and Swordfish Larvae Across Mesoscale Features in the Gulf of Mexico. PloS One 7 (4), e34180. doi: 10.1371/journal.pone.0034180
Ruiz J., Abascal F. J., Bach P., Baez J. C., Cauquil P., Grande M., et al. (2018). Bycatch of the European, and Associated Flag, Purse-Seine Tuna Fishery in the Indian Ocean for the Period 2008-2017. IOTC-2018-WPEB14-15. Indian Ocean Tuna, Mahé, 1–15.
Saranya J., Roxy M. K., Dasgupta P., Anand A. (2021). Genesis and Trends in Marine Heatwaves Over the Tropical Indian Ocean and Their Interaction With the Indian Summer Monsoon. J. Geophys. Res. Oceans 127, doi: 10.1029/2021JC017427
Schott F. A., McCreary J. P. (2001). The Monsoon Circulation of the Indian Ocean. Prog. Oceanog 51 (1), 1–123. doi: 10.1016/S0079-6611(01)00083-0
Schott F. A., Xie S.-P., McCreary J. J. P. (2009). Indian Ocean Circulation and Climate Variability. Rev. Geophys 47 (1), 8755–1209. doi: 10.1029/2007RG000245
Sequeira A. M. M., Mellin C., Fordham D. A., Meekan M. G., Bradshaw C. J. A. (2014). Predicting Current and Future Global Distributions of Whale Sharks. Global Change Biol. 20 (3), 778–789. doi: 10.1111/gcb.12343
Setiawati M., Sambah A., Miura F., Tanaka T., As-syakur A. R. (2015). Characterization of Bigeye Tuna Habitat in the Southern Waters Off Java–Bali Using Remote Sensing Data. Adv. Space Res. 55, 732–746. doi: 10.1016/j.asr.2014.10.007
Shi W., Wang M. (2021). A Biological Indian Ocean Dipole Event in 2019. Sci. Rep. 11 (1), 2452. doi: 10.1038/s41598-021-81410-5
Su N.-J., Sun C.-L., Punt A. E., Yeh S.-Z., DiNardo G. (2011). Modelling the Impacts of Environmental Variation on the Distribution of Blue Marlin, Makaira Nigricans, in the Pacific Ocean. ICES J. Mar Sci. 68 (6), 1072–1080. doi: 10.1093/icesjms/fsr028
Tolotti M. T., Bach P., Hazin F., Travassos P., Dagorn L. (2015a). Vulnerability of the Oceanic Whitetip Shark to Pelagic Longline Fisheries. PloS One 10 (10), e0141396. doi: 10.1371/journal.pone.0141396
Tolotti M. T., Bauer R., Forget F., Bach P., Dagorn L., Travassos P. (2017). Fine-Scale Vertical Movements of Oceanic Whitetip Sharks (Carcharhinus Longimanus). Fisher Bull. 115 (3), 380–395. doi: 10.7755/fb.115.3.8
Tolotti M. T., Filmalter J. D., Bach P., Travassos P., Seret B., Dagorn L. (2015b). Banning is Not Enough: The Complexities of Oceanic Shark Management by Tuna Regional Fisheries Management Organizations. Global Ecol. Conserv. 4, 1–7. doi: 10.1016/j.gecco.2015.05.003
Tremblay-Boyer L., Carvalho F., Neubauer P., Pilling G. (2019). Stock assessment for oceanic whitetip shark in the Western and Central Pacific Ocean. WCPFC Scientific Committee 15th Regular Session. WCPFC-SC15-2019/SA-WP-06. Pohnpei, p. 99
Urbisci L. C., Stohs S. M., Piner K. R. (2016). From Sunrise to Sunset in the California Drift Gillnet Fishery: An Examination of the Effects of Time and Area Closures on the Catch and Catch Rates of Pelagic Species. Mar. Fish Rev. 78 (3-4), 1–11.
Venables W. N., Dichmont C. M. (2004). GLMs, GAMs and GLMMs: An Overview of Theory for Applications in Fisheries Research. Fisher Res. 70 (2), 319–337. doi: 10.1016/j.fishres.2004.08.011
Watson J. T., Essington T. E., LENNERT-CODY C. E., Hall M. A. (2009). Trade-Offs in the Design of Fishery Closures: Management of Silky Shark Bycatch in the Eastern Pacific Ocean Tuna Fishery. Conserv. Biol. 23 (3), 626–635. doi: 10.1111/j.1523-1739.2008.01121.x
WCPFC (2019). Conservation and Management Measure for Sharks, Vol. 2019-04. Western and Central Pacific Fisheries Commission, Kolonia
Welch H., Brodie S., Jacox M. G., Bograd S. J., Hazen E. L. (2020). Decision-Support Tools for Dynamic Management. Conserv. Biol. 34 (3), 589–599. doi: 10.1111/cobi.13417
Wiggert J. D., Murtugudde R. G., Christian J. R. (2006). Annual Ecosystem Variability in the Tropical Indian Ocean: Results of a Coupled Bio-Physical Ocean General Circulation Model. Deep Sea Res. Part II: Top Stud. Oceanog 53 (5), 644–676. doi: 10.1016/j.dsr2.2006.01.027
Williamson M. J., Tebbs E. J., Dawson T. P., Jacoby D. M. P. (2019). Satellite Remote Sensing in Shark and Ray Ecology, Conservation and Management. Front. Mar Sci. 6 (135). doi: 10.3389/fmars.2019.00135
Wood S. (2006). Generalized Additive Models: An Introduction With R. Chapman Hall/CRC, Boca Raton. doi: 10.1201/9781420010404
Yang K., Cai W., Huang G., Wang G., Ng B., Li S. (2020). Oceanic Processes in Ocean Temperature Products Key to a Realistic Presentation of Positive Indian Ocean Dipole Nonlinearity. Geophys Res. Lett. 47 (16), e2020GL089396. doi: 10.1029/2020GL089396
Young C., Carlson J. (2020). The Biology and Conservation Status of the Oceanic Whitetip Shark (Carcharhinus Longimanus) and Future Directions for Recovery. Rev. Fish Biol. Fisher 30, 293–312. doi: 10.1007/s11160-020-09601-3
Young J., Hunt B., Cook T., Llopiz J., Hazen E., Pethybridge H., et al. (2015). The Trophodynamics of Marine Top Predators: Current Knowledge, Recent Advances and Challenges. Deep Sea Res. Part II Top Stud. Oceanog. 113, 170–187. doi: 10.1016/j.dsr2.2014.05.015
Zhang L., Du Y., Cai W. (2018). Low-Frequency Variability and the Unusual Indian Ocean Dipole Events in 2015 and 2016. Geophys Res. Lett. 45 (2), 1040–1048. doi: 10.1002/2017GL076003
Keywords: oceanic whitetip shark, species distribution model, bycatch species, tropical tuna purse-seine fishery, Western Indian Ocean
Citation: Lopetegui-Eguren L, Poos JJ, Arrizabalaga H, Guirhem GL, Murua H, Lezama-Ochoa N, Griffiths SP, Gondra JR, Sabarros PS, Báez JC and Juan-Jordá MJ (2022) Spatio-Temporal Distribution of Juvenile Oceanic Whitetip Shark Incidental Catch in the Western Indian Ocean. Front. Mar. Sci. 9:863602. doi: 10.3389/fmars.2022.863602
Received: 27 January 2022; Accepted: 28 March 2022;
Published: 01 June 2022.
Edited by:
David M. P. Jacoby, University of Lancaster, United KingdomReviewed by:
Gonzalo Mucientes Sandoval, Institute of Marine Research (CSIC), SpainCopyright © 2022 Lopetegui-Eguren, Poos, Arrizabalaga, Guirhem, Murua, Lezama-Ochoa, Griffiths, Gondra, Sabarros, Báez and Juan-Jordá. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Leire Lopetegui-Eguren, bGxvcGV0ZWd1aUBhenRpLmVz
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.