 Luisa Marques
Luisa Marques Ricardo Calado
Ricardo Calado Ana Isabel Lillebø
Ana Isabel Lillebø- ECOMARE, CESAM–Centre for Environmental and Marine Studies, Department of Biology, University of Aveiro, Aveiro, Portugal
Ascidians are considered as filter-feeder biofouling pests that negatively affect aquaculture facilities. However, they can also be recognized as a potential co-cultured/extractive species for integrated multi-trophic aquaculture (IMTA) with potential added value as bioresources. A systematic review aiming to understand the ecological importance of ascidians as efficient filter-feeders [What?]; their potential contribution as extractive species [How?]; and to set the benchmark for their nutritional value and potential added value to the aquaculture industry [For what?] is a timely contribution to advance the state of the art on these largely overlooked bioresources. In the last two decades, there has been an overall increase in publications addressing ascidians in aquaculture, namely, their negative impacts through biofouling, as well as their role in IMTA, environmental status, and microbiology. While Ciona intestinalis, a solitary ascidian, has been the most studied species, overall, most ascidians present high filtration and fast-growth rates. As ascidians perform well under IMTA, competition for resources and space with other filter-feeders might occur, which may require additional management actions to optimize production. Studies addressing their bioactive products show that ascidians hold great potential as premium ingredients for aquafeed formulations, as well as dietary supplements (e.g., amino acids, fatty acids). Further research on the potential use of ascidians in IMTA frameworks should focus on systems carrying capacity.
Introduction
Aquaculture is an important source of food, nutrition, income, and livelihoods for hundreds of millions of people worldwide (FAO, 2020). With the continuous increase of the world’s population, aquaculture production needs to increase by 21–44 million tons by the year 2050 (Costello et al., 2020). An idealistic scenario, and a major challenge for the aquaculture industry, is to be profitable, product-diversified, socially beneficial, and yet ecologically efficient and environmentally friendly, i.e., to cope with the principles of sustainable development. Integrated multi-trophic aquaculture (IMTA) has the potential to achieve such a goal.
An IMTA framework is a nature-based solution in which the by-products, wastes, uneaten feed, and nutrients from one species are recycled and converted to become fertilizer, feed, and energy for the growth of another (Naylor and Burke, 2005). These systems can be land-based or open-water, use marine, brackish or freshwater, and may include several different combinations of co-cultured species (Neori et al., 2004). IMTA aims at mimicking a natural ecosystem by combining and incorporating complementary species from different trophic or nutritional levels in the same productive environment. In an operational IMTA system, extractive species uptake organic and inorganic matter contributing to reduce costs and comply with environmental regulations (Reid et al., 2020).
In addition, their potential market value (e.g., food, feed, pharma) might provide extra economic benefits to farmers (Barrington et al., 2009; Béné et al., 2015). Selected species should be cultured at densities that optimize nutrient uptake, promote a stable balance between biological and chemical processes improving the ecosystem’s health, and should be economically important as aquaculture products (Alexander et al., 2016).
However, implementing a healthful and balanced concept can present multiple challenges to farmers. Nonetheless, an IMTA framework also presents numerous benefits, including the decrease in waste outputs from overall farming activities, the additional production of a marketable product for little or no additional input cost, and more importantly, environmentally sustainable farming operations (Barrington et al., 2009; Troell et al., 2009).
The open key question concerns the optimization of the uptake of particulate and dissolved organic matter from uneaten/undigested feed and feces. Organic nutrients can nitrify the benthic-pelagic community (Albert et al., 2021) and the excess of inorganic nutrients (ammonia, nitrate, nitrite) may ultimately create a bio-deposit and lead to eutrophicated waters (Chopin et al., 2001) and/or represent an economic burden to fish farmers (Fry et al., 2016).
Ascidians, commonly known as sea squirts or tunicates, are found in all marine habitats from shallow water to the deep sea (Shenkar and Swalla, 2011) and there are approximately 3000 described species (Shenkar and Swalla, 2011). Currently classed under Phylum Chordata, ascidians hold a unique evolutionary position as the sister group of vertebrates. These organisms are benthic suspension feeders that filter particulate organic matter from the water column via an oral siphon and expelled filtered water through the atrial siphon (Jørgensen and Goldberg, 1953; Jørgensen, 1954). They present a wide variety of forms (from small colonies to big solitary forms), colors, shapes (from cone-shaped, elongated, globular, or oval), and sizes (generally from 0.5 to 200 mm) (Petersen, 2007; Shenkar and Swalla, 2011).
The body is always covered with a tunic, a protective layer that may be translucid, brightly colored or dull, covered by various kinds of spines, and contain calcareous spicules (Lambert and Lambert, 1987), or even be covered by a dense layer of sand grains (Young, 1989). Most solitary ascidians are hermaphrodites and reproduce by external fertilization (Honegger, 1986). They develop a free-swimming tadpole-like larva that swims during a short period, settles on a wide variety of habitats, and finally matures into a sessile adult (Shenkar and Swalla, 2011). Colonial specimens can reproduce both sexually and asexually (Gasparini et al., 2015). Ascidians often present an invasive behavior, representing the most dominant fouling species worldwide, colonizing natural and artificial substrates (Ordóñez et al., 2013).
While several ascidian species present a preference to settle on natural substrates (Hirose and Sensui, 2021), others settle on artificial structures, such as ship hulls, floating docks (Zvyagintsev et al., 2007), and aquaculture infrastructures (Hodson et al., 2000; Khalaman, 2001; Bullard et al., 2013; Rosa et al., 2013), process known as biofouling. At times some species even grow on other organisms being farmed, such as on the shells of mollusks (Dijkstra and Nolan, 2017; Casso et al., 2018).
Hereupon, these organisms hold great potential as co-cultured/extractive species in IMTA frameworks, with potential to contribute to more efficient, profitable, and sustainable aquaculture systems. Benthic fish contribute to sediment resuspension while searching for food or shelter (Yahel et al., 2008; Carvajalino-Fernández et al., 2020). Although these resuspension events can be brief and localized (Yahel et al., 2002), in an IMTA scenario ascidians, as excellent filter-feeders, can rapidly uptake nutrient recycling and contribute to a positive outcome.
The main objective of this systematic review is to understand how ascidians may no longer be regarded as pest organisms, who’s biofouling negatively impacts aquaculture ventures, but rather as important extractive species in IMTA frameworks that yield premium biomass for high-end uses. To this purpose, we surveyed the scientific literature to answer the following three questions: [What do we know?] To better understand the biological and ecological importance of ascidians as filter-feeders in an IMTA framework; [How do ascidians perform in IMTA?] to evaluate which combination of species will contribute the most to enhance the performance of ascidians in IMTA frameworks; and [For what kind of bioactive products?] to recognize ascidians as potential bioresources in different high-end fields, namely, blue biotechnology and human nutrition.
Here, special attention will be given to fatty acids, as both omega-3 (n-3) and omega-6 (n-6) fatty acids are essential components for food, feed, and pharma industries. The analysis of these three questions will enable us to discuss and conclude on the potential of ascidians as extractive species and their added value in marine IMTA frameworks.
Literature Review
In January 2020, a systematic literature review, with no year restriction, was performed using the databases Thomson Reuters Web of Science (Core Collection) (Topic) and Scopus (Article title, Abstract, Keywords). The strategy used was to search within a combination of specific terms: Filtration AND (tunicate OR ascidian OR “sea squirt*”); Aquaculture AND (tunicate OR ascidian OR “sea squirt*”); Fatty acid* AND (tunicate OR ascidian OR “sea squirt*”) to achieve the review’s goal.
A schematic representation of the selection process is summarized in Figure 1. Overall, a total of 566 publications were retrieved (after excluding duplicates from the two databases), and a spreadsheet with the bibliographic information of each reference was created for further analysis to ascertain their relevance for this study. The review selection consists of two sections.
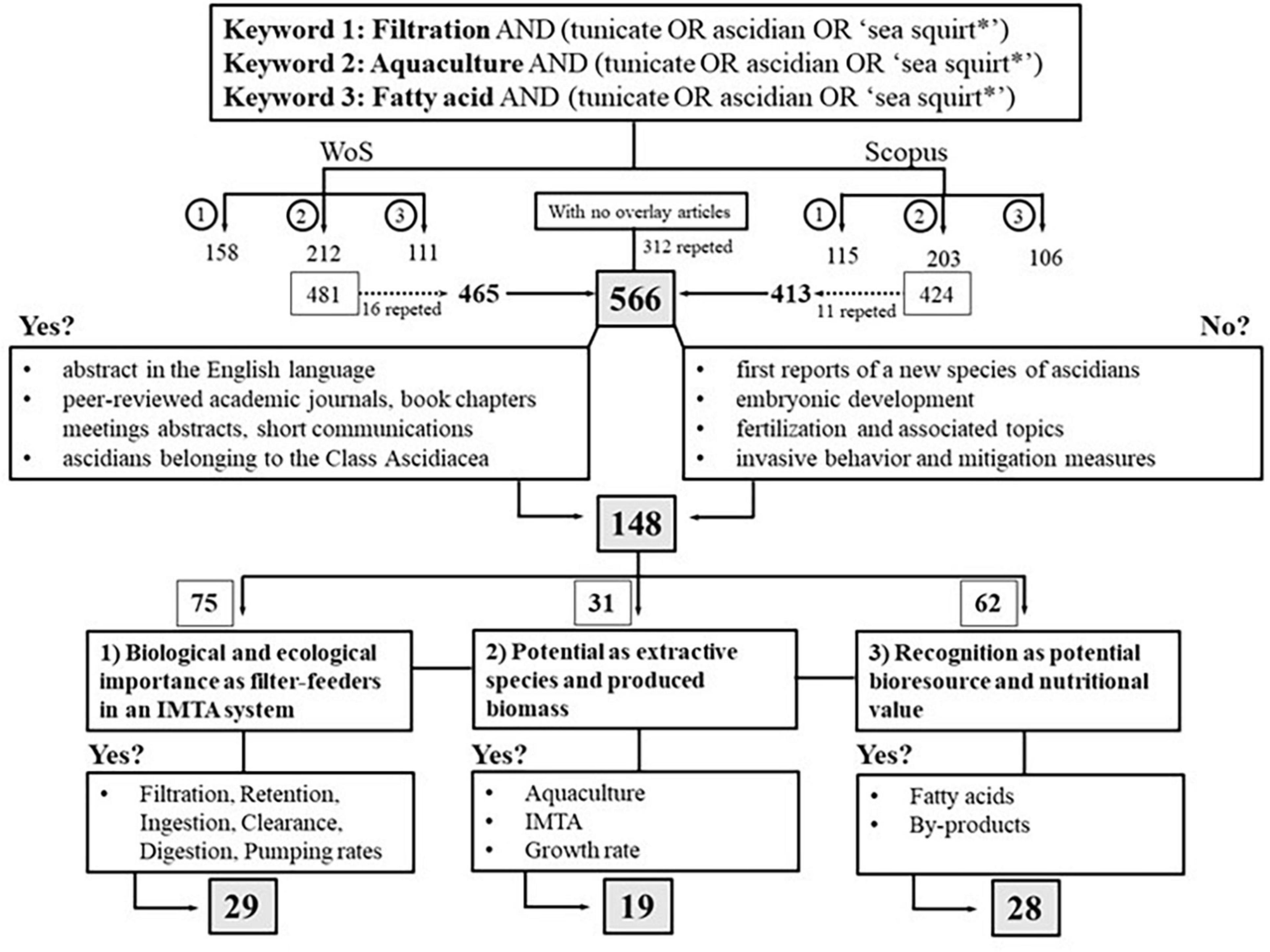
Figure 1. Schematic representation of the process employed for the selection of relevant publications retrieved from each database (Web of Science, WoS; Scopus).
The first article selection aimed to retain publications with, at least, the abstract in the English language, peer-reviewed academic journals, book chapters, meetings abstracts, short communications, and reports on ascidians within Class Ascidiacea. When studies addressed ascidians in a general way, these were registered as “Ascidiacea.” Furthermore, studies addressing the following topics were excluded from the present review: (i) first reports on the occurrence of a new species of ascidian in a given location and their geographic distributions, (ii) embryonic development, (iii) fertilization, reproduction, and associated topics, (iv) invasive behavior of ascidians and mitigation measures.
A total of 148 publications (Supplementary Table 1) were considered relevant and selected for further analysis. Ten research categories (aquaculture, biochemistry, biofouling, biology, biotechnology/methods, diseases, environmental, IMTA, microbiology, and review) were created and assigned to each of the 148 publications, with a maximum of four categories being attributed per publication.
The rationale for this procedure is detailed in Table 1. Additionally, each publication was also assigned to one of the three questions (occasionally two) initially established: (question 1 [What?]: 75 publications, question 2 [How?]: 31 publications, and question 3 [For what?]: 62 publications). Subsequently, each of the publications assigned to each of the three questions was further screened as detailed in Figure 1.

Table 1. Research categories considered and their respective criteria.
Briefly, concerning question 1, only publications addressing filtration, retention, ingestion, clearance, digestion, and water pumping rates were selected, for a total of 29 publications. Regarding question 2, only publications addressing topics such as aquaculture, IMTA, and growth rates were included, for a total of 19 publications. Finally, for question 3, publications referring to fatty acids and other potential co-products were considered, for a total of 28 publications. Blue biotechnology may focus on a plethora of potentially bioactive compounds (Vieira et al., 2020).
As this review targets marine species and they can be seen as sources of the essential omega-3 (n-3) and omega-6 (n-6) fatty acids, special attention will be given to these bioactive compounds as they can represent an added value for food, feed, and pharma industries.
Results
Out of the 148 publications, 80 fell into the research category “Biology,” hence demonstrating the importance of understanding morphology, biology, and anatomy of ascidians in a general manner (Figure 2). Since the 21st century, an overall increase in all research categories is noted, but it is worth mentioning a gradual and joint increase of publications in “Aquaculture,” “Biofouling,” and “IMTA” categories (44, 25, and 7%, respectively) as these are correlated with each other.
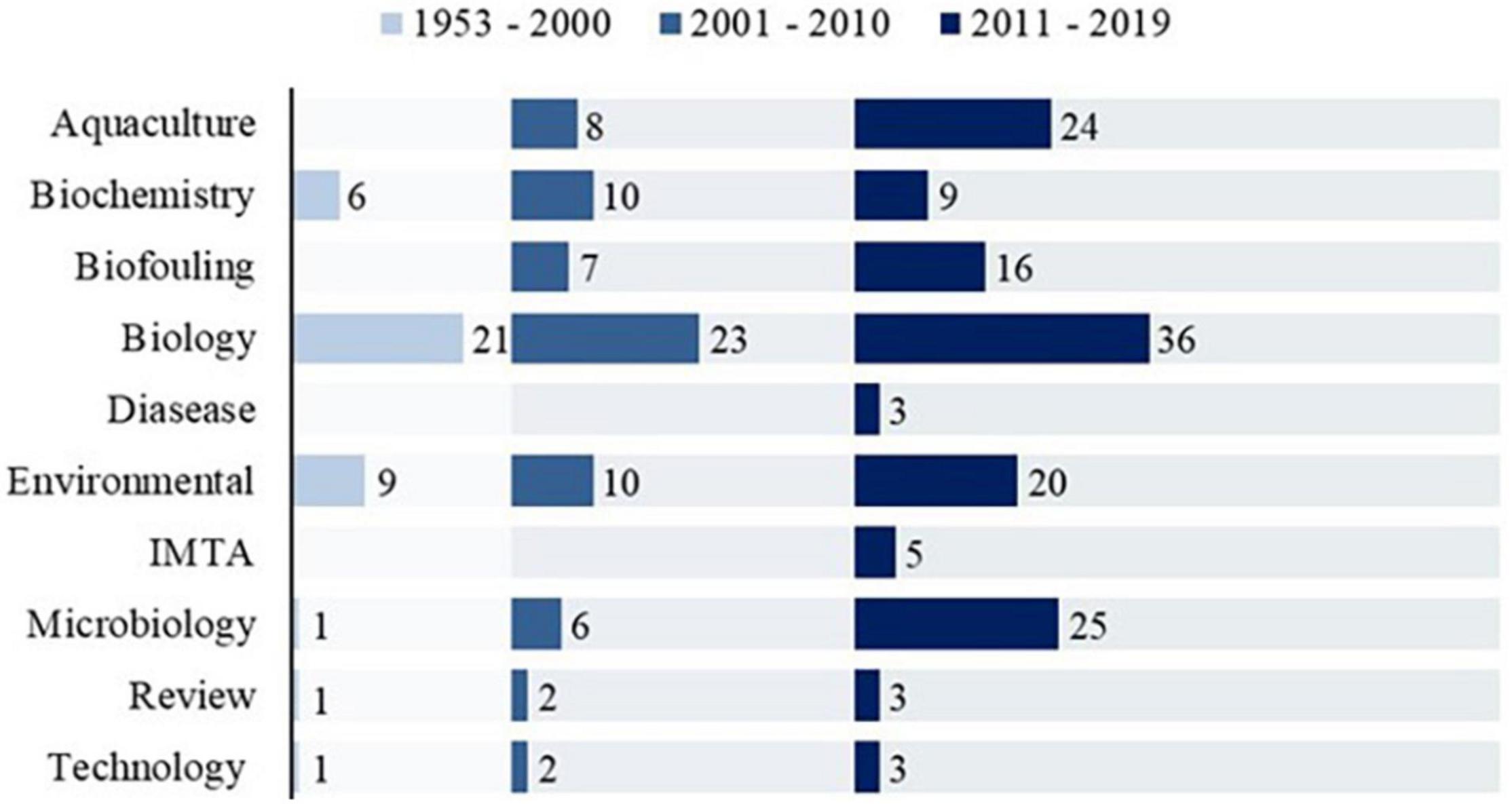
Figure 2. Number of publications (n = 148), from 1953 to 2019, assigned to each research category.
In addition, bacteria and associated diseases with ascidians are a growing concern, as seen with the increase in the number of publications within the category “Microbiology.” A total of 45 species, belonging to 3 orders and 12 families (Supplementary Table 2) were present in this review, in which solitary ascidians represented 72% and merely 28% were colonial ascidians (Figure 3).
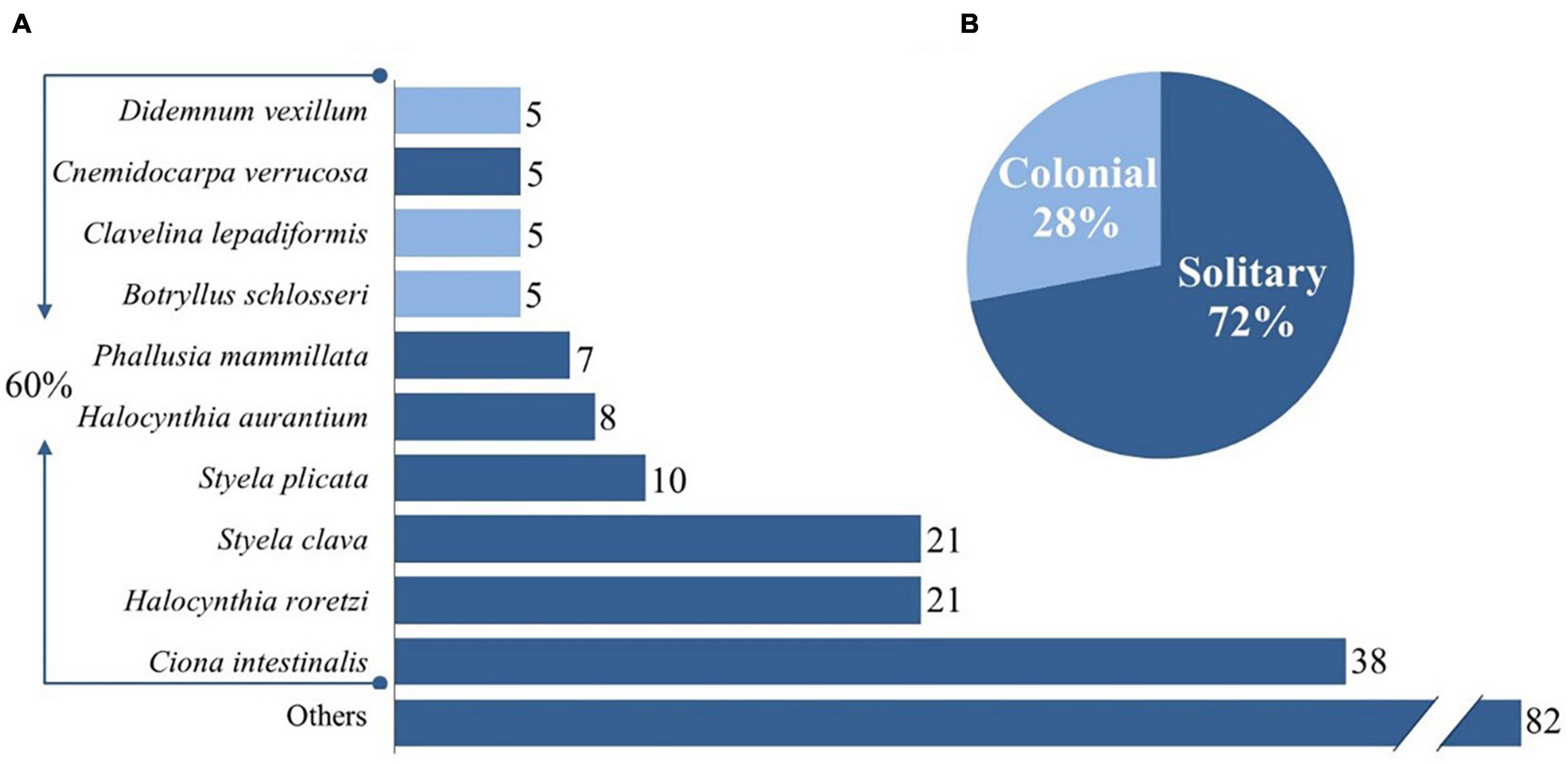
Figure 3. (A) Top ten studied ascidian species and total number of publications addressing them. Light blue represents colonial species, dark blue represents solitary species; (B) Percentage of solitary and colonial ascidians addressed in the 148 publications surveyed. See Supplementary Material for complete list of studied species (Supplementary Table 1).
Despite the high number of ascidians from the marine realm, solely three species dominated the focus of scientists throughout the years. Ciona intestinalis, a translucent column-like tunicate, was by far the most studied species, followed by Halocynthia roretzi and Styela clava (Figure 3). A detailed analysis was performed regarding the three questions (Table 2). Ascidians C. intestinalis and S. clava were the two most studied species for their biological and ecological importance as filter-feeders [What?] and on the most effective combination of species for IMTA [How?]; while H. roretzi and Halocynthia aurantium, were mostly studied for their potential as bioresource [For what?] (26 and 11%, respectively).
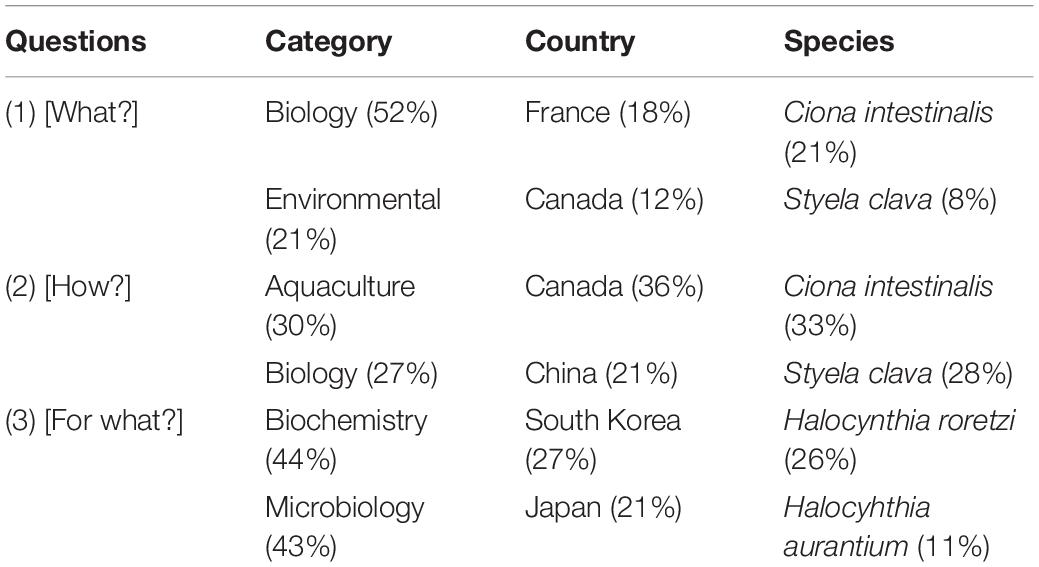
Table 2. Percentages of the top two research categories, countries, and ascidian species that most contributed to each of the questions [What?], [How?], [For what?], addressed in the 148 publications surveyed.
[What Do We Know?] (Question 1)
To comprehend the role of ascidians as filter-feeders and their importance, a better understanding of basic biology is needed. Essentially, water filtrations rates were present in most of the 29 publications analyzed (Figure 1), with these referring to 31 different species (allocated to 17 different genera). Solitary ascidians, such as C. intestinalis, Phallusia mammillata, and Styela plicata, were the most investigated species accounting for 17.5, 9.5, and 7.9% of the publications, respectively. France is in the leadership both in the number of ascidians species being studied, as well as in the number of studies performed (Table 3).

Table 3. Summary of the main features of filtration, pumping, and retention rate of the studied ascidians addressed in the 29 publications selected regarding question 1 [What?].
Filtration rates presented a great variability between the different species of ascidians addressed, with intraspecific variability also being recorded, for example, for C. intestinalis with values ranging from 3.5 to 11.9 L h–1 (Fiala-Médioni, 1974; Petersen and Riisgärd, 1992) and for P. mammillata with values ranging from 4.4 to 11.9 L h–1 (Fiala-Médioni, 1973; Hily, 1991; Table 3). Nakai et al. (2018) demonstrated that water filtration rate increases with size, while Ribes et al. (1998) showed that filtration rates may vary seasonally, displaying an increase with rising water temperatures. Just 30% of publications registered retention rate values. A total of 17 species were investigated, with only two species being colonial ascidians. Particle retention varied from 1.7 to 4.71 μm (mean value).
[How Do Ascidians Perform in Integrated Multi-Trophic Aquaculture?] (Question 2)
Question 2 focused on understanding which combination of species with ascidians contributes the most to enhance the performance of IMTA frameworks, their extractive ability, and their impacts on other cultured species. Only five out of the 19 relevant publications (Figure 1) specifically addressed IMTA. These five publications presented similar aspects, as they were all performed in China and addressed S. clava.
The main goal of these publications was to optimize the commercial production and growth of the sea cucumber Apostichopus japonicus (Figure 4). The remaining 14 publications addressed issues associated with the impact of biofouling promoted by ascidians on cultured species, namely, mussels, oysters, and scallops were addressed (Table 4). C. intestinalis was the most discussed ascidian regarding this topic.

Figure 4. Schematic representation of the main features of integrated multitrophic aquaculture (IMTA) frameworks using ascidians addressed in the 5 of the 19 publications selected regarding question 2 [How?].
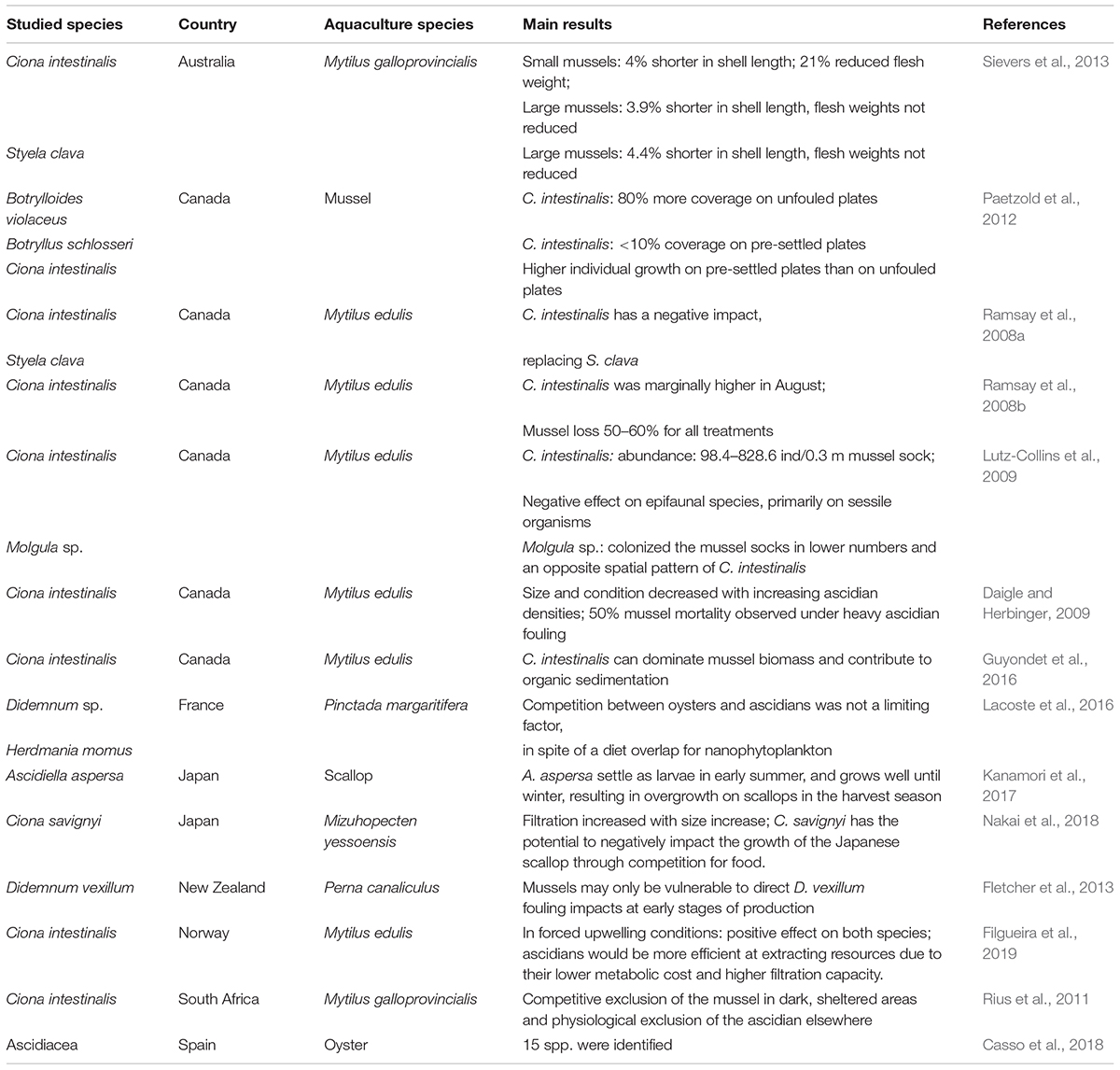
Table 4. Summary of the main impacts produced by ascidians in aquaculture scenarios addressed in the 19 publications selected regarding question 2 [How?].
[For What Kind of Bioactive Products?] (Question 3)
The recognition of the nutritional value sensu lato of ascidians and their potential as bioresources was considered in 28 publications (Figure 1), addressing 25 species belonging to 15 genera.
Ascidians being addressed under this scope mostly originated from Asian countries (China, Japan, South Korea, and North Korea), with a major focus on C. intestinalis, Halocynthia sp., and Styela sp. Amongst the various studies, 16 of them addressed specifically the fatty acid composition of ascidians, including 20 species belonging to 13 genera. From these, few analyzed the tunic and inner body separately (Zhao et al., 2015; Zhao and Li, 2016), with the remaining analyzing the whole body of ascidians.
A wide range of fatty acids was identified with percentages varying from 0.06 to 44% total fatty acid (Jeong et al., 1996; Zlatanos et al., 2009), nonetheless, palmitic acid (16:0), stearic acid (18:0), arachidonic acid [AA-20:4 (n-6)], eicosapentaenoic acid (EPA–20:5n-3) and docosahexaenoic acid (DHA–22:6n-3) were consistently recorded (Figure 5), see Supplementary Table 3 for further detail. Fatty acids 16:0 and 18:0 were constantly higher in all studied ascidians, however, in several species, EPA and DHA presented high values as well (Carballeira et al., 1995; Jeong et al., 1996; Zhao and Li, 2016).
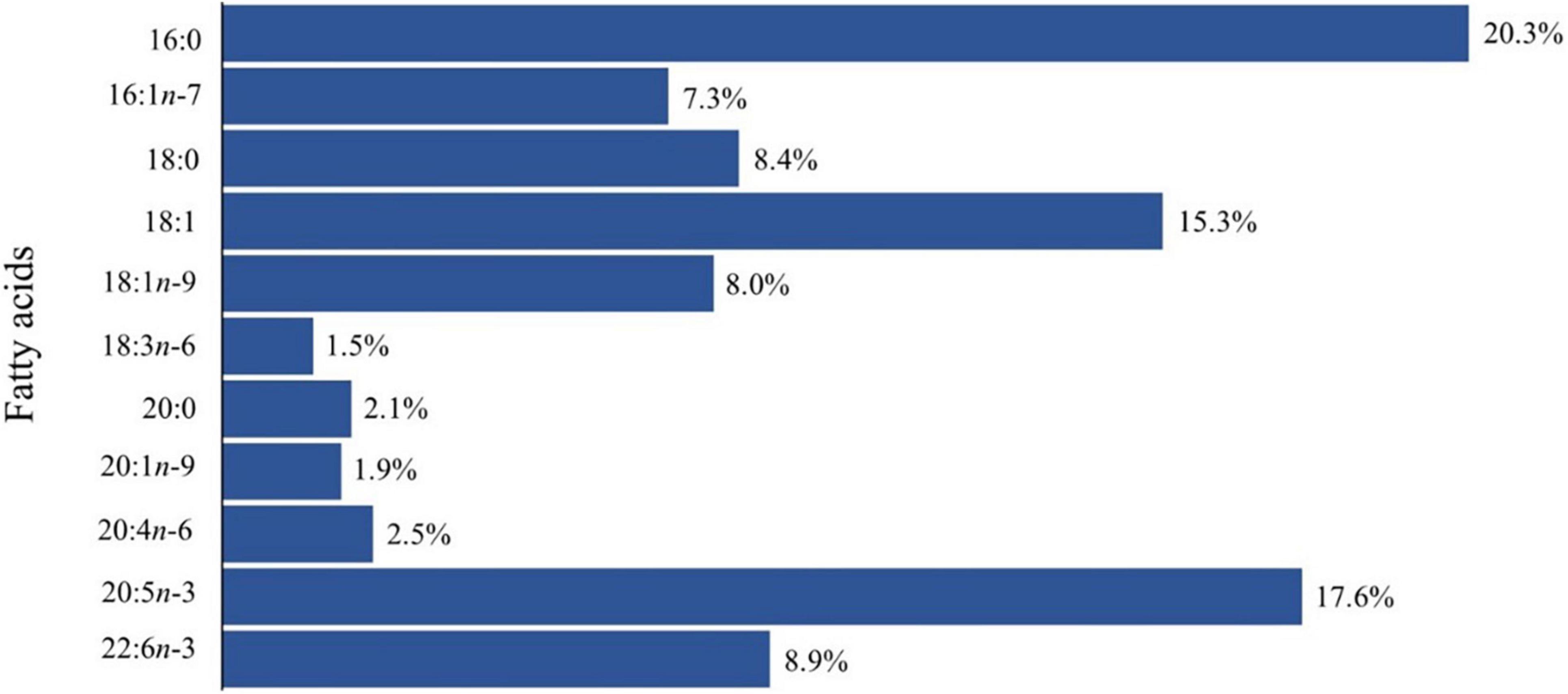
Figure 5. Mean relative percentage values of total fatty acids of the ascidians addressed in this review. See Supplementary Table 2 for further details.
Out of the studies analyzed, biocompounds such as didemnilactones A and B and neodidemnilactone (Niwa et al., 1994), 2,3-dihydroxy fatty acid glycosphingolipids (Aiello et al., 2003), anticancer ecteinascidin 743 (Mendola, 2003), pentylphenols, cyclopropane fatty acid, and cyclopentenones (Rob et al., 2011) were proven to originate from ascidians (Table 5). Cytotoxicity against human solid tumor cell lines (Bao et al., 2009), against HCT116 cells (human colon cancer cells), and inhibition of the division of fertilized sea urchin eggs (Rob et al., 2011) are just some examples of these compound functionalities.
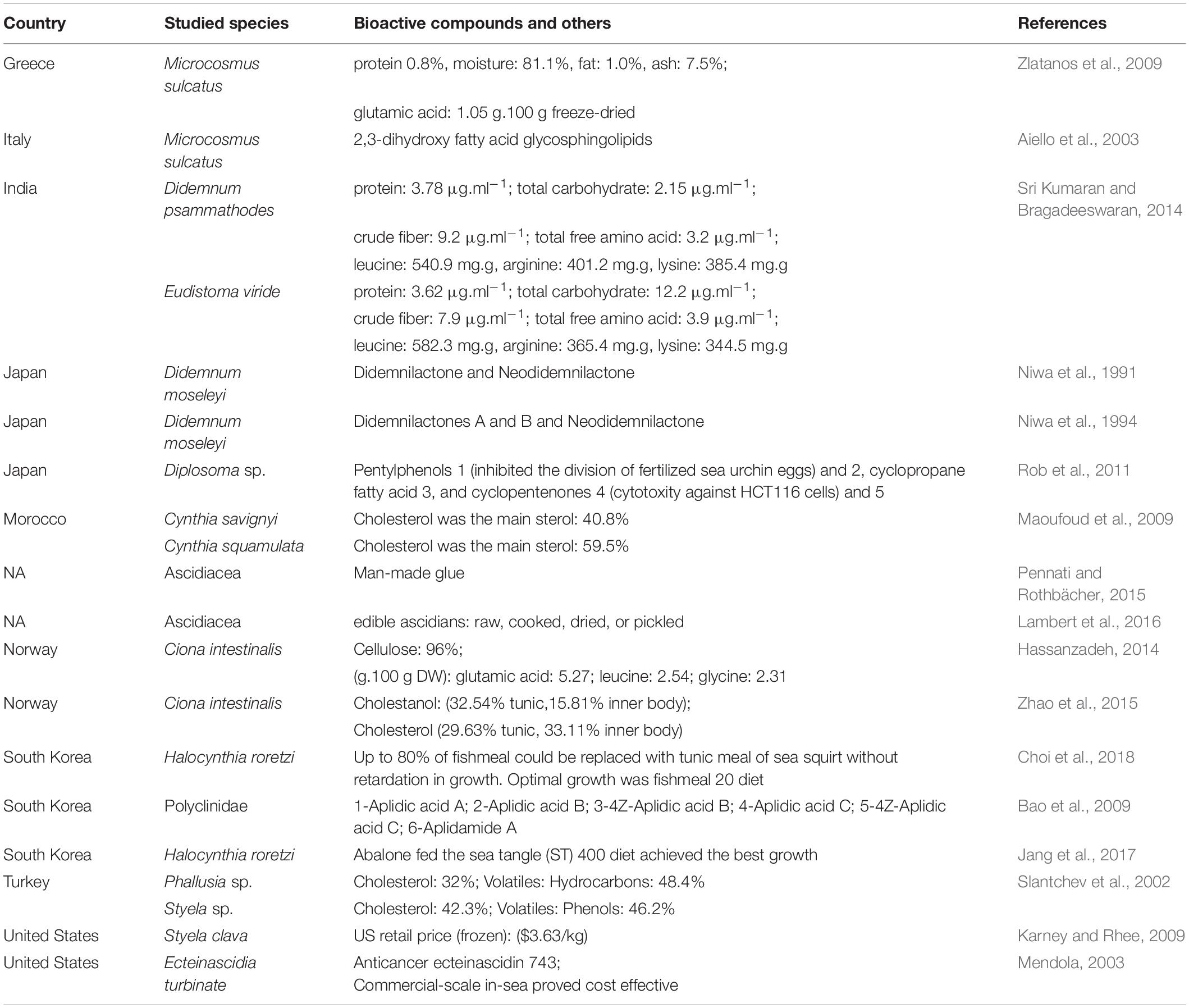
Table 5. Summary of the main attributes of the bioactive compounds of ascidians and other features addressed in the 28 publications selected regarding question 3 [For what?].
Discussion
Ascidians as Organic Matter Extractive Species
Over the years, ascidian’s biology and functionality have been of growing interest and several studies have addressed water filtration, clearance, retention, pumping, ingestion, and digestion rates. According to Fiala-Médioni (1978a), the definition of filtration rate is the volume of water that has been cleared of particles in a given time frame. Authors have gradually replaced the term “filtration rate” for clearance rate and although this topic has been widely addressed, previous reports have shown considerable variation in the results being reported.
Petersen (2007) complied information on the suspension-feeding of ascidians and concluded that “filtration rates in different species at identical conditions will not vary more than within the same species of different sizes” and also suggests that ascidians are more efficient in non-turbid conditions. Moreover, this present review revealed that since Petersen’s (2007) work, there is a generalized lack of studies on this topic. In the last decade, only six new publications have addressed filtration rates, mainly on genus Styela and Ciona (Draughon et al., 2010; Sumerel and Finelli, 2014; Kang et al., 2015; Stabili et al., 2016; Hoxha et al., 2018; Nakai et al., 2018), and therefore further research is urgently needed.
Testing filtration rates can be very complex in several ways and several variables must be taken into consideration. Robbins (1983) suggested that with an increase in food concentration, the filtration rate would decrease. Randløv and Riisgärd (1979) observed that the presence of a folded pharynx in Molgula manhattensis increased the area of the water transporting structure, thus allowing for higher filtration rates. The lag-phase phenomenon was not perceived by Randløv and Riisgärd (1979) leading to lower rates being reported and ultimately to an overall misinterpretation of their findings and not allowing comparison with other studies. Therefore, the need for a lag phase with an appropriate time (20–140 min) is highly recommended (Petersen and Riisgärd, 1992). Moreover, Petersen and Svane (2002) measured the filtration rate of seven ascidians and concluded that the area of the branchial basket and the length of the ciliary band lining the stigmata openings also contributes to higher filtration rates.
Ascidians are very sensitive organisms to any chemical or mechanical disturbance, which can cause them to close their siphons and thereby stop filtration, thus generating unrealistic filtration rates. Several studies in the 1970s (Fiala-Médioni, 1973, 1974, 1978a,b; Randløv and Riisgärd, 1979) concluded that undisturbed ascidians filter water very efficiently and at constant rates, a feature that will unquestionably optimize their performance if these are employed in the IMTA framework.
Most often, it is not easy to evaluate if filtration rates are at their optimal by merely recording the appearance of ascidians (unlike what occurs for some bivalves, such as mussels) (Petersen and Riisgärd, 1992). As environmental variables play an important role in the filtration process, several investigations aimed to elucidate the relationship between filtration rate, body size, temperature, and particle concentration (Fiala-Médioni, 1978b; Petersen and Riisgärd, 1992; Kang et al., 2015). In sum, the standardization of the methodology used to investigate filtration rate is at a high demand to better evaluate and compare data from different research.
Consistent results were observed allowing to affirm that as ascidians increase in size, their filtration rate will also increase, and filtration rate declined with temperatures above 20–21°C, this being true for ascidians from temperate waters. Moreover, the optimal temperature for ascidians’ filtration rate may vary with the species being addressed and with the local conditions. Nakai et al. (2018) registered an optimal filtration at temperatures of 24–25°C for Ciona savignyi. Several reports focus the deleterious effects of biofouling by ascidians on mussel farming and their potential competition as filter feeders for trophic resources. One study compared ascidian and mussel filtration rates and highlighted that at 16 and 19°C these are similar (Daigle and Herbinger, 2009).
Conversely, allied with the filtration process is particle retention efficiency. Various approaches have shown that the diet of ascidians mainly comes from smaller particles [particulate organic matter (POM) < 20 μm] (Ju et al., 2015, 2016), picophytoplankton (<2 μm), and phytoplankton biomass (Riisgärd and Larsen, 2016). Moreover, Lacoste et al. (2016) verified an overall lack of food selectivity. The retention efficiency increased for particle sizes 2–5 μm (Armsworthy et al., 2001), in which particles from 2 to 3 μm were completely retained (Randløv and Riisgärd, 1979) and retention efficiency decreased for particles above 4.5 μm (Pascoe et al., 2007).
In general, a threshold of 2–4 μm is observed. In a more recent study, with an in situ experiment using 6 different ascidian species, a 95% retention efficiency was registered for 1 μm particles and 50% efficiency for submicron particles (0.3 μm), thus widening ascidians scope (Jacobi, 2018). The ability of S. plicata and Polyandrocarpa zorritensis to remove Vibrio alginolyticus from seawater has also been tested, with S. plicata showing a higher efficiency for bioremediation and restoring seawater quality (Stabili et al., 2016). The same authors also demonstrated that retention efficiency was higher in solitary ascidians (81%) than in colonial ones (41%). Lefebvre et al. (2000) used oysters in a land-based fish-farm effluent and confirmed that suspended feeders can improve water quality and add economic value. However, the retention efficiency of filter-feeders in an IMTA scenario must be dealt with caution as many parameters may influence the settling velocity of the suspended particles of organic matter (Reid et al., 2009).
Nonetheless, further research is much needed considering that our systematic review revealed the existence of few publications addressing this topic in colonial ascidians.
Ascidians Incorporated in Integrated Multi-Trophic Aquaculture Frameworks
Despite the increase in interest in IMTA frameworks over the last years, ascidians have rarely been addressed under this scope. Most publications on aquaculture mostly focus on -ascidians as pests due to biofouling features and negative impacts on aquaculture facilities, mainly on shellfish productions (Carver et al., 2003). Cultured shellfish can be negatively affected by ascidian fouling in many ways, with these causing a reduction in mussel growth, flesh weight, and reduced overall size and condition (Daigle and Herbinger, 2009; Sievers et al., 2013; Guyondet et al., 2016; Nakai et al., 2018).
In extreme conditions, this may even lead to mussel mortality (Daigle and Herbinger, 2009). However, this scenario cannot be generalized, as Cordell et al. (2013) did not record any negative effects on mussel growth at four different locations and Sievers et al. (2013) observed no reduction of flesh weight was seen in larger mussels. Moreover, Lacoste et al. (2016) found that food competition between oysters and ascidians was not a limiting factor, which advises caution on making generalized assumptions on the negative impacts of ascidians on the farming of bivalves.
Indeed, several factors such as location, species involved, environmental parameters, sampling, and experiment conditions, among others must also be considered (Fletcher et al., 2013). Furthermore, some ascidians present invasive traits, growing quickly and therefore must be supervised to not overwhelm and overgrow the other culture species.
The solitary ascidian, C. intestinalis, was investigated in 60% of publications in this field, given that this is one of the most studied ascidian species. As an example, they present high tolerance to a wide range of salinities and temperatures (Lutzen, 1999; Shenkar and Swalla, 2011), allowing them a worldwide spatial distribution. This biofouling ascidian, with a fast-growing rate (Ramsay et al., 2008b,Lutz-Collins et al., 2009), that contributes to organic sedimentation (Guyondet et al., 2016), and prefers unfouled sites, dark and sheltered areas (Paetzold et al., 2012) does not necessarily have negative impacts on all bivalves or other organisms, further research is needed. Recently, some studies investigated the impacts of the presence of ascidian S. clava in an IMTA framework to optimize the growth of the sea cucumber A. japonicus (Zhen et al., 2014; Ju et al., 2015, 2016). These studies have shown that an IMTA framework consisting of ascidian-sea cucumbers-microalgae, not only has the potential to reduce organic matter in the surrounding sediment (Ju et al., 2015), it can also reduce harmful bacteria (Lin et al., 2016) and purify the water body from dissolved nutrients such as nitrogen and phosphorus (Ju et al., 2015).
Moreover, this framework can also have a positive impact on the growth performance of these sea cucumber species (Zhen et al., 2014; Chen et al., 2015; Ju et al., 2016). Available literature shows that only one ascidian species (S. clava) was addressed in these studies, and yet with very positive results.
How to incorporate and manage ascidians in an IMTA framework is an important issue with many critical factors that must be considered. Growth rate, spawning season, number of generations, settlement locations, and life span are some of these factors. As an example of how contrasting can these factors be for different ascidians, C. intestinalis can produce from 12000 to 100000 eggs over different spawning periods, whereas the colonial ascidian Botryllus schlosseri can only produce up to 50 eggs in 3 months (Paetzold et al., 2012). Solitary ascidians Ascidiella aspersa, C. intestinalis (Millar, 1952), and Corella willmeriana (Lambert, 1968) can develop into mature adults in just 3 months reach up to 50, 120, and 12 mm, respectively, with 1 or 2 generations and a life span of 12–18 months (A. aspersa and C. intestinalis) and 3 months (C. willmeriana).
The difficulty arises in the management of these biological and ecological characteristics due to the range of intra and interspecific variability and the potential environmental impacts that using ascidians may bring (e.g., biofouling). The existence of a specific area that may promote the settlement of ascidians, such as longlines or PVC plates, can be a simple solution to foster the production of biomass of these organisms and allow to easily remove their biomass for multiple applications.
Exploring the possibility of using multiple combinations of different ascidian species with other taxa, such as fish, shellfish, or echinoderms (namely, sea cucumbers) is paramount to test innovative IMTA frameworks with enhanced socio-economic and environmental performance.
Ascidians as Bioresources for High-End Uses
Considering the increase of wild-harvested or cultured ascidians for human consumption, mainly in Japan, South Korea, and Chile, knowledge on the proximate composition, biocompounds, food safety issues are of greater relevance.
Over the last decade, an increasing concern on food safety issues associated with ascidians has led prompt several studies on the identification of bacteria associated with edible ascidians such as H. aurantium (Chen et al., 2018) and H. roretzi (Kumagai et al., 2011). Bacteria associated with ascidians can also be a source of bioactive secondary metabolites and biosurfactants with diverse biotechnology applications in the food-processing industry, among other high-end markets (Achieng et al., 2017).
Several natural products have been isolated from ascidians, for example, the cellulose that is present almost exclusively in the ascidian’s tunic and it is rich in carbohydrate contents (Zhao and Li, 2016), whereas the inner body is protein-rich (Berrill and Ray Society, 2005; Hassanzadeh, 2014). Many other compounds, for example, alkaloids, cyclic peptides, and polyketides, collagens, sulfated polysaccharides, glycosaminoglycans, sterols, among others, can be exploited as by-products in the pharmaceutical and chemical industry (Hassanzadeh, 2014; Monmai et al., 2018) due to their antibacterial, antifungal, antitumor and anti-inflammatory activities (Chen et al., 2018). Numerous biocompounds have successfully been retrieved from ascidians, a recent review on this matter describes “about 160 molecules endowed with antimicrobial activity produced by ascidians and/or by their associated microorganisms” (Casertano et al., 2020).
In recent years, the search for new chemical constituents derived from marine invertebrates has increased intensity (Datta et al., 2015). For instance, Pennati and Rothbächer (2015) investigated ascidian’s larval bioadhesion properties to develop man-made glues and fouling resistant surfaces from solitary and colonial ascidians. Nowadays, ascidians are used in multiple applications such as fishing bait, health supplement tablets (Lambert et al., 2016), and as ornamental species for marine aquaria, fetching high prices online.12
Looking at fatty acids in more detail, our review revealed that approximately 70% of publications regarding fatty acids focused on solitary ascidians and once again ascidian C. intestinalis was the main focus. Many studies have drawn their attention to establishing ascidians as a new bioresource for n-3 fatty acids-rich marine lipids (Hassanzadeh, 2014; Zhao et al., 2015; Zhao and Li, 2016). Nonetheless, the profiling of fatty acids in ascidians, in general, is still poorly explored. Our study retrieved information from 20 species, with 13 ascidian species being addressed only once.
The overall results suggest that ascidians can be a good source of n-3 polyunsaturated fatty acids, namely, essential fatty acids such as EPA and DHA, which were detected in most ascidians surveyed (Dagorn et al., 2010; Zhao et al., 2015). Therefore, ascidians present a high nutritional value, they are a healthy seafood choice due to their high protein levels and low calories (Lee et al., 1995; Kang et al., 2011). Hassanzadeh (2014) concluded that the composition profile of ascidian fatty acids seems to be similar to fish oil. Therefore, ascidians biomass may eventually be a good alternative to fish oil and fish meal in formulated aquafeeds.
Moreover, ascidians present amino acid composition similar to egg albumin, suggesting a great potential and capability to be weighed as marine organisms’ feed (Hassanzadeh, 2014). Indeed, the replacement of fish meal with ascidian’s biomass in aquafeeds has already started being addressed with Jang et al. (2017) and Choi et al. (2018) having partially or fully replaced the fish meal with the tunic of the ascidian H. roretzi in aquafeeds for the abalone Haliotis discus with compromising its growth performance.
Conclusion
In the past two decades, considerable insights have been achieved on ascidians’ ecology and biology, including filtration and retention efficiencies. Their nutritional value and potential role in IMTA frameworks are also starting to be thoroughly investigated.
Despite the intra and interspecific variability recorded for ascidians filtration rates, there is a consensus that these organisms do display high filtration rates, that they can retain submicron and picoplankton particles, and they also present a fast-growing rate. As available scientific evidence suggests that these organisms are capable to perform well under an IMTA framework, however, it is important to investigate if competition with other filter-feeders for trophic resources and space can occur, namely, with mussels, scallops, and oysters.
Furthermore, available studies to date suggest that ascidians achieve higher growth performances in IMTA frameworks when in the presence of sea cucumbers and fish. The development of innovative IMTA frameworks is important to maximize the systems carrying capacity.
Finally, among other potentially bioactive compounds, ascidians represent a rich source of EPA and DHA, both being essential fatty acids paramount for human consumption, marine fish, and shrimp nutrition. Despite some cultural barriers in western countries, ascidians are increasingly regarded as a healthy seafood for human consumption, being an interesting source of essential amino and fatty acids. The use of ascidians as an alternative ingredient for the formulation of aquafeeds also looks promising and will certainly deserve further attention in coming years.
Author Contributions
LM contributed to the main investigation, writing the original draft, reviewing, and editing the final version. RC provided supervision of the writing, reviewing, editing, and validation. AL contributed to supervision of the writing, reviewing and editing process, validation, and funding acquisition. All authors contributed to the article and approved the submitted version.
Funding
We are thankful to the Integrated Program of SR&TD “Smart Valorization of Endogenous Marine Biological Resources Under a Changing Climate” (reference Centro-01-0145-FEDER-000018) and to FCT (Portuguese Foundation for Science and Technology) for the financial support through a Ph.D. grant attributed to LM (PD/BD/127918/2016). We acknowledge financial support to CESAM by FCT/MEC (UIDP/50017/2020 + UIDB/50017/2020) through national funds.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.849870/full#supplementary-material
Footnotes
References
Achieng, A. O., Rawat, M., Ogutu, B., Guyah, B., Ong’echa, J. M., Perkins, D. J., et al. (2017). Ascidians: an emerging marine model for drug discovery and screening. Curr. Top. Med. Chem. 17, 2056–2066. doi: 10.2174/1568026617666170130
Aiello, A., Fattorusso, E., Mangoni, A., and Menna, M. (2003). Three new 2, 3-dihydroxy fatty acid glycosphingolipids from the Mediterranean tunicate Microcosmus sulcatus. Eur. J. Org. Chem. 4, 734–739.
Albert, S., Bonaglia, S., Stjärnkvist, N., Winder, M., Thamdrup, B., and Nascimento, F. J. A. (2021). Influence of settling organic matter quantity and quality on benthic nitrogen cycling. Limnol. Oceanogr. 66, 1882–1895. doi: 10.1002/LNO.11730
Alexander, K. A., Angel, D., Freeman, S., Israel, D., Johansen, J., Kletou, D., et al. (2016). Improving sustainability of aquaculture in Europe: stakeholder dialogues on Integrated Multi-trophic Aquaculture (IMTA). Environ. Sci. Policy 55, 96–106. doi: 10.1016/j.envsci.2015.09.006
Armsworthy, S. L., Macdonald, B. A., and Ward, J. E. (2001). Feeding activity, absorption efficiency and suspension feeding processes in the ascidian, Halocynthia pyriformis (Stolidobranchia: Ascidiacea): responses to variations in diet quantity and quality. J. Exp. Biol. Ecol. 260, 41–69. doi: 10.1016/s0022-0981(01)00238-6
Bao, B., The, H., Zhang, P., Hong, J., Lee, C., Young, H., et al. (2009). Bicyclic α,ω-dicarboxylic acid derivatives from a colonial tunicate of the family Polyclinidae. Bioorg. Med. Chem. Lett. 19, 6205–6208. doi: 10.1016/j.bmcl.2009.08.094
Barrington, K., Chopin, T., and Robinson, S. (2009). “Integrated multi-trophic aquaculture (IMTA) in marine temperate waters,” in Integrated Mariculture: a Global Review. (ed.) D. Soto, (Rome: FAO).
Béné, C., Barange, M., Subasinghe, R., Pinstrup-Andersen, P., Merino, G., Hemre, G. I., et al. (2015). Feeding 9 billion by 2050 - putting fish back on the menu. Food Secur. 7, 261–274. doi: 10.1007/s12571-015-0427-z
Berrill, N. J., Ray Society (2005). The Tunicata With an Account of the British Species. Lymington: Pisces Conservation.
Bullard, S. G., Davis, C. V., and Shumway, S. E. (2013). Seasonal patterns of ascidian settlement at an aquaculture facility in the Damariscotte River, Maine. J. Shellfish Res. 32, 255–264. doi: 10.2983/035.032.0202
Carballeira, N. M., Shalabi, F., Stefanov, K., and Dimitrov, K. (1995). Comparison of the fatty acids of the tunicate Botryllus schlosseri from the Black Sea with two associated bacterial strains. Lipids 30, 677–679. doi: 10.1007/BF02537006
Carlisle, D. B. (1966). The ciliary current of Phallusia [Ascidiacea] and the squirting of sea squirts. J. Mar. Biol. Assoc. United Kingdom 46, 125–127.
Carvajalino-Fernández, M. A., Sævik, P. N., Johnsen, I. A., Albretsen, J., and Keeley, N. B. (2020). Simulating particle organic matter dispersal beneath Atlantic salmon fish farms using different resuspension approaches. Mar. Pollut. Bull. 161:111685. doi: 10.1016/J.MARPOLBUL.2020.111685
Carver, C. E., Chisholm, A., and Mallet, A. L. (2003). Strategies to mitigate the impact of Ciona intestinalis (L.) biofouling on shellfish production. J. Shellfish Res. 22, 621–631.
Casertano, M., Menna, M., and Imperatore, C. (2020). The ascidian-derived metabolites with antimicrobial properties. Antibiotics 9:510. doi: 10.3390/antibiotics9080510
Casso, M., Navarro, M., Ordóñez, V., and Fernández-tejedor, M. (2018). Seasonal patterns of settlement and growth of introduced and native ascidians in bivalve cultures in the Ebro Delta (NE Iberian Peninsula). Reg. Stud. Mar. Sci. 23, 12–22. doi: 10.1016/j.rsma.2017.11.002
Chen, L., Hu, J., Xu, J., Shao, C., and Wang, G. (2018). Biological and chemical diversity of ascidian-associated microorganisms. Mar. Drugs 743:362. doi: 10.3390/md16100362
Chen, L., Xing, R., Jiang, A., Teng, L., and Wang, C. (2015). A preliminary study on the potential value of a novel integrated aquaculture system on water purification. Aquac. Int. 24, 879–892. doi: 10.1007/s10499-015-9958-9958
Choi, D. G., Kim, J., Yun, A., and Cho, S. H. (2018). Dietary substitution effect of fishmeal with tunic meal of sea squirt, Halocynthia roretzi, Drasche, on growth and soft body composition of juvenile abalone, Haliotis discus, Reeve 1846. J. World Aquac. Soc. 49, 1095–1104. doi: 10.1111/jwas.12537
Chopin, T., Buschmann, A. H., Halling, C., Troell, M., Kautsky, N., Neori, A., et al. (2001). Integrating seaweeds into marine aquaculture systems: a key toward sustainability. J. Phycol. 37, 975–986.
Clos, K. T. D., Jones, I. T., Carrier, T. J., Damian, C., and Jumars, P. A. (2017). Model-assisted measurements of suspension-feeding flow velocities. J. Exp. Biol. 220, 2096–2107. doi: 10.1242/jeb.147934
Cordell, J. R., Levy, C., and Toft, J. D. (2013). Ecological implications of invasive tunicates associated with artificial structures in Puget Sound, Washington, USA. Biol. Invasions 15, 1303–1318. doi: 10.1007/s10530-012-0366-y
Costello, C., Cao, L., Gelcich, S., Cisneros-mata, M. Á., Free, C. M., Froehlich, H. E., et al. (2020). The future of food from the sea. Nature 588, 95–100. doi: 10.1038/s41586-020-2616-y
Dagorn, F., Dumay, J., Monniot, C., Rabesaotra, V., and Barnathan, G. (2010). Phospholipid distribution and phospholipid fatty acids of the tropical tunicates Eudistoma sp. and Leptoclinides uniorbis. Lipids 45, 253-261. doi: 10.1007/s11745-010-3389-3380
Daigle, R. M., and Herbinger, C. M. (2009). Ecological interactions between the vase tunicate (Ciona intestinalis) and the farmed blue mussel (Mytilus edulis) in Nova Scotia, Canada. Aquat. Invasions 4, 177–187. doi: 10.3391/ai.2009.4.1.18
Datta, D., Nath Talapatra, S., and Swarnakar, S. (2015). Bioactive compounds from marine invertebrates for potential medicines - an overview. Int. Lett. Nat. Sci. 34, 42–61.
Dijkstra, J. A., and Nolan, R. (2017). Potential of the invasive colonial ascidian, Didemnum vexillum, to limit escape response of the sea scallop, Placopecten magellanicus. Aquactic Invasions 6, 451–456. doi: 10.3391/ai.2011.6.4.10
Draughon, L. D., Scarpa, J., Hartmann, J. X., Draughon, L. D., Scarpa, J., and Hartmann, J. X. (2010). Are filtration rates for the rough tunicate Styela plicata independent of weight or size? J. Environ. Sci. Heal. Part A 45, 168–176. doi: 10.1080/10934520903429816
Fiala-Médioni, A. (1973). Ethologie alimentaire d’ invertébrés benthiques filtreurs (ascidies). I. Dispositif expérimental. Taux de filtration et de digestion chez Phallusia mammillata. Mar. Biol. 23, 137–145.
Fiala-Médioni, A. (1974). Ethologie alimentaire d’ invertébrés benthiques filtreurs (ascidies). II. variations des taux de filtration et de digestion en fonction de l’ espèce. Mar. Biol. 28, 199–206. doi: 10.1007/BF00387298
Fiala-Médioni, A. (1978a). Filter-feeding ethology of benthic invertebrates (Ascidians). IV. pumping rate, filtration rate, filtration efficiency. Mar. Biol. 48, 243–249.
Fiala-Médioni, A. (1978b). Filter-feeding ethology of benthic invertebrates (Ascidians). V. Influence of temperature on pumping, filtration and digestion rates and rhythms in Phallusia mamillata. Mar. Biol. 48, 251–259.
Fiala-Médioni, A. (1979). Effects of oxygen tension on pumping, filtration and oxygen uptake in the ascidian Phallusia mammillata. Mar. Ecol. Prog. Ser. 1, 49–53.
Filgueira, R., Strople, L. C., Strohmeier, T., Rastrick, S., and Strand, O. (2019). Mussels or tunicates: that is the question. evaluating efficient and sustainable resource use by low-trophic species in aquaculture settings. J. Clean. Prod. 231, 132–143. doi: 10.1016/j.jclepro.2019.05.173
Fletcher, L. M., Forrest, B. M., and Bell, J. J. (2013). Impacts of the invasive ascidian Didemnum vexillum on green-lipped mussel Perna canaliculus aquaculture in New Zealand. Aquac. Environ. Interact. 4, 17–30. doi: 10.3354/aei00069
Fry, J. P., Love, D. C., MacDonald, G. K., West, P. C., Engstrom, P. M., Nachman, K. E., et al. (2016). Environmental health impacts of feeding crops to farmed fish. Environ. Int. 91, 201–214. doi: 10.1016/j.envint.2016.02.022
Gasparini, F., Manni, L., Cima, F., Zaniolo, G., Burighel, P., Caicci, F., et al. (2015). Sexual and asexual reproduction in the colonial ascidian Botryllus schlosseri. Genesis 53, 105–120. doi: 10.1002/dvg.22802
Guyondet, T., Patanasatienkul, T., Comeau, L. A., Landry, T., and Davidson, J. (2016). Preliminary model of tunicate infestation impacts on seston availability and organic sedimentation in longline mussel farms. Aquaculture 465, 387–394. doi: 10.1016/j.aquaculture.2016.09.026
Hassanzadeh, M. (2014). Unique marine organism: identification of some methods for biomaterial production. Chem. Eng. Trans. 37, 385–390. doi: 10.3303/CET1437065
Hily, C. (1991). Is the activity of benthic suspension feeders a factor controlling water quality in the Bay of Brest? Mar. Ecol. Prog. Ser. 69, 179–188.
Hily, C., Potin, P., and Floc’h, J. Y. (1992). Structure of subtidal algal assemblages on soft-bottom sediments: fauna/flora interactions and role of disturbances in the Bay of Brest, France. Mar. Ecol. Prog. Ser. 85, 115–130. doi: 10.3354/meps085115
Hirose, E., and Sensui, N. (2021). Substrate selection of ascidian larva: wettability and nano-structures. J. Mar. Sci. Eng. 9:634. doi: 10.3390/JMSE9060634
Hodson, S. L., Burke, C. M., and Bissett, A. P. (2000). Biofouling of fish-cage netting: the efficacy of a silicone coating and the effect of netting colour. Aquaculture 184, 277–290. doi: 10.1016/S0044-8486(99)00328-322
Honegger, T. G. (1986). Fertilization in ascidians: studies on the egg envelope, sperm and gamete interactions in Phallusia mammillata. Dev. Biol. 118, 118–128. doi: 10.1016/0012-1606(86)90079-5
Hoxha, T., Crookes, S., Lejeusne, C., Dick, J. T. A., Chang, X., Bouchemousse, S., et al. (2018). Comparative feeding rates of native and invasive ascidians. Mar. Pollut. Bull. 135, 1067–1071. doi: 10.1016/j.marpolbul.2018.08.039
Jacobi, Y. (2018). Efficient filtration of micron and submicron particles by ascidians from oligotrophic waters. Limnol. Oceanogr. 63, 267–279. doi: 10.1002/lno.10736
Jang, B., Kim, P. Y., Kim, H. S., Lee, K. W., Kim, H. J., Choi, D. G., et al. (2017). Substitution effect of sea tangle (ST) (Laminaria japonica) with tunic of sea squirt (SS) (Halocynthia roretzi) in diet on growth and carcass composition of juvenile abalone (Haliotis discus, Reeve 1846). Aquac. Nutr. 1–8. doi: 10.1111/anu.12593
Jeong, B. Y., Ohshima, T., and Koizumi, C. (1996). Hydrocarbon chain distribution of ether phospholipids of the ascidian Halocynthia roretzi and the sea urchin Strongylocentrotus intermedius. Lipids 31, 9–18. doi: 10.1007/BF02522404
Jørgensen, B. Y. C. B. (1954). Quantitative aspects of filter feeding in invertebrates. Biol. Rev. 30, 391–453. doi: 10.1128/AEM.02007-20
Jørgensen, B., and Goldberg, E. D.. (1953). Particle filtration in some ascidians and lamellibranchs. Biol. Bull. 105, 477–489.
Ju, B., Chen, L., Xing, R., and Jiang, A. (2015). A new integrated multi-trophic aquaculture system consisting of Styela clava, microalgae, and Stichopus japonicus. Aquac. Int. 23, 471–497. doi: 10.1007/s10499-014-9829-9828
Ju, B., Jiang, A., Xing, R., Chen, L., and Teng, L. (2016). Optimization of conditions for an integrated multi-trophic aquaculture system consisting of sea cucumber Apostichopus japonicus and ascidian Styela clava. Aquac. Int. 25, 265–286. doi: 10.1007/s10499-016-0027-28
Kanamori, M., Baba, K., Natsuike, M., and Goshima, S. (2017). Life history traits and population dynamics of the invasive ascidian, Ascidiella aspersa, on cultured scallops in Funka Bay, Hokkaido, northern Japan. J. Mar. Biol. Assoc. United Kingdom 97, 387–399. doi: 10.1017/S0025315416000497
Kang, C., Jung, E., Chan, W., Jung, N., Park, H., and Choi, K. (2011). Physiological energetics and gross biochemical composition of the ascidian Styela clava cultured in suspension in a temperate bay of Korea. Aquaculture 319, 168–177. doi: 10.1016/j.aquaculture.2011.06.016
Kang, C., Lee, Y., Han, E., Park, H., Yun, S., and Chan, W. (2015). Effects of temperature and body size on the physiological energetics of the stalked sea squirt Styela clava. J. Exp. Mar. Bio. Ecol. 462, 105–112. doi: 10.1016/j.jembe.2014.10.026
Karney, R. C., and Rhee, W. Y. (2009). Market potential for Styela clava, a non-indigenous pest invading New England coastal waters. Aquactic Invasions 4, 295–297. doi: 10.3391/ai.2009.4.1.31
Khalaman, V. V. (2001). Fouling communities of mussel aquaculture installations in the White Sea. Russ. J. Mar. Biol. 27, 227–237.
Kowalke, J. (1999). Filtration in antarctic ascidians - striking a balance. J. Exp. Mar. Bio. Ecol. 242, 233–244. doi: 10.1016/s0022-0981(99)00108-2
Kumagai, A., Suto, A., Ito, H., Tanabe, T., Song, J., Kitamura, S., et al. (2011). Soft tunic syndrome in the edible ascidian Halocynthia roretzi is caused by a kinetoplastid protist. Dis. Aquat. Organ. 95, 153–161. doi: 10.3354/dao02372
Lacoste, É., Raimbault, P., Harmelin-vivien, M., and Gaertner-mazouni, N. (2016). Trophic relationships between the farmed pearl oyster Pinctada margaritifera and its epibionts revealed by stable isotopes and feeding experiments. Aquac. Environ. Interact. 8, 55–66. doi: 10.3354/aei00157
Lambert, G. (1968). The general ecology and growth of a solitary ascidian, Corella willmeriana. Biol. Bull. 135, 296–307. doi: 10.2307/1539783
Lambert, G., and Lambert, C. C. (1987). Spicule formation in the solitary ascidian, Herdmania momus. J. Morphol. 192, 145–159. doi: 10.1002/jmor.1051920206
Lambert, G., Karney, R. C., Rhee, W. Y., Carman, M. R., Shellfish, V., Box, P. O., et al. (2016). Wild and cultured edible tunicates: a review. Manag. Biol. Invasions 7, 59–66.
Lee, K. H., Hong, B. I., Jung, B. C., and Cho, H. S. (1995). Seasonal variations of nutrients in warty sea squirt (Styela clava). J. Korean Soc. Food Nutr. (Korea Republic) 24, 268–273.
Lefebvre, S., Barillé, L., and Clerc, M. (2000). Pacific oyster (Crassostrea gigas) feeding responses to a fish-farm effluent. Aquaculture 187, 185–198. doi: 10.1016/S0044-8486(99)00390-397
Lesser, M. P., and Slattery, M. (2015). Picoplankton consumption supports the ascidian Cnemidocarpa verrucosa in McMurdo Sound, Antarctica. Mar. Ecol. Prog. Ser. 525, 117–126. doi: 10.3354/meps11215
Lin, J., Ju, B., Yao, Y., Lin, X., Xing, R., Teng, L., et al. (2016). Microbial community in a multi-trophic aquaculture system of Apostichopus japonicus, Styela clava and microalgae. Aquac. Int. 24, 1119–1140. doi: 10.1007/s10499-016-9975-9972
Lohrer, A. M., Hewitt, J. E., and Thrush, S. F. (2006). Assessing far-field effects of terrigenous sediment loading in the coastal marine environment. Natl. Inst. Water Atmos. Res. 315, 13–18. doi: 10.3354/meps315013
Lutz-Collins, V., Ramsay, A., Quijón, P. A., and Davidson, J. (2009). Invasive tunicates fouling mussel lines: evidence of their impact on native tunicates and other epifaunal invertebrates. Aquat. Invasions 4, 213–220. doi: 10.3391/ai.2009.4.1.22
Lutzen, J. (1999). Styela clava Herdman (Urochordata, Ascidiacea), a successful immigrant to North West Europe: ecology, propagation and chronology of spread. Helgoländer Meeresuntersuchungen 52, 383–391.
Maoufoud, S., Abdelmjid, A., Abboud, Y., and Tarik, A. (2009). Chemical composition of fatty acids and sterols from tunicates Cynthia savignyi, Cynthia squamulata and from the brown alga Cystoseira tamariscifolia. Phys. Chem. News 47, 115–119.
Mendola, D. (2003). Aquaculture of three phyla of marine invertebrates to yield bioactive metabolites: process developments and economics. Biomol. Eng. 20, 441–458. doi: 10.1016/S1389-0344(03)00075-73
Millar, R. H. (1952). The annual growth and reproductive cycle in four ascidians. J. Mar. Biol. Assoc. United Kingdom 31, 41–61. doi: 10.1017/S0025315400003672
Monmai, C., Go, S. H., Shin, I. -S., You, S. G., Lee, H., Kang, S. B., et al. (2018). Immune-enhancement and anti-inflammatory activities of fatty acids extracted from Halocynthia aurantium tunic in RAW264.7 cells. Mar. Drugs 16:309. doi: 10.3390/md16090309
Nakai, S., Shibata;, J., Umehara, A., Okuda, T., and Nishijima, W. (2018). Filtration rate of the ascidian Ciona savignyi and its possible impact. Thalassas 34, 271–277. doi: 10.1007/s41208-017-0061-y
Naylor, R., and Burke, M. (2005). Aquaculture and ocean resources: raising tigers of the sea. Annu. Rev. Environ. Resour. 30, 185–218. doi: 10.1146/annurev.energy.30.081804.121034
Neori, A., Chopin, T., Troell, M., Buschmann, A. H., Kraemer, G. P., Halling, C., et al. (2004). Integrated aquaculture: rationale, evolution and state of the art emphasizing seaweed biofiltration in modern mariculture. Aquaculture 231, 361–391. doi: 10.1016/j.aquaculture.2003.11.015
Niwa, H., Inagaki, H., and Yamada, K. (1991). Didemnilactone and neodidemnilactone, two new fatty acid metabolites possessing a 10-membered lactone from the tunicate Didemnum moseleyi (Herdman). Tetrahedron Lett. 32, 5127–5128.
Niwa, H., Watanabe, M., Inagaki, H., and Yamada, K. (1994). Didemnilactones A and B and Neodidemnilactone, three new fatty acid metabolites isolated from the tunicate Didemnum moseleyi (Herdman). Tetrahedron 50, 7385–7400.
Ordóñez, V., Pascual, M., Rius, M., and Turon, X. (2013). Mixed but not admixed: a spatial analysis of genetic variation of an invasive ascidian on natural and artificial substrates. Mar. Biol. 160, 1645–1660. doi: 10.1007/s00227-013-2217-2215
Paetzold, S. C., Giberson, D. J., Hill, J., Davidson, J. D. P., and Davidson, J. (2012). Effect of colonial tunicate presence on Ciona intestinalis recruitment within a mussel farming environment. Manag. Biol. Invasions 3, 15–23.
Pascoe, P. L., Parry, H. E., and Hawkins, A. J. S. (2007). Dynamic filter-feeding responses in fouling organisms. Aquat. Biol. 1, 177–185. doi: 10.3354/ab00022
Pennati, R., and Rothbächer, U. (2015). Bioadhesion in ascidians: a developmental and functional genomics perspective. Interface Focus 2, 1–10. doi: 10.1098/rsfs.2014.0061
Petersen, J. K. (2007). Ascidian suspension feeding. Rome 342, 127–137. doi: 10.1016/j.jembe.2006.10.023
Petersen, J. K., and Riisgärd, H. U. (1992). Filtration capacity of the ascidian Ciona intestinalis and its grazing impact in a s hallow fjord. Mar. Ecol. Prog. Ser. 88, 9–17. doi: 10.3354/meps088009
Petersen, J. K., and Svane, I. (2002). Filtration rate in seven Scandinavian ascidians: implications of the morphology of the gill sac. Mar. Biol. 140, 397–402. doi: 10.1007/s002270100706
Pile, A. J. (2005). Overlap in diet between co-occuring active suspension feeders on tropical and temperate reefs. Bull. Mar. Sci. 76, 743–749.
Ramsay, A., Davidson, J., Landry, T., and Arsenault, G. (2008a). Process of invasiveness among exotic tunicates in Prince Edward Island, Canada. Biol. Invasions 10, 1311–1316. doi: 10.1007/s10530-007-9205-y
Ramsay, A., Davidson, J., Landry, T., and Stryhn, H. (2008b). The effect of mussel seed density on tunicate settlement and growth for the cultured mussel, Mytilus edulis. Aquaculture 275, 194–200. doi: 10.1016/j.aquaculture.2008.01.024
Randløv, A., and Riisgärd, H. U. (1979). Efficiency of particle retention and filtration rate in four species of ascidians. Mar. Ecol. Prog. Ser. 1, 55–59.
Reid, G. K., Lefebvre, S., Filgueira, R., Robinson, S. M. C., Broch, O. J., Dumas, A., et al. (2020). Performance measures and models for open-water integrated multi-trophic aquaculture. Rev. Aquac. 12, 47–75. doi: 10.1111/RAQ.12304
Reid, G. K., Liutkus, M., Robinson, S. M. C., Chopin, T. R., Blair, T., Lander, T., et al. (2009). A review of the biophysical properties of salmonid faeces: implications for aquaculture waste dispersal models and integrated multi-trophic aquaculture. Aquac. Res. 40, 257–273. doi: 10.1111/J.1365-2109.2008.02065.X
Ribes, M., Coma, R., and Gili, J. (1998). Seasonal variation of in situ feeding rates by the temperate ascidian Halocynthia papillosa. Mar. Ecol. Prog. Ser. 175, 201–213.
Riisgärd, H. U., and Larsen, P. S. (2016). “Filter-feeding zoobenthos and hydrodynamics,” in Marine Animal Forests (ed.) S. Rossi (Switcherland: Springer International Publishing)
Rius, M., Heasman, K. G., and Mcquaid, C. D. (2011). Long-term coexistence of non-indigenous species in aquaculture facilities. Mar. Pollut. Bull. 62, 2395–2403. doi: 10.1016/j.marpolbul.2011.08.030
Rob, T., Ogi, T., Maarisit, W., Taira, J., and Ueda, K. (2011). Isolation of C11 compounds and a cyclopropane fatty acid from an Okinawan ascidian, Diplosoma sp. Molecules 16, 9972–9982. doi: 10.3390/molecules16129972
Robbins, I. J. (1983). The effects of body size, temperature, and suspension density on the filtration and ingestion of inorganic particulate suspensions by ascidians. J. Exp. Mar. Bio. Ecol. 70, 65–78. doi: 10.1016/0022-0981(83)90149-1
Robbins, J. (1984). The regulation of ingestion rate, at high suspended particulate concentrations, by some phleobranchiate ascidians. J. Exp. Mar. Bio. Ecol. 82, 1–10.
Rosa, M., Holohan, B. A., Shumway, S. E., Bullard, S. G., Wikfors, G. H., Morton, S., et al. (2013). Biofouling ascidians on aquaculture gear as potential vectors of harmful algal introductions. Harmful Algae 23, 1–7. doi: 10.1016/j.hal.2012.11.008
Shenkar, N., and Swalla, B. J. (2011). Global diversity of Ascidiacea. PLoS One 6:e20657. doi: 10.1371/journal.pone.0020657
Sievers, M., Fitridge, I., Dempster, T., and Keough, M. J. (2013). Biofouling leads to reduced shell growth and flesh weight in the cultured mussel Mytilus galloprovincialis. Biofouling 29, 97–107. doi: 10.1080/08927014.2012.749869
Slantchev, K., Yalçın, F., Ersöz, T., Nechev, J., Çalis, I., Stefanov, K., et al. (2002). Composition of lipophylic extracts from two tunicates, Styela sp. and Phallusia sp. from the Eastern Mediterranean. Z. Naturforsch 57, 534–540. doi: 10.1515/znc-2002-5-623
Sri Kumaran, N., and Bragadeeswaran, S. (2014). Nutritional composition of the colonial ascidian Eudistoma viride and Didemnum psammathodes. Biosci. Biotechnol. Res. Asia 11, 331–338. doi: 10.13005/bbra/1427
Stabili, L., Licciano, M., Flavia, M., and Giangrande, A. (2016). Filtering activity on a pure culture of Vibrio alginolyticus by the solitary ascidian Styela plicata and the colonial ascidian Polyandrocarpa zorritensis: a potential service to improve microbiological seawater quality economically. Sci. Total Environ. 573, 11–18. doi: 10.1016/j.scitotenv.2016.07.216
Sumerel, A. N., and Finelli, C. M. (2014). Particle size, flow speed, and body size interactions determine feeding rates of a solitary ascidian Styela plicata: a flume experiment. Mar. Ecol. Prog. Ser. 495, 193–204. doi: 10.3354/meps10571
Troell, M., Joyce, A., Chopin, T., Neori, A., Buschmann, A. H., and Fang, J. G. (2009). Ecological engineering in aquaculture - potential for integrated multi-trophic aquaculture (IMTA) in marine offshore systems. Aquaculture 297, 1–9. doi: 10.1016/j.aquaculture.2009.09.010
Vieira, H., Leal, M. C., and Calado, R. (2020). Fifty shades of blue: how blue biotechnology is shaping the bioeconomy. Trends Biotechnol. 38, 940–943. doi: 10.1016/j.tibtech.2020.03.011
Yahel, G., Yahel, R., Katz, T., Lazar, B., Herut, B., and Tunnicliffe, V. (2008). Fish activity: a major mechanism for sediment resuspension and organic matter remineralization in coastal marine sediments. Mar. Ecol. Prog. Ser. 372, 195–209. doi: 10.3354/MEPS07688
Yahel, R., Yahel, G., and Genin, A. (2002). Daily cycles of suspended sand at coral reefs: a biological control. Limnol. Oceanogr. 47, 1071–1083. doi: 10.4319/LO.2002.47.4.1071
Young, C. M. (1989). Distribution and dynamics of an intertidal ascidian pseudopopulation. Bull. Mar. Sci. 45, 288–303.
Zhao, Y., and Li, J. (2016). Ascidian bioresources: common and variant chemical compositions and exploitation strategy - examples of Halocynthia roretzi, Styela plicata, Ascidia sp. and Ciona intestinalis. Zeitschrift Naturforsch. - Sect. C J. Biosci. 71, 165–180. doi: 10.1515/znc-2016-2012
Zhao, Y., Wang, M., Lindström, M. E., and Li, J. (2015). Fatty acid and lipid profiles with emphasis on n-3 fatty acids and phospholipids from Ciona intestinalis. Lipids 50, 1009–1027. doi: 10.1007/s11745-015-4049-4041
Zhen, Y., Aili, J., and Bao, J. (2014). Effect of the ascidian Styela clava on the growth of small holothurians of the species Apostichopus japonicus. Mar. Freshw. Behav. Physiol. 47, 211–226. doi: 10.1080/10236244.2014.929800
Zlatanos, S., Laskaridis, K., and Sagredos, A. (2009). Determination of proximate composition, fatty acid content and amino acid profile of five lesser-common sea organisms from the Mediterranean Sea. Int. J. Food Sci. Technol. 44, 1590–1594. doi: 10.1111/j.1365-2621.2008.01870.x
Keywords: tunicates, bioresource, IMTA, filtration rate, fatty acids, retention efficiency
Citation: Marques L, Calado R and Lillebø AI (2022) Potential of Ascidians as Extractive Species and Their Added Value in Marine Integrated Multitrophic Aquaculture Systems–From Pests to Valuable Blue Bioresources. Front. Mar. Sci. 9:849870. doi: 10.3389/fmars.2022.849870
Received: 06 January 2022; Accepted: 24 January 2022;
Published: 08 March 2022.
Edited by:
Sílvia Lourenço, Escola de Turismo e Tecnologia Marítima, Instituto Politécnico de Leiria, PortugalReviewed by:
Patrick Cahill, Cawthron Institute, New ZealandAdriana Giangrande, University of Salento, Italy
Copyright © 2022 Marques, Calado and Lillebø. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luisa Marques, bHVpc2EubWFycXVlc0B1YS5wdA==; Ana Isabel Lillebø, bGlsbGVib0B1YS5wdA==