 Rachel Wachtendonk
Rachel Wachtendonk John Calambokidis
John Calambokidis Kiirsten Flynn
Kiirsten Flynn
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Mar. Sci., 17 May 2022
Sec. Marine Megafauna
Volume 9 - 2022 | https://doi.org/10.3389/fmars.2022.847032
This article is part of the Research TopicWhales and ClimateView all 14 articles
Large marine mammals can serve as an indicator of the overall state of the environment due to their apex position in marine food webs and their functions as sentinels of change. Reductions in prey, driven by changes in environmental conditions can manifest in reduced fat stores that are visible on whales. We developed a non-invasive technique using photographs of blue whales taken on the US west coast from 2005-2018 (n=3,660) and scored body condition based on visible undulations from the vertebral processes and body shape. We analyzed patterns in the body condition of whales across years and their relation to oceanographic conditions. Females with calves had significantly poorer body conditions and calves had significantly better body conditions compared to other adult whales (Chi-Square, x2 = 170.36, df=6, p<2.2e-16). Year was a significant factor in body condition (Chi-Square, x2 = 417.73, df=39, p<0.001). The highest proportion of whales in poor body condition was observed for 2015 (one of the only two years along with 2017 where >50% had poor body condition) coincides with the marine heat wave that affected the NE Pacific 2014-2016. A cumulative mixed model examining the relationship between body condition and environmental variables revealed that negative Pacific Decadal Oscillation and longer upwelling seasons correlated with better blue whale body condition, likely to be due to higher primary productivity and prey availability. This study indicates that with an adequate scoring method, photographs collected during boat based surveys can be used to effectively evaluate whale health in response to a changing ocean.
Large marine mammals can serve as an indicator of the overall state of the environment due to their apex position in marine food webs and their functions as sentinels of change (Moore, 2008; Williams et al., 2013). Changes in environmental conditions can result in decreased prey availability at the base of the food web, which impacts predators further up the trophic chain (Acevedo-Whitehouse and Duffus, 2009; Benoit-Bird and McManus, 2012; Soledade Lemos et al., 2020; Rasmussen et al., 2021). In the case of whales, reduced food availability results in visible reductions in fat stores (Lockyer, 1986; Konishi, 2006; Christiansen et al., 2013; Braithwaite et al., 2015). Whales with a compromised body condition may be unable to take on the challenges of their extreme life history, such as their long migrations and periods of fasting, which can lead to reduced survivorship and reproductive fitness (Greene et al., 2003; Lockyer, 2007; Bradford et al., 2012; Soledade Lemos et al., 2020). This can have adverse impacts on the population or species as a whole, which is compounded in recovering populations (Acevedo-Whitehouse and Duffus, 2009).
These impacts are magnified in pregnant and lactating females who rely on bountiful foraging seasons in order to sustain themselves and their calf during the different phases of pregnancy (Lockyer, 1984; Miller et al., 2012; Christiansen et al., 2013; Christiansen et al., 2014). Body condition affects the fertility of many mammalian species, including whales, where a minimum threshold of body fat is needed for ovulation, and therefore, pregnancy (Young, 1976; Frisch, 1984; Lockyer, 1987; Miller et al., 2011). Poor foraging can also influence future pregnancies through a process known as the carry-over effect- where a reduced body condition of a reproductive female can decrease her ability to carry a pregnancy to term in future years, ultimately impacting population growth rates (Soledade Lemos et al., 2020).
Blue whales (Balaenoptera musculus) are the largest animal on Earth and thus have the highest prey demands (Savoca et al., 2021). These large animals feed almost exclusively on euphausiids (krill) and consume up to 22 tons of prey per day, often employing energetically costly maneuvers to obtain their prey (Acevedo-Gutiérrez et al., 2002; Croll et al., 2005; Calambokidis et al., 2008; Goldbogen et al., 2011; Goldbogen et al., 2015; Barlow et al., 2020; Savoca et al., 2021). Blue whales were hunted extensively during the whaling era, facing reductions in populations of 90% or more (Jefferson et al., 2015; Thomas et al., 2015). After they became a protected species by the International Whaling Commission in 1966 blue whale populations started to increase, but they are still considered endangered. The eastern North Pacific population of blue whales is currently estimated to be at 1,898 (lower and upper 20th percentile values of 1,767 to 2,038) individuals (Calambokidis and Barlow, 2020) and is considered to be at or near pre-whaling estimates (Monnahan et al., 2015).
Due to their narrowly focused prey regime, blue whale health is closely linked with environmental factors that determine prey abundance and concentration (Croll et al., 2005; Calambokidis et al., 2007; Silber et al., 2017). The eastern North Pacific blue whales forage partially in the highly productive California Current System which spans from British Columbia, Canada (~50°N) to Baja California, Mexico (~15-25°N) (Huyer, 1983; Di Lorenzo et al., 2008; McClatchie et al., 2008; Checkley and Barth, 2009; Palacios et al., 2019). Productivity in the California Current System is driven by patterns of variability on various spatial and temporal scales from local and seasonal upwelling to ocean basin level interannual/decadal Pacific Decadal Oscillation. From 2014 to 2016 the California Current System also experienced a large marine heat wave with sea surface temperature anomalies exceeding 3°C, which was the warmest three- year stretch on record (Schwing et al., 2006; McClatchie et al., 2008; Checkley and Barth, 2009; Di Lorenzo and Mantua, 2016; Gentemann et al., 2017; Thompson et al., 2018).
Body condition is commonly used to assess the health of individuals in both terrestrial and aquatic ecosystems (Lockyer et al., 1985; Lockyer, 1986; Batzli and Esseks, 1992; Pettis et al., 2004; Konishi, 2006; Acevedo-Whitehouse and Duffus, 2009; Bradford et al., 2012; Williams et al., 2013; Braithwaite et al., 2015; Soledade Lemos et al., 2020; Akmajian et al., 2021; Rasmussen et al., 2021). Body condition studies are important to gauge the health of recovering populations of animals and the environment as a whole (Williams et al., 2013). However, measuring large whale body condition at sea is logistically challenging due to their size, sighting frequency, and remoteness (Pettis et al., 2004; Konishi, 2006). Previous studies have used blubber thickness and whaling measurements of girth in carcasses to estimate health (Lockyer et al., 1985; Lockyer, 1986; Konishi, 2006; Williams et al., 2013; Braithwaite et al., 2015). But these studies have limitations as few countries have allowed the lethal removal of whales since the 1980s when international whaling was outlawed. Other studies have used ultrasound to measure the blubber thickness on live right whales (Miller et al., 2011), but such methodology would be challenging for faster, more streamlined whales.
Alternatively, the use of photographic data has been developed as a low cost, easy to implement, and non-invasive way to monitor whale health. Qualitative visual body condition assessments of target areas make this methodology valuable for long-term studies (Pettis et al., 2004; Bradford et al., 2012; Akmajian et al., 2021). Photographic studies show temporal trends in the body condition of baleen whales over years for species such as North Atlantic right whales (Eubalaena glacialis) (Pettis et al., 2004), and gray whales (Eschrichtius robustus) (Bradford et al., 2012; Soledade Lemos et al., 2020; Akmajian et al., 2021).
The aims of this study were (1) to determine if photographs, especially those used for individual identification (ID) of blue whales can be used to assess body condition, (2) to determine if body condition varied by year or reproductive class, and (3) to investigate the relationships between blue whale body condition and environmental indices representative of upwelling strength and productivity in the California Current System. This research provided a non-invasive model for monitoring blue whale body condition in the future and determining how blue whales may respond to a quickly changing ocean.
This project analyzed photos collected yearly by Cascadia Research Collective and contributors from 2005 to 2018 from the US West Coast which represented 3,660 sightings of 1,112 unique blue whales that were deemed fit for assessment. Each image was scored on a scale of 1 to 3 for photo quality and proportion visible (see Supplementary Material for details). Sightings took place in all months and between 25-47°N. Most sightings were in the summer feeding season between June and October (97%), and between Ensenada, Mexico and the Gulf of the Farallones, US (30-39°N, 93%). (Calambokidis et al., 2007; Calambokidis, 2009; Calambokidis and Barlow, 2020).
Methods in this study were modified from the protocol developed for determining the body condition of North Atlantic right whales (Pettis et al., 2004) and western gray whales (Bradford et al., 2012) and were originally developed by Cascadia Research Collective to study the impact of tags on whales. One analyst (RKW) scored all images, an approach known to improve the consistency of qualitative scoring (Pettis et al., 2004). The lateral flanks forward of the dorsal fin were visually assessed for several features that were potentially indicative of the overall body conditions of the whale: 1) visible undulations along the ridge of the back reflecting the vertebral processes and 2) degree of depression (dorsal ridge) or rotundness along the lateral flanks. Body condition was scored on a scale from 0 to 3 where a score of 0 indicated that the whale had rounded sides, no undulations from vertebrae visible, and presumed to be in the best body condition (Figure 1A). A score of 1 indicated that the whale had a well-defined dorsal ridge but whose vertebrae were hardly visible (Figure 1B). A score of 2 indicated that the whale had well defined dorsal ridge and multiple vertebrae were slightly visible (Figure 1C). Finally, a score of 3 indicated that the whale had well defined dorsal ridge, multiple vertebrae are visible, and presumed to reflect the poorest body condition (Figure 1D).

Figure 1 Examples of whales in each category of body condition. (A) Score 0, good body condition, rounded sides and no vertebrae are seen. (B) Score 1, moderately good body condition, a definite dorsal ridge and possible detectable vertebrae. (C) Score 2, a moderately poor body condition, a definite dorsal ridge and slight but multiple detectable vertebrae. D) Score 3, poor body condition, a definite dorsal ridge and obvious multiple vertebrae seen. Arrows indicate rounded sides (A), dorsal ridge (B), and detectable vertebrae (C, D). The red box indicates the area assessed for body condition and proportion of image seen. All images have scores of 1 for both image quality and proportion of image seen. All images were taken by Cascadia Research Collective.
Several environmental indices are known to reflect prey availability in the California Current System on different spatial and temporal scales (Brinton and Townsend, 2003; Bograd et al., 2009; Checkley and Barth, 2009). In this study, we included the Pacific Decadal Oscillation (PDO) and the length of the upwelling season (LUSI) in the creation of our model to see if they can be linked to blue whale body condition. The PDO is the primary driver of sea surface temperature in the ocean basin and influences regime shifts (1-20 years) of many marine organisms from primary producers to marine mammals (Mantua and Hare, 2002). We looked at both the average annual PDO value for each year and PDO as a binomial factor (positive or negative). On a smaller temporal and spatial scale, seasonal upwelling occurs in the spring/summer and varies in duration (LUSI) at different latitudes along the current, which leads to high levels of primary and secondary production (Huyer, 1983; Fiedler et al., 1998; Croll et al., 2005; Bograd et al., 2009; Checkley and Barth, 2009). The California Current System also experienced a heat wave from 2014-2016 and we considered all months of those years to be under the influence of the heat wave.
We tested the effect of year and reproductive class on blue whale body condition determined through photographic scoring. First the number of whales in each body condition score ranging from 0 to 3 (healthy, moderately healthy, moderately unhealthy, and unhealthy) were summed for each reproductive class (lactating female, calf, or other) and each year (2005-2018). Lactating females and calves were determined to be in those classes for the entire year after they had been observed as a mother-calf pair. We did not otherwise consider the sex and age class of other whales since this information was only known for a small proportion of individuals. Then Chi-squared goodness of fit tests were run in the program R (R Core Team, 2019) to examine the difference in proportion of body condition scores among reproductive classes and year to determine if body condition scores differed. In addition, Kruskal-Wallis tests were run to determine if there were significant annual variations in the PDO and LUSI during the study period.
Then, we used ordinal regression models to explore what environmental variables had an effect on blue whale body condition (see Supplementary Material for details). Using the Cumulative Mixed Model test from the Ordinal package (Christensen, 2019) within the program R, we determined the effect that two categorical variables (reproductive class and proportion of image seen) and two environmental indices (PDO, LUSI) had on body condition score (0 to 3). In this analysis reproductive class is lactating female, calf, or other (IDs that did not have a known sex or age class assigned); and proportion of image seen is the score on the three-point scale (see Supplementary Material for details) for the primary image being scored. Since blue whales are highly mobile (Calambokidis et al., 1990; Mate et al., 1999; Calambokidis et al., 2009; Busquets-Vass et al., 2021) and body condition would be determined by feeding success over an extended period and range, we used a single annual average value for both the PDO and the LUSI. In each model the year was included as random effects to account for pseudoreplication.
To determine the most parsimonious model, first a full model was created with the two categorical variables (reproductive class and proportion of image seen) and then complexity (interactions between variables) and each covariate were singularly removed. Additional covariates were removed from selective models until we were able to determine the model with the lowest Akaike information criterion (AIC) score using the performance package (Lüdecke et al., 2021). A Pearson’s cross-correlation analysis was performed for all environmental variables and highly correlated variables were not included in the same models. Then environmental indices were added to the model until the model with the lowest AIC score was found. This was repeated with a one-and two-year lag for environmental variables to determine if environmental changes had a delayed effect on body condition. The final models were then compared and the best fitting model was chosen (all models tested can be found in Supplementary Material).
The lateral flanks forward of the dorsal fin showed variations through differences in visible undulations reflecting the vertebral processes and the degree of depression or rotundness along the lateral flanks, confirming photographic assessments of body condition can be used for blue whales (Figure 1). The distribution of scores for the study period showed that 34.5% of photographed whales were in good body condition (score 0), 32% of photographed whales were in moderately good body condition (score 1), 18.6% of photographed whales were in moderately poor body condition (score 2), and 14.9% of photographed whales were in poor body condition (score 3). The reproductive class had a significant impact on overall body condition (Figure 2; Chi-Square, x2 = 170.36, df=6, p<2.2e-16). Lactating females had a higher probability of being in poor body condition (60.5% score 3) and calves had a higher probability of being in good body condition (76% score 0) compared to the general population that displayed fairly evenly spread body condition scores, with more whales being in good body condition (34.4% score 0, 32.6% score 1, 19% score 2, 14% score 3).
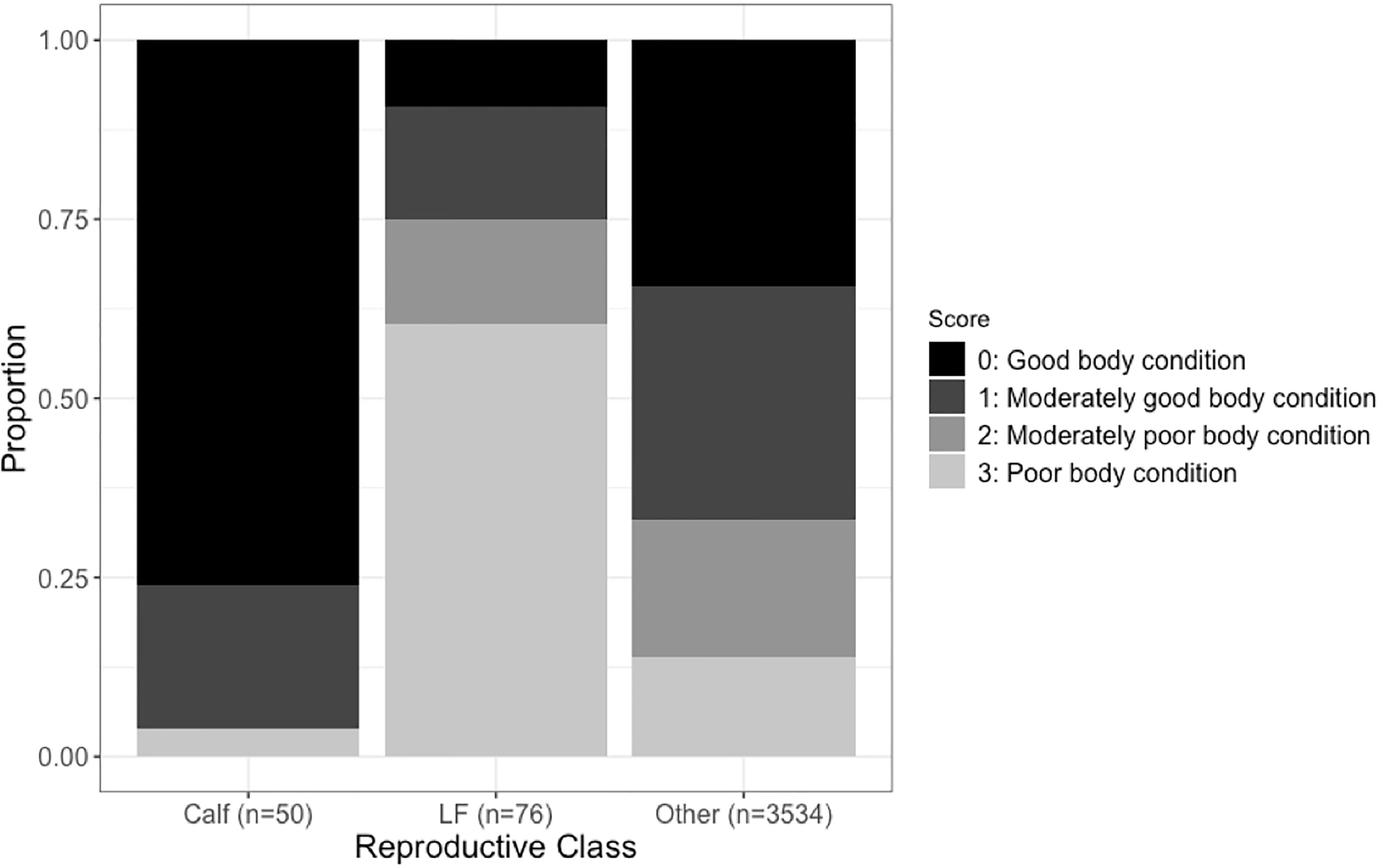
Figure 2 The proportion of each body condition score by reproductive class as a stacked bar graph (LF, lactating female).
The year was also a highly significant factor in the overall body condition for all scores (Figure 3A; Chi-Square, x2 = 417.73, df=39, p<0.001). For any given year the proportion of whales in moderately poor to poor body condition (scores 2 & 3) was 33%. But this varied widely across years with a low of 18% in 2008 to a high of 55% of whales in poor body condition in 2015. The percentage of whales in moderately poor to poor body condition did not exceed 50% except for 2015 and 2017.
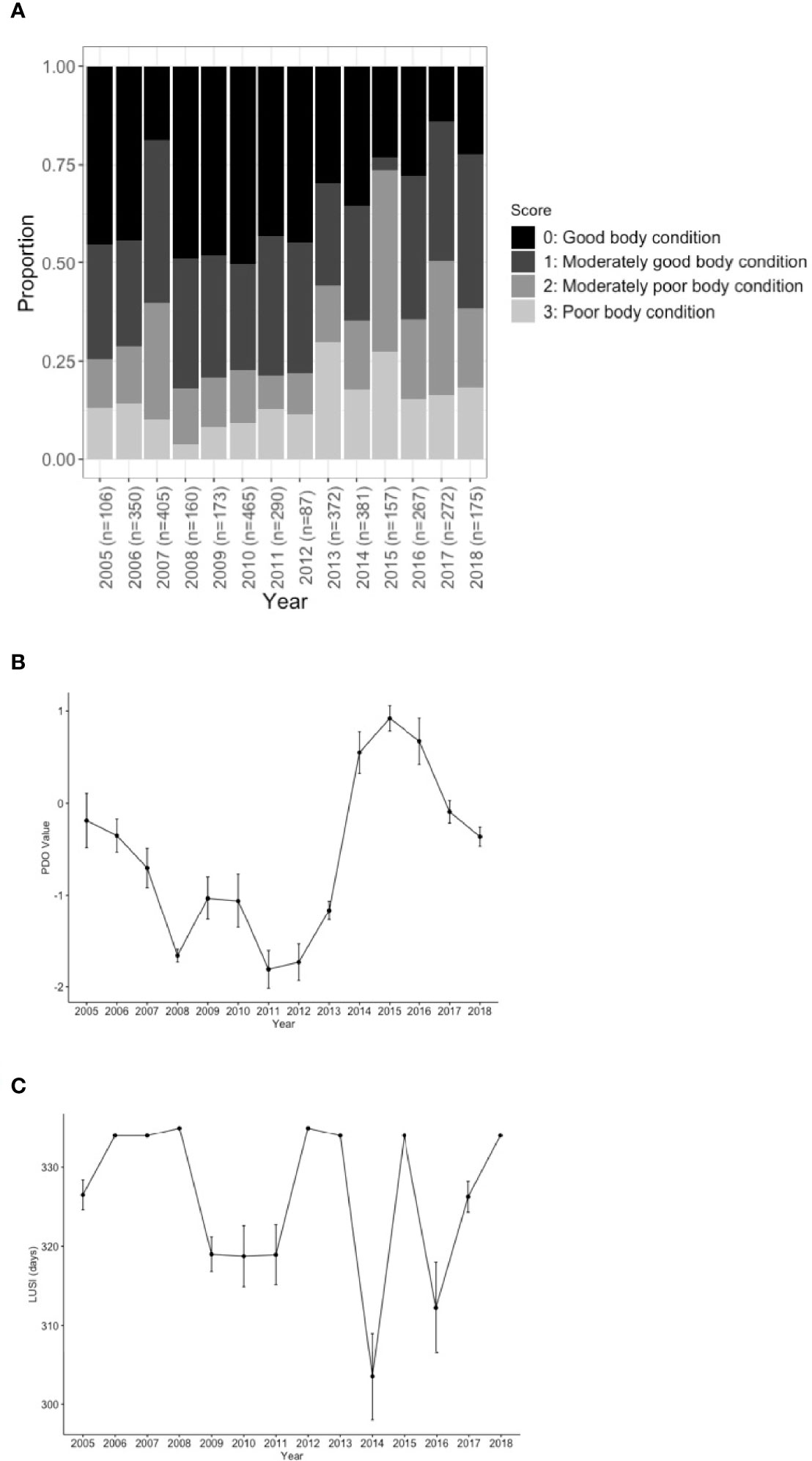
Figure 3 (A) the proportion of each body condition by year as a stacked bar graph compared to (B) average Pacific Decadal Oscillation (PDO) value (C) and the length of the upwelling season (LUSI). The marine heat wave occurred from 2014-2016. Error bars show standard error.
Two environmental indicators varied significantly by year, PDO (Kruskal-Wallis, χ2 = 105.37, df=13, p < 2.2e-16) and LUSI (Kruskal-Wallis, χ2 = 358.14, df=14, p < 2.2e-16). Only three of the fifteen years (2014-2016) had positive annual mean PDO values, and on average, the PDO was negative during the study period (-0.57 ∓ 1.09). PDO was lowest in 2011 with a value of -1.81 and highest in 2015 with a value of 0.92. On average the length of the upwelling season was 326∓21 days. LUSI was the shortest in 2014 at 304 days and longest in 2008 and 2012 at 335 days.
Of the five final models, the model incorporating the proportion of image seen, PDO value, and interaction between reproductive class and LUSI was most parsimonious (Table 1). The interaction between reproductive class and LUSI means that the length of the upwelling season impacted the groups differently. Lactating females were only observed during times that had the same LUSI value, which may explain this interaction. The significant parameters in the model were the reproductive class being a lactating female (p=0.019), the proportion of image seen (linear, p< 2e-16), the proportion of image seen (quadratic, p=2.64e-08), PDO value (p=5.10e-05), and the interaction between lactating females and the LUSI value (p=0.010, see Supplementary Material for details).

Table 1 Results comparing the three final qumulative mixed models of blue whale body condition.
In general, peaks in PDO value coincided with dips in LUSI (except for 2015) and the two variables were moderately correlated (Pearson’s cross-correlation -0.3653, t=-23.734, df=3657, p<2.2e-16)). Years that had positive PDO values (2014-2016) had an increase in the number of whales in poor body condition (scores 2 and 3; Figure 3B). The opposite was true for LUSI, with an increase in the length of the upwelling season leading to an increase in whales with good body condition (Figure 3C).
This study shows that visual health assessments based on foundational studies (Pettis et al., 2004; Bradford et al., 2012) can be applied to blue whales to track body condition over time. The lateral flanks varied in the amount of subcutaneous fat in that area which made differentiation between condition scores possible. This region had the added benefit of being the target for photo identification images, making it possible to use historical data in our analysis. Our study could be used as a basis to investigate other streamlined whales such as the minke (Balaenoptera acutorostrata), sei (Balaenoptera borealis), and fin (Balaenoptera physalus) whales.
The reproductive class of blue whales had a highly significant impact on body condition. Calves had the best body condition scores in the population and females who were seen with dependent calves had the worst scores, though this study did not account for variation among other demographic groups. Other body condition studies have found that cows who are nursing their calves are in the worst condition of observed whales (Pettis et al., 2004; Bradford et al., 2012; Soledade Lemos et al., 2020). The high reproductive costs of nursing a mysticete whale calf has been well documented, as cows must consume enough during their feeding season to sustain themselves and their quickly growing calf while in calving grounds, although all North Pacific blue whales forage year-round (Busquets-Vass et al., 2021). Blue whale calves are nursed over a period of 7-8 months during which their size more than doubles (Mackintosh and Wheeler, 1929; Jefferson et al., 2015) and this period of lactation is the costliest part of reproduction, requiring 3-5 times more energy than gestation (Miller et al., 2012).
Like Bradford et al., 2012, we saw small variations in the body condition of calves across years with 76% being scored as having good body condition scores (score 0) despite a variation of conditions being observed in the other groups. This could be due to the link between health and reproductive success seen in many species (Lockyer, 1984; Lockyer, 1986; Greene et al., 2003; Christiansen et al., 2013; Williams et al., 2013; Soledade Lemos et al., 2020). Females in poor body condition may be unable to become pregnant (Young, 1976; Frisch, 1984; Lockyer, 1987; Miller et al., 2011), and those that do then must minimize the energy expended on their fetus to maximize their own chance of survival (Christiansen et al., 2014). This means poor foraging conditions can lead to drops in calving rates- something that is especially concerning for an endangered species (Greene et al., 2003). A year of bad foraging can also influence more than just the pregnancies for that year, as a reduced body condition of a reproductive female can decrease their ability to carry a pregnancy to term in future years (carry-over effect; Christiansen et al., 2014; Soledade Lemos et al., 2020). If foraging conditions are poor for consecutive years, the long term recovery of endangered populations, especially if they are dealing with the effects of human activities, can be in jeopardy.
Blue whale body condition varied by year, but overall the proportion of whales in poor body condition remained generally low, except for two years (2015 and 2017) when more than half of the whales were in poor health. While this study focused on the blue whales feeding on the US west coast, the eastern North Pacific population feed year-round and has a much broader feeding range that extends from the Eastern Tropical Pacific to Alaska (Mate et al., 1999; Stafford et al., 1999; Calambokidis et al., 2009; Monnahan et al., 2014). After their numbers were greatly reduced from whaling, blue whales were rarely seen in Alaska; but starting in the late 1990s an increase of whales in more northern waters coincided with a decrease of blue whales off California (Calambokidis et al., 2009). It has been hypothesized that when foraging around California is poor due to decreased krill availability or increased competition, some blue whales may continue elsewhere to look for better foraging opportunities. It is likely that the pattern in blue whale body condition we observed results from a combination of some whales shifting foraging grounds and others being influenced by the feeding conditions off of California (Calambokidis et al., 2009; Busquets-Vass et al., 2021).
Variation in energy reserves (body fat) has been linked to prey availability, where oftentimes environmental conditions are used as a proxy (Braithwaite et al., 2015; Soledade Lemos et al., 2020). We looked at several environmental indices that we believed would drive prey availability and therefore body condition. PDO and LUSI were the environmental drivers selected in our final model. While our final model included data from environmental indices taken in the same year as the sightings, other models that performed well had one- or two-year lags for the environmental indices, which may reflect that blue whale body condition is a function of both conditions in the current year, as well as previous years.
PDO varied significantly by year over our study period with high values in 2014-2016. Only four years of our study period had positive PDO values (2014-2017), which followed the same pattern of poor body condition. PDO reflects sea surface temperature in the Pacific Ocean basin with positive PDO values signifying warmer ocean temperatures (Mantua and Hare, 2002; Brinton and Townsend, 2003; Di Lorenzo and Mantua, 2016). We also looked at LUSI since upwelling occurs on a smaller temporal scale than PDO, seasonally leading to high levels of primary and secondary production (Huyer, 1983; Fiedler et al., 1998; Croll et al., 2005; Checkley and Barth, 2009). The seasonal upwelling coincides with the summer feeding period for blue whales (Croll et al., 2005; Barlow et al., 2020) and therefore its length could drive overall foraging success. The worst body conditions occurred in years with the shortest upwelling seasons. Our results suggest that warm sea surface temperatures driven by positive PDO values and short upwelling seasons are detrimental to blue whale health by reducing food resources for the whales feeding off the US west coast. PDO was linked to changes in blue whale distribution and their use of areas off the US west coast versus further north into the Gulf of Alaska for present day as well as for historical whaling data showing that PDO may predict broad shifts of blue whale distribution in the eastern North Pacific (Calambokidis et al., 2009).
While not all environmental indices are highly correlated, they do play off one another and have a cumulative impact on the California Current System and the organisms that live there. Future studies may be required to quantitatively link specific indices and environmental events to whale body condition. It would be interesting to perform a similar study on other whales that forage in the same region, like humpback whales (Megaptera novaeangliae). Unlike blue whales, humpbacks forage only seasonally so the environmental factors may have a larger impact on their body condition.
We provided a basis for evaluating large whale health using historic records that can provide context for present and future conditions. This methodology can also be used to monitor populations over the long term which will only become more important in the context of a changing climate. Marine heat waves are expected to become more frequent and more intense in the future (Oliver et al., 2018) due to anthropogenically driven climate change (Frölicher et al., 2018; Oliver et al., 2018). This study indicates that with an adequate scoring method, photographs collected during boat based surveys can be used to effectively evaluate whale health in response to a changing ocean.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was reviewed and approved by Cascadia Research IACUC.
The authors confirm contribution to the paper as follows: RW, JC, and KF conceived the study. JC and KF collected original data. RW scored the data and created the model. RW was the lead author on the draft and revisions. JC and KF gave assistance in analysis and editing the manuscript. All authors contributed to the article and approved the submitted version.
Support for long term photo ID effort was provided through various contracts and grants over the years from the National Oceanic and Atmospheric Administration (NOAA) as well as support for this project through a NOAA Section 6 grant through the Washington Department of Fish and Wildlife. Support for initial assessment of blue whale body condition provided through a grant from the Office of Naval Research with funding from the Bureau of Ocean Energy Management through the National Oceanographic Partnership Program.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank the following people and institutions: NOAA for providing the principle support for the long term photo ID project over the years; Ben Miner for guidance in developing the models; Jarrod Santora, Karin Forney, and Elizabeth Becker on advice on environmental variables to use; the other staff, interns, and collaborators of Cascadia Research Collective who aided in obtaining, processing and matching photographs of blue whales especially Annie Douglas, Elana Dobson, and Azucena Ugalde; and José Pedro Andrade for his helpful advice during this project.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.847032/full#supplementary-material
Acevedo-Gutiérrez A., Croll D., Tershy B. (2002). High Feeding Costs Limit Dive Time in the Largest Whales. J. Exp. Biol. 205, 1747–1753. doi: 10.1242/jeb.205.12.1747
Acevedo-Whitehouse K., Duffus A. (2009). Effects of Environmental Change on Wildlife Health. Philos. Trans. R. Soc. B: Biol. Sci. 364, 3429–3438. doi: 10.1098/rstb.2009.0128
Akmajian A., Scordino J., Gearin P., Gosho M. (2021). Body Condition of Gray Whales (Eschrichtius robustus) Feeding on the Pacific Coast Reflects Local and Basinwide Environmental Drivers and Biological Parameters. J. Cetacean Res. Manage. 22, 87–110. doi: 10.47536/jcrm.v22i1.223
Barlow D., Bernard S., Escobar-Flores P., Palacios D., Torres L. (2020). Links in the Trophic Chain: Modeling Functional Relationships Between in Situ Oceanography, Krill, and Blue Whale Distribution Under Different Oceanographic Regimes. Marine Mammal Prog. Ser. 624, 207–225. doi: 10.3354/meps13339
Batzli G., Esseks E. (1992). Body Fat as an Indicator of Nutritional Condition for the Brown Lemming. Am. Soc. Mamma. 73 (2), 431–439. doi: 10.1098/rsbl.2012.0232
Benoit-Bird K., McManus M. (2012). Bottom-Up Regulation of a Pelagic Community Through Spatial Aggregations. Biol. Lett. 8 (5), 813–816. doi: 10.1098/rsbl.2012.0232
Bograd S., Schroeder I., Sarkar N., Qiu X., Sydeman W., Schwing F. (2009). Phenology of Coastal Upwelling in the California Current. Geophy. Res. Lett. 36, L01602. doi: 10.1029/2008GL035933
Bradford A., Weller D., Punt A., Ivashchenko Y., Burdin A., VanBlaricom G., et al. (2012). Leaner Leviathans: Body Condition Variation in a Critically Endangered Whale Population. J. Mammal 93 (1), 251–266. doi: 10.1644/11-mamm-a-091.1
Braithwaite J., Meeuwig J., Letessier T., Jenner K., Brierley A. (2015). From Sea Ice to Blubber: Linking Whale Condition to Krill Abundance Using Historical Whaling Records. Polar Biol. 38 (8), 1195–1202. doi: 10.1007/s00300-015-1685-0
Brinton E., Townsend A. (2003). Decadal Variability in Abundances of the Dominant Euphausiid Species in Southern Sectors of the California Current. Deep-Sea Res. Part II 50, 2449–2472. doi: 10.1016/S0967-0645(03)00126-7
Busquets-Vass G., Newsome S., Pardo M., Calambokidis J., Aguíñiga-García S., Páez-Rosas D., et al. (2021). Isotope-Based Inferences of the Seasonal Foraging and Migratory Strategies of Blue Whales in the Eastern Pacific Ocean. Marine Environ. Res. 163, 105201. doi: 10.1016/j.marenvres.2020.105201
Calambokidis J. (2009). Abundance Estimates of Humpback and Blue Whales Off the US West Coast Based on Mark-Recapture of Photo-Identified Individuals Through 2008 (San Diego: Pacific Scientific Review Group), 3–5. Report # PSRG-2009-07 toCA.
Calambokidis J., Barlow J. (2020). Updated Abundance Estimates for Blue and Humpback Whales Along the U.S. West Coast Using Data Through 2018. U.S. Department of Commerce:NOAA Technical Memorandum NMFS-SWFSC-634 Available at: https://repository.library.noaa.gov/view/noaa/27104
Calambokidis J., Barlow J., Ford J., Chandler T., Douglas A. (2009). Insights Into the Population Structure of Blue Whales in the Eastern North Pacific From Recent Sightings and Photographic Identifications. Marine Mammal Sci. 25, 816–832. doi: 10.1111/j.1748-7692.2009.00298.x
Calambokidis J., Douglas A., Falcone E., Schlender L. (2007). Abundance of Blue Whales Off the US West Coast Using Photo Identification. Southwest Fish. Sci. Center, AB133F06SE3906S.
Calambokidis J., Falcone E., Douglas A., Schlender L., Huggins J., et al (2009). Photographic identification of humpback and blue whales off the US West Coast: Results and updated abundance estimates from 2008 field season. Final Report for Contract AB133F08SE2786 from Southwest Fisheries Science Center, La Jolla, CA.
Calambokidis J., Schorr G., Steiger G., Francis J., Bakhtiari M., Marshall G., et al. (2008). Insights Into the Underwater Diving, Feeding, and Calling Behavior of Blue Whales From a Suction-Cup Attached Video-Imaging Tag (Crittercam). Marine Technol. Soc. J. 41, 15–25. doi: 10.4031/002533207787441980
Calambokidis J., Steiger G.H. , Cubbage J.C., Balcomb K., Ewald C., Kruse S., et al. (1990). Sightings and Movements of Blue Whales Off Central California 1986-88 From Photo-Identification of Individuals. Rep. Int. Whaling Commission 343348, (Special Issue 12), 343–348
Checkley D., Barth J. (2009). Patterns and Processes in the California Current System. Prog. Oceanog. 83, 49–64. doi: 10.1016/j.pocean.2009.07.028
Christensen R. (2019). Ordinal - Regression Models for Ordinal Data. R Package Version 2019. Available at: https://CRAN.R-project.org/package=ordinal.
Christiansen F., Víkingsson G., Rasmussen M., Lusseau D. (2013). Minke Whales Maximise Energy Storage on Their Feeding Grounds. J. Exp. Biol. 216, 427–436. doi: 10.1242/jeb.074518
Christiansen F., Víkingsson G., Rasmussen M., Lusseau D. (2014). Female Body Condition Affects Foetal Growth in a Capital Breeding Mysticete. Funct. Ecol. 28 (3), 579–588. doi: 10.1111/1365-2435.12200
Croll D., Marinovic B., Benson S., Chavez F., Black N., Ternullo R., et al. (2005). From Wind to Whales: Trophic Links in a Coastal Upwelling System. Mar. Ecol. Prog. Ser. 289, 117–130. doi: 10.3354/meps289117
Di Lorenzo E., Mantua N. (2016). Multi-Year Persistence of the 2014/15 North Pacific Marine Heat Wave. Nat. Climate Change 6 (11), 1042–1047. doi: 10.1038/nclimate3082
Di Lorenzo E., Schneider N., Cobb K., Franks P., Chhak K., Miller A., et al. (2008). North Pacific Gyre Oscillation Links Ocean Climate and Ecosystem Change. Geophy. Res. Lett. 35, L08607. doi: 10.1029/2007GL032838
Fiedler P., Reilly S., Hewitt R., Demer D., Philbrick V., Smith S., et al. (1998). Blue Whale Habitat and Prey in the California Channel Islands. Deep-Sea Res. II 45, 1781–1801. doi: 10.1016/S0967-0645(98)80017-9
Frisch R. (1984). Body Fat, Puberty, and Fertility. Biol. Rev. Cambr Philos. Soc. 59 (2), 161–188. doi: 10.1111/j.1469-185x.1984.tb00406.x
Frölicher T., Fischer E., Gruber N. (2018). Marine Heat Waves Under Global Warming. Nature 360 (560), 360–364. doi: 10.1038/s41586-018-0383-9
Frölicher T., Laufkötter C. (2018). Emerging Risks From Marine Heat Waves. Nat. Commun. 9, 650. doi: 10.1038/s41467-018-03163-6
Gentemann C., Fewings M., Garcia-Reyes M. (2017). Satellite Sea Surface Temperatures Along the West Coast of the United States During the 2014–2016 Northeast Pacific Marine Heat Wave. Geophy. Res. Lett. 44, 312–319. doi: 10.1002/2016GL071039
Goldbogen J., Calambokidis J., Oleson E., Potvin J., Pyenson N., Schorr G., et al. (2011). Mechanics, Hydrodynamics and Energetics of Blue Whale Lunge Feeding: Efficiency Dependence on Krill Density. J. Exp. Biol. 214, 131–146. doi: 10.1242/jeb.054726
Goldbogen J., Hazen E., Friedlaender A., Calambokidis J., DeRuiter S., Stimpert A., et al. (2015). Prey Density and Distribution Drive the Three-Dimensional Foraging Strategies of the Largest Filter Feeder. Funct. Ecol. 29, 951–961. doi: 10.1111/1365-2435.12395
Greene C., Pershing A., Kenney R., Jossi J. (2003). Impact of Climate Variability on the Recovery of Endangered North Atlantic Right Whales. Oceanography 16 (4), 98–103. doi: 10.5670/oceanog.2003.16
Huyer A. (1983). Coastal Upwelling in the California Current System. Prog. Oceanog. 12, 259–284. doi: 10.1016/0079-6611(83)90010-1
Jefferson T., Webber M., Pitman R. (2015). “Blue Whale- Balaenoptera Musculus (Linnaeu),” in Marine Mammals of the World: A Comprehensive Guide to Their Identification, 2nd ed. (San Diego, CA: Elsevier).
Konishi K. (2006). Characteristics of Blubber Distribution and Body Condition Indicators for Antarctic Minke Whales (Balaenoptera Bonaerensis). Mammal Study 31, 15–22. doi: 10.3106/1348-6160(2006)31[15:cobdab]2.0.co;2
Leslie M., Perkins-Taylor C., Durban J., Moore M., Miller C., Chanarat P., et al. (2020). Body Size Data Collected non-Invasively From Drone Images Indicate Morphologically Distinct Chilean Blue Whale (Balaenoptera Musculus Linnaeus 1758). Endangered Species Res. 43, 291–304. doi: 10.3354/esr01066
Lockyer C. (1984). Review of Baleen Whale (Mysticeti) Reproduction and Implications for Management. Rep. Int. Whaling Commission 6, 27–50.
Lockyer C. (1986). Body Fat Condition in Northeast Atlantic Fin Whales, Balaenoptera Physalus, and its Relationship With Reproduction and Food Resource. Can. J. Fish. Aquat. Sci. 43, 142–147. doi: 10.1139/f86-015
Lockyer C. (1987). The Relationship Between Body Fat, Food Resource and Reproductive Energy Costs in North Atlantic Fin Whales (Balaenoptera Physalus). Symp. Zool. Soc. Lond. 57, 343–361.
Lockyer C. (2007). All Creatures Great and Smaller: A Study in Cetacean Life History Energetics. J. Mar. Biol. Assoc. United Kingdom 87 (4), 1035–1045. doi: 10.1017/S0025315407054720
Lockyer C., McConnel L., Waters T. (1985). Body Condition in Terms of Anatomical and Biochemical Assessment of Body Fat in North Atlantic Fin and Sei Whales. Can. J. Zool. 63, 2328–2338. doi: 10.1139/z85-345
Lüdecke D., Makowski D., Ben-Shachar M., Patil I., Waggoner P., Wiernik B., et al. (2021). Performance: An R Package for Assessment, Comparison and Testing of Statistical Models. J. Open Source Softw. 6 (60), 3139. doi: 10.21105/joss.03139
Mantua N., Hare S. (2002). The Pacific Decadal Oscillation. J. Oceanog. 58, 35–44. doi: 10.1023/A:1015820616384
Mate B., Lagerquist B., Calambokidis J. (1999). Movements of North Pacific Blue Whales During the Feeding Season Off Southern California and Their Southern Fall Migration. Mar. Mammal Sci. 15 (4), 1246–1257. doi: 10.1111/j.1748-7692.1999.tb00888.x
McClatchie S., Goericke R., Koslow J., Schwing F., Bograd S., Charter R., et al. (2008). The State of the California Curren-2008: La Niña Conditions and Their Effects on the Ecosystem. California Cooperative Ocean. Fish. Investig. Rep. 49, 39–76.
Miller C., Best P., Perryman W., Baumgartner M., Moore M. (2012). Body Shape Changes Associated With Reproductive Status, Nutritive Condition and Growth in Right Whales Eubalaena Glacialis and E. Australis. Marine Ecol. Prog. Ser. 459, 135–156. doi: 10.3354/meps09675
Miller C., Reeb D., Best P., Knowlton A., Brown M., Moore M. (2011). Blubber Thickness in Right Whales Eubalaena Glacialis and Eubalaena Australis Related With Reproduction, Life History Status and Prey Abundance. Mar. Ecol. Prog. Ser. 438, 267–283. doi: 10.3354/meps09174
Monnahan C., Branch T., Punt A. (2015). Do Ship Strikes Threaten the Recovery of Endangered Eastern North Pacific Blue Whales? Marine Mammal Sci. 31 (1), 279–297. doi: 10.1111/mms.12157
Monnahan C., Branch T., Stafford K., Ivashchenko Y., Oleson E. (2014). Estimating Historical Eastern North Pacific Blue Whale Catches Using Spatial Calling Patterns. PloS One 9 (6), e98974. doi: 10.1371/journal.pone.0098974
Moore S. (2008). Marine Mammals as Ecosystem Sentinels. J. Mammal 89 (3), 534–540. doi: 10.1644/07-MAMM-S-312R1.1
Oliver E., Donat M., Burrows M., Moore P., Smale D., Alexander L., et al. (2018). Longer and More Frequent Marine Heat Waves Over the Past Century. Nat. Commun. 9, 1324. doi: 10.1038/s41467-018-03732-9
Palacios D., Bailey H., Becker E., Bograd S., DeAngeis M., Forney K., et al. (2019). Ecological Correlates of Blue Whale Movement Behavior and its Predictability in the California Current Ecosystem During the Summer-Fall Feeding Season. Movement Ecol. 7 (26), 1–21. doi: 10.1186/s40462-019-0164-6
Pettis H., Rolland R., Hamilton P., Brault S., Knowtown A., Kraus S. (2004). Visual Health Assessment of North Atlantic Right Whales (Eubalaena Glacialis) Using Photographs. Can. J. Zool 82, 8–19. doi: 10.1139/z03-207
Rasmussen G., Smultea M., Cloutier T., Giordano A., Kaplin B., Willey L. (2021). Quantitative Photogrammetric Methodology for Measuring Mammalian Belly Score in the Painted Dog. PloS One 16 (12), e0261171. doi: 10.1371/journal.pone.0261171
R Core Team. (2019). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: http://www.R-project.org/.
Savoca M., Czapanskiy M., Kahane-Rapport S., Gough W., Fahlbusch J., Bierlich K. (2021). Baleen Whale Prey Consumption Based on High-Resolution Foraging Measurements. Nature 599 (7883), 85–90. doi: 10.1038/s41586-021-03991-5
Schwing F., Bond N., Bograd S., Mitchell T., Alexander M., Mantua N. (2006). Delayed Coastal Upwelling Along the U.S. West Coast in 2005: A Historical Perspective. Geophys. Res. Lett. 33, L22S01. doi: 10.1029/2006GL026911
Silber G., Lettrich M., Thomas P., Baker J., Baumgartner M., Becker E., et al. (2017). Projecting Marine Mammal Distribution in a Changing Climate. Front. Mar. Sci. 4. doi: 10.3389/fmars.2017.00413
Soledade Lemos L., Burnett J., Chandler T., Sumich J., Torres L. (2020). Intra- and Inter-Annual Variation in Grey Whale Body Condition on a Foraging Ground. Ecosphere 11 (4), 1–19. doi: 10.1002/ecs2.3094
Stafford K., Nieukirk S., Fox C. (1999). An Acoustic Link Between Blue Whales in the Eastern Tropical Pacific and the Northeast Pacific. Mar. Mammal Sci. 15 (4), 1258–1268. doi: 10.1111/j.1748-7692.1999.tb00889.x
Thomas P., Reeves R., Brownell R. (2015). Status of the World's Baleen Whales. Mar. Mammal Sci. 32 (2), 682–734. doi: 10.1111/mms.12281
Thompson A., Fisher J., Jacobson K., Schroeder I., Bograd S., Hazen E, et al. (2018). State of the California Current 2017–18: Still Not Quite Normal in the North and Getting Interesting in the South. CalCOFI Rep. 59, 1–66.
Williams R., Vikingsson G., Gislason A., Lockyer C., New L., Thomas L., et al. (2013). Evidence for Density-Dependent Changes in Body Condition and Pregnancy Rate of North-Atlantic Fin Whales Over Four Decades of Varying Environmental Conditions. ICES J. Marine Sci. 70 (6), 1273–1280. doi: 10.1093/icesjms/fst059
Keywords: blue whale, body condition, environmental variability, photo-identification, marine heat wave
Citation: Wachtendonk R, Calambokidis J and Flynn K (2022) Blue Whale Body Condition Assessed Over a 14-Year Period in the NE Pacific: Annual Variation and Connection to Measures of Ocean Productivity. Front. Mar. Sci. 9:847032. doi: 10.3389/fmars.2022.847032
Received: 31 December 2021; Accepted: 12 April 2022;
Published: 17 May 2022.
Edited by:
Solène Derville, Oregon State University, United StatesReviewed by:
Amy Richardson Knowlton, New England Aquarium, United StatesCopyright © 2022 Wachtendonk, Calambokidis and Flynn. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rachel Wachtendonk, cmFjaGVsLndhY2h0ZW5kb25rQGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.