
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 31 March 2022
Sec. Marine Conservation and Sustainability
Volume 9 - 2022 | https://doi.org/10.3389/fmars.2022.812144
This article is part of the Research TopicTracking Marine Megafauna for Conservation and Marine Spatial PlanningView all 35 articles
 A. R. Patrício1,2†*
A. R. Patrício1,2†* M. Beal1,3†
M. Beal1,3† C. Barbosa4D. Diouck5
C. Barbosa4D. Diouck5 B. J. Godley2
B. J. Godley2 F. M. Madeira6
F. M. Madeira6 A. Regalla4M. S. Traoré7C. Senhoury8E. Sidina8
A. Regalla4M. S. Traoré7C. Senhoury8E. Sidina8 P. Catry1
P. Catry1Networks of marine protected areas (MPAs) are invaluable for the protection of species with high dispersal capacity, yet connectivity within networks is poorly understood. We demonstrate the connectivity within the regional MPA network in West Africa (RAMPAO), mediated by the largest green turtle population in the eastern Atlantic Ocean. We equipped with satellite tags 45 female green turtles nesting in the Bijagós Archipelago, Guinea-Bissau, and tracked them during internesting, migration, and foraging to quantify the degree of coverage the RAMPAO network provides during each of these critical periods. During the internesting period, turtles were largely concentrated around the nesting islands, with a mean of 94.8% (SD 0.1%, range: 46% - 100%, n = 40 turtles) of tracking positions falling within MPA limits. Among the 35 turtles successfully tracked into the foraging period, we identified variable migratory strategies, with 12 turtles remaining near-resident at distances of 40-90 km from breeding sites, 10 turtles migrating 300-400 km to The Gambia and Senegal, and 13 turtles traveling >1000 km to northern Mauritania. Of the 35 foraging turtles, 26 used MPAs, with a mean of 78.0% (SD 34.8%, range: 3.7% - 100%) of their tracking positions falling within the limits of RAMPAO MPAs, across Guinea-Bissau, Senegal and Mauritania. Migration corridors with high concentrations of passing turtles were mostly located nearshore, and 21% of these high passage areas fell within the MPA network. Overall, we found that this population connects five RAMPAO MPAs, yet some foraging sites (e.g., in the Bijagós) and important migration areas (e.g., Cap-Vert peninsula) described here are currently unprotected. These results are relevant to any considerations of MPA extension or establishment within the regional network, which would contribute towards meeting the Convention on Biological Diversity targets for national marine protected area estate coverage. By documenting biological connectivity across RAMPAO, this study represents an important example of the relevance of international protected area networks for green turtle conservation and for wider conservation action at a regional scale.
Marine protected areas (MPAs) are a major pillar of global ocean conservation (Laffoley et al., 2019), and the current proposed target under the Convention on Biological Diversity (CBD) for national protected marine area is 30% by 2030 (IUCN, 2016). To meet the large-scale challenges of marine conservation, regional networks of MPAs are becoming preferred management tools over standalone MPAs (Gaines et al., 2010). MPA networks can improve conservation outcomes by increasing the spatial coverage of species distributions, thereby providing safe areas for population persistence or replacement in the face of variable anthropogenic pressures (Gaines et al., 2010, Xu et al., 2020), as well as potentially increasing resilience to climate change (McLeod et al., 2009). Additionally, to be effective, MPA networks should share common conservation targets so that efforts are coordinated and outcomes are achieved across a larger area. Yet the reality is that MPAs within a network often act as isolated management units, partly because connectivity between them is non-existent or has not been empirically demonstrated (Assis et al., 2021). Hence, assessing the degree of biological connectivity across existing and proposed MPA designs is central to ensuring the effectiveness of protected area networks (Santos et al., 2021).
Connectivity within protected area networks is particularly important for the conservation of migratory species that perform cyclical movements between breeding and non-breeding habitats (Hays et al., 2014; Runge et al., 2015; Santos et al., 2021), which are often hundreds or thousands of kilometers apart (Schuster et al., 2019), and bound to different jurisdictions and under different anthropogenic pressures (Lascelles et al., 2014; Thornton et al., 2018; Dunn et al., 2019). However, gathering information on the spatial distribution and movements of marine migrants can be difficult, due to the logistical, technological, and economic challenges involved in monitoring these animals over the vast areas that they move (Hussey et al., 2015). Despite recent advances (e.g. Doherty et al., 2017; Hays and Hawkes, 2018; Irvine et al., 2019; Gredzens and Shaver, 2020), our knowledge of the spatial distributions of marine megafauna is geographically biased, in large part due to the unequal availability of research funding (Amano and Sutherland, 2013) leaving areas of the world understudied. West Africa is generally a data-poor region, particularly when it comes to the marine realm (Failler et al., 2020). Nonetheless, marine regions of West Africa, a major upwelling zone (i.e., the Canary current) with rich coastal ecosystems including seagrass meadows, mangroves and tidal flats, are recognized as hosting globally important populations of coastal birds, marine mammals (e.g., Atlantic humpback dolphins and West African manatee), and marine turtles (e.g. Barnett et al., 2004; Cunha and Araújo, 2009; Campredon and Catry, 2016; Pottier et al., 2021).
To protect these critical ecosystems and conserve their biodiversity, several MPAs were established in the region, most from the year 2000 onwards (Failler et al., 2020), followed by the creation of a regional network of MPAs in 2007 called RAMPAO (from the French ‘Réseau Régional d’Aires Marines Protégées en Afrique de l’Ouest’). The RAMPAO network extends across seven West African nations: Cabo-Verde, Mauritania, Senegal, The Gambia, Guinea-Bissau, Guinea, and Sierra Leone (https://rampao.org/). Although the existence of migratory species and shared resources were considered in the establishment of RAMPAO, to date, there is scant empirical evidence that the MPA network contributes to the connectivity of any particular animal population (but see Godley et al., 2010). The green turtle is a marine migratory species of conservation concern (Godley et al., 2020), well represented in the region, which hosts a nesting population among the top six worldwide (SWOT, 2011), with over 25,000 clutches laid annually (Patrício et al., 2019). The core-nesting site is Poilão Island, within the João Vieira - Poilão Marine National Park (JVPMNP), an MPA within the RAMPAO, located in the Bijagós Archipelago of Guinea-Bissau (Catry et al., 2009; Patrício et al., 2019). A previous study tracked the post-breeding migrations of four green turtles from Poilão Island to the National Park of the Banc d’Arguin in Mauritania (PNBA), also in RAMPAO, ca. 1000 km north of Poilão (Godley et al., 2010). This was the first indication of biological connectivity within this network, but sample size was limited, and recent research suggested that this population may migrate to different locations after breeding (Patricio et al., 2019), thus requiring further tracking to elucidate the full seasonal distribution.
Here, we investigate the seasonal distributions of reproductive female green turtles to inform international efforts to conserve this globally important breeding population, as well as other biodiversity, which depends on this protected area network in West Africa. Specifically, we tracked turtles to map their spatial distribution throughout the internesting, migration, and foraging periods in relation to the RAMPAO, and contribute essential baseline information and recommendations for marine spatial planning in the region.
We equipped 45 nesting green turtles with satellite tracking devices from 2018 to 2020 at the João-Vieira Poilão Marine National Park (JVPMNP, Figure 1), in the Bijagós Archipelago, Guinea-Bissau (hereafter “Bijagós”). The park includes four main islands (João Vieira, Cavalos, Meio and Poilão) and an islet (Cabras) where green turtle nesting occurs, with Poilão hosting the great majority of the nests (Patrício et al., 2019). Satellite tags were deployed in Poilão (n=43) and Meio (n=2), either when turtles were laying or when they were stranded in intertidal rock pools in the morning after nesting or attempting to nest, due to the low tide exposing vast areas of rocks preventing turtles from returning to the sea (Patrício et al., 2018). In the first case, we waited for the turtles to start laying eggs, and then executed the attachment procedure within 20 minutes. All turtles continued with their nesting activity throughout the tag application, therefore we are confident that the procedure did not disturb them. In the second case, we approached stranded turtles, started sanding their second vertebral scute and observed their reaction. Most turtles did not react, as if unaware of our presence, thus, we proceeded with attaching tags. If a turtle was startled, we left her undisturbed. We observed all tagged turtles normally returning to the sea with the coming of the high tide. For details on attachment protocol, see Supplemental Methods. The green turtle nesting season at these rookeries extends from July to December (Catry et al., 2002), with few nests year-round (Barbosa et al., 2018). About 80% of the clutches are laid from August to end of October (Barbosa et al., 2018), and we deployed tags from August to November, to cover the period when most turtles are nesting (see Table S1 for details on deployment date). We used two models of Platform Transmitter Terminal (PTT) devices; SPOT-375B from Wildlife Computers, Seattle, Washington (n = 19, 2018), which transmit Argos locations, and F6G - 376B from Lotek, Havelock North, New Zealand (n = 11, 2019; n = 15, 2020), which transmit both Argos and Fast GPS (hereafter ‘FGPS’) location information via the Argos satellite system. Both tag types were set to transmit already collected positions every 15s.
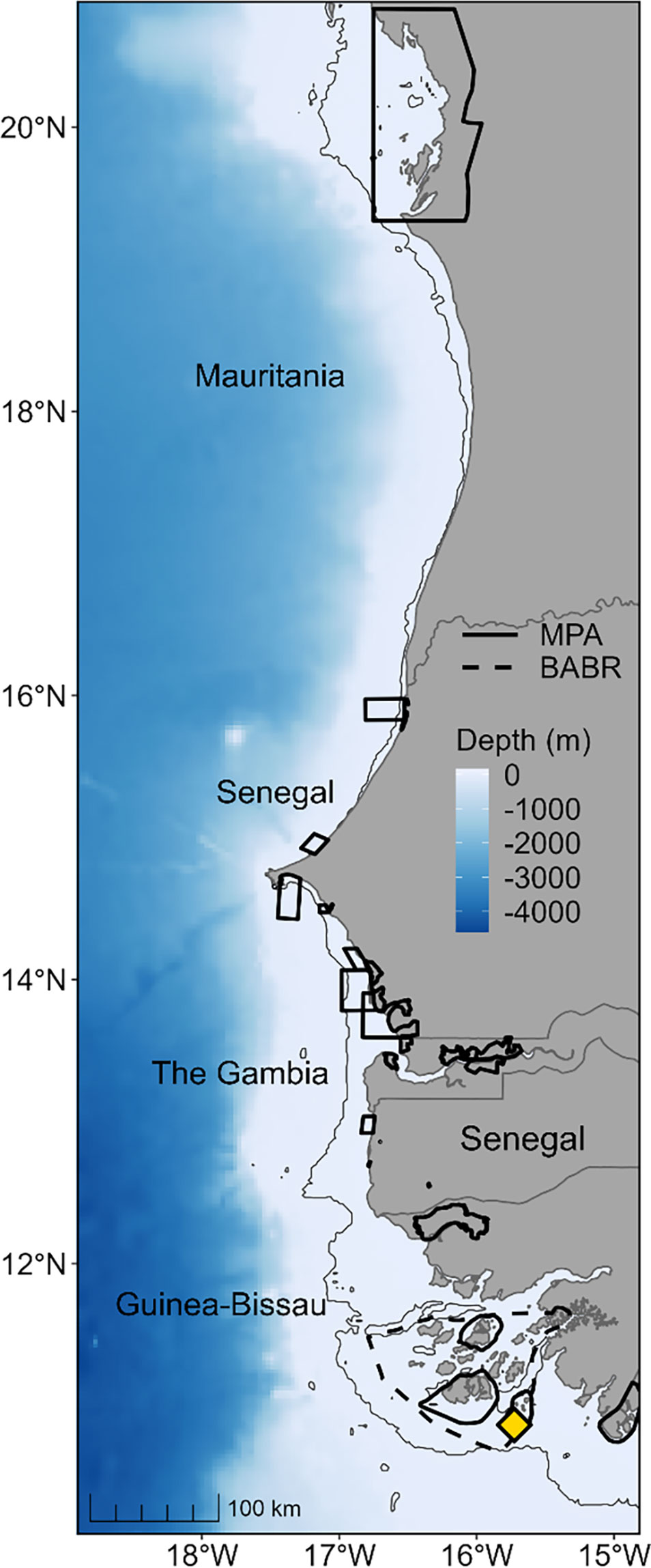
Figure 1 Map of West Africa from the north of Guinea in the south to Mauritania in the north. Solid black polygons show the limits of existing marine protected areas (MPAs). Dashed black line shows the limit of the UNESCO Bolama-Bijagós Archipelago Biosphere Reserve. Yellow diamond indicates the main nesting site of the study population of green turtles (Chelonia mydas); Poilão Island. The bathymetry is shown in a scale of blue, and a thin grey line is the 15 m depth contour.
We split tracking data into the behaviorally distinct seasonal periods of internesting, migration, and post-nesting foraging (hereafter “foraging”) based on close inspection of individual tracks (Figure S1). We classified turtles which stayed in the immediate region around the Bijagós archipelago as near-resident and those which performed long-distance movements (i.e, >100 km from the nesting islands) as migratory. For turtles classified as migrating, we defined the start of migration (i.e., the ‘migration’ portion of a track) as directional and persistent movements at least 10 km from the nesting islands. The 10 km buffer was selected as this is approximately the range in which the turtles spent most time during internesting, so tracking positions outside this area are clearly distinguishable between short forays and long-distance migratory movements. Arrival at the foraging site was identified as localized movements after leaving the internesting area. Foraging turtles did not show stopover behavior (i.e., short stops), and had at least 10 days of localized movements post-nesting. Next, we filtered the raw tracking data to remove positions with high spatial error. For Argos PTT data we retained only positions with a quality index of 1, 2 or 3 (with respective horizontal spatial error radii of <250 m, 250-500 m, and 500-1500 m; Douglas et al., 2012) while for FGPS data we retained positions fixed with 6 or more satellites (Dujon et al., 2014). For data from both sensor types we used a data-driven speed and angle filter (Shimada et al., 2012) to remove biologically unrealistic positions. A total of 28.2% of positions were retained after quality filtering. We used the raw, filtered positions for subsequent analysis, as a sensitivity test showed that interpolation of the data to a regular time interval had little effect on the final outcome (Table S2). For each seasonal period, we retained only individuals with at least 10 tracking locations for analysis.
To estimate how the spatial distribution of adult females during the internesting and foraging periods falls in relation to marine protected areas, we downloaded MPA spatial polygons from the Protected Planet database (UNEP-WCMC, 2021) and calculated: (1) the number of tracked turtles visiting MPAs, and (2) the percentage of tracking positions falling within MPA borders. The average percentage of positions within MPAs was calculated first per individual then across individuals to give an estimate at the population level. The breeding site is located within the UNESCO Bolama-Bijagós Archipelago Biosphere Reserve (BABR, Figure 1). Despite the current lack of any management regulations specific to this reserve area, we additionally assessed the proportion of locations within the BABR limits to demonstrate the potential benefits if relevant protection measures were introduced.
To be able to infer patterns at the population level based on tracking data, it is important to assess the representativeness of the tracked sample of individuals (Sequeira et al., 2019). Therefore, we estimated the representativeness of our samples during the internesting and foraging periods using an established method for informing conservation (Beal et al., 2021b). Briefly, this method identifies how close the space use of the tracked sample of individuals comes to an information asymptote, a point at which most of the population-level space use has been captured. We estimated that our data were 99% (internesting, n = 40) and 76% (foraging, n = 35) representative of the home range used by the wider population (Figures S2, S3).
To assess whether the spatial error of tracking positions affected our calculations regarding MPA coverage, we performed a sensitivity analysis comparing results using only PTT data (which tend to have higher error) against only FGPS data. For the turtles transmitting both PTT and FGPS positions during internesting (n = 21) and foraging (n = 20) respectively, we compared the percentage of points within and outside MPAs separately for each geolocation method. We found little difference in the percentage of positions within MPAs between the geolocation methods, suggesting our filters removed positions with large enough error to affect MPA overlap (Table S3).
To visualize the home ranges and core areas of the tracked turtles during internesting and foraging, we used kernel density estimation (KDE). KDE is a non-parametric technique for smoothing point densities, and can be used to derive probabilistic animal home ranges known as utilization distributions (UD) based on location data (Worton, 1989). The UD of a tracked animal represents space use intensity. An important step when running KDE is determining the smoothing parameter for analysis, which affects the scale of the resulting spatial estimate (Gitzen et al., 2006). We calculated the canonical reference bandwidth for each individual, which reflects the number and spatial variance of tracking positions, and then took the median across individuals as the parameter value for analysis (Beal et al., 2021b). Given that the movement behavior of female green turtles differs between internesting and foraging periods, we made this calculation separately for each period resulting in a smoothing parameter value of 0.9 km for internesting data and 2.1 km for foraging data. To get an estimate of the population-level distribution for each period, we averaged together the individual UDs.
To estimate migration routes for each turtle and identify population-level high-passage corridors, we used dynamic Brownian bridge movement models (dBBMM). Brownian bridge movement models estimate the probability of occurrence of animals in space during the tracking period by using a conditional random walk to approximate potential movement paths connecting tracking positions, while taking into account the time gaps between them. Brownian bridge models are able to handle irregular sampling intervals and incorporate positional errors into the occurrence estimation. dBBMMs extend classic Brownian bridge models by detecting behavioral changes within “moving windows” along the movement path and dynamically assigning appropriate Brownian motion variance values (σ2m) to the random walk procedure. This allows for more accurate estimations of occurrence over lengthy tracking periods, during which the movement behavior of animals may change (Kranstauber et al., 2012).
Following Buechley et al., 2018, we estimated migratory corridors for the adult female green turtle population by calculating the percentage of migration routes (i.e., tracked turtles) overlapping across a gridded surface. A ‘migration route’ was defined as the 95% occurrence probability as estimated by the dBBMM, fitted using filtered locations and their estimated horizontal spatial error for each individual turtle. To estimate the Brownian motion variance, we used a sliding window of seven locations with a margin of three locations. We used a grid cell resolution of 10 x 10 km, which is large enough to identify similar route selection among the turtles, while remaining of relevance for informing spatial protection measures, such as MPA border extension or temporal fisheries restrictions in RAMPAO, where MPAs cover a median of 200 km2 (range 0.5 – 12,006 km2). Finally, we identified “high-passage corridors” as grid cells visited by at least 25% of the turtles which migrated north of the Bijagós after nesting. We then calculated the percentage of high passage cells falling within established MPAs, when the center of a high turtle passage grid cell (i.e., cells through which ≥ 25% of turtles likely passed) fell inside an MPA border, the MPA was considered as covering that cell.
We obtained spatial data from 44 of the 45 tagged individuals. Not all 44 turtles contributed data for the three seasonal periods analyzed (Table S1 and Figure S1), due to premature device failure, to a lack of good locations retained after filtering, to nearly resident foraging behavior (i.e., no migratory period) or to leaving the breeding area immediately after tag deployment (i.e., no data on internesting period). After filtering, we retained for analysis the tracks of 40 individuals during internesting (88.9%, n=45), 18 individuals during migration (48.9%, n=45), and 35 during foraging (77.8%, n=45). Of the 40 turtles that contributed with data for the internesting period, 18 were re-sighted at the beach during multiple nesting events/attempts, thus we are confident that they were in their breeding period (see Table S4). In total, we analyzed 4,358 tracking days, with each individual tracked between 15 - 332 days (mean 99 days ± SD 73 days). Mean curved-carapace-length of tagged turtles was 98.5 cm (SD 8.9 cm, range: 78.0 – 121.5), with no significant differences in curved-carapace-length between years (ANOVA: F1,42 = 0.383, p = 0.540).
The green turtles tracked in this study were distributed between 10.5°N and 20.5°N, an area within which there are 28 MPAs (Figure 1 and Table S5).
Tracking during the internesting period extended from August to early December (Figures S1, S4). We found that the 95% home ranges and 50% core-use areas of individual turtles during internesting were highly concentrated around their respective nesting islands (Figure 2). The spatial distribution of breeding females was well covered by MPAs during internesting, with all 40 individuals using MPAs and a mean of 94.8% (SD 0.1%, range: 46% - 100%) of tracking positions falling within MPAs. The mean percent coverage increased to 99.5% (SD 0.01%) of positions when considering the limits of the BABR.
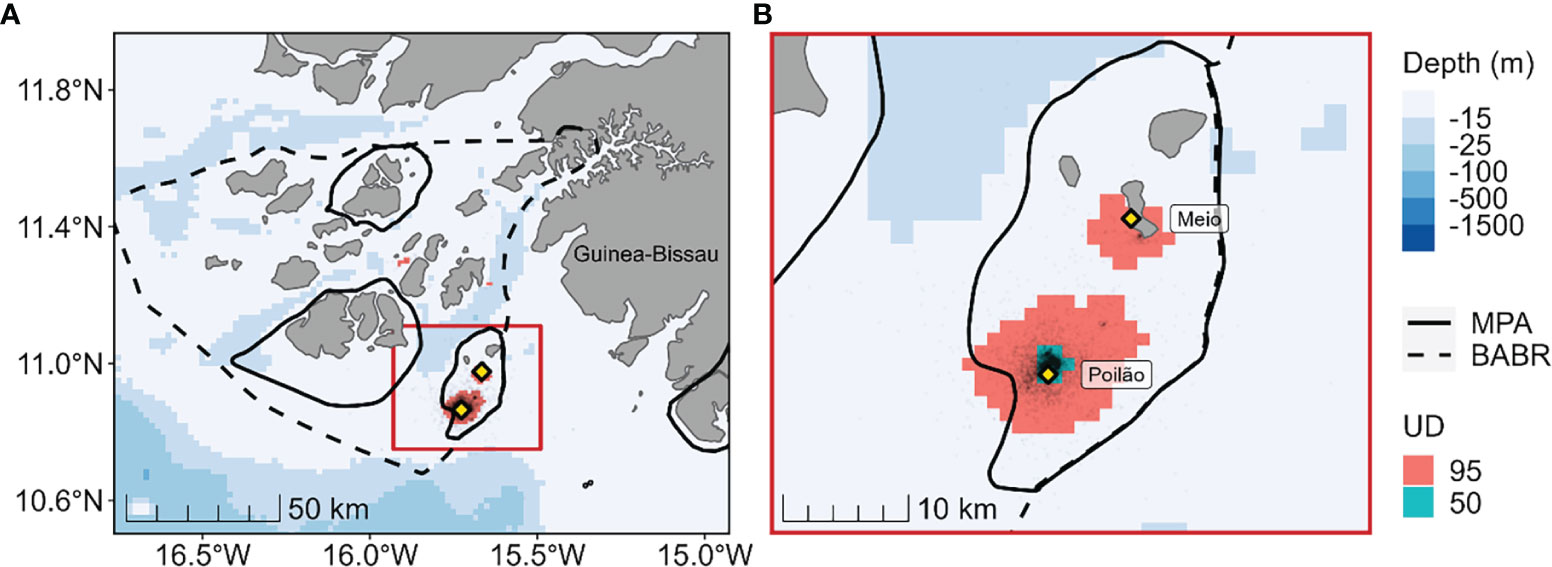
Figure 2 (A) Distribution of 40 female green turtles (Chelonia mydas) tracked during internesting. Yellow diamonds indicate the nesting sites where satellite tags were deployed. The 95% (red) and 50% (blue-green) utilization distribution areas are shown, respectively signifying the population-level home range and core-use areas during internesting. The bold solid lines show the limits of existing marine protected areas (MPAs) and the dashed line shows the extent of the Bolama-Bijagós Archipelago Biosphere Reserve (BABR). (B) Inset showing close-up of distribution around the nesting sites.
The migration period of the tracked turtles extended from September to December (Figure S1). The routes of the 18 individuals which migrated north of the Bijagós followed the shoreline for the most part, with some individuals travelling several hundred kilometers offshore (Figures 3, S5). The areas of highest migration route concentration were used by 61% (11 individuals) of migrating turtles. With 25% of turtles (i.e., 5 individuals) set as a minimum threshold to define high-passage migration corridors, we identified three main coastal stretches as particularly important: from the outer Bijagós to The Gambia, off the city of Dakar in Senegal, and along the south of Mauritania to the south of the Banc d’Arguin (Figure 3). We calculated that RAMPAO MPAs overlapped 18 of the 84 (21%) high-passage corridor cells identified in our analysis. If the BABR area was included, the coverage rose to 30 of 84 (36%) of high-passage cells.
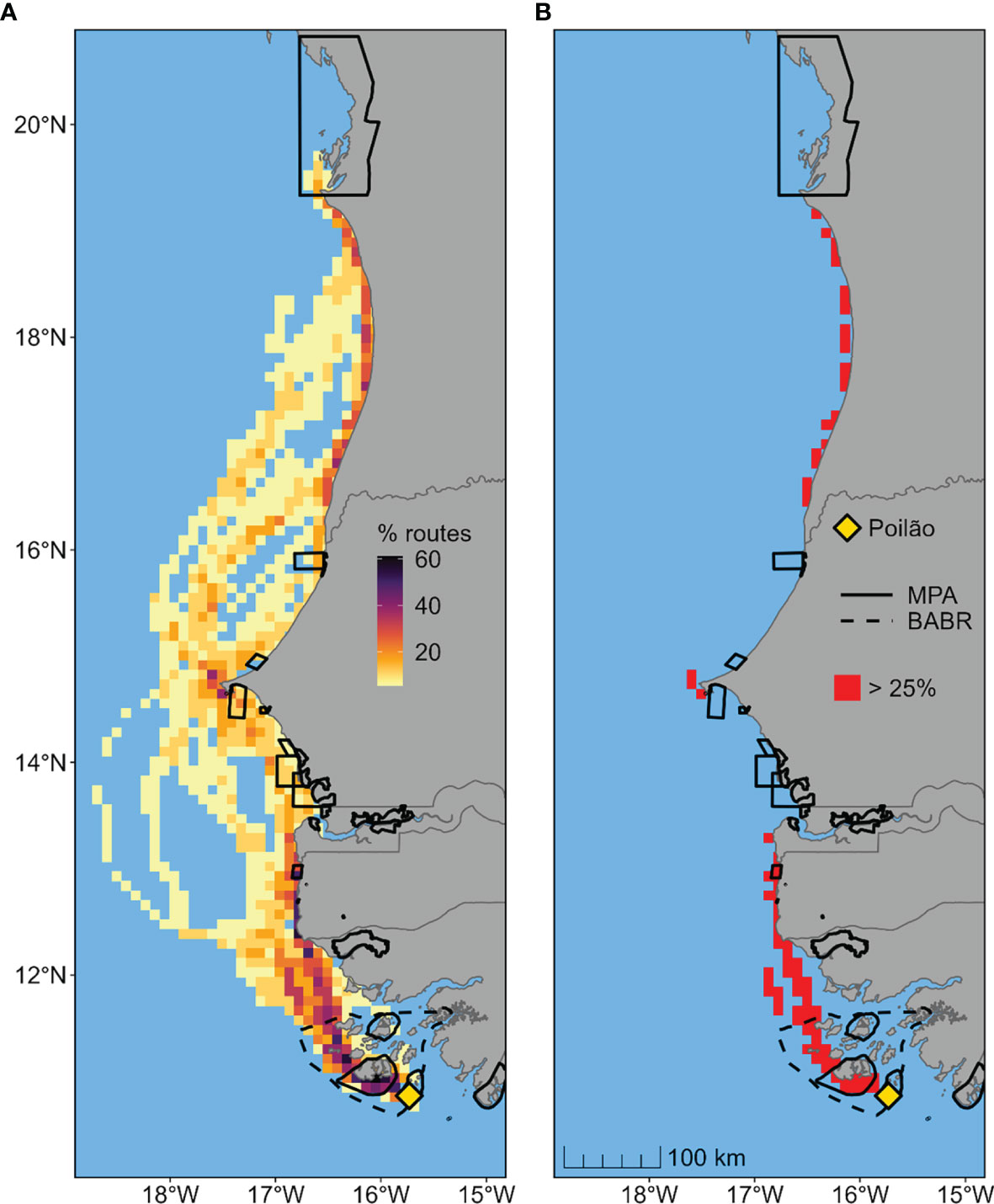
Figure 3 Migration routes of 18 female green turtles (Chelonia mydas) travelling from the nesting island in Guinea-Bissau (i.e., Poilão Island) to foraging areas in Senegal, The Gambia or Mauritania. (A) Percentage of overlapping migration routes, defined as 95% probability areas estimated using dynamic Brownian bridge movement models. (B) Identified high-passage corridors, defined as grid cells used by at least 25% of migrating turtles. Black solid and dotted lines represent existing conservation areas in the region. (Marine Protected Areas [MPA] and UNESCO Bolama-Bijagós Biosphere Reserve [BABR]).
The nesting population of the Bijagós displayed variable migratory strategies, with 12 turtles remaining resident in the Bolama-Bijagós region (34.2%, n=35), foraging at 40-90 km from the breeding sites, including one that relocated ca. 80km southeast of Poilão Island, 10 migrating 300-400 km to The Gambia and Senegal (28.6%, n=35), and 13 migrating >1000 km to the north of Mauritania (37.1%, n=35, Figure S6). The turtles analyzed here engaged in foraging behavior (i.e., localized movements) for a range of 10 to 288 days (Table S1 and Figure S1) before device failure, and no turtle that migrated north of the Bijagós (i.e. non-resident) stopped to forage within the archipelago.
The 95% home ranges and 50% core-use areas of individual turtles during the foraging period were more dispersed than during internesting (Figure 4). During the foraging period, we found lower MPA coverage than during internesting, with 26 of 35 individuals (74%) using MPAs at some point (i.e., at least 3 locations within an MPA, Table 1). Of the individuals using MPAs, a mean of 78.0% (SD 34.8%, range: 3.7% - 100%) of their tracking positions fell within protected area borders. Disaggregating this result by foraging destination, a mean of 93.3% (SD 16.5%) of positions from turtles which migrated to Mauritania fell within MPAs, 83.9% (SD 28.7%) of positions of those which travelled to The Gambia and Senegal, and 15.1% (SD 20.7%) for turtles which remained in the wider Bijagós region (Table 1 and Figure 4). With the BABR included in the coverage calculation, the percentage of tracking positions occurring within all conservation areas for post-nesting turtles in the Bijagós increased to 99.3% (SD 0.008%).
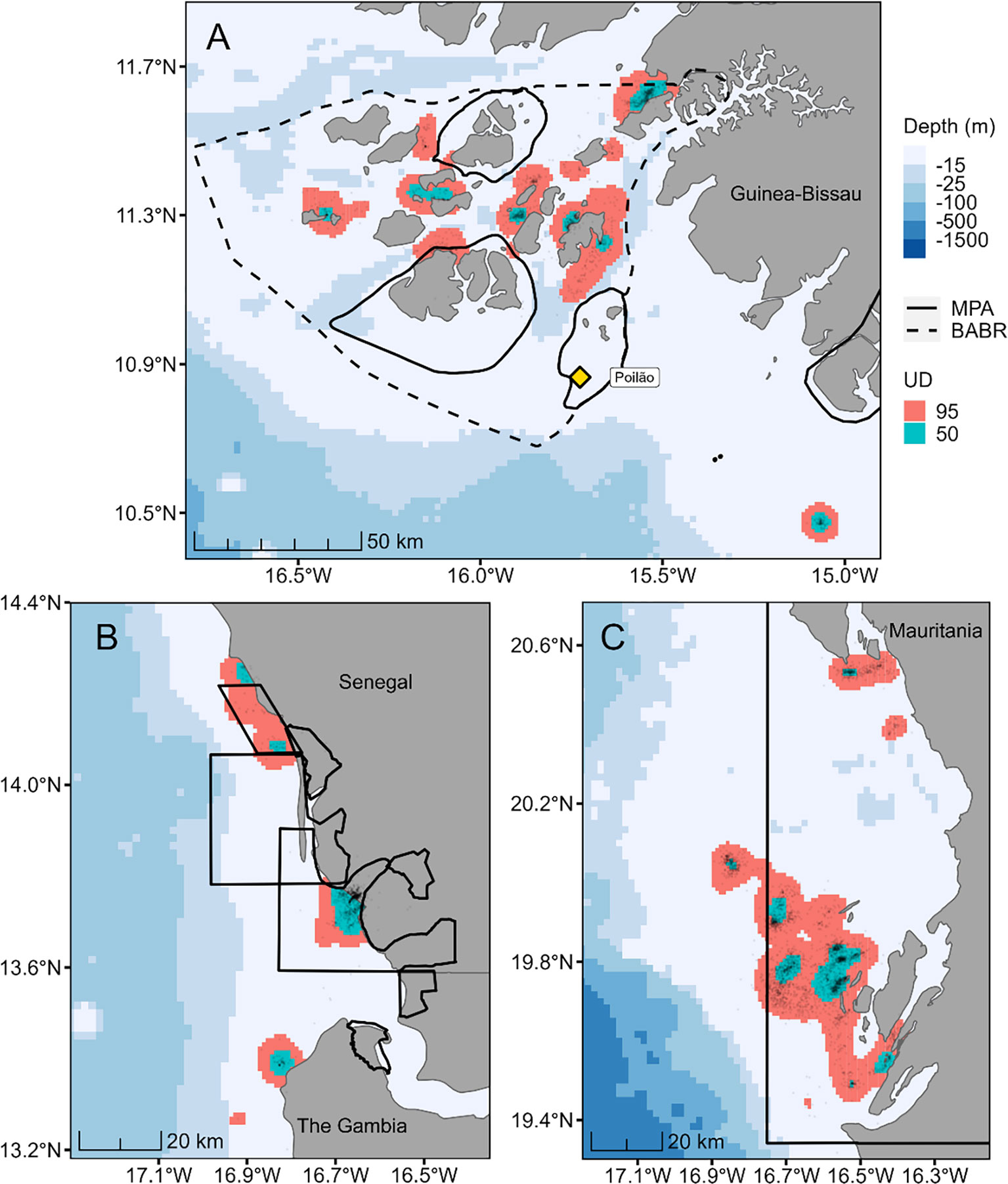
Figure 4 Distribution of 35 female green turtles (Chelonia mydas) tracked during the post-nesting foraging period in (A) the Bijagós Archipelago of Guinea-Bissau and south of the Bijagós (n = 12), (B) the Saloum and Gambia river deltas of Senegal and The Gambia (n = 10) and (C) the Bay of Arguin in Mauritania (n = 13). Black dots represent tracking locations, and the underlying colored areas represent the 95% (pink) and 50% (blue-green) utilization distribution areas, respectively signifying the population-level home range and core-use areas during foraging. The bold solid lines indicate the location of existing marine protected areas (MPAs) and the dashed line shows the extent of the BABR biosphere reserve.

Table 1 Coverage of adult female turtles during the post-nesting foraging period in West Africa by marine protected areas (MPA).
Assessing the seasonal distributions of migratory species is a key step to informing marine spatial planning and to identifying opportunities for cross-regional collaboration for conservation (Beal et al., 2021a). By analyzing the movements of reproductive female green turtles across three seasonal periods, we provide novel information on the spatial distribution of this globally significant rookery, which is highly relevant to regional conservation and management efforts. Using tracking data, we identified multiple post-nesting migratory strategies, novel foraging sites, and high-density migration corridors. We then quantified the spatial coverage of seasonally important areas by the West Africa MPA network, RAMPAO. Our findings highlight the biological relevance of the RAMPAO, and its importance for the conservation of this charismatic species and wider biodiversity in the region. We provide recommendations for MPA managers, to improve both the protection coverage and knowledge of this important population.
During the internesting period, breeding females concentrated near the nesting beaches, remaining within the limits of the João Vieira – Poilão Marine National Park (JVPMNP). This supports findings by Godley et al. (2010), based on the internesting movements of seven turtles tracked in 2001. Indeed, the protection of this globally significant rookery was one of the main motivations for the creation of this MPA (Catry et al., 2002, Table 2), but conservation efforts are mostly concentrated on the terrestrial phase (i.e. nesting and egg incubation), particularly at Poilão Island. Illegal fishing is known to occur inside the MPA (186 seized fishing boats from 2011 to 2019, Catry et al., 2018), with fishers mostly from neighboring countries (ca. 62%, Senegal, Guinea, Ghana, Sierra Leone, Catry et al., 2018), where marine turtles have commercial value (notably not the case in Guinea-Bissau, Barbosa et al., 2018). Still, marine surveillance at the JVPMNP and enforcement of fines, contribute to deter most illegal activities, and enhance protection during the nesting season.
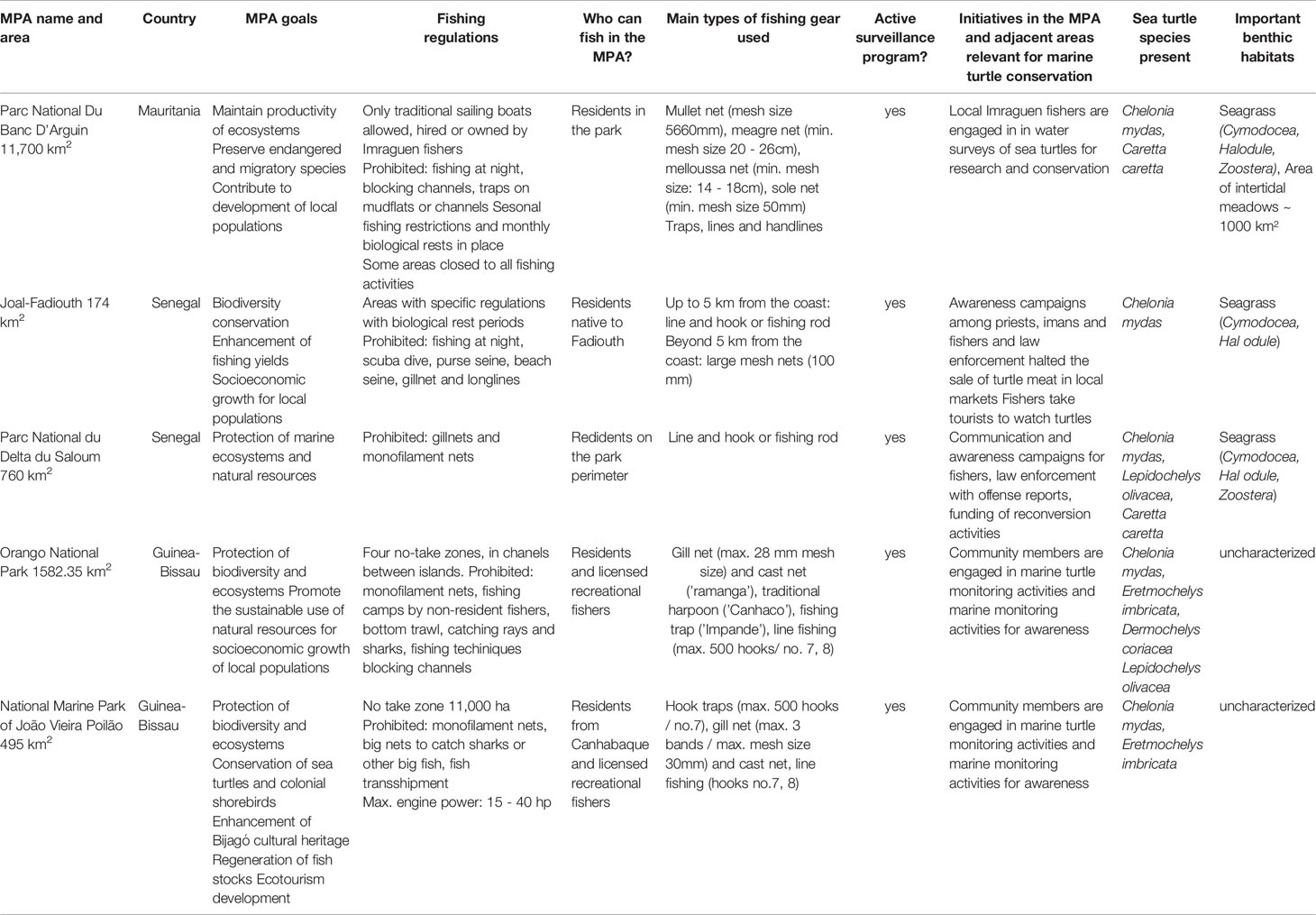
Table 2 Summary of relevant management characteristics of five marine protected areas (MPAs) of the West Africa MPA network (RAMPAO), used by the green turtle population breeding in the Bijagós Archipelago, Guinea-Bissau.
Small home ranges and high MPA coverage during internesting have been found among other green turtle populations, e.g., at the Chagos Archipelago, Indian Ocean (Hays et al., 2014) and in the Gulf of Mexico, south Florida, USA (Hart et al., 2013). This was also observed among hawksbill turtles from the Solomon Islands, West Pacific, spending 98.5% of their internesting time within the Arnavon Community Marine Park (Hamilton et al., 2021), and from the Dominican Republic, Caribbean Sea, with 82.7% of their internesting time within the Del Este National Park (Revuelta et al., 2015). Further studies reported restricted home ranges during the internesting period among loggerhead (Schofield et al., 2010) and green turtles (Snape et al., 2018; Shimada et al., 2021). Thus, implementing relevant measures around rookeries of conservation value (e.g. protected areas, fishing restrictions) may be a good strategy to protect vulnerable populations during internesting (Dryden et al., 2008).
Turtles passed through several RAMPAO MPAs during the migration period, but overall, MPA coverage during migration was relatively low (21%), with a large portion of coverage provided by the Orango National Park (ONP), located near the nesting islands in the Bijagós. Most turtles followed a similar path after leaving the nesting grounds potentially because they are using the same local environmental cues, such as ocean currents (Hays et al., 2010), wave-direction (Papi et al., 2000) or coastlines (Lohmann et al., 2008), to orient themselves. Yet some individuals diverted this corridor, and other populations display highly dispersed post-nesting migratory paths (e.g. Hays et al., 2014). Similar to our results, high-passage migratory corridors near the nesting beach have been described in several marine turtle populations (Pendoley et al., 2014; Stokes et al., 2015; Hamilton et al., 2021; Santos et al., 2021), as well as near foraging locations (Baudouin et al., 2015; Stokes et al., 2015; Santos et al., 2021), suggesting that these areas can be effective places to implement conservation measures.
The majority of the turtles that migrated to Mauritania converged around the Cap-Vert Peninsula (where the city of Dakar is situated). This prominent promontory acts as a migratory bottleneck (sensu Bayly et al., 2018), funneling huge numbers of migratory seabirds, which are compelled by the promontory to approach the shore (Dubois et al., 2009; Péron and Grémillet, 2013; Paiva et al., 2015). Given our findings, combined with the apparent importance of this site for other taxa, increasing spatial protection here may have a broad impact on biodiversity in the region. The National Park of the Madeleine Islands is a terrestrial only protected area located just offshore of the peninsula in an area of high turtle passage, suggesting that extension of the limits of the park to the marine zone would increase protection coverage for migrating green turtles. As migration is restricted to a well-defined period of the year (e.g. Hart et al., 2019; Santos et al., 2021; September to December in our study), migratory bottlenecks are seasonal and therefore temporal measures in these areas could be effective. We recommend implementing seasonal, spatially explicit bans on fishing practices that cause turtle bycatch.
We found that after nesting, females had three distinct migratory strategies, with some turtles remaining resident in the Bolama – Bijagós region and others performing short or long migrations to foraging grounds in other parts of West Africa. Non-migratory green turtles have also been reported from the Cocos Keeling Islands, in the eastern Indian Ocean (Whiting et al., 2008), and a similar pattern of both near-residents and migratory individuals was observed among green turtles in the Indian Ocean (Hays et al., 2014), and in the Caribbean (Esteban et al., 2015).
We found that this nesting population from the Bijagós connects at least five RAMPAO MPAs, across three countries, which have a marine area with fishing regulations and active marine surveillance programs (Table 2). Overall, the MPA coverage of foraging areas was good (over 70%), but it varied greatly between the foraging regions. At the Bay of Arguin, in Mauritania, most of the foraging occurred within the Banc d’Arguin National Park (PNBA). In Senegal, a very high proportion of the foraging period was spent within the limits of either the Saloum Delta National Park (SDNP) or the Joal Fadiouth MPA (JFMPA). Some turtles foraged around the Bijol Islands of The Gambia, where green turtle nesting is also known to occur (Barnett et al., 2004), and although these islands are part of the Reserve of the Tanji Bijol Island Banks, this MPA does not currently include a marine area. Extending the limits of this reserve to include nearshore waters would enhance protection of both the turtles which nest on the Bijol Islands during internesting, as well as turtles from the Bijagós during foraging. In the Bijagós, the majority of turtles foraged outside MPAs (88%, n=12), but within the limits of the UNESCO Bolama-Bijagós Archipelago Biosphere Reserve (BABR, 98.1%). Notably, the BABR also hosts foraging aggregations of immature green turtles (Monteiro et al., 2021), therefore, creating and implementing relevant regulations within the BABR would greatly enhance the protection of green turtles in West Africa. In this study, our tracking effort was focused during the time of year when the large majority of the Bijagós green turtle population nest, as this is the most relevant period for informing for conservation. However, the tracking data obtained here might not represent the full variability of post-nesting migrations and foraging grounds used by this population. For example, it is possible that the few green turtles that nest outside of the principal nesting season use different foraging areas, a point which may be investigated in the future.
Foraging habitats were not uniformly distributed along the West African coast, which could suggest that food resources are patchy. The PNBA of Mauritania, and the SDNP and JF MPA of Senegal, have vast areas of seagrass (Cymodocea nodosa, Halodule wrightii, Zoostera noltii, Vermaat et al., 1993, Cunha and Araújo, 2009), which is one of the food items most consumed by green turtles, globally (Esteban et al., 2020). At the PNBA, green turtles were found to consume seagrasses, along with animal prey, with larger turtles relying more on seagrass than smaller juveniles (Cardona et al., 2009), thus, the distribution of this resource could in part explain the locations of turtle foraging areas. However, although Cunha and Araújo (2009) found the same species of seagrasses along the unprotected stretch of coast from the south of the Bay of Arguin to Nouakchott, Mauritania, none of the turtles we tracked foraged in this area, suggesting that other factors influence their choice of foraging area. An alternative explanation is that the protection afforded by MPAs is defining the spatial distribution of this population. In the Bijagós seagrass is scarce (Campredon and Catry, 2016), although more benthic surveys are needed for a thorough characterization of the foraging areas in the archipelago. A study using DNA metabarcoding from esophageal swabs of immature green turtles foraging in the westernmost islands of the Bijagós archipelago, also used by adult female green turtles as revealed by our tracking data, identified red algae as their main food source (Rhodymeniophycidae subclass, Díaz-Abad et al., 2022). Seagrass was also detected in the diet, along with invertebrates, but in smaller proportions (Díaz-Abad et al., 2022). Green turtles foraging in São Tomé, Gulf of Guinea, also consumed more macroalgae (mostly red algae Polysiphonia sp., Hancock et al., 2018) than seagrass, which was less abundant (Hancock et al., 2018). Future studies will further investigate the diet of green turtles in West Africa, across their main foraging areas.
Identifying green turtles as a common conservation target for several of the RAMPAO MPAs can motivate and inform collaboration. We recommend that stakeholders from each interested country together define standard monitoring parameters for systematic data collection to increase the knowledge of this globally important population, and reinforce information exchange to support conservation actions and management. Additionally, for a complete picture of the MPA coverage and connectivity of this breeding population, future research should address the spatial distribution of reproductive males. Spatial analyses should also extend to other taxa, to maximize effective MPA design encompassing key ecosystems and a broad range of species of ecological and conservation value.
This study unequivocally demonstrates the importance of an established MPA network, the RAMPAO, for the conservation of a marine migratory species. Ongoing research indicates that connectivity across the RAMPAO is also established by migratory coastal waders (Rocha pers comm). Notably, the green turtle rookery at Poilão Island is growing (Broderick and Patrício, 2019), which can be a possible indication that, despite existing threats (Catry et al., 2002), RAMPAO MPAs are effectively protecting this population. We argue that lessons can be learned from the RAMPAO process, and applied globally. West African countries (and other countries around the world) are well below the previously set 2020 Aichi Target 11 of 10% of marine areas under nature protection (Failler et al., 2020). Therefore, studies of this kind provide highly relevant information to countries of ways they can expand their protected area estate to both meet the targets of the upcoming post-2020 Biodiversity Framework and contribute to international conservation efforts (Hays et al., 2019).
All analysis scripts and data derivatives produced in this study are available for download at https://doi.org/10.5281/zenodo.6346610. Tracking data analysed here may be found on Movebank (https://www.movebank.org/cms/movebank-main) using the accession numbers 1988749082 and 1988757157 and 1988638748. Further inquiries can be directed to the corresponding authors.
The animal study was reviewed and approved by Orgão Responsável pelo Bem-estar Animal do ISPA (ORBEA - ISPA). Research permits were granted by the Instituto da Biodiversidade e das Áreas Protegidas, Dr. Alfredo Simão da Silva (IBAP), of Guinea-Bissau. Fieldwork protocols involving sea turtles followed recommended guidelines (NMFS-SFC, 2008), to enhance efficiency and reduce possible disturbance to the turtles, and were conducted by trained personnel, in collaboration with national authorities for biodiversity management.
ARP, FM, CB and PC conducted fieldwork. ARP, PC, MB and BG contributed to conception and design of the study. MB conducted spatial analysis. ARP and MB wrote the first draft of the manuscript. PC, BG wrote sections of the manuscript. CB, AR, CS and ES helped with fieldwork logistics and permits. DD and MT contributed with crucial information on marine protected areas in the region. All authors contributed to manuscript revision, read, and approved the submitted version.
This study was funded by: the MAVA Foundation through the projects “Consolidation of sea turtle conservation at the Bijagós Archipelago, Guinea-Bissau” and “Tortue d’Arguin”; the Regional Partnership for Coastal and Marine Conservation (PRCM), through the project “Survies des Tortues Marines”; the “La Caixa” Foundation (ID 100010434) through a fellowship awarded to ARP (LCF/BQ/PR20/11770003); the Fundação para a Ciência e a Tecnologia, Portugal, through a grant (UIDB/04292/2020 and UIDP/04292/2020) awarded to MARE; the European Union’s Horizon 2020 Research and Innovation Programme under the Marie Skłodowska-Curie grant agreement no.766417 - this communication reflects only the authors’ view, and the Research Executive Agency of the European Union is not responsible for any use that may be made of the information it contains.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Research permits and logistic support were provided by the Instituto da Biodiversidade e das Áreas Protegidas, Dr. Alfredo Simão da Silva (IBAP). We conducted fieldwork in collaboration with IBAP technicians and IBAP rangers, and locals from the villages of Ambeno and Bine, on Canhabaque Island, Bijagós, and we are deeply thankful to all. This manuscript benefited from the comments of two reviewers, to whom we are thankful.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.812144/full#supplementary-material
Amano T., Sutherland W. J. (2013). Four Barriers to the Global Understanding of Biodiversity Conservation: Wealth, Language, Geographical Location and Security. Proc. R. Soc. B 280, p.20122649. doi: 10.1098/rspb.2012.2649
Assis J., Fragkopoulou E., Serrão E. A., e Costa B. H., Gandra M., Abecasis D. (2021). Weak Biodiversity Connectivity in the European Network of No-Take Marine Protected Areas. Sci. Total Environ. 773, 145664. doi: 10.1016/j.scitotenv.2021.145664
Barbosa C., Patrício R., Ferreira B., Sampaio M., Catry P. (2018). “Tartarugas Marinhas,” in Parque Nacional Marinho João Vieira E Poilão: Biodiversidade E Conservação. Eds. Catry P., Regalla A. (Bissau: IBAP – Instituto da Biodiversidade e das Áreas Protegidas).
Barnett L. K., Emms C., Cham A. M., Mortimer J. A. (2004). The Distribution and Conservation Status of Marine Turtles in The Gambia, West Africa: A First Assessment. Oryx 38, 203–208. doi: 10.1017/S0030605304000353
Baudouin M., de Thoisy B., Chambault P., Berzins R., Entraygues M., Kelle L., et al. (2015). Identification of Key Marine Areas for Conservation Based on Satellite Tracking of Post-Nesting Migrating Green Turtles (Chelonia mydas). Biol. Conserv. 184, 36–41. doi: 10.1016/j.biocon.2014.12.021
Bayly N. J., Rosenberg K. V., Easton W. E., Gomez C., Carlisle J. A. Y., Ewert D. N., et al. (2018). Major Stopover Regions and Migratory Bottlenecks for Nearctic-Neotropical Landbirds Within the Neotropics: A Review. Bird Conserv. Int. 28, 1–26. doi: 10.1017/S0959270917000296
Beal M., Dias M. P., Phillips R. A., Oppel S., Hazin C., Pearmain E. J., et al. (2021a). Global Political Responsibility for the Conservation of Albatrosses and Large Petrels. Sci. Adv. 7, eabd7225. doi: 10.1126/sciadv.abd7225
Beal M., Oppel S., Handley J., Pearmain E. J., Morera-Pujol V., Carneiro A. P. B., et al. (2021b). Track2kba: An R Package for Identifying Important Sites for Biodiversity From Tracking Data. Methods Ecol. Evol. doi: 10.1111/2041-210X.13713
Broderick A., Patricio A.. (2019). Chelonia Mydas (South Atlantic Subpopulation). The IUCN Red List of Threatened Species 2019: e.T142121866A142086337. doi: 10.2305/IUCN.UK.2019-2.RLTS.T142121866A142086337.en
Buechley E. R., Oppel S., Beatty W. S., Nikolov S. C., Dobrev V., Arkumarev V., et al. (2018). Identifying Critical Migratory Bottlenecks and High-Use Areas for an Endangered Migratory Soaring Bird Across Three Continents. J. Avian Bio. 49, e01629. doi: 10.1111/jav.01629
Campredon P., Catry P. (2016). Bijagos Archipelago (Guinea-bissau). The Wetland Book. II. Distribution, Description and Conservation (Dordrecht:Springer), 1–8.
Cardona L., Aguilar A., Pazos L. (2009). Delayed Ontogenic Dietary Shift and High Levels of Omnivory in Green Turtles (Chelonia Mydas) From the NW Coast of Africa. Mar. Biol. 156 (7), 1487–1495. doi: 10.1007/s00227-009-1188-z
Catry P., Barbosa C., Indjai B., Almeida A., Godley B. J., Vié J. C. (2002). First Census of the Green Turtle at Poilão, Bijagós Archipelago, Guinea-Bissau: The Most Important Nesting Colony on the Atlantic Coast of Africa. Oryx 36, 400–403. doi: 10.1017/S0030605302000765
Catry P., Barbosa C., Paris B., Indjai B., Almeida A., Limoges B., et al. (2009). Status, Ecology, and Conservation of Sea Turtles in Guinea-Bissau. Chelonian Conserv. Biol. 8, 150–160. doi: 10.2744/CCB-0772.1
Catry P., Pires A. J., Barbosa C., Cordeiro J. S., Tchantchalam Q., Regalla A. (2018). “Ameaças E Conservação,” in Parque Nacional Marinho João Vieira E Poilão: Biodiversidade E Conservação. Ed. Catry P., Regalla A. (Bissau: IBAP – Instituto da Biodiversidade e das Áreas Protegidas).
Cunha A. H., Araújo A. (2009). New Distribution Limits of Seagrass Beds in West Africa. J. Biogeogr. 36, 1621–2. doi: 10.1111/j.1365-2699.2009.02135.x
Díaz-Abad L., Bacco-Mannina N., Madeira F. M., Neiva J., Aires T., Serrao E. A., et al. (2022). eDNA Metabarcoding for Diet Analyses of Green Sea Turtles (Chelonia Mydas). Mar. Biol. 169, 1–12.
Doherty P. D., Baxter J. M., Godley B. J., Graham R. T., Hall G., Hall J., et al. (2017). Testing the Boundaries: Seasonal Residency and Inter-Annual Site Fidelity of Basking Sharks in a Proposed Marine Protected Area. Biol. Conserv. 209, 68–75. doi: 10.1016/j.biocon.2017.01.018
Douglas D. C., Weinzierl R., C. Davidson S., Kays R., Wikelski M., Bohrer G. (2012). Moderating A Rgos Location Errors in Animal Tracking Data. Methods Ecol. Evol. 3 (6), 999–1007. doi: 10.1111/j.2041-210X.2012.00245.x
Dryden J., Grech A., Moloney J., Hamann M. (2008). Rezoning of the Great Barrier Reef World Heritage Area: Does It Afford Greater Protection for Marine Turtles? Wildl. Res. 35, 477–485. doi: 10.1071/WR07087
Dubois J. P., Holmström N., Verneau A. (2009). La Péninsule Du Cap-Vert a Dakar, Sénégal, Est-Elle La ‘‘Mecque’’ Du Seawatching. Ornithos 4, 220–232.
Dujon A. M., Lindstrom R. T., Hays G. C. (2014). The Accuracy of Fastloc-GPS Locations and Implications for Animal Tracking. Methods Ecol. Evol. 5, 1162–1169. doi: 10.1111/2041-210X.12286
Dunn D. C., Harrison A. L., Curtice C., DeLand S., Donnelly B., Fujioka E., et al. (2019). The Importance of Migratory Connectivity for Global Ocean Policy. Proc. R. Soc B 286, 20191472. doi: 10.1098/rspb.2019.1472
Esteban N., Mortimer J. A., Stokes H. J., Laloë J. O., Unsworth R. K., Hays G. C. (2020). A Global Review of Green Turtle Diet: Sea Surface Temperature as a Potential Driver of Omnivory Levels. Mar. Biol. 167, 1–17. doi: 10.1007/s00227-020-03786-8
Esteban N., van Dam R. P., Harrison E., Herrera A., Berkel J. (2015). Green and Hawksbill Turtles in the Lesser Antilles Demonstrate Behavioural Plasticity in Inter-Nesting Behaviour and Post-Nesting Migration. Mar. Biol. 162, 1153–1163. doi: 10.1007/s00227-015-2656-2
Failler P., Touron-Gardic G., Traoré M. S., Phang S. C. (2020). Evaluating the Official Achievement of Aichi Target 11 for West African Countries: A Twofold Challenge of Accuracy and Catching-Up. Sci. Total Environ. 698, 134284. doi: 10.1016/j.scitotenv.2019.134284
Gaines S. D., White C., Carr M. H., Palumbi S. R. (2010). Designing Marine Reserve Networks for Both Conservation and Fisheries Management. Proc. Natl. Acad. Sci. 107, 18286–18293. doi: 10.1073/pnas.0906473107
Gitzen R. A., Millspaugh J. J., Kernohan B. (2006). Bandwidth Selection for Fixed-Kernel Analysis of Animal Utilization Distributions. J. Wildl. Manage. 70, 1334–1344. doi: 10.2193/0022-541X(2006)70[1334:BSFFAO]2.0.CO;2
Godley B. J., Barbosa C., Bruford M., Broderick A. C., Catry P., Coyne M. S., et al. (2010). Unravelling Migratory Connectivity in Marine Turtles Using Multiple Methods. J. Appl. Ecol. 47, 769–778. doi: 10.1111/j.1365-2664.2010.01817.x
Godley B. J., Broderick A. C., Colman L. P., Formia A., Godfrey M. H., Hamann M., et al. (2020). Reflections on Sea Turtle Conservation. Oryx 54, 287–289. doi: 10.1017/S0030605320000162
Gredzens C., Shaver D. J. (2020). Satellite Tracking can Inform Population-Level Dispersal to Foraging Grounds of Post-Nesting Kemp’s Ridley Sea Turtles. Front. Mar. Sci. 7, 559. doi: 10.3389/fmars.2020.00559
Hamilton R. J., Desbiens A., Pita J., Brown C. J., Vuto S., Atu W., et al. (2021). Satellite Tracking Improves Conservation Outcomes for Nesting Hawksbill Turtles in Solomon Islands. Biol. Conserv. 261, 109240. doi: 10.1016/j.biocon.2021.109240
Hancock J. M., Vieira S., Jimenez V., Rio J. C., Rebelo R. (2018). Stable Isotopes Reveal Dietary Differences and Site Fidelity in Juvenile Green Turtles Foraging Around São Tomé Island, West Central Africa. Mar. Ecol. Prog. Ser. 600, 165–177. doi: 10.3354/meps12633
Hart K. M., Iverson A. R., Benscoter A. M., Fujisaki I., Cherkiss M. S., Pollock C., et al (2019). Satellite Tracking of Hawksbill Turtles Nesting at Buck Island Reef National Monument, US Virgin Islands: Inter-Nesting and Foraging Period Movements and Migrations. Biol. Conserv. 229, 1–13. doi: 10.1016/j.biocon.2018.11.011
Hart K. M., Zawada D. G., Fujisaki I., Lidz B. H. (2013). Habitat Use of Breeding Green Turtles Chelonia Mydas Tagged in Dry Tortugas National Park: Making Use of Local and Regional MPAs. Biol. Conserv. 161, 142–154. doi: 10.1016/j.biocon.2013.03.019
Hays G. C., Bailey H., Bograd S. J., Bowen W. D., Campagna C., Carmichael R. H., et al. (2019). Translating Marine Animal Tracking Data Into Conservation Policy and Management. Trends Ecol. Evol. 34, 459–473. doi: 10.1016/j.tree.2019.01.009
Hays G. C., Fossette S., Katselidis K. A., Mariani P., Schofield G. (2010). Ontogenetic Development of Migration: Lagrangian Drift Trajectories Suggest a New Paradigm for Sea Turtles. J. R. Soc Interface 7, 1319–1327. doi: 10.1098/rsif.2010.0009
Hays G. C., Hawkes L. A. (2018). Satellite Tracking Sea Turtles: Opportunities and Challenges to Address Key Questions. Front. Mar. Sci. 5, 432. doi: 10.3389/fmars.2018.00432
Hays G. C., Mortimer J. A., Lerodiaconou D., Esteban N. (2014). Use of Long-Distance Migration Patterns of an Endangered Species to Inform Conservation Planning for the World’s Largest Marine Protected Area. Conserv. Biol. 28, 1636–1644. doi: 10.1111/cobi.12325
Hussey N. E., Kessel S. T., Aarestrup K., Cooke S. J., Cowley P. D., Fisk A. T., et al. (2015). Aquatic Animal Telemetry: A Panoramic Window Into the Underwater World. Science 348 (6240), 1255642. doi: 10.1126/science.1255642
Irvine L. M., Palacios D. M., Lagerquist B. A., Mate B. R. (2019). Scales of Blue and Fin Whale Feeding Behavior Off California, USA, With Implications for Prey Patchiness. Front. Ecol. Evol. 7, 338. doi: 10.3389/fevo.2019.00338
IUCN. (2016). Increasing Marine Protected Area Coverage for Effective Marine Biodiversity Conservation Motion, Vol. 53.
Kranstauber B., Kays R., LaPoint S. D., Wikelski M., Safi K. (2012). A Dynamic Brownian Bridge Movement Model to Estimate Utilization Distributions for Heterogeneous Animal Movement: The Dynamic Brownian Bridge Movement Model. J. Anim. Ecol. 81, 738–746. doi: 10.1111/j.1365-2656.2012.01955.x
Laffoley D., Baxter J. M., Day J. C., Wenzel L., Bueno P., Zischka K. (2019). “Marine Protected Areas,” in World Seas: An Environmental Evaluation (Academic Press), 549–569.
Lascelles B., Notarbartolo Di Sciara G., Agardy T., Cuttelod A., Eckert S., Glowka L., et al. (2014). Migratory Marine Species: Their Status, Threats and Conservation Management Needs. Aquat. Conserv. Mar. Freshw. Ecosyst. 24, 111–127. doi: 10.1002/aqc.2512
Lohmann K. J., Luschi P., Hays G. C. (2008). Goal Navigation and Island-Finding in Sea Turtles. J. Exp. Mar. Biol. Ecol. 356, 83–95. doi: 10.1016/j.jembe.2007.12.017
McLeod E., Salm R., Green A., Almany J. (2009). Designing Marine Protected Area Networks to Address the Impacts of Climate Change. Front. Ecol. Environ. 7, 362–370. doi: 10.1890/070211
Monteiro J., Duarte M., Amadou K., Barbosa C., El Bar N., Madeira F. M., et al. (2021). Fibropapillomatosis and the Chelonid Alphaherpesvirus 5 in Green Turtles From West Africa. EcoHealth 18 (2), 229–240. doi: 10.1007/s10393-021-01526-y
Paiva V. H., Geraldes P., Rodrigues I., Melo T., Melo J., Ramos J. A. (2015). The Foraging Ecology of the Endangered Cape Verde Shearwater, A Sentinel Species for Marine Conservation Off West Africa. PloS One 10, e0139390. doi: 10.1371/journal.pone.0139390
Papi F., Luschi P., Akesson S., Capogrossi S., Hays G. C. (2000). Open-Sea Migration of Magnetically Disturbed Sea Turtles. J. Exp. Biol. 203, 3435–3443. doi: 10.1242/jeb.203.22.3435
Patrício A. R., Varela M. R., Barbosa C., Broderick A. C., Airaud M. B. F., Godley B. J. (2018). Nest Site Selection Repeatability of Green Turtles, Chelonia mydas, and Consequences for Offspring. Anim. Behav. 139, 91–102. doi: 10.1016/j.anbehav.2018.03.006
Patrício A. R., Varela M. R., Barbosa C., Broderick A. C., Catry P., Hawkes L. A., et al. (2019). Climate Change Resilience of a Globally Important Sea Turtle Nesting Population. Glob. Change Biol. 25, 522–535. doi: 10.1111/gcb.14520
Pendoley K. L., Schofield G., Whittock P. A., Ierodiaconou D., Hays G. C. (2014). Protected Species Use of a Coastal Marine Migratory Corridor Connecting Marine Protected Areas. Mar. Biol. 161, 1455–1466. doi: 10.1007/s00227-014-2433-7
Péron C., Grémillet D. (2013). Tracking Through Life Stages: Adult, Immature and Juvenile Autumn Migration in a Long-Lived Seabird. PLoS One 8, e72713. doi: 10.1371/journal.pone.0072713
Pottier A., Catry T., Trégarot E., Maréchal J. P., Fayad V., David G., et al. (2021). Mapping Coastal Marine Ecosystems of the National Park of Banc D’arguin (PNBA) in Mauritania Using Sentinel-2 Imagery. Int. J. Appl. Earth Obs. Geoinf. 102, 102419. doi: 10.1016/j.jag.2021.102419
Revuelta O., Hawkes L., León Y., Godley M., Raga B. J., Tomás J. A., et al. (2015). Evaluating the Importance of Marine Protected Areas for the Conservation of Hawksbill Turtles Eretmochelys Imbricata Nesting In the Dominican Republic. Endanger. Species Res. 27, 169–80. doi: 10.3354/esr00660
Runge C. A., Watson J. E. M., Butchart S. H. M., Hanson J. O., Possingham H. P., Fuller R. A. (2015). Protected Areas and Global Conservation of Migratory Birds. Science 350 (6265), 1255–1258. doi: 10.1126/science.aac9180
Santos A. J., Bellini C., Santos E. A. P., Sales G., Ramos R., Vieira D. H. G., et al. (2021). Effectiveness and Design of Marine Protected Areas for Migratory Species of Conservation Concern: A Case Study of Post-Nesting Hawksbill Turtles in Brazil. Biol. Conserv. 261, 109229. doi: 10.1016/j.biocon.2021.109229
Schofield G., Hobson V. J., Lilley M. K., Katselidis K. A., Bishop C. M., Brown P., et al. (2010). Inter-Annual Variability in the Home Range of Breeding Turtles: Implications for Current and Future Conservation Management. Biol. Conserv. 143, 722–30. doi: 10.1016/j.biocon.2009.12.011
Schuster R., Wilson S., Rodewald A. D., Arcese P., Fink D., Auer T., et al. (2019). Optimizing the Conservation of Migratory Species Over Their Full Annual Cycle. Nat. Commun. 10, 1–8. doi: 10.1038/s41467-019-09723-8
Sequeira A. M., Heupel M. R., Lea M. A., Eguíluz V. M., Duarte C. M., Meekan M. G., et al. (2019). The Importance of Sample Size in Marine Megafauna Tagging Studies. Ecol. Appl. 29, e01947. doi: 10.1002/eap.1947
Shimada T., Duarte C. M., Al-Suwailem A. M., Tanabe L. K., Meekan M. G. (2021). Satellite Tracking Reveals Nesting Patterns, Site Fidelity, and Potential Impacts of Warming on Major Green Turtle Rookeries in the Red Sea. Front. Mar. Sci. 8, 608. doi: 10.3389/fmars.2021.633814
Shimada T., Jones R., Limpus C., Hamann M. (2012). Improving Data Retention and Home Range Estimates by Data-Driven Screening. Mar. Ecol. Progr. Ser. 457, 171–180. doi: 10.3354/meps09747
Snape R. T., Bradshaw P. J., Broderick A. C., Fuller W. J., Stokes K. L., Godley B. J. (2018). Off-The-Shelf GPS Technology to Inform Marine Protected Areas for Marine Turtles. Biol. Conserv. 227, 301–309. doi: 10.1016/j.biocon.2018.09.029
Stokes K. L., Broderick A. C., Canbolat A. F., Candan O., Fuller W. J., Glen F., et al. (2015). Migratory Corridors and Foraging Hotspots: Critical Habitats Identified for Mediterranean Green Turtles. Divers. Distrib. 21, 665–674. doi: 10.1111/ddi.12317
SWOT, State of the World’s Sea Turtles. (2011). The Most Valuable Reptile in the World, the Green Turtle. Report Vol. VI. Available at: http://seaturtlestatus.org/.
Thornton D. H., Wirsing A. J., Lopez-Gonzalez C., Squires J. R., Fisher S., Larsen K. W., et al. (2018). Asymmetric Cross-Border Protection of Peripheral Transboundary Species. Conserv. Lett. 11, e12430. doi: 10.1111/conl.12430
UNEP-WCMC, IUCN. (2020). Protected Planet: The World Database on Protected Areas (WDPA). Available at: https://www.protectedplanet.net/en.
Vermaat J. E., Beijer J. A. J., Gijlstra R., Hootsmans M. J. M., Philippart C. J. M., Van Den Brink N. W., et al. (1993). Leaf Dynamics and Standing Stocks of Intertidal Zostera Noltii Hornem. And Cymodocea Nodosa (Ucria) Ascherson on the Banc D’arguin (Mauritania). Hydrobiologia 258 (1), 59–72.
Whiting S. D., Murray W., Macrae I., Thorn R., Chongkin M., Koch A. U. (2008). Non-Migratory Breeding by Isolated Green Sea Turtles (Chelonia Mydas) in the Indian Ocean: Biological and Conservation Implications. Naturwissenschaften 95, 355–360. doi: 10.1007/s00114-007-0327-y
Worton B. J. (1989). Kernel Methods for Estimating the Utilization Distribution in Home-Range Studies. Ecology 70, 164–168. doi: 10.2307/1938423
Keywords: Chelonia mydas, protected area network, connectivity, MPA, satellite tracking, West Africa, GPS, sea turtle
Citation: Patrício AR, Beal M, Barbosa C, Diouck D, Godley BJ, Madeira FM, Regalla A, Traoré MS, Senhoury C, Sidina E and Catry P (2022) Green Turtles Highlight Connectivity Across a Regional Marine Protected Area Network in West Africa. Front. Mar. Sci. 9:812144. doi: 10.3389/fmars.2022.812144
Received: 09 November 2021; Accepted: 02 March 2022;
Published: 31 March 2022.
Edited by:
Ana M. M. Sequeira, University of Western Australia, AustraliaReviewed by:
Nicole Esteban, Swansea University, United KingdomCopyright © 2022 Patrício, Beal, Barbosa, Diouck, Godley, Madeira, Regalla, Traoré, Senhoury, Sidina and Catry. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: A. R. Patrício, YXBhdHJpY2lvQGlzcGEucHQ=
†These authors share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.