
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 14 February 2022
Sec. Aquatic Physiology
Volume 9 - 2022 | https://doi.org/10.3389/fmars.2022.797517
This article is part of the Research TopicThe Cellular Stress Response and Physiological Adaptations of Corals Subjected to Environmental Stressors and PollutantsView all 19 articles
 Jeremy B. Axworthy1*
Jeremy B. Axworthy1* Emma Timmins-Schiffman2
Emma Timmins-Schiffman2 Tanya Brown1
Tanya Brown1 Lisa J. Rodrigues3
Lisa J. Rodrigues3 Brook L. Nunn2†
Brook L. Nunn2† Jacqueline L. Padilla-Gamiño1†
Jacqueline L. Padilla-Gamiño1†
Coral bleaching events are increasing with such frequency and intensity that many of the world’s reef-building corals are in peril. Some corals appear to be more resilient after bleaching but the mechanisms underlying their ability to recover from bleaching and persist are not fully understood. We used shotgun proteomics to compare the proteomes of the outer layer (OL) tissue and inner core (IC) tissue and skeleton compartments of experimentally bleached and control (i.e., non-bleached) colonies of Montipora capitata, a perforate Hawaiian species noted for its resilience after bleaching. We identified 2,361 proteins in the OL and IC compartments for both bleached and non-bleached individuals. In the OL of bleached corals, 63 proteins were significantly more abundant and 28 were significantly less abundant compared to the OL of non-bleached corals. In the IC of bleached corals, 22 proteins were significantly more abundant and 17 were significantly less abundant compared to the IC of non-bleached corals. Gene ontology (GO) and pathway analyses revealed metabolic processes that were occurring in bleached corals but not in non-bleached corals. The OL of bleached corals used the glyoxylate cycle to derive carbon from internal storage compounds such as lipids, had a high protein turnover rate, and shifted reliance on nitrogen from ammonia to nitrogen produced from the breakdown of urea and betaine. The IC of bleached corals compartmentalized the shunting of glucose to the pentose phosphate pathway. Bleached corals increased abundances of several antioxidant proteins in both the OL and IC compartments compared to non-bleached corals. These results highlight contrasting strategies for responding to bleaching stress in different compartments of bleached M. capitata and shed light on some potential mechanisms behind bleaching resilience.
Rising seawater temperatures threaten the future of coral reefs and the ecosystem services they provide. Increased temperatures can result in the breakdown of mutualistic symbiosis between corals and Symbiodiniaceae (i.e., coral bleaching), causing a severe energy deficit to the coral that impairs critical physiological functions and can lead to mortality. However, if temperature stress abates before the coral’s energy stores are consumed, symbiosis can be restored, and the coral can recover. To survive and recover from bleaching, corals must rely on internal energy and nutrient stores and/or employ alternative strategies to sustain themselves until the stressor abates and their Symbiodiniaceae population can recover (Grottoli et al., 2006; Rodrigues and Grottoli, 2007). For example, some corals exploit energy from stored lipids following bleaching, and some corals increase feeding on zooplankton to make up the energy deficit (Grottoli et al., 2006; Rodrigues and Grottoli, 2007; Ferrier-Pagès et al., 2010; Hughes and Grottoli, 2013). Bleaching comes with physiological costs and important consequences for coral fitness, such as decreased fecundity and skeletal growth, shifts in metabolic activity, increased antioxidant response and susceptibility to diseases and mortality (Lesser et al., 1990; Ward et al., 2000; Rodrigues and Grottoli, 2007; Cantin et al., 2010; Muller et al., 2018). While these processes are well documented, and we are beginning to understand their underlying cellular mechanisms, many knowledge gaps remain, including how a coral’s proteome responds to bleaching stress and how variable the response is in different parts of the colony. Identifying such proteins and patterns can help to develop physiological biomarkers of coral resilience and improve our mechanistic understanding of recovery from bleaching.
The coral host has two distinct compartments: the tissue, where cellular functions involved in the overall maintenance and fitness of the coral occur; and the skeleton, which provides structural foundation for individual corals and entire reef ecosystems. The coral’s polyps and interconnecting tissue or coenosarc between polyps make up the tissue fraction where physiological functions such as feeding, nutrient acquisition and reproduction occur, and it is in this compartment where the majority of Symbiodiniaceae reside (Yost et al., 2013). The coral’s skeleton consists of an inorganic aragonitic calcium carbonate structure deposited by the coral tissue and an intra-skeletal organic matrix, including sugars, lipids and proteins (Isa and Okazaki, 1987; Dauphin, 2001; Puverel et al., 2005). The interface between coral tissue and skeleton depends on a coral’s skeletal architecture. In corals with a perforate (porous) skeleton, tissue penetrates the cavities of the skeletal matrix. In contrast, tissue does not typically penetrate imperforate coral skeletons. It has been suggested that perforate corals are more resistant to external stressors than imperforate corals because they provide refuge for coral tissue and Symbiodiniaceae (Brown et al., 1994; Santos et al., 2009; Yost et al., 2013). Furthermore, tissue that penetrates perforate coral skeletons can host Symbiodiniaceae which may enhance calcification (Pearse and Muscatine, 1971; Gladfelter, 1983; Yost et al., 2013). These benefits of having a perforate skeleton suggest that there may be differences in cellular functions and biological processes that occur between a corals outer tissue layer and intra-skeletal tissue. However, to our knowledge, no studies have attempted to uncover these potential differences. Nor has there been a comparison of how these different compartments respond to bleaching stress.
Proteomics is a powerful tool that has advantages over other “omics” technologies for exploring how an organism responds to stress. While genomics and epigenomics provide the blueprint for all potential proteins, revealing the metabolic flexibility of an organism, they do not reveal metabolic pathways that are actively responding to the environment at a time of interest. In contrast, transcriptomics reveals which genes are transcribed in response to a specific stress, yet it has been demonstrated to be a magnified response that does not often correlate to protein translation (Gygi et al., 1999; Maier et al., 2009; Mayfield et al., 2016; Mayfield, 2020). As a result, there are challenges with relating how genes and transcripts equate to biological processes occurring in cells at the time of collection. High-throughput discovery-based proteomics circumvents these challenges by directly identifying and quantifying proteins in the cells at the time of collection, which are closer to the realized function of mRNA and have been shown to be environmentally sensitive (Pandey and Mann, 2000; Tomanek, 2014; Stuhr et al., 2018). Recent advances in proteomics allow for the identification of potentially thousands of proteins by coupling liquid chromatography with tandem mass spectrometry, i.e., “shotgun” proteomics.
In this study, we used shotgun proteomics to explore how a reef-building perforate coral, Montipora capitata, responds at the proteomic level to experimental bleaching and to investigate whether the outer layer (OL) tissue and inner core (IC) intra-skeletal tissue and skeleton compartments respond to bleaching differently. By examining proteins from these two distinct compartments, we aimed to uncover connections between metabolic functions of the OL that support overall maintenance of the coral, and of the IC that support skeletal processes. Specifically, we aimed to address these questions: (1) Are there differential abundances of metabolic proteins in the OL and IC? and (2) Which metabolic pathways differ between these different coral compartments in response to bleaching? We hypothesized that the proteomes of coral OL and IC reflect different functions specific to these two compartments, and that the proteomes and metabolic pathways of coral’s OL and IC respond differently to bleaching.
Montipora capitata is locally abundant, dominant reef-building coral in Hawai´i, United States. It is a small-polyp species (ca. 0.8 mm) that grows in branching and plating forms, or it can exhibit both morphologies in a single colony. Only the branching form was collected for this study. M. capitata was chosen because it has a highly perforate skeleton, and because it is often noted for its resilience (Ritson-Williams and Gates, 2020) indicated by its shift from auto- to heterotrophy (Grottoli et al., 2006) and its ability to reproduce following thermally induced bleaching (Cox, 2007).
Between August 22–25, 2017, 10 colonies of Montipora capitata with a diameter of ∼ 24 cm were collected from the inner lagoon of Kāne´ohe Bay, surrounding the Hawai´i Institute of Marine Biology (HIMB, 21.428° N, 157.792° W). Individual colonies were collected more than 3 m apart and although no genetic analyses were conducted, they were considered to be different genets. Upon collection, each colony was halved using a hammer and chisel to produce two genetically identical colonies (ramets). The two halves from each colony were randomly assigned to one of three outdoor flow-through tanks for the ambient temperature control group or one of three tanks for the bleaching treatment group. Colonies were allowed to acclimate for 7–10 days prior to temperature adjustments. The thermal stress treatment started in September during the period when coral bleaching events may occur. Maximum monthly mean temperature for the main Hawaiian Islands is 27°C and occurs between August–September (NOAA Coral Reef Watch, 2018). On September 1, the temperature in the bleaching treatment tanks was elevated (600 W Titanium Aquarium Heater, Bulk Reef Supply, United States) 0.6°C day–1 over 4 days, reaching a mean (±SD) temperature of 30.4 ± 0.5°C (range: 29.4–31°C). The control tanks had a temperature of 28 ± 0.9°C. On September 26, the heat in the bleaching treatment tanks was gradually decreased to ambient levels by September 29. Twenty-four hours later, two small fragments (ca. 2 cm) were sampled from each colony, one to quantify chlorophyll a and Symbiodiniaceae density, and one for proteomics. Samples were cut from the colonies using toe-nail clippers (Revlon Inc., United States) and were immediately frozen in liquid nitrogen. They were stored at −80°C at HIMB until they were shipped on dry ice to the University of Washington, WA, United States.
All tanks were maintained with a volume of 400 L of ambient sand-filtered seawater from Kāne´ohe Bay. Throughout the experiment, corals were randomly rotated among tanks weekly to minimize any tank effects. Water was circulated using 100 W submersible pumps (Rio® 26HF HyperFlow Water Pump 6019 LPH, TAAM, United States). Mean daytime photosynthetic active radiation (PAR) levels in the tanks were 584 μmol photons m–2 s–1, and mean PAR at 12:00 was 1,249 μmol photons m–2 s–1, measured using a waterproof PAR logger (Odyssey®, Dataflow Systems Limited, NZ). Corals were not given any supplemental food during the experiment.
Bleaching was measured by quantifying chlorophyll a and Symbiodiniaceae densities. Chlorophyll a was extracted by first grinding each sample in separate glass mortar and pestle. Extractions were then conducted for two consecutive 24 h-periods with fresh 100% acetone used at the start of each period. The two-part extraction allowed us to extract total chlorophyll per coral fragment. At the end of each 24-h period absorbances were measured at 630, 663, and 750 nm. Equations from Jeffrey and Humphrey (1975) were used to calculate chlorophyll concentration for each period and summed to determine total chlorophyll per sample. Symbiodiniaceae were separated from ground coral tissue by centrifugation (two times for 5 min at 4,000 rpm). Symbiodiniaceae pellets were resuspended in filtered seawater with 1% formalin and 2–3 drops of Lugol’s iodine, then homogenized using a Tissue Tearer™ (Model# 985–370). Three subsamples were counted using a hemocytometer and the mean was determined. Chlorophyll a and Symbiodiniaceae densities were standardized to grams of ash-free dry weight (gdw) of coral tissue.
Coral branches for proteomics were sub-sectioned into two sample types: an outer layer (OL) of coral tissue and an inner core (IC) consisting of intra-skeletal tissue and skeleton (Figure 1). Samples were collected at least 1 cm from the tip of the fragment. A cross-section, about 5 mm thick, was cut from the fragment, then the OL and IC were separated using a razor blade. An effort was made to collect OL samples of tissue from the oral surface of the coral to the base of the polyps where tissue meets the surface of the skeleton. The remaining skeleton and intra-skeletal tissue material made up the IC samples. All samples were crushed with a metal spatula, placed in 1.5 ml microcentrifuge tubes and frozen at −80°C until proteomic analyses.
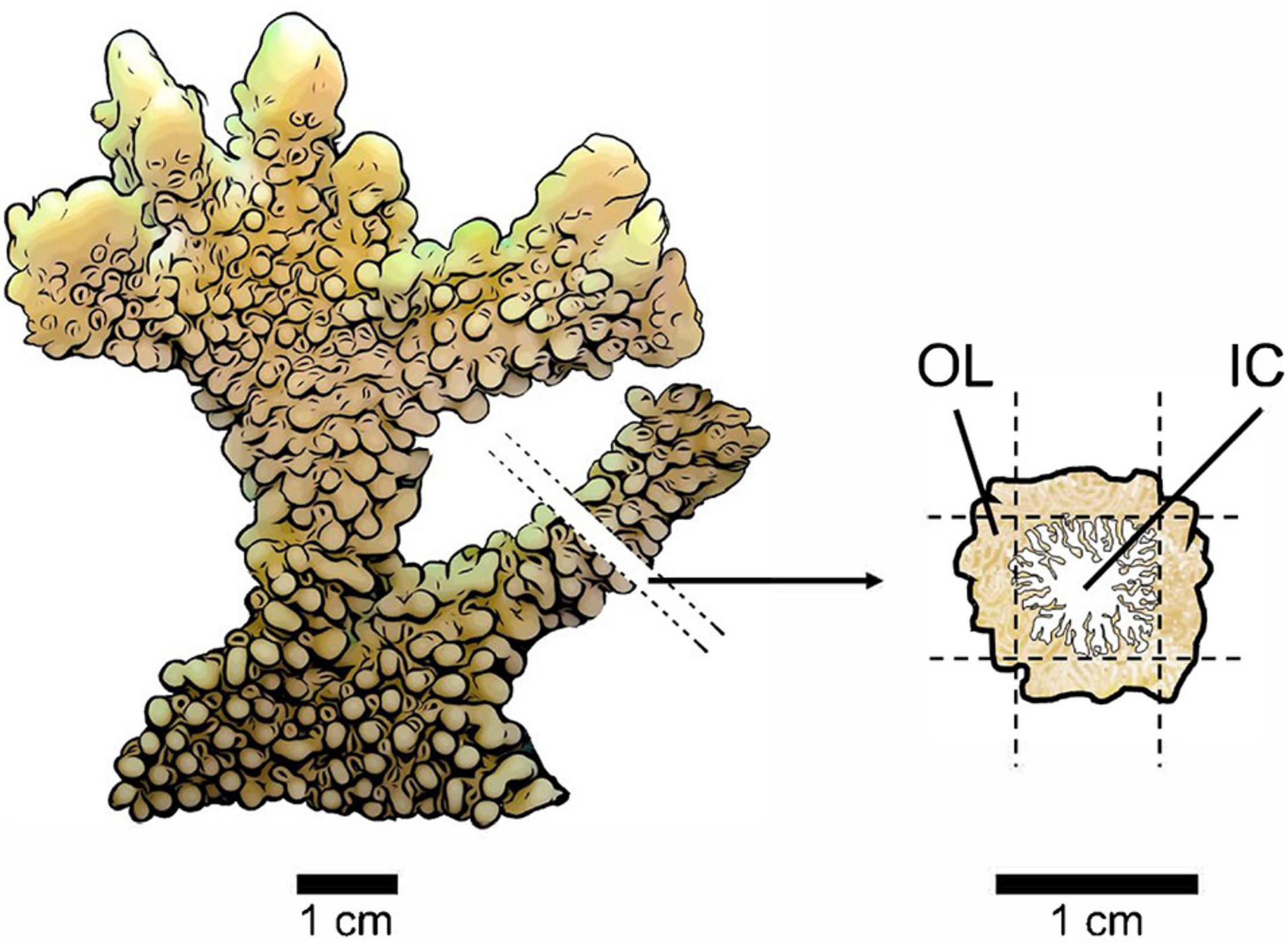
Figure 1. Sampling method for protein extractions from bleached and non-bleached Montipora capitata colonies. A small cross-section (∼5 mm thick) was removed from one branch of each coral fragment at least 1 cm from the tip. The cross-section was laid flat, and a thin layer of tissue (∼1–2 mm) was removed from four sides using a sterilized razor blade producing an outer layer (OL) of tissue and an inner core (IC) of intra-skeletal tissue and skeleton. The cross-section diagram shows the composition of our two sample types as the tissue of M. capitata penetrates the perforate skeleton (coral tissue indicated in brown and skeleton indicated in white).
To lyse cells, coral samples were sonicated in a 100 μl solution of 50 mM sodium bicarbonate (NH4HCO3) with 6 M urea three times (for 15 s then placed on ice for 20–30 s each time), using a titanium micro-probe sonicator (Branson 250 Sonifier; 20 kHz), then flash frozen in a dry ice bath for 30 s and stored at −80°C. Samples were thawed on ice and centrifuged at 4°C at 5,000 rpm for 10 min to separate and remove solid skeletal material to prevent particles from clogging the chromatography system and from disrupting pH balance during the digestion step. Protein concentrations were quantified from the supernatant in triplicate using a BCA assay (Pierce™), following the manufacturer’s instructions for limited sample size (microplate procedure). Samples from one coral colony yielded extremely low protein concentration and were removed from further analyses. For each sample, 50 μg of protein were aliquoted into a new microcentrifuge tube, then brought to a total volume of 100 μl using a solution of 50 mM NH4HCO3 with 6 M urea. Samples were frozen at −80°C until further processing.
Enzymatic digestions of the 50 μg protein lysate aliquots were performed following Nunn et al. (2015). Briefly, samples were reduced with T tris(2-carboxyethyl)phosphine, alkylated with iodoacetamide, and diluted with a 4:1 ratio of ammonium bicarbonate to methanol prior to enzymatic digestion with Promega Trypsin (1:20; enzyme: protein) overnight at 37°C. To stop the digestion, each sample’s pH was modified with 10% formic acid to a pH ≤ 2. Samples were evaporated to near dryness (<20 μl) in a speed vacuum (CentriVap® Refrigerated Centrifugal Concentrator Model 7310021) prior to desalting. To remove buffer salts, urea and other non-peptide molecules prior to mass spectrometry analysis, samples were desalted with MicroSpin™ Columns (Nest Group) following the manufacturer’s instructions. The resulting peptide samples were evaporated to near dryness, then reconstituted in 5% acetonitrile (ACN) with 0.1% formic acid to final concentration of 0.5 μg μl–1 and stored at −80°C prior to analysis.
Liquid chromatography tandem mass spectrometry was performed on Thermo Fisher QExactive according to Nunn et al. (2015). Liquid chromatography was performed using a 28 cm, 75 μm i.d., fused silica capillary column (Magic C18AQ, 100 Å, 5; Michrom, Bioresources, CA) with a 4 cm, 100 mm i.d., precolumn (Magic C18AQ, 100 Å, 5; Michrom). Peptide samples were randomized in the autosampler and loaded on the precolumn for 10 min with 5% ACN, followed by elution onto the analytical column using a 90 min gradient of 5–35% ACN with 0.1% formic acid. Top 20 data dependent acquisition (DDA) was used to collect tandem mass spectrometry data. MS1 data was collected on the mass range of 400–1,400 m/z and collision energy was set to 25. The 20 most intense ions with + 2 to + 4 charge states were selected for collision induced fragmentation and subsequent data acquisition in the MS2 using a dynamic exclusion of 10 s. The column was then washed for 10 min with 80% ACN and 0.1% formic acid and equilibrated for 10 min in 5% ACN and 0.1% formic acid prior to the next sample. Quality control standards were introduced every five runs to monitor chromatography and MS performance and visualized using Skyline (MacLean et al., 2010).
To generate a high quality protein database, the transcriptome of M. capitata (Frazier et al., 2017; GSE97888_Montiporacapitata_transcriptome.fasta) was translated using Transdecoder v 2.0.1 (Haas et al., 2013). Peptide tandem mass spectra were searched against the resulting protein database of M. capitata concatenated with 50 common contaminant proteins (Mellacheruvu et al., 2013). Spectra were searched using Comet version 2017.01 rev. 4 (Eng et al., 2013). Search parameters included a concatenated decoy search, peptide mass tolerance of 10 ppm, trypsin as the search enzyme, oxidized methionine as a variable modification (+ 15.9949 Da), alkylated cysteine (57.021464 Da), and up to three missed cleavages. PeptideProphet (Keller et al., 2002) and ProteinProphet (Nesvizhskii et al., 2003) were used to assign probabilities to peptide and protein identifications. Adjusted normalized spectral abundance factor (ADJNSAF) for each sample was determined using Abacus (Fermin et al., 2011). Abacus parameters included minimum PeptideProphet score “maxIniProbTH” = 0.99 and “miniProbTH” = 0.50, and a combined ProteinProphet score > 0.88 (FDR of 0.01) (Supplementary File 1). For all downstream analyses, the dataset included only proteins with at least 2 unique peptides across all samples and proteins identified at FDR < 0.01. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE [1] partner repository with the dataset identifier PXD021243.
To investigate the role of proteins identified by the proteomics experiments we used Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) (Kanehisa and Goto, 2000) pathway analyses. To recover GO terms for each protein, protein sequences were BLASTed against the UniProtKB Swiss-Prot non-redundant protein database (downloaded 10.15.2018). Top results are reported with cutoff E-value < 1E-10. Due to a low annotation rate retrieved from the UniProtKB database (∼49%), a second BLAST analysis was performed against the National Center for Biotechnology Information (NCBI) nr database on proteins that lacked detailed UniProt annotations. KEGG ID numbers were retrieved from BlastKoala (Kanehisa et al., 2016) using M. capitata predicted protein sequence data. GO terms and KEGG data were used to organize proteins into functional categories (e.g., carbon metabolism, response to oxidative stress, etc.; Supplementary File 2).
Proteomics differences among all sample types were visualized using non-metric multidimensional scaling (NMDS) plots based on log (base 10) transformed adjusted normalized spectral abundance factor (ADJNSAF) values. NMDS was performed with distance = “bray,” trymax = 100 and autotransform = FALSE, using the vegan package in R (Oksanen et al., 2020). Bray-Curtis dissimilarity was used to account for a high amount of zero values in the dataset. Analysis of similarity (ANOSIM) was used to test for significant differences among all sample types and between the following comparisons: bleached outer layer (BOL) vs. non-bleached outer layer (NBOL), bleached inner core (BIC) vs. non-bleached inner core (NBIC), and NBOL vs. NBIC. Statistical differences of protein abundances between sample types were determined using Qspec, a program that computes differential protein abundance (Choi et al., 2015). Analyses were performed on (n = 9) paired samples (ramets) using the qspec-paired command (burn-in = 2,000, iterations = 10,000, normalized = 1) for the following comparisons: BOL vs. NBOL, BIC vs. NBIC, and NBOL vs. NBIC. Qspec was performed using raw spectral counts with a sum of at least two unique peptides for each protein in each comparison. Qspec output included a protein-length corrected log-fold change analysis (base 2) and a false discovery rate (FDR)-corrected z-score based on the posterior distribution of the LFC parameter, guided by the FDR estimated by a well-known Empirical Bayes method. Differential expression was reported using a LFC ≥ | 0.5| and z-score ≥ | 2| (Supplementary File 3). Enrichment analysis of biological processes were compared between NBOL and NBIC using MetaGOmics ver. 0.1.1 (Riffle et al., 2017). MetaGOmics quantifies functional differences among treatments based on peptide, not protein, abundance using GO. The M. capitata proteome identified in this study was set as the background reference. Then, for each treatment group spectral counts were pooled across all samples (n = 9) and compared against the reference. GO terms were considered significant when Laplace corrected LFC > | 0.5| and when Laplace corrected and Bonferroni corrected p-value < 0.01. The Laplace correction adds one to every spectral count for each GO term to account for spectral counts that equal zero when calculating log fold ratios, while the Bonferroni correction controls for type I errors due to multiple hypothesis testing (Riffle et al., 2017). As above, GO term data were used to organize proteins into functional categories.
Photobiological data confirmed that the bleaching treatment corals were bleached (Figure 2). Symbiodiniaceae density (number of cells gdw–1) decreased ∼71% in bleached corals compared to non-bleached control corals (Wilcoxon rank sum test, W = 8, p-value = 0.002756). Chlorophyll a concentration (μg gdw–1) decreased ∼78% in bleached corals compared to non-bleached corals (T-test, t = −8.0304, df = 13.91, p-value = 1.368e-06).
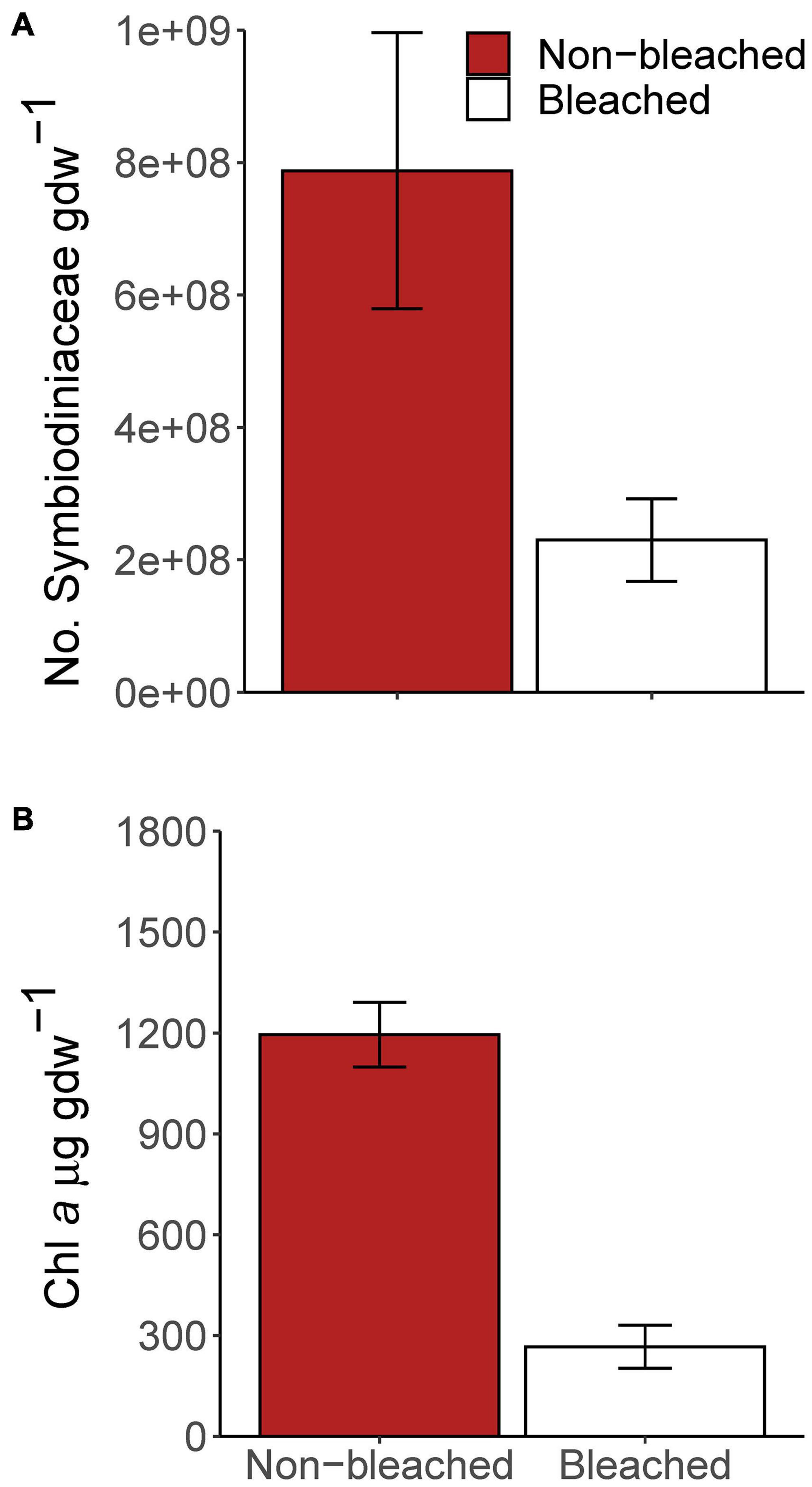
Figure 2. Photobiological measurements of experimental corals. Bars indicate the mean and error bars indicate 1 standard error, for (A) Symbiodiniaceae density (number of cells gdw– 1) and (B) chlorophyll a concentration (μg gdw– 1).
Proteomic analyses revealed a total of 2,361 proteins that were identified across all treatments and sample types. Overall, ∼90% (2,128) of the proteins identified in this study were annotated with either National Center for Biotechnology Information (NCBI) (n = 2,082) or UniProtKB IDs (n = 1,167) and ∼81% (1,918) were associated with GO terms. Of all 2,361 proteins, ∼71% (1,687) had associated KEGG terms. This high percentage of annotated proteins allowed for more detailed analysis of active metabolic pathways utilized in each sample type across both treatments. About 70% (1,660 proteins) of all proteins were found in every sample group and the number of unique proteins per treatment or sample type ranged from 0.5–2%, with the highest number of unique proteins observed in the OL of bleached corals (Figure 3A). NMDS ordination of protein profiles of all sample types show a distinction between the OL and IC compartments but not between bleached and non-bleached samples for either OL or IC (Figure 3B). ANOSIM revealed that there was a significant difference of protein profiles among all sample types (R = 0.5701; p = 9.999e-05).
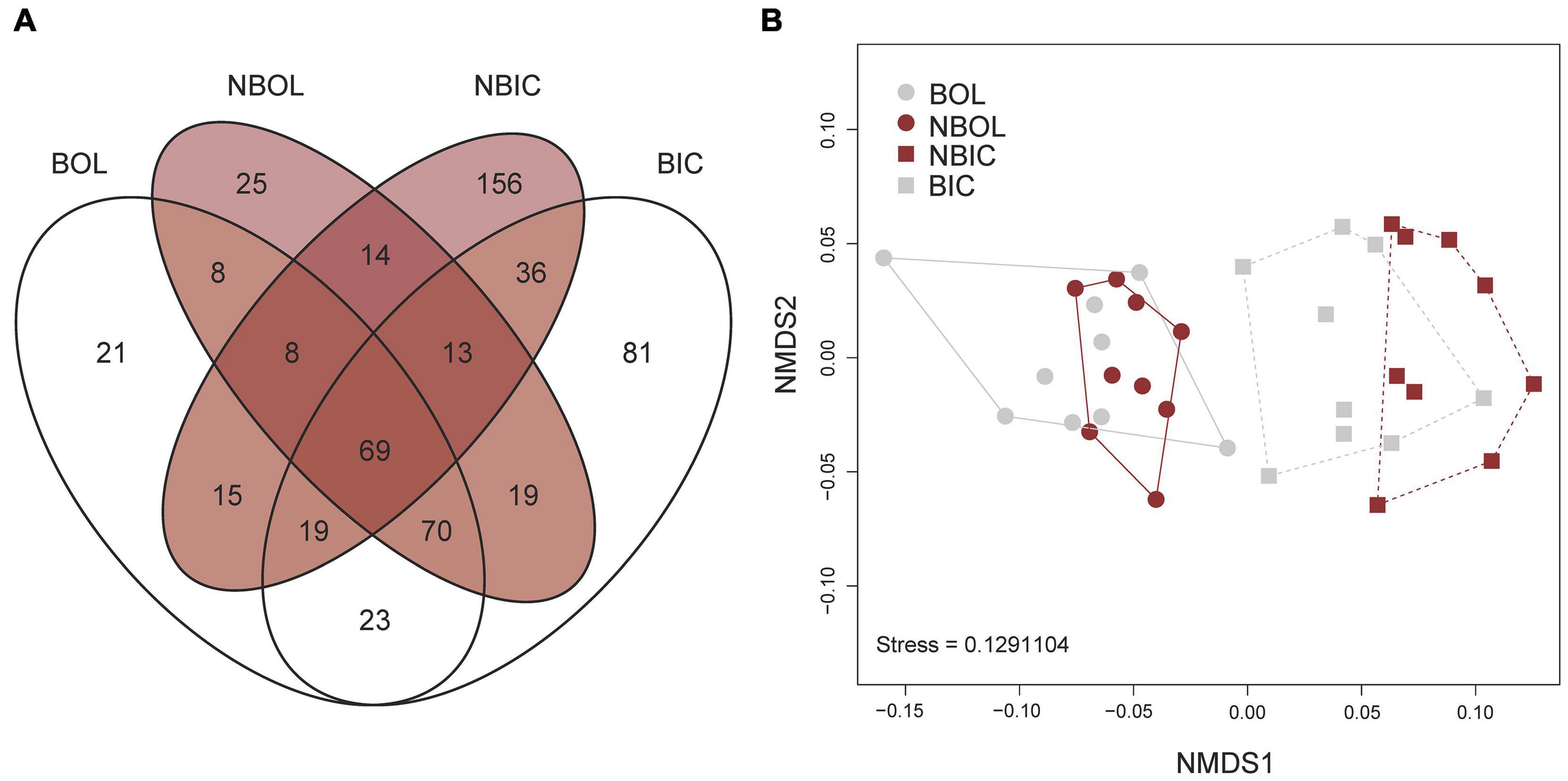
Figure 3. Global results of proteomics analysis of bleached and non-bleached Montipora capitata outer layer (OL) tissue and inner core (IC) intra-skeletal tissue and skeleton. (A) Venn diagram depicts number of proteins identified in each sample type, and (B) non-metric dimensional scaling (NMDS) plot of all treatments. BOL, bleached outer layer; NBOL, non-bleached outer layer; NBIC, bleached inner core; BIC, bleached inner core.
To gain a general understanding of the overall protein differences between M. capitata OL and IC, we compared the proteomes of the OL and IC of non-bleached corals. Across the two sample types, we identified 2,242 proteins. ANOSIM revealed that there was a significant difference of protein profiles between the OL and IC of non-bleached samples (R = 0.5936; p = 0.001). Further analysis revealed that 187 proteins were significantly more abundant in the OL and 132 proteins were more abundant in the IC (Supplementary File 3). Based on enrichment analysis, the OL of non-bleached M. capitata was enriched in biological processes involved in anatomical development, carbohydrate metabolism, cell differentiation, endocytosis, lipid metabolism, protein metabolism, RNA processing and small molecule processing (Figure 4A and Supplementary File 4). Peroxidasin (PER2; m.25079; LFC−3.7) was detected at the highest abundance in the OL and is part of cellular response to oxidative stress. Several other proteins related to carbon and nitrogen metabolism were also present in high abundances such as GDP-mannose 4,6 dehydratase (GMD; m.26053; LFC−1.2), pyruvate dehydrogenase (PDH; m.4559; LFC−1.1), aconitate hydratase (ACN; m.20259; LFC−0.7), and isocitrate dehydrogenase (IDH; m.11767; LFC−0.7) (Figure 4B). The IC was characterized by an enrichment in GO biological processes involved in cellular structure and adhesion, carbohydrate metabolism, immune response, lipid metabolism, oxidoreductase activity and response to stimulus (Figure 4A and Supplementary File 4). The protein identified in the OL to have the highest fold change difference compared to the IC was a lipase-related protein (LIP1; m.21324, LFC + 2.1) that is involved in lipid metabolism (Figure 4B). Von Willebrand factor type A (VWA; m.31548, LFC + 1.5), an important adhesion structural protein, was also present at significantly high concentration in the IC (Figure 4B).
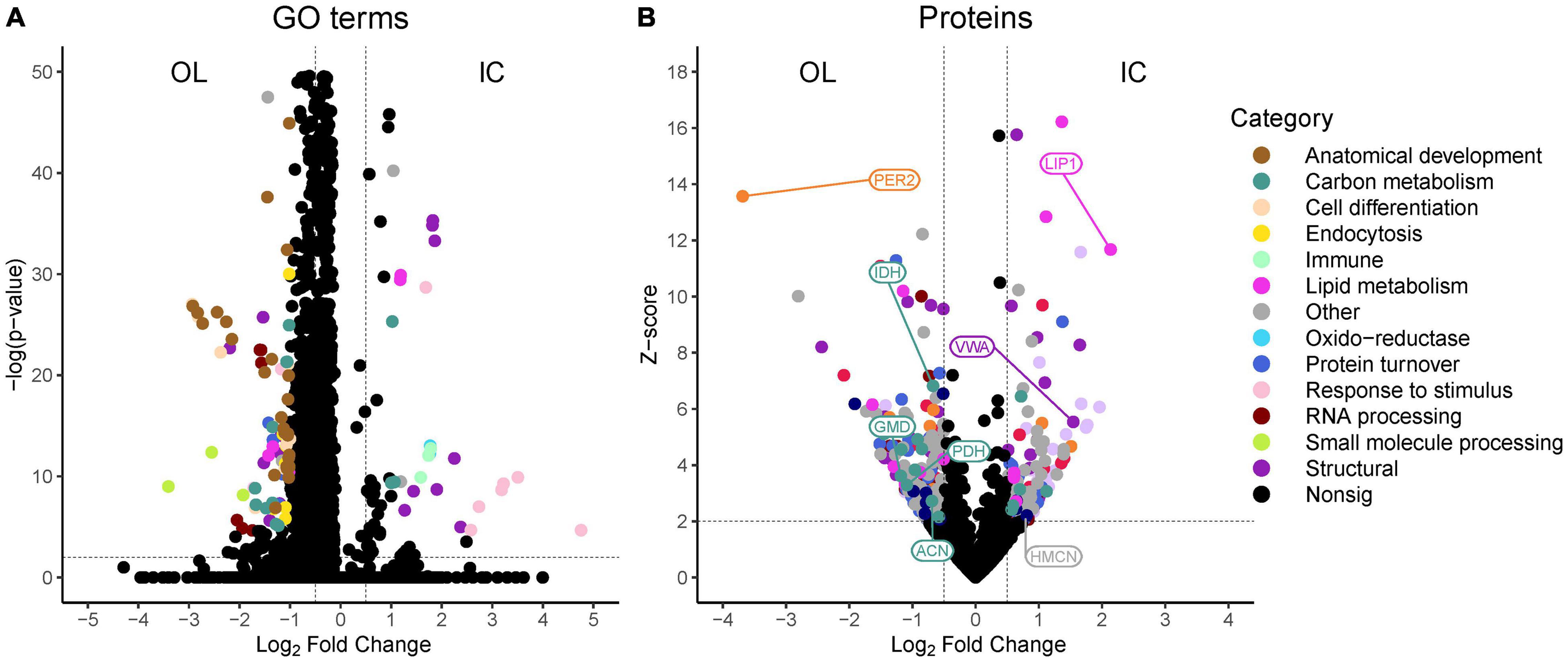
Figure 4. Volcano plots of (A) associated gene ontology (GO) terms derived from MetaGOmics and (B) proteins identified in non-bleached outer layer (OL) tissue and inner core (IC) intra-skeletal tissue and skeleton of Montipora capitata. Points represent the magnitude of the log2 fold change and the -log(p-value) or z-score for each GO term or protein, respectively. Dashed lines indicate significance thresholds. Select proteins that are discussed in the text are indicated by their abbreviated protein name.
In the OL of bleached and non-bleached M. capitata, 2,281 proteins were identified. ANOSIM did not reveal a significant difference of protein profiles between OL of bleached and non-bleached corals (R = 0.05316; p = 0.159). Qspec analysis of differential abundances between the OL of bleached and non-bleached corals revealed 63 significant proteins at higher, and 28 at lower, abundance in bleached corals compared to non-bleached corals (Supplementary File 3). Biological enrichment analysis of GO terms identified that the OL of bleached corals have a higher abundance of proteins involved in carbon metabolism, lipid metabolism, nitrogen metabolism, protein metabolism, response to oxidative stress, RNA processing and cellular signaling and transport than non-bleached corals (Figures 5A, 6). Proteins with the highest log-fold change included calumenin (CALU; m.10914, LFC + 1.8), betaine-homocysteine S-methyltransferase (BHMT; m.1453, LFC + 1.6), peroxidasin (PER1; m.28022, LFC + 1.5), urease (URE; m.24102, LFC + 1.4), and catalase (CAT; m.4762, LFC + 1) (Figures 5A, 6). Key cellular pathways that increased in the OL of bleached corals include the glyoxylate cycle, fatty acid beta oxidation, beta alanine metabolism, protein degradation and synthesis, betaine degradation and urea degradation. GO terms associated with less abundant proteins in the OL of bleached corals are associated with biological processes involved in lipid metabolism, amino acid synthesis, protein metabolism, RNA processing, cellular signaling and transport, and cellular structure (Figures 5A, 6). Proteins that were significantly lower in abundance in the OL of bleached corals included cholesterol transporter (CHLT; m.15955, LFC −1.8), glutamine synthetase (GS; m.30399, LFC −1.6), phospholipase B (PLIP; m.27749, LFC −1.1), and mucin protein (MUC; m.29491, LFC −1) (Figures 5A, 6).
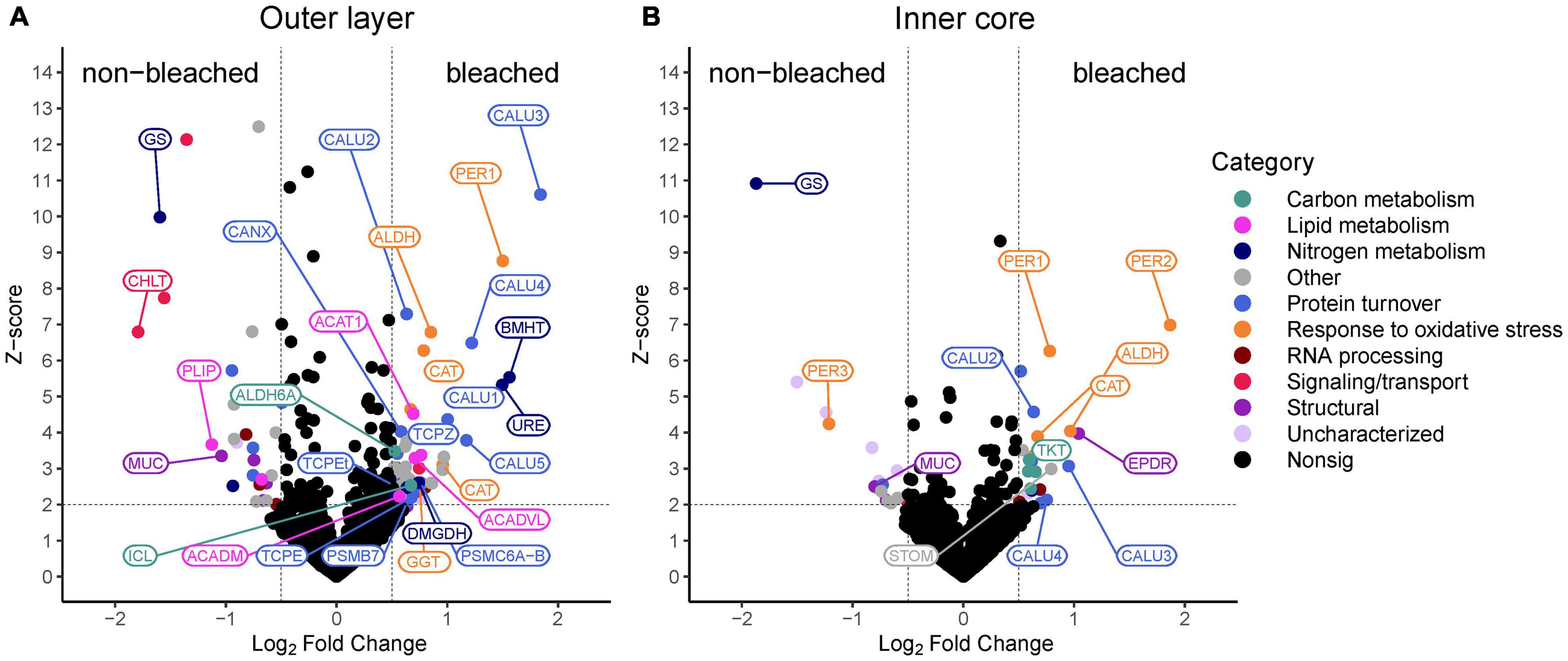
Figure 5. Volcano plots of proteins identified in non-bleached and bleached (A) outer layer (OL) tissue and (B) inner core (IC) intra-skeletal tissue and skeleton of Montipora capitata. Points represent the magnitude of the log2 fold change and the z-score for each protein. Dashed lines indicate significance thresholds. Select proteins that are discussed in the text are indicated by their abbreviated protein name.
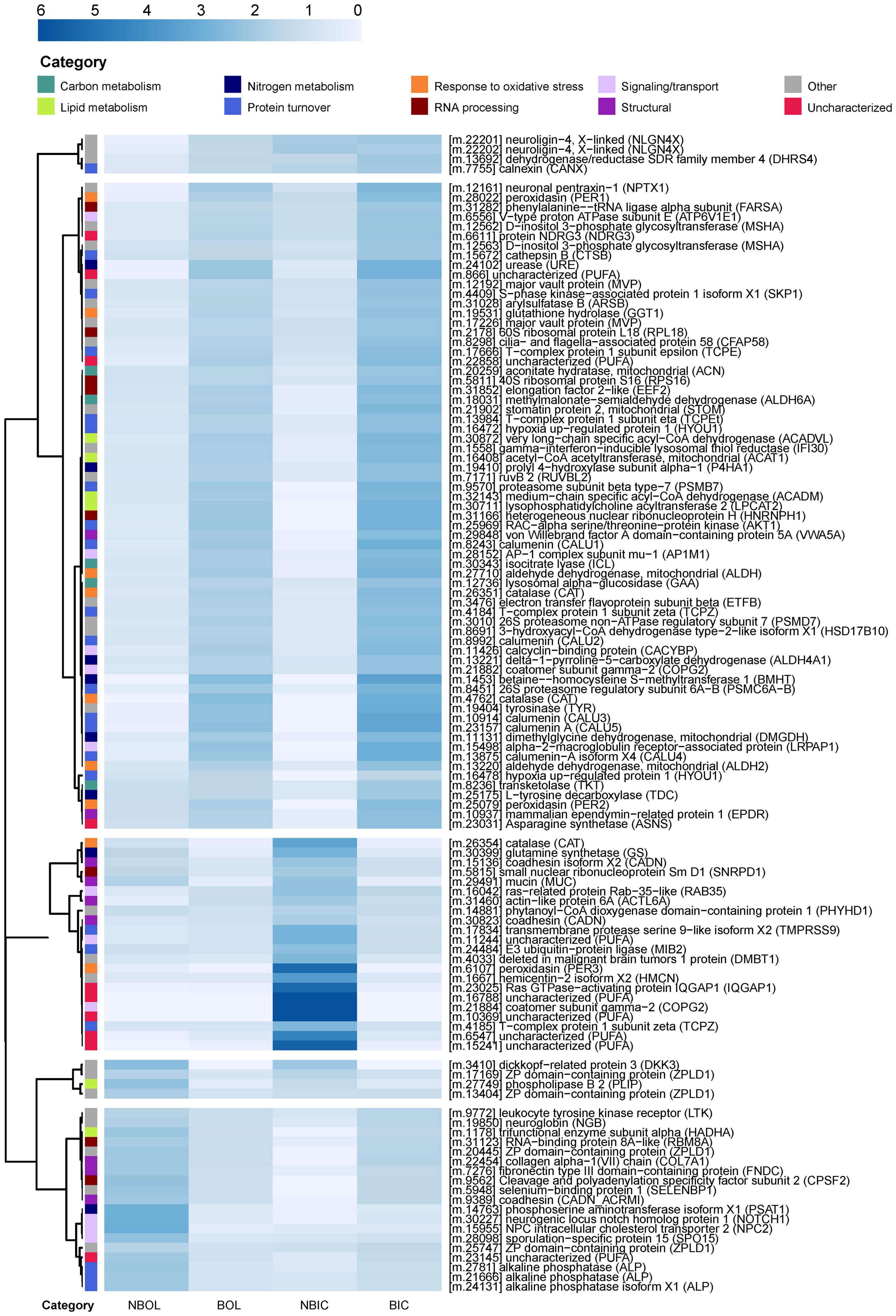
Figure 6. Heatmap of differentially abundant proteins in non-bleached and bleached Montipora capitata outer layer (OL) tissue and inner core (IC) intra-skeletal tissue and skeleton. Cell shading represents the mean NSAF value for each treatment normalized by the row mean. Rows are clustered using the “correlation” method of the pheatmap function in R. The dendrogram was set to cut 5 distinct clusters. The row annotations (categories) represent broad biological function categories based on GO and KEGG terms associated with each protein. NBOL, non-bleached outer layer; BOL, bleached outer layer; NBIC, non-bleached inner core; BIC, bleached inner core.
In the IC of bleached and non-bleached M. capitata, 2,181 proteins were identified. ANOSIM did not reveal a significant difference of protein profiles between the IC of bleached and non-bleached corals (R = −0.04287; p = 0.636). Qspec analysis of relative abundances of the proteins detected in the IC revealed 22 proteins at significantly higher abundance and 17 at lower abundance in the IC of bleached corals compared to non-bleached corals. Associated GO terms indicate that proteins at higher abundance in the IC of bleached corals are involved in biological functions including carbon metabolism, protein metabolism, response to oxidative stress and RNA processing (Figures 5B, 6). The top 5 proteins with the highest positive log-fold change in the IC of bleached corals included peroxidasin (PER2; m.25079, LFC 1.9), ependymin (EPDR; m.10937, LFC 1), aldehyde dehydrogenase (ALDH; m.27710, LFC 1), calumenin (CALU; m.10914, LFC 0.9), and stomatin (STOM; m.21902, LFC 0.8) (Figures 5B, 6). Additionally, a protein involved in the pentose phosphate pathway was also represented in this dataset with significantly increased abundance (Figures 5B, 6). Biologically enriched GO terms associated with the IC of non-bleached corals were involved in nitrogen metabolism, cellular signaling and cellular structure (Figures 5B, 6). The proteins that were lower in abundance in the IC of bleached corals compared to the IC of non-bleached corals included glutamine synthetase (GS; m.30399, LFC −1.9), peroxidasin (PER3; m.6107, LFC −1.2), and mucin (MUC; m.29491, LFC −0.8) (Figures 5B, 6).
Proteomics has proven to be a valuable tool for uncovering active cellular processes and elucidating the response of different biological compartments of symbiotic cnidarians in response to stress (Weston et al., 2015; Ricaurte et al., 2016; Cziesielski et al., 2018; Hernández-Elizárraga et al., 2019; Mayfield, 2020; Mayfield et al., 2021; Petrou et al., 2021). Our high-throughput discovery-based proteomic analyses of the OL and IC of bleached and non-bleached corals support our hypothesis that proteomic signatures differ between M. capitata OL and IC, reflecting different biological functions associated with these two compartments. Additionally, comparisons of bleached and non-bleached proteomes support our hypothesis that M. capitata OL and IC respond in different metabolic ways to bleaching stress. We observed bleaching induced differences in carbon metabolism, response to oxidative stress, protein turnover and nitrogen metabolism between corals’ OL and IC compartments. Our results corroborate several previous transcriptomics studies (DeSalvo et al., 2010, 2012; Barshis et al., 2013; Kenkel et al., 2013; Polato et al., 2013; Aguilar et al., 2019) by providing direct evidence of protein translation in response to bleaching stress. To our knowledge, our analysis was the first to compare the simultaneous bleaching proteomic response in both OL and IC. Our results highlight the differential metabolic response between OL and IC in M. capitata that may reflect the compartmentalization of specific physiological functions and energy acquisition and allocation in each compartment.
Among our four sample types, the most significant proteomic difference was found when comparing the OL and IC (see Figure 3B). Biological enrichment analysis of GO terms using MetaGOmics highlight how the coral OL is characterized by routine cellular activities such as metabolic processes, cellular maintenance, and growth, while the IC was significantly enriched in proteins related to structural maintenance, cell adhesion, immune and stimulus response, as well as carbon and lipid metabolism. These differences in biological processes suggest that there are compartmentalized pathways between the OL and IC, with IC proteins playing a larger role in skeletal formation than OL proteins. Previous proteomic studies of coral skeleton report the presence of structural and cell adhesion proteins that were also found in this study in the M. capitata IC compartment, including von Willebrand factor type A (VWA), hemicentin (HMCN), and collagens (COL) (Drake et al., 2013; Ramos-Silva et al., 2013). The von Willebrand factor type A containing proteins have been suggested to play a role in connecting the layer of cells at the interface of the skeleton organic matrix to the skeleton, and collagen has been suggested to serve as a site for proteins to bind where minerals, such as calcium carbonate, can nucleate (Drake et al., 2013).
A considerable amount of redundancy was revealed between the OL and IC of M. capitata as ∼2,000 proteins were detected with similar abundances between the two compartments, which is likely due in part to our sampling method of a perforate coral. In corals with a perforate skeleton, tissue penetrates the complex matrix of crevices and cavities and can host Symbiodiniaceae that can drive biological activity (Pearse and Muscatine, 1971; Gladfelter, 1983; Yost et al., 2013). In contrast, the skeleton of imperforate corals is more dense and typically does not contain coral tissue. Evidence suggests that perforate corals are more robust to stressful events than imperforate corals (Krupp et al., 1992; Jokiel et al., 1993; Santos et al., 2009), which may be due to higher protein and Symbiodiniaceae densities in their deep tissue compared to imperforate corals (Schlöder and D’Croz, 2004). It has also been suggested that perforate skeletons can provide refuge for deep coral tissue and Symbiodiniaceae that reside deep in the skeleton during stressful conditions (Santos et al., 2009). Given that the IC samples collected here contain both coral tissue and skeleton, it is likely that there is some overlap in metabolic processes and pathways that occur between the OL and IC compartments, which would explain some of the similarity in proteins between them. Furthermore, proteins from tissue within the skeleton that do not play a role in calcification may have been at a higher abundance and were more likely to be sampled with our mass spectrometry methods than proteins that are directly involved in calcification, such as carbonic anhydrase, calcium ion pumps or the coral acid rich proteins identified by other studies (Drake et al., 2013; Ramos-Silva et al., 2013).
Carbon is a critical nutrient for corals that is primarily provided in the form of glucose-based photosynthate that is passed to the host from Symbiodiniaceae (Muscatine et al., 1981). As corals bleach and the association with Symbiodiniaceae is weakened and carbon-based photosynthate decreases, it has been proposed that corals maintain their energetic needs through the degradation of stored lipids (Grottoli and Rodrigues, 2011). Here, we provide direct evidence of modifications to carbon metabolic pathways through quantitative analyses of proteins that are differentially abundant in bleached corals compared to non-bleached corals. Our results reveal mitigation strategies for bleached corals to generate the required energy needed to persist after bleaching stress.
In the OL of bleached M. capitata, increased abundance of isocitrate lyase (ISL) suggests an increased capacity to produce glyoxylate. Typically produced in the glyoxylate cycle, glyoxylate is a two-carbon metabolite and a precursor to glucose and many other C-storage molecules. The glyoxylate cycle, a variation of the Krebs cycle, uses acetyl-CoA as a source of carbon and conserves carbon by bypassing two decarboxylation steps of the Krebs Cycle (Kornberg and Krebs, 1957). Although it was once thought to only occur in plants and bacteria, recent transcriptomics studies have shown that glyoxylate cycle enzymes also occur in corals (DeSalvo et al., 2010; Kenkel et al., 2013; Polato et al., 2013). Bleached corals may upregulate the glyoxylate cycle, or the production of glyoxylate, to metabolize energy in stored lipids, breaking down triacylglycerol into their component fatty acids for release of acetyl-CoA, when carbohydrate supply is low (Polato et al., 2013; Wright et al., 2015; Petrou et al., 2021). Malate synthetase (MS), the other key enzyme of the glyoxylate cycle that converts glyoxylate and acetyl-CoA to malate, was present in the proteomes identified in this study, but no significant difference was detected between bleached and non-bleached corals. The same trend of increased ISL abundance and no increase in MS abundance has been reported for Porites asteroides (Kenkel et al., 2013) and Acropora millepora (Petrou et al., 2021). We also detected three key enzymes involved in lipid metabolism in higher abundance in the OL of bleached corals compared to the OL of non-bleached corals: very long-chain specific acyl-CoA dehydrogenase (ACADVL), medium chain specific acyl-CoA dehydrogenase (ACADM) and acetyl-CoA acetyltransferase (ACAT1). The presence of these enzymes further supports the hypothesis that bleached corals utilize the glyoxylate cycle to extract energy from lipids when Symbiodiniaceae-derived photosynthate is decreased (Figure 7).
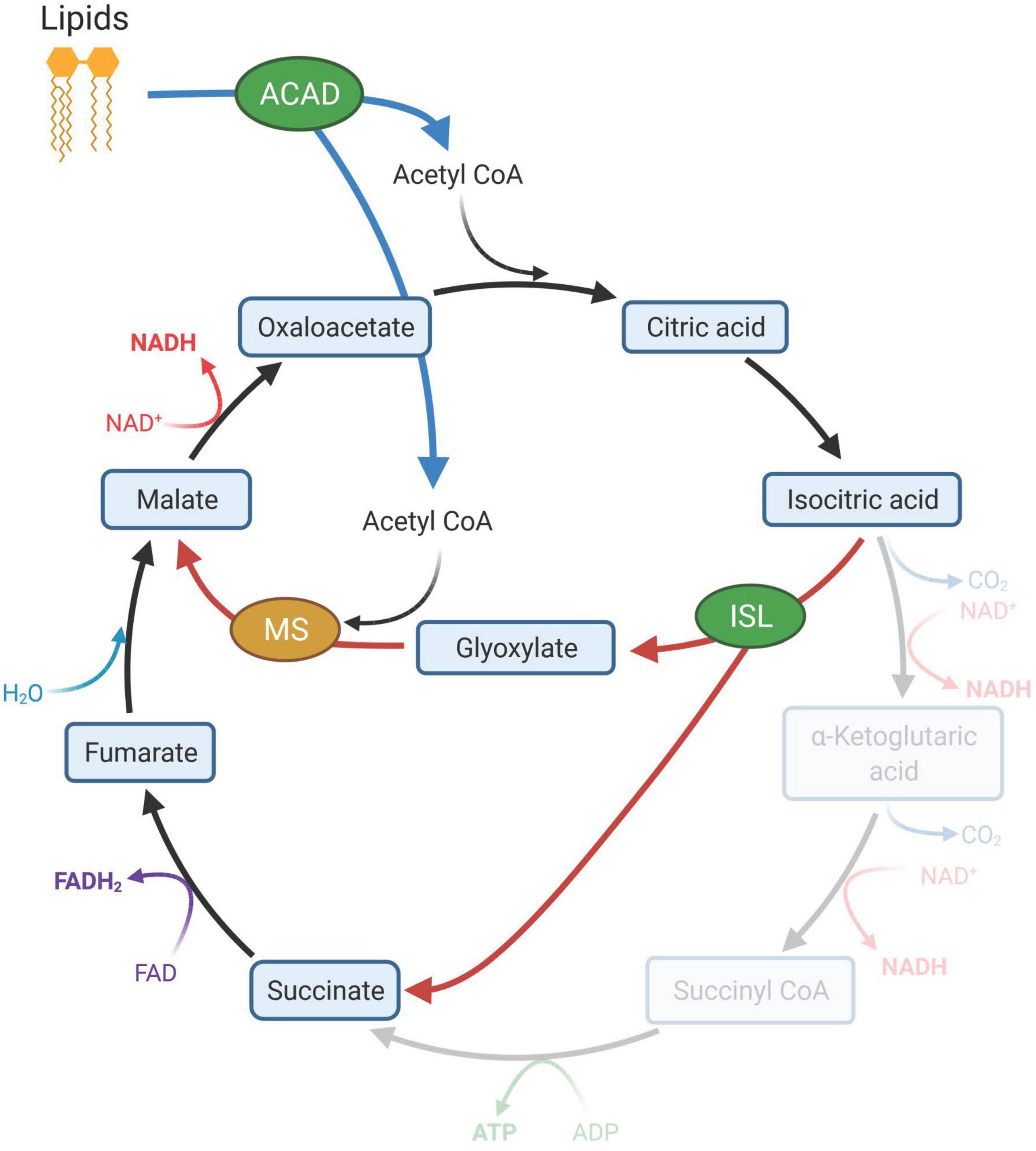
Figure 7. Lipid degradation and the glyoxylate cycle. Increased abundance of acyl-CoA dehydrogenases (ACAD) in the outer layer (OL) tissue of bleached coral indicated the breakdown of lipids that produce acetyl CoA (blue arrows). Acetyl CoA is the primary source of carbon in the glyoxylate cycle (red arrows). The glyoxylate cycle, a modified version of the Kreb’s cycle (black arrows), utilizes isocitrate lyase (ISL) and malate synthetase (MS) and can produce additional intermediate molecules of the Krebs cycle that can be used to generate glucose. ISL was significantly elevated in the OL of bleached corals while MS was elevated but not significantly. Created with BioRender.com.
Malonate-semialdehyde dehydrogenase (ALDH) was another enzyme that was significantly increased in bleached corals and can provide an additional source of acetyl-CoA that could be used in the glyoxylate cycle. ALDH is involved in several metabolic pathways including carbon metabolism, beta-alanine metabolism and propanoate metabolism where it catalyzes the conversion of malonate semialdehyde (3-oxopropanoate) to acetyl-CoA. Enzymes associated with the glyoxylate cycle and fatty acid beta-oxidation were higher in the OL of bleached corals than the OL of non-bleached corals. However, ALDH was more abundant in both the OL and IC of bleached corals compared to these compartments in non-bleached corals (Figure 5).
An enzyme involved in the pentose phosphate pathway (PPP), transketolase (TKT), was lower in the IC of non-bleached corals compared to the OL of non-bleached corals, but higher in the IC of bleached corals than the IC of non-bleached corals, suggesting that this pathway is initially prioritized in the OL but becomes more essential in the IC during bleaching. The PPP helps maintain carbon homeostasis, generates precursors to nucleotides, and produces reducing molecules (e.g., NADPH) to fight oxidative stress. This pathway has been elevated in stressed cnidarians suffering from disease (Wright et al., 2015; Garcia et al., 2016) as well as thermal stress (Oakley et al., 2017; Fonseca et al., 2019). Hernández-Elizárraga et al. (2019) found proteomic evidence that bleached fire coral, Millepora complanata, redirected carbohydrate flux from glycolysis to the PPP, which would help the coral alleviate oxidative stress through the production of NADPH. Additionally, the generation of nucleotides could assist with multiple other cellular processes. Future research should investigate potential links between the pentose phosphate pathway and coral calcification.
Oxidative stress, due to production of excess reactive oxygen species (ROS), has been implicated as a key underlying mechanism in the breakdown of coral-algal symbiosis leading to bleaching (Lesser et al., 1990; Lesser, 1997; Downs et al., 2002). Under environments favoring physiological homeostasis, ROS produced during metabolic processes of the coral host, Symbiodiniaceae, and in the chloroplasts of Symbiodiniaceae, are kept in check by cellular antioxidants. Under environmental stress, however, this tightly regulated process can become unbalanced resulting in excess ROS, leading to cellular damage and bleaching (Lesser et al., 1990; Downs et al., 2002). One possible mechanism for a coral’s resilience to bleaching may be explained by the timing, magnitude, and types of antioxidants regulated during stress (Gardner et al., 2017).
Several enzymes involved in oxidative stress response were at higher abundance in both the OL and IC of bleached M. capitata compared to samples from non-bleached corals. This observation is consistent with proteomics (e.g., Weston et al., 2015; Cziesielski et al., 2018; Petrou et al., 2021) and transcriptomics (e.g., DeSalvo et al., 2008, 2010; Voolstra et al., 2009) studies that have demonstrated increased antioxidant activity in response to thermal stress in cnidarians. Multiple catalases (CAT) were elevated in the OL and IC of bleached corals. This antioxidant enzyme is an efficient scavenger of hydrogen peroxide and is commonly observed in high quantities in bleached corals (Lesser et al., 1990; Seneca et al., 2010; Krueger et al., 2015; Gardner et al., 2017). Peroxidasins (PER1, PER2, PER3) that also neutralize hydrogen peroxide were detected at significantly higher abundances in both compartments from bleached corals. In coral embryos, significant upregulation of peroxidasin genes, a multi-domain peroxidase, has been observed after 48 h of thermal stress (Voolstra et al., 2009), while in adult corals it was downregulated after 11 days of thermal stress exposure (DeSalvo et al., 2008). Here we report that peroxidases are present in an adult coral following nearly 1 month of thermal stress. Peroxidasin has been proposed as a biomarker for heat stress in coral embryos and adults (Voolstra et al., 2009) and may be important for resilience to high temperatures (Barshis et al., 2013).
Another enzyme higher in the OL of bleached corals, gamma-glutamyltranspeptidase 1 (GGT1), likely plays an indirect role in oxidative stress response through its role in the glutathione cycle. Glutathione, in its reduced form, is important for various metabolic processes where it can neutralize ROS and help break down toxins (Mailloux et al., 2013; Bachhawat and Yadav, 2018). GGTs break down oxidized glutathione that has been transported out of the cell into glutamate and cysteine-glycine. Cysteine-glycine can then be transported back into the cell to produce more glutathione in its reduced form, ready to neutralize ROS (Mailloux et al., 2013; Bachhawat and Yadav, 2018).
Environmental stressors that have the potential to induce cellular damage, or significantly alter homeostatic metabolisms, drive high protein turnover rates (Downs et al., 2002; Oakley et al., 2017). Several pathways, including protein degradation, translation, and protein folding, play key roles in the turnover of proteins and maintaining proteostasis. Significantly higher abundances of proteins involved in each of those pathways were quantified in bleached corals compared to non-bleached controls, indicating a significantly higher level of protein turnover. In the OL of bleached corals, protein degradation was indicated by higher abundance of two proteins associated with proteasomes, the protein complexes responsible for degrading extraneous or damaged proteins: 26S proteasome regulatory subunit 6A-B (PSMC6A-B) and proteasome subunit beta type-7-like (PSMB7). The observed increase in proteasome components in this study is consistent with transcriptomic (Traylor-Knowles et al., 2017) and proteomic (Petrou et al., 2021) research investigating short term heat stress in corals.
Multiple chaperone subunits of the T-complex protein Ring Complex (TRiC: TCPE, TCPEt, TCPZ), as well as calnexin (CANX) were in high abundance in the OL of bleached corals, suggesting that bleached corals were undergoing endoplasmic reticulum (ER) stress. ER stress occurs due to accumulation of unfolded or misfolded proteins and initiates the unfolded protein response which, under prolonged or severe stress, can lead to cell death (Xu et al., 2005). The TRiC assists the folding of up to 10% of eukaryotic cytosolic proteins involved in a broad range of functions (Yam et al., 2008). It is suggested to be well-equipped for folding complex, slow-folding proteins that are prone to aggregation (Yam et al., 2008; Gestaut et al., 2019), making it critical for maintaining proteostasis and reducing ER stress. Calnexin is recognized for its role in processing glycoproteins in the ER where it binds to unfolded or misfolded proteins and prohibits their release (Ou et al., 1993). Eight different calumenins (CALU) were in higher abundance in bleached corals, including five in the OL (CALU1-5) and three in the IC (CALU2, CALU3, CALU4). Although its role as a chaperone in the ER has not been directly confirmed, increased abundance of calumenin resulted in down-regulation of proteins involved in ER stress reduction, suggesting it functions similarly to chaperones (Lee et al., 2013). Chaperones, most notably heat shock proteins, are commonly detected in heat-stressed and bleached corals (Black et al., 1995; Brown et al., 2002; Barshis et al., 2013; Weston et al., 2015; Ricaurte et al., 2016; Traylor-Knowles et al., 2017; Seveso et al., 2020) because of their role in stress response. However, no heat shock proteins were identified at significant levels in this study. Few coral bleaching studies have reported the presence of the chaperone proteins detected in the current study (Bellantuono et al., 2012; Maor-Landaw et al., 2014; Chakravarti et al., 2020).
The increased abundances of proteins related to high protein turnover in our bleached samples, one day after the heat stress was removed, may be the result of a rapid adjustment to a change in the coral’s environment (e.g., a recovery phase) via alteration of its proteome. High rates of protein turnover have been associated with an organism’s ability to acclimatize (Hawkins, 1991). For corals it has been suggested that slow growth rates and high metabolic rates correlate to high protein turnover and acclimatization potential (Gates and Edmunds, 1999). An analysis of protein turnover rates in bleached and healthy states between M. capitata and other species would allow more direct testing of the hypothesis that protein turnover is directly linked to thermal acclimation.
Nitrogen (N) is a critical element used for synthesizing nucleotides, amino acids, proteins and other molecules in both the coral host and Symbiodiniaceae. Living in oligotrophic, tropical waters, the coral holobiont has adapted to efficiently acquire N from the surrounding environment via heterotrophy, or assimilation of dissolved organic and inorganic N by the host and the Symbiodiniaceae (Grover et al., 2006, 2008; Pernice et al., 2012). Although ammonia is the primary N-based metabolite (Grover et al., 2008), corals also have the enzymatic capacity for transport and assimilation of nitrate, dissolved free amino acids and, to a much lesser degree, urea from the surrounding water, providing them with a range of mechanisms to adapt to changing nutrient conditions (Grover et al., 2008). Nitrogen is transferred between the host and Symbiodiniaceae (Rahav et al., 1989; Atkinson et al., 1994; Hoegh-Guldberg and Williamson, 1999), however, when symbiosis breaks down due to prolonged thermal stress, mutualistic N-exchange decreases, demanding the coral host independently acquire N from the environment or intracellularly recycle it. Here we show that bleached M. capitata switches its dominant N- acquisition strategy from ammonia uptake to the degradation of urea and betaine (Figure 8).
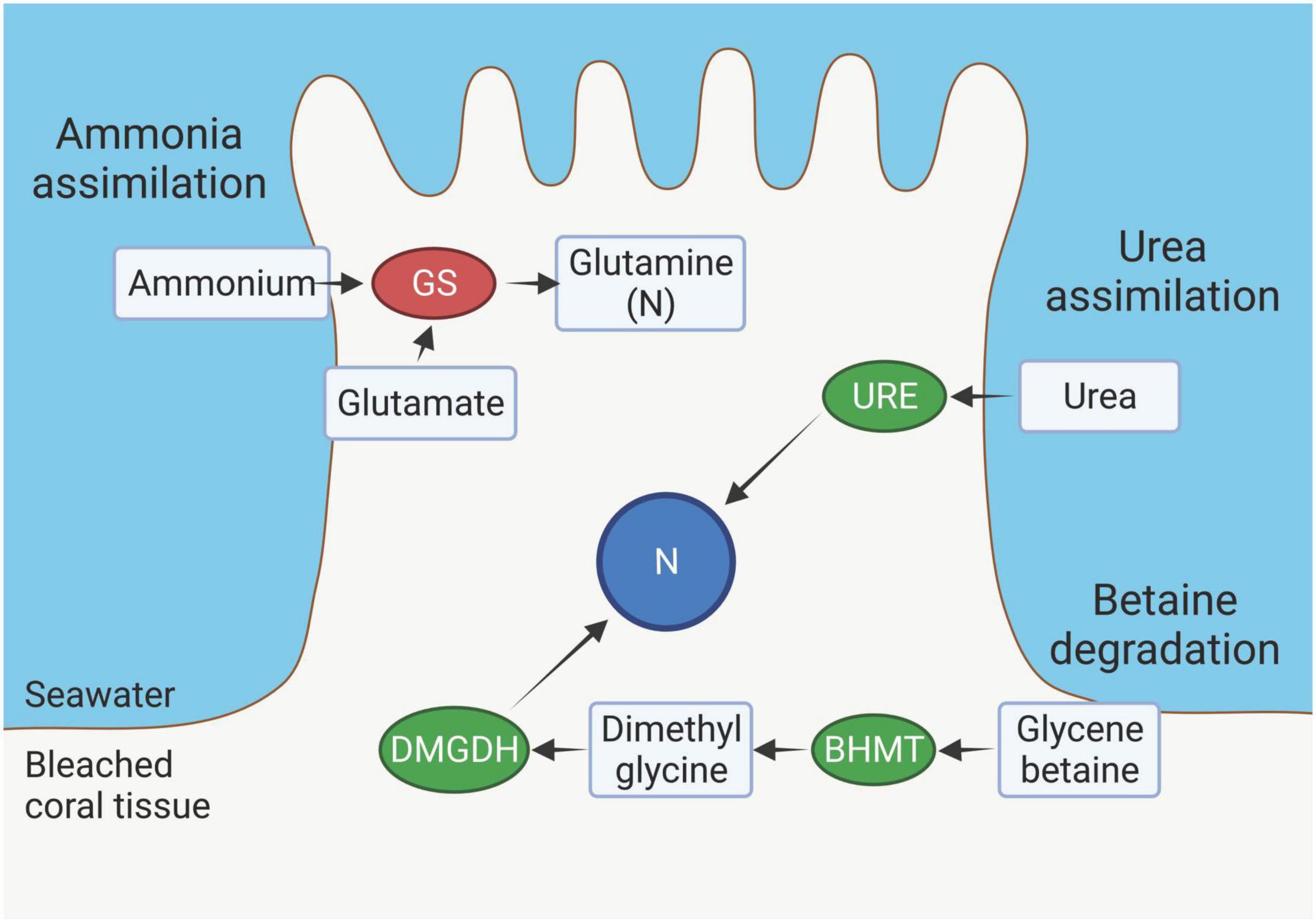
Figure 8. Nitrogen (N) source pathways in the outer layer (OL) tissue of bleached Montipora capitata. Decreased abundance of glutamine synthetase (GS) suggests a decrease in ammonium assimilation from surrounding seawater. An increased abundance of urease (URE) suggests an increase in assimilation of urea from surrounding seawater that is broken down by URE to ammonia and carbon dioxide. Increased abundances of dimethylglycine dehydrogenase (DMGDH) and betaine–homocysteine S-methyltransferase (BHMT) indicate an increase in the betaine degradation pathway. Created with BioRender.com.
Enzymes involved in the typical pathway of ammonia assimilation decreased in bleached corals. Both the coral host and Symbiodiniaceae are able to assimilate ammonia via catalysis by glutamine synthetase (GS) or glutamine dehydrogenase in the host (Yellowlees et al., 1994; Wang and Douglas, 1998; Su et al., 2018) and via the glutamine synthetase/glutamine:2-oxoglutarate aminotransferase (GS/GOGAT) cycle in Symbiodiniaceae (D’Elia et al., 1983; Roberts et al., 2001). In bleached corals, GS was present at significantly lower abundances in both the OL and the IC compared to non-bleached corals, suggesting an alternate route for N acquisition must be utilized (Figures 5, 6). This result is consistent with recent studies in corals (Petrou et al., 2021; Rädecker et al., 2021), however, increased GS activity has been reported for other cnidarians (Wang and Douglas, 1998; Lipschultz and Cook, 2002; Oakley et al., 2016). Both Petrou et al. (2021) and Rädecker et al. (2021) also observed concurrent increases in glutamate dehydrogenase (GDH) in bleached corals, which catabolizes glutamate to α-ketoglutarate, an intermediate of the Krebs Cycle. Thus, GDH indicates the degradation of amino acids for use as energy while producing ammonia as a byproduct. While we did not observe any change in GDH in bleached corals in this study, we propose two alternative nitrogen sources in bleached corals.
Urease (URE), an enzyme that catalyzes the conversion of urea to ammonia and carbon dioxide, provides another metabolic pathway for acquiring nitrogen and was significantly more abundant in the OL of bleached corals than non-bleached corals. Urea is thought to be an important source of nitrogen for corals (Grover et al., 2006; Crandall and Teece, 2012) because they can easily assimilate it from the surrounding environment where it accumulates from anthropogenic runoff and is a naturally produced metabolite from a variety of biological sources (Conover and Gustavson, 1999; Lomas et al., 2002; Glibert et al., 2006; McDonald et al., 2006). Grover et al. (2008), however, calculated that urea only constitutes about three percent of nitrogen uptake in Stylophora pistillata. Corals may also produce urea on their own, but the mechanism remains unclear because the complete enzymatic toolkit of the urea cycle has yet to be observed in corals (Streamer, 1980). Genes encoding four of the five enzymes of the urea cycle are present in the M. capitata genome, including: carbamoyl phosphate synthetase I (CPS1), ornithine transcarbamylase (OTC), argininosuccinate synthase (ASS) and argininosuccinate lyase (ASL), but the fifth enzyme, arginase (ARG1), has not been identified. Three of those enzymes, OTC, ASS, and ASL, were detected in the proteome identified in this study. Regardless, the elevated level of urease provides direct evidence that bleached corals rely on urea as a source of nitrogen, which may help to counteract the effects of decreased GS activity described above.
Here, we reveal that a critical source of nitrogen in bleached corals likely comes from the intracellular degradation of glycine betaine. Two enzymes involved in betaine degradation pathway, betaine-homocysteine S-methyltransferase (BHMT) and dimethylglycine dehydrogenase (DMGDH), were significantly more abundant in the OL of bleached corals than non-bleached corals. Enzymatic components of this pathway have been observed in multiple marine organisms, including marine invertebrates and corals (DeSalvo et al., 2012; Aguilar et al., 2019; Sproles et al., 2019). Glycine betaine is an important osmolyte because of its role in counteracting osmotic and other abiotic stressors (Rathinasabapathi, 2000; Jahn et al., 2006) and may have been produced in abundance in response to bleaching (DeSalvo et al., 2012). Recently, however, betaine has been proposed as a major source of nitrogen in reef-building corals where it can account for up to 16% of total nitrogen biomass (Ngugi et al., 2020). A recent metabolomics study on the bleaching history of M. capitata demonstrates that betaine-lipids are depleted in bleached samples, whereas historically non-bleached M. capitata is enriched in this metabolite (Roach et al., 2021). These proteomic profiles on the same coral species complement the metabolomic profiles of bleached vs. non-bleached M. capitata and provide direct evidence of active molecular pathways utilized by bleached corals to acquire N from stored betaine-lipids, potentially leading to its long-term persistence on reefs.
Through analyzing the proteomes of the OL and IC of bleached and non-bleached M. capitata, we investigated: (1) whether there were differential abundances of metabolic proteins in the OL and IC compartments, and (2) if metabolic pathways differ between these two compartments in response to bleaching. We identified several metabolic pathways and biological processes that are used in specific compartments or shared across compartments following bleaching stress. These included different strategies for metabolizing carbon, lipids and nitrogen by each compartment and high oxidative stress response and protein turnover in both compartments following bleaching. Our findings suggest that some molecular responses to bleaching are compartmentalized, which may be the most efficient way to continue functions specific to each compartment following bleaching. Future research should assess whether these compartmentalized responses to bleaching are linked to compartment-specific physiological functions. For example, does energy produced from lipids via the glyoxylate cycle help fuel reproductive output in the coral OL compartment? Or are there links between the products of the pentose phosphate pathway, such as NADPH and nucleotide generation, and calcification? Additionally, employing these compartmentalized strategies may convey resilience to bleaching and studies targeting these pathways, such as the glyoxylate cycle, the pentose phosphate pathway, protein turnover, urea assimilation and betaine degradation, should be undertaken to better understand their influence on coral resilience. Answers to questions like these will further our understanding of how and where distinct physiological functions in corals are affected by thermal stress and bleaching. Furthermore, elucidating the precise role of these metabolic pathways and cellular responses to bleaching could provide management with molecular-based tools, such as biomarkers, for conservation and restoration.
Data are available via ProteomeXchange with identifier PXD021243.
JA performed the experiments, analyzed the data, and wrote the manuscript. ET-S performed the experiments, analyzed the data, and edited the manuscript. TB performed the experiments and edited the manuscript. LR and JP-G designed, performed the experiments, and edited the manuscript. BN designed, performed the experiments, and wrote the manuscript. All authors contributed to the article and approved the submitted version.
This work was supported in part by the University of Washington’s Proteomics Resource (UWPR95794), NSF IOS-IEP 1655682 awarded to JP-G and BN, NSF IOS-IEP 1655888 to LR, and NSF GFRP DGE1762114 awarded to JA.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We offer our warmest gratitude to the Gates Coral Lab for hosting and supporting us during this experiment at the Hawai´i Institute of Marine Biology. We thank Brenner Wakayama, Gavin Kreitman, Melissa Jaffe and Sean Frangos for help with collecting and culturing the experimental corals. We thank Callum Backstrom for assistance with processing Symbiodiniaceae and chlorophyll data. We also thank Hyungwon Choi for assistance with statistical software.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.797517/full#supplementary-material
Aguilar, C., Raina, J.-B., Fôret, S., Hayward, D. C., Lapeyre, B., Bourne, D. G., et al. (2019). Transcriptomic analysis reveals protein homeostasis breakdown in the coral Acropora millepora during hypo-saline stress. BMC Genomics 20:148. doi: 10.1186/s12864-019-5527-2
Atkinson, M. J., Kotler, E., and Newton, P. (1994). Effects of water velocity on respiration, calcification, and ammonium uptake of a Porites compressa community. Pac. Sci. 48, 296–303.
Bachhawat, A. K., and Yadav, S. (2018). The glutathione cycle: glutathione metabolism beyond the γ-glutamyl cycle. IUBMB Life 70, 585–592.
Barshis, D. J., Ladner, J. T., Oliver, T. A., Seneca, F. O., Traylor-Knowles, N., and Palumbi, S. R. (2013). Genomic basis for coral resilience to climate change. Proc. Natl. Acad. Sci. U.S.A. 110, 1387–1392. doi: 10.1073/pnas.1210224110
Bellantuono, A. J., Granados-Cifuentes, C., Miller, D. J., Hoegh-Guldberg, O., and Rodriguez-Lanetty, M. (2012). Coral thermal tolerance: tuning gene expression to resist thermal stress. PLoS One 7:e50685. doi: 10.1371/journal.pone.0050685
Black, N. A., Voellmy, R., and Szmant, A. M. (1995). Heat shock protein induction in Montastraea faveolata and Aiptasia pallida exposed to elevated temperatures. Biol. Bull. 188, 234–240. doi: 10.2307/1542301
Brown, B., Downs, C., Dunne, R., and Gibb, S. (2002). Exploring the basis of thermotolerance in the reef coral Goniastrea aspera. Mar. Ecol. Prog. Ser. 242, 119–129. doi: 10.3354/meps242119
Brown, B. E., Le Tissier, M. D. A., and Dunne, R. P. (1994). Tissue retraction in the scleractinian coral Coeloseris mayeri, its effect upon coral pigmentation, and preliminary implications for heat balance. Mar. Ecol. Prog. Ser. 105, 209–218.
Cantin, N. E., Cohen, A. L., Karnauskas, K. B., Tarrant, A. M., and McCorkle, D. C. (2010). Ocean warming slows coral growth in the central red sea. Science 329, 322–325. doi: 10.1126/science.1190182
Chakravarti, L. J., Buerger, P., Levin, R. A., and van Oppen, M. J. H. (2020). Gene regulation underpinning increased thermal tolerance in a laboratory-evolved coral photosymbiont. Mol. Ecol. 29, 1684–1703. doi: 10.1111/mec.15432
Choi, H., Kim, S., Fermin, D., Tsou, C.-C., and Nesvizhskii, A. I. (2015). QPROT: statistical method for testing differential expression using protein-level intensity data in label-free quantitative proteomics. J. Proteomics 129, 121–126. doi: 10.1016/j.jprot.2015.07.036
Conover, R., and Gustavson, K. (1999). Sources of urea in arctic seas: zooplankton metabolism. Mar. Ecol. Prog. Ser. 179, 41–54. doi: 10.3354/meps179041
Cox, E. F. (2007). Continuation of sexual reproduction in Montipora capitata following bleaching. Coral Reefs 26, 721–724. doi: 10.1007/s00338-007-0251-9
Crandall, J. B., and Teece, M. A. (2012). Urea is a dynamic pool of bioavailable nitrogen in coral reefs. Coral Reefs 31, 207–214.
Cziesielski, M. J., Liew, Y. J., Cui, G., Schmidt-Roach, S., Campana, S., Marondedze, C., et al. (2018). Multi-omics analysis of thermal stress response in a zooxanthellate cnidarian reveals the importance of associating with thermotolerant symbionts. Proc. R. Soc. B Biol. Sci. 285:20172654. doi: 10.1098/rspb.2017.2654
Dauphin, Y. (2001). Comparative studies of skeletal soluble matrices from some Scleractinian corals and Molluscs. Int. J. Biol. Macromol. 28, 293–304. doi: 10.1016/S0141-8130(01)00124-6
D’Elia, C. F., Domotor, S. L., and Webb, K. L. (1983). Nutrient uptake kinetics of freshly isolated zooxanthellae. Mar. Biol. 75, 157–167. doi: 10.1007/BF00405998
DeSalvo, M., Sunagawa, S., Voolstra, C., and Medina, M. (2010). Transcriptomic responses to heat stress and bleaching in the elkhorn coral Acropora palmata. Mar. Ecol. Prog. Ser. 402, 97–113. doi: 10.3354/meps08372
DeSalvo, M. K., Estrada, A., Sunagawa, S., and Medina, M. (2012). Transcriptomic responses to darkness stress point to common coral bleaching mechanisms. Coral Reefs 31, 215–228. doi: 10.1007/s00338-011-0833-4
DeSalvo, M. K., Voolstra, C. R., Sunagawa, S., Schwarz, J. A., Stillman, J. H., Coffroth, M. A., et al. (2008). Differential gene expression during thermal stress and bleaching in the Caribbean coral Montastraea faveolata. Mol. Ecol. 17, 3952–3971. doi: 10.1111/j.1365-294X.2008.03879.x
Downs, C. A., Fauth, J. E., Halas, J. C., Dustan, P., Bemiss, J., and Woodley, C. M. (2002). Oxidative stress and seasonal coral bleaching. Free Radic. Biol. Med. 33, 533–543. doi: 10.1016/S0891-5849(02)00907-3
Drake, J. L., Mass, T., Haramaty, L., Zelzion, E., Bhattacharya, D., and Falkowski, P. G. (2013). Proteomic analysis of skeletal organic matrix from the stony coral Stylophora pistillata. Proc. Natl. Acad. Sci. U.S.A. 110, 3788–3793. doi: 10.1073/pnas.1301419110
Eng, J. K., Jahan, T. A., and Hoopmann, M. R. (2013). Comet: an open-source MS/MS sequence database search tool. Proteomics 13, 22–24. doi: 10.1002/pmic.201200439
Fermin, D., Basrur, V., Yocum, A. K., and Nesvizhskii, A. I. (2011). Abacus: a computational tool for extracting and pre-processing spectral count data for label-free quantitative proteomic analysis. Proteomics 11, 1340–1345. doi: 10.1002/pmic.201000650
Ferrier-Pagès, C., Rottier, C., Beraud, E., and Levy, O. (2010). Experimental assessment of the feeding effort of three scleractinian coral species during a thermal stress: effect on the rates of photosynthesis. J. Exp. Mar. Biol. Ecol. 390, 118–124. doi: 10.1016/j.jembe.2010.05.007
Fonseca, S. J., Marangoni, B. L. F., Marques, J. A., and Bianchini, A. (2019). Energy metabolism enzymes inhibition by the combined effects of increasing temperature and copper exposure in the coral Mussismilia harttii. Chemosphere 236:124420. doi: 10.1016/j.chemosphere.2019.124420
Frazier, M., Helmkampf, M., Bellinger, M. R., Geib, S. M., and Takabayashi, M. (2017). De novo metatranscriptome assembly and coral gene expression profile of Montipora capitata with growth anomaly. BMC Genomics 18:710. doi: 10.1186/s12864-017-4090-y
Garcia, G. D., Santos, E. O., Sousa, G. V., Zingali, R. B., Thompson, C. C., and Thompson, F. L. (2016). Metaproteomics reveals metabolic transitions between healthy and diseased stony coral Mussismilia braziliensis. Mol. Ecol. 25, 4632–4644. doi: 10.1111/mec.13775
Gardner, S. G., Raina, J.-B., Nitschke, M. R., Nielsen, D. A., Stat, M., Motti, C. A., et al. (2017). A multi-trait systems approach reveals a response cascade to bleaching in corals. BMC Biol. 15:117. doi: 10.1186/s12915-017-0459-2
Gates, R. D., and Edmunds, P. J. (1999). The physiological mechanisms of acclimatization in tropical reef corals. Am. Zool. 39, 30–43. doi: 10.1093/icb/39.1.30
Gestaut, D., Limatola, A., Joachimiak, L., and Frydman, J. (2019). The ATP-powered gymnastics of TRiC/CCT: an asymmetric protein folding machine with a symmetric origin story. Curr. Opin. Struct. Biol. 55, 50–58. doi: 10.1016/j.sbi.2019.03.002
Gladfelter, E. H. (1983). Circulation of fluids in the gastrovascular system of the reef coral Acropora cervicornis. Biol. Bull. 165, 619–636. doi: 10.2307/1541469
Glibert, P. M., Harrison, J., Heil, C., and Seitzinger, S. (2006). Escalating worldwide use of Urea – a global change contributing to coastal eutrophication. Biogeochemistry 77, 441–463. doi: 10.1007/s10533-005-3070-5
Grottoli, A. G., and Rodrigues, L. J. (2011). Bleached Porites compressa and Montipora capitata corals catabolize δ13C-enriched lipids. Coral Reefs 30:687. doi: 10.1007/s00338-011-0756-0
Grottoli, A. G., Rodrigues, L. J., and Palardy, J. E. (2006). Heterotrophic plasticity and resilience in bleached corals. Nature 440, 1186–1189. doi: 10.1038/nature04565
Grover, R., Maguer, J.-F., Allemand, D., and Ferrier-Pagès, C. (2006). Urea uptake by the scleractinian coral Stylophora pistillata. J. Exp. Mar. Biol. Ecol. 332, 216–225. doi: 10.1016/j.jembe.2005.11.020
Grover, R., Maguer, J.-F., Allemand, D., and Ferrier-Pages, C. (2008). Uptake of dissolved free amino acids by the scleractinian coral Stylophora pistillata. J. Exp. Biol. 211, 860–865. doi: 10.1242/jeb.012807
Gygi, S. P., Rochon, Y., Franza, B. R., and Aebersold, R. (1999). Correlation between Protein and mRNA abundance in Yeast. Mol. Cell. Biol. 19, 1720–1730. doi: 10.1128/MCB.19.3.1720
Haas, B. J., Papanicolaou, A., Yassour, M., Grabherr, M., Blood, P. D., Bowden, J., et al. (2013). De novo transcript sequence reconstruction from RNA-Seq: reference generation and analysis with Trinity. Nat. Protoc. 8:10.1038/nrot.2013.084. doi: 10.1038/nprot.2013.084
Hawkins, A. J. S. (1991). Protein turnover: a functional appraisal. Funct. Ecol. 5:222. doi: 10.2307/2389260
Hernández-Elizárraga, V. H., Olguín-López, N., Hernández-Matehuala, R., Ocharán-Mercado, A., Cruz-Hernández, A., Guevara-González, R. G., et al. (2019). Comparative analysis of the soluble proteome and the cytolytic activity of unbleached and bleached Millepora complanata (“Fire Coral”) from the Mexican Caribbean. Mar. Drugs 17:393. doi: 10.3390/md17070393
Hoegh-Guldberg, O., and Williamson, J. (1999). Availability of two forms of dissolved nitrogen to the coral Pocillopora damicornis and its symbiotic zooxanthellae. Mar. Biol. 133, 561–570. doi: 10.1007/s002270050496
Hughes, A. D., and Grottoli, A. G. (2013). Heterotrophic compensation: a possible mechanism for resilience of coral reefs to global warming or a sign of prolonged stress? PLoS One 8:e81172. doi: 10.1371/journal.pone.0081172
Isa, Y., and Okazaki, M. (1987). Some observations on the Ca2 +-binding phospholipid from scleractinian coral skeletons. Comp. Biochem. Physiol. Part B Comp. Biochem. 87, 507–512. doi: 10.1016/0305-0491(87)90045-9
Jahn, M. P., Cavagni, G. M., Kaiser, D., and Kucharski, L. C. (2006). Osmotic effect of choline and glycine betaine on the gills and hepatopancreas of the Chasmagnathus granulata crab submitted to hyperosmotic stress. J. Exp. Mar. Biol. Ecol. 334, 1–9. doi: 10.1016/j.jembe.2006.01.006
Jeffrey, S. W., and Humphrey, G. F. (1975). New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pflanz. 167, 191–194. doi: 10.1016/S0015-3796(17)30778-3
Jokiel, P. L., Hunter, C. L., Taguchi, S., and Watarai, L. (1993). Ecological impact of a fresh-water “reef kill” in Kaneohe Bay, Oahu, Hawaii. Coral Reefs 12, 177–184. doi: 10.1007/BF00334477
Kanehisa, M., and Goto, S. (2000). KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 28, 27–30. doi: 10.1093/nar/28.1.27
Kanehisa, M., Sato, Y., and Morishima, K. (2016). BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J. Mol. Biol. 428, 726–731. doi: 10.1016/j.jmb.2015.11.006
Keller, A., Nesvizhskii, A. I., Kolker, E., and Aebersold, R. (2002). Empirical statistical model to estimate the accuracy of peptide identifications made by MS/MS and database search. Anal. Chem. 74, 5383–5392. doi: 10.1021/ac025747h
Kenkel, C. D., Meyer, E., and Matz, M. V. (2013). Gene expression under chronic heat stress in populations of the mustard hill coral (Porites astreoides) from different thermal environments. Mol. Ecol. 22, 4322–4334. doi: 10.1111/mec.12390
Kornberg, H. L., and Krebs, H. A. (1957). Synthesis of cell constituents from C2-units by a modified tricarboxylic acid cycle. Nature 179, 988–991. doi: 10.1038/179988a0
Krueger, T., Hawkins, T. D., Becker, S., Pontasch, S., Dove, S., Hoegh-Guldberg, O., et al. (2015). Differential coral bleaching—Contrasting the activity and response of enzymatic antioxidants in symbiotic partners under thermal stress. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 190, 15–25. doi: 10.1016/j.cbpa.2015.08.012
Krupp, D. A., Jokiel, P. L., and Chartrand, T. S. (1992). “Asexual reproduction by the solitary scleractinian coral Fungia scutaria on dead parent coralla in Kaneohe Bay, Oahu, Hawaiian Islands,” in Proceeding of the 7th International. Coral Reef Symposium, Vol. 1, (Guam University Guam Press RH Richmond), 527–534
Lee, J. H., Kwon, E. J., and Kim, D. H. (2013). Calumenin has a role in the alleviation of ER stress in neonatal rat cardiomyocytes. Biochem. Biophys. Res. Commun. 439, 327–332. doi: 10.1016/j.bbrc.2013.08.087
Lesser, M. P. (1997). Oxidative stress causes coral bleaching during exposure to elevated temperatures. Coral Reefs 16, 187–192. doi: 10.1007/s003380050073
Lesser, M. P., Stochaj, W. R., Tapley, D. W., and Shick, J. M. (1990). Bleaching in coral reef anthozoans: effects of irradiance, ultraviolet radiation, and temperature on the activities of protective enzymes against active oxygen. Coral Reefs 8, 225–232. doi: 10.1007/BF00265015
Lipschultz, F., and Cook, C. (2002). Uptake and assimilation of 15N-ammonium by the symbiotic sea anemones Bartholomea annulata and Aiptasia pallida: conservation versus recycling of nitrogen. Mar. Biol. 140, 489–502. doi: 10.1007/s00227-001-0717-1
Lomas, M. W., Mark Trice, T., Glibert, P. M., Bronk, D. A., and McCarthy, J. J. (2002). Temporal and spatial dynamics of urea uptake and regeneration rates and concentrations in Chesapeake Bay. Estuaries 25, 469–482. doi: 10.1007/BF02695988
MacLean, B., Tomazela, D. M., Shulman, N., Chambers, M., Finney, G. L., Frewen, B., et al. (2010). Skyline: an open source document editor for creating and analyzing targeted proteomics experiments. Bioinform. Oxf. Engl. 26, 966–968. doi: 10.1093/bioinformatics/btq054
Maier, T., Güell, M., and Serrano, L. (2009). Correlation of mRNA and protein in complex biological samples. FEBS Lett. 583, 3966–3973.
Mailloux, R. J., McBride, S. L., and Harper, M.-E. (2013). Unearthing the secrets of mitochondrial ROS and glutathione in bioenergetics. Trends Biochem. Sci. 38, 592–602. doi: 10.1016/j.tibs.2013.09.001
Maor-Landaw, K., Karako-Lampert, S., Ben-Asher, H. W., Goffredo, S., Falini, G., Dubinsky, Z., et al. (2014). Gene expression profiles during short-term heat stress in the red sea coral Stylophora pistillata. Glob. Change Biol. 20, 3026–3035. doi: 10.1111/gcb.12592
Mayfield, A. B. (2020). Proteomic signatures of corals from thermodynamic reefs. Microorganisms 8:1171. doi: 10.3390/microorganisms8081171
Mayfield, A. B., Aguilar, C., Kolodziej, G., Enochs, I. C., and Manzello, D. P. (2021). Shotgun proteomic analysis of thermally challenged reef corals. Front. Mar. Sci. 8:547. doi: 10.3389/fmars.2021.660153
Mayfield, A. B., Wang, Y., Chen, C., Chen, S., and Lin, C. (2016). Dual-compartmental transcriptomic + proteomic analysis of a marine endosymbiosis exposed to environmental change. Mol. Ecol. 25, 5944–5958.
McDonald, M. D., Smith, C. P., and Walsh, P. J. (2006). The physiology and evolution of Urea transport in fishes. J. Membr. Biol. 212, 93–107. doi: 10.1007/s00232-006-0869-5
Mellacheruvu, D., Wright, Z., Couzens, A. L., Lambert, J.-P., St-Denis, N. A., Li, T., et al. (2013). The CRAPome: a contaminant repository for affinity purification–mass spectrometry data. Nat. Methods 10, 730–736. doi: 10.1038/nmeth.2557
Muller, E. M., Bartels, E., and Baums, I. B. (2018). Bleaching causes loss of disease resistance within the threatened coral species Acropora cervicornis. eLife 7:e35066. doi: 10.7554/eLife.35066
Muscatine, L., McCloskey, L. R., and Marian, R. E. (1981). Estimating the daily contribution of carbon from zooxanthellae to coral animal respiration. Limnol. Oceanogr. 26, 601–611. doi: 10.4319/lo.1981.26.4.0601
Nesvizhskii, A. I., Keller, A., Kolker, E., and Aebersold, R. (2003). A statistical model for identifying proteins by Tandem mass spectrometry. Anal. Chem. 75, 4646–4658. doi: 10.1021/ac0341261
Ngugi, D. K., Ziegler, M., Duarte, C. M., and Voolstra, C. R. (2020). Genomic blueprint of glycine betaine metabolism in coral metaorganisms and their contribution to reef nitrogen budgets. iScience 23:101120. doi: 10.1016/j.isci.2020.101120
NOAA Coral Reef Watch (2018). NOAA Coral Reef Watch Virtual Station for the Main Hawaiian Islands, Jan. 1, 2007-Dec. 31, 2017. College Park, MD: NOAA Coral Reef Watch.
Nunn, B. L., Slattery, K. V., Cameron, K. A., Timmins-Schiffman, E., and Junge, K. (2015). Proteomics of Colwellia psychrerythraea at subzero temperatures - a life with limited movement, flexible membranes and vital DNA repair: proteomics of microbes at subzero temperatures. Environ. Microbiol. 17, 2319–2335. doi: 10.1111/1462-2920.12691
Oakley, C. A., Ameismeier, M. F., Peng, L., Weis, V. M., Grossman, A. R., and Davy, S. K. (2016). Symbiosis induces widespread changes in the proteome of the model cnidarian Aiptasia. Cell. Microbiol. 18, 1009–1023.
Oakley, C. A., Durand, E., Wilkinson, S. P., Peng, L., Weis, V. M., Grossman, A. R., et al. (2017). Thermal shock induces host proteostasis disruption and endoplasmic reticulum stress in the model symbiotic cnidarian Aiptasia. J. Proteome Res. 16, 2121–2134. doi: 10.1021/acs.jproteome.6b00797
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., et al. (2020). vegan: Community Ecology Package. Available online at: https://CRAN.R-project.org/package=vegan (accessed February 11, 2021).
Ou, W.-J., Cameron, P. H., Thomas, D. Y., and Bergeron, J. J. M. (1993). Association of folding intermediates of glycoproteins with calnexin during protein maturation. Nature 364, 771–776. doi: 10.1038/364771a0
Pandey, A., and Mann, M. (2000). Proteomics to study genes and genomes. Nature 405, 837–846. doi: 10.1038/35015709
Pearse, V. B., and Muscatine, L. (1971). Role of symbiotic algae (zooxanthellae) in coral calcification. Biol. Bull. 141, 350–363. doi: 10.2307/1540123
Pernice, M., Meibom, A., Van Den Heuvel, A., Kopp, C., Domart-Coulon, I., Hoegh-Guldberg, O., et al. (2012). A single-cell view of ammonium assimilation in coral–dinoflagellate symbiosis. ISME J. 6, 1314–1324.
Petrou, K., Nunn, B. L., Padula, M. P., Miller, D. J., and Nielsen, D. A. (2021). Broad scale proteomic analysis of heat-destabilised symbiosis in the hard coral Acropora millepora. Sci. Rep. 11:19061. doi: 10.1038/s41598-021-98548-x
Polato, N. R., Altman, N. S., and Baums, I. B. (2013). Variation in the transcriptional response of threatened coral larvae to elevated temperatures. Mol. Ecol. 22, 1366–1382. doi: 10.1111/mec.12163
Puverel, S., Tambutté, E., Pereira-Mouriès, L., Zoccola, D., Allemand, D., and Tambutté, S. (2005). Soluble organic matrix of two Scleractinian corals: partial and comparative analysis. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 141, 480–487. doi: 10.1016/j.cbpc.2005.05.013
Rädecker, N., Pogoreutz, C., Gegner, H. M., Cárdenas, A., Roth, F., Bougoure, J., et al. (2021). Heat stress destabilizes symbiotic nutrient cycling in corals. Proc. Natl. Acad. Sci. U.S.A. 118:e2022653118. doi: 10.1073/pnas.2022653118
Rahav, O., Dubinsky, Z., Achituv, Y., and Falkowski, P. G. (1989). Ammonium metabolism in the zooxanthellate coral, Stylophora pistillata. Proc. R. Soc. Lond. B Biol. Sci. 236, 325–337. doi: 10.1098/rspb.1989.0026
Ramos-Silva, P., Kaandorp, J., Huisman, L., Marie, B., Zanella-Cléon, I., Guichard, N., et al. (2013). The skeletal proteome of the coral Acropora millepora: the evolution of calcification by co-option and domain shuffling. Mol. Biol. Evol. 30, 2099–2112. doi: 10.1093/molbev/mst109
Rathinasabapathi, B. (2000). Metabolic engineering for stress tolerance: installing osmoprotectant synthesis pathways. Ann. Bot. 86, 709–716.
Ricaurte, M., Schizas, N. V., Ciborowski, P., and Boukli, N. M. (2016). Proteomic analysis of bleached and unbleached Acropora palmata, a threatened coral species of the Caribbean. Mar. Pollut. Bull. 107, 224–232. doi: 10.1016/j.marpolbul.2016.03.068
Riffle, M., May, D., Timmins-Schiffman, E., Mikan, M., Jaschob, D., Noble, W., et al. (2017). MetaGOmics: a web-based tool for peptide-centric functional and taxonomic analysis of metaproteomics data. Proteomes 6:2. doi: 10.3390/proteomes6010002
Ritson-Williams, R., and Gates, R. D. (2020). Coral community resilience to successive years of bleaching in Kāne‘ohe Bay, Hawai‘i. Coral Reefs 39, 757–769. doi: 10.1007/s00338-020-01944-4
Roach, T. N. F., Dilworth, J., Martin H, C., Jones, A. D., Quinn, R. A., and Drury, C. (2021). Metabolomic signatures of coral bleaching history. Nat. Ecol. Evol. 5, 495–503. doi: 10.1038/s41559-020-01388-7
Roberts, J. M., Fixter, L. M., and Davies, P. S. (2001). Ammonium metabolism in the symbiotic sea anemone Anemonia viridis. Hydrobiologia 461, 25–35. doi: 10.1023/A:1012752828587
Rodrigues, L. J., and Grottoli, A. G. (2007). Energy reserves and metabolism as indicators of coral recovery from bleaching. Limnol. Oceanogr. 52, 1874–1882. doi: 10.4319/lo.2007.52.5.1874
Santos, S. R., Toyoshima, J., and Kinzie, R. A. 3rd. (2009). Spatial and temporal dynamics of symbiotic dinoflagellates (Symbiodinium: dinophyta) in the perforate coral Montipora capitata. Galaxea J. Coral Reef Stud. 11, 139–147.
Schlöder, C., and D’Croz, L. (2004). Responses of massive and branching coral species to the combined effects of water temperature and nitrate enrichment. J. Exp. Mar. Biol. Ecol. 313, 255–268. doi: 10.1016/j.jembe.2004.08.012
Seneca, F. O., Forêt, S., Ball, E. E., Smith-Keune, C., Miller, D. J., and van Oppen, M. J. H. (2010). Patterns of gene expression in a scleractinian coral undergoing natural bleaching. Mar. Biotechnol. 12, 594–604.
Seveso, D., Arrigoni, R., Montano, S., Maggioni, D., Orlandi, I., Berumen, M. L., et al. (2020). Investigating the heat shock protein response involved in coral bleaching across scleractinian species in the central Red Sea. Coral Reefs 39, 85–98. doi: 10.1007/s00338-019-01878-6
Sproles, A. E., Oakley, C. A., Matthews, J. L., Peng, L., Owen, J. G., Grossman, A. R., et al. (2019). Proteomics quantifies protein expression changes in a model cnidarian colonised by a thermally tolerant but suboptimal symbiont. ISME J. 13, 2334–2345. doi: 10.1038/s41396-019-0437-5
Streamer, M. (1980). Urea and arginine metabolism in the hard coral, Acropora acuminata. Comp. Biochem. Physiol. Part B Comp. Biochem. 65, 669–674. doi: 10.1016/0305-0491(80)90177-7
Stuhr, M., Blank-Landeshammer, B., Reymond, C. E., Kollipara, L., Sickmann, A., Kucera, M., et al. (2018). Disentangling thermal stress responses in a reef-calcifier and its photosymbionts by shotgun proteomics. Sci. Rep. 8:3524. doi: 10.1038/s41598-018-21875-z
Su, Y., Zhou, Z., and Yu, X. (2018). Possible roles of glutamine synthetase in responding to environmental changes in a scleractinian coral. Mol. Biol. Rep. 45, 2115–2124. doi: 10.1007/s11033-018-4369-3
Tomanek, L. (2014). Proteomics to study adaptations in marine organisms to environmental stress. J. Proteomics 105, 92–106. doi: 10.1016/j.jprot.2014.04.009
Traylor-Knowles, N., Rose, N. H., Sheets, E. A., and Palumbi, S. R. (2017). Early transcriptional responses during heat stress in the coral Acropora hyacinthus. Biol. Bull. 232, 91–100. doi: 10.1086/692717
Voolstra, C. R., Schnetzer, J., Peshkin, L., Randall, C. J., Szmant, A. M., and Medina, M. (2009). Effects of temperature on gene expression in embryos of the coral Montastraea faveolata. BMC Genomics 10:627. doi: 10.1186/1471-2164-10-627
Wang, J., and Douglas, A. E. (1998). Nitrogen recycling or nitrogen conservation in an alga-invertebrate symbiosis? J. Exp. Biol. 201, 2445–2453.
Ward, S., Harrison, P., and Hoegh-Guldberg, O. (2000). “Coral bleaching reduces reproduction of scleractinian corals and increases susceptibility to future stress,” in Proceeding of the 9th International Coral Reef Symposium. Bali Indones. 23-27 Oct. 2000, 7, (Bali).
Weston, A. J., Dunlap, W. C., Beltran, V. H., Starcevic, A., Hranueli, D., Ward, M., et al. (2015). Proteomics links the redox state to calcium signaling during bleaching of the scleractinian coral acropora microphthalma on exposure to high solar irradiance and thermal stress. Mol. Cell. Proteomics 14, 585–595. doi: 10.1074/mcp.M114.043125
Wright, R. M., Aglyamova, G. V., Meyer, E., and Matz, M. V. (2015). Gene expression associated with white syndromes in a reef building coral, Acropora hyacinthus. BMC Genomics 16:371. doi: 10.1186/s12864-015-1540-2
Xu, C., Bailly-Maitre, B., and Reed, J. C. (2005). Endoplasmic reticulum stress: cell life and death decisions. J. Clin. Invest. 115, 2656–2664. doi: 10.1172/JCI26373
Yam, A. Y., Xia, Y., Lin, H.-T. J., Burlingame, A., Gerstein, M., and Frydman, J. (2008). Defining the TRiC/CCT interactome links chaperonin function to stabilization of newly made proteins with complex topologies. Nat. Struct. Mol. Biol. 15, 1255–1262. doi: 10.1038/nsmb.1515
Yellowlees, D., Rees, T. A. V., and Fiti, W. K. (1994). Effect of ammonium-supplemented seawater on glutamine synthetase and glutamate dehydrogenase activities in host tissue and zooxanthellae of Pocillopora damicornis and on ammonium uptake rates of the zooxanthellae. Pac. Sci. 48, 291–295.
Keywords: coral bleaching, eco-physiology, metabolic pathways, calcification, glyoxylate, betaine, proteins, perforate skeleton
Citation: Axworthy JB, Timmins-Schiffman E, Brown T, Rodrigues LJ, Nunn BL and Padilla-Gamiño JL (2022) Shotgun Proteomics Identifies Active Metabolic Pathways in Bleached Coral Tissue and Intraskeletal Compartments. Front. Mar. Sci. 9:797517. doi: 10.3389/fmars.2022.797517
Received: 18 October 2021; Accepted: 11 January 2022;
Published: 14 February 2022.
Edited by:
Davide Seveso, University of Milano-Bicocca, ItalyReviewed by:
Rowan H. McLachlan, Oregon State University, United StatesCopyright © 2022 Axworthy, Timmins-Schiffman, Brown, Rodrigues, Nunn and Padilla-Gamiño. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jeremy B. Axworthy, amVyZW15YXhAdXcuZWR1
†These authors have contributed equally to this work and share last authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.