
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci. , 12 January 2023
Sec. Marine Pollution
Volume 9 - 2022 | https://doi.org/10.3389/fmars.2022.1025387
This article is part of the Research Topic Novel Insights into the Circulation, Biogeochemistry, and Ecosystem Function of the Gulf of Mexico: A Perspective from Mexican Waters in Support of Oil Spill Preparedness View all 17 articles
 Oscar Gabriel Hernández-Sánchez1†
Oscar Gabriel Hernández-Sánchez1† Víctor F. Camacho-Ibar2†
Víctor F. Camacho-Ibar2† Leticia Barbero3†
Leticia Barbero3† Juan Carlos Herguera4†
Juan Carlos Herguera4† Sharon Z. Herzka1*
Sharon Z. Herzka1*A synoptic gulf-wide isoscape of carbon (δ13C) and nitrogen (δ15N) in the Gulf of Mexico based on mesozooplankton (335-1000 um) was used as a proxy for the isotopic baseline and for calculating regional contributions of dissolved inorganic nitrogen sources. Mesozooplankton were sampled at 0-200 m (depth permitting) during the XIXIMI-06 and GOMECC-3 cruises held during the summer of 2017. A striking latitudinal gradient was found in δ15N values of zooplankton, with the highest values (10.4 ± 1.2‰) found over the northern shelf, and lowest values in the central, oligotrophic gulf (1.9 ± 0.5‰). To estimate the fractional contribution of potential nitrogen sources, the gulf was divided into six regions based on the spatial distribution of surface Chl-a, SST from remote sensing products and likely region-specific source contributions. A literature survey of (δ13C and δ15N values of particulate organic matter was used to characterize region-specific endpoint isotope ratios for use in a Bayesian isotope mixing model. Regional differences in δ15N values and the results of mixing models indicated nitrogen fixation is most likely an important source (45-74% contribution) of new nitrogen in the oceanic regions of the Gulf, the Loop Current and the Yucatan Shelf. In the oligotrophic gulf, the potential input of relatively light nitrate that reflects remineralization of surface layer POM or the excretion of light nitrogen by heterotrophs was insufficient to explain the low δ15N values found in the central Gulf, although it could account for about 40% of the N supporting secondary production. The high nitrogen isotope ratios found in the northern shelf were attributed to denitrified N (60%) and the inflow of heavy nitrogen from the Mississippi-Atchafalaya river system. Our results support the potential importance of fixed nitrogen in the deep waters of the Gulf of Mexico during the summer, characteristic for its highly stratified surface waters.
Stable isotope analysis (SIA) of carbon (δ13C) and nitrogen (δ15N) in organic matter, including organisms, is an ecological and biogeochemical tool that provides information on food web trophic structure, sources of primary production and nutrient inputs (Post, 2002; Layman et al., 2012; Sigman and Fripiat, 2019). δ13C values can be used to track carbon sources in the ocean, allowing for discrimination between phytoplankton, macroalgae and terrestrial C3 and C4 plants due to differences in photosynthetic pathways and the isotopic composition of the inorganic carbon pool, as well as growth rates (Fry and Sherr, 1989; Yamamuro et al., 1995; Ohkouchi et al., 2015 and references therein).
Variations in the isotopic composition of dissolved inorganic nitrogen (DIN) result from differences in the extent of isotope discrimination that takes place during assimilation, nitrification, denitrification, N2 fixation and remineralization (Sigman and Fripiat, 2019), and can also vary due to the mixing of water masses (Marconi et al., 2015). In turn, the isotopic composition of DIN is reflected in the composition of primary producers. Nitrate (NO3-) from subsurface waters, or new nitrogen, has a global mean isotopic composition (δ15N-NO3) of ca. 5‰ (Sigman and Casciotti, 2001), and is enriched in 15N compared with fixed N2 (-2 to 0‰; Carpenter et al., 1999; Montoya et al., 2002). Riverine DIN discharged in coastal areas has δ15N values around 5‰ due to terrestrial denitrification and inputs of waste waters and manure, but can occasionally be higher (Wissel and Fry, 2003; Bryantmason et al., 2013). Denitrification, which predominates in waters with low oxygen concentrations, has high isotope discrimination (~15-25‰) that leads to residual nitrate enriched in 15N compared with subsurface NO3- (Kritee et al., 2012). The assimilation of NO3- by primary producers has an isotope discrimination of ~5‰ when only part of the NO3- pool is assimilated, and the residual nitrate is therefore enriched in 15N. However, when all of the NO3- is assimilated, as is frequently the case in the euphotic layer of oligotrophic systems (Somes et al., 2010), the water column behaves like a closed system and the isotopic composition of primary producers will reflect that of the DIN pool. Remineralization of organic matter leads to lower δ15N-NO3 values than the isotopic composition of particulate organic matter (δ15N-POM), with isotope discrimination of around 3‰ (Sigman and Fripiat, 2019). Also, zooplankton excrete ammonium with lower δ15N values compared with the isotopic composition of zooplankton ~2-3‰ (Checkley Jr. and Miller, 1989; Vanderklift and Ponsard, 2003).
In marine systems, δ15N-POM has been used to infer sources and geochemical processes underlying the nutrient pool supporting primary and secondary production and is considered a proxy for δ15N-phytoplankton (Waite et al., 2007; Kolasinski et al., 2012). However, the isotopic composition of POM varies over small spatial and temporal scales due to changes in phytoplankton community composition, the rapid assimilation of pulsed nutrient inputs and rapid isotope turnover rates of small organisms, with isotope integration times of the order of days (Kürten et al., 2013; Lorrain et al., 2015). This implies intensive sampling is necessary for its adequate characterization of carbon and nitrogen sources at the base of the food web and can complicate the interpretation of the sources of variation.
In comparison, mesozooplankton integrate the isotopic composition of their food sources through time, smoothing the variation in POM (Hou et al., 2013). Zooplankton consume phytoplankton and microzooplankton and hence feed at the base of the food web (Turner, 2015). The δ13C and δ15N values of mesozooplankton can therefore serve as a proxy for the isotopic baseline since they integrate the isotopic composition over longer periods (weeks) than POM (Gorokhova and Hansson, 1999; Schmidt et al., 2004). The δ13C and δ15N values of zooplankton have been successfully used to infer nitrogen sources and estimate their fractional contribution to secondary production using mixing models (Landrum et al., 2011; Chen et al., 2018; Hernández-Sanchez et al., 2022). They have also been used to establish regional and latitudinal patterns in the biogeochemical processes that dominate nitrogen cycling (Mompeán et al., 2013; McMahon et al., 2013 and references therein), examine seasonal changes in N sources (El-Sabaawi et al., 2013; Kurle and McWhorter, 2017; Troina et al., 2020) and evaluating the importance of N derived from N2 fixation vs. subsurface nitrogen (Landrum et al., 2011). For example, copepods have higher δ13C values onshore compared with offshore, which has been linked to differences in the community composition of phytoplankton (Perry et al., 1999). In the Red Sea, high δ13C and low δ15N values of zooplankton in the northern region are associated with the presence of the nitrogen-fixing Trichodesmium, whereas lower δ13C and higher δ15N values toward the south are associated with N fluxes of subsurface water (Kürten et al., 2016). Differences in zooplankton δ15N values have been found between anticyclonic and cyclonic eddies, which have been attributed to whether subsurface DIN reaches the euphotic layer and differences in the contribution of N2 fixation (Waite et al., 2007; Henschke et al., 2015; Hernández-Sánchez et al., 2022).
Estimating source contributions based on zooplankton SIA and isotope mixing models requires estimates of the trophic discrimination factor (TDF), which is an empirically derived quantitative estimate of the isotope discrimination between consumers and their food sources. TDFs are consistently small (~1‰) in the case of δ13C values; carbon sources can thus be tracked through the food web (Post, 2002). On the other hand, δ15N values can be used to track N sources as well as for estimating trophic level, with given TDFs of around 2-4‰ (Post, 2002; McCutchan et al., 2003; Vanderklift and Ponsard, 2003). The trophic enrichment in the heavy isotope observed in consumer tissues implies that the nitrogen excreted by heterotrophs is lighter (Vanderklift and Ponsard, 2003).
The Gulf of Mexico (GM) is a marginal ocean basin surrounded by Mexico, the United States, and Cuba. The circulation and hydrography of the central gulf’s upper waters (0-1000 m) are strongly influenced by the Loop Current (LC), which forms from the Yucatan Current and transports water from the Caribbean Sea through the Yucatan Straight and which exits the gulf through the Straits of Florida (Hamilton et al., 2018). The LC detaches anticyclonic mesoscale eddies periodically, and these Loop Current eddies (LCEs) transport water masses into the central and western GM that mix with gulf waters during their transit and when they dissipate near the western slope (Oey et al., 2005, Sosa-Gutiérrez et al., 2020, Cervantes-Díaz et al., 2022). During the summer, high SST and weaker winds lead to stratification and a shallow mixed layer, limiting vertical fluxes of subsurface the NO3- in the central GM (Müller-Karger et al., 2015; Pasqueron De Fommervault et al., 2017). Within the Bay of Campeche in the southern gulf, there is a semi-permanent cyclonic eddy that pumps subsurface water toward the euphotic layer due to a shallowing of the pycnocline (Klein and Lapeyre, 2009; Pérez-Brunius et al., 2013; Durán-Campos et al., 2017).
Some studies indicate N2 fixation may be the most important source of new N supporting secondary production in the oceanic GM, particularly during the summer months or during blooms of diazotrophs such as Trichodesmium (Mulholland et al., 2006; Holl et al., 2007; Landrum et al., 2011; Hunt et al., 2016). Over the Yucatan shelf, positive anomalies of chl a have been reported for the inner shelf during the summer months (Zavala-Hidalgo et al., 2006) when upwelled water resulting from the interaction between the Loop Current and slope is then transported westward across the shelf (Merino, 1997; Jouanno et al., 2018). Along the shelves, nutrient inputs from rivers, in particular, the Mississippi-Atchafalaya River System (MARS) in the north and the Grijalva-Usumacinta in the south, can lead to high inputs of terrestrial carbon and inorganic nitrogen from various sources that include manure, wastewater treatment, and fertilizers (Alexander et al., 2008). In MARS, high nutrient inputs have led to phytoplankton blooms and high water column respiration, which coupled with water column stratification leads to decreases dissolved oxygen concentrations in the subsurface and in sediments that favor denitrification (DiMarco et al., 2000; Rabalais et al., 2002).
Specifically, low δ15N values of zooplankton (1.8‰ to 4‰) have been associated with high N2 fixation rates measured in the GM during the summer (Holl et al., 2007), and Dorado et al. (2012) reported low δ15N values of zooplankton in the northern oceanic GM (2.8 ± 1.4‰) suggesting that N2 fixation by Trichodesmium is an important N source in the LC and in LCE. In contrast, Knapp et al. (2021) and Kelly et al. (2021), based on indirect estimates, reported low N2 fixation rates in the GM as a whole, suggesting that N2 fixation is too small to be considered as a relevant N source; these authors also suggested that the low δ15N values of POM were due to remineralization of primary producers supported by subsurface nitrate (Knapp et al., 2021; Kelly et al., 2021). On the other hand, cyclonic eddies with divergent conditions at their core lead to a shallower nitracline and pumping of subsurface NO3- to the euphotic zone (Lee-Sánchez et al., 2022), while transport of NO3- toward the surface can also occur due to a deepening of the mixed layer in winter (Müller-Karger et al., 2015; Damien et al., 2018). Important inputs of N into the gulf also occur through the discharge of the Mississippi-Atchafalaya river system, which is the major source of terrestrial organic matter and nutrients to the northern GM shelf. Low δ13C values of POM associated with the contribution of terrestrial C3 plants have been reported compared with those for the open ocean of the northern shelf GM (Bianchi et al., 2007; Dorado et al., 2012; Cai et al., 2015; Wells et al., 2017). In the northern GM shelf, a shift from terrestrial to marine sources has been reported with lower δ13C values of POC close to the shore and higher δ13C values of POC with increased salinity (Cai et al., 2012). On the other hand, the higher δ15N values from the Mississippi-Atchafalaya River have been associated with different sources including manure and wastewater in the upper Mississippi River and tributaries, and denitrification in the lower basin (Chang et al., 2002; Bryantmason et al., 2013). Hypoxic conditions have been reported in the Texas and Louisiana coasts during the spring and summer (Rabalais et al., 2002; Rabalais et al., 2001; Bianchi et al., 2010), which favors denitrification in the shelf sediments and increases the δ15N values of the remnant NO3- pool (Heaton, 1986). In the Mississippi Sound, δ15N values of between 8.2‰ and 11.1‰ have been reported for phytoplankton and POM (Chanton and Lewis, 1999; Moncreiff and Sullivan, 2001). On the western Florida Shelf, a latitudinal pattern in δ15N values of POM has been documented, with higher values on the northernmost samples (~6-7‰) compared with those from the south (~3-5‰) and δ13C values with a nearshore-offshore pattern with higher values (~-26‰) in the coastal samples compared with oceanic samples (~-24‰; Radabaugh et al., 2013). In the southern GM, the Grijalva-Usumacinta River system represents the most important source of land-derived freshwater and nutrients and a mean δ15N value of POM = 4.6 ± 0.5‰ in the lower river region has been reported (Sepúlveda-Lozada et al., 2015).
Isoscapes, or maps that reflect the spatial distribution of isotope ratios, can reveal biogeochemical gradients that allow for inferences regarding nutrient cycling, migration patterns of large organisms and the detection of isotopic baseline shifts due to changes in nutrient sources (Hobson et al., 2010; McMahon et al., 2013; Radabaugh, et al., 2013). For example, zooplankton sampled in the Southern Ocean showed a latitudinal gradient, with low δ15N values in the north that reflected a productivity gradient due to iron limitation (Brault et al., 2018). McMahon et al. (2013) generated isoscapes for the Atlantic Ocean basin based on the δ13C and δ15N values of zooplankton and found low (~0-2‰) δ15N values in the subtropical western Atlantic and the Caribbean Sea that were attributed to previously documented inputs of fixed nitrogen (Montoya et al., 2002; Capone et al., 2005). Recently, Le-Alvarado et al. (2021) used basin-wide isoscapes for the GM based on δ13C and δ15N values of zooplankton collected during the summer of 2017 to infer the foraging habitat and trophic position of migratory yellowfin tuna caught in the southern GM. They found a pronounced latitudinal gradient in the δ15N values of zooplankton, with the highest δ15N values in the northern gulf and lower values in the central oligotrophic gulf, but did not examine source contributions. More generally, studies in the GM have focused on the region of influence of the Loop Current (Howe et al., 2020; Knapp et al., 2021), the northwestern gulf (Holl et al., 2007), the west Florida Shelf (Mullholand et al., 2006; Mullholand et al., 2014), and the central, oligotrophic region (Hernández-Sánchez et al., 2022).
We estimated the fractional contribution of different N sources throughout the GM based on the δ13C and δ15N values of zooplankton reported by Le-Alvarado et al. (2021) for the summer of 2017. The GM was divided into regions based on likely N sources, known predominant circulation patterns, and previous regionalization’s of the gulf based on the distribution of surface chl a concentrations. The relative importance of each N source was estimated using Bayesian isotope mixing models in which isotopic endpoints were obtained from literature-derived regional POM isotope ratios. Given that subsurface δ15N-NO3 is lower in the western gulf compared with the eastern gulf and western Atlantic Ocean (Holl et al., 2007; Marconi et al., 2015; Howe et al., 2020), we evaluated how these differences impact regional source contributions. We hypothesized that in the northern shelf of the GM denitrification is the most important source, while in the central GM and the region of influence of the Loop Current, N2 fixation is the most important source. In contrast, in the Bay of Campeche NO3- should be the predominant source due to regional upwelling, cross-shelf transport of river inputs and the semi-permanent cyclonic eddy. Lastly, we examine whether isotope ratios of zooplankton reflect those of POM and nitrate integrated based on samples collected at the same stations in which zooplankton were sampled. Our study provides the first gulf-wide assessment of the relative importance of nitrogen sources in the GM and provides a baseline against which to compare future regional changes in nutrient loads and increased stratification due to near-surface warming linked to climate change.
Two oceanographic cruises, XIXIMI-06 and GOMECC-3, were conducted concurrently during August and September 2017. A total of 93 mesozooplankton samples were collected throughout GM with oblique tows using a 60 cm diameter bongo equipped with 335 µm mesh nets (Figure 1, see also Supplementary Figure 1S for sea surface height anomalies during the cruises). Tows were deployed to a depth of 200 m except for shallower shelf stations, where tows reached about 20 m off the bottom. Once on board, 20% by volume was subsampled with a Hempel-Stempel pipette and frozen in Whirl-Pak bags at -20°C. The second sample for the bongo net was fixed in ethanol and subsequently used to estimate zooplankton biovolume (ml 100 m-3).
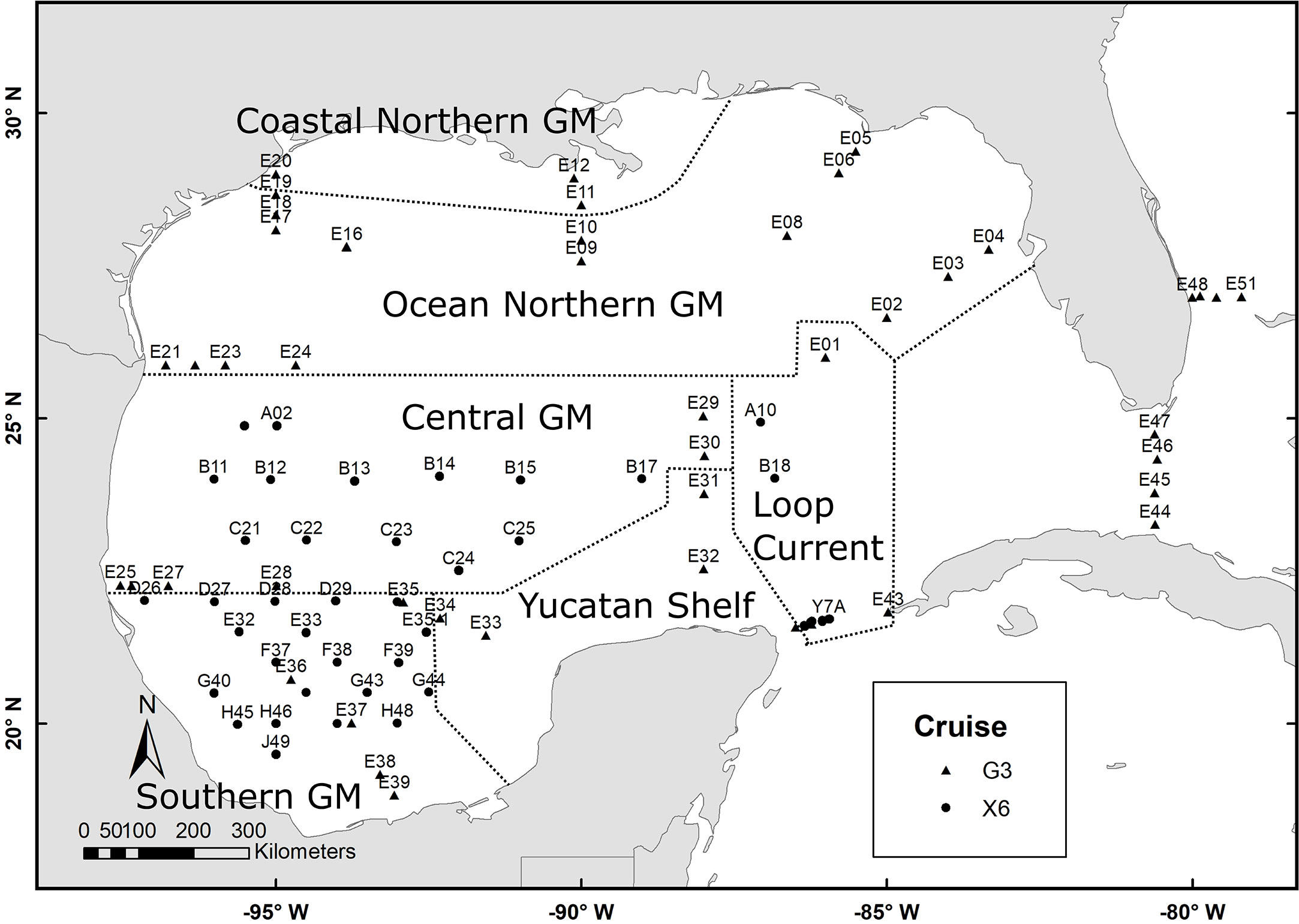
Figure 1 Zooplankton sampling stations in the Gulf of Mexico during the XIXIMI-06 (X6) and GOMECC-03 (G3) cruises were held during the summer of 2017. The six regions for which fractional nitrogen source contributions o were calculated are depicted.
Water samples for the measurement of the isotopic composition of suspended POM were collected during XIXIMI-06 (covering the central and southern GM’s deep water region; Figures 1; Supplementary Figure 1S) as a proxy for phytoplankton. Water was obtained at each of three depths (10, 20 and 50 m or 10, 50 and the depth of maximum of fluorescence depending on the station) with Niskin bottles mounted on a SeaBird rosette. Samples from each depth were pooled (7-29 L total) and filtered onto pre-combusted (500° C for 4 h) GF/F filters and frozen at -20°C. Water samples for measurements of nitrate concentrations were collected and processed as described in Camacho-Ibar et al. (2021).
Zooplankton samples were prepared for SIA as described in Le-Alvarado et al. (2021). Briefly, zooplankton were defrosted and rinsed in distilled water to remove dissolved inorganic carbon. Mesozooplankton <1000 µm were separated using nitex mesh sieves cleaned with ethanol, dried, ground and placed in tin capsules and sent to the Stable Isotope Facility at the University of California, Davis. The upper size of the zooplankton analyzed was limited to 1000 µm to minimize the contribution of higher trophic level organisms. Samples were processed with a PDZ Europa ANCA-GSL elemental analyzer interfaced with a PDZ Europa 20-20 isotope ratio mass spectrometer. The standard deviation of internal standards (glutamic acid, bovine liver, enriched alanine, and nylon) was between 0.04‰ and 0.07‰ for δ13C, and 0.05‰, 0.08‰, for δ15N, respectively. For suspended POM samples, filters were lyophilized and one half was treated with acid by fuming (1 M HCl treatment) to eliminate carbonates and analyzed for δ13C, while the other half was analyzed untreated for δ15N values. Isotope ratios of suspended POM were analyzed with an elemental analyzer interfaced with a DELTA V isotope ratio mass spectrometer at CICESE. The standard deviation of the internal standard (glutamic acid and calcium carbonate) was 0.05‰ for δ13C and 0.06‰ for δ15N, respectively.
Isotope ratios are reported in delta (δ) notation calculated relative to Vienna Pee Dee Belemnite for δ13C and atmospheric nitrogen for δ15N, using the following equation:
where X is 13C or 15N, and Rsample and Rstandard are the relative abundance of heavy to light isotope ratio (13C/12C or 15N/14N) for the sample and the standard, respectively. δ13C and δ15N values are reported in parts per thousand (‰).
We used the δ13C and δ15N values of zooplankton to create isoscapes that allowed for a spatially explicit visualization of the isotopic baseline. Zooplankton isoscapes were generated using ArcMap version 10.1 using an ordinary point kriging interpolation and a tetraspherical semivariogram model (Supplementary Table 1). The δ13C and δ15N values of suspended POM were also used to create an isoscape. These isoscapes were generated with QGIS version 3.16.6 using IWD interpolation rather than ordinary point kriging because the semivariogram indicated a poor fit.
The GM has been regionalized based on the spatial distribution of surface chlorophyll-a and SST from remote sensing products. Most studies coincide in differentiating the shelf of the northern and southern GM from the oligotrophic central and southern (Bay of Campeche) oceanic regions (Salmerón-García and Zavala-Hidalgo, 2011; Callejas-Jiménez et al., 2012; Damien et al., 2018). The deep water region of the central GM is generally considered oligotrophic because NO3- concentrations are near or below the detection limit in the upper euphotic layer and chl-a concentrations are relatively low, with values of around 0.1-0.7 mg m-3 at the deep chlorophyll maximum that is 50-80 m deep (Biggs, 1992; Salas-de-León et al., 2004; Pasqueron De Fommervault et al., 2017).
The GM was divided into 6 regions based on likely region-specific source contributions and considering similarities in the spatial distribution of zooplankton isotope ratios (Figure 1). This regionalization is based on the premise that while nitrate and fixed N2 may support primary and secondary production throughout the gulf, other sources have a more limited regional contribution. Inputs from the Mississippi-Atchafalaya river system, rivers that drain onto the western Florida shelf, N from denitrification in the hypoxic region of the northern shelf, and the Grijalva-Usumacinta River were considered as regional sources with limited spatial extent. Although studies indicate that river inputs can reach the central GM under certain conditions, such as when LCE’s interact with the gulf’s northern slope (Otis et al., 2019) and during the fall in the Bay of Campeche due to the convergence of currents over the continental shelf (Martínez-López and Zavala-Hidalgo, 2009), our approach considers these inputs to be limited on a regional scale.
The regional source contributions were calculated based on published literature on values of the isotopic composition of suspended POM. The northern GM (north of 26° N) was divided into the coastal (NGMc) and oceanic northern Gulf of Mexico (NGMo; Table 1). The likely N sources in these two regions are the Mississippi-Atchafalaya river system, river inputs to the western Florida shelf, denitrified N from the hypoxic region and the two Gulf-wide sources (fixed N2 and subsurface nitrate). For the southern GM (SGM), the Grijalva-Usumacinta river system (GUS) and the two gulf-wide sources were included in the model. Finally, for the central GM (CGM), the Yucatan Shelf (YS) and the region of influence of the Loop Current (LC), only the Gulf-wide sources were included (Table 1).
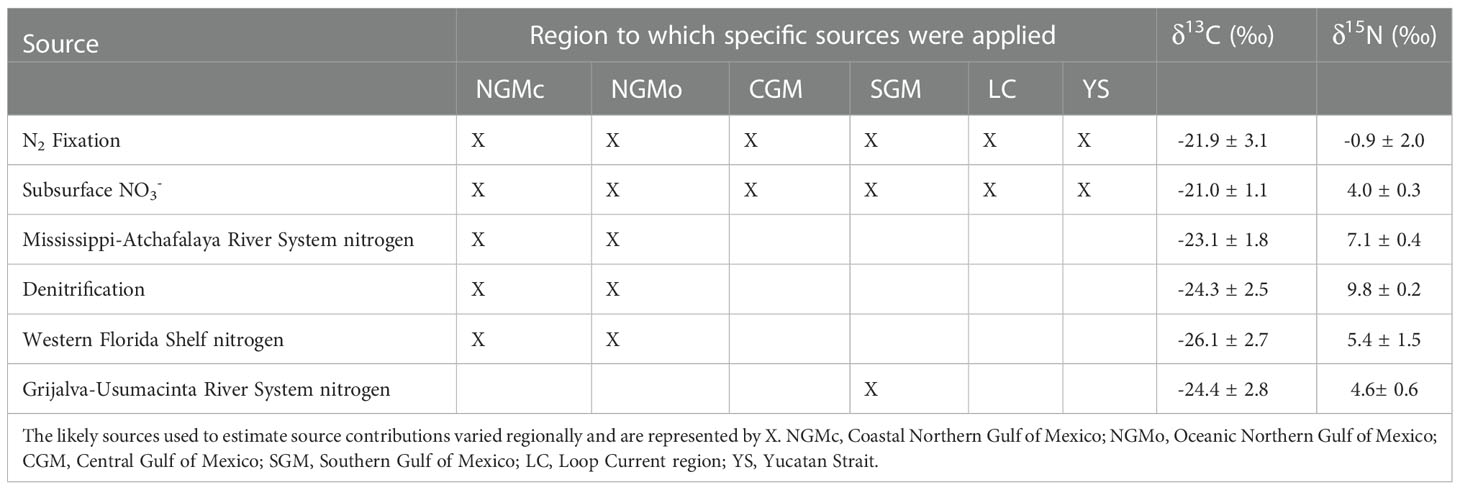
Table 1 Mean ± SD carbon and nitrogen isotope ratios of source POM used as endpoints in isotope mixing models.
The mean δ13C and δ15N values of suspended POM for each region were obtained from a literature review (see Tables 1; 2S and references therein). Data were available for Mississippi-Atchafalaya river system inputs (Macko et al., 1984; Wissel and Fry, 2005; Bianchi et al., 2007; Dorado et al., 2012), regions with high inputs of denitrified N (Chanton and Lewis, 1999; Moncreiff and Sullivan, 2001), the West Florida shelf (Gu et al., 2001; Radabaugh et al., 2013), the Grijalva-Usumacitna river system (Sepulveda-Lozada et al., 2015), POM that reflects subsurface NO3- (Dorado et al., 2012), and POM linked to N2 fixation (Holl et al., 2007; Wells and Rooker, 2009; Dorado et al., 2012). Isotope ratios presented in the figures were extracted using Plot Digitizer software.
To test for differences in δ13C and δ15N values of zooplankton between regions, Shapiro and Levene’s tests were performed to assess the normality and homoscedasticity of the data, respectively. Since the data did not meet the assumptions required for parametric analysis, a Kruskall-Wallis test was used. A Wilcoxon post-hoc test was applied to evaluate differences between regions. Sampling stations in the Florida Strait and east Florida shelf from the GOMECC-3 cruise were not included in statistical analyses as they lie outside the six regions considered. The relationships between δ15N values of zooplankton v.s suspended POM and integrated NO3- concentrations (0-200 m) was examined For the XIXIMI region with a correlation analysis. In addition, we examined the correlation between δ15N values of zooplankton, surface chl a concentrations obtained from Marine Copernicus Service (https://marine.copernicus.eu/), and %C and % N and C: ratios of zooplankton for the entire Gulf.
Bayesian mixing models were applied to estimate the contribution of different N sources to zooplankton using the SIMMR package in R (Parnell et al., 2010). Bayesian mixing models incorporate estimates of the variability in the isotope composition of the sources and trophic discrimination factors. The model inputs were (1) the δ15N and δ13C values of zooplankton, (2) the isotope ratios of suspended POM (mean and SD values, see Table 1) of the potential N sources for each region, and (3) the trophic discrimination factor for crustacean zooplankton of δ13C = 1.0 ± 0.6‰, δ15N= 2.0 ± 0.5‰ (Davenport and Bax, 2002; Vanderklift and Ponsard, 2003; Henschke et al., 2015); crustacean zooplankton are typically the dominant component of the community in the GM (Hopkins, 1982; Martinez et al., 2021). The model was applied to the zooplankton isotope ratios for all stations within each region. Model inputs for SIMMR did not include concentration dependence and they were run with 100,000 iterations and 10,000 burnins (Phillips et al., 2014). Subsequently, maps with the fractional contribution for each source and station within a region were generated using the IDW (Inverse Distance Weighting) interpolation method in QGIS version 3.16.6 for visualization purposes. Interpolations are meant to depict general patterns and may not add to 100% if all source contributions are considered. The mean ± standard deviation (SD) of the estimated contribution for each station was also calculated for each region.
To evaluate the potential effect of variations in the isotopic composition of nitrate reaching the euphotic layer, the Bayesian mixing model was applied using δ15N-NO3 values of subsurface nitrate considering three scenarios: scenario 1 with a value of 4.0 ± 0.3 ‰ based on POM isotope ratios for the deep waters of the GM (see above), and which is within the range of subsurface δ15N-NO3 values between 400 to 600 m reported for the western Atlantic Ocean (Knapp et al., 2008); scenario 2 with a value of 3.5 ± 1.1 ‰ corresponding to nitrate of water samples collected in the northern GM at 26° N and ~91-92° W at depths of 100-200 m (Howe et al., 2020); and scenario 3 with a value of 1.9 ± 0.8 ‰ reported for nitrate in the western GM (26-27° N and 95-96° W) for samples collected at ~200 m depth (Holl et al., 2007).
Average carbon isotope ratios of zooplankton exhibited a limited variability spanning 2.1‰ and were mostly consistent with phytoplankton-derived primary production (Figure 2A). Mean δ13C values were lowest (-21.7 ‰) in the NGMc, followed by NGMo with a mean of -20.9‰. Relatively low values (-21.3‰) were also observed in the SGM, especially at stations close to Grijalva-Usumacinta river. Higher δ13C values (-17.3 to -18.4‰) were found at some stations in the CGM (northwest of the Yucatan Peninsula), which could be due to the contribution of remineralization of C from seagrass or macroalgae from the Yucatan shelf or Trichodesmium that has high δ13C values compared with phytoplankton (Carpenter et al., 1997).
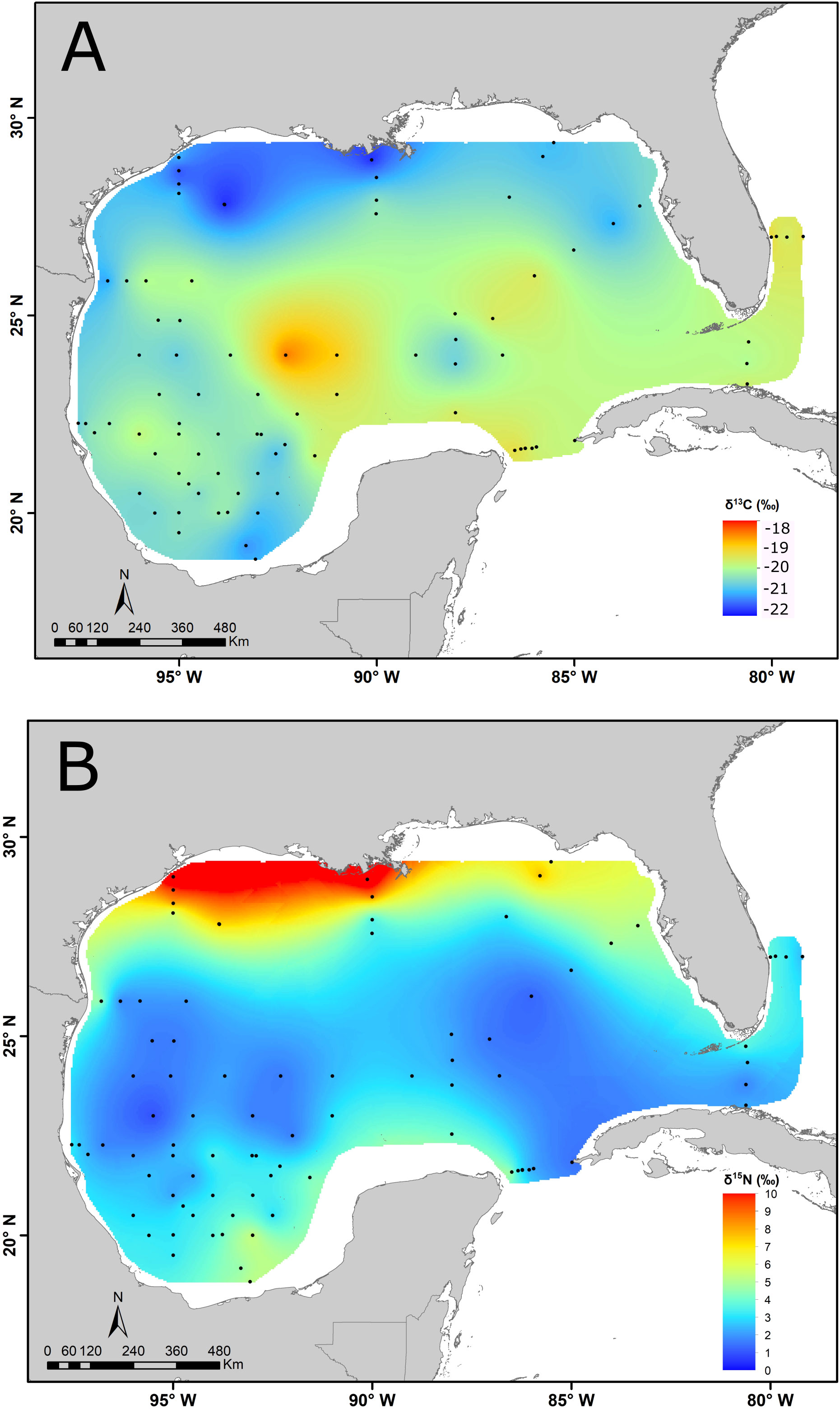
Figure 2 δ13C (A) and δ15N (B) isoscapes of zooplankton < 1000 um sampled in the Gulf of Mexico during the summer of 2017. Dots indicate sampling stations. Maps adapted from Le-Alvarado et al. (2021). (A) represent the d13C-POM values and B represent te d15N-POM values.
Mean δ13C values of zooplankton varied significantly among regions (KW, X2 = 23.887, df = 5, p-value < 0.001). However, post-hoc pairwise comparisons indicated that only stations from the LC region had δ13C values that were significantly higher than those from the NGMo and SGM (1.3 and 0.8‰, respectively); all other comparisons did not differ statistically (Table 2).
Zooplankton δ15N values showed a strong latitudinal gradient and differed significantly among regions (X2 = 42.723, df = 5, p = <0.001, Table 2). The highest mean δ15N values were observed in the NGMc and NGMo (10.4 ± 1.2‰ and 5.0 ± 2.2‰, respectively; Figure 2 and Table 3) suggesting the assimilation of N from MARS runoff and denitrification. In contrast, the lowest values were for CGM and LC (1.9 ± 0.5‰ and 2.3 ± 0.6‰, respectively), which indicates the contribution of N2 fixation. The SGM and YS had significantly higher values (3.1 ± 0.8‰ and 3.8 ± 0.9‰) than the central gulf. The significant differences in zooplankton isotope ratios among regions support the need of considering regional N sources to estimate contributions.
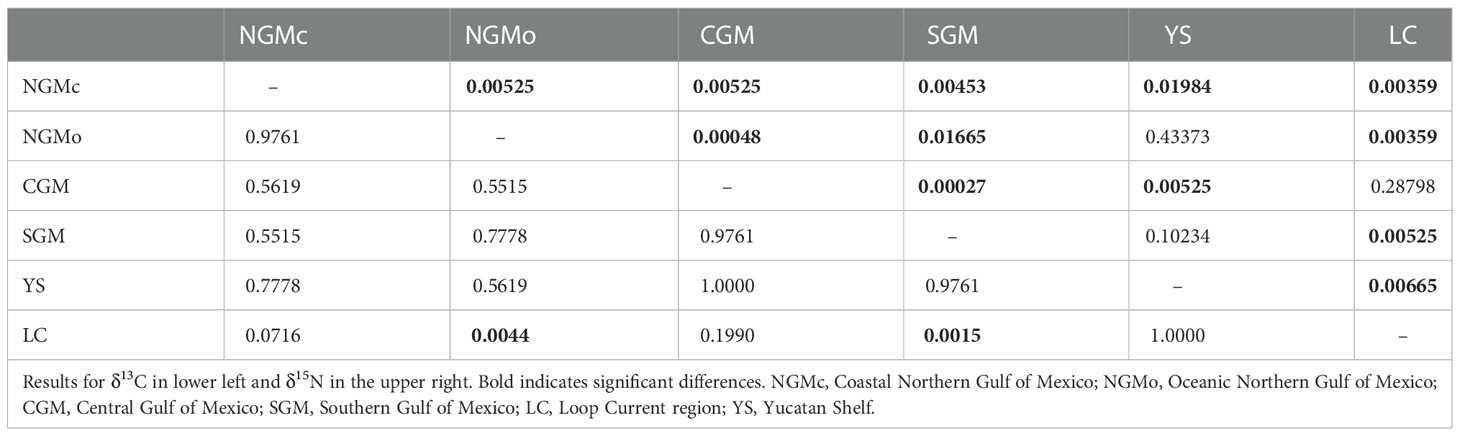
Table 3 Mean ± SD carbon and nitrogen stable isotope ratios of zooplankton collected in six regions of the Gulf of Mexico during the summer of 2017.
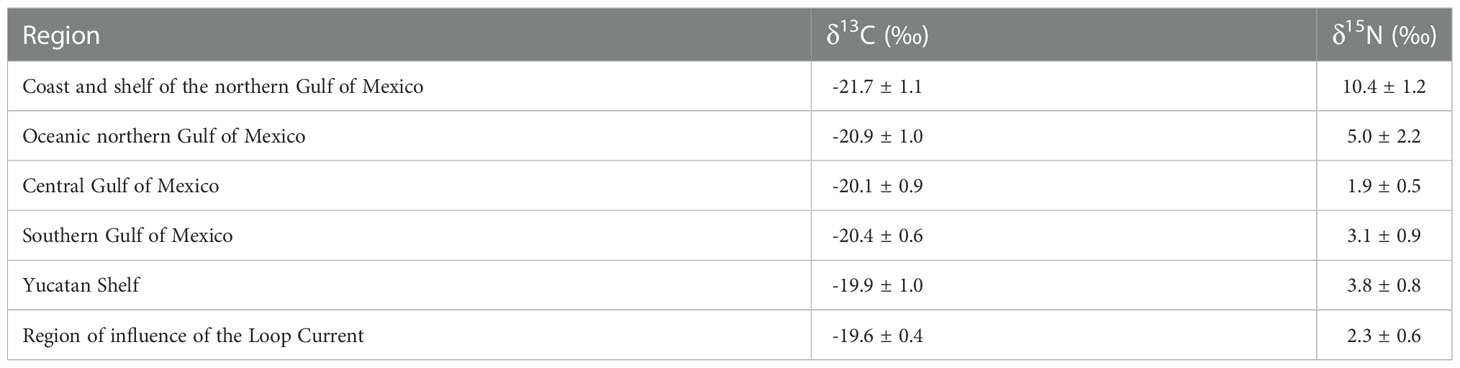
Table 2 Wilcoxon signed-rank post-hoc pairwise comparisons test of the isotopic composition of zooplankton collected in different regions of the Gulf of Mexico.
The δ13C suspended POM values during the XIXIMI-06 cruise ranged from -25.3 to -21.3‰, but a spatial pattern was not evident (Figure 3A; Table 3S). Mean δ13C suspended POM values did not show statistical differences among CGM, SGM and LC (F= 0.584, p=0.563). In contrast, the δ15N suspended POM values ranged from -1.9 to 3.2‰ (Figure 3B), with lower values in the western GM, in stations with influence by the remnants of an LC anticyclonic eddy (Poseidon; Figure 1S). In the SGM the values were higher compared with the western GM, especially for stations close to the shelf that could reflect cross-shelf transport of DIN from Grijalva-Usumacinta River or subsurface nitrate supply induced by cyclonic circulation.
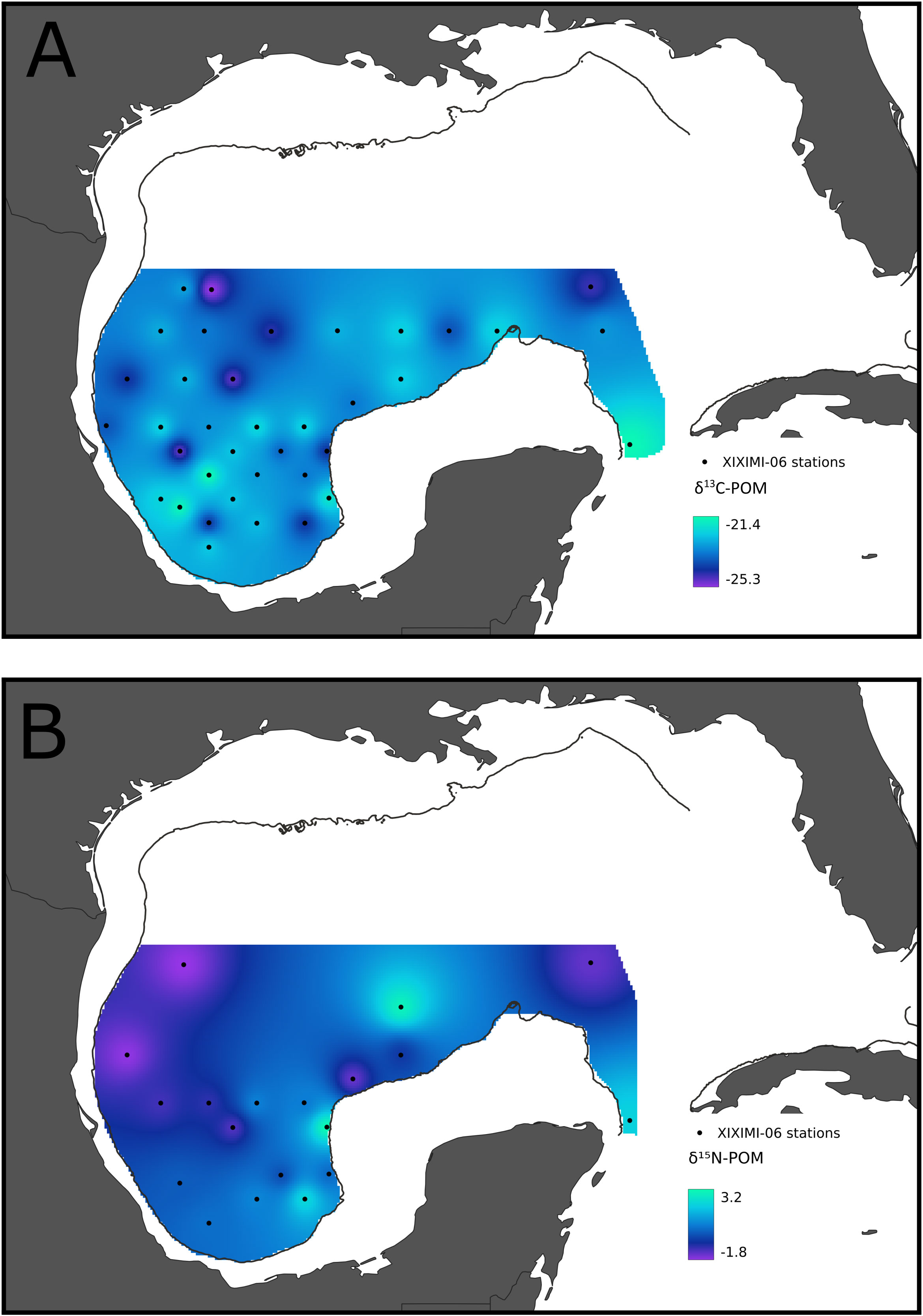
Figure 3 δ13C (A) and δ15N (B) isoscapes of suspended POM sampled in the euphotic layer of the Gulf of Mexico during the summer of 2017. Dots indicate sampling stations. Color gradient is consistent with Figure 2.
There was a significant and positive correlation (r = 0.62, p=0.0065) between δ15N values of suspended POM and those of zooplankton in the deep water region sampled during the XIXIMI-06 cruise (Figure 4A). There was also a positive correlation (r = 0.63, p=0.0065) between the isotopic composition of suspended POM and integrated NO3- concentrations (Figure 4B), which suggests fluxes of NO3- from the subsurface led to higher δ15N values of suspended POM. Also, there was a positive and highly significant correlation (r=0.53, p=0.00035) between δ15N values of zooplankton and surface chl a for samples from the deep water region collected during XiXIMI-06 (Figure 4C). The correlation between surface chla and zooplankton δ15N values that included the deep waters, slope and shelf samples was also positive and significant (Figure 2S). Lastly, there was a weaker correlation between zooplankton δ15N values and biovolumes (r=0.41, p = 0.0071; Figure 3S), and the relationships with %C, %N and C:N were not significant (Figures 4S–6S).
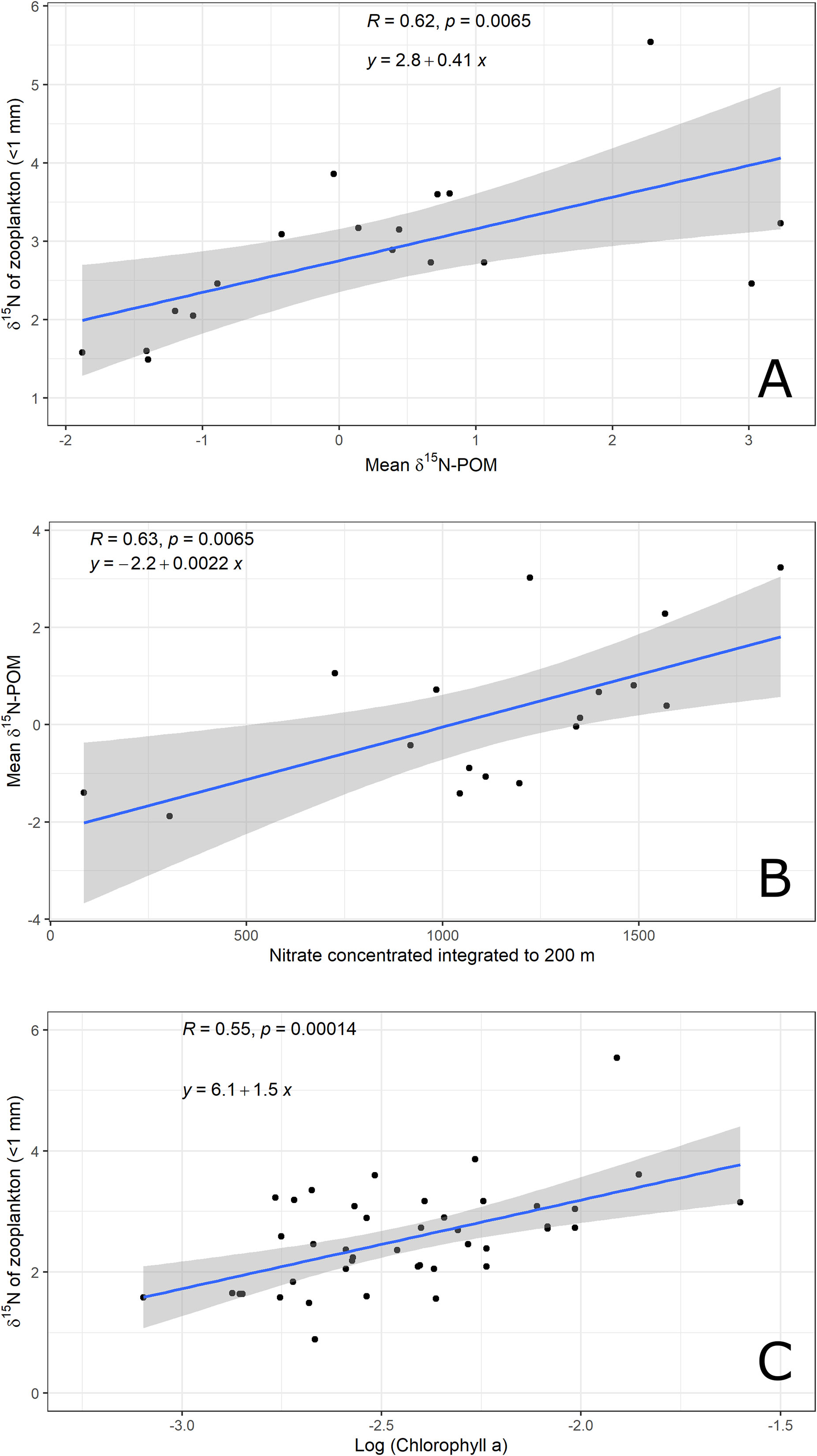
Figure 4 Least-squares correlation analysis between δ15N of suspended POM from pooled water samples and zooplankton, F= 9.797, df= 16, (A), nitrate concentration integrated between 0-200 m at XIXIMI-06 stations, F = 9.967, df=15 for (B) and Log (Chlorophyll a) and δ15N values of zooplankton, F=15.25 df=40. Gray areas indicate 95% confidence intervals. Least-squares correlation analysis between logarithm of chlorophyll a and δ15N values of zooplankton (C), F=15.25 df=40.
For simplicity, we present region-specific source contributions from north to south. In the NGMc the largest contribution was attributed to N from denitrification, with a mean value for all stations within the region of 60 ± 23% (Figures 5, 6A); in the most coastal stations (E12 and E20) denitrification contributes ~ 80% of the N (Figure 5D). Nitrogen from MARS contributed with a mean of 17.8 ± 11.7% (Figures 5C, 6A). In the NGMo, the highest contribution was from N2 fixation, with 45 ± 21.3% (Figure 5), although at some stations (E08, E09 and E10) fixation contributed up to ~ 60%. The contribution of nitrate supporting secondary production in the NGMo was estimated at 20 ± 8.4%, (Figure 5B); MARS, denitrification and WFS N contributed less than 15% (Figures 5D, E). In the CGM and LC, N2 fixation was the most important N source supporting zooplankton production, with a mean contribution of 73 ± 4.8% (Figures 5A, 6A), and contributions as high as 80% were estimated for two stations (B14 and C21; Figure 5A). Nitrate was the most important N source in the YS with an estimated mean contribution of 45.5 ± 11.4% (Figure 5).
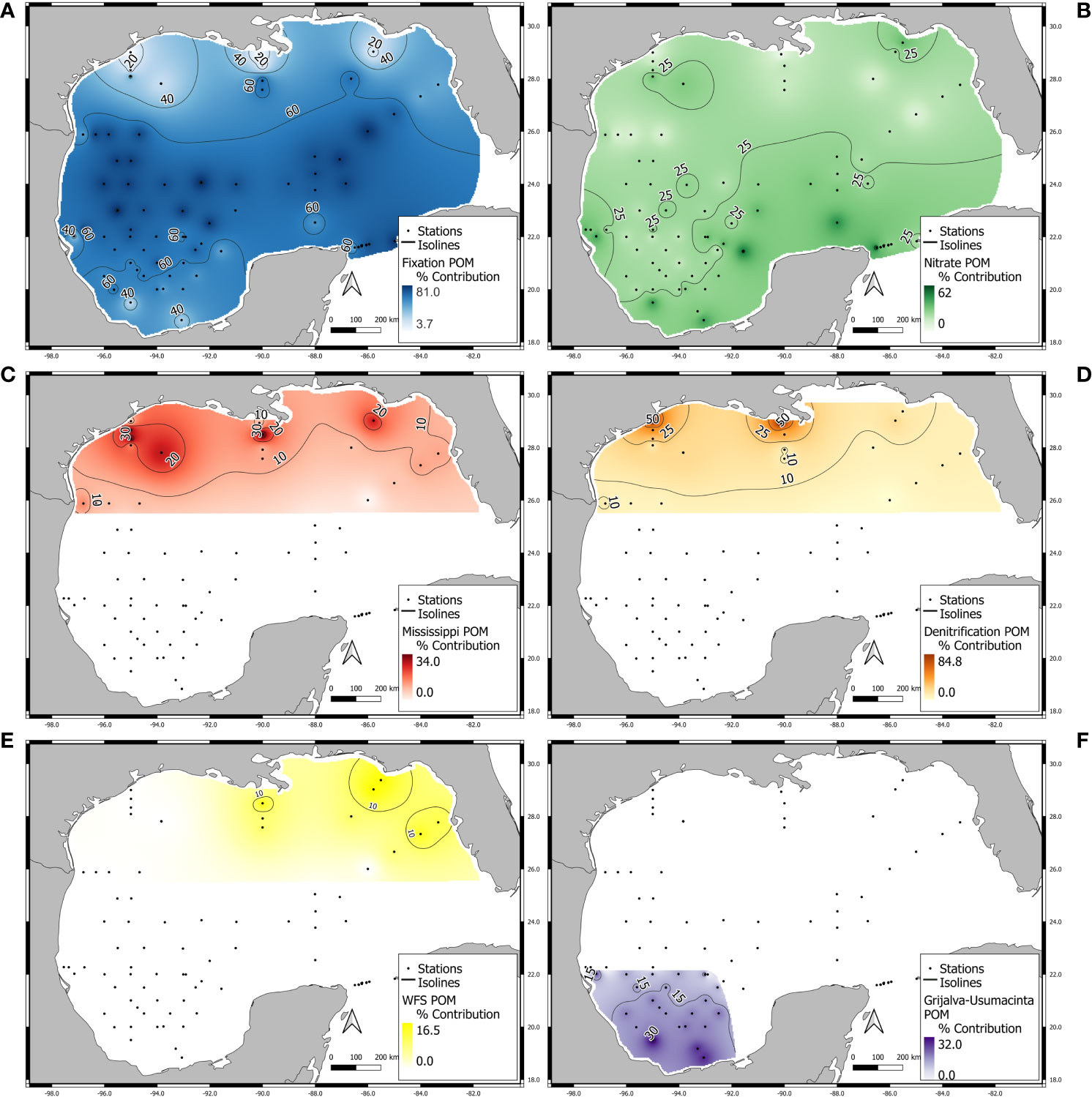
Figure 5 Percent contribution of (A) N2 fixation, (B) nitrate, (C) Mississippi-Atchafalaya River System ‘‘MARS’’, (D) denitrification, (E) Western Florida shelf ‘‘WFS’’ and (F) Grijalva-Usumacinta System ‘‘GUS’’ for different regions of in the Gulf of Mexico. Black dotted lines represent the isolines of fractional contributions and numbers show the percent contribution. White areas indicate a particular source was not considered important in the region and was therefore not included in isotope mixing models. Source contributions may not sum to 100% in interpolated areas. See Table 4S for source contribution estimates for each station.
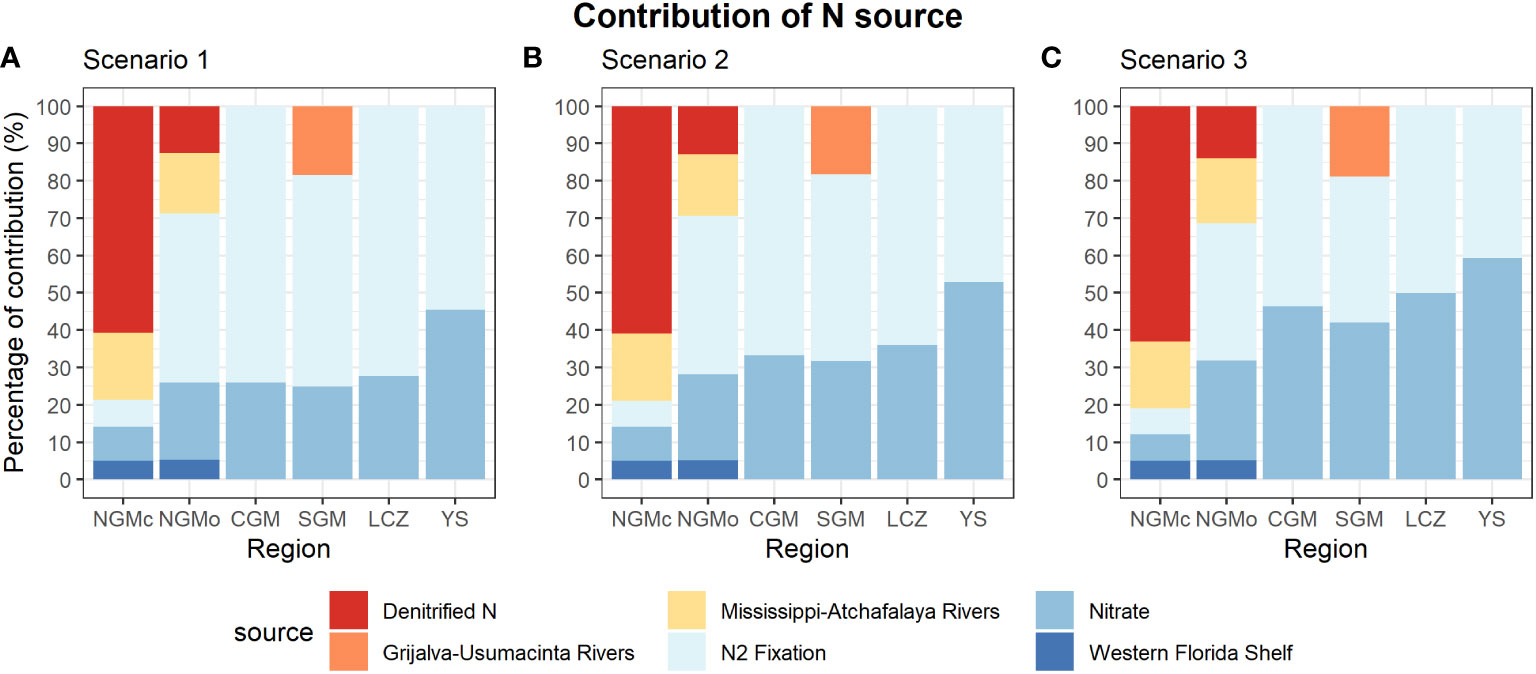
Figure 6 Percent contribution of N sources calculated by varying the isotopic composition of nitrate. (A) Scenario 1: δ15N-NO3 = 4.0 ± 0.3‰ of subsurface waters for the western Atlantic, (B) Scenario 2: δ15N-NO3 = 3.5± 1.1‰ from Howe et al. (2020), (C) Scenario 3: δ15N-NO3 = 1.9± 0.8‰ from Holl et al. (2007). NGMc, Coastal Northern Gulf of Mexico; NGMo, Oceanic Northern Gulf of Mexico; CGM, Central Gulf of Mexico; SGM, Southern Gulf of Mexico, LC, Loop Current region; YS, Yucatan Shelf.
For the SGM, N2 fixation was the most important source, with a mean of 56.6 ± 11.8% (Figures 5A, 6A), in contrast, the contribution of nitrate was about half of fixed N2, with a mean of 25 ± 7.8%. On the other hand, the N associated with the GUS discharge had a contribution of 18.5 ± 5.8% (Figure 5), although the contribution at some stations of the inner shelf was as high as ~ 30%.
A difference of 0.5‰ in the δ15N-NO3 values applied for source contribution estimates in scenario 2 (δ15N-NO =3.5 ± 1.1 ‰ corresponding to subsurface nitrate in the northeastern GM) and scenario 1 (δ15N-NO3 = 4.0 ± 0.3 ‰ corresponding to subsurface nitrate in the western Atlantic) was too small to substantially change source contributions. The mean gulf-wide contribution of N2 fixation decreased from 58.1% in scenario 2 to 52.1% in scenario 1, and the mean gulf-wide contribution of subsurface nitrate increased from 25.2% to 31.0%. For the other N sources, the changes were limited to <1% (Figure 6B).
The largest changes in source contributions (up to 22%) were found between scenario 1 and scenario 3 (δ15N-NO3 = 1.9 ± 0.8 ‰ corresponding to subsurface nitrate from the western Gulf) and were greater for the CGM, SGM, YS and LC. Under scenario 3 (Figure 6), the mean gulf-wide contribution of fixed N2 decreased from 58.1 to 42.1%, and the mean gulf-wide contribution of nitrate increased from 25.2 to 40.3%. The regions with the highest decrease in the contribution of N2 fixation were the LC (72.2 to 50.0%), CGM (74.0 to 53.5%), SGM (56.6 to 39.1%), and YS (54.5 to 40.6%). In contrast, the estimated contribution of subsurface nitrate increased for all of these regions but decreased slightly in the NGMc (from 9.1 to 7.0%; Figure 6).
We found a broad range of δ15N values for mesozooplankton sampled throughout the GM (0.9 to 11.6‰), which indicates that differences in regional DIN source contributions strongly control the isotopic baseline. In turn, the isotopic composition of DIN reflects the biological, biogeochemical and physical processes that characterize the basin.
The isotopic composition of mesozooplankton showed a strong latitudinal gradient, with the highest δ15N values (8.9 to 11.6‰) at stations of the coastal northern GM. This is the region of influence of MARS, and where the so-called “dead zone” is, where suboxic and anoxic conditions caused by high nutrient inputs and stratification are prevalent (Rabalais et al., 2001; Bianchi et al., 2010). The northern shelf is known for isotopically heavy N inputs, with δ15N-NO3 values of 7.3 ± 0.3‰ (Bryantmason et al., 2013), as well as regional denitrification that leads to enrichment in residual nitrate pools (Heaton, 1986; Ledford et al., 2020). In contrast, the lowest values were found in the central gulf and in the region of influence of the LC (0.9 to 3.6‰), which have well-described oligotrophic conditions (Biggs et al., 1992; Müller-Karger et al., 2015; Pasqueron De Fommervault et al., 2017). These lower nitrogen isotope ratios are consistent with inputs of N2 fixation (see below).
A basin-wide isoscape for the Atlantic Ocean based on a meta-analysis of δ15N values of zooplankton showed a marked regional pattern (McMahon et al., 2013), with low δ15N values (0 to 2‰) in the subtropical western region that has been attributed to N2 fixation by diazotrophic organisms (Montoya et al., 2002; Landrum et al., 2011), compared with higher values (6 to 8‰) for the temperate and Arctic regions, where NO3- is the major source of DIN for phytoplankton. The δ15N values for the central and LC regions in this study are consistent with those reported by McMahon et al. (2013) for the subtropical western Atlantic Ocean. However, very limited data for the GM were included in their isoscape and their interpolation was based on a few stations of the northwestern GM and the Yucatan Channel and Strait of Florida, with δ15N values of zooplankton 5-7‰ and 2-4‰, respectively. Hence, the isoscape did not capture the lower zooplankton and suspended POM isotope ratios reported here for the central and southern gulf. Our results, therefore, complement their basin-wide zooplankton-based isoscape for the Atlantic Ocean by providing higher spatial resolution values for one of its two marginal seas. Similar to what we found, a zooplankton-based isoscape for the subtropical southwestern Atlantic Ocean by Troina et al. (2020) reported a strong latitudinal pattern in δ15N values. They found a north-to-south gradient, with lower δ15N values (2.9 ± 1.0‰) in the more oligotrophic northern region that was attributed to the influence of N fixed by Trichodesmium, compared with higher δ15N values (4.0 ± 1.5‰) in the southern region where there are higher fluxes of subsurface NO3- and inputs from continental runoff. All of these studies highlight the importance of sampling zooplankton at an adequate spatial resolution to capture the isotopic fingerprint of regional biogeochemical processes and source contributions.
In contrast to what was observed for the isotopic composition of nitrogen, in this study, the carbon isotope ratios of zooplankton did not show a clear spatial pattern. The range of values for the GM (-22 to -17‰) is generally consistent with those of marine phytoplankton (-25 to -18‰, Fry and Sherr, 1984). However, at shelf stations close to the discharge of rivers, there was a gradient toward lower values inshore. Carbon isotope ratios in the coastal northern GM (-21.7‰) and the Grijalva-Usumacinta river plumes (-21.3‰) were ca. 1‰ lower than for the deep water region (-20.4‰). This likely reflects the input of C3 terrestrial organic matter (-27 to 30‰) that is typical of freshwater systems and river runoff, as well as low DIC δ13C values due to remineralization of terrestrial organic matter (Druffel et al., 2005; Cai et al., 2015; Sepúlveda-Lozada et al., 2015). Troina et al. (2020) also reported an inshore-offshore gradient, with low δ13C values (ca. 21.8‰) for the shelf-break region compared with offshore stations (ca. -20.5‰), which they attributed to inputs of terrestrial organic matter, riverine water and upwelling of subsurface DIC depleted in 13C. Hence, the δ13C values of zooplankton collected throughout the GM largely reflect phytoplankton production, although limited but detectable inputs of C3 terrestrial carbon or DIC in coastal areas close to river plumes were evident.
The Bayesian isotope mixing model indicated that, excluding the coastal northern Gulf of Mexico region, nitrogen fixation was the most important source supporting secondary production during the summer, with an estimated mean regional contribution between 45 and 74 (see also source contributions for each station in Table 4S). The lowest nitrogen isotope ratios were found in the CGM, LC and SGM, with mean values of 1.9 ± 0.5‰, 2.3 ± 0.6‰ and 3.1 ± 0.9‰, respectively. These values led to high estimates of the contribution of fixed N2 (mean 56-74%, Figures 5A, 6A). Low δ15N values have been associated with inputs of fixed N in the tropical and subtropical Atlantic (Montoya et al., 2002; Holl et al., 2007; Landrum et al., 2011; Kürten et al., 2016), as well as in temperate oceans (Loick-Wilde et al., 2019). For example, for the western tropical and subtropical Atlantic Ocean, a contribution of diazotrophs to suspended POM and zooplankton as high as 65% has been estimated for the mixed layer (Landrum et al., 2011), and high rates of nitrogen fixation (150 to 850 μmol N m-2 d-1) have been reported (Capone et al., 2005; Montoya et al., 2007; Table 4). Also, Holl et al. (2007) measured increasing N2 fixation rates (from 47 to 119 μmol N m-2 d-1) along a transect running from the continental shelf to the deep water region in the northwestern GM and used an isotope mixing model to estimate that 60% of the C and N supporting zooplankton production during the summer in the deep water region was from Trichodesmium. Hernández-Sánchez et al. (2022) also reported low δ15N values of copepods (3.7 ± 1.0‰) and mesozooplankton (<1000 μm; 2.8 ± 0.8‰) for the CGM and SGM sampled during 4 cruises held between 2010 and 2016, and estimated a 50-63% contribution of fixed N2 during the summer, consistent with estimates for the same regions based on this study’s more extensive sampling in 2017. Low δ15N values (ca. 1.8 to 2.8‰) of zooplankton have also been reported within the northern GM in anticyclonic eddies and the Loop Current and attributed to the presence of fixed N by Trichodesmium during the summer (Dorado et al., 2012; Wells et al., 2017). Landry and Swalethorp (2022) sampled the northeastern oceanic GM in the region of influence of the Loop Current. They found slightly lower isotope ratios for their small size fraction (0.2-0.5 mm ~3.4‰) compared with the larger fraction (0.5-1.0 mm, with ~4‰). A difference of 0.5‰ corresponds to about a fourth of a trophic level (assuming a TDF of 2‰ for crustacean zooplankton). Importantly, the isotope ratios they report for their small size fractions were higher (δ15N = ca. 3.0‰) than what we report for the GoM’s deep water region’s zooplankton in the 335-1000 µm size range. Given that larger size fractions of zooplankton typically reflect higher trophic levels, the fact that we found lower values is also indicative of the contribution of nitrogen fixation. The authors also calculated that the percentage of carnivores in the two fractions was only about ~15%, which implies that the skew in our isotope data would be minimized due to the presence of carnivores if our samples contain a similar proportion of higher-level consumers. Hence, as has been documented for the oligotrophic Atlantic, fixed nitrogen plays a significant role in supporting secondary production in oligotrophic regions of the GM.
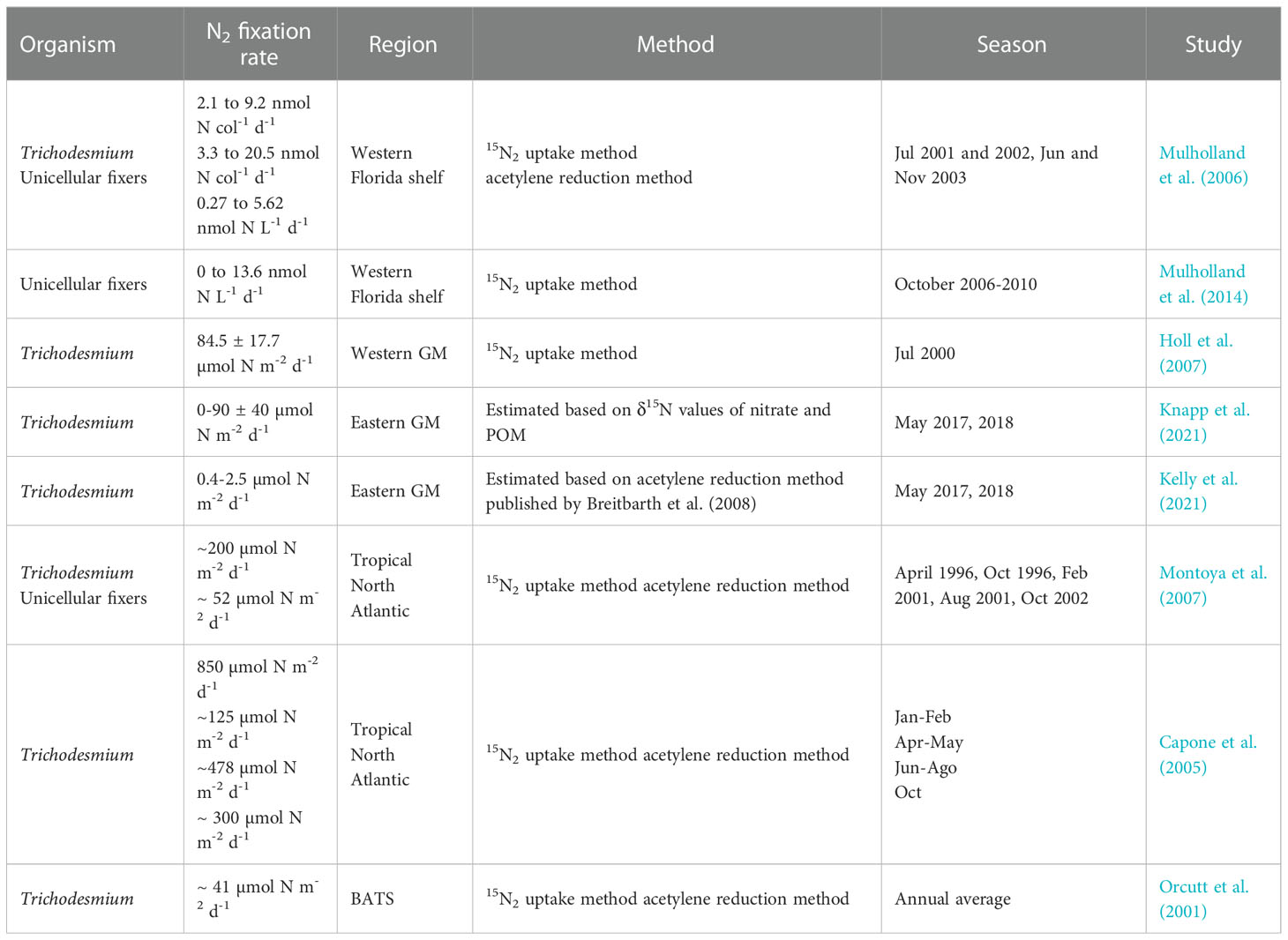
Table 4 Dinitrogen fixation rates in oligotrophic waters and Gulf of Mexico.
Subsurface NO3- was the second most important N source, with mean contributions between 25-27% in the deep water regions of the Gulf (CGM, LC and SGM). The moderate contribution of nitrate during the summer is likely due to the strong stratification and shallowing of the mixed layer (<30 m), which limits the vertical flux of subsurface NO3- to the euphotic zone (Müller-Karger et al., 2015; Damien et al., 2018). Anticyclonic eddies and the Loop Current show a deepening of the nitracline (Lee-Sánchez et al., 2022), which limits the transport of subsurface nitrate toward the euphotic layer. However, the transport of nitrate toward the euphotic layer occurs in cyclonic eddies, due to a shallowing of the nitracline, resulting in higher NO3- available for assimilation by primary producers (Biggs and Müller-Karger, 1994; Seki et al., 2001). In addition, the semipermanent cyclonic eddy found in the SGM (Pérez-Brunius et al., 2013) should enhance nitrate availability in the upper layer (see Figure 1S).
For the central and southern deep water region of the Gulf, we found positive correlations between the integrated nitrate concentrations and δ15N values of POM, and between δ15N values of POM and those of zooplankton (Figures 4A and 4B). Given that the isotopic composition of POM serves as a proxy for that of phytoplankton in oceanic systems, this indicates that nitrate assimilation by primary producers is propagated to zooplankton. This has been reported for oligotrophic regions in which nitrogen isotope ratios tend to be low due to N2 fixation (Montoya et al., 2002) compared with regions with higher nitrate concentrations and availability, which show higher δ15N values for POM and zooplankton (Lorrain et al., 2015; Kürten et al., 2016). In addition, the strong relationship between surface chl a and the δ15N values of zooplankton suggest that higher nitrate availability is reflected in higher primary producer biomass, at least near the surface (Figures 4C), as well as in zooplankton biomass (Figure 2S).
Over the shelves, various mechanisms can lead to inputs of nitrate. In the southern Bay of Campeche, convergent currents during the peak of the rainy season (July through November), cause cross-shelf transport of river waters that likely leads to increases in [NO3-] near the surface, which is reflected in surface chlorophyll plumes (Martínez-López and Zavala-Hidalgo, 2009). Coastal upwelling in the Tamaulipas-Veracruz shelf in the western GM during the summer and cross-shelf transport in the southwestern Louisiana-Texas (LATEX) shelf during May would increase the transport of subsurface nitrate to the euphotic zone in the western gulf close to the slope (Martínez-López and Zavala-Hidalgo, 2009; Mateos-Jasso et al., 2012; Zavala-Hidalgo et al., 2014). For the Yucatan Shelf, the isotope mixing model indicated that on average 50% of the N was from subsurface nitrate, although values as high as 60% were calculated for some stations. This higher contribution is likely due to the upwelling of water from depths of 200-250 m that occurs in the eastern Yucatan shelf and upper slope due to the interaction of the intense western boundary Yucatan Current with the Yucatan Channel, as well as the westward winds that can contribute to upwelling of water with high NO3- concentrations (8 to 14 μmol; Merino, 1997; Reyes-Mendoza et al., 2016; Jouanno et al., 2018). This regional upwelling produces positive anomalies in surface chl-a concentrations, which has been reported along the inner YS during the summer months due to the transport by the westward current over the shelf (Zavala-Hidalgo et al., 2006). Indeed, higher nitrogen isotope ratios and nitrate contributions can in fact be observed at some of the stations over the shelves and near the shelves of LATEX, the Bay of Campeche and the Yucatan Shelf (Figures 2, 5).
Although nitrate was estimated to have a moderate contribution (~25%) to zooplankton nitrogen in the deep water region of the gulf during the summer, its contribution is higher during the winter. Hernández-Sánchez et al. (2022) reported higher δ15N values (4.3 ± 0.6‰) of copepods sampled during the winter in the CGM and SGM compared to summer (3.7 ± 1.0‰), and estimated a 56 ± 2% contribution of subsurface NO3-, which they attributed to wind-driven mixing of cooler waters and the consequent deepening of the mixed layer. Indeed, a deepening of the mixed layer during the winter has been shown to increase the nitrate supply to the euphotic layer, leading to higher depth-integrated production (Müller-Karger et al., 2015; Damien et al., 2018). Hence, there is a seasonal pattern of the relative contribution of subsurface nitrate to secondary production in the Gulf’s deep waters that are not considered in our synoptic summer survey and that should be examined in greater detail.
The residual N resulting from denitrification was the most important N source in the coastal northern GM, contributing 60% of the nitrogen supporting zooplankton (Figure 6A). Hypoxic conditions have been reported downstream of the flow of MARS during the spring and summer (Rabalais et al., 2001; Rabalais et al., 2002; Bianchi et al., 2010), where high nutrient and organic matter loads lead to phytoplankton blooms and high water column and sediment respiration. Coupled with the strong stratification of the water column during the summer months, it exacerbates the hypoxia close to the bottom along the coast and shelves off Louisiana and Texas (Bianchi et al., 2010; McCarthy et al., 2015). Under these conditions, denitrification leads to remnant NO3- enriched in 15N, since it discriminates against 15N with fractionation values as high as ~30-35‰ (Heaton, 1986; Kritee et al., 2012). A high contribution of enriched N resulting from denitrification was estimated for the NGMc stations, particularly for the innermost stations (E12 and E20), which showed the highest δ15N values of zooplankton measured in this study (11.6 ± 0.1‰) and the highest source contributions (81-85%). Likewise, high δ15N values of zooplankton have been reported for the northern GM shelf (8.9 ± 0.9‰; Macko et al., 1984) and values between 8.2‰ and 9.9‰ have also been reported for phytoplankton and suspended POM in the Mississippi Sound (Chanton and Lewis, 1999; Moncreiff and Sullivan, 2001). Taken together, these measurements indicate that residual N from denitrification is the most important source of N for zooplankton in the coastal stations of the northern GM, particularly in the region of influence of MARS.
Our results indicate that the nitrogen inputs of the Mississippi and Atchafalaya river systems have a moderate contribution (mean 17%) in the northern shelf compared with denitrification, although their discharge is the major source of terrestrial organic matter and nutrient inputs to the GM. MARS contributions were > 15% only in the outer shelf stations of NGMc and inner stations of NGMo (26-34%; Figure 5C), and more limited in the deep water stations of the oceanic northern gulf (5-15%; see Figure 5C).
Relatively high δ15N-NO3 values (~7.0‰) and δ15N suspended POM values (6.5-7.2‰) from MARS have been associated with different N sources, including manure, treated wastewater in the upper Mississippi River and its tributaries and denitrification (Chang et al., 2002; Bryantmason et al., 2013). If NO3- with these higher δ15N values was the main N source supporting the food web in the NGM, the δ15N values of zooplankton would be ~8-9‰ (considering a TDF ~2‰). However, at two of the inner stations of the NGMc (E12 and E20), the δ15N values were 11.6‰, which is almost 3‰ higher than what would be expected if MARS were the sole N source. The low contribution of MARS N to the deep water stations may also reflect the timing of our sampling; discharge is mostly transported toward the LATEX shelf during the autumn, winter and early spring, while eastward transport can occur during the late spring and summer months (Schiller et al., 2011). Offshore transport of low salinity water from MARS due to interactions with mesoscale eddies can occur (Schiller et al., 2011), but this occurs as filaments and it is not a region-wide phenomenon. In addition, the highest discharge of MARS occurs during the spring and early summer, and decreases in the late summer and autumn (Walker et al., 2005), and our zooplankton samples were collected during late summer.
The inputs of the Grijalva-Usumacinta River system only had moderate contributions at the stations closest to the river mouths, accounting for ~30% of zooplankton N. In the coastal southern GM, these rivers are the most important in terms of freshwater inflow and nutrient inputs, and mean δ15N values of suspended POM 4.6 ± 0.5‰ have been reported for their lower reaches (Sepúlveda-Lozada et al., 2015). The maximum discharge from Grijalva-Usumacinta rivers is during the summer (August-October; Muñoz-Salinas and Castillo, 2015), and chl a produced by this discharge may be transported toward the SGM by cross-shelf transport (Martínez-López and Zavala-Hidalgo, 2009). Zavala-García et al. (2016) evaluated the relationship between zooplankton biomass on the shelf and deep water region of the southern Bay of Campeche and discharge from rivers in the states of Veracruz, Tabasco and Campeche during the annual cycle. They found a positive relationship between zooplankton biomass and the river discharge and attributed the high zooplankton biomass to increased productivity due to high river discharge in summer. This is consistent with the contributions estimated with the Bayesian isotope mixing model, which indicated that suspended POM from the Grijalva-Usumacinta River has moderate contributions only at the stations closest to the coast of Campeche.
The gradients in zooplankton stable isotope ratios over the WFS and the results of the regional mixing model indicate that multiple nitrogen sources and processes are involved in supporting secondary production. The N source from WFS rivers had a relatively low N contribution to the northeastern gulf (with an average of ~5% and up to 16% contribution at the northern shelf stations). Zooplankton N isotope ratios showed a latitudinal pattern within the WFS, with higher values at northern stations associated with a higher contribution of heavy N likely resulting from denitrification and intermediate values in the central WFS that were associated with increased contributions of N2 fixation, respectively. Radabaugh et al. (2013) evaluated the isotopic composition of suspended POM, primary producers and fish along a gradient from eutrophic to oligotrophic waters on the WFS, and also reported a latitudinal pattern in δ15N suspended POM values that was reflected at higher trophic levels, with higher values at the northernmost stations (~6-7‰) compared with the southern shelf (~3-5‰). Del Castillo et al. (2001) evaluated river runoff, dissolved and particulate organic matter and chl a concentrations using multispectral fluorescence and satellite sensing in the eastern GM, and found that high chl a was associated with relatively low salinities in the inner WFS. Lower surface chl a concentrations in the central shelf was attributed to the limited influence of river runoff. This is consistent with our results, which suggest the river contribution of N to the WFS was relatively low (8-15%). Del Castillo et al. (2001) also reported high surface chl a concentrations that were transported eastward from MARS toward the outer WFS by LC anticyclonic eddies when the LC presented a high intrusion into the GM, and during the cruise, an anticyclonic eddy was observed close to the shelf of the Northern Eastern GM (Figure 1S) that could transport the runoff with suspended POM with higher isotopic values from MARS and which in our data are reflected in an estimated 15-25% contribution of MARS N.
On the other hand, we estimate a relatively high contribution of fixed N in the central region of the WFS (45-68%) resulting from relatively low δ15N values of zooplankton (2.4 to 4.8‰). N2 fixation rates (1.32 to 8.2 μmol N m-2 d-1) associated with Trichodesmium have been reported for the central region of the western Florida shelf and may be linked to blooms of the toxic dinoflagellate Karenia brevis (Mulholland et al., 2006).
The estimates of the subsurface nitrate contribution increased with decreasing δ15N-NO3 values. This pattern was more marked in the central and western GM where nitrogen fixation estimates were highest. When using the lowest δ15N-NO3 value reported by Holl et al. (2007) under scenario 3 (1.9 ± 0.8‰), the contribution of N2 fixation decreased in all regions relative to scenario 1 (δ15N-NO3 = 4.0 ± 0.3‰), and its fractional contribution was similar to that of nitrate (ca. 42%; Figure 6C). In the YS region, the major N source changed from N2 fixation to subsurface NO3-. However, using a δ15N-NO3 value of 1.9 ± 0.8‰ is not realistic for the Yucatan Strait and the eastern gulf, since the measurement corresponds to the western gulf and for waters that have been within the gulf for months or years. Although the fractional contribution of nitrate did increase under scenarios 2 (δ15N-NO3 = 3.5 ± 1.1‰) and 3, the contribution of N2 fixation remained substantial for all regions (mean = 37 to 53%), except for NGMc (7%) under scenario 3.
The subsurface nitrate isotope ratios measured to date within the GM decrease westward, indicating the contribution of a relatively light source of DIN. Three possible explanations are not mutually exclusive: (1) the remineralization of organic matter with low nitrogen isotope ratios that reflects the uptake of fixed N by primary producers, (2) the remineralization of nitrate-based organic matter that contributes dissolved inorganic nitrogen depleted in 15N, and (3) the contribution of light N excreted by heterotrophs.
First, remineralization of diazotroph-derived organic matter would contribute light DIN to the subsurface nutrient pool, lowering δ15N-NO3 values. Remineralization has an estimated N isotope discrimination of ~3‰ (Sigman and Fripiat, 2019). Based on the mean δ15N values of suspended POM measured in this study for the gulf’s deep waters (0.3 ± 1.4‰), the remineralization of this organic matter would result in dissolved inorganic nitrogen with an isotopic composition of about -3‰. The contribution of remineralized DIN from diazotroph-derived organic matter hinges on the magnitude of nitrogen fixation within the gulf.
During our study, we did not measure nitrogen fixation rates or the abundance of diazotrophs. However, Trichodesmium has been reported for the western and eastern GM at abundances of 10 to 1 x 104 trichomes L-1 (Holl et al., 2007) and 300 to ~106 trichomes L-1 (Lenes and Heil, 2010) during the summer months. Assuming that a colony of Trichodesmium has 200 trichomes (Carpenter, 1983; Lenes and Heil, 2010), a conservative amount of 2 colonies L-1 for the western GM during the summer and using an average fixation rate of 8.8 nmol N col-1 d-1 reported by Mulholland et al. (2014), we estimate 176 µmol N m-2 d-1 of fixed N for the first 10 m depth in the GM, which is the range of N2 fixation rates for the western Atlantic Ocean (Capone et al., 2005, see Table 4), where relatively low δ15N values of zooplankton have also been documented (Montoya et al., 2002; Lamdrum et al., 2011). Hence, fixation appears to be an important N source supporting secondary production in the deep water region of the GM. We nevertheless recognize that data on Trichodesmium abundance and distribution throughout the GoM are scarce and that additional sampling to quantify the abundance of colonies and in situ measurements of fixation rates would be of great value. Second, the remineralization of organic matter supported by subsurface NO3- would contribute to isotopically light DIN. Given that the euphotic layer behaves as a closed system in stratified oligotrophic waters, the complete assimilation of subsurface nitrate as a single N source with δ15N values of 4.0 ‰ (under scenario 1) or 3.5 ‰ (under scenario 2) would lead to phytoplankton with similar isotope ratios, and herbivorous zooplankton with values of 5.5-6.0‰, which is substantially higher than measured in this study and by Hernández-Sánchez et al. (2022). The values reported here for the central GM and LC regions are at least 3‰ lower, which implies that subsurface nitrate could not have been the sole N source supporting secondary production. Importantly, the highest δ15N values of zooplankton caught in the deep water region of the GM were at stations in which integrated nitrate concentration was highest (Figure 4). This indicates that zooplankton isotope ratios do reflect that of nitrate when it is available.
In addition, the suspended POM measured in the central and southern deep waters of the GM had a range of -1.9 to 3.2‰ (0.2 ± 1.4‰ mean± SD); most values were lower than 1‰ (Figure 4). These low δ15N suspended POM values cannot be explained solely by the remineralization of organic matter supported by subsurface NO3-. DIN produced in the euphotic zone by the remineralization of phytoplankton supported solely by subsurface nitrate would have an isotope ratio of about 0.5-1‰. An additional source of low δ15N, such as N2 fixation is required (-2 to 0‰; Carpenter et al., 1997). Given the high level of stratification that characterizes the gulf’s deep waters during the summer, and the low δ15N values of zooplankton and suspended POM, the contributions of subsurface nitrate are likely limited.
Third, heterotroph excretion could contribute to the low values of suspended POM. Checkley Jr. and Miller (1989) reported a discrimination factor of 3‰ between zooplankton bodies and excreted ammonium and found this to be consistent across the eutrophic sub-boreal to oligotrophic subtropical regions. The lowest mean δ15N values of zooplankton we sampled were for the CGM (1.9‰). Assuming a discrimination factor of 3‰, the δ15N value of excreted ammonium would be -1.1‰. Applying the Bayesian mixing model including excreted ammonium as an additional source (-1.1‰) under scenario 1 for the deepwater regions of the GM, the excreted ammonium could contribute ~45% of the N, while N2 fixation would decrease to 23-27% (a 60% decrease), and the contribution of NO3- would remain similar (Table 4S).
While we do not discount subsurface nitrate inputs to the euphotic layer of the deep water region of the GM, zooplankton δ15N values in our study suggest that nitrate cannot be the sole N source supporting primary and secondary production, contrary to what has been suggested by Knapp et al. (2021) and Kelly et al. (2021) for the northeastern gulf in the region of influence of the Loop Current. Knapp et al. (2021) used sediment traps drifting below the euphotic layer in the north-central GM for 2-4 days during late spring to sample sinking particles, and measured their isotopic composition as well as that of subsurface nitrate. They used a box model approach and an δ15N budget to evaluate the relative importance of N2 fixation and subsurface NO3- to export production. They specifically targeted the region of influence of the LC, an area well-known for providing suitable habitat for several species of tuna (Thunnus spp.) larvae. Highest larval densities are often found at the edges of the LC or in frontal zones where eddies converge, leading to shear and upwelling (Lindo-Atichati et al., 2012; Rooker et al., 2012; Cornic et al., 2018). High zooplankton biomass has also been reported for the region (Wormuth et al., 2000; Biggs and Ressler, 2001).
Knapp et al. (2021) reported δ15N values in suspended POM of 1.0 to 2.5‰ for the upper 100 m (excluding their station C5, which was influenced by coastal inputs) which were higher compared to our average of 0.2‰; these values were similar or higher than those of δ15N-NO3 measured in their water samples collected below the euphotic layer in their offshore traps. They attributed their δ15N suspended POM values to nitrate-based production (subsurface NO3- had values between 2.0 to 3.8‰), remineralization of DON and zooplankton excretion. For one trap, however, they reported δ15N-POM-sink values that were lower than δ15N-NO3 values and estimated fixation rates of 90 ± 40 μmol m-2 d-1, which represented a contribution of only 10-18% of the exported N. Hence, they concluded that subsurface NO3- was the dominant N source supporting exported production in the GM. Although they estimated the contribution of N2 fixation indirectly based on δ15N-NO3 and δ15N values of POM-sink and considered it negligible in most cases, they did not measure in situ fixation rates.
A complementary study by Kelly et al. (2021) used data from the same sediment traps to develop a C and N budget using a biogeochemical model, remote-sensing observations and in situ measurements of Trichodesmium abundance to estimate fixation rates, lateral N transport and the contribution of upwelled NO3- to POM exported out of the euphotic layer. They attributed the low N2 fixation rates (<0.4-2.8 μmol N m-2 d-1) to a low abundance of Trichodesmium (range 0-19 trichomes L-1; Selph et al., 2022). In addition, they estimated that between 90-100% of the exported particulate N was supported by lateral transport of organic matter based on nitrate, and concluded that N2 fixation and subsurface NO3- fluxes do not play an important role. While lateral transport of subsurface nitrate is likely important in the context of Loop Current eddies that transport large volumes of water into the Gulf, our data indicate that it is not the only source of nitrogen throughout the extensive deep water region of the GoM. Strong stratification during the summer, when we sampled, also limits the availability of nitrate in the euphotic layer. In addition, their fixation rates were lower than estimates for the northwestern GM (49 to 119 μmol N m-2 d-1; Holl et al., 2007) and close to estimates for the Loop Current (<20 μmol N m-2 d-1; Knapp et al., 2021).
The large differences in the contribution of fixed nitrogen between the studies of Knapp et al. (2021) and Kelly et al. (2021) and this study are likely attributed to differences in methodological approaches, sampling season and spatial coverage. Their studies estimated the flux and isotopic composition of POM that sank from the euphotic zone, while we inferred nitrogen sources based on zooplankton isotope ratios that integrated that of phytoplankton and microzooplankton prey. The organic matter that sinks tends to have higher δ15N values than suspended POM because remineralization favors the discrimination against 15N (Sigman and Fripiat, 2019). Additionally, Fawcett et al. (2014) reported that in the Sargasso Sea region, small eukaryotic phytoplankton (<35 μm) show higher δ15N values than the prokaryotic phytoplankton (Prochlorococcus and Synechococcus), as they strongly rely on nitrate as a source of N while prokaryotes likely rely on recycled N (Fawcett et al., 2011). These authors also suggested that eukaryotic phytoplankton significantly contributes to new production and carbon export in this oligotrophic region, thus Knapp et al. (2021) sediment trap data may be reflecting a higher contribution of eukaryotes to sinking particles. This evidence suggests the N2 fixation contribution is potentially underestimated based on measurements of sinking POM alone.
In addition, Knapp et al. (2021) and Kelly et al. (2021) sampled during late spring (April and May) and reported very low Trichodesmium abundance (< trichomes L-1). Much higher Trichodesmium abundances (mean ± SD of 360 ± 157 trichomes L-1) were reported by Holl et al. (2007) for the summer, and they estimated an average fixation rate of 85 ± 17 μmol N m-2 d-1 with up to 119 μmol N m-2 d-1 at the most oceanic station. According to the authors, these estimates accounted for 60% of secondary production. Direct fixation rate measurements for the GM reported by Holl et al. (2007) are consistent with those reporteds by Jickells et al. (2017) based on a global biogeochemical model, ranging 82-137 μmol N m-2 d-1 through most of the central and southern gulf and up to 274 μmol N m-2 d-1 toward the north. Furthermore, much higher abundances of Trichodesmium have been reported for the WFS for both the summer (~106 trichomes L-1) and winter (~103 trichomes L-1) (Lenes and Heil, 2010). For the WFS, Mulholland et al. (2014) reported very high N2 fixation rates for Trichodesmium compared to those reported by Knapp et al. (2021) based on the sinking particulate N export in their 60 m traps (0.59-1.53 mmol N m-2 d-1). By considering an average fixation value per colony of 8.8 nmol N d-1, an average abundance of 20 colonies L-1 during summer (June to September) and a 20 m integration depth (Mulholland et al., 2014), an integrated N2 fixation value of 3.5 mmol m-2 d-1 is obtained. Mulholland et al. (2014) also reported an average fixation for other planktonic diazotrophs of 6 nmol L-1 d-1, which integrated to 20 m results in 120 μmol N m-2 d-1. While direct N2 fixation measurements are clearly needed for the central and southern GM, fixation rates previously reported for the summer months strongly suggest that this process is an important source of new N, likely representing a significant source of N for primary and secondary producers.
Based on the Bayesian isotope mixing model, regional sources strongly control the isotopic baseline and therefore mesozooplankton isotope nitrogen values during the summer in the GM. The Bayesian mixing model showed that N2 fixation supported up to 68% of the mesozooplankton production in the upper euphotic layer (~50 m) in the deep water region of GM where the nitrate is limited due to the high stratification during the summer. Also, the isotopic composition of zooplankton suggests that there is some feeding in the lower euphotic layer, where nitrate is more available, which is consistent with the vertical migration of some zooplankton taxa. Nitrogen sources from river runoff were limited to shelf stations. This was particularly notable in the case of the Mississippi-Atchafalaya Rivers, where residual N resulting from denitrification also provides an important source of nitrogen along the shelf. If the runoff from Mississippi-Atchafalaya Rivers and the residual N form denitrification are taken together as representative of the contribution of MARS, the contribution rises up to 50% to the secondary production for the NGMc and NGMo. This study provides the first gulf-wide assessment of nitrogen source contributions to secondary production that includes the open waters of the GM as well as coastal regions, and which may serve as baseline against which to assess the impact of anthropogenic disturbances such as changes in nutrient inputs and increased stratification due to global warming.
Publicly available datasets were analyzed in this study. This data can be found here: Bulk zooplankton data are presented in: Le-Alvarado M, Romo-Curiel AE, Sosa- Nishizaki O, OH-S, BL, SH (2021) Yellowfin tuna (Thunnus albacares) foraging habitat and trophic position in the Gulf of Mexico based on intrinsic isotope tracers. PLoS ONE 16(2): e0246082. https://doi.org/10.1371/journal.pone.0246082. Other data are presented within the article.
SH and JH obtained funding, SH, VC-I and OH-S designed the study, OS-H processed samples and analyzed data, OS-H, SH and VC-I wrote the manuscript, LB was chief scientist for the GOMECC-3 cruise, JH generated the isotope POM data. All authors contributed to the article and approved the submitted version.
This research has been funded by the Mexican National Council for Science and Technology (CONACYT)- Mexican Ministry of Energy - Hydrocarbon Fund, project 201441. This is a contribution of the Gulf of Mexico Research Consortium (CIGoM). We acknowledge PEMEX’s specific request to the Hydrocarbon Fund to address the environmental effects of oil spills in the Gulf of Mexico. OH-S was a doctoral student in the PhD program in Marine Ecology at CICESE and received a fellowship from CONACYT.
Data reported in this work were also partially collected under the auspices of the National Oceanic and Atmospheric Administration (NOAA) ‘s Ocean Acidification Program (OAP) GOMECC-3 cruise. We thank the scientific participants and crew of R/V Justo Sierra and the R/V Ronald Brown for their assistance in sample collection. Jesús C Compaire collected zooplankton samples during GOMECC-3. María Reyna Barradas helped with sorting of zooplankton and sample preparation for isotope ratio analysis.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.1025387/full#supplementary-material
Alexander R. B., Smith R. A., Schwarz G. E., Boyer E. W., Nolan J. V., Brakebill J. W. (2008). Differences in phosphorus and nitrogen delivery to the gulf of Mexico from the Mississippi river basin. Environ. Sci. Technol. 42, 822–830. doi: 10.1021/es0716103
Bianchi T. S., DiMarco S. F., Cowan J. H., Hetland R. D., Chapman P., Day J. W., et al. (2010). The science of hypoxia in the northern gulf of Mexico: A review. Sci. Total Environ. 408, 1471–1484. doi: 10.1016/j.scitotenv.2009.11.047
Bianchi T. S., Wysocki L. A., Stewart M., Filley T. R., McKee B. A. (2007). Temporal variability in terrestrially-derived sources of particulate organic carbon in the lower Mississippi river and its upper tributaries. Geochimica Cosmochimica Acta 71, 4425–4437. doi: 10.1016/j.gca.2007.07.011
Biggs D. C. (1992). Nutrients, plankton, and productivity in a warm-core ring in the western gulf of Mexico. J. Geophys. Res. 97, 2143–2154. doi: 10.1029/90JC02020
Biggs D. C., Müller-Karger F. E. (1994). Ship and satellite observations of chlorophyll stocks in interacting cyclone-anticyclone eddy pairs in the western gulf of Mexico. J. Geophys. Res. 99, 7371–7384. doi: 10.1029/93JC02153
Biggs D. C., Ressler P. H. (2001). Distribution and abundance of phytoplankton, zooplankton, ichthyoplankton, and micronekton in the deepwater gulf of Mexico. Gulf Mex. Sci. 19, 2. doi: 10.18785/goms.1901.02
Brault E. K., Koch P. L., McMahon K. W., Broach K. H., Rosenfield A. P., Sauthoff W., et al. (2018). Carbon and nitrogen zooplankton isoscapes in West Antarctica reflect oceanographic transitions. Mar. Ecol. Prog. Ser. 593, 29–45. doi: 10.3354/meps12524
Breitbarth E., Wohlers J., Kläs J., LaRoche J., Peeken I. (2008). Nitrogen fixation and growth rates of trichodesmium IMS-101 as a function of light intensity. Mar. Ecol. Prog. Ser. 359, 25–36. doi: 10.3354/meps07241
Bryantmason A., Xu Y. J., Altabet M. (2013). Isotopic signature of nitrate in river waters of the lower Mississippi and its distributary, the atchafalaya. Hydrol. 27, 2840–2850. doi: 10.1002/hyp.9420
Cai Y., Guo L., Wang X., Aiken G. (2015). Abundance, stable isotopic composition, and export fluxes of DOC, POC, and DIC from the lower Mississippi river during 2006-2008. J. Geophys. Res. Biogeosci. 120, 2273–2288. doi: 10.1002/2015JG003139
Cai Y., Guo L., Wang X., Mojzis A. K., Redalje D. G. (2012). The source and distribution of dissolved and particulate organic matter in the bay of st. Louis, northern gulf of Mexico. Estuar. Coast. Shelf Sci. 96, 96–104. doi: 10.1016/j.ecss.2011.10.017
Callejas-Jimenez M., Santamaria-del-Angel E., Gonzalez-Silvera A., Millan-Nuñez R., Cajal-Medrano R. (2012). Dynamic regionalization of the gulf of Mexico based on normalized radiances (nLw) derived from MODIS-aqua. Cont. Shelf Res. 37, 8–14. doi: 10.1016/j.csr.2012.01.014
Camacho-Ibar V. F., Muñóz-Anderson M., Ávila López M. C., Hakspiel-Segura C. (2021). Water column nitrate+nitrite and phosphate from samples collected during the XIXIMI-5 oceanographic cruise in the gulf of Mexico in June 2016. PANGAEA. doi: 10.1594/PANGAEA.934036
Capone D. G., Burns J. A., Montoya J. P., Subramaniam A., Mahaffey C., Gunderson T., et al. (2005). Nitrogen fixation by trichodesmium spp.: An important source of new nitrogen to the tropical and subtropical north Atlantic ocean. Glob. Biogeochem. Cycles 19, 1–17. doi: 10.1029/2004GB002331
Carpenter E. J. (1983). Nitrogen fixation by marine oscillatoria. Nitrogen Mar. Environ. 65–104. doi: 10.3354/meps185273
Carpenter E. J., Harvey H. R., Brian F., Capone D. G. (1997). Biogeochemical tracers of the marine cyanobacterium Trichodesmium. Deep-Sea Res. I: Oceanogr. Res. Pap. 44, 27–38. doi: 10.1016/S0967-0637(96)00091-X
Carpenter E., Montoya J., Burns J., Mulholland M., Subramaniam A., Capone D. (1999). Extensive bloom of a N2-fixing diatom/cyanobacterial association in the tropical Atlantic ocean. Mar. Ecol. Prog. Ser. 185, 273–283. doi: 10.3354/meps185273
Cervantes-Díaz G. Y., Hernández-Ayón J. M., Zirino A., Herzka S. Z., Camacho-Ibar V., Norzagaray O., et al. (2022). Understanding upper water mass dynamics in the gulf of Mexico by linking physical and biogeochemical features. J. Mar. Syst. 225, 103647. doi: 10.1016/j.jmarsys.2021.103647
Chang C. C. Y., Kendall C., Silva S. R., Battaglin W. A., Campbell D. H. (2002). Nitrate stable isotopes: Tools for determining nitrate sources among different land uses in the Mississippi river basin. Can. J. Fish. Aquat. Sci. 59, 1874–1885. doi: 10.1139/f02-153
Chanton J. P., Lewis F. G. (1999). Plankton and dissolved inorganic carbon isotopic composition in a river-dominated estuary: Apalachicola bay, Florida. Estuaries 22, 575–583. doi: 10.2307/1353045
Checkley D. M. Jr., Miller C. A. (1989). Nitrogen isotope fractionation by oceanic zooplankton. Deep-Sea Res. I: Oceanogr. Res. Pap. 36, 1449–1456. doi: 10.1016/0198-0149(89)90050-2
Chen M., Kim D., Liu H., Kang C. K. (2018). Variability in copepod trophic levels and feeding selectivity based on stable isotope analysis in gwangyang bay of the southern coast of the Korean peninsula. Biogeosciences 15, 2055–2073. doi: 10.5194/bg-15-2055-2018
Cornic M., Smith B. L., Kitchens L. L., Alvarado Bremer J. R., Rooker J. R. (2018). Abundance and habitat associations of tuna larvae in the surface water of the gulf of Mexico. Hydrobiologia 806, 29–46. doi: 10.1007/s10750-017-3330-0
Damien P., Pasqueron de Fommervault O., Sheinbaum J., Jouanno J., Camacho-Ibar V. F., Duteil O. (2018). Partitioning of the open waters of the gulf of Mexico based on the seasonal and interannual variability of chlorophyll concentration. J. Geophys. Res. Oceans 123, 2592–2614. doi: 10.1002/2017JC013456
Davenport S. R., Bax N. J. (2002). A trophic study of a marine ecosystem off southeastern Australia using stable isotopes of carbon and nitrogen. Can. J. Fish. Aquat. Sci. 59, 514–530. doi: 10.1139/f02-031
Del Castillo C. E., Coble P. G., Conmy R. N., Müller-Karger F. E., Vanderbloemen L., Vargo G. A. (2001). Multispectral in situ measurements of organic matter and chlorophyll fluorescence in seawater: Documenting the intrusion of the Mississippi river plume in the West Florida shelf. Limnol. Oceanogr. 46, 1836–1843. doi: 10.4319/lo.2001.46.7.1836
DiMarco S. F., Howard M. K., Reid R. O. (2000). Seasonal variation of wind-driven diurnal current cycling on the Texas-Louisiana continental shelf. Geophys. Res. Lett. 27, 1017–1020. doi: 10.1029/1999GL010491
Dorado S., Rooker J. R., Wissel B., Quigg A. (2012). Isotope baseline shifts in pelagic food webs of the gulf of Mexico. Mar. Ecol. Prog. Ser. 464, 37–49. doi: 10.3354/meps09854
Druffel E. R. M., Bauer J. E., Griffin S. (2005). Input of particulate organic and dissolved inorganic carbon from the Amazon to the Atlantic ocean. Geochem. Geophys. Geosyst. 6, Q03009. doi: 10.1029/2004GC000842
Durán-Campos E., Salas-de-León D. A., Monreal-Gómez M. A., Coria-Monter E. (2017). Patterns of chlorophyll-a distribution linked to mesoscale structures in two contrasting areas campeche canyon and bank, southern gulf of Mexico. J. Sea Res. 123, 30–38. doi: 10.1016/j.seares.2017.03.013
El-Sabaawi R., Trudel M., Mazumder A. (2013). Zooplankton stable isotopes as integrators of bottom-up variability in coastal margins: A case study from the strait of georgia and adjacent coastal regions. Prog. Oceanogr. 115, 76–89. doi: 10.1016/j.pocean.2013.05.010
Fawcett S. E., Lomas M. W., Casey J. R., Ward B. B., Sigman D. M. (2011). Assimilation of upwelled nitrate by small eukaryotes in the Sargasso Sea. Nat. Geosci. 4, 717–722. doi: 10.1038/ngeo1265
Fawcett S. E., Lomas M. W., Ward B. B., Sigman D. M. (2014). The counterintuitive effect of summer-to-fall mixed layer deepening on eukaryotic new production in the Sargasso Sea, global biogeochem. Cycles 28, 86–102. doi: 10.1002/2013GB004579
Fry B., Sherr E. B. (1989). “δ13C measurements as indicators of carbon flow in marine and freshwater ecosystems,” in Stable isotopes in ecological research. Eds. Rundel P. W., Ehleringer J. R., Nagy K. A. (New York, NY: Springer) 196–229. doi: 10.1007/978-1-4612-3498-2_12
Gorokhova E., Hansson S. (1999). An experimental study on variations in stable carbon and nitrogen isotope fractionation during growth of Mysis mixta and Neomysis integer. Can. J. Fish. Aquat. Sci. 56, 2203–2210. doi: 10.1139/f99-149
Gu B., Schell D. M., Frazer T., Hoyer M., Chapman F. A. (2001). Stable carbon isotope evidence for reduced feeding of gulf of Mexico sturgeon during their prolonged river residence period. Est. Coast. Shelf Sci. 53, 275–280. doi: 10.1006/ecss.2001.0816
Hamilton P., Leben R., Bower A., Furey H., Pérez-Brunius P. (2018). Hydrography of the gulf of Mexico using autonomous floats. J. Phys. Oceanogr. 48, 773–794. doi: 10.1175/JPO-D-17-0205.1
Heaton T. H. E. (1986). Isotopic studies of nitrogen pollution in the hydrosphere and atmosphere: A review. Chem. Geology: Isotope Geosci. Section 59, 87–102. doi: 10.1016/0168-9622(86)90059-X
Henschke N., Everett J. D., Suthers I. M., Smith J. A., Hunt B. P. V. V., Doblin M. A., et al. (2015). Zooplankton trophic niches respond to different water types of the western Tasman Sea: A stable isotope analysis. Deep-Sea Res. I: Oceanogr. Res. Pap. 104, 1–8. doi: 10.1016/j.dsr.2015.06.010
Hernández-Sánchez O. G., Camacho-Ibar V. F., Fernández Álamo M. A., Herzka S. Z. (2022). Nitrogen sources (NO3– vs N2 fixation) inferred from bulk δ15N values of zooplankton from the deep water region of the gulf of Mexico. J. Plankton Res. 44, 48–67. doi: 10.1093/plankt/fbab089
Hobson K. A., Barnett-Johnson R., Cerling T. (2010). “Using isoscapes to track animal migration,” in Isoscapes: understanding movement, pattern, and process on earth through isotope mapping. Eds. West J. B., Bowen G. J., Dawson T. E., Tu K. P. (Dordrecht: Springer) 273–298. doi: 10.1007/978-90-481-3354-3_13
Holl C. M., Villareal T. A., Payne C. D., Clayton T. D., Hart C., Montoya J. P. (2007). Trichodesmium in the western gulf of Mexico: 15N2 -fixation and natural abundance stable isotopic evidence. Limnol. Oceanogr. 52, 2249–2259. doi: 10.4319/lo.2007.52.5.2249
Hopkins T. L. (1982). The vertical distribution of zooplankton in the eastern gulf of Mexico. Deep-Sea Res. I: Oceanogr. Res. Pap. 29, 1069–1083. doi: 10.1016/0198-0149(82)90028-0
Hou W., Gu B., Zhang H., Gu J., Han B. P. (2013). The relationship between carbon and nitrogen stable isotopes of zooplankton and select environmental variables in low-latitude reservoirs. J. Limnol. 14, 97–104. doi: 10.1007/s10201-012-0388-2
Howe S., Miranda C., Hayes C. T., Letscher R. T., Knapp A. N. (2020). The dual isotopic composition of nitrate in the gulf of Mexico and Florida straits. J. Geophys. Res. Oceans 125, 1–17. doi: 10.1029/2020JC016047
Hunt B. P. V., Bonnet S., Berthelot H., Conroy B. J., Rachel A. F., Pagano M. (2016). Contribution and pathways of diazotroph-derived nitrogen to zooplankton during the VAHINE mesocosm experiment in the oligotrophic new Caledonia lagoon. Biogeosciences 13, 3131–3145. doi: 10.5194/bg-13-3131-2016
Jickells T. D., Buitenhuis E., Altieri K., Baker A. R., Capone D., Duce R. A., et al. (2017). A reevaluation of the magnitude and impacts of anthropogenic atmospheric nitrogen inputs on the ocean. Glob. Biogeochem. Cycles 31, 289–305. doi: 10.1002/2016gb005586
Jouanno J., Pallàs-Sanz E., Sheinbaum J. (2018). Variability and dynamics of the Yucatan upwelling: High-resolution simulations. J. Geophys. Res. Oceans 123, 1251–1262. doi: 10.1002/2017JC013535
Kelly T. B., Knapp A. N., Landry M. R., Selph K. E., Shropshire T. A., Thomas R. K., et al. (2021). Lateral advection supports nitrogen export in the oligotrophic open-ocean gulf of Mexico. Nat. Commun. 12, 3325. doi: 10.1038/s41467-021-23678-9
Klein P., Lapeyre G. (2009). The oceanic vertical pump induced by mesoscale and submesoscale turbulence. Annu. Rev. Mar. Sci. 1, 351–375. doi: 10.1146/annurev.marine.010908.163704
Knapp A. N., DiFiore P. J., Deutsch C., Sigman D. M., Lipschultz F. (2008). Nitrate isotopic composition between Bermuda and Puerto Rico: Implications for N2 fixation in the Atlantic ocean. Glob. Biogeochem. Cycles. 22 (3), 1–14. doi: 10.1029/2007gb003107
Knapp A. N., Thomas R. K., Stukel M. R., Kelly T. B., Landry M. R., Selph K. E., et al. (2021). Constraining the sources of nitrogen fueling export production in the gulf of Mexico using nitrogen isotope budgets. J. Plankton Res. 44, 1–19. doi: 10.1093/plankt/fbab049
Kolasinski J., Kaehler S., Jaquemet S. (2012). Distribution and sources of particulate organic matter in a mesoscale eddy dipole in the Mozambique channel (south-western Indian ocean): Insight from C and N stable isotopes. J. Mar. Syst. 96, 122–131. doi: 10.1016/j.jmarsys.2012.02.015
Kritee K., Sigman D. M., Granger J., Ward B. B., Jayakumar A., Deutsch C. (2012). Reduced isotope fractionation by denitrification under conditions relevant to the ocean. Geochim. Cosmochim. Acta 92, 243–259. doi: 10.1016/j.gca.2012.05.020
Kurle C. M., McWhorter J. K. (2017). Spatial and temporal variability within marine isoscapes: Implications for interpreting stable isotope data from marine systems. Mar. Ecol. Prog. Ser. 568, 31–45. doi: 10.3354/meps12045
Kürten B., Al-Aidaroos A. M., Kürten S., El-Sherbiny M. M., Devassy R. P., Struck U., et al. (2016). Carbon and nitrogen stable isotope ratios of pelagic zooplankton elucidate ecohydrographic features in the oligotrophic red Sea. Prog. Oceanogr. 140, 69–90. doi: 10.1016/j.pocean.2015.11.003
Kürten B., Painting S. J., Struck U., Polunin N. V. C., Middelburg J. J. (2013). Tracking seasonal changes in north Sea zooplankton trophic dynamics using stable isotopes. Biogeochemistry 113, 167–187. doi: 10.1007/s10533-011-9630-y
Landrum J. P., Altabet M. A., Montoya J. P. (2011). Basin-scale distributions of stable nitrogen isotopes in the subtropical north Atlantic ocean: Contribution of diazotroph nitrogen to particulate organic matter and mesozooplankton. Deep-Sea Res. I: Oceanogr. Res. Pap. 58, 615–625. doi: 10.1016/j.dsr.2011.01.012
Landry M. R., Swalethorp R. (2022). Mesozooplankton biomass, grazing and trophic structure in the bluefin tuna spawning area of the oceanic gulf of Mexico. J. Plankton Res. 44, 677–691. doi: 10.1093/plankt/fbab008
Layman C. A., Araujo M. S., Boucek R., Hammerschlag-Peyer C. M., Harrison E., Jud Z. R., et al. (2012). Applying stable isotopes to examine food-web structure: An overview of analytical tools. Biol. Rev. 87, 545–562. doi: 10.1111/j.1469-185X.2011.00208.x
Le-Alvarado M., Romo-Curiel A. E., Sosa-Nishizaki O., Hernández-Sánchez O., Barbero L., Herzka S. Z. (2021). Yellowfin tuna (Thunnus albacares) foraging habitat and trophic position in the gulf of Mexico based on intrinsic isotope tracers. PLoS One 16, e0246082. doi: 10.1371/journal.pone.0246082
Ledford T. C., Mortazavi B., Tatariw C., Mason O. U. (2020). Elevated nutrient inputs to marshes differentially impact carbon and nitrogen cycling in two northern gulf of Mexico saltmarsh plants. Biogeochemistry 149, 1–16. doi: 10.1007/s10533-020-00656-9
Lee-Sánchez E., Camacho-Ibar V. F., Velásquez-Aristizábal J. A., Valencia-Gasti J. A., Samperio-Ramos G. (2022). Impacts of mesoscale eddies on the nitrate distribution in the deep-water region of the gulf of Mexico. J. Mar. Syst. 229, 103721. doi: 10.1016/j.jmarsys.2022.103721
Lenes J. M., Heil C. A. (2010). A historical analysis of the potential nutrient supply from the N2 fixing marine cyanobacterium Trichodesmium spp. to Karenia brevis blooms in the eastern gulf of Mexico. J. Plankton Res. 32, 1421–1431. doi: 10.1093/plankt/fbq061
Lindo-Atichati D., Bringas F., Goni G., Muhling B., Muller-Karger F. E., Habtes S. (2012). Varying mesoscale structures influence larval fish distribution in the northern gulf of Mexico. Mar. Ecol. Prog. Ser. 463, 245–257. doi: 10.3354/meps09860
Loick-Wilde N., Fernández-Urruzola I., Eglite E., Liskow I., Nausch M., Schulz-Bull D., et al. (2019). Stratification, nitrogen fixation, and cyanobacterial bloom stage regulate the planktonic food web structure. Glob. Change Biol. 25, 794–810. doi: 10.1111/gcb.14546
Lorrain A., Graham B. S., Popp B. N., Allain V., Olson R. J., Hunt B. P. V., et al. (2015). Nitrogen isotopic baselines and implications for estimating foraging habitat and trophic position of yellowfin tuna in the Indian and pacific oceans. Deep-Sea Res. II: Top. Stud. Oceanogr. 113, 188–198. doi: 10.1016/j.dsr2.2014.02.003
Macko S. A., Entzeroth L., Parker P. L. (1984). Regional differences in nitrogen and carbon isotopes on the continental shelf of the gulf of Mexico. Naturwissenschaften 71, 374–375. doi: 10.1007/BF00410746
Marconi D., Alexandra Weigand M., Rafter P. A., McIlvin M. R., Forbes M., Casciotti K. L., et al. (2015). Nitrate isotope distributions on the US GEOTRACES north Atlantic cross-basin section: Signals of polar nitrate sources and low latitude nitrogen cycling. Mar. Chem. 177, 143–156. doi: 10.1016/j.marchem.2015.06.007
Martinez M. A., Hereu C. M., Arteaga M. C., Jiménez-Rosenberg S. P., Herzka S. Z., Saavedra-Flores A., et al (2021). Epipelagic zooplankton diversity in the deep water region of the gulf of Mexico: a metabarcoding survey. ICES J. Mar. Sci. 78, 3317–3332. doi: 10.1093/icesjms/fsab090
Martínez-López B., Zavala-Hidalgo J. (2009). Seasonal and interannual variability of cross-shelf transports of chlorophyll in the gulf of Mexico. J. Mar. Syst. 77, 1–20. doi: 10.1016/j.jmarsys.2008.10.002
Mateos-Jasso A., Zavala-Hidalgo J., Romero-Centeno R., Allende-Arandía M. E. (2012). Variability of the thermohaline structure in the northern veracruz coral reef system, Mexico. Cont. Shelf Res. 50, 30–40. doi: 10.1016/j.csr.2012.10.001
McCarthy M. J., Newell S. E., Carini S. A., Gardner W. S. (2015). Denitrification dominates sediment nitrogen removal and is enhanced by bottom-water hypoxia in the northern gulf of Mexico. Estuaries Coast. 38, 2279–2294. doi: 10.1007/s12237-015-9964-0
McCutchan J. H., Lewis W. M., Kendall C., McGrath C. C. (2003). Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos 102, 378–390. doi: 10.1034/j.1600-0706.2003.12098.x
McMahon K. W., Hamady L. L., Thorrold S. R. (2013). A review of ecogeochemistry approaches to estimating movements of marine animals. Limnol. Oceanogr. 58, 697–714. doi: 10.4319/lo.2013.58.2.0697
Merino M. (1997). Upwelling on the Yucatan shelf: Hydrographic evidence. J. Mar. Syst. 13, 101–121. doi: 10.1016/S0924-7963(96)00123-6
Mompeán C., Bode A., Benítez-Barrios V. M., Domínguez-Yanes J. F., Escánez J., Fraile-Nuez E. (2013). Spatial patterns of plankton biomass and stable isotopes reflect the influence of the nitrogen-fixer Trichodesmium along the subtropical north Atlantic. J. Plankton Res. 35, 513–525. doi: 10.1093/plankt/fbt011
Moncreiff C., Sullivan M. (2001). Trophic importance of epiphytic algae in subtropical seagrass beds: Evidence from multiple stable isotope analyses. Mar. Ecol. Prog. Ser. 215, 93–106. doi: 10.3354/meps215093
Montoya J. P., Carpenter E. J., Capone D. G. (2002). Nitrogen fixation and nitrogen isotope abundances in zooplankton of the oligotrophic north Atlantic. Limnol. Oceanogr. 47, 1617–1628. doi: 10.4319/lo.2002.47.6.1617
Montoya J. P., Voss M., Capone D. G. (2007). Spatial variation in N2-fixation rate and diazotroph activity in the tropical Atlantic. Biogeosciences 4, 369–376. doi: 10.5194/bg-4-369-2007
Mulholland M. R., Bernhardt P. W., Heil C. A., Bronk D. A., O'Neil J. M. (2006). Nitrogen fixation and release of fixed nitrogen by Trichodesmium spp. in the gulf of Mexico. Limnol. Oceanogr. 51, 1762–1776. doi: 10.4319/lo.2006.51.4.1762
Mulholland M. R., Bernhardt P. W., Ozmon I., Procise L. A., Garrett M., O’Neil J. M., et al. (2014). Contribution of diazotrophy to nitrogen inputs supporting Karenia brevis blooms in the gulf of Mexico. Harmful Algae 38, 20–29. doi: 10.1016/j.hal.2014.04.004
Müller-Karger F. E., Smith J. P., Werner S., Chen R., Roffer M., Liu Y., et al. (2015). Natural variability of surface oceanographic conditions in the offshore gulf of Mexico. Prog. Oceanogr. 134, 54–76. doi: 10.1016/j.pocean.2014.12.007
Muñoz-Salinas E., Castillo M. (2015). Streamflow and sediment load assessment from 1950 to 2006 in the usumacinta and grijalva rivers (Southern Mexico) and the influence of ENSO. Catena 127, 270–278. doi: 10.1016/j.catena.2015.01.007
Oey L. Y., Ezer T., Lee H. C. (2005). Loop current, rings and related circulation in the gulf of Mexico: A review of numerical models and future challenges. Circ. Gulf Mexico: Observations Models 161, 31–56. doi: 10.1029/161GM04
Ohkouchi N., Ogawa N. O., Chikaraishi Y., Tanaka H., Wada E. (2015). Biochemical and physiological bases for the use of carbon and nitrogen isotopes in environmental and ecological studies. Prog. Earth Planet. Sci. 2, 1–17. doi: 10.1186/s40645-015-0032-y
Otis D. B., Le Hénaff M., Kourafalou V. H., McEachron L., Muller-Karger F. E. (2019). Mississippi River and campeche bank (Gulf of Mexico) episodes of cross-shelf export of coastal waters observed with satellites. Remote Sens. 11, 1–14. doi: 10.3390/RS11060723
Orcutt K. M., Lipschultz F., Gundersen K., Arimoto R., Michaels A. F., Knap A. H., et al. (2001). A seasonal study of the significance of N2 fixation by trichodesmium spp. at the Bermuda Atlantic time-series study (BATS) site. Deep Sea Res. Part II Top. Stud. Oceanogr. 48, 1583–1608. doi: 10.1016/S0967-0645(00)00157-0
Parnell A. C., Inger R., Bearhop S., Jackson A. L. (2010). Source partitioning using stable isotopes: Coping with too much variation. PLoS One 5, 1–5. doi: 10.1371/journal.pone.0009672
Pasqueron De Fommervault O., Perez-Brunius P., Damien P., Camacho-Ibar V. F., Sheinbaum J. (2017). Temporal variability of chlorophyll distribution in the gulf of Mexico: Bio-optical data from profiling floats. Biogeosciences 14, 5647–5662. doi: 10.5194/bg-14-5647-2017
Pérez-Brunius P., García-Carrillo P., Dubranna J., Sheinbaum J., Candela J. (2013). Direct observations of the upper layer circulation in the southern gulf of Mexico. Deep-Sea Res. II: Top. Stud. Oceanogr. 85, 182–194. doi: 10.1016/j.dsr2.2012.07.020
Perry R. I., Thompson P. A., Mackas D. L., Harrison P. J., Yelland D. R. (1999). Stable carbon isotopes as pelagic food web tracers in adjacent shelf and slope regions off British Columbia, Canada. Can. J. Fish. Aquat. Sci. 56, 2477–2486. doi: 10.1139/f99-194
Phillips D. L., Inger R., Bearhop S., Jackson A. L., Moore J. W., Parnell A. C., et al. (2014). Best practices for use of stable isotope mixing models in food-web studies. Can. J. Zoology 92, 823–835. doi: 10.1139/cjz-2014-0127
Post D. M. (2002). Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 83, 703–718. doi: 10.1890/0012-9658(2002)083[0703:USITET]2.0.CO;2
Rabalais N. N., Turner R. E., Wiseman W. J. (2001). Hypoxia in the gulf of Mexico. J. Environ. Qual. 30, 320–329. doi: 10.2134/jeq2001.302320x
Rabalais N. N., Turner R. E., Wiseman W. J. (2002). Gulf of Mexico hypoxia, a.k.a. “The dead zone”. Annu. Rev. Ecol. Evol. Syst. 33, 235–263. doi: 10.1146/annurev.ecolsys.33.010802.150513
Radabaugh K. R., Hollander D. J., Peebles E. B. (2013). Seasonal δ13C and δ15N isoscapes of fish populations along a continental shelf trophic gradient. Cont. Shelf Res. 68, 112–122. doi: 10.1016/j.csr.2013.08.010
Reyes-Mendoza O., Mariño-Tapia I., Herrera-Silveira J., Ruiz-Martínez G., Enriquez C., Largier J. L. (2016). The effects of wind on upwelling off cabo catoche. J. Coast. Res. 32, 638–650. doi: 10.2112/JCOASTRES-D-15-00043.1
Rooker J. R., Simms J. R., Wells R. D., Holt S. A., Holt G. J., Graves J. E., et al. (2012). Distribution and habitat associations of billfish and swordfish larvae across mesoscale features in the gulf of Mexico. PLoS One 7, e34180. doi: 10.1371/journal.pone.0034180
Salas-de-León D. A., Monreal-Gómez M. A., Signoret M., Aldeco J. (2004). Anticyclonic-cyclonic eddies and their impact on near-surface chlorophyll stocks and oxygen supersaturation over the campeche canyon, gulf of Mexico. J. Geophys. Res. Oceans 109, 1–10. doi: 10.1029/2002JC001614
Salmerón-García O., Zavala-Hidalgo J. (2011). Regionalization of the gulf of Mexico from space-time chlorophyll- a concentration variability. Ocean Dyn. 61, 439–448. doi: 10.1007/s10236-010-0368-1
Schiller R. V., Kourafalou V. H., Hogan P., Walker N. D. (2011). The dynamics of the Mississippi river plume: Impact of topography, wind and offshore forcing on the fate of plume waters. J. Geophys. Res. 116, C06029. doi: 10.1029/2010JC006883
Schmidt K., McClelland J. W., Mente E., Montoya J. P., Atkinson A., Voss M. (2004). Trophic-level interpretation based on δ15N values: Implications of tissue-specific fractionation and amino acid composition. Mar. Ecol. Prog. Ser. 266, 43–58. doi: 10.3354/meps266043
Seki M. P., Polovina J. J., Brainard R. E., Bidigare R. R., Leonard C. L., Foley D. G. (2001). Biological enhancement at cyclonic eddies tracked with GOES thermal imagery in Hawaiian waters. Geophys. Res. Lett. 28, 1583–1586. doi: 10.1029/2000GL012439
Selph E., Swalethorp R., Stukel R., Kelly B., Knapp N., Fleming K., et al. (2022). Phytoplankton community composition and biomass in the oligotrophic gulf of Mexico. J. Plankton Res. 44, 618–637. doi: 10.1093/plankt/fbab006
Sepúlveda-Lozada A., Mendoza-Carranza M., Wolff M., Saint-Paul U., Ponce-Mendoza A. (2015). Differences in food web structure of mangroves and freshwater marshes: evidence from stable isotope studies in the southern gulf of Mexico. Wetl. Ecol. Manage. 23, 293–314. doi: 10.1007/s11273-014-9382-2
Sigman D. M., Casciotti K. L. (2001). Nitrogen isotopes in the ocean. Encyclopedia Ocean Sci. 3, 1884–1894. doi: 10.1006/rwos.2001.0172
Sigman D. M., Fripiat F. (2019). Nitrogen isotopes in the ocean. Encyclopedia Ocean Sci., 263–278. doi: 10.1016/B978-0-12-409548-9.11605-7
Somes C. J., Schmittner A., Galbraith E. D., Lehmann M. F., Altabet M. A., Montoya J. P., et al. (2010). Simulating the global distribution of nitrogen isotopes in the ocean. Glob. Biogeochem. Cycles 24, 1–16. doi: 10.1029/2009GB003767
Sosa-Gutiérrez R., Pallàs-Sanz E., Jouanno J., Chaigneau A., Candela J., Tenreiro M. (2020). Erosion of the subsurface salinity maximum of the loop current eddies from glider observations and a numerical model. J. Geophys. Res. Oceans 125, e2019JC015397. doi: 10.1029/2019JC015397
Troina G. C., Dehairs F., Botta S., Tullio J.C.Di, Elskens M., Secchi E. R. (2020). Zooplankton-based δ13C and δ15N isoscapes from the outer continental shelf and slope in the subtropical western south Atlantic. Deep-Sea Res. I: Oceanogr. Res. Pap. 159, 103235. doi: 10.1016/j.dsr.2020.103235
Turner J. T. (2015). Zooplankton fecal pellets, marine snow, phytodetritus and the ocean’s biological pump. Prog. Oceanogr. 130, 205–248. doi: 10.1016/j.pocean.2014.08.005
Vanderklift M. A., Ponsard S. (2003). Sources of variation in consumer-diet δ15N enrichment: A meta-analysis. Oecologia 136, 169–182. doi: 10.1007/s00442-003-1270-z
Waite A. M., Muhling B. A., Holl C. M., Beckley L. E., Montoya J. P., Strzelecki J., et al. (2007). Food web structure in two counter-rotating eddies based on δ15N and δ13C isotopic analyses. Deep-Sea Res. II: Top. Stud. Oceanogr. 54, 1055–1075. doi: 10.1016/j.dsr2.2006.12.010
Walker N. D., Wiseman W. J., Rouse L. J., Babin A. (2005). Effects of river discharge, wind stress, and slope eddies on circulation and the satellite-observed structure of the Mississippi river plume. J. Coast. Res. 216, 1228–1244. doi: 10.2112/04-0347.1
Wells R. J. D., Rooker J. R., Quigg A., Wissel B. (2017). Influence of mesoscale oceanographic features on pelagic food webs in the gulf of Mexico. Mar. Biol. 164, 1–11. doi: 10.1007/s00227-017-3122-0
Wells R. J. D., Rooker J. R. (2009). Feeding ecology of pelagic fish larvae and juveniles in slope waters of the gulf of Mexico. J. Fish Biol. 75, 1719–1732. doi: 10.1111/j.1095-8649.2009.02424.x
Wissel B., Fry B. (2003). Sources of particulate organic matter in the Mississippi river, USA. Large Rivers 15, 105–118. doi: 10.1127/lr/15/2003/105
Wissel B., Fry B. (2005). Tracing Mississippi river influences in estuarine food webs of coastal Louisiana. Oecologia 144, 659–672. doi: 10.1007/s00442-005-0119-z
Wormuth J. H., Ressler P. H., Cady R. B., Harris E. J. (2000). Zooplankton and micronekton in cyclones and anticyclones in the northeast gulf of Mexico. Gulf Mexico Sci. 18, 3. doi: 10.18785/goms.1801.03
Yamamuro M., Kayanne H., Minagawao M. (1995). Carbon and nitrogen stable isotopes of primary producers in coral reef ecosystems. Limnol. Oceanogr. 40, 617–621. doi: 10.4319/lo.1995.40.3.0617
Zavala-García F., Flores-Coto C., Espinosa-Fuentes M.de l. (2016). Relación entre la biomasa zooplanctónica y las descargas de aguas continentales, en el sur del golfo de méxico, (1984-2001). Rev. Biol. Mar. Oceanogr 51, 21–31. doi: 10.4067/S0718-19572016000100003
Zavala-Hidalgo J., Gallegos-García A., Martínez-López B., Morey S. L., O’Brien J. J. (2006). Seasonal upwelling on the Western and southern shelves of the gulf of Mexico. Ocean Dyn. 56, 333–338. doi: 10.1007/s10236-006-0072-3
Keywords: mesozooplankton, isoscape, nitrogen sources, Bayesian mixing model, N2 fixation, Gulf of Mexico (GOM)
Citation: Hernández-Sánchez OG, Camacho-Ibar VF, Barbero L, Herguera JC and Herzka SZ (2023) A gulf-wide synoptic isoscape of zooplankton isotope ratios reveals the importance of nitrogen fixation in supporting secondary production in the central Gulf of Mexico. Front. Mar. Sci. 9:1025387. doi: 10.3389/fmars.2022.1025387
Received: 22 August 2022; Accepted: 21 December 2022;
Published: 12 January 2023.
Edited by:
Travis Elsdon, Chevron, United StatesReviewed by:
Thomas Bryce Kelly, University of Alaska Fairbanks, United StatesCopyright © 2023 Hernández-Sánchez, Camacho-Ibar, Barbero, Herguera and Herzka. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sharon Z. Herzka, c2hlcnprYUBjaWNlc2UubXg=
†ORCID: Oscar Gabriel Hernández-Sánchez, orcid.org/0000-0003-3525-4003
Víctor F. Camacho-Ibar, orcid.org/0000-0003-2424-1979
Leticia Barbero, orcid.org/0000-0002-8858-5247
Juan Carlos Herguera, orcid.org/0000-0001-8335-2607
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.