
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Mar. Sci., 02 November 2022
Sec. Marine Conservation and Sustainability
Volume 9 - 2022 | https://doi.org/10.3389/fmars.2022.1018118
This article is part of the Research TopicAquatic One Health—The Intersection of Marine Wildlife Health, Public Health, and Our OceansView all 10 articles
 Casandra Gálvez1
Casandra Gálvez1 Mónica Tenorio-Osorio2,3
Mónica Tenorio-Osorio2,3 Isabel Hernández-Candelario3
Isabel Hernández-Candelario3 Christian Alejandro Delfín-Alfonso3,4
Christian Alejandro Delfín-Alfonso3,4 Eduardo Morteo3*
Eduardo Morteo3*Mycotic diseases are considered a worldwide growing concern related to public health. Lobomycosis like disease (LLD) (Lacazia loboi) is a chronic and progressive infection in skin of humans and small cetaceans present in both sides of the Americas, including Mexico but information is still limited. Marine predators are indicators of potential risks for human and wildlife health, including fungal diseases like LLD in bottlenose dolphins. Here we report the first findings of an initial assessment in LLD epidemiology, pathology, and behavioral constraints of coastal bottlenose dolphins (Tursiops truncatus) from the Southwestern Gulf of Mexico (SWGM). Overall, LLD prevalence in the population was low, within ranges reported for the species, and only in highly associated unisexual pairs near the Alvarado coastal waters. Photo-identified individuals exhibited an annual increase in average progression for LLD skin lesions. Gross lesions and skin biopsy evidenced mycotic structures and subcutaneous alterations associated to LLD. Habitat quality, demographic, and social characteristics of bottlenose dolphins are likely influencing LLD geographical expansion and temporal prevalence, but global and local climate variability may influence LLD epidemiology, implying a potential risk for human and dolphin health from coastal communities at the SWGM.
Lacaziosis like disease (LLD), formerly known as Lobomycosis, is a mycotic (Lacazia loboi) chronic skin disease that affects several species of small cetaceans with potential transmission to humans (Reif et al., 2013). Therefore, coastal bottlenose dolphins (Tursiops truncatus) are useful indicators of risk of emerging diseases in human populations (Bossart, 2007, Bossart et al., 2019). Gross findings in the skin of T. truncatus affected by LLD are consistent with white to gray nodules that may ulcerate and form large plaques, particularly in fins, head, fluke, and caudal peduncle (Reif et al., 2006; Van Bressem et al., 2007; Murdoch et al., 2008; Ueda et al., 2013). The impact of LLD in marine mammals is unclear, nonetheless, there is a growing concern about its persistence and progression as well as high prevalence, especially in T. truncatus populations (Van Bressem et al., 2007; Van Bressem et al., 2015; Siciliano et al., 2008; Daura-Jorge and Simões-Lopes, 2011; Bessesen et al., 2014; Ramos et al., 2018). Immunodeficiency seems to facilitate its occurrence acting as an opportunistic infection (Reif et al., 2009) and as potential death cause due to an eventual immunologic dysfunction in individuals (Bossart et al., 2019). Although, the etiology and epidemiology of LLD in most cetaceans worldwide are still largely unknown and recently associated with Paracoccidioides brasiliensis var. ceti in T. truncatus (Vilela et al., 2016), LLD transmission among individuals has been linked to social behavior in coastal T. truncatus, suggesting horizontal contagion and geographic dissemination related to sex (Tenorio-Osorio, 2015; Félix et al., 2019).
Additionally, chemical (e.g., pollutants) and biological (e.g., pathogens) characteristics of marine habitats are thought to play a role in LLD presence and prevalence, particularly in small cetaceans inhabiting sites impacted by anthropogenic activities like inshore or estuarine populations (Bessesen et al., 2014; Rotstein et al., 2009; Van Bressem et al., 2009a b; Félix et al., 2019). However, LLD has also been identified in offshore T. truncatus, suggesting an increase in its geographical range (Rotstein et al., 2009). This is relevant considering that mycotic diseases are a worldwide growing public health concern in the face of climate change (e.g., ocean warming) (Seyedmousavi et al., 2015; Gnat et al., 2021). For instance, the expansion of marine mammal foraging areas due to prey shortage during marine warm conditions may increase individual exposure to new and polluted environments (chemical and biological), including pathogens like L. loboi (Learmonth et al., 2006; Van Bressem et al., 2009b),
Currently, LLD has been recorded in T. truncatus from both sides of the Americas; in the Pacific (Van Bressem et al., 2015, Ramos et al., 2018) and Atlantic (Vilela et al., 2016), including South Africa (Van Bressem et al., 2015), but apparently there are no records of LLD from the Southwestern Gulf of Mexico (SWGM). Therefore, this research aims to provide an epidemiological baseline of LLD in T. truncatus from SWGM, including the progression of gross skin lesions in the dorsal fin and their relation to the social behavior of infected individuals, to fill in the geographic gap and improve our overall understanding of LLD presence in small odontocetes.
Coastal T. truncatus from Veracruz state (SWGM portion) have been studied intermittently since the early 1990’s using primarily photo-identification (Heckel, 1992; Martínez-Serrano et al., 2011; Morteo et al., 2014, Morteo et al., 2017, Morteo et al., 2019; Bolaños-Jiménez et al., 2021; Bolaños-Jiménez et al., 2022). The studied area comprises roughly three sites in the coastal strip from shore to the 30 – 40 m isobath along 230 km off the waters of 1) Nautla, 2) Veracruz Reef System (VRS), and 3) Alvarado lagoon system (ALV) (Figure 1). Weather variability follows a tropical pattern with three periods: rainy (July-October), windy (November-February), and dry season (March-June) (Bolaños-Jiménez et al., 2022) that provide a variable influx of organic material discharged from the continent to the sea that, in addition to tidal exchange, produce important differences in salinity and temperature (Morán-Silva et al., 2005). Rivers and lagoons sustain local patches of mangrove habitats, where marine sediments and water contain traces of organochlorine pesticides that affect water quality and are toxic to marine species (Vázquez-Botello et al., 2019). Also, artisanal riverine fishing is one of the most important commercial activities carried out all year in the area and antagonistic interactions with T. truncatus are frequently recorded (Rechimont et al., 2018; Morales-Rincon et al., 2019).
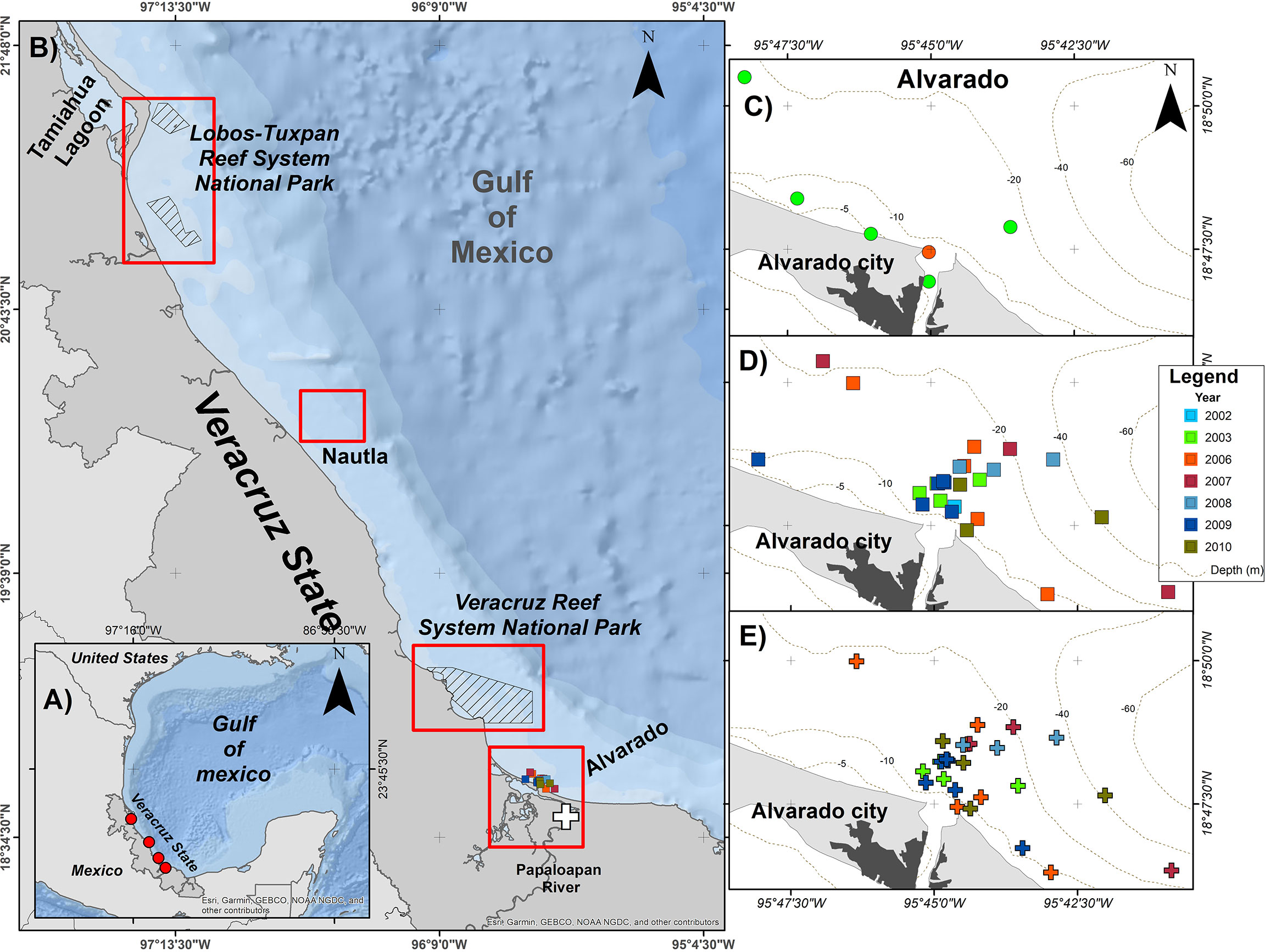
Figure 1 Study area along the coastal waters off Veracruz. (A) General location of bottlenose dolphin study sites in the Southwestern Gulf of Mexico (red circles). (B) Location of the sites with photo-ID catalogues and studies on skin lesions in bottlenose dolphins. (C) Recorded sightings of individual EN05. (D) individual EN10. (E) individual TN15. The white cross shows the location of the stranding of the dead male EN10 in the Papaloapan river.
T. truncatus from the SWGM were photo-identified using their dorsal fins during different survey campaigns at the three sites, during 2002 – 2003 (Nautla and ALV) (photographic negatives), over 2005 – 2007 (digital pictures at the VRS), and 2006 – 2010 (digital pictures at ALV). Individuals were identified by their natural marks according to standard photo-identification procedures using SLR cameras equipped with 70 – 300 mm lenses (Morteo et al., 2014; Bolaños-Jiménez et al., 2022). Only long-lasting marks such as cuts, nicks, and deformities were used for individual identification. All photographs were graded for technical quality (e.g., focus, sharpness, lighting, angle, and proportion of dorsal fin exposure) and distinctiveness. We followed a previously tested protocol for the selection of photographs (Urian et al., 2015), and selected only good-quality images (Q1 and Q2) of individuals with conspicuous permanent markings (D1 and D2) to minimize misidentification. Catalogs were kept and analyzed at the Marine Mammal Laboratory (LabMMar, IIB-ICIMAP) at Universidad Veracruzana (UV). Population estimates for T. truncatus of the studied area have yielded between 69 and 636 individuals, with daily abundances of 187 (±132 SD) dolphins for Nautla, over 45 (±23 SD) for the VRS (Morteo et al., 2019), and monthly abundances between 70 (±14 SD) (Bolaños-Jiménez et al., 2022) and 125 (±52 SD) (Morteo et al., 2017) for ALV, when considering the more resident fraction of the population and the whole sample (residents and transients), respectively.
The presence of LLD was visually analyzed when gross lesions were observed (i.e., raised light grey whitish to pinkish granulomas, nodules, and ulcers) (Van Bressem et al., 2007; Daura-Jorge and Simões-Lopes, 2011). Relative size and progression of LLD were determined by measuring the lesion areas and dividing them by the total surface of the dorsal fin (Daura-Jorge and Simões-Lopes, 2011). Thus, successive images of each individual were projected onto millimeter sheets and the areas were measured. The first and last images of each animal were used to determine changes in the proportion of the affected area. We computed the prevalence (P) of the LLD lesions as the proportion of the animals in the population that have potentially acquired this disease, by dividing the number of affected animals (NI), by the total number of photo-identified individuals (FI) and multiplied by 100.
Also, whenever individuals with LLD were identified, photographic records of these animals were used to measure the level of association between them and the other individuals identified within each study area, using SocProg 2.4 (Whitehead, 2009). We used a half-weight index which calculates coefficients of association (COA) between pairs of animals within each social network, considering the total number of times each dolphin was observed and the number of times each pair of dolphins were observed together (Cairns and Schwager, 1987). Values range from zero for a pair of animals that were never seen together to one for animals that are always seen together. The mean association rate for all identified individuals observed on more than 5 opportunities/encounters (Morteo et al., 2014) was used to determine the number of significant pairs by a randomization test (Bejder et al., 1998) using 1 000 iterations. The mean values of non-null associations (i.e., >0) for all animals were calculated and compared against those exclusively from individuals with LLD through a test for differences in means (Tenorio-Osorio, 2015; Félix et al., 2019). All significance levels were set at α= 0.05.
Finally, in February 2022 a dead old (>20 y) (based on photo-ID records and teeth attrition) male T. truncatus was found at the Papaloapan River in Tlacotalpan, Veracruz (Figure 1), which exhibited diffuse dermatological gross lesions presumptively linked to LLD and matched one of the adult dolphins in our photo-ID catalog since 2002. Thus, a skin biopsy was collected and preserved in 10% neutral buffered formalin for histology description with hematoxylin and eosin stain. Fungal identification with Methenamine Silver stain was carried out by the Diagnostic Laboratory at Centro Veterinario de Xalapa, A.C. in Xalapa, Veracruz, Mexico.
We analyzed 20, 665 images (Nautla=2,779, VRS=8,664, ALV=9, 212) from 524 individuals at SWGM, only 283 images from three dolphins (EN10, EN05, and TN10) exhibited gross lesions in the dorsal fins presumably related to LLD (Table 1). These individuals were initially classified as males based on their behavioral cues (i.e., synchronized swimming and no affiliation with calves) and later confirmed through visual inspection of their genital area on the field. Thus, the prevalence for all identified dolphins at SWGM was estimated between 0.47 and 0.57% considering the estimated total abundance for the area (i.e., 636 dolphins) and the total of photo-identified individuals (i.e., 524 individuals) (Table 1). However, considering that only individuals within the ALV area were affected with LLD (Figures 2A, B), thus according to photo identified individuals the prevalence ranged from 4.6% (from photo-identified dolphins in 2002 – 2003) to 1.4% (identified dolphins in 2006 – 2010) (Table 1), and between 2.4 and 4.3% when considering the total abundances at Alvarado area (Min-Max: 70 – 125 individuals, see Materials And methods).
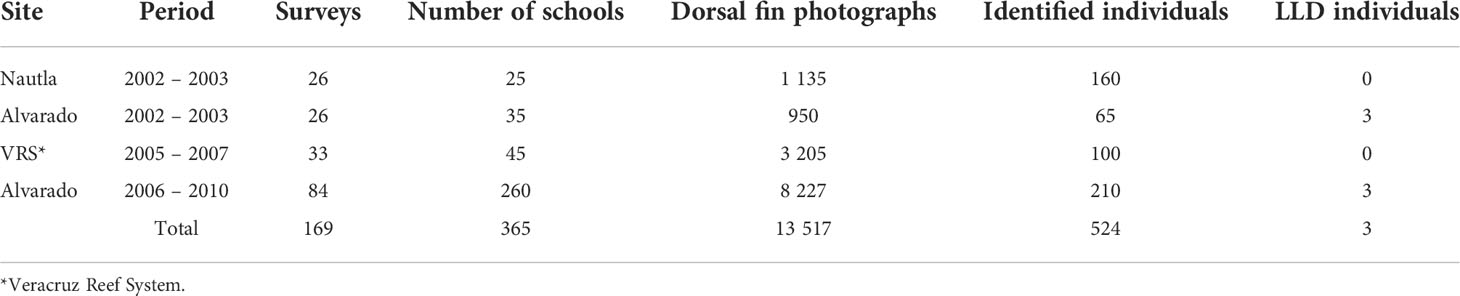
Table 1 Surveys and sightings of coastal bottlenose dolphins with and without gross LLD skin lesions at Veracruz, Gulf of Mexico.

Figure 2 Wild-bottlenose dolphins from Alvarado affected by LLD. (A) Presence of individual EN05 at Alvarado lagoon close to rural communities exhibiting skin gross lesion possible related to LLD (2006), (B) pair of males, EN10 (right) and TN15 (left) with early LLD gross lesions in dorsal fins and dorsum (2006), (C) re-sighting of individual EN10 (left) and TN15 (right) with progression of skin lesions in dorsal fins and lateral flanks (2019), (D) dorsal fin of individual EN10 stranded at Papaloapan River with severe skin gross lesions, (E) multiple nodules ulcerated in head, anterior dorsum near to blowhole and lateral flank (left), (F) Microscopic image of dermal tissue of individual EN10. Note presence of rounded deep purple structures connected by a tube-shaped forms, typical of fungus Lacazia loboi. Methenamine Silver stain. Magnification, 100x.
Early gross lesions described in dorsal fins of the three male T. truncatus (EN10, EN05, and TN10) presumably affected by LLD were whitish and small nodular areas in dorsal fins and dorsum (100%; 2003 – 2007) that later raised in middle plaques of verrucous lesions and to light grey whitish and pinkish granulomas (66.6%; 2009 – 2010) in dorsal fins, including edges, anterior dorsum and middle flanks (Figures 2B, C) that finally progressed into whitish extensive plaques of serpiginous aspect, ulcerated with deformation of dorsal fin and presence of raised firm nodules at the beak, head, anterior dorsum, flanks and fluke (33.3%; 2019 – 2022) (Figures 2B–E). Area measurements of the same images from each of the three individuals were repeated three times with an average variation of only 0.25% (± 0.037 SD); also, the measurement error (average coefficient of variation) of different images of each individual within the same sighting was 4% ( ± 4.1 SD). LLD skin gross lesions in dorsal fins persisted and progressed during the study in all three individuals with an increasing average annual rate estimated at 0.61 ( ± 0.61 SD) times the size of the original lesion (Table 2). The highest progression rate occurred in individual TN15, followed by EN10 and EN05 photo-identified during 2003-2010 (Figures 2A–E).
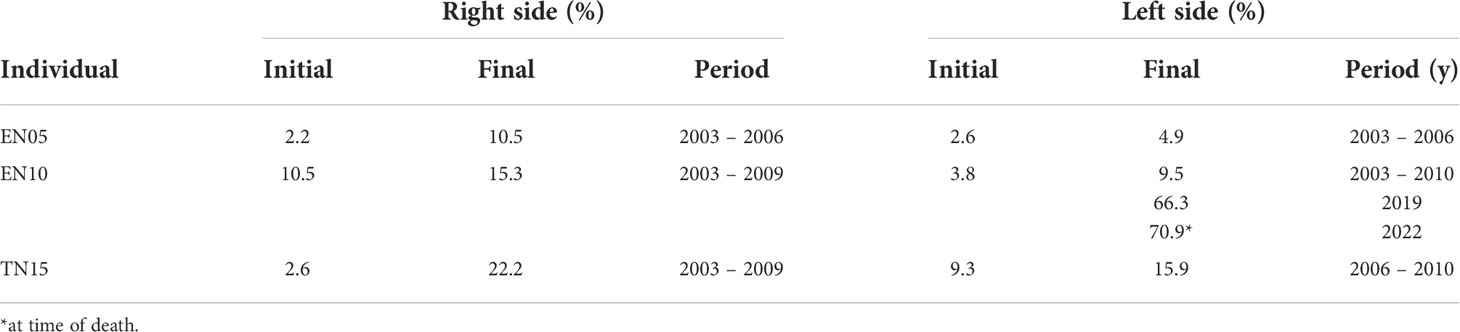
Table 2 Temporal progression of gross skin lesions (%) linked to LLD in dorsal fins of photo-identified bottlenose dolphins from Alvarado area.
Interestingly, the stranded dolphin found in the Papaloapan River in 2022, was recorded 24.5 km upstream (in freshwater) and identified as individual EN10 from the ALV population since 2002, and re-sighted alive in the ALV area in 2019. Total length was 276 cm and body weight was estimated between 280 – 300 kg. The cause of death was undetermined, but the aged male exhibited poor body condition and severe LLD skin gross lesions in the head, dorsal fin, flukes, and caudal peduncle (Figures 2D, E), as well as old injuries likely related to fisheries interactions. The temporal progression and severity of skin lesions in this individual were evident until his death (in 2022) (Table 2 and Figures 2B–D). Histological analyses of biopsied skin revealed granulomatous, nodular, multifocal dermatitis with ulceration and epithelial hyperplasia as well infiltration of neutrophils, macrophages, and giant cells but scarce lymphocytes and plasmatic cells infiltration. The Methenamine silver stain evidenced multiple deep blue round-shaped structures of 6 – 15 µm in diameter interconnected by short tubular structures, which are typical of L. loboi (Figure 2F).
Overall, COA values (i.e., among all identified individuals within each study area) were generally low (), and in the ALV population (the only with LLD), from the 53 542 possible combinations, only 237 pairs (0.04%) were non-random. From the latter, 11% had low or very low association values (<0.4), 68% were moderate (0.4 – 0.6), and the rest were high (7%, 0.6 – 0.8) or very high (3%, 0.8 – 1.0). Specific COA among males affected by LLD (EN05, EN10, and TN10) yielded high values and thus stable associations (), particularly between EN10-TN15 (0.80 from 2003 to 2010) (Figures 2B, C). Individuals EN05 and TN15 were also paired with LLD-negative individuals within the population with higher-than-average association rates (e.g., a possible female named TN10, COA= 0.73 from 2006 to 2010, and other adult individual of undetermined sex named UP22, COA= 0.35, from 2007 to 2010). Therefore, mean COA values for non-null paired combinations of all individuals identified on multiple occasions within the same study area (i.e., ALV) were similar to the overall mean for two of the three individuals (EN10 and TN15) with LLD lesions (, , p > 0.05), but it was very low in individual EN05 ().
Our study confirmed the presence of LLD in T. truncatus at the SWGM, supporting the value of photo-identification surveys as a non-invasive research tool to monitor skin gross lesions progression and histological analyses in stranded individuals as an alternative for epidemiological surveillance studies contributing to health assessment programs in Veracruz, Mexico.
The overall prevalence of LLD in T. truncatus from SWGM, which only included individuals from ALV, was within of the ranges observed in other coastal dolphins from the east and west coasts of Florida (<1 to 16.9%) (Wagner et al., 2003; Murdoch et al., 2008), as well Tursiops spp. from South America, such as Ecuador (2.33 – 14.3%) (Van Bressem et al., 2015; Félix et al., 2019), but considerably lower than populations from Brazil (9 – 16.7%; Daura-Jorge and Simões-Lopes, 2011; Van Bressem et al., 2015), Costa Rica (13.2 – 16.1%; Bessesen et al., 2014) and cetaceans from Japan (15.8 – 23.1%; Van Bressem et al., 2013), the Indian Ocean (8.4%; Kiszka et al., 2009), and Guerrero, Mexico (9.8%; Ramos et al., 2018). Such geographical and temporal differences are expected for infectious diseases both in human and wild animal populations due to their relation to environmental, ecological, and socio-economic factors (Daszak et al., 2001; Jones et al., 2008), including mycotic diseases like LLD (Seyedmousavi et al., 2015; Gnat et al., 2021).
Studies of LLD in T. truncatus are mostly from inshore (coastal and lagoon) populations, and diagnosis is often limited or inconclusive because samples are hard to collect (Van Bressem et al., 2007; Van Bressem et al., 2009b; Kiszka et al., 2009; Ramos et al., 2018). In fact, other photo-ID studies on the species at the north of our study area (i.e., Lobos-Reef System to Nautla, see Figure 1), found no evidence of LLD skin lesions in the 85 photo-identified individuals during 27 surveys in 2016 (Alvízar-Cruz, 2018). However, the presence of LLD in T. truncatus at the SWGM during the current research, even at a low prevalence reflects population vulnerability and potentially subjacent health problems (e.g., immunosuppression) in individuals that should be explored.
In cetaceans, LLD susceptibility has been associated to chemical pollution (agricultural, industrial contaminants) that reduces immunological efficiency and facilitates opportunistic infections (Van Bressem et al., 2009b; Reif et al., 2006; Reif et al., 2009; Van Bressem et al., 2009a; Bossart et al., 2019). However, it is not clear whether other chemical and physical characteristics of the habitat (e.g., lower salinity and warmer marine conditions) may favor susceptibility among small cetaceans populations (Reif et al., 2006; Bessesen et al., 2014; Van Bressem et al., 2015; Ramos et al., 2018). All our study areas are characterized by freshwater intrusion derived from the influence of rivers that flow near the coastal habitat of T. truncatus at SWGM, particularly, the ALV region, having the second largest hydrographic basin (354 km) in Mexico (Morteo et al., 2019). Toxicological studies have identified organochlorine pesticides, aromatic hydrocarbons, and heavy metals (Pb, Hg) from petrogenic origin (Vázquez-Botello et al., 2018) in both fish and hair from local human communities, especially at the margins of the Papaloapan River in ALV (Guentzel et al., 2007). The ALV T. truncatus population is mostly composed of small and single-sex groups, with very fluid and open affiliations and generally low levels of association (Morteo et al., 2014; García-Vital et al., 2015), with a core community of resident individuals compared to Nautla and the VRS (Bolaños-Jiménez et al., 2021; Bolaños-Jiménez et al., 2022). However, coefficients of association among infected T. truncatus from ALV were high in contrast to non-infected individuals. It is possible that temporal and spatial exposure to chemical pollutants enhanced by population residency and social structure, may favor LLD, similar to what was suggested in resident T. truncatus from Mexico and Belize (Ramos et al., 2018). However, in both cases, ecotoxicological studies are necessary to clarify the relation between habitat pollution and disease epidemiology.
In small cetaceans, chronic stress increases vulnerability to LLD and other skin diseases (Reif et al., 2009; Van Bressem et al., 2013). Different immune response suggests sex-biased susceptibility (Van Bressem et al., 2018), as LLD severity and the highest prevalence has been reported in males of the Indo-Pacific bottlenose dolphins (Tursiops aduncus) (Reif et al., 2009; Van Bressem et al., 2013), and T. truncatus from Mexico (Tenorio-Osorio, 2015) and Ecuador (Félix et al., 2019). In these last cases, 100% and 86% of individuals with LLD were adult males, closely associated in small groups (pairs) with higher association coefficients. These findings may imply social vulnerability related to sex (males) and age class structure (adults) in T. truncatus at the SWGM.
The close associations among LLD positive T. truncatus at ALV suggest horizontal transmission during their physical interactions, given the typical social affiliations of the community (Tenorio-Osorio, 2015; Morteo et al., 2017), and the likelihood to present superficial dermal wounds over their development (Paniz-Mondolfi et al., 2007; Murdoch et al., 2008; Kiszka et al., 2009; Murdoch et al., 2010; Daura-Jorge and Simões-Lopes, 2011). Furthermore, LLD in our study suggests a geographical pattern (Daura-Jorge and Simões-Lopes, 2011; Félix et al., 2019); for instance, Ruiz-Hernández (2014) and Morteo et al. (2019) already showed limited individual exchange (<1 individual y-1) among the SWGM. Therefore, behavioral patterns (e.g., social and feeding) and individual residency, combined with local habitat characteristics (e.g., pollution) could be limiting contact among infected individuals and preventing LLD transmission to T. truncatus from Nautla and VRS. However, extended epidemiological efforts in communities (dead and alive) are warranted to explore this hypothesis, as well as the potential differences in age classes and sex.
LLD skin gross lesions described in T. truncatus from ALV are similar to macroscopic findings reported in small cetaceans worldwide (Moreno et al., 2008; Murdoch et al., 2008; Daura-Jorge and Simões-Lopes, 2011; Van Bressem et al., 2013; Van Bressem et al., 2015). Also, microscopic characteristics of these lesions in the stranded dolphin at Papaloapan River matched the histological description in the skin of small cetaceans affected by LLD (Migaki et al., 1971; Haubold et al., 2000; Moreno et al., 2008; Rotstein et al., 2009; Ueda et al., 2013). Thus, according to macroscopic and microscopic findings in skin of individual EN10 and due to the lack of molecular evidence, it is plausible that the other two males in ALV were affected by L. loboi. Admittedly, other nodular and granulomatous progressive skin diseases reported in small cetaceans, including Tursiops sp., that are associated to fungus such as Fusarium spp., Paracoccidioides brasiliensis and Trichophyton spp., and bacteria like Streptococcus iniae may be causing these lesions (Van Bressem et al., 2008; Van Bressem et al., 2013; Vilela et al., 2016).
Progressive development of LLD skin gross lesions progression in cetaceans is variable within similar and different species, ranging from 5 to 15 y (Murdoch et al., 2008; Ramos et al., 2018). This reflects the chronicity of long-lived species exposed to LLD, linked to immune system activation and the inability to eliminate prolonged inflammation that could favor the persistency and progression of the disease (Bossart et al., 2019). This seems to be the case for the photographed dead male (EN10) found at the Papaloapan River over the course of 20 y. This reflects the chronicity of LLD in small cetaceans and an increase of skin gross lesion progression during aging, possibly linked to immune senescence, implying a limited disease control and vulnerability to opportunistic disease infections (Venn-Watson et al., 2011; Venn-Watson et al., 2020) like LLD. In some cases, this may be aggravated by subjacent conditions, such as immunosuppression by chronic exposure to heavy metals and hydrocarbons (De Guise et al., 2002; Desforges et al., 2016) present in ALV (Guentzel et al., 2007; Vázquez-Botello et al., 2018) and could influence LLD skin gross lesion progression in individuals.
Increasing LLD prevalence in dolphin populations could represent a potential threat for long-term survival (Van Bressem et al., 2015); however, this does not seem to be the case within the SWGM, since within the timeframe of the photo-ID surveys (i.e., 2002 – 2003, 2005 – 2007, and 2006 – 2010) there were no new infected individuals. Thus, social structure, behavior, and residency seem to conform to a positive mechanism for disease control through potential social isolation and the natural death of affected T. truncatus. It is also possible that LLD negative (not gross skin lesions) individuals such as UP22 have higher contagion risk because of their regular and long-lasting associations with infected individuals. In fact, individual EN10 was spotted in subsequent surveys (2019) with a new and unidentified individual with LLD skin lesions. Therefore, intrusion and contact with new individuals could represent another source of LLD exposure.
Furthermore, attention must be drawn to the fact that marine pollution remains a health risk for T. truncatus in the Gulf of Mexico, due to the elevated level of oil-related activities (Vázquez-Botello et al., 2004, Vázquez-Botello et al., 2018) and the explosion of the Deepwater Horizon oil spill in 2010 with short- and long-term consequences in the health and survival of T. truncatus in the northern Gulf of Mexico (Schwacke et al., 2014; De Guise et al., 2017). These and other human activities (e.g., interaction with fisheries) have resulted in injuries in up to 11.5% of the individuals at SWGM (Morteo et al., 2017) and may play an important role in the onset of skin diseases (Rowe et al., 2010) like LLD, which is a typical cutaneous disease from rural, humid and tropical environments with abundant vegetation and soil; these last two factors are considered potential sources for human infection in localities near to rivers and creeks, including local marine wildlife such as T. truncatus. The latter suggests L. loboi as a hydrophilic fungus (Lupi et al., 2005; Queiroz-Telles et al., 2011) that could be already present within the tropical and humid environment of the ALV lagoon (García, 1973). Potential LLD cases reported in T. truncatus from ALV add to the reports of LLD diagnosed in humans from the Gulf of Mexico (Pech-Ortiz et al., 2020). Therefore, T. truncatus could be considered as local sentinel of LLD disease risk for the rural ALV community, reinforcing the vulnerability and interconnectivity between humans, wildlife, and environmental health in low-income countries such as Mexico, where 40% of infectious diseases (i.e., zoonoses) emerged from animals (Grace et al., 2012). Since exploitation of coastal marine resources is crucial for human livelihood in Veracruz (Sánchez-Gil et al., 2004), and both species have overlapping diets (Chávez-Martínez et al., 2022), LLD presence in the ALV marine ecosystem is a potential risk to public health.
Additionally, long-term multidisciplinary studies in T. truncatus at the SWGM are necessary to define the interindividual and environmental characteristics involved in LLD susceptibility. Molecular diagnosis of the agent linked to LLD and disease surveillance related to climate change and other skin problems (e.g., trauma) at ALV are warranted, while studying the epidemiology and diagnosis of LLD in rural communities from the ALV lagoon.
Finally, we highlight the importance of a regional marine mammal stranding database that helps identifying health and survival threats, and their relation to environmental hazards at the SWGM (Chan et al., 2017) using both traditional veterinary protocols (e.g., necropsy) and near future modern technology (e.g., virtopsy) to improve individual diagnosis (Tsui et al., 2020) and hence to implement appropriate conservation strategies based in health programs for T. truncatus at Veracruz.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
CG - Conceptualization, Methodology, Manuscript Writing and reviewing. MT-O - Data collection, Sample processing, Data analysis, Manuscript Writing. IH-C - Conceptualization, Methodology, Data collection, Data analysis, Manuscript reviewing. CD-A - Methodology, Data collection, Data analysis, Manuscript reviewing. EM - Conceptualization, Funds acquisition, Project manager, Methodology, Data collection, Manuscript writing and reviewing. All authors contributed to the article and approved the submitted version.
The project was financed through the New Fulltime Professor Support Program (PRODEP) by the Public Education Secretariat (EM) and the Academic Group for Management and Conservation of Aquatic Resources (EM and H. Pérez-España), as well as projects No. 45468 (E. Velarde) and 221750 (EM) by the National Council for Science and Technology (CONACyT), and also Acuario de Veracruz, A.C.
This work was part of the undergraduate thesis of MT-O. Field data were obtained through federal permits SGPA/DGVS/00351/06 (EM), SGPA/DGVS/00870/07, 02788/07, 01344/08 and 01649/08 (C. Bazúa), and (EM). We also acknowledge the availability and diligence of Eduardo Gazol at Centro Veterinario Xalapa, for his assistance on histopathology. Finally, we thank I. Martínez, E. Suárez, and M. Páez-Rodríguez for their contributions to the thesis, and M.F. Van Bressem for her comments on the earlier version of this manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Alvízar-Cruz I. (2018). Indicadores de saluid en el delfín naríz de botella (Tursiops truncatus) en las costas de tamiahua-tecolutla, veracruz. tesis de licenciatura tuxpan (Tuxpan, Veracruz, México: Veracruz, México Universidad Veracruzana. Facultad de Biología).
Bejder L., Fletcher D., BräGer S. (1998). A method for testing association patterns of social animals. Anim. Behav. 56 (3), 719–725. doi: 10.1006/anbe.1998.0802
Bessesen B. L., Oviedo L., Hart L. B., Herra-Miranda D., Pacheco-Polanco J. D., Baker L., et al. (2014). Lacaziosis-like disease among bottlenose dolphins Tursiops truncatus photographed in golfo dulce, Costa Rica. Dis. Aquat. Org. 107 (3), 173–180. doi: 10.3354/dao02692
Bessesen B., Oviedo L., Hart L., Herra- Miranda D., Pacheco-Polanco J., Baker L., et al. (2014). Lacaziosis-like disease among bottlenose dolphins Tursiops truncatus photographed in golfo dulce, Costa Rica. Dis. Aquat. Organisms 107, 173–180. doi: 10.3354/dao02692
Bolaños-Jiménez J., Morteo E., Delfín-Alfonso C. A., Fruet P. F., Secchi E. R., Bello-Pineda J. (2021). Population dynamics reveal a core community of the common bottlenose dolphin (Tursiops truncatus) in open waters of the south-Western gulf of Mexico. Front. Mar. Sci. 1610. doi: 10.3389/fmars.2021.753
Bolaños-Jiménez J., Morteo E., Fruet P., Secchi E., Delfín-Alfonso C., et al. (2022). Seasonal population parameters reveal sex-related dynamics of bottlenose dolphins off open waters of the southwestern gulf of Mexico. Mar. Mamm. Sci. 38 (2), 705–724. doi: 10.1111/MMS.12897
Bossart G. D. (2007). Emerging diseases in marine mammals: from dolphins to manatees. Microbe 2, 544–549. doi: 10.1128/microbe.2.544.1
Bossart G. D., Romano T. A., Peden-Adams M. M., Schaefer A. M., Rice C. D., Fair P. A., et al. (2019). Comparative innate and adaptive immune responses in Atlantic bottlenose dolphins (Tursiops truncatus) with viral, bacterial, and fungal infections. Front. Immunol. 10. doi: 10.3389/fimmu.2019.01125
Cairns S. J., Schwager S. J. (1987). A comparison of association indices. Anim. Behav. 35, 1454–1469. doi: 10.1016/S0003-3472(87)80018-0
Chan D. K. P., Tsui H. C. L., Kot B. C. W. (2017). Database documentation of marine mammal stranding and mortality: current status review and future prospects. Dis. Aquat Org. 126, 247–256. doi: 10.3354/dao03179
Chávez-Martínez K., Morteo E., Hernández-Candelario I., Herzka S. Z., Delfín-Alfonso C. A. (2022). Opportunistic gillnet depredation by common bottlenose dolphins in the southwestern gulf of Mexico: Testing the relationship with ecological, trophic, and nutritional characteristics of their prey. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.870012
Daszak P., Cunningham A. A., Hyatt A. D. (2001). Anthropogenic environmental change and the emergence of infectious diseases in wildlife. Acta tropica 78, 103–116. doi: 10.1016/S0001-706X(00)00179-0
Daura-Jorge F. G., Simões-Lopes P. C. (2011). Lobomycosis-like disease in wild bottlenose dolphins Tursiops truncatus of Laguna, southern Brazil: monitoring of a progressive case. Dis. Aquat. Org. 93, 163–170.
De Guise S., Beckmen K. B., Holladay S. D. (2002). “Contaminants and marine mammal immunotoxicology and pathology,” in Toxicology of marine mammals (London: CRC Press), 50–66.
De Guise S., Levin M., Gebhard E., Jasperse L., Hart L. B., Smith C. R., et al. (2017). Changes in immune functions in bottlenose dolphins in the northern gulf of Mexico associated with the deepwater horizon oil spill. Endang. Species. Res. 33, 291–303. doi: 10.3354/esr00814
Desforges J. P. W., Sonne C., Levin M., Siebert U., De Guise S., Deitz R., et al. (2016). Immunotoxic effects of environmental pollutants in marine mammals. Environ. Int. 86, 126–139. doi: 10.1016/j.envint.2015.10.007
Félix F., Van Bressem M. F., Van Waerebeek K. (2019). Role of social behaviour in the epidemiology of lobomycosis-like disease (LLD) in estuarine common bottlenose dolphins from Ecuador. Dis. Aquat. Org. 134, 75–87. doi: 10.3354/dao03356
García E. (1973). Modificaciones al sistema de clasificación de climática de köppen (Universidad Nacional Autónoma de México: Instituto de Geografía), 522.
García-Vital M., Morteo Ortiz E., Martínez-Serrano I., Delgado-Estrella A., Bazúa-Durán M. D. C. (2015). Las asociaciones entre individuos se correlacionan con la diversidad de las conductas en delfines costeros (Tursiops truncatus) del sureste del golfo de méxico. Therya 6, 337–349. doi: 10.12933/therya-15-270
Gnat S., Łagowski D., Nowakiewicz A., Dyląg M. (2021). A global view on fungal infections in humans and animals: infections caused by dimorphic fungi and dermatophytoses. J. Appl. Microbiol. 131, 2688–2704. doi: 10.1111/jam.15084
Grace D., Gilbert J., Randolph T., Kang´ethe E., et al. (2012). The multiple burdens of zoonotic disease and an ecohealth approach to their assessment. Trop. Anim. Health Prod 44, 67–73. doi: 10.1007/s11250-012-0209-y
Guentzel J. L., Portilla E., Keith K. M., Keith E. O. (2007). Mercury transport and bioaccumulation in riverbank communities of the alvarado lagoon system, veracruz state, Mexico. Sci. Total Environ. 388, 316–324. doi: 10.1016/j.scitotenv.2007.07.060
Haubold E. M., Cooper C. R., Wen J. W., McGinnis M. R., Cowan D. F. (2000). Comparative morphology of Lacazia loboi (syn. loboa loboi) in dolphins and humans. Med. Mycol. 38, 9–14. doi: 10.1080/mmy.38.1.9.14
Heckel G. (1992). Fotoidentificación de tursiones (Tursiops truncatus) (Montagu, 1821) en la boca de corazones de la Laguna de Tamiahua, Veracruz, México (Cetacea: Delphinidae). B.Sc. dissertation, (Mexico: Universidad Nacional Autónoma de Mexico)
Jones K. E., Patel N. G., Levy M. A., Storeygard A., Balk D., Gittleman J. L., et al. (2008). Global trends in emerging infectious diseases. Nature 451, 990–993. doi: 10.1038/nature06536
Kiszka J., Van Bressem M. F., Pusineri C. (2009). Lobomycosis-like disease and other skin conditions in indo-pacific bottlenose dolphins tursiops aduncus from the Indian ocean. Dis. Aquat Org. 84, 151–157. doi: 10.3354/dao02037
Learmonth J. A., Colin D., MacLeod M., Begoña S., Pierce G. J., Crick H. Q. P., et al. (2006). “Potential effects of climate change on marine mammals,” in Oceanography and marine biology: An Annual Review, vol. .p. (CRC Press), 431–464. doi: 10.1016/j.envint.2009.10.008
Lupi O., Tyring S. K., McGinnis M. R. (2005). Tropical dermatology: fungal tropical diseases. J. Am. Aca. Derma. 53, 931–951. doi: 10.1016/j.jaad.2004.10.883
Martínez-Serrano I., Serrano A., Heckel G., Schramm Y. (2011). Distribution and home range of bottlenose dolphins (Tursiops truncatus) off veracruz, Mexico. Cienc. Marinas 37, 379–392. doi: 10.7773/cm.v37i4A.1860
Migaki G., Valerio M. G., Irvine B., Garner F. M. (1971). Lobo’s disease in an Atlantic bottle-nosed dolphin. J. Am. Vet. Med. Assoc. 159, 578–582.
Morales-Rincon N., Morteo E., Delfín-Alfonso C. A. (2019). Influence of artisanal fisheries on the behaviour and social structure of Tursiops truncatus in the south-western gulf of Mexico. J. Mar. Biol. Assoc. U. K. 99, 1841–1849. doi: 10.1017/S002531541900078X
Morán-Silva Á., Martinez Franco L. A., Chávez-López R., Franco-López J., Bedia-Sánchez C. M., Contreras Espinosa F., et al. (2005). Seasonal and spatial patterns in salinity, nutrients, and chlorophyll α in the alvarado lagoonal system, veracruz, Mexico. Gulf Caribbean Res. 17, 133–143. doi: 10.18785/gcr.1701.14
Moreno I. B., Ott P. H., Tavares M., Oliveira L. R., Borba M. R., Driemeier D., et al. (2008). “Mycotic dermatitis in common bottlenose dolphins (Tursiops truncatus) from southern Brazil, with a confirmed record of lobomycosis disease,” in Paper SC/60/DW1 presented to the IWC Scientific Committee, Santiago, Chile, IWC, Cambridge, United Kingdom, 30 May– 27 June 2008.
Morteo E., Ramírez y Romero T., Del Castillo-Olvera V., Delfín-Alfonso C., Bello-Pineda J., González-Christen A. (2019). “Ecología poblacional de delfines (Tursiops truncatus) en el corredor arrecifal del suroeste del golfo de méxico. pages 191-214,”. Eds. Granados-Barba A., Ortiz-Lozano L. D., González-Gándara C., y Salas-Monreal D. (Universidad Autónoma de Campeche: Estudios Científicos en el Corredor Arrecifal del Suroeste del Golfo de México), 376. doi: 10.26359/epomex.0319
Morteo E., Rocha-Olivares A., Abarca-Arenas L. G. (2014). Sexual segregation in coastal bottlenose dolphins (Tursiops truncatus) in the south-western gulf of Mexico. Aquat. Mammals 40, 375–385. doi: 10.1578/AM.40.4
Morteo E., Rocha-Olivares A., Abarca-Arenas L. G. (2017). Abundance, residency and potential hazards for coastal bottlenose dolphins (Tursiops truncatus) off a productive lagoon in the gulf of Mexico. Aquat. Mammals 43, 308–319. doi: 10.1578/AM.43.3.2017.308
Murdoch M. E., Mazzoil M., McCulloch S., Bechdel S., O’Corry-Crowe G., Bossart G. D., et al. (2010). Lacaziosis in bottlenose dolphins Tursiops truncatus along the coastal Atlantic ocean, Florida, USA. Dis. Aquat. Org. 92 (1), 69–73. doi: 10.3354/dao02282
Murdoch M. E., Reif J. S., Mazzoil M., McCulloch S. D., Fair P. A., Bossart G. D. (2008). Lobomycosis in bottlenose dolphins (Tursiops truncatus) from the Indian river lagoon, Florida: estimation of prevalence, temporal trends, and spatial distribution. EcoHealth 5 (3), 289–297. doi: 10.1007/s10393-008-0187-8
Paniz-Mondolfi A. E., Reyes-Jaimes O., Dávila-Jones L. (2007). Lobomycosis in Venezuela. Int. J. Derma. 46 (2), 180–185. doi: 10.1111/j.1365-4632.2007.02937.x
Pech-Ortiz L., Maya-Aranda S., Hernández-Castro R., Xicohtencatl-Cortes J., Tirado-Sánchez A., Bonifaz A.. (2020). Lacaziosis (Lobomycosis) from southern Mexico: A case confirmed by molecular biology. Mycopathologia 185, 737–739. doi: 10.1007/s11046-020-00464-7
Queiroz-Telles F., Nucci M., Colombo A. L., Tobón A., Restrepo A. (2011). Mycoses of implantation in Latin America: An overview of epidemiology, clinical manifestations, diagnosis and treatment. Med. mycology 49 (3), 225–236. doi: 10.3109/13693786.2010.539631
Ramos E., Castelblanco-Martínez N., Garcia J., Rojas-Arias J., Foley J., Audley K., et al. (2018). Lobomycosis-like disease in bottlenose dolphins in Belize and Mexico: Bridging the gap between the americas. Dis. Aquat. Organisms 128, 1–12. doi: 10.3354/dao03206
Rechimont M. E., Lara-Domínguez A. L., Morteo E., Martínez-Serrano I., Equihua M. (2018). Depredation by coastal bottlenose dolphins (Tursiops truncatus) in the southwestern gulf of Mexico in relation to fishing techniques. Aqua. Mammals 44 (5), 469. doi: 10.1578/AM.44.5.2018.469
Reif J. S., Mazzoil M. S., McCulloch S. D., Varela R. A., Goldstein J. D., Fair P. A., et al. (2006). Lobomycosis in Atlantic bottlenose dolphins from the Indian river lagoon, Florida. J. Am. Vet. Med. Assoc. 228 (1), 104–108. doi: 10.2460/javma.228.1.104
Reif J. S., Peden-Adams M. M., Romano T. A., Rice C. D., Fair P. A., Bossart G. D. (2009). Immune dysfunction in Atlantic bottlenose dolphins (Tursiops truncatus) with lobomycosis. Med. Mycology 47 (2), 125–135. doi: 10.1080/13693780802178493
Reif J. S., Schaefer A. M., Bossart G. D. (2013). Lobomycosis: risk of zoonotic transmission from dolphins to humans. Vector-Borne Zoonotic Dis. 13 (10), 689–693. doi: 10.1089/vbz.2012.1280
Rotstein D. S., Burdett L. G., McLellan W., Schwacke L., Rowles T., Terio K. A., et al. (2009). Lobomycosis in offshore bottlenose dolphins (Tursiops truncatus), north Carolina. Emerg. Infect. Dis. 15 (4), 588–590. doi: 10.3201/eid1504.081358
Rowe L., Currey R., Dawson S., Johnson D. (2010). Assessment of epidermal condition and calf size of fiordland bottlenose dolphin Tursiops truncatus populations using dorsal fin photographs and photogrammetry. Endang. Species Res. 11, 83–89. doi: 10.3354/esr00256
Ruíz-Hernández I. A. (2014). “Desplazamientos de toninas (Tursiops truncatus) en la costa central de veracruz, méxico,” in Tesis de licenciatura. facultad de biología(Universidad Veracruzana. Xalapa, Ver., México), 58.
Sánchez-Gil P., Yáñez-Arancibia A., Ramírez-Gordillo J., Day J. W., Templet P. H. (2004). Some socio-economic indicators in the Mexican states of the gulf of Mexico. Ocean Coast. Management. 47 (11-12), 581–596. doi: 10.1016/j.ocecoaman.2004.12.003
Schwacke L. H., Smith C. R., Townsend F. I., Wells R. S., Hart L. B., Balmer B. C., et al. (2014). Health of common bottlenose dolphins (Tursiops truncatus) in barataria bay, Louisiana, following the deepwater horizon oil spill. Environ. Sci. Technol. 48 (1), 93–103. doi: 10.1021/es403610f
Seyedmousavi S., Guillot J., Tolooe A., Verweij P. E., De Hoog G. S. (2015). Neglected fungal zoonoses: hidden threats to man and animals. Clin. Microbiol. Infect. 21 (5), 416–425. doi: 10.1016/j.cmi.2015.02.031
Siciliano S., Van Bressem M.-F., Moreno I., Ott P., Tavares M., André P., et al. (2008). Review of lobomycosis and lobomycosis-like disease (LLD) in cetacea from south America. Workshop cetacean skin diseases Int. Whaling Commission SC/60/DW13.
Tenorio-Osorio M. I. (2015). Lesiones potencialmente provocadas por lacaziosis (Lacazia loboi) en tursiones (Tursiops truncatus) de las aguas costeras de alvarado, veracruz, méxico. tesis de licenciatura xalapa, veracruz (México Universidad Veracruzana: Facultad de Biología).
Tsui H. C. L., Kot B. C. W., Chung T. Y. T., Chan D. K. P. (2020). Virtopsy as a revolutionary tool for cetacean stranding programs: Implementation and management. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.542015
Ueda K., Sano A., Yamate J., Itano Nakagawa E., Kuwamura M., Izawa T., et al. (2013). Two cases of lacaziosis in bottlenose dolphins (Tursiops truncatus) in Japan. Case Rep. Vet. Med. 2013, 318548. doi: 10.1155/2013/318548
Urian K., Gorgone A., Read A., Balmer B., Wells R. S., Berggren P., et al. (2015). Recommendations for photoidentification methods used in capture-recapture models with cetaceans. Mar. Mam Sci. 31, 298–321. doi: 10.1111/mms.12141
Van Bressem M. F., de Oliveira Santos M. C., de Faria Oshima J. E. (2009a). Skin diseases in Guiana dolphins (Sotalia guianensis) from the paranaguá estuary, Brazil: A possible indicator of a compromised marine environment. Mar. Environ. Res. 67 (2), 63–68. doi: 10.1016/j.marenvres.2008.11.002
Van Bressem M. F., Raga J. A., Di Guardo G., Jepson P. D., Duignan P. J., Siebert U., et al. (2009b). Emerging infectious diseases in cetaceans worldwide and the possible role of environmental stressors. Dis. Aquat. Org. 86 (2), 143–157. doi: 10.3354/dao02101
Van Bressem M. F., Reyes J. C., Félix F., Echegaray M., Siciliano S., Di Beneditto A. P., et al. (2007). A preliminary overview of skin and skeletal diseases and traumata in small cetaceans from south American waters. Lat. J. Aquat. Mammals 6 (1), 7–42. doi: 10.5597/lajam00108
Van Bressem M. F., Shirakihara M., Amano M. (2013). Cutaneous nodular disease in a small population of indo-pacific bottlenose dolphins, Tursiops aduncus, from Japan. Mar. Mamm. Sci. 29, 525–532. doi: 10.1111/j.1748-7692.2012.00589.x
Van Bressem M. F., Simões-Lopes P. C., Félix F., Kiszka J. J., Daura-Jorge F. G., Avila I. C., et al. (2015). Epidemiology of lobomycosis-like disease in bottlenose dolphins tursiops spp. from south America and southern Africa. Dis. Aquat. Org. 117 (1), 59–75. doi: 10.3354/dao02932
Van Bressem M. F., Van Waerebeek K., Duignan P. J. (2018). Epidemiology of tattoo skin disease in captive common bottlenose dolphins (Tursiops truncatus): are males more vulnerable than females? J. Appl. Anim. Welfare Sci. 21 (4), 305–315. doi: 10.1080/10888705.2017.1421076
Van Bressem M. F., Van Waerebeek K., Flach L., Reyes J. C., de Oliveira Santos M. C., Siciliano S., et al. (2008). “Skin diseases in cetaceans,” in Scientific Committee document SC/60/DW8, International Whaling Commission, Santiago, Chile, June 2008.
Vázquez-Botello A., Villanueva S., Rosales L. (2004). “Distribución de metales pesados en el golfo de méxico,” in Diagnóstico ambiental del golfo de méxico. secretaría de medio ambiente y recursos naturales, instituto nacional de ecología, harte research institute for gulf of Mexico studies. Eds. Caso M., Pisanty I., Ezcurra E., 681–706.
Vázquez-Botello A., de la Lanza Espino G., Fragoso S. V., Velez G. P. (2019). Pollution issues in coastal lagoons in the gulf of mexico. lagoon environments around the world - A scientific perspective. IntechOpen. doi: 10.5772/intechopen.86537
Vázquez-Botello A., Villanueva F. S., Rivera R. F., Velandia A. L., de la Lanza G. E. (2018). Analysis and tendencies of metals and POPs in a sediment core from the alvarado lagoon system (ALS), veracruz, Mexico. Arch. Environ. Contamination Toxicol. 75, 157–173. doi: 10.1007/s00244-018-0516-z
Venn-Watson S., Jensen E. D., Schork N. J. (2020). A 25-y longitudinal dolphin cohort supports that long-lived individuals in same environment exhibit variation in aging rates. Proc. Natl. Acad. Sci. 117 (34), 20950–20958. doi: 10.1073/pnas.1918755117
Venn-Watson S., Smith C. R., Gomez F., Jensen E. D.. (2011). Physiology of aging among healthy, older bottlenose dolphins (Tursiops truncatus): comparisons with aging humans. J. Comp. Physiol. B 181, 667–680. doi: 10.1007/s00360-011-0549-3
Vilela R., Bossart G. D., St. Leger J. A., Dalton L. M., Reif J. S., Schaefer A. M., et al. (2016). Cutaneous granulomas in dolphins caused by novel uncultivated paracoccidioides brasiliensis. Emerg. Infect. Dis. 22 (12), 2063–2069. doi: 10.3201/eid2212.160860
Wagner B., Gardner I., Cameron A., Doherr M. G. (2003). “Statistical analysis of data from surveys, monitoring, and surveillance systems,” in Animal disease surveillance and survey systems: methods and applications. Ed. Salman M. D. (Ames, Iowa: Iowa State Press), 67–86.
Keywords: zoonosis, infectious diseases, marine mammals, wildlife, mycosis, cetaceans, public health
Citation: Gálvez C, Tenorio-Osorio M, Hernández-Candelario I, Delfín-Alfonso CA and Morteo E (2022) Lobomycosis-like disease epidemiology, pathology and social affiliations in bottlenose dolphins from Southwestern Gulf of Mexico. Front. Mar. Sci. 9:1018118. doi: 10.3389/fmars.2022.1018118
Received: 12 August 2022; Accepted: 10 October 2022;
Published: 02 November 2022.
Edited by:
Stephanie Norman, Marine-Med: Marine Research, Epidemiology and Veterinary Medicine, United StatesReviewed by:
Brian Kot, City University of Hong Kong, Hong Kong SAR, ChinaCopyright © 2022 Gálvez, Tenorio-Osorio, Hernández-Candelario, Delfín-Alfonso and Morteo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eduardo Morteo, ZWR1YXJkby5tb3J0ZW9AZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.