 Thais Martins
Thais Martins Paula Santana
Paula Santana Ítalo Lutz
Ítalo Lutz Raimundo da Silva
Raimundo da Silva Aurycéia Guimarães-Costa
Aurycéia Guimarães-Costa Marcelo Vallinoto
Marcelo Vallinoto Iracilda Sampaio
Iracilda Sampaio Grazielle Evangelista-Gomes*
Grazielle Evangelista-Gomes*
- Instituto de Estudos Costeiros, Universidade Federal do Pará, Bragança, Brazil
Elasmobranchs represent a well-defined group, composed of about 1,150 species inhabiting diverse aquatic environments. Currently, several of these species have been classified as threaten due to overexploitation. Therefore, we used DNA barcode to identify traded species of sharks and stingrays in the municipality of Bragança (Amazon coastal region), a major fishery landing site in northern Brazil. We collected a total of 127 samples labeled into 24 commercial nomenclatures over 1-year period. Twenty species were discriminated and 13 of them are recognized under some threatening status. In relation to sharks, Carcharhinidae showed the highest number of species, half of them classified as endangered, followed by hammerhead sharks (Sphyrnidae), with four species also regarded as threatened with extinction. The Rhinopteridae and Dasyatidae rays were the most abundant groups, with trade records of the following threatened species: Rhinoptera brasiliensis, Rhinoptera bonasus, Hypanus berthalutzae, and Fontitrygon geijskesi. It is noteworthy that threatened species of elasmobranchs have been frequently and regularly exploited because of inefficient fishery management policies. Therefore, effective inspection practices need to be incorporated in fisheries, including the use of DNA barcode to enable a reliable method of species authentication and to assure the proper commercialization.
Introduction
Sharks and rays (Subclass Euselachii) are elasmobranchs whose origin dates back to the Devonian (∼400 million years ago) (Compagno, 1990). Currently, this group is represented by nearly 1,150 valid species, found mainly in marine habitats even though some of them are typical of freshwater environments (Nelson et al., 2016).
In spite of their wide distribution, sharks and rays have been severely threatened with extinction, mainly because of overfishing (Stevens et al., 2000; Pacoureau et al., 2021). The fishing pressure over this fish group has determined remarkable reductions of population size particularly because elasmobranchs present peculiar biological traits, such as longevity, late sex maturation and low fecundity rates (Stevens et al., 2000; Oliver et al., 2015), making this group even more vulnerable to overfishing. Global data provided by Food and Agriculture Organization of United Nations pointed out that nearly 30% of elasmobranchs are vulnerable, endangered or critically threatened with extinction in nature (IUCN, 2021). The population drops of sharks and rays by overfishing are directly associated with the commercialization of fins in Asian markets (Pough et al., 2008), bycatch and artisanal fishing (Robbins et al., 2006; Graham et al., 2010; Ferrette et al., 2019).
Both artisanal and industrial fishing practices to supply the demand of elasmobranch-derived products in regional markets take place regularly in several points along the northern coast of Brazil, where nearly 69 species of shark and rays are recognized, many of them classified as threatened (Feitosa et al., 2018; Marceniuk et al., 2019; Rodrigues-Filho et al., 2020). As a matter of fact, many species of threatened sharks and rays are commercialized in coastal Amazon region (Feitosa et al., 2018). In the municipality of Bragança, northeastern Pará, these species are locally sold under the denomination of “cação” (sharks) and “arraias” (rays) in street markets, representing an important source of income (Freire et al., 2011; Martins et al., 2021).
During commercialization, the morphological identification of sharks and rays is usually jeopardized because body parts, such as fins, are removed right after catches in open sea (Rodrigues-Filho et al., 2009; Feitosa et al., 2018). In addition, the Brazilian laws lack a standardized nomenclature for the commercialization of these species, thus favoring illegal trade since the commercial common names often refer to a group of species (Ardura et al., 2010; Feitosa et al., 2018; Martins et al., 2021).
Inasmuch as useful meristic and morphometric traits are unavailable in the commercialized sharks and rays, alternative approaches should be implemented to assure the unambiguous identification of species. Accordingly, DNA barcode based on the sequencing of a portion of the mitochondrial gene Cytochrome C oxidase—subunit I (COI) has been widely used to discriminate and identify species of several groups, including fishes and their products that could not be properly validated by morphological traits (Ward et al., 2005; Steinke et al., 2017; Feitosa et al., 2018; Gomes et al., 2019; Carvalho et al., 2020).
Bragança represents a major fishing market in northeastern Pará in Amazon coastal region, northern Brazil (Freire et al., 2011; Santana et al., 2020; Martins et al., 2021). Fish landing occur daily in this municipality (Isaac et al., 2008) and most of products are sold in the street market of Bragança that encompasses indoor (“Mercado”) and outdoor (“Feirinha”) sites where a high diversity and number of species are commercialized (Freire et al., 2011; Martins et al., 2021). Over more than a decade, studies in this region have shown that Bragança lands and commercializes a large diversity of elasmobranchs with variation in the abundance and richness of sharks and rays species sold in the street market (Carmona et al., 2008; Rodrigues-Filho et al., 2009, 2020; Guimarães-Costa et al., 2020; Feitosa et al., 2018; Marceniuk et al., 2019; Souza-Araujo et al., 2021). The elasmobranchs labeled as “cação” and “arraia” are among the most commercialized fish groups in this region but the lack of standard commercial common names and the mischaracterization of specimens constrain their morphological identification and justify the utilization of other methods based on molecular validation of species, like DNA barcode (Feitosa et al., 2018; Rodrigues-Filho et al., 2020; Martins et al., 2021).
Therefore, in the present study, the dynamics of commercialization of sharks and rays was investigated in northeastern Pará, in order to identify the commercialized species based on DNA barcode and potential records of threatened species in local markets.
Materials and Methods
Sampling
The collections were carried out by incursions in 15-day intervals to the street market of Bragança, state of Pará, Brazil over a year (February 2017 to January 2018) to cover the species that are traded throughout time space. The elasmobranchs traded in the Bragança street market are caught, mainly, as bycatch, in fisheries of acoupa weakfish Cynoscion acoupa (Lacèpede, 1801), pink-shrimp Penaeus sp., piramutaba catfish Brachyplatystoma vaillantii (Valenciennes, 1840) and several fish (Mourão et al., 2018; Marceniuk et al., 2019). These fisheries are conducted on a small and large scale by different fishing systems operating in different areas of the northern Brazilian coast (Amapá, Pará, and Maranhão), using a wide variety of fishing gear (Braga et al., 2006; Espírito Santo and Isaac, 2012).
The collection schedule was defined based on a previous report in the same location (Martins et al., 2021). A total of 127 samples of elasmobranchs were collected, 35 labeled as “ray” and 92 under the label “shark” (Supplementary Material 1). The samples were acquired in indoor and outdoor market (“feirinha”) with available supply of rays and sharks on collection expeditions, avoiding duplication of samples from a single individual, especially in the case of large sharks that are usually sold by more than one fishmonger.
Specimens were previously identified using the commercial nomenclatures used by traders at the time of collection (Supplementary Material 1) to verify if there was a relationship between genetic identification and commercial nomenclatures for sharks and rays (Supplementary Material 1).
A large portion of the biological tissue samples collected were from uncharacterized specimens (sharks: headless or finless samples; rays: headless samples, filets, or only the pectoral fins) (Supplementary Figure 1). Only a small portion of samples encompassed whole individuals.
The whole specimens of rays (Hypanus guttatus, n = 2; Gymnura micrura, n = 3) and shark (Rhizoprionodon porosus, n = 1) were identified by specialized literature (Compagno, 1984; Mc Eachran and Carvalho, 2002) and used as vouchers to compose a reference sample database to assist in the identification process from public sequence banks (Genbank and BOLD Systems). These specimens were deposited in the ichthyological collection of the Laboratório de Genética Aplicada (LAGA), Instituto de Estudos Costeiros (IECOS), UFPA-Bragança.
Laboratory Procedures
Total DNA was extracted using the Wizard Genomic DNA Purification kit (Promega Corporation, Madison, WI, United States), according to the manufacturer’s instructions. To amplify the barcode portion of the COI gene we used the primers FishF1 (5′ TCAACCAACCACAAAGACATTGGCAC 3′) and FishR1 (5′ TAGACTTCTGGGTGGCCAAAGAATCA 3′) (Ward et al., 2005).
Amplification was performed via polymerase chain reaction (PCR) including 2.4 μL of dNTP (1.25 mM), 1.5 μL of buffer (10×), 0.6 μL of MgCl2 (25 mM), 0.4 μL of each primer (5 pmol/μL), 0.1 μL of Taq Polymerase (5 U/μL), 0.6–1 μL of template DNA and ultrapure water to a final volume of 15 μL. The amplification conditions were: initial denaturation at 95°C for 3 min, 35 cycles of denaturation at 95°C for 30 sec, hybridization at 56°C for 45 sec, extension at 72°C for 1 min and a final extension at 72°C for 3 min.
Positive PCRs were purified with PEG (Polyethylene Glycol) according to Paithankar and Prasad (1991). The sequencing was performed by the dideoxy terminal method (Sanger et al., 1977) with the Big Dye kit (ABI Prism Dye Terminator Cycle Sequencing Reading Reaction–Thermo Fisher Scientific) according to the manufacturer’s instructions, using FishF1 primer. After precipitation, the purified PCR products were submitted to capillary electrophoresis in ABI 3500 Genetic Analyzer automatic sequencer (Thermo Fisher Scientific).
Database and Analysis
The sequences were edited in the software BioEdit v. 7.1.3.0 (Hall, 1999), by visual inspection of chromatogram files in ABI format, using a threshold of ≥30 for PHRED quality score of base calling for trimming. DNA sequences were aligned using CLUSTAL W (Thompson et al., 1994; Larkin et al., 2007), available in the software BioEdit v. 7.1.3.0 (Hall, 1999). Presence of stop codons was evaluated in the BOLD Systems platform (Ratnasingham and Hebert, 2007)1.
To help identify the taxa, the haplotypes were defined in the software DNAsp (Librado and Rozas, 2009). This identification was performed based on the similarity of our sequences with those available in GenBank (NCBI--National Center for Biotechnology Information)2 and BOLD (Barcode of Life Data Systems--reference libraries),3 using the highest similarity values (99–100%) (Supplementary Material 1) to discriminate species (Hebert et al., 2003). Nineteen reference sequences from GenBank (Supplementary Material 2) were used. The maximum and average barcode gap distances were evaluated in the BOLD Systems platform using the Barcode Gap Analysis tool (Ratnasingham and Hebert, 2019).
Furthermore, phylogenetic analyses based on the tree topology using Bayesian inference were carried out to verify the species validation within clades. The evolutionary model was defined according to the jModelTest 2 (Darriba et al., 2012), available in CIPRES Science Gateway v3.3 (Miller et al., 2010), based on the Bayesian Information Criterion (BIC).
Bayesian inference (BI) was performed in the software BEAST v. 1.8.4 (Drummond et al., 2012) assuming the following parameters: strict clock, Yule speciation process, HKY + I + G evolutionary model as selected by jModelTest2, 50 million generations and 10% of burn-in. The log files were checked in Tracer v. 1.7 (Rambaut et al., 2018) to assess the burn-in length and the convergence based on ESS (Effective Sample Size) parameters above 200. The generated trees were summarized in TreeAnnotator v. 1.8.4 to obtain the consensus tree, which was visualized in FigTree v. 1.4.4 (Rambaut, 2018). The p distance was calculated in MEGA X (Kumar et al., 2018) to compare intra and interspecific distances and to assess the genetic divergence of closely related species (Kumar et al., 2018).
After the genetic identification of traded species, the conservation status was verified in the IUCN Red List of Threatened Species (IUCN, 2021) and Brazil Red Book of Threatened Species of Fauna (ICMBio, 2018a). The graph of threatened categories and the graph relating the traded categories and the identified species were built using the online tool RAWGraphs4.
Results
Molecular Identification and Genetic Similarity
A dataset of 127 samples (92 of sharks and 35 of rays) comprising COI fragments of 599 base pairs were obtained, referring to 24 commercial nomenclatures and totaling 31 haplotypes (Supplementary Material 1). All generated sequences were stored in GenBank under accession numbers MZ303812–MZ303842.
A total of 20 species was discriminated by DNA barcode. Nineteen of them were identified based on their genetic similarity with sequences from public datasets, most of them exhibiting values equal to 100% (Table 1).
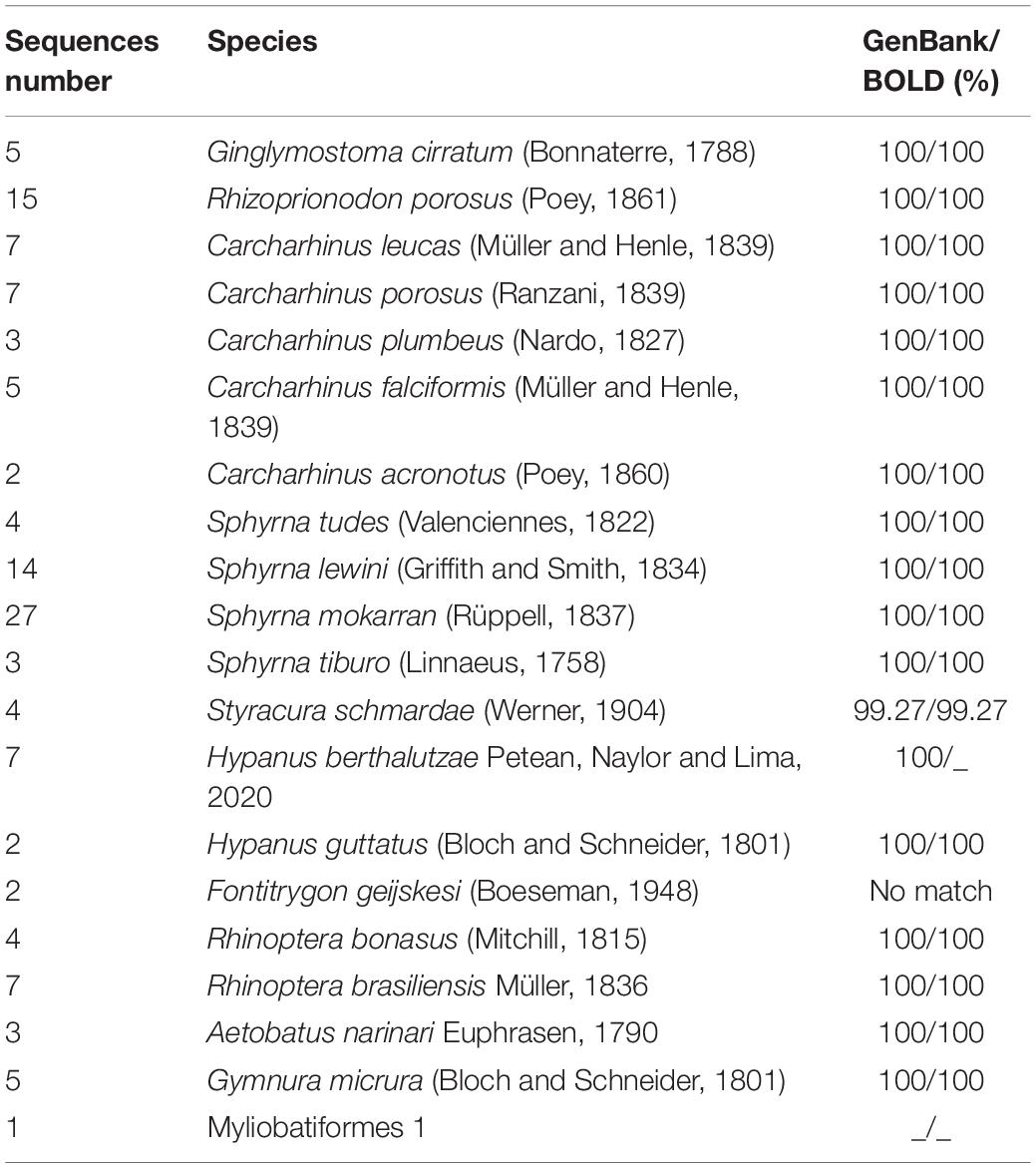
Table 1. Genetic similarity of the 127 samples of elasmobranchs collected from the Amazon coast. The samples that showed incompatibility in the identification are indicated by the symbol “_”.
Identification Based on Phylogenetic Inference
The Bayesian tree topology showed 20 clades, being 9 of them related to rays (order Myliobatiformes) and 11 shark taxa from orders Orectolobiformes and Carcharhiniformes. The representatives of Myliobatiformes encompassed two species of Rhinopteridae, three species of Dasyatidae, one species of Potamotrygonidae, one species of Gymnuridae and one species of Aetobatidae, as well as specimen identified as Myliobatiformes 1. The order Orectolobiformes was represented only by species of family Ginglymostomatidae (one species) while the Carcharhiniformes were represented by six species of Carcharhinidae and four species of Sphyrnidae.
In the case of rays, the most representative families were: Rhinopteridae (N = 11) with two species–Rhinoptera brasiliensis (N = 7) and Rhinoptera bonasus (N = 4)–and Dasyatidae (N = 11) including the species Hypanus berthalutzae (N = 7), Hypanus guttatus (N = 2) and Fontitrygon geijskesi (N = 2). The families Potamotrygonidae, Gymnuridae and Aetobatidae encompassed the species Styracura schmardae (N = 4), Gymnura micrura (N = 5) and Aetobatus narinari (N = 3), respectively (Figure 1A).
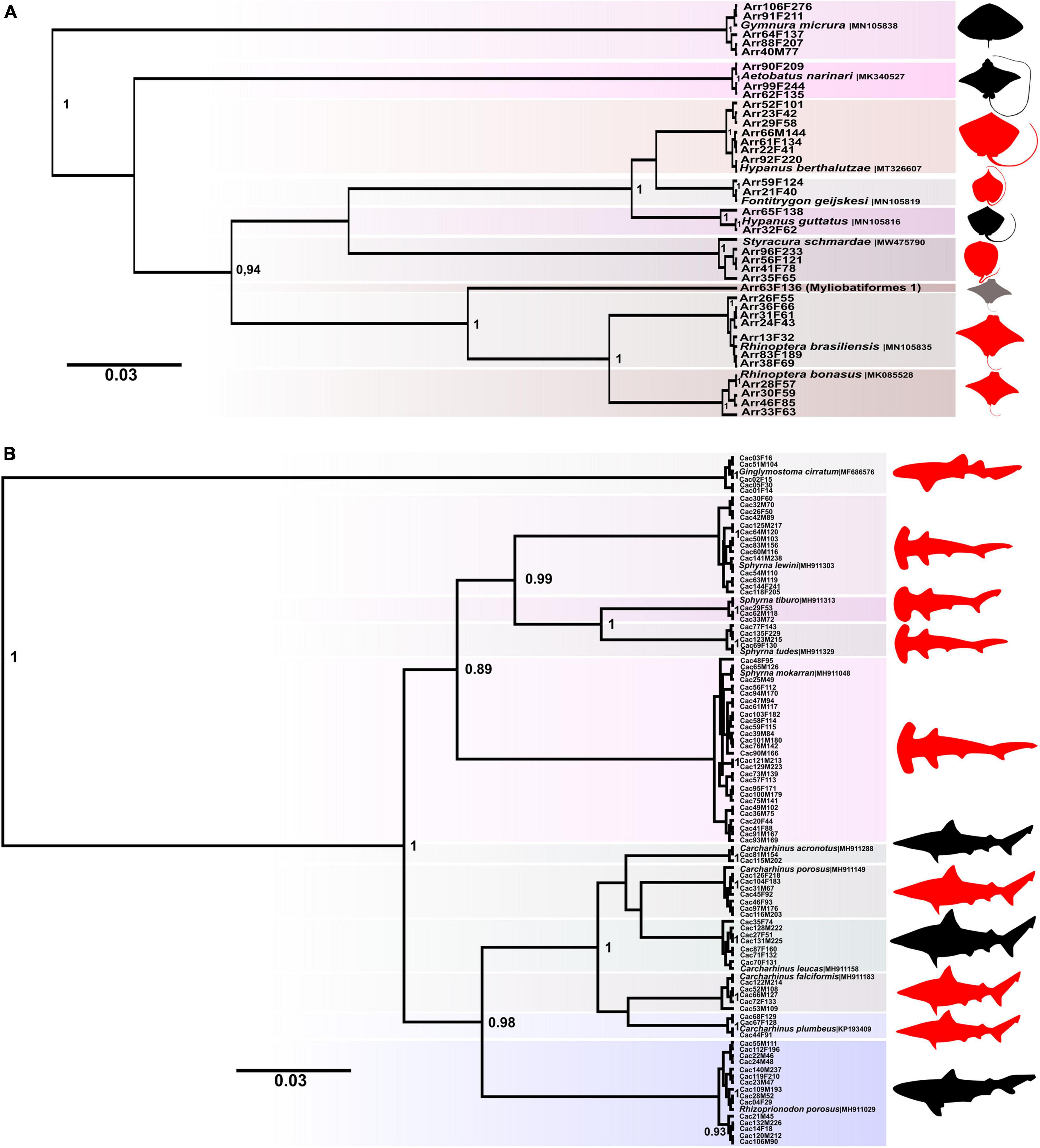
Figure 1. Bayesian inference of species tree performed in BEAST showing the diversity of commercialized Elasmobranchii at the Street Market of Bragança-PA. (A) Endangered and Myliobatiformes 1 species are indicated in red and gray, respectively. (B) The red color indicates the endangered species.
The family Sphyrnidae comprised the largest number of samples (N = 48) distributed into four species: Sphyrna mokarran (N = 27), Sphyrna lewini (N = 14), Sphyrna tudes (N = 4) and Sphyrna tiburo (N = 3). The family Carcharhinidae encompassed 39 samples and six species: Rhizoprionodon porosus (N = 15), Carcharhinus porosus (N = 7), Carcharhinus leucas (N = 7), Carcharhinus falciformis (N = 5), Carcharhinus plumbeus (N = 3), Carcharhinus acronotus (N = 2) while the family Ginglymostomatidae was represented by Ginglymostoma cirratum (N = 5) (Figure 1B).
The specimen designated here as Myliobatiformes 1 was not suitable for molecular identification at the species level because it showed genetic similarity with sequences from public banks of up to 88% with species of the families Rhinopteridae and Mobulidae. The IB tree showed the grouping of this taxon with the species Mobula birostris (Walbaum, 1792) and Mobula hypostoma (Bancroft, 1831) and R. brasiliensis, R. bonasus (Supplementary Material 2). However, the distance p recorded for Myliobatiformes 1 to M. birostris and M. hypostoma was 13.11 and 12.57%, respectively, and from Myliobatiformes 1 to R. brasiliensis and R. bonasus was 13, 98%, and 12.62%, respectively.
Commercial Labels and the Supply of Threatened Species
Fifteen commercial nomenclatures were recorded for sharks, representing 11 species, while nine commercial labels were recorded for rays (Figure 2) with discrimination of nine taxa (eight identified at species level and one of them at the order level). Thirteen out of the 20 listed species are considered endangered and five of them are regarded as nearly threatened (ICMBio, 2018a; IUCN, 2021). Amongst the identified species, five are classified as data deficient (DD), one as not evaluated (NE), one as endangered (EN), one as vulnerable (VU) according to Brazilian lists (ICMBio, 2018a), while according to international red lists, the samples comprised one least concern (LC), five near threatened (NT), and five critically endangered (CR) species (IUCN, 2021; Figure 3).
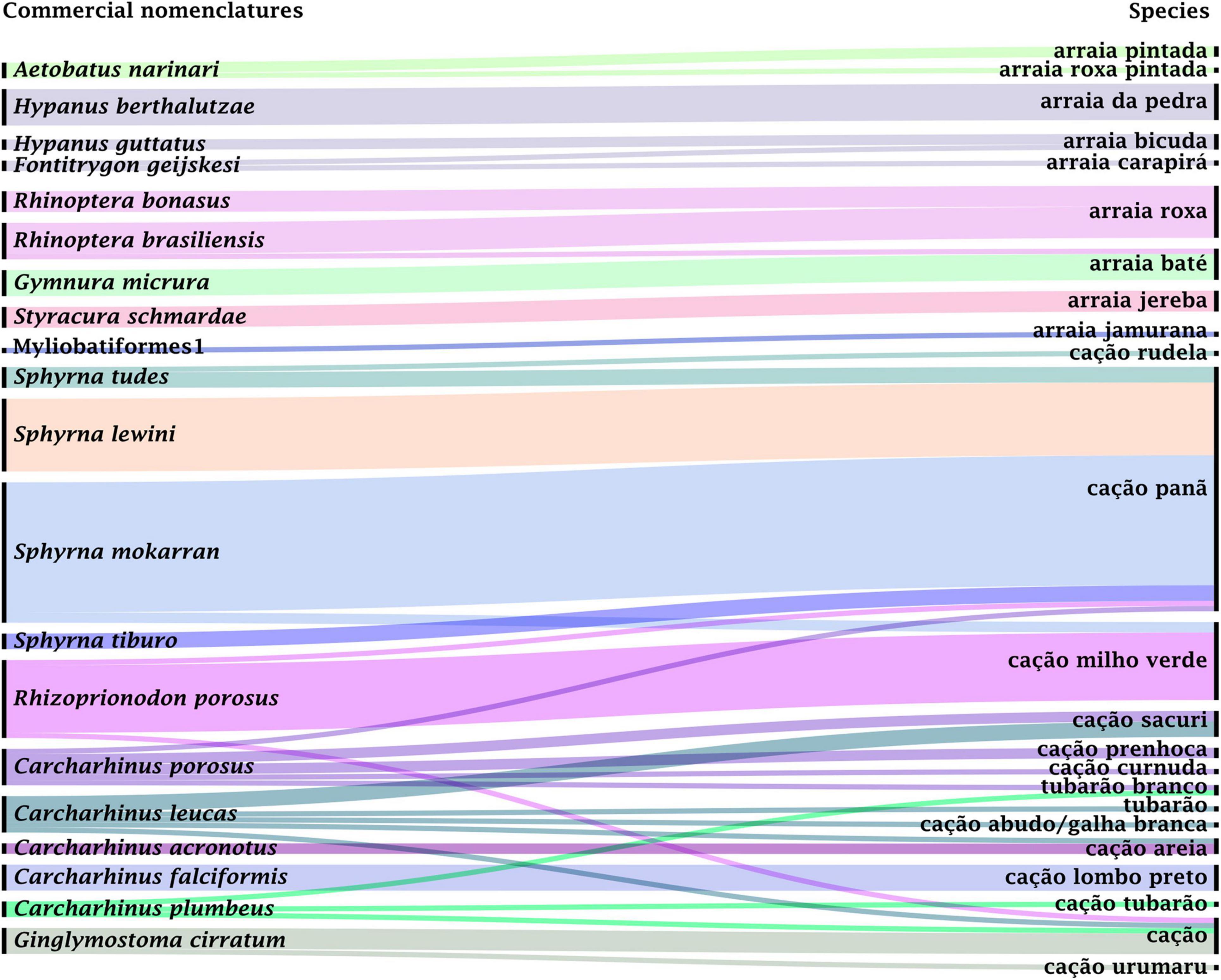
Figure 2. Graph relating the categories or names of sharks and rays and their molecular identification. The colored lines connect the commercial popular names to each species and the line thickness refers to the number of samples collected under each denomination. The graph was generated using RAWGraphs (https://rawgraphs.io/).
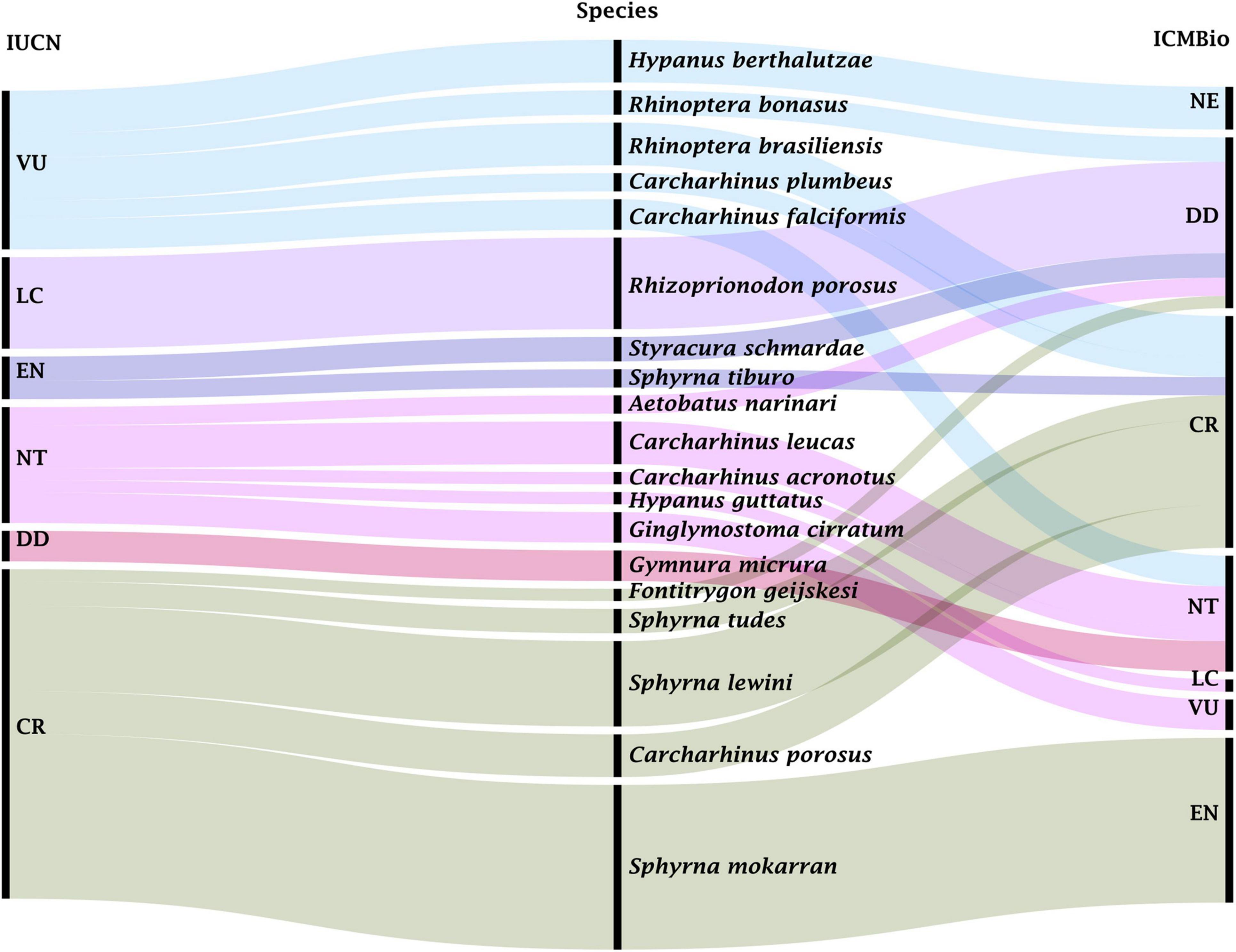
Figure 3. Alluvial diagram relating the status of analyzed species (NE, not evaluated; DD, data deficient; LC, least concern; NT, near threatened; VU, vulnerable; EN, endangered; CR, critically endangered) according to the IUCN Red List of Threatened Species, on the left, with the Brazil Red Book of Threatened Species of Fauna (ICMBio), on the right. The line thickness indicates the frequency of the species sampled. The diagram was generated using RAWGraphs (https://rawgraphs.io/).
Distinct commercial labels often referred to the same species, such as “arraia pintada” and “arraia roxa-pintada” stingrays, which are nomenclatures used for A. narinari, as well as “cação prenhoca” and “cação-curnuda” that refer to C. porosus and “cação abudo” and “cação galha-branca” used for the commercialization of C. leucas (Figure 2). On the other hand, the label “cação panã” encompassed six species with different threatening status: R. porosus, C. porosus, S. tiburo, S. tudes, S. mokarran, and S. lewini (Figure 3).
The collected of sharks and rays over a 1-year period revealed regular supplies for some species, such as S. mokarran and S. lewini, sampled nearly every month. Other species were collected occasionally, including R. brasiliensis, C. plumbeus, C. porosus, S. tudes, S. tiburo, and C. falciformis.
Discussion
Diversity of Elasmobranchs in the Amazon Coast
A total of 20 taxa of elasmobranchs were discriminated via DNA barcode among the samples collected over a year in the street market of Bragança. Nineteen of them could be identified at species-level revealing the high efficiency of this method to assess the diversity of commercialized sharks and rays. Previous molecular reports based on molecular markers carried out in northern Brazilian coast identified similar numbers of elasmobranch species in fish landings and local markets (N = 26), comprising 17 taxa of sharks and nine species of rays (Feitosa et al., 2018; Rodrigues-Filho et al., 2020).
Ten out of the 17 shark species reported in northern Brazilian coast (Feitosa et al., 2018) were found in the street Market of Bragança. In addition, we recorded for the first time the presence of Carcharhinus plumbeus in local markets from this region (Rodrigues-Filho et al., 2009; Feitosa et al., 2018).
A total of 11 species of sharks are commercialized in the street market of Bragança, as similarly reported by Rodrigues-Filho et al. (2009) using 12S and 16S mitochondrial DNA markers. Nonetheless, the species composition between both reports differed inasmuch as we identified some species that have never been reported before such as C. plumbeus, G. cirratum, S. lewini, and R. porosus. On the other hand, Carcharhinus perezi and Galeocerdo cuvier (Rodrigues-Filho et al., 2009) were not identified in the present study.
As for rays, nine species have been commercialized in Bragança, including two unrecorded species in previous studies (Carmona et al., 2008): H. berthalutzae and R. brasilenses. However, in both reports some taxa of rays could not be precisely identified. A potential obstacle to a proper identification of the species was the lack of whole specimens to be used as a voucher for morphological identification as well as the absence of sequences in public datasets for comparative analyses (Rodrigues-Filho et al., 2020).
In recent years, studies based on molecular identification of stingray species have allowed both the identification of valid species and the registration of new occurrences and new species (Guimarães-Costa et al., 2020; Pentean et al., 2020). In a recent study, Rodrigues-Filho et al. (2020) identified nine ray species on the Amazon coast using DNA barcodes. However, the unidentified stingray taxon (Myliobatiformes 1) discovered here has no genetic match with any of the species presented by Rodrigues-Filho et al. (2020), nor with any other species previously reported in this region with COI sequences in public banks.
Even though 69 species of elasmobranchs are recognized for the Amazon coast (Marceniuk et al., 2019), the diversity of sharks and rays from this region remains overlooked. As a matter of fact, as long as the number of studies in this region has increased, new discoveries have been reported. Recently, morphological analyses reported the following species for the first time in Amazon coast: Narcine bancroftii, Pseudobatos horkelii, Urobatis jamaicensis, and Narcine sp. (Marceniuk et al., 2019). Similarly, molecular data identified Narcine brasiliensis and Mustelus canis (Guimarães-Costa et al., 2020; Rodrigues-Filho et al., 2020) while integrative analyses based on morphological, genetic and ecological studies recorded H. berthalutzae (Pentean et al., 2020) for this region. These data highlight the importance of focusing on the diversity of elasmobranchs in the north coast of Brazil, as recent studies have identified distinct results on the species composition in this group of fishes.
Commercialization of Threatened Species
The present results showed that 13 out of the 20 taxa of elasmobranchs are threatened with extinction and five of them are nearly threatened according to the Red List of Threatened Brazilian Fauna (ICMBio, 2018a) and the International Red List of Threatened Species (IUCN, 2021). When both national and international lists are compared, we noticed species classified under different threatening statuses. For instance, R. bonasus is regarded as DD by ICMBio (2018a) but as VU according to IUCN (2021), while C. falciformis is classified as NT and VU by ICMBio (2018a) and IUCN (2021), respectively.
Elasmobranchs tend to be sold uncharacterized and often using commercial names, usually referring to a general fish category. The lack of standardization in nomenclature can also make it easier for species with different threat status to be sold together. A case, frequently observed in the present study, was the utilization of the local term “cação panã” to refer to the following species: R. porosus, C. porosus, S. tiburo, S. tudes, S. mokarran, and S. lewini. Similarly, the name long-nose stingray or “arraia bicuda” was indiscriminately used to refer to H. berthalutzae, H. guttatus and F. geijskesi. These data highlight that commercial labels are not a reliable source of information for the species identification, as previously reported by other authors (Clarke et al., 2006; Rodrigues-Filho et al., 2009; Domingues et al., 2013).
Commercialization of Shark Species
Six species of the family Carcharhinidae (R. porosus, C. porosus, C. leucas C. falciformis, C. plumbeus, and C. acronotus) were commercialized, presenting the highest species richness but with lower frequency in relation to those representatives of the family Sphyrnidae.
Three out of the six Carcharhinidae species herein identified are classified as threatened, like C. porosus whose status is defined as “critically endangered.” This species was rarely reported in the present study, thus differing from a previous report (Rodrigues-Filho et al., 2009) that placed C. porosus as the most commercialized shark species in fish markets of Bragança. The population decline of C. porosus, formerly abundant in catches along the northern Brazilian coast, has increased from 1990 to 2004 (Rodrigues-Filho et al., 2009; ICMBio, 2018b), associated with cases of local extinction (ICMBio, 2018b; Dulvy et al., 2021).
The species R. porosus was observed being commercialized, similarly other reports have detected the increased commercialization of this species (Feitosa et al., 2018; Martins et al., 2018). Even though R. porosus is not considered a threatened species, population approaches are encouraged since most of samples collected in the present study referred to newborns removed from pregnant females.
On the other hand, C. falciformis, C. plumbeus, and C. leucas, usually exploited in fin markets (Clarke et al., 2006), encompassed a few samples in this study, differing from other reports in southeastern Brazilian coast, where C. falciformis was the most abundant shark species (51%) in fish landings (Domingues et al., 2013). These data show that the supply of these species is sporadic, possibly because they derive from bycatches or these species may be being marketed in other locations (Lessa et al., 1999).
In relation to the family Ginglymostomatidae, the species G. cirratum is regarded as vulnerable according to Brazilian official reports (ICMBio, 2018a). Other reports have indicated that stocks of G. cirratum along the Brazilian coast have decreased over the last years (SBEEL, 2005).
Based on these results, we observed a change in the pattern of shark commercialization with the highest abundance for Sphyrnidae while previous reports indicated representatives of Carcharhinidae as the most commercialized species (Rodrigues-Filho et al., 2009; Feitosa et al., 2018). Within the family Sphyrnidae, S. mokarran was the most frequently exploited species as also observed in another study carried out along the northern Brazilian coast that placed this shark as this shark is the fourth most found and sampled species (Feitosa et al., 2018). These data are worrying since all species of the genus Sphyrna identified in the present study are threatened with extinction.
For instance, during the development of this study the status of S. mokarran has changed from endangered to critically endangered (IUCN, 2021). These results are worrisome, since overexploited species tend to have their populations reduced and may lead to extinction (Dulvy et al., 2014).
Commercialization of Ray Species
We identified two species belonging to the family Rhinopteridae, being Rhinoptera brasiliensis considered threatened for being sensitive species to fishing pressure, as the females of this species have only one offspring per gestation (Vooren et al., 2005; IUCN, 2021). The occurrence of the species on the market was not regular, suggesting an occasional trade. This scenario corroborates with previous studies that reported a low number of samples found in Bragança (Rodrigues-Filho et al., 2020). These data are in agreement with previous studies showing that R. brasiliensis, which was previously considered restricted to the southern coast of America (Lessa et al., 1999), actually presents a wider range of occurrence, reaching the northern coast of Brazil (Marceniuk et al., 2019; Rodrigues-Filho et al., 2020).
Another representative of the family Rhinopteridae, Rhinoptera bonasus, was also represented by a few samples in the present work. This species was also reported in another study along the northern coast (Rodrigues-Filho et al., 2020).
The species G. micrura in the present study showed a genetic divergence of 10.10% in relation to the sequences of the G. micrura populations from the Mexican coast (Sarmiento-Camacho and Valdez-Moreno, 2018), suggesting a restriction of the gene flow between populations from the north and south of the Atlantic. On the other hand, the genetic similarity was equal to 100% when the present sequences of this species were compared to other samples from northern Brazil (Rodrigues-Filho et al., 2020). This result might indicate that this taxon encompasses two distinct groups (or even species). In fact, Yokota and De Carvalho (2017) suggested a taxonomic revision of G. micrura, recommending the denomination Gymnura lessae for the populations distributed along United States coast and Gulf of Mexico, while G. micrura would be restricted to central and southern regions of Western Atlantic (Brazil, northern Venezuela, Trinidad, and Tobago).
Dasyatidae family was represented by the commercialization of H. berthalutzae, H. guttatus, and F. geijskesi, species that have been previously considered as data deficient but after a recent reevaluation (IUCN, 2021) are presently regarded as vulnerable, nearly endangered and critically endangered, respectively. According to ICMBio (2018a), H. guttatus is a species of least concern by presenting no significant threats, while H. berthalutzae remains not evaluated (NE) because of its recent description (Pentean et al., 2020) and F. geijskesi was classified as DD (ICMBio, 2018a).
In the present study, we recorded the commercialization of H. berthalutzae (formerly recognized as H. americanus) for the first time. The geographic distribution of this species ranges from the mouth of Amazonas River to the coast of São Paulo, southeastern Brazil, probably representing an endemic species to the Brazilian coast (Pentean et al., 2020). We also identified the trade of the endangered species (IUCN, 2021) S. schmardae belonging to the Potamotrygonidae family.
In the sample labeled as ray, we also found a specie that could not be attributed to any taxon Myliobatiformes 1 (Arr63F136). After evaluating and comparing with all those stingray species described for the North coast with COI sequences available in public banks, Myliobatiformes 1 cannot be precisely identified (Supplementary Figure 2).
Impacts Caused by the Exploitation of Elasmobranchs
When we compared our results with previous studies, the reported species diversity changed. For example, Sphyrna zygaena, Rhizoprionodon terraenovae, and Negaprion brevirostris were found to be traded in the open market in Bragança (Rodrigues-Filho et al., 2009), while the landing of Pristis pristis in Bragança was reported by Palmeira et al. (2013). These referenced species were not recorded in this study throughout the sampling period. The absence of these species can be explained by their commercialization in other places or even that they are showing a possible decline in their natural populations, since these taxa are classified as critically threatened by overfishing (IUCN, 2021).
The evaluation of exploitation of sharks and rays, whether at a regional level or on a global scale, is difficult, once catch volume estimates and landing statistical data in fisheries are often incomplete and superficial (Dent and Clarke, 2015). Fishing practices are particularly harmful to these fishes since most elasmobranchs are k-strategist animals that are more susceptible to exploitation and have slow recoveries after stock depletions (Stevens et al., 2000; Oliver et al., 2015; Pacoureau et al., 2021).
In order to avoid overexploitation of these fisheries resources, deep changes are necessary, such as strict monitoring of what species have been caught and landed, implementation of on-board and landing inspectors, as well as the use of more selective fishing tools to target species thus reducing bycatches (Barreto et al., 2017). Popular names also need to be re-evaluated in future fisheries statistics as they vary greatly among regions and fail in providing reliable information about which species are being exploited (Ardura et al., 2010; Feitosa et al., 2018).
Moreover, surveillance strategies need to be expanded since a large number of elasmobranchs are widely distributed, including nearly 150 species of shark that migrate across national borders, while a quarter of threatened species can migrate across at least 18 countries (Dulvy et al., 2014). Therefore, global public policies are required to the proper conservation of these organisms.
In this context, DNA-based methodologies, such as the use of DNA barcode, as well as the enrichment of public banks with reference DNA sequences for several taxa are particularly useful to obtain accurate data. This information can be further used to support reliable statistical data for the development of efficient management plans in fisheries (Dulvy et al., 2014; Oliver et al., 2015; Barreto et al., 2016).
Conclusion
Twenty-four commercial nomenclatures for elasmobranchs were recorded in the street market of Bragança, referring to 11 species of sharks and 9 taxa of rays, as discriminated by DNA barcode. Sphyrnidae and Carcharhinidae were the most sampled families of sharks, while Rhinopteridae and Dasyatidae were the most represented groups of rays. We also identified the regular trade of threatened species, particularly in the family Sphyrnidae (e.g., S. mokarran and S. lewini). This scenario is largely related to the lack of standardized commercial nomenclatures and mischaracterization of traded specimens thus revealing the importance of using molecular tools, especially DNA barcode in species validation.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Ethics Statement
Ethical review and approval was not required for the animal study because the animals were bought at the fish market. They were already dead.
Author Contributions
TM and GE-G participated in the study design and structure. TM, PS, and GE-G obtained and organized the database. IS and GE-G generated the computational data. TM, AG-C, and RS performed the statistical analysis. TM, PS, ÍL, AG-C, and GE-G wrote the draft manuscript. TM, PS, ÍL, and GE-G reviewed the manuscript. TM and ÍL performed the final formatting of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Financial Code 001, by granting a master’s scholarship to TM, to CNPq for financing this study (Universal project #439113/2018-0).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We are grateful to the fishmongers at the street market of Bragança for donating most of samples, and to the Universidade Federal do Pará (UFPA), Pró-Reitoria de Pesquisa e Pós-Graduação (PROPESP) for the financial assistance in publication fees.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2021.769908/full#supplementary-material
Footnotes
- ^ http://www.boldsystems.org/index.php/MAS_Management_DataConsole?codes=CA
- ^ http://www.ncbi.nlm.nih.gov
- ^ http://www.boldsystems.org/
- ^ https://rawgraphs.io/
References
Ardura, A., Linde, A. R., Moreira, J. C., and Garcia-Vazquez, E. (2010). DNA barcoding for conservation and management of Amazonian commercial fish. Biol. Conserv. 143, 1438–1443. doi: 10.1016/j.biocon.2010.03.019
Barreto, R. R., Bornatowski, H., Motta, F. S., Santander-Neto, J., Vianna, G. M. S., and Lessa, R. (2017). Rethinking use and trade of pelagic sharks from Brazil. Mar. Policy 85, 114–122. doi: 10.1016/j.marpol.2017.08.016
Barreto, R., Ferretti, F., Flemming, J. M., Amorim, A., Andrade, H., Worm, B., et al. (2016). Trends in the exploitation of South Atlantic shark populations. Conserv. Biol. 30, 792–804. doi: 10.1111/cobi.12663
Braga, C. F., Espírito-Santo, R. V., Silva, B. B., Giarrizzo, T., and Castro, E. R. (2006). Considerações sobre a comercialização do pescado no município de Bragança-PA. Bol. Téc. Cient. Cepnor 6, 105–120. doi: 10.17080/1676-5664/btcc.v6n1p105-120
Carmona, N., Sampaio, I., Santos, S., Souza, R. F. C., and Schneider, H. (2008). Identificação de arraias marinhas comerciais da costa norte brasileira com base em sequências de DNA mitocondrial. Bol. Téc. Cient. Cepnor 8, 51–58. doi: 10.17080/1676-5664/btcc.v8n1p51-58
Carvalho, S. C., Sampaio, I., and Santos, S. (2020). DNA barcoding reveals mislabeling and commercial fraud in the marketing of fillets of the genus Brachyplatystoma Bleeker, 1862, the Amazonian freshwater catfishes economically important in Brazil. Heliyon 6:e04888. doi: 10.1016/j.heliyon.2020.e04888
Clarke, S. C., Magnussen, J. E., Abercrombie, D. L., McAllister, M. K., and Shivji, M. S. (2006). Identification of shark species composition and proportion in the Hong Kong shark fin market based on molecular genetics and trade records. Conserv. Biol. 20, 201–211. doi: 10.1111/j.1523-1739.2005.00247.x
Compagno, L. J. V. (1990). Alternative life-history styles of cartilaginous fishes in time and space. Environ. Biol. Fish. 28, 33–75. doi: 10.1007/BF00751027
Darriba, D., Taboada, G. L., Doallo, R., and Posada, D. (2012). JModelTest 2: more models, new heuristics and parallel computing. Nat. Methods. 9:772. doi: 10.1038/nmeth.2109
Dent, F., and Clarke, S. (2015). State of the Global Market for Shark Products. FAO Fisheries and Aquaculture Technical Paper. Rome: FAO.
Domingues, R. R., de Amorim, A. F., and Hilsdorf, A. W. S. (2013). Genetic identification of Carcharhinus sharks from the southwest Atlantic Ocean (Chondrichthyes: Carcharhiniformes). J. Appl. Ichthyol. 29, 738–742. doi: 10.1111/jai.12154
Drummond, A. J., Suchard, M. A., Xie, D., and Rambaut, A. (2012). Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 29, 1969–1973. doi: 10.1093/molbev/mss075
Dulvy, N. K., Fowler, S. L., Musick, J. A., Cavanagh, R. D., Kyne, P. M., Harrison, L. R., et al. (2014). Extinction risk and conservation of the world’s sharks and rays. Elife 3, 1–34. doi: 10.7554/elife.00590
Dulvy, N. K., Pacoureau, N., Rigby, C. L., Pollom, R. A., Jabado, R. W., Ebert, D. A., et al. (2021). Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Curr. Biol. 31, 4773–4787.e8. doi: 10.1016/j.cub.2021.08.062
Espírito Santo, R. V., and Isaac, V. J. (2012). Desembarques da pesca de pequena escala no município de Bragança – PA, Brasil: esforço e produção. Bol. Lab. Hidrobiol. 25, 31–48.
Feitosa, L. M., Martins, A. P. B., Giarrizzo, T., MacEdo, W., Monteiro, I. L., Gemaque, R., et al. (2018). DNA-based identification reveals illegal trade of threatened shark species in a global elasmobranch conservation hotspot. Sci. Rep. 8, 1–11. doi: 10.1038/s41598-018-21683-5
Ferrette, B. L. F., Domingues, R. R., Rotundo, M. M., Miranda, M. P., Bunholi, I. V., De Biasi, J. B., et al. (2019). DNA barcode reveals the bycatch of endangered batoids species in the southwest Atlantic: implications for sustainable fisheries management and conservation efforts. Genes 10, 1–15. doi: 10.3390/genes10040304
Freire, J. L., Silva, B. B., and Souza, A. S. (2011). Aspectos econômicos e higiênico-sanitários da comercialização do pescado no município de bragança (pa). Biota Amaz. 1, 17–28. doi: 10.18561/2179-5746/biotaamazonia.v1n2p17-28
Gomes, G., Correa, R., Veneza, I., da Silva, R., da Silva, D., Miranda, J., et al. (2019). Forensic analysis reveals fraud in fillets from the “Gurijuba” Sciades parkeri (Ariidae–Siluriformes): a vulnerable fish in brazilian coastal Amazon. Mitochondrial DNA Part A 30, 721–729. doi: 10.1080/24701394.2019.1622694
Graham, N. A. J., Spalding, M. D., and Sheppard, C. R. C. (2010). Reef shark declines in remote atolls highlight the need for multi-faceted conservation action. Aquat. Conserv. Mar. Freshw. Ecosyst. 20, 543–548. doi: 10.1002/aqc.1116
Guimarães-Costa, A., Machado, F. S., Reis-Filho, J. A., Andrade, M., Araújo, R. G., Corrêa, E. M. R., et al. (2020). DNA barcoding for the assessment of the taxonomy and conservation status of the fish bycatch of the Northern Brazilian shrimp trawl fishery. Front. Mar. Sci. 7:566021. doi: 10.3389/fmars.2020.566021
Hall, T. A. (1999). BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 41, 95–98.
Hebert, P. D. N., Cywinska, A., Ball, S. L., and DeWaard, J. R. (2003). Biologicalidentifications through DNA barcodes. Proc. R. Soc. B Biol. Sci. 270, 313–321. doi: 10.1098/rspb.2002.2218
ICMBio (2018a). Instituto Chico Mendes de Conservação da Biodiversidade. Livro Vermelho da Fauna Brasileira Ameaçada de Extinção, Vol. 1. Brasília: ICMBio.
ICMBio (2018b). Instituto Chico Mendes de Conservação da Biodiversidade. Livro Vermelho da Fauna Brasileira Ameaçada de Extinção - Peixes, Vol. 6. Brasília: ICMBio.
Isaac, V. J. I., Espírito Santo, R. V., and Nunes, J. L. G. (2008). A estatística pesqueira no litoral do Pará: resultados divergentes. Pan Am. J. Aquat. Sci. 3, 205–213.
IUCN (2021). International Union for Conservation of Nature, Red List of Threatened Species 2021. Available online at: https://www.iucnredlist.org (accessed 10 June, 2021).
Kumar, S., Stecher, G., Li, M., Knyaz, C., and Tamura, K. (2018). MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 35, 1547–1549. doi: 10.1093/molbev/msy096
Larkin, M. A., Blackshields, G., Brown, N. P., Chenna, R., McGettigan, P. A., McWilliam, H., et al. (2007). Clustal W and clustal X version 2.0. Bioinformatics 23, 2947–2948. doi: 10.1093/bioinformatics/btm404
Lessa, R. S., Rincon, G., Gadig, O. B., and El-Deir, A. C. A. (1999). Biodiversidade de Elasmobrânquios no Brasil, Projeto de Conservação e utilização da Diversidade Biológica Brasileira (PROBIO), Ministério do Meio Ambiente. Brasília: Ministério do Meio.
Librado, P., and Rozas, J. (2009). DnaSP v5: a software for comprehensive analysis of DNA polymorphism data. Bioinformatics 25, 1451–1452. doi: 10.1093/bioinformatics/btp187
Marceniuk, A. P., Barthem, R. B., Wosiacki, W. B., Klautau, A. D. M., Junior, T. V., Rotundo, M. M., et al. (2019). Sharks and batoids (Subclass Elasmobranchii) caught in the industrial fisheries off the Brazilian North coast. Rev. Nord. Biol. 27, 120–142. doi: 10.22478/ufpb.2236-1480.2019v27n1.47112
Martins, A. P. B., Feitosa, L. M., Lessa, R. P., Almeida, Z. S., Heupel, M., Silva, W. M., et al. (2018). Analysis of the supply chain and conservation status of sharks (Elasmobranchii: Superorder Selachimorpha) based on fisher knowledge. PLoS One 13:e0193969. doi: 10.1371/journal.pone.0193969
Martins, T., Da Silva, R., Veneza, I., Santana, P., Correa, R., Santa Brígida, N., et al. (2021). Diversity and abundance of commercialized fish in northeastern Pará, coastal amazon: the case of the street market in Bragança-PA. Arq. Cienc. Mar. 54, 27–43. doi: 10.32360/acmar.v54i1.43960
Mc Eachran, J. D., and Carvalho, M. R. (2002). “FAO species identification guide for fishery purposes. The living marine resources of the Western Central Pacific,” in Introduction, Molluscs, Crustaceans, Hagfishes, Sharks, Batoid fishes and Chimaeras, Vol. 1, eds K. E. Carpenter and V. H. Niem (Rome: FAO).
Miller, M. A., Pfeier, W., and Schwartz, T. (2010). “Creating the CIPRES Science Gateway for inference of large phylogenetic trees,” in Proceedings of the 2010 Gateway Computing Environments Workshop (GCE) (Piscataway, NJ: IEEE).
Mourão, K. R. M., Frédou, F. L., Espírito-Santo, R. V., de Almeida, M. C., da Silva, B. B., Frédou, T., et al. (2018). Sistema de produção pesqueira pescada amarela-Cynoscion acoupa Lacèpede (1802): um estudo de caso no litoral nordeste do Pará-Brasil. Bol. Inst.Pesca 35, 497–511.
Nelson, J. S., Grande, T. C., and Wilson, M. V. H. (2016). Fishes of the World. New Jersey, NJ: John Wiley & Sons.
Oliver, S., Braccini, M., Newman, S. J., and Harvey, E. S. (2015). Global patterns in the bycatch of sharks and rays. Mar. Policy. 54, 86–97. doi: 10.1016/j.marpol.2014.12.017
Pacoureau, N., Rigby, C. L., Kyne, P. M., Sherley, R. B., Winker, H., Carlson, J. K., et al. (2021). Half a century of global decline in oceanic sharks and rays. Nature 589, 567–571. doi: 10.1038/s41586-020-03173-9
Paithankar, K. R., and Prasad, K. S. N. (1991). Precipitation of DNA by polyethylene glycol and ethanol. Nucleic Acids Res. 19:1346. doi: 10.1093/nar/19.6.1346
Palmeira, C. A. M., da Silva Rodrigues-Filho, L. F., de Luna Sales, J. B., Vallinoto, M., Schneider, H., and Sampaio, I. (2013). Commercialization of a critically endangered species (largetooth sawfish, Pristis perotteti) in fish markets of northern Brazil: authenticity by DNA analysis. Food Control 34, 249–252. doi: 10.1016/j.foodcont.2013.04.017
Pentean, F. F., Naylor, G. J., and Lima, S. M. (2020). Integrative taxonomy identifies a new stingray species of the genus Hypanus Rafinesque, 1818 (Dasyatidae. Myliobatiformes), from the Tropical Southwestern Atlantic J. Fish Biol. 97, 1120–1142. doi: 10.1111/jfb.14483
Pough, F. H., Janis, C. M., and Heiser, J. B. (2008). A Vida Dos Vertebrados, Fourth. São Paulo: Atheneu.
Rambaut, A. (2018). FigTree v1. 4.4, a Graphical Viewer of Phylogenetic Trees. Available online at: https://github.com/rambaut/figtree (accessed 18 April, 2021).
Rambaut, A., Drummond, A. J., Xie, D., Baele, G., and Suchard, M. A. (2018). Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 67, 901–904. doi: 10.1093/sysbio/syy032
Ratnasingham, S., and Hebert, P. (2007). The barcode of life data system. Mol. Ecol. Notes 7, 355–364. doi: 10.1111/j.1471-8286.2006.01678.x
Ratnasingham, S., and Hebert, P. D. N. (2019). Barcode of Life Data Systems Handbook A web-Based Bioinformatics Platform Supporting The Dna Barcoding Of Animal, Plant, And Fungal Species. Available online at: http://www.boldsystems.org/index.php/MASAnalysis2BarcodeGapAnalysis/preprocess/ (accessed 27 September, 2020).
Robbins, W. D., Hisano, M., Connolly, S. R., and Choat, J. H. (2006). Ongoing collapse of coral-reef shark populations. Curr. Biol. 16, 2314–2319. doi: 10.1016/j.cub.2006.09.044
Rodrigues-Filho, L. F., da Rocha, T. C., do Rêgo, P. S., Schneider, H., Sampaio, I., and Vallinoto, M. (2009). Identification and phylogenetic inferences on stocks of sharks affected by the fishing industry off the Northern coast of Brazil. Genet. Mol. Biol. 32, 405–413. doi: 10.1590/s1415-47572009005000039
Rodrigues-Filho, L. F., da, S., Feitosa, L. M., Silva Nunes, J. L., Onodera Palmeira, A. R., Martins, A. P. B., et al. (2020). Molecular identification of ray species traded along the Brazilian Amazon coast. Fish. Res. 223:105407. doi: 10.1016/j.fishres.2019.105407
Sanger, F., Nicklen, S., and Coulson, A. R. (1977). DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. U.S.A. 74, 5463–5467. doi: 10.1073/pnas.74.12.5463
Santana, P., da Silva, R., Veneza, I., Martins, T., Santa Brígida, N., Gonçalves, J., et al. (2020). Prospecção de Crustáceos e Moluscos comercializados na Feira livre de Bragança, Pará, Amazônia costeira. Bogota, D.C: Editora Científica.
Sarmiento-Camacho, S., and Valdez-Moreno, M. (2018). DNA barcode identification of commercial fish sold in Mexican markets. Genome 61, 457–466. doi: 10.1139/gen-2017-0222
SBEEL (2005). Sociedade Brasileira para o Estudo de Elasmobrânquios Plano de Ações para Conservação e Manejo dos Estoques dos Recursos Pesqueiros. Rio Grande, RS: SBEEL
Souza-Araujo, J., Souza-Junior, O. G., Guimarães-Costa, A., Hussey, N. E., Lima, M. D. O., and Giarrizzo, T. (2021). The consumption of shark meat in the Amazon region and its implications for human health and the marine ecosystem. Chemosphere 265:129132. doi: 10.1016/j.chemosphere.2020.129132
Steinke, Di, Bernard, A. M., Horn, R. L., Hilton, P., Hanner, R., and Shivji, M. S. (2017). DNA analysis of traded shark fins and mobulid gill plates reveals a high proportion of species of conservation concern. Sci. Rep. 7, 5–10. doi: 10.1038/s41598-017-10123-5
Stevens, J. D., Bonfil, R., Dulvy, N. K., and Walker, P. A. (2000). The effects of fishing on sharks, rays, and chimaeras (chondrichthyans), and the implications for marine ecosystems. ICES J. Mar. Sci. 57, 476–494. doi: 10.1006/jmsc.2000.0724
Thompson, J. D., Higgins, D. G., and Gibson, T. J. (1994). CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22, 4673–4680. doi: 10.1093/nar/22.22.4673
Vooren, C. M., Klippel, S., and Galina, A. B. (eds). (2005). “Os elasmobrânquios das águas costeiras da Plataforma Sul,” in Ações Para A Conservação De Tubarões E Raias No Sul Do Brasil. Sandro Klippel. (Porto Alegre: Igarapé), 262.
Ward, R. D., Zemlak, T. S., Innes, B. H., Last, P. R., and Hebert, P. D. N. (2005). DNA barcoding Australia’s fish species. Philos. Trans. R. Soc. B Biol. Sci. 360, 1847–1857. doi: 10.1098/rstb.2005.1716
Keywords: Bragança, northeastern Pará, northern coast, threatened species, COI
Citation: Martins T, Santana P, Lutz Í, da Silva R, Guimarães-Costa A, Vallinoto M, Sampaio I and Evangelista-Gomes G (2021) Intensive Commercialization of Endangered Sharks and Rays (Elasmobranchii) Along the Coastal Amazon as Revealed by DNA Barcode. Front. Mar. Sci. 8:769908. doi: 10.3389/fmars.2021.769908
Received: 02 September 2021; Accepted: 25 November 2021;
Published: 14 December 2021.
Edited by:
Zhijun Dong, Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Juan Andrés López, University of Alaska Fairbanks, United StatesJose Julian Tavera, University of Valle, Colombia
Copyright © 2021 Martins, Santana, Lutz, da Silva, Guimarães-Costa, Vallinoto, Sampaio and Evangelista-Gomes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Grazielle Evangelista-Gomes, Z3JhemllbGxlZmVnQGdtYWlsLmNvbQ==, Z3JhemllbGxlQHVmcGEuYnI=