 Mariana Cravo1*
Mariana Cravo1* Armando J. Almeida1,2Hamilton Lima3
Armando J. Almeida1,2Hamilton Lima3 João Azevedo e Silva1
João Azevedo e Silva1 Salomão Bandeira4Vilma Machava-António4José Paula1,5
Salomão Bandeira4Vilma Machava-António4José Paula1,5
- 1Department of Animal Biology, Faculty of Sciences, University of Lisbon, Lisbon, Portugal
- 2Maritime Laboratory of Guia, Faculty of Sciences, Marine and Environmental Sciences Centre, University of Lisbon, Lisbon, Portugal
- 3Príncipe Island UNESCO Biosphere Reserve, Autonomous Region of Príncipe, Santo António, São Tomé and Príncipe
- 4Department of Biological Sciences, Eduardo Mondlane University, Maputo, Mozambique
- 5Faculty of Sciences, Marine and Environmental Sciences Centre, University of Lisbon, Lisbon, Portugal
Mangroves are highly productive ecosystems with complex adaptations to the transition between freshwater and sea. Mangroves function as nursery habitats for many organisms, providing protection and food sources for early developmental stages of crustaceans and fish, helping to maintain adjacent marine stocks. Mangroves in São Tomé and Príncipe remain poorly studied. This study addresses the importance of a small mangrove stand to ichthyofauna. The main goal of the study was to describe the fish assemblages of the Praia Salgada mangrove stand on Príncipe Island, and assess if variations in the season, tide, and mangrove zone affected fish distribution. Fish assemblages were sampled with mosquito nets during the rainy and dry seasons, and neap and spring tides, while environmental parameters such as water depth, temperature, pH, and salinity measurements were taken. The characteristics of the water column were affected by a sandbank that developed between sampling seasons, impacting on the dynamics of the water and biological exchanges between the mangrove stand and the adjacent marine environment. The study identified 14 fish species occurring in the Praia Salgada mangrove stand from a total of 772 specimens caught. Five species were recorded for the first time as occurring in the country’s mangrove areas, namely Caranx latus, Ethmalosa fimbriata, Mugil curema, Gobioides cf. africanus, and Citharus cf. linguatula. Most of the reported species are of commercial interest, and were predominantly juveniles, suggesting that the mangrove ecosystem provides a nursery function for several species. Some species revealed preferences for either the upper or lower part of the mangrove forest. The size of fish sampled tended to be bigger during the dry season, especially for the Mugilidae, Aplocheilichthys spilauchen and Gobiidae groups. E. fimbriata and Eucinostomus melanopterus displayed similar sizes between seasons. The average quantity of fish caught per day in the rainy season was three times higher than in the dry season. The fish species distribution in the mangrove stand varied significantly according to the season and mangrove zone. The overall results suggest that the Praia Salgada mangrove stand provides a nursery function for several of the studied fish species.
Introduction
Mangroves are defined as tropical trees restricted to intertidal and adjacent ecosystems or subjected to indirect tidal influence (Tomlinson, 2016). Mangrove environments have unique characteristics, both environmental and biological, attracting high biodiversity and providing important ecological services.
These aquatic forests benefit coastal resources by supporting the early stages of commercial and non-commercial fauna such as crustaceans, mollusks, and fishes (Tomlinson, 2016). Mangroves provide food sources mainly via the detritus chain (Tomlinson, 2016) and provide shelter through the complex tangling of roots and associated water turbidity. Young stages of fish take advantage of these conditions that provide a favorable and safe environment for their development. The density of several fish species on coral reefs appear to be related to nearby bays containing mangroves that function as nurseries (Nagelkerken et al., 2002). In addition, declines in local fish catches result when mangrove forests are destroyed (FAO, 2007).
Besides the nursery, breeding, and feeding areas that mangroves provide for juvenile fishes, other ecosystem services such as shoreline protection, provisioning of wood and non-wood products, climate regulation, water purification, cultural services, and several ecosystem supporting benefits such as nutrient cycling, soil formation, and primary production are provided by these forests (MEA, 2005).
São Tomé and Príncipe is an insular African country composed of two main islands and several islets. The islands are of volcanic origin and are part of the volcanic chain known as the Cameroon line, which extends for 1600 km from the ocean in the Gulf of Guinea to the African continent. The islands were never connected to the mainland, resulting in significant differentiation of fauna and flora and a high degree of endemism (Bonfim and Carvalho, 2009).
São Tomé has at least four mangrove forests while Príncipe supports three small mangrove stands (Praia Salgada, Praia Grande, and Praia Caixão). These aquatic forests in São Tomé are made of two families of mangrove trees; black mangroves Avicennia germinans (Linnaeus, 1764), and red mangroves of the genus Rhizophora (Afonso, 2019). Mangrove stands on Príncipe Island are composed of monostands of Rhizophora harrisonii individuals.
In the past years, more than 20% of the mangrove area worldwide has been lost (FAO, 2007). Even though mangrove forests have been proved to be essential to the well-being of human populations and coastal fish stocks (FAO, 2007; UNEP, 2010), there is still much to describe and monitor so that sustainable management of these environments is achieved, particularly for insular tropical systems. São Tomé and Príncipe has almost no records relating to its mangroves. The only published work on the mangroves of Príncipe Island is the report by Herrero-Barrencua et al. (2017), describing the mangrove ecosystems with faunal lists and flora characterization, but with little emphasis on the ichthyofauna.
The present study aimed to describe the mangrove environment of Praia Salgada on Príncipe Island, including the hydrodynamic aspects and fish assemblages. The specific objectives were to describe the abundance, species richness and diversity of the fish fauna, correlate the fish abundance and size with environmental variables, and assess the potential nursery role of the mangrove stands for the fish community.
The present work is the first in the country to compare mangrove fish abundance between the seasons. It has also determined the exact tree coverage of R. harrisonii, allowing future comparisons of mangrove cover and detect possible regressions or growth. This study covered two seasons between October 2019 and February 2020, and determined the influence of seasonality on water column characteristics, species richness and abundance of fish. Variations in temperature, salinity, pH, and depth between the tides (spring and neap) and seasons (rainy and dry) in the upper and lower mangrove zones were also measured and related to fish catches in order to assess if the distribution of fish in the mangrove stand was related to these factors.
Materials and Methods
Study Area
This study is focused on a small mangrove system in Príncipe Island, São Tomé and Príncipe (Gulf of Guinea). The Praia Salgada mangrove stand (Figure 1), located in Abade Bay on the east coast of the island, is a small brackish water system receiving freshwater from the Água Grande River and salt water from the Atlantic Ocean. Four sampling sites were established, the first three being within the main mangrove creek (upper, middle, and lower mangrove) surrounded by R. harrisonii trees, and the fourth between the end of the mangrove vegetation and the mouth of the estuary.
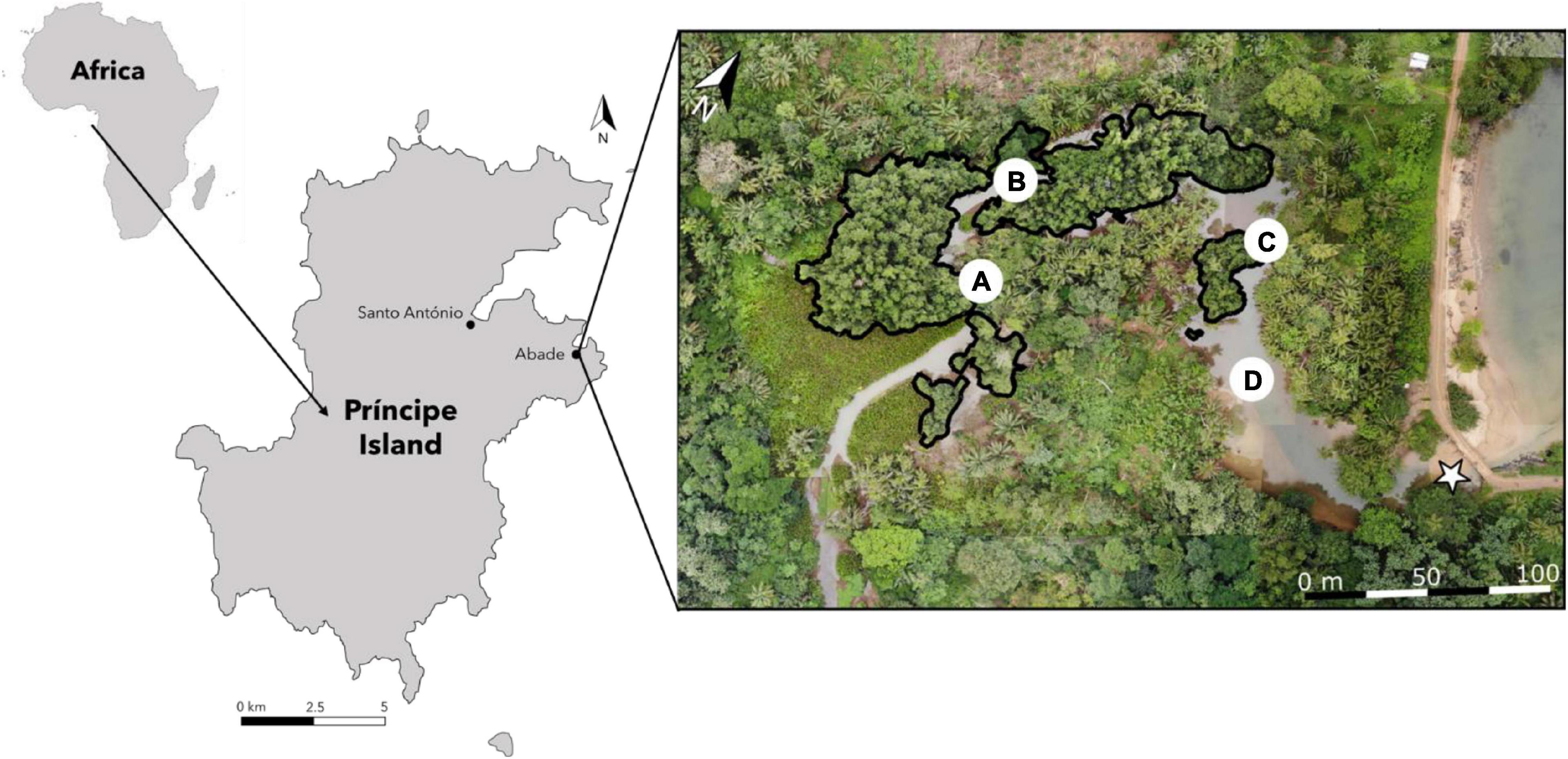
Figure 1. Map of the Praia Salgada mangrove stand (on the right) and respective location in Príncipe Island and African continent (on the left). Black perimeter in the mangrove indicates the distribution of the mangrove tree Rhizophora harrisonii. Sampling sites: A – upper; B – middle; C – lower; and D – outer. Points A–D represent the sampling points for the water column characterization, and points A and C correspond to the fishing points. ✩ – location where the sandbank was formed. The map of Praia Salgada mangrove stand (on the right) was constructed with aerial photography obtained with a drone.
The country has a humid tropical climate with four seasons alternating between rainier and dryer phases of different intensities. The less pronounced dry season falls between January and March, followed by the less pronounced rainy season in April–June with (Chou et al., 2020). The main dry season from July to September is called “gravana” (Pisoni et al., 2015; Chou et al., 2020), followed by the most intense rainy season from October–December (Chou et al., 2020). The average annual rainfall ranges from 2000 to 3000 mm, reaching 7000 mm in the cloud forests (NBSAP II, 2015; Chou et al., 2020). Temperatures range from 18–21°C minimum to 30–35°C maximum (Herrero-Barrencua et al., 2017) and with an annual average of 26°C (NBSAP II, 2015). The relative humidity of the air is very high, reaching more than 90% at higher altitudes (NBSAP II, 2015).
Water Column Characterization
To characterize the water column, sampling was performed with a multimeter probe (YSI Multi 350i multimeter) measuring temperature and salinity. The water column depth was measured with a measuring tape or a depth probe, depending on the sampling method (by foot or by boat, respectively). During high tide, the measurements were taken twice, at the top and bottom of the water column, and during low tide only once at mid-depth due to the reduced water column height.
The sampling took place in the seasons: rainy (October, November, and December) and dry (January and February), with 6 days of sampling in each season. These 6 days were evenly distributed between neap and spring tides, with fieldwork during low and high tide on each day. Four working stations (see Figure 1A–D) were established in the mangrove stand. The water column was characterized (temperature, salinity, and depth) for each day in non-successive triplicates for each of the four points.
A sandbank at the river mouth (see Figure 1✩) developed between December and January and increased until February, obstructing water circulation and the mobility of organisms across the estuarine boundary. This different topography was due to the transport and deposition of sediments originating upstream, together with the transport of sand by sea waves. This accumulation of sediments that concentrated at the river mouth significantly reduced the width and depth of the river channel at this narrowest point. During the rainy season the strength of the estuarine discharge precludes the formation of these deposits.
Assessment of Fish Assemblages
Sampling to assess the species richness and abundance of the community took place in the same months as the water column characterization. Sampling was performed at low tide (the only time possible due to the width and depth of the water column) at upper and lower mangrove sites (Figure 1A,C), with two fishing attempts during the same low tide at each site per day.
The sampling was carried out with mosquito nets (1.5 mm mesh size, 2.0 m wide, and 1.2 m high, with a sack shape) and operations involved at least four people. Two people held the net open, with the other two splashing water from a starting point about 10 m upstream to chase organisms into the net. After capture the fish were photographed on-site with a reference scale to later determine the total length (TL) of each individual using the software ImageJ™. The procedure was undertaken as rapidly as possible so that the individuals could be returned to the environment without jeopardizing their survival. Fish sub-samples for identification purposes were retained and transported to the Biosphere Reserve and fixed in a 6% formaldehyde solution, and transferred to a new solution of 80% alcohol after about 24 h.
The fish were identified to species level when possible, except when the lack of adequate descriptions prevented an accurate identification, such as in the case of late larval and juvenile stages. The fish guides by Oren (1981), Lévêque et al. (1990a), Guatier and Hussenot (2005), and FAO (2016a,b), and complemented with the consultation of FishBase (Froese and Pauly, 2019) were used for identification purposes. The lists of fish taxa are presented in taxonomic order according to Nelson (2006).
This study did not require ethics approval or specific consent procedures. The work did not involve animal experimentation and did not compromise animal welfare. The field work involving fish was simple and returned the vast majority of individuals to the environment without harming them, given that the procedure was quick and simple.
Data Analysis
The water column parameters data were normalized, and a resemblance matrix constructed using Euclidean distance. Water column parameters were then compared using principal coordinates analysis (PCO), equivalent do principal component analysis (PCA), considering three factors: season (rainy and dry), tide (neap and spring) and mangrove zone (upper and lower).
Diversity of the fish community between seasons was ascertained by the Shannon–Wiener index (H′) given as , where s is the number of species in the community and Pi is the proportion of individuals belonging to species i in the community (Krebs, 1999). The evenness or equitability component of diversity was calculated from Pielou’s index (Pielou, 1966) given as J′ = H′/Hmax where Hmax = ln s.
Fish assemblage data were logarithmized, and a resemblance matrix was constructed using the “zero-adjusted” Bray–Curtis similarity coefficient (Clarke et al., 2006). Fish assemblages were also compared with the three factors using PCO and PERMANOVA with crossed design. In this PCO, the vectors of fish species correlated >0.4 were overlapped for comparison.
PERMANOVA analysis with crossed design, based on Euclidean distances was applied to the data on fish captures per day, after the assumptions of homogeneity of variances were not fulfilled even with logarithmic transformation.
The non-parametric Mann–Whitney test was used to assess the differences in the size of the taxa between seasons, as some of the data groups were not homoscedastic (even with logarithmic transformation).
All analyses were performed using the software PRIMER-E™ v. 6.1.11, PERMANOVA+ v. 1.0.1 and IBM SPSS™ Statistics v. 26.0.
Results
Water Column Characterization
The water column depth in the mangrove stand ranged between 0.22 and 1.90 m. As expected, the amplitude between low and high tide for spring tides was larger than that of neap tides (Supplementary Figure 1). The sandbank that emerged during the dry season affected the water column parameters, as the average low tide depth (0.63 m) was not as low as in rainy season (0.47 m). In fact, the opposite was expected, where the depth of the water column would be lower in the dry season. The water column depth reached the lowest values in the rainy season during spring tide event when there was no obstruction of water flow to the sea (by the sandbank) resulting in a very low tide with only a few centimeters of water in the lower part of the mangrove stand. The spring tide in the dry season did not reach this low water level due to the presence of the sandbank.
The salinity of the mangrove creek (0.0–30.0 PSU) increased closer to the sea. The stratification of the water column, interpreted by the difference between the high tide top and bottom values, confirmed the expectation that salinity would be higher at the bottom (Supplementary Figure 2). The observed temperature (21.8–32.2°C) generally increased with proximity to the sea and the patterns observed were similar to those of salinity, mainly because lower salinities are associated with lower temperatures of water descending from upstream.
The intrusion of saltwater is more intense during spring tides, explaining the higher temperatures registered in this situation at the upper and middle sites (Supplementary Figure 3). In terms of water column stratification, the expected tendency of lower temperature and salinity in the upper layer, and higher temperature and salinity in the bottom layers was evident. Temperature was generally higher during the dry season.
The PCO of the environmental data (Figure 2) presents the distribution of points according to the similarity of the environmental parameters. This graph segregates three major groups of points: (1) rainy season; (2) dry season, upper zone; and (3) dry season lower zone. The group (1), rainy season, is characterized by the lowest values of depth, salinity, and temperature, and within this group, the spring tide differs from the neap tide by having the lowest values. The group (3), dry season, lower zone, presents the highest values of depth, salinity, and temperature. Lastly, the group (2) is characterized by being in the middle of the gradient in the x axis of the PCO and by the low pH values, compared with the other two groups. The points of the rainy season are separated in the same group by tide (neap or spring). On the other hand, the points of the dry season are not separated by tide, but by zone, forming distinct groups in the PCO.
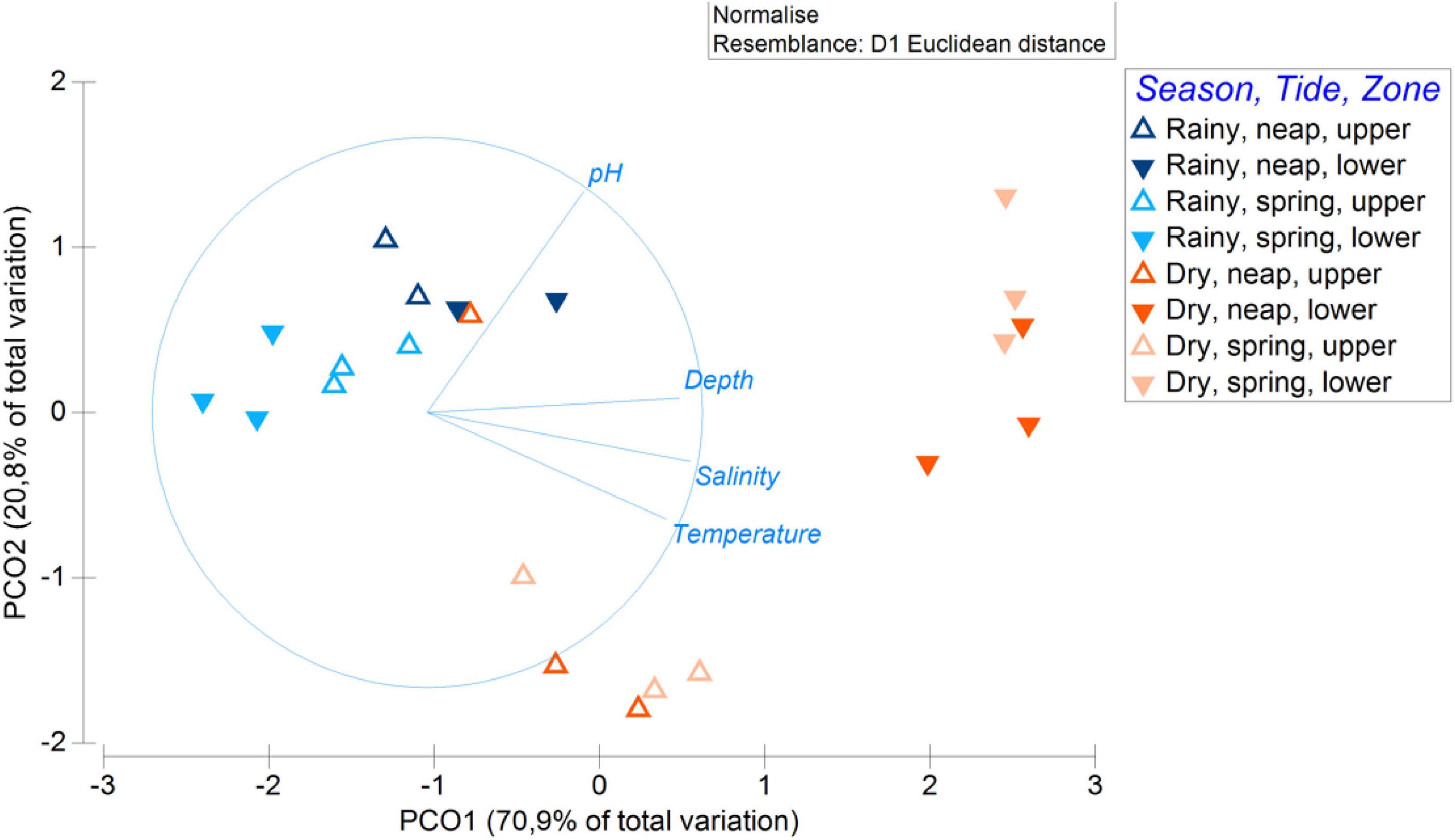
Figure 2. Principal coordinates ordination (PCO) plot of the environmental data with differentiated points according to season (rainy and dry), tide (spring and neap), and mangrove zone (upper and lower). Environmental vectors inside the circle represent the gradients that characterize the points distribution.
Fish Communities
A total of 772 individuals of at least 14 species were collected in the Praia Salgada mangrove stand (Table 1). The Atlantic Mudskipper Periophthalmus barbarus was very abundant on the banks and margins of the mangrove, however, they were never caught during the sampling due to their ability to avoid the nets.
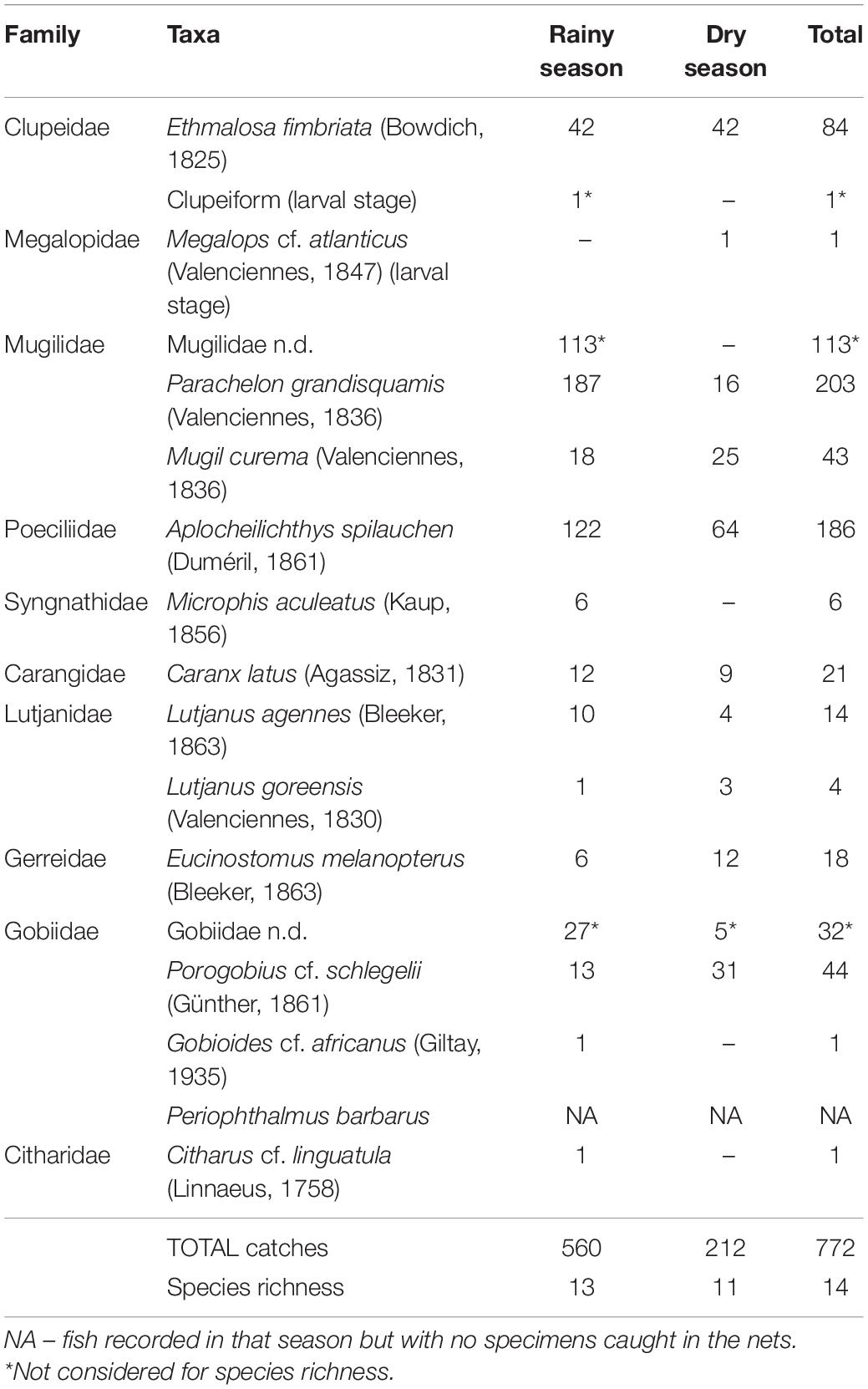
Table 1. Fish taxa, respective number of specimens caught, and species richness (number of species) according to the season.
During this field survey, two species of mullets (Mugil curema and Parachelon grandisquamis) were caught in the Praia Salgada mangrove stand. The designation “Mugilidae n.d.” was used when the size of the individual was too small to determine the species. This occurred during one sampling event (26th November 2019), when 113 individuals of this family were caught simultaneously with an average length of only 1.48 cm.
Two species from the family Gobiidae were identified (Porogobius cf. schlegelii and Gobioides cf. africanus). When “Gobiidae n.d.” designation was used, these individuals may be one of these two species, or another. The diversity of this family and the lack of adequate descriptions for the area made it difficult to accurately identify some individuals. The probability of these individuals included as “Mugilidae n.d.” and “Gobiidae n.d.” belonging to the recorded species led to them being excluded for species richness calculations.
The diversity of the fish community was slightly higher in dry season (rainy season H′ = 1.86; dry season H′ = 1.96). Distribution of the individuals among the various species was more even in dry season (J′ = 0.82) than in rainy season (J′ = 0.72).
A total of seven fishing days occurred in the rainy season and eight in the dry season. The average number of fish caught per day was 80.0 for the rainy season and 26.5 for the dry season. The dry season included two fishing attempts where no catches were made. The average number of fish caught per day was higher in the rainy season and lower but more stable in the dry season (Figure 3). These results may be influenced by the shoaling behavior of some taxa such as the Mugilidae, Aplocheilichthys spilauchen and Ethmalosa fimbriata. However, these groups represented more than half of the catches, and performing this analysis excluding them was not deemed to be justified. To test the significance of differences in the catches between the seasons, tides, and mangrove zones, a three-factor PERMANOVA was performed. The results (Table 2) show a significant difference between season, and non-significance between zone and tide. The interactions of the factors were not significant.
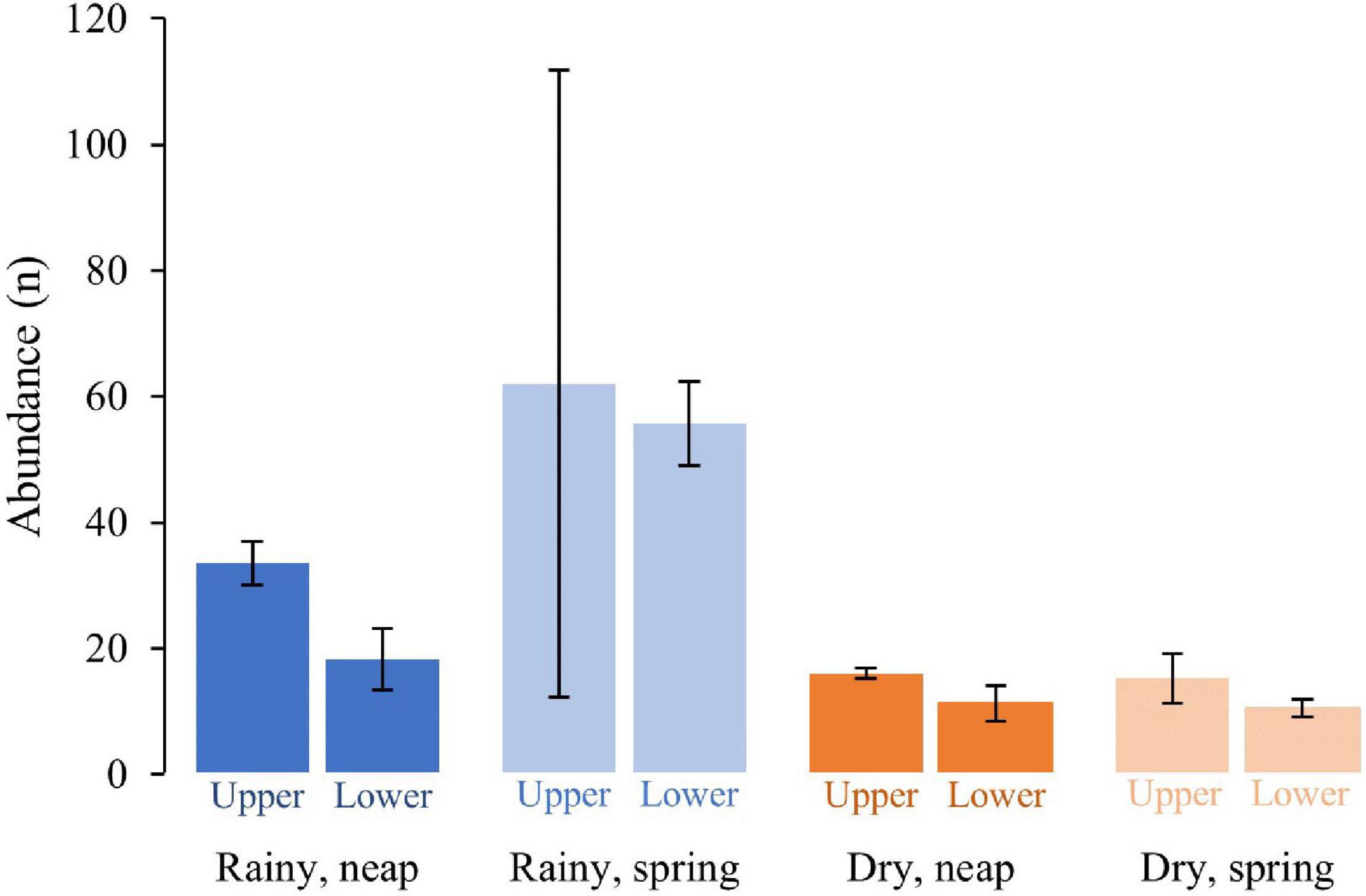
Figure 3. Average abundance of fish caught per day according to the season, tide, and zone. Error bars: ±SE.
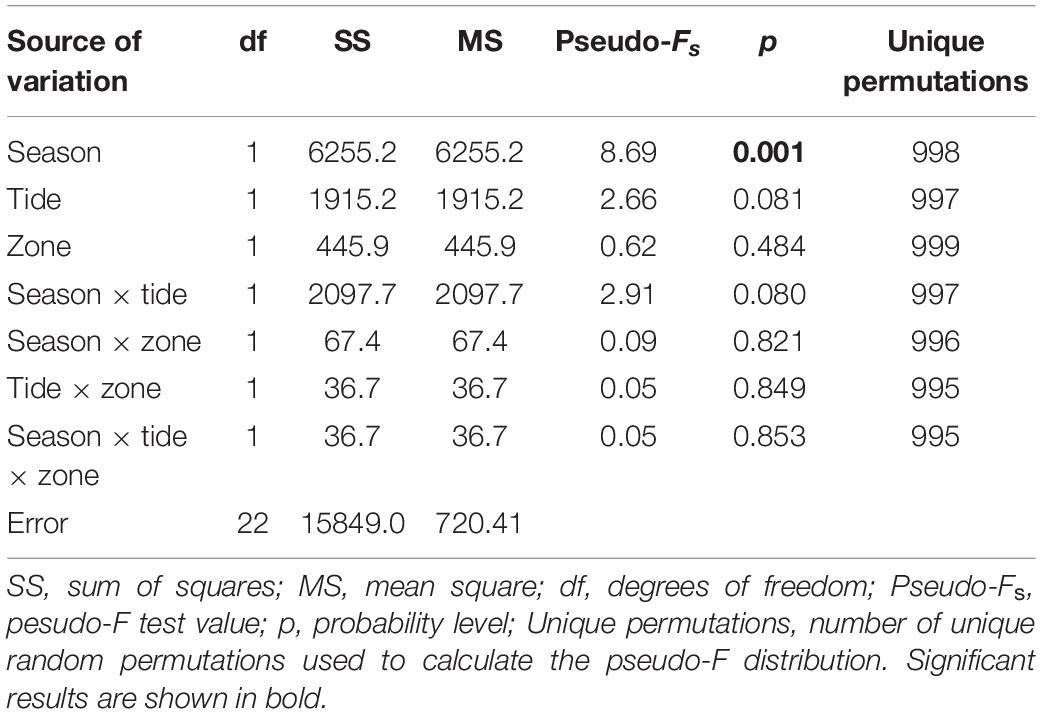
Table 2. PERMANOVA of the total fish catches per day (abundance) according to season, tide, and zone, based on non-transformed data.
The sizes of individuals tended to be higher during the dry season (Figure 4). Mann–Whitney U tests (at 95% confidence) were performed to assess the significance of the differences. Significant differences (U = 761.000, p < 0.001) were found for the Mugilidae as well as for A. spilauchen (U = 2612.000, p < 0.001) and the Gobiidae (U = 337.000, p < 0.001). Even though the graphical results for Caranx latus (Figure 4) show a tendency for larger individuals during the dry season, the Mann–Whitney U test results did not reveal significant differences (U = 36.000, p = 0.219) due to the variable distribution of the data from the rainy season. E. fimbriata and Eucinostomus melanopterus did not reveal a tendency for larger individuals in the dry season, and the Mann–Whitney U results indicate non-significant differences between the seasons (E. fimbriata: U = 955.000, p = 0.514; E. melanopterus: U = 47.000, p = 0.335).
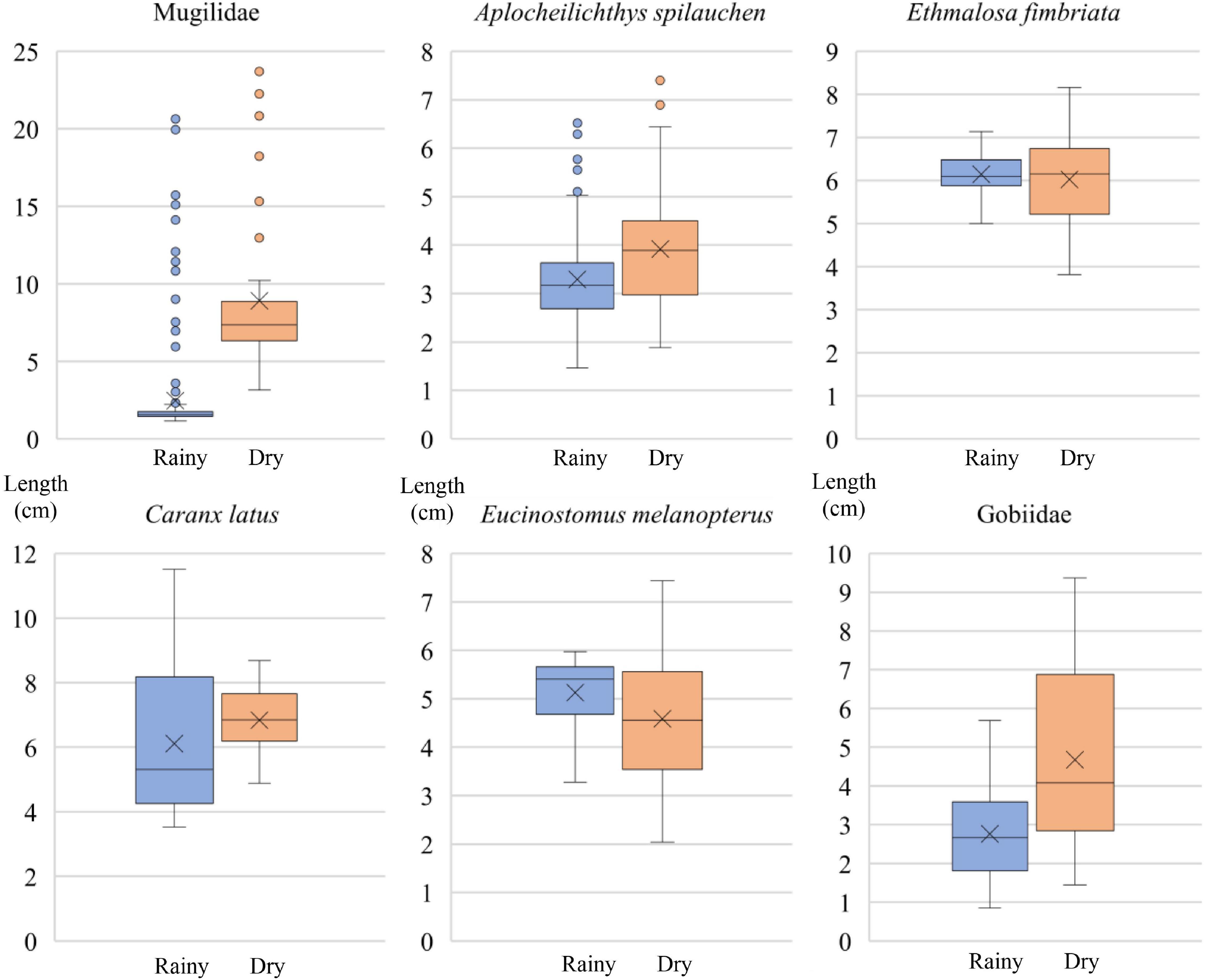
Figure 4. Fish lengths (TL), during the rainy and dry seasons. The boxplots represent the four quartiles of the data and the “X” the mean. The outliers represent a data point that exceeds 1.5 times the interquartile range (IQR) from the top or bottom of the box.
The other fish species (Lutjanus agennes, Lutjanus goreensis, Microphis aculeatus, Citharus linguatula, Clupeiforme larva, and Megalops atlanticus) were not analyzed for length according to season because their abundance was considered too low (14). The average length of individuals of Mugilidae, considering the separate groups of P. grandisquamis (rainy = 2.966 cm; dry = 12.515 cm), M. curema (rainy = 3.406 cm; dry = 6.603 cm), and Mugilidae n.d. (rainy = 1.484 cm; dry = NA) shows the same pattern of higher lengths in the rainy season. The Mugilidae boxplot is therefore presented with the lengths combined for all groups to synthesize the results. The same situation was apparent for the Gobiidae, where the pattern of higher average length in the dry season was present (Gobiidae n.d.: rainy = 2.196 cm, dry = 2.239 cm; P. cf. schlegelii: rainy = 3.706 cm, dry = 5.064 cm; and G. cf. africanus: rainy = 5.692 cm, dry = NA). However, when looking at the average length of Gobiidae n.d., similar lengths are apparent due to the difficulty in determining the species in these smaller individuals (until approximately 2.2 cm); a situation that occurred in the rainy and dry season with this family.
The Lutjanidae family were present on 14 occasions by specimens of L. agennes and 4 by L. goreensis. L. agennes were of an average length of 14.5 cm with minimum and maximum values of 2.01 and 43.7 cm, respectively. L. goreensis showed less variability values with an average of 13.3 cm and minimum and maximum values of 11.3 and 16.5 cm, respectively.
The distribution of the fish community in relation to location within the mangrove stand (Figure 5) reveals that the preference of the species does not varied between the studied seasons. The Mugilidae species, C. latus, and E. melanopterus, showed no significant preference between the upper and lower locations in the mangrove creek. Gobiidae and A. spilauchen populations seemed to prefer the upper, and E. fimbriata the lower mangrove creek.
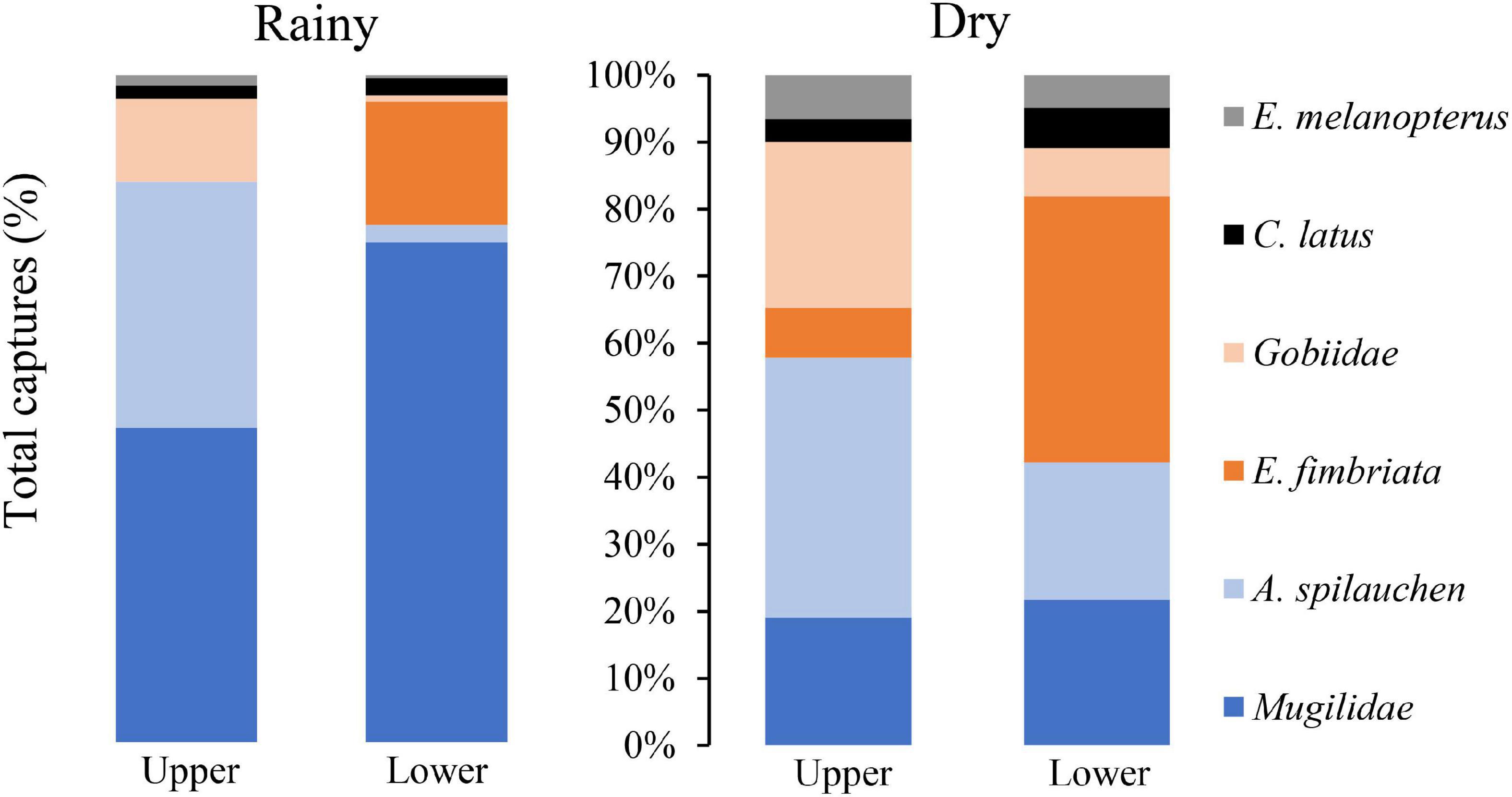
Figure 5. Distribution of the fish taxa (%) according to mangrove location (upper or lower) in the rainy and dry season.
The PERMANOVA results (Table 3) showed statistical differences in the fish community composition between season and zone, which are visible in the PCO plot (Figure 6), separating samples from rainy and dry season and upper and lower location. The results also showed a significant interaction between tide and zone, suggesting that the differences in the zone depend on the tide. Samples from each season clustered according to their species composition, where P. grandisquamis and M. aculeatus were associated with the rainy season samples (Figure 6), characterized by lower salinity, depth and temperature. On the other hand, E. fimbriata was associated with the lower zone of the mangrove stand (Figure 6), occurring only from 1.6 PSU or higher salinities in this study. M. aculeatus was recorded six times and only in the rainy season under low salinity conditions, with an average length and standard error of 9.55 and 1.67 cm, respectively. P. schlegelii and A. spilauchen were associated with the upper mangrove zone samples (Figure 6).
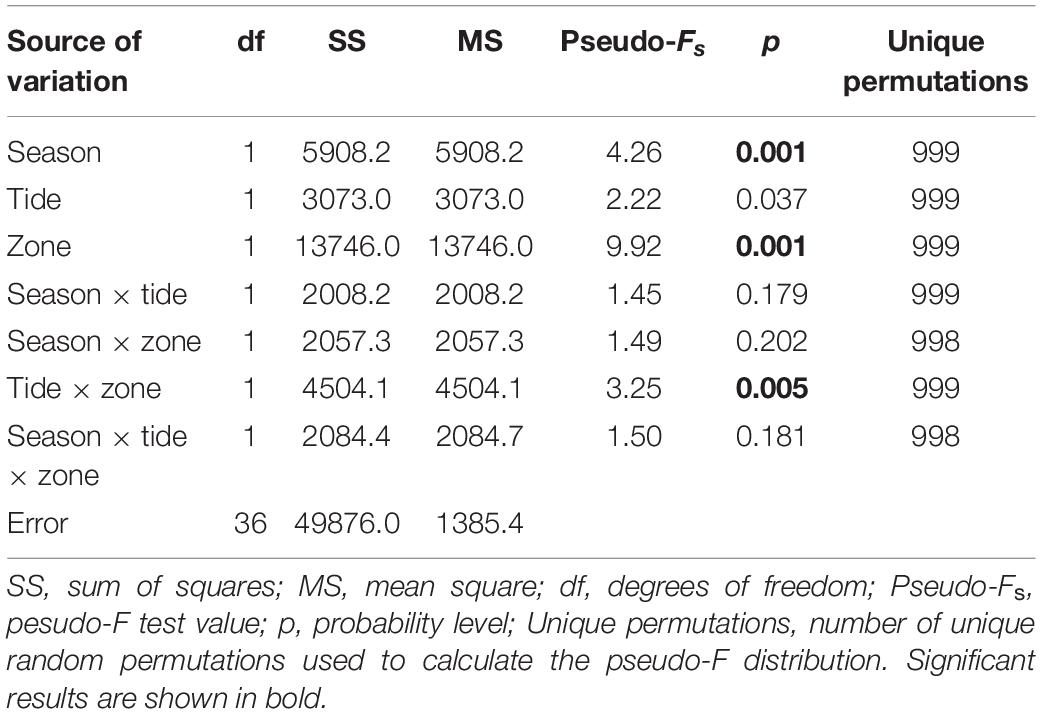
Table 3. PERMANOVA of the fish community composition according to season, tide, and zone, based on logarithmically transformed data.
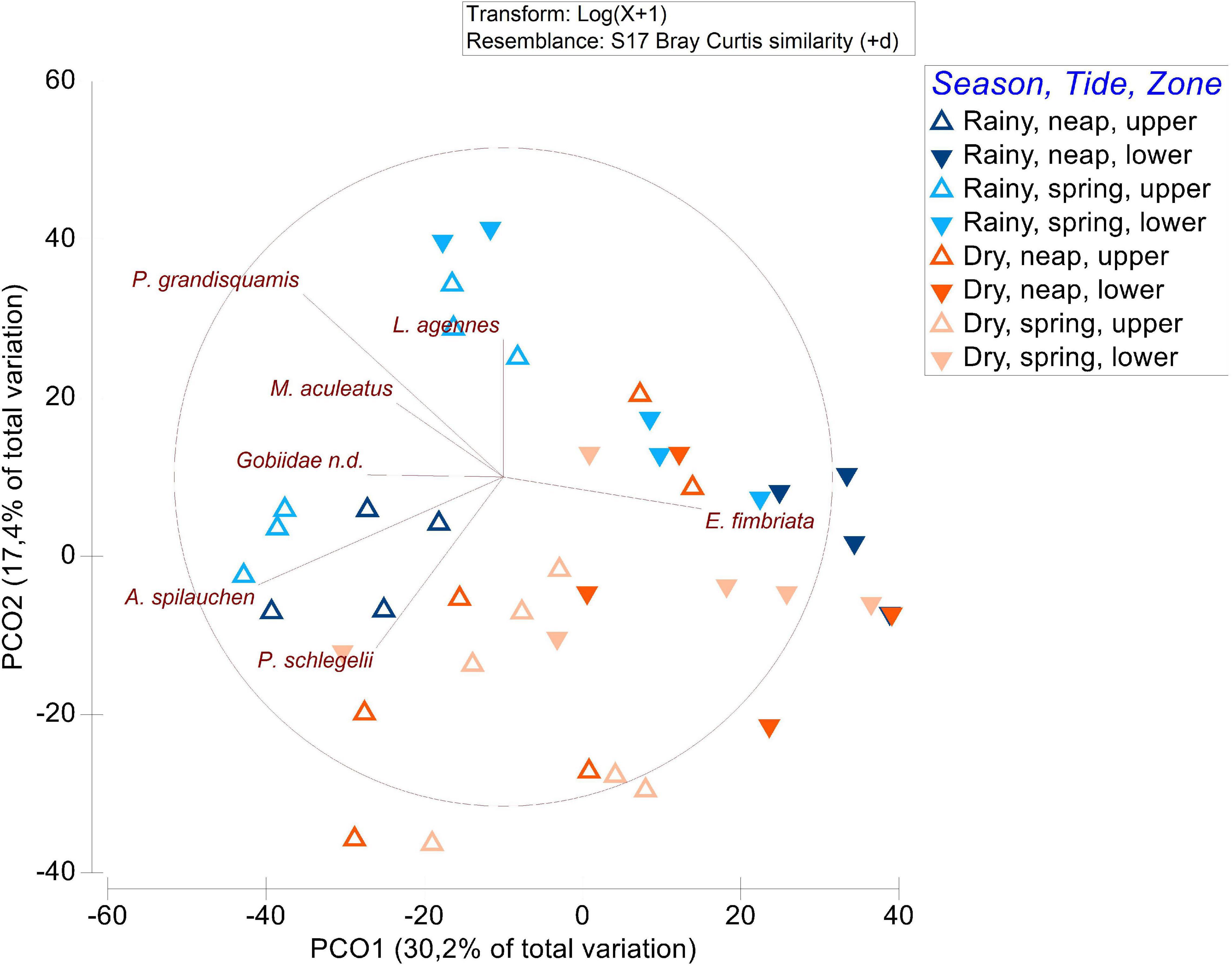
Figure 6. Principal coordinates ordination (PCO) plot of the fish assemblages data with differentiated points according to season (rainy and dry), tide (spring and neap), and mangrove zone (upper and lower). Vectors inside the circles represent the species that characterize the distribution of the points.
Discussion
Water Mass Characterization
The water mass parameters found in this study were as to be expected with regards to patterns of the gradients with proximity to the sea, the influence of irradiation, rains, and interaction between fresh and saltwater. The more intense water column stratification observed in the dry season could be explained by the increase in radiation at this time due to less cloudy skies, less precipitation, more stagnation of water due to the sandbank, and less flow from the river upstream. There are two published works with water mass parameter measurements for São Tomé and Príncipe, respectively (Pisoni et al., 2015; Herrero-Barrencua et al., 2017). They were developed in September–December of different years, including only the rainy season. The characteristics of the top and bottom layers of the water column were different. However, limited comparison is possible since the water mass characteristics of a given estuary are mainly related to the specific geomorphology and characteristics of the basin. Moreover, these studies did not distinguish between measurements taken at high, low, neap, and spring tide. Despite these differences, the values reported previously are within the same range as those reported in this study, and the variation patterns are similar.
The saline wedge is the result of the lack of mixing between saltwater from the ocean and fresh water from the river that creates a line between the two and represents the limit of saltwater intrusion. This happens due to the differences in density and temperature of the waters that do not mix due to low tidal motion. This situation was detected during the rainy season neap tide, where the salinity wedge is between the middle and lower points of the mangrove stand. In all other situations the salinity wedge was above the sampling points.
The water column depth results show the expected influence of the tide on the water dynamics in the mangrove system, with increasing and decreasing water level with tidal changes. However, during the dry season, the presence of the sand bank affected these water exchanges and, consequently, the biological communities of the mangrove system. The dry season did not reveal differences between tides, as the tidal effect was heavily reduced by the sand bank at the entrance of the mangrove creek. On the other hand, the points of the dry season were strongly separated by zone, also explained by the sandbank that blocked the exchanges of water between the creek and the sea and created a more lagoon-like environment.
The obstruction of the sand bank raised the water level inside the mangrove stand during the dry season, decreased the water exchange, and enlarged the mangrove creek. The depth was always greater for the three lowest sampling points (middle, lower, and outer) during the dry season at low spring tides, with tidal differences of approximately 19 cm in the middle site and 43 cm in lower and outer sites. These differences could have led to variations in the effectiveness of sampling and explain why the average number of fish caught per day was 80 for the rainy season and only 26.5 for the dry season.
The water column depth seemed to influence fish catches, with the shallowest depth associated with the greatest abundance of fish (spring tide during the rainy season). In the dry season, when the low tides were identical between spring and neap tides, the abundance of fish caught was the same. The abundance of fish during neap tides in the rainy season had intermediate abundance values, being coincident with also intermediate values for the water column depth. However, the data obtained are not strong enough to assess with certainty the factors that most contribute to the variation in fish abundance, especially in view of the effects of the variable environmental conditions on sampling effectiveness.
Fish Composition and Abundance
The total of 772 individuals of fish caught in the Praia Salgada mangrove stand comprised at least 14 different species. The majority of these species have already been reported from the country’s mangrove systems (Pisoni et al., 2015; Félix et al., 2017; Herrero-Barrencua et al., 2017; Hauron et al., 2018), except for E. fimbriata, M. curema, G. cf. africanus, and Citharus cf. linguatula. The genus Caranx had already been reported for São Tomé Island (Pisoni et al., 2015; Félix et al., 2017), however, the present study is the first to identify a species of this genus in a mangrove system.
The recorded species richness is similar to previous works to other small mangroves of the country: 13 in Praia das Conchas, São Tomé (Félix et al., 2017); 13 in Praia Salgada (Herrero-Barrencua et al., 2017); 11 in Praia Caixão (Herrero-Barrencua et al., 2017); and 10 in Praia Grande (Herrero-Barrencua et al., 2017). However, the species richness recorded in this work is smaller compared to Malanza, the most extensive mangrove system in the country, where were recorded 21 species (Félix et al., 2017).
The species richness was slightly lower in the dry season than the rainy season, which may be related to the abundance of fish caught being lower in the dry season. From the 14 different species reported, four are new records for the island mangrove system (E. fimbriata, M. curema, Gobioides africanus, and C. linguatula). The abundance of E. fimbriata and M. curema was significant, with total captures of 11 and 6%, respectively, revealing the common use of the mangrove system by this species. C. linguatula, caught only once, has been reported for the country (Krakstad et al., 2010) and can occur widely in tropical estuaries (Munroe, 2016).
Despite the species richness being similar to the other studies, this is not based on precisely the same species assemblages. For example, Elops senegalensis, Plectorhinchus macrolepis, Galeoides decadactylus, Monodactylus sebae, and Bostrychus africanus were all reported in the other two works (Félix et al., 2017; Herrero-Barrencua et al., 2017) but not in this survey.
Although Praia Salgada constitutes a small mangrove, the diversity and evenness of the fish community revealed to be relatively similar to other studies in more extensive mangroves the Gulf of Guinea (Aheto et al., 2014; Okyere, 2018).
The distribution of the fish species according to the different factors was shown to be statistically significant between season and zone. The non-significant effect of the factor tide alone is probably influenced by presence of the sand bank that modified the water dynamics of the mangrove system between the two seasons and attenuated the tide effect inside the mangrove creek. However, the interaction between tide and zone was significant, indicating that the effect of the tide might be relevant to fish distribution depending on the mangrove zone.
The lack of connectivity imposed by the sand bank during the dry season restricted the entry and exit of fish with the sea. In a small mangrove such as Praia Salgada, the connection to the sea probably has major importance to the fish community and the emergence of this barrier presumably affected the fish diversity and abundace. Season was the factor with more statistical significance on the captures per day with lower captures in dry season, and the sand bank occurred in the dry season, highlighting this possible effect.
All the reported species are found in brackish systems, some living in these environments permanently (Lévêque et al., 1990b) while others come to feed or seek refuge in their juvenile stages (FAO, 2016b; Ferraris and Smith, 2016). Although some of the species recorded in very low salinity situations are marine, the majority are juveniles. Juvenile fish tend to be more tolerant to low salinity waters (Bæuf and Payan, 2001).
The most important factor affecting the fish population in a mangrove swamp is often salinity (Wright, 1986). Salinity affects the distribution patterns and survival of fishes in estuaries, but may even affect metabolic processes (Nagelkerken et al., 2008). The species most commonly ascribed to freshwater, like A. spilauchen and the Gobiidae, preferred the upper part of the mangrove creek, while E. fimbriata preferred the lower location near the sea and more salty waters. The species that were found to be more common on the upper part of the mangrove creek could be classified as residents (Wright, 1986; Félix et al., 2017).
Most of the fish caught in the Praia Salgada mangrove system are of commercial importance. E. fimbriata, P. grandisquamis, M. curema, C. latus, L. agennes, L. goreensis, and E. melanopterus have all been reported as food fish resources for the country (Horemans et al., 1994; Afonso et al., 1999; Wirtz et al., 2007; Krakstad et al., 2010; Direção das Pescas, 2015; OMALI, 2019). This list excludes some species of freshwater fish and those species that are too small to be important for consumption such as A. spilauchen, M. aculeatus, P. schlegelii, and G. africanus. C. linguatula and M. atlanticus are reported as fish resources in the Eastern Central Atlantic (Ferraris and Smith, 2016; Munroe, 2016), but are not explicitly found in references for São Tomé and Príncipe.
A single M. atlanticus leptocephalus larva was captured in the Praia Salgada mangrove system. This amphi-Atlantic species is classified as vulnerable in the IUCN Red List (Adams et al., 2016) and existing records from mangrove systems only include adults in Malanza, São Tomé (Pisoni et al., 2015; Félix et al., 2017). This species spawns offshore and the leptocephalus larva, when ready, migrate to shallow coastal waters and lagoons to undergo metamorphosis (Taylor et al., 2011). The turbid low-oxygen waters in mangrove marshes offer protection to the young. The occurrence of this larval phase within the mangrove systems has been reported by others (e.g., Harrington, 1966), although these are most commonly juveniles, leading to the conclusion that this record from the current study was an isolated individual about to begin metamorphosis. Nevertheless, is an important record because the species has not yet been identified from the mangroves of Príncipe Island.
Fish Size and Life Stage
The Bonga shad E. fimbriata matures at 22.0 cm in Ghanaian waters according to Blay and Eyeson (1982). Two other works indicate that size at maturity is between 12.0 and 17.5 cm, with larger sizes for females (Facade and Olaniyan, 1972; Faye et al., 2014). Although the values in the literature vary, the individuals caught in this study all belong to the juvenile phase, with sizes from 3.8 to 8.2 cm.
The size at maturity for the mugilid P. grandisquamis is 15.0 cm, and of M. curema 19.7 cm, according to FishBase (Froese and Pauly, 2019), and 18–20.8 cm according to Aguirre and Gallardo-Cabello (2004). The sizes of specimens from the current study averaged between 2.5 and 8.9 cm, with standard deviations of 8.9 and 4.9 cm for the rainy and dry season, respectively. All captured individuals of the family Mugilidae were juveniles, except for a few outliers.
Caranx latus was recorded in this study with a maximum length of 11.5 cm. The species becomes mature at a length of 37.0 cm (Froese and Pauly, 2019), indicating that all recorded individuals were juveniles. The same applied to E. melanopterus, with a maximum length recorded of 7.4 cm and a size at maturity of 7.5 cm (Ramos et al., 2012), and for C. linguatuta, which the only specimen captured being 6.5 cm in length while the size at maturity of this species is reported at 14.5 cm (Cengiz et al., 2014).
Lutjanus agennes was recorded on 14 occasions in this study. The size at first maturity from literature is uncertain. However, the variability in the length recorded ranging from 2.0 to 43.7 cm with the average of 14.5 cm, indicate that the specimens caught were mainly juveniles, with few adults. L. goreensis occurred four times, and all were juveniles with an average length of 13.3 cm. This species is recorded with mean size at first sexual maturity at around 34.4 cm (Fakoya and Anetekha, 2019).
Aplocheilichthys spilauchen is a non-migratory brackish water species, occurring in swamps, river mouths, lagoons, and mangrove swamps (Wildekamp et al., 1986; Lévêque et al., 1990b). The length distribution of the specimens collected is similar to that reported by Okyere (2012), suggesting that the sampling included juveniles and adults. Immature individuals made up 9.2% of the total population in the study by Okyere (2012).
After analyzing the size at first maturity of the species with commercial interest, it was determined that they were all recorded exclusively as juvenile stages within the mangrove system, except for some adults of L. agennes and the larva of M. atlanticus. This strongly suggests a nursery role for the Praia Salgada mangrove system, where early stages of marine species can find food and shelter from predators.
Conclusion
The present study of the fish assemblages of the Praia Salgada mangrove system provides a fish species list for this brackish system, a description of water column parameters, and patterns of distribution of the fish community along the mangrove creek, in the context of variation in season between October and February, tides and mangrove zone.
Although Praia Salgada is a small mangrove system, its importance to fish recruitment is clearly suggested by the results of this study. After analyzing the size at first maturity of the species with commercial interest, it was observed that the majority of fish collected in the system were juveniles. This suggests a nursery role for the Praia Salgada mangrove system, where early stages of marine species can find food, and shelter from predators. The accumulation of sediments forming a sandbank in different sampling seasons constrained the water exchange between the creek and the sea at certain times of the year, consequently affecting fish movements. With this constraint, a decrease in fish catches was observed. The distribution of fish in the mangrove creek varied significantly according to the season and mangrove zone. The preservation of this habitat is vital to maintain its benefits to local fisheries, as well as the other environmental services it provides.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics Statement
Ethical review and approval were not required for the animal study since the work did not involve animal experimentation and did not compromise animal welfare. The fieldwork involving fish was simple and returned the vast majority of individuals to the environment without harming them, being the procedure quick and simple. A small portion of the fish was reserved to be integrated into the first natural history collection of Príncipe Island.
Author Contributions
MC was responsible for the work and took lead in all phases. JP contributed to the conception, progress of the work, and guidance, as well as data analysis and discussion. AA, HL, JA, SB, and VM-A all took part in the sampling, laboratory procedures, and analyses of data. All authors have contributed to the main components of the work.
Funding
This study was financed by AKDN – Fundação Aga Khan and FCT – Fundação para a Ciência e a Tecnologia IP, in the context of the research project COBIONET. This publication was also funded by the European Union’s Horizon 2020 Research and Innovation Programme under grant agreement N810139: Project Portugal Twinning for Innovation and Excellence in Marine Science and Earth Observation – PORTWIMS.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We acknowledge all the help provided by those that assisted during sampling work in the mangrove system, namely Ciciliano Bom Jesus, Jony Moreno Aurélio, and Elimesio do Rosário. We are also thankful to Pedro Ré, from Faculdade de Ciências da Universidade de Lisboa, for assisting with the fish larvae identification.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2021.721692/full#supplementary-material
Supplementary Figure 1 | Average water column depth at the four sampling sites in the mangrove system (upper, middle, lower, and outer) during the rainy and dry seasons and spring or neap tides. The points represent the average depth over the 3 days measured for each tidal situation and respective standard error.
Supplementary Figure 2 | Salinity at the four study sites in the mangrove system (upper, middle, lower, and outer) during the rainy and dry seasons and spring or neap tides. The points represent the average salinity over the 3 days measured for each tidal situation and respective standard error.
Supplementary Figure 3 | Water temperature at the four study points in the mangrove system (upper, middle, lower, and outer) during the rainy and dry seasons and spring or neap tides. The points represent the average temperature over the 3 days measured for each tidal situation and respective standard error.
References
Adams, A., Guindon, K., Horodysky, A., MacDonald, T., McBride, R., Shenker, J., et al. (2016). Megalops cyprinoides. The IUCN Red List of Threatened Species 2016: e.T166868A46642796. Available online at: https://dx.doi.org/10.2305/IUCN.UK.2016-3.RLTS.T166868A46642796.en (accessed November 23, 2021).
Afonso, F. (2019). A Importância dos Mangais de São Tomé?: Perceções e Serviços Ecossistémicos. Master’s thesis. Lisbon: University of Lisbon.
Afonso, P. F., Porteiro, F. M., Santos, R. S., Barreiros, J. P., Worms, J., and Wirtz, P. (1999). Coastal marine fishes of São Tomé island (Gulf of Guinea). Arquipél. Life Mar. Sci. 17A, 65–92.
Aguirre, A. L. I., and Gallardo-Cabello, M. (2004). Reproduction of Mugil cephalus and M. curema (Pisces: Mugilidae) from a coastal lagoon in the Gulf of Mexico. Bull. Mar. Sci. 75, 37–49.
Aheto, D. W., Okyere, I., Asare, N. K., Dzakpasu, M. F., Wemegah, Y., Tawiah, P., et al. (2014). A survey of the benthic macrofauna and fish species assemblages in a mangrove habitat in Ghana. West Afr. J. Appl. Ecol. 22, 1–15.
Blay, J. Jr., and Eyeson, K. N. (1982). Observations on the reproductive biology of the shad, Ethmalosa fimbriata (Bowdich), in the coastal waters of Cape Coast, Ghana. J. Fish Biol. 21, 485–496. doi: 10.1111/j.1095-8649.1982.tb02854.x
Bæuf, G., and Payan, P. (2001). How should salinity influence fish growth? Comp. Biochem. Physiol. C Toxicol. Pharmacol. 130, 411–423. doi: 10.1016/s1532-0456(01)00268-x
Bonfim, F., and Carvalho, S. (2009). Fourth National Report on the Biodiversity (1st Draft). República Democráática de São Tomé e Príncipe. São Tomé: Ministry of Natural Resources, Energy and Environment Directorate, General of the Environment.
Cengiz, Ö., Ismen, A., and Ozekinci, U. (2014). Reproductive biology of the spotted flounder, Citharus Linguatula (Actinopterygii: Pleuronectiformes: Citharidae), from Saros Bay (Northern Aegean Sea, Turkey). Acta Ichthyol. Piscatoria 44, 123–129. doi: 10.3750/aip2014.44.2.06
Chou, S. C., de Arruda Lyra, A., Gomes, J. L., Rodriguez, D. A., Alves Martins, M., Costa Resende, N., et al. (2020). Downscaling projections of climate change in Sao Tome and Príncipe Islands, Africa. Clim. Dyn. 54, 4021–4042. doi: 10.1007/s00382-020-05212-7
Clarke, K. R., Somerfield, P. J., and Chapman, M. G. (2006). On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray–Curtis coefficient for denuded assemblages. J. Exp. Mar. Biol. Ecol. 330, 55–80. doi: 10.1016/j.jembe.2005.12.017
Direção das Pescas (2015). Relatório do Estudo de Seguimento de Espécies Demersais da Zona Costeira de S. Tomé e Príncipe. São Tomé: Fisheries Department, Ministry of Economy and International Co-Operation.
Facade, S. O., and Olaniyan, C. I. O. (1972). The biology of the West African shad Ethmalosa fimbriata (Bowdich) in the Lagos Lagoon, Nigeria. J. Fish Biol. 4, 519–533. doi: 10.1111/j.1095-8649.1972.tb05699.x
Fakoya, K. A., and Anetekha, M. A. (2019). Macroscopic gonad staging and reproductive seasonality in the Gorean snapper, Lutjanus goreensis a gonochoristic West African Lutjanid. West Afr. J. Appl. Ecol. 27, 1–22.
FAO (2016a). The Living Marine Resources of the Eastern Central Atlantic: Volume 4: Bony Fishes Part 2 (Perciformes to Tetradontiformes) and Sea Turtles. FAO Species Identification Guide for Fisheries Purposes. Rome: FAO, 2343–3124.
FAO (2016b). The Living Marine Resources of the Eastern Central Atlantic. Volume. 3: Bony Fishes Part 1 (Elopiformes to Scorpaeniformes). FAO Species Identification Guide for Fisheries Purposes. Rome: FAO, 1511–2350.
Faye, A., Sarr, A., Thiaw, M., Ndiaye, I., Fall, J., Diouf, M., et al. (2014). Reproductive biology of Ethmalosa fimbriata (Bowdich) in Senegalese coastal waters. J. Biol. Life Sci. 5, 57–71. doi: 10.5296/jbls.v5i1.4903
Félix, P. M., Chainho, P. A., Lima, R. F. B., Costa, J. L., Almeida, A. J., Domingos, I., et al. (2017). Mangrove fish of Sao Tome Island (Gulf of Guinea): new occurrences and habitat usage. Mar. Freshw. Res. 68, 123–130. doi: 10.1071/MF15392
Ferraris, C. J. Jr., and Smith, D. G. (2016). “Megalopidae,” in The Living Marine Resources of the Eastern Central Atlantic. Vol. 3: Bony Fishes Part 1 (Elopiformes to Scorpaeniformes Turtles, eds K. E. Carpenter and N. De Angelis (Rome: FAO Species Identification Guide for Fishery Purposes, FAO), 1588–1589.
Guatier, D., and Hussenot, J. (2005). Les Mulets des Mers d’Éurope. Synthèse des Connaissances sur les Bases Biologiques e les Techniques D’aquaculture. Plouzane: Éditions Quae, 119.
Harrington, R. W. (1966). Changes through one year in the growth rates of tarpon, Megalops atlanticus Valenciennes, reared from mid-metamorphosis. Bull. Mar. Sci. 16, 863–883.
Hauron, R., Barrencua, A. H., and Abreu, A. D. (2018). “Mangrove habitats in São Tomé and Príncipe (Gulf of Guinea, Africa): conservation and management status,” in Threats to Mangrove Forests (Cham: Springer), 589–605. doi: 10.1007/978-3-319-73016-5_27
Herrero-Barrencua, A., Haroun, R., and Abreu, A. D. (2017). Caracterización Preliminar de los Manglares de la Isla de Príncipe (São Tomé e Príncipe). Las Palmas: IU-ECOAQUA, Universidad de Las Palmas de Gran Canaria, 70.
Horemans, B., Galléne, J., and Njock, J. (1994). Revista Sectorial da Pesca Artesanal a São Tomé e Príncipe. Programa de Desenvolvimento Integrado das Pescas Artesanais na África Ocidental (DIPA). São Tomé: Project DIPA, FAO.
Krakstad, J. O., Alvheim, O., Lopes, J., and Iwamoto, T. (2010). Surveys of the Fish Resources of São Tomé & Príncipe: Survey of the Demersal Resources (Issue May). Bergen: Institute of Marine Research.
Krebs, C. J. (1999). Ecological Methodology. Boston, MA: Addison-Welsey Educational Publishers, 620.
Lévêque, C., Paugy, D., and Teugels, G. G. (1990a). The Fresh and Brackish Water Fishes of West Africa, Vol. 1. Paris: Institut Français de Recherche Scientifique Pour le Developpement en Cooperation, 1–386.
Lévêque, C., Paugy, D., and Teugels, G. G. (1990b). The Fresh and Brackish Water Fishes of West Africa. Vol. 2. Paris: Institut Français de recherche scientifique pour le Developpement en cooperation, 389–902.
MEA (2005). Ecosystems and Human Well-being: Synthesis. Washington, DC: Millennium Ecosystem Assessment (MEA), Island Press.
Munroe, T. A. (2016). “Citharidae,” in The Living Marine Resources of the Eastern Central Atlantic. Vol. 4: Bony Fishes Part 2 (Perciformes to Tetradontiformes) and Sea Turtles, eds K. E. Carpenter and N. De Angelis (Rome: FAO Species Identification Guide for Fishery Purposes, FAO), 2952–2955.
Nagelkerken, I., Blaber, S. J. M., Bouillon, S., Green, P., Haywood, M., Kirton, L. G., et al. (2008). The habitat function of mangroves for terrestrial and marine fauna: a review. Aquat. Bot. 89, 155–185. doi: 10.1016/j.ejmech.2020.112957
Nagelkerken, I., Roberts, C. M., van der Velde, G., Dorenbosh, M., van Riel, M. C., Cocheret de la Morinièrel, E., et al. (2002). How important are mangroves and seagrass beds for coral-reef fish? The nursery hypothesis tested on an island scale. Mar. Ecol. Prog. Ser. 244, 299–305. doi: 10.3354/meps244299
NBSAP II (2015). National Biodiversity Strategy and Action Plan 2015–2020, National Commission for Monitoring and Evaluation, Ministry of Infrastructure. São Tomé: Natural Resources and Environment, 120.
Okyere, I. (2012). Some ambient environmental conditions, food and reproductive habits of the banded lampeye killifish Aplocheilichthys spilauchen in the Kakum Estuary Wetland, Ghana. Environ. Biol. Fish. 94, 639–647. doi: 10.1007/s10641-011-9970-4
Okyere, I. (2018). Influence of diurnal tides and other physico-chemical factors on the assemblage and diversity of fish species in River Pra Estuary, Ghana. Trop. Ecol. 59, 83–90.
OMALI (2019). Sumário de Atividades de Projeto e Resultados Preliminares – Março, 2019. Santo António City: Omali Vida Nón.
Pielou, E. C. (1966). The measurement of diversity in different types of biological collections. J. Theor. Biol. 13, 131–144. doi: 10.1016/0022-5193(66)90013-0
Pisoni, T., de Lima, R. F., Brito, A. C., Chainho, P., Félix, P. M., Caçador, I., et al. (2015). Planos de Gestão Participativa para Dois Sítios de Mangal na Ilha de S. Tomé: Praia das Conchas e Malanza. Abordagem Ecossistémica Integrada para a Conservação e Gestão da Biodiversidade na Zona Tampão dos Parques Naturais obô de São Tomé e Príncipe. São Tomé: ALISEI.
Ramos, J. A. A., Barletta, M., and Costa, M. F. (2012). Ingestion of nylon threads by gerreidae while using a tropical estuary as foraging grounds. Aquat. Biol. 17, 29–34. doi: 10.3354/ab00461
Taylor, S. M., Loew, E. R., and Grace, M. S. (2011). Developmental shifts in functional morphology of the retina in Atlantic tarpon, Megalops atlanticus (Elopomorpha: Teleostei) between four ecologically distinct life-history stages. Vis. Neurosci. 28, 309–323. doi: 10.1017/S0952523810000362
Tomlinson, P. (2016). The Botany of Mangroves, 2nd Edn. Cambridge: Cambridge University Press. doi: 10.1017/CBO9781139946575
UNEP (2010). Global Mangrove Extent Much Smaller than Previously Estimated, Thematic Focus: Ecosystem Management, Disaster and Conflicts, and Climate Change. Nairobi: United Nations Environmental Programme (UNEP).
Wildekamp, R. H., Romand, R., and Scheel, J. J. (1986). “Cyprinodontidae,” in Check-List of the Freshwater Fishes of Africa (CLOFFA), eds J. Daget, J.-P. Gosse, and D. F. E. Thys van den Audenaerde (Paris: ORSTOM), 165–276.
Wirtz, P., Ferreira, C. E. L., Floeter, S. R., Fricke, R., Gasparini, J. L., Iwamoto, T., et al. (2007). Coastal fishes of São Tomé and Príncipe islands, Gulf of Guinea (Eastern Atlantic Ocean) – an update. Zootaxa 1523, 1–48. doi: 10.11646/zootaxa.1523.1.1
Keywords: fish checklist, brackish system, season, São Tomé and Príncipe, West Africa
Citation: Cravo M, Almeida AJ, Lima H, Azevedo e Silva J, Bandeira S, Machava-António V and Paula J (2021) Fish Assemblages in a Small Mangrove System on Príncipe Island, Gulf of Guinea. Front. Mar. Sci. 8:721692. doi: 10.3389/fmars.2021.721692
Received: 07 June 2021; Accepted: 09 November 2021;
Published: 07 December 2021.
Edited by:
Janine Barbara Adams, Nelson Mandela University, South AfricaReviewed by:
Marina Dolbeth, University of Porto, PortugalIsaac Okyere, University of Cape Coast, Ghana
Copyright © 2021 Cravo, Almeida, Lima, Azevedo e Silva, Bandeira, Machava-António and Paula. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mariana Cravo, bWFyaWFuYW1hcnRpbnNjcmF2b0BnbWFpbC5jb20=