
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci. , 21 June 2021
Sec. Marine Fisheries, Aquaculture and Living Resources
Volume 8 - 2021 | https://doi.org/10.3389/fmars.2021.669176
This article is part of the Research Topic The Approach to Complex Systems in Fisheries View all 9 articles
 Enrique Morales-Bojórquez1
Enrique Morales-Bojórquez1 Manuel O. Nevárez-Martínez2*
Manuel O. Nevárez-Martínez2* Gabriela García-Alberto3Héctor Villalobos3Hugo Aguirre-Villaseñor4
Gabriela García-Alberto3Héctor Villalobos3Hugo Aguirre-Villaseñor4 Estefani Larios-Castro1Sergio S. González-Peláez1Dana I. Arizmendi-Rodríguez2María de los Ángeles Martínez-Zavala2
Estefani Larios-Castro1Sergio S. González-Peláez1Dana I. Arizmendi-Rodríguez2María de los Ángeles Martínez-Zavala2The first steps have been taken to analyze the effects of small pelagic harvests on the Gulf of California ecosystem. The primary goal of this study was to estimate the latitudinal differences in regionalized baselines of endangered and threatened marine species attracted to a purse seine in a Mexican fishery of small pelagic fish. We also analyzed the spatial patterns in fishing effort in the Gulf of California. Seven zones were analyzed in the Gulf of California, including the east and west coasts, and a total of 3,051 fishing sets were analyzed during January 2013 and July 2014; the data provided a comprehensive picture of the distribution of the fishing effort, small pelagic fishes harvested, and the presence of species attracted to the fishing sets. The region in the upper Gulf of California showed a low presence of individuals, and the east coast recorded more sightings than the west coast; consequently the fishing effort was mainly distributed in the east coast. The number of individuals for several species sighted and counted by fishing set was used for each zone in the Gulf of California, and a conservative baseline based on the Pennington estimator was computed. It provided an average value of endangered and threatened marine species attracted to a purse seine for each zone. The Pennington estimator is recommended due to the precision of the confidence intervals and the nature of the uncertainty in the data collection based on sightings.
The first Pacific sardine fishery (Sardinops sagax) in Mexico was established in the port of Ensenada, Baja California, in the late 1920s, and there were significant records of catches there since the late 1940s, as well as later in Isla Cedros, Baja California, and Bahía Magdalena, Baja California Sur (Pedrín and Ancheita, 1976; Lluch-Belda et al., 1986; Cisneros-Mata et al., 1995; Nevárez-Martínez et al., 2014). However, as a consequence of significant declines in Pacific sardine in the Mexican Pacific, and the discovery of a significant stock of Pacific sardine and other small pelagic species in the Gulf of California, this fishing activity also moved to the Gulf in the late 1960s, becoming the basis for what is now a small pelagic fishery whose base ports are in Guaymas and Yavaros, Sonora. These ports are now extensively developed, with important fleets and the most significant industrial plant in the country (Cisneros-Mata et al., 1995; Nevárez-Martínez et al., 2014). Although this is a multi-species fishery, Pacific sardine is on average the largest proportion of total catches (Nevárez-Martínez et al., 2001, 2014; Morales-Bojórquez and Nevárez-Martínez, 2005).
Small pelagic fish are commercially caught using large vessels (23–30 meters in length and 120–220 t ship’s hold) equipped with a purse seine, an auxiliary boat (skiff) and a maximum crew size of 10 (Nevárez-Martínez et al., 2014). Fishing activities take place over a period of 22–26 days, centered on the new moon. The fishing set begins by searching for the location of schools of fish. Once a school has been detected, the purse seine is set around it, the skiff is released, and the purse seine is quickly maneuvered to surround the school. The lower part of the net is closed, pulling the purse line through the lower rings of the net. Most of the net is then recovered; leaving a bag on one side of the ship, from which the catch is uploaded to the ship’s hold using a suction pump. The incidental catch in this fishery, although very low, is mostly composed of other fish species (e.g., wavyline grunt, rough triggerfish, Pacific sierra), but jumbo squid, cannonball jellyfish, and shrimp have also been found (Nevárez-Martínez et al., 2014). Larger animals, such as marlin or dolphins, can usually be released alive by lowering a section of the net or using a dip net, and birds are kept out of the fishing gear using a jet of water and by making a lot of noise.
It is important to analyze the structure of marine communities in order to understand the regional processes and the functioning of ecosystems. There is concern over the effect of fishing on benthic and demersal environments, mainly by shrimp trawl fisheries (Jennings and Kaiser, 1998; Hall, 1999; Gislason et al., 2000). Fishing has both direct (e.g., removal of individuals) and indirect (e.g., habitat modification) effects on ecosystems. These effects include the capture of non-target species, collectively known as bycatch, which includes species that are unwanted and thrown away (discards), and species that are retained and sold (byproducts). This issue is of particular concern in the small pelagic fishery in the Gulf of California, Mexico, given that the Marine Stewardship Council has certified this fishery as sustainable (Izquierdo et al., 2020). The interaction between marine fauna and this small pelagic fishery must therefore be evaluated to understand the effects of the harvest on the coastal marine ecosystem.
The large volume of bycatch in various fisheries has contributed to the widespread belief that this practice may be causing detrimental changes to ecosystems. The effects of global fisheries on bycatch species are little known, although some studies have documented changes in specific bycatch species (Casey and Myers, 1998; Pope et al., 2000). There are also reports of the maximum sustainable yield being reached in some species of the families Lutjanidae and Haemulidae, where juveniles are present in the bycatch of shrimp trawl fisheries (Espino-Barr et al., 1997). Løkkeborg and Robertson (2002) have reported the negative effects of longline fisheries on seabirds. Zador et al. (2008) and Kimberly et al. (2009) reported the mortality of albatrosses in an Alaskan trawl fishery and a demersal longline fishery, respectively. Darby and Dawson (2000) reported the bycatch of yellow-eyed penguins in gillnets in New Zealand. Best practice seabird bycatch mitigation has been proposed by Zador et al. (2008), Lescrauwaet et al. (2013), and Melvin et al. (2014). The bycatch for combined species has also been analyzed, including seabirds and turtles (Burgoni et al., 2008), seabirds and marine mammals (Norman, 2000; Majluf et al., 2002), and fish (Madrid-Vera et al., 2007).
Mortality due to bycatch is unavoidable for several types of marine fisheries. Dillingham and Fletcher (2008) estimated the likelihood of seabirds being killed by human activity, based on the annual growth rates of the populations, current status, and management objectives. Zydelis et al. (2009) evaluated the effects of additive mortality on bird populations, applying the potential biological removal (PBR) concept to three species with the most extensive bycatch information. PBR is a viable management tool, for example, PBR thresholds can be readily estimated to populations with limited demographic information, additionally, PBR estimates are easy to update once new data becomes available.
Sampling programs are required in the Gulf of California in order to obtain reliable estimates of the effects of the small pelagic fishery harvest on marine fauna. Estimates of discards are also needed to evaluate the impact of fishing on both non-commercial species and the ecosystem as a whole (Alverson et al., 1994; Madrid-Vera et al., 2007), as well as to assess the effectiveness of technical management measures aimed at reducing incidental mortality. In the Gulf of California, the first steps have been taken in the small pelagic fishery to analyze the effects of the harvest on the ecosystem, recognizing that the fishing gear (purse seine) risks incidentally catching, killing, or injuring non-target species. In response, the Mexican fleet has incorporated several actions to avoid mortality due to bycatch, including a water curtain to prevent birds from entering the purse seine, the release of small marine mammals or birds trapped by the purse seine, and best practices among the fleet and crew to carefully handle marine fauna, thereby improving the survival of non-target species. Thus, the primary goal of this study was to estimate the latitudinal differences in regionalized baselines of endangered and threatened marine species attracted to a purse seine in a Mexican fishery of small pelagic fish. We also analyzed the spatial patterns in fishing effort in the Gulf of California.
Many marine fauna are attracted to bait, offal, and discards from fishing vessels (Zador et al., 2008), and this feeding behavior often brings these species into dangerous contact with a variety of fisheries, because fishing vessels provide a potential food source (Kimberly et al., 2009). This phenomenon occurs during the fishing sets of the small pelagic fishery, where several species are attracted. Onboard observers therefore collected marine fauna data during 2013 and 2014. For each fishing trip, observers recorded basic information on the vessels, daily fishing activities (including fishing geographical position), and catch information, which was mainly the number of species retained, discarded, and live-released, including elasmobranch and teleost fishes and ecologically related species such as sea turtles, seabirds, and marine mammals. For this group of species, the observers recorded whether the individuals were released, and whether they were alive, dead, or in poor condition. Digital photographs were taken of individuals not immediately identified, to be determined later in the laboratory.
The study area was along the continental shelf of the east and west coasts of the Gulf of California. The Mexican National Fisheries Institute (MNFI), the Mexican agency for fisheries management, conducts fisheries acoustics surveys here in the Gulf of California, analyzing the most important zones, where the occurrence, abundance and recruitment of small pelagic species are traditionally observed. The sampled region was based on a spatial modification of the areas proposed by Nevárez-Martínez et al. (2001). The information was collected in these areas, and the marine stations were surveyed over 19 months from January 2013 to July 2014 on board commercial vessels using trained observers. The fishing sets were predominantly inshore. Data analysis was focused on endangered and threatened marine species that were attracted to the purse seine in the small pelagic fishery (hereafter called “individuals attracted”) in the Gulf of California, and the status of each species was defined according to Mexican laws1 and international organizations such as the International Union for Conservation of Nature (IUCN) and the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES).
The Gulf of California is a region of high biological productivity, which is affected by a combination of bottom topography and a high degree of wind-induced mixing and upwelling from strong predominantly northwesterly winds, particularly in the midriff islands region (Lanz et al., 2009). According to Lavín et al. (1997) the ocean circulation in the Gulf of California is determined mainly by the tide and winds; whilst the residual currents in the Gulf of California promote the net transport of nutrients. These oceanographic conditions are favorable for the existence of a variety of populations of small pelagic fishes (e.g., sardines, anchovies), mainly distributed on the east coast of the Gulf of California where their abundance is greater (Sonora and Sinaloa states). In comparison, the west coast of the Gulf of California includes a low abundance of small pelagic fishes (Baja California Peninsula). A regionalization was proposed by Nevárez-Martínez et al. (2001), based on the spatial distribution of the main species harvested in the Gulf of California (Pacific sardine Sardinops sagax, Pacific mackerel Scomber japonicus, thread herring Opisthonema libertate, and northern anchovy Engraulis mordax), thus classifying 11 subzones. This study is based on a slight modification of the regionalization initially proposed, such that the data obtained from the onboard fisheries observer program was according to the spatial coverage of the different species of small pelagic fishes. The zones were identified as follows:
Zone I: This zone is close to the Biosphere Reserve of the upper Gulf of California; where a coastal area is traditionally authorized for fishing activities, and some vessels harvest small pelagic species.
Zone II: This includes an extensive oceanic area where small pelagic species are occasionally harvested. Their boundaries are defined from the south limit of Zone I to the north of Isla Tiburón (Sonora state).
Zone III: This traditional fishing ground is very well-identified and close to the port of Guaymas, Sonora state, where an important fishing industry has been established since 1960.
Zone IV: This covers the southern region of Sonora state, which is also a productive fishing ground, and where Yavaros port is located.
Zone V: This is defined from the south boundary of Sonora state to the north area of Sinaloa state; in this zone two species of thread herring Opisthonema libertate and Cetengraulis mysticetus are the most important in the fishery.
Zone VI: This zone is identified as a spawning and nursery area for Pacific sardine Sardinops sagax and anchovy Engraulis mordax; the zone is characterized by the adjacent waters to the Isla Ángel de la Guarda (Baja California).
Zone VII: This is defined from the south boundary of Baja California state to the north area of Baja California Sur state. This zone traditionally exhibits a high interannual variability in fishing pressure, which depends on the spatial availability of small pelagic fish.
The data provided a comprehensive picture of the distribution of the fishing effort, small pelagic fishes harvested, and the presence of species attracted to the fishing sets. The number of individuals for several species sighted and counted by fishing set was thus used. Hsiang-Wen and Kwang-Ming (2010) and Guy et al. (2013) used this approach based on combined data. A monthly average value for species attracted was estimated for the different fishing areas defined by the MNFI. A basic assumption of the species-attracted data obtained by the onboard observers was that it had the same ratio of species attracted as the small pelagic fishery, because they were collected using the same boats and the same fishing gear used at the commercial fishery, hence their catchability and fishing power would be similar during 2013 and 2014. Given that each species identified in this study depended on the level of experience and training of the onboard observers, and given that the fishing maneuver takes place at night, when the small pelagic species are available to the fishing fleet, it was possible that the observers would miss, or fail to identify and count correctly, the number of species and organisms for each species; therefore, there is some degree of uncertainty linked to the attracted species data. Nevertheless, it was possible to estimate from the database the number and percentage of endangered and threatened marine fauna attracted, released and killed by fishing sets in the small pelagic fishery on the Gulf of California.
The Kappenman distribution estimated the average value (φ) of the species attracted, using the number of positive fishing sets according to the zones defined by the MNFI. This estimator computed the expected values of all observations, and if these were of the same magnitude as those computed by a kernel estimator, the procedure was concluded. Otherwise, the Kappenman estimator performed a replacement by recalculating the abundance of the unusually large fishing sets, estimating the average value based on a normal probability density function (Kappenman, 1999). This avoided the risk of an average value being biased by unusually large fishing sets. Pennington (1996) proposed a general model incorporating Δ-distribution and log-normal distribution into a new estimator which is useful for estimating the mean (δ) and the variance from highly skewed marine data. Pennington’s proposal is based on the product of two independent components: the probability of non-zero observations associated with the probability of positive highly abundant fishing sets (Ortiz et al., 2000). This general model is effective when the marine data shows a predominance of zero values, the presence of highly aggregated observations, or both (Pennington, 1983, 1986, 1996; Pennington and Stromme, 1998; Folmer and Pennington, 2000).
This is a generalized probabilistic density function supported by a Δ distribution and a log normal distribution. In this study, the Pennington estimator was used for computing the average number and variance of individuals attracted by a fishing set, and the estimators are described as follows:
and
where: ρ and θ are the minimum variance of unbiased estimates of the mean and its variance of the Δ-distribution, n is the number of fishing sets, δ is the number of positive fishing sets, zi = ln (yi), and s2 are the sample mean and variance of positive fishing sets logged values, y1 is a single (untransformed) fishing set when δ = 1, and Gδ(t) is a function of δ and t (e.g., t = s2/2) shown as:
The minimum variance unbiased estimator of the variance of the mean (varest(ρ)) is given by:
when δ = n, the estimator assumes a lognormal distribution.
The Kappenman estimator performs a replacement by recalculating the abundance of the unusually large fishing sets, estimating an average value based on a normal probability density function (Kappenman, 1999). The basic equations for computing average value from Kappenman distribution include the kernel estimator expressed as (Larios-Castro et al., 2021):
where and zi = lnyi, for i = 1,…n. The number h in the above equation is the value of h which maximizes:
When the statistical procedure concludes and the kernel estimator did not detect significant statistical differences between the unusually abundant fishing set and the expected values, then the analysis concludes by estimating the average value as . Otherwise, the unusually abundant fishing set or sets are estimated and replaced as ; consequently, the average value was estimated as:
Both statistical procedures are widely documented in the literature, and were solved here using the functions cpuekapp (Kappenman estimator and gap function) and deltadist (for Pennington estimator) contained in the “fishmethods” package (version 1.11-1; Nelson, 2019) in the R programming language (R Core Team, 2020); the gap function finds unusually large values in vectors of random samples (Tukey, 1971; Kappenman, 1999). Finally, once the average values (φi and δi) of each zone were estimated, the baselines for each zone i, and each statistical estimator (Kappenman and Pennington) were computed as and (Morales-Bojórquez et al., 2008), where n was the number of monthly average values for each zone i analyzed. The baselines for each zone i therefore represented the number of animals attracted to the purse seine by a fishing set; thus, the attracted animals were those counted within the net and those that swam or flew outside. According to Blackhart et al. (2006) the baseline is defined as a set of references used for comparative purposes; it can be based on a set of standard conditions, and in this study this was established from φi and δi, as suggested by Folmer and Pennington (2000).
The geographical location of fishing sets was recorded. For visualization purposes, the fishing sets were mapped into the regions analyzed in the Gulf of California. Kernel maps for representing the fishing effort in the Gulf of California were estimated based on two-dimensional (Gaussian) kernel density estimation with bandwidth selected by pilot estimation derivatives (Sheather and Jones, 1991; Venables and Ripley, 2002).
The fishing ground on the east coast of the Gulf of California includes Sonora and Sinaloa states, where Zones II, III, IV, and V were the most important fishing areas in both years analyzed, whereas Zone I presented a reduced number of fishing sets (Figures 1, 2). The kernel maps showing the spatial concentration of the fishing effort also indicated the east coast as the predominant region. During 2013, the fishing sets exhibited high frequency, mainly toward the north region in Zone III, and a lower quantity of fishing sets at the southern limit of this same zone (Figure 3). A different spatial pattern was observed for the fishing effort during 2014, although Zone III remained a suitable region for harvest, and Zones IV and V showed a concentration of fishing sets greater that observed in Zone III (Figure 4). In comparison, the fishing effort during 2013 was concentrated on the central east coast of the Gulf of California, and during 2014 the fishing effort changed its location toward southern coasts, identifying a more extended region where the fishing sets were recorded, and even the boundary between Zones II and III were suitable for harvest. Zones I, II, VI, and VII were not identifiable as suitable regions for harvesting Pacific sardine in either year.
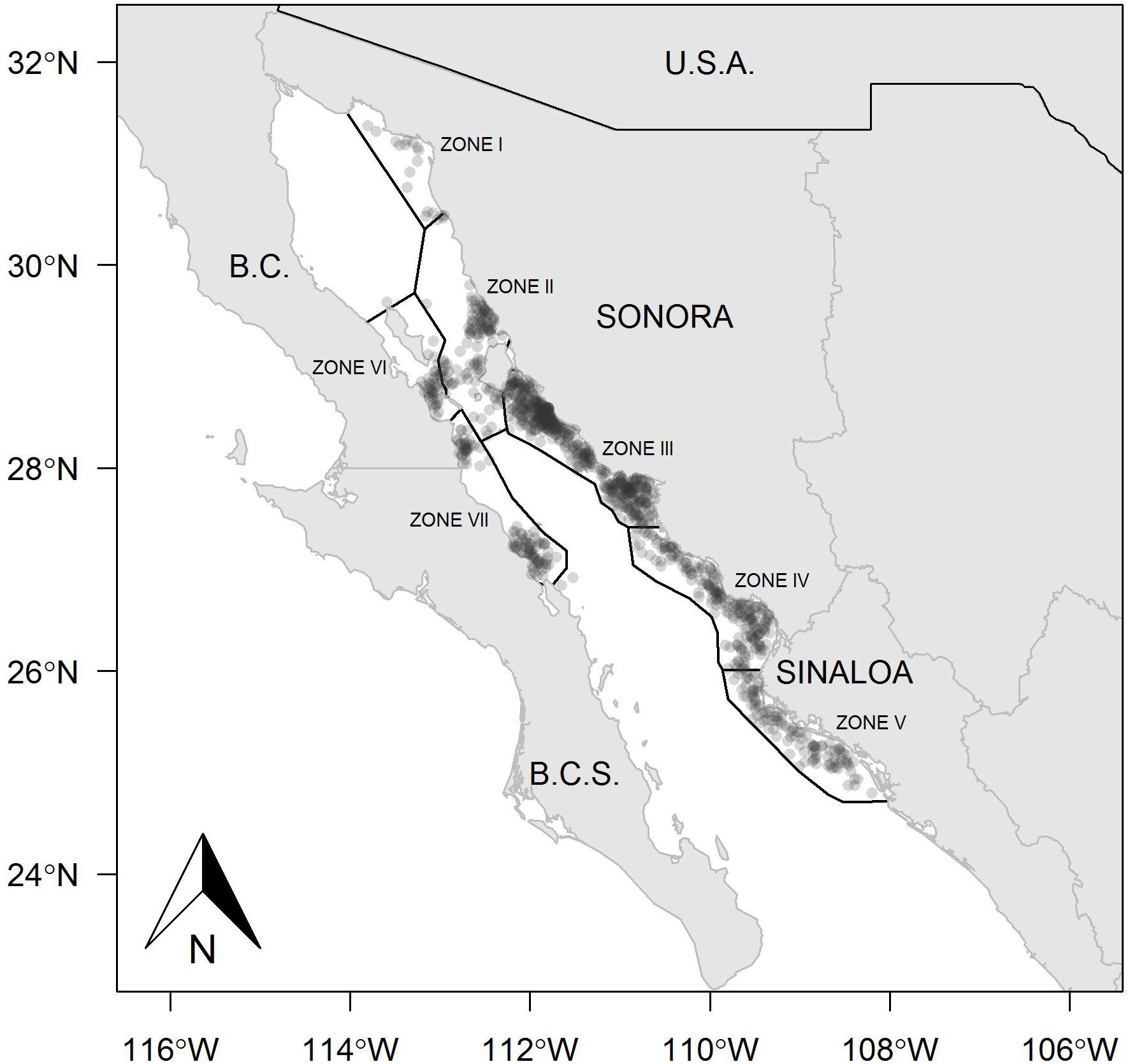
Figure 1. Zones and geographical locations of fishing sets in the small pelagic fishery in the Gulf of California, Mexico, during 2013.
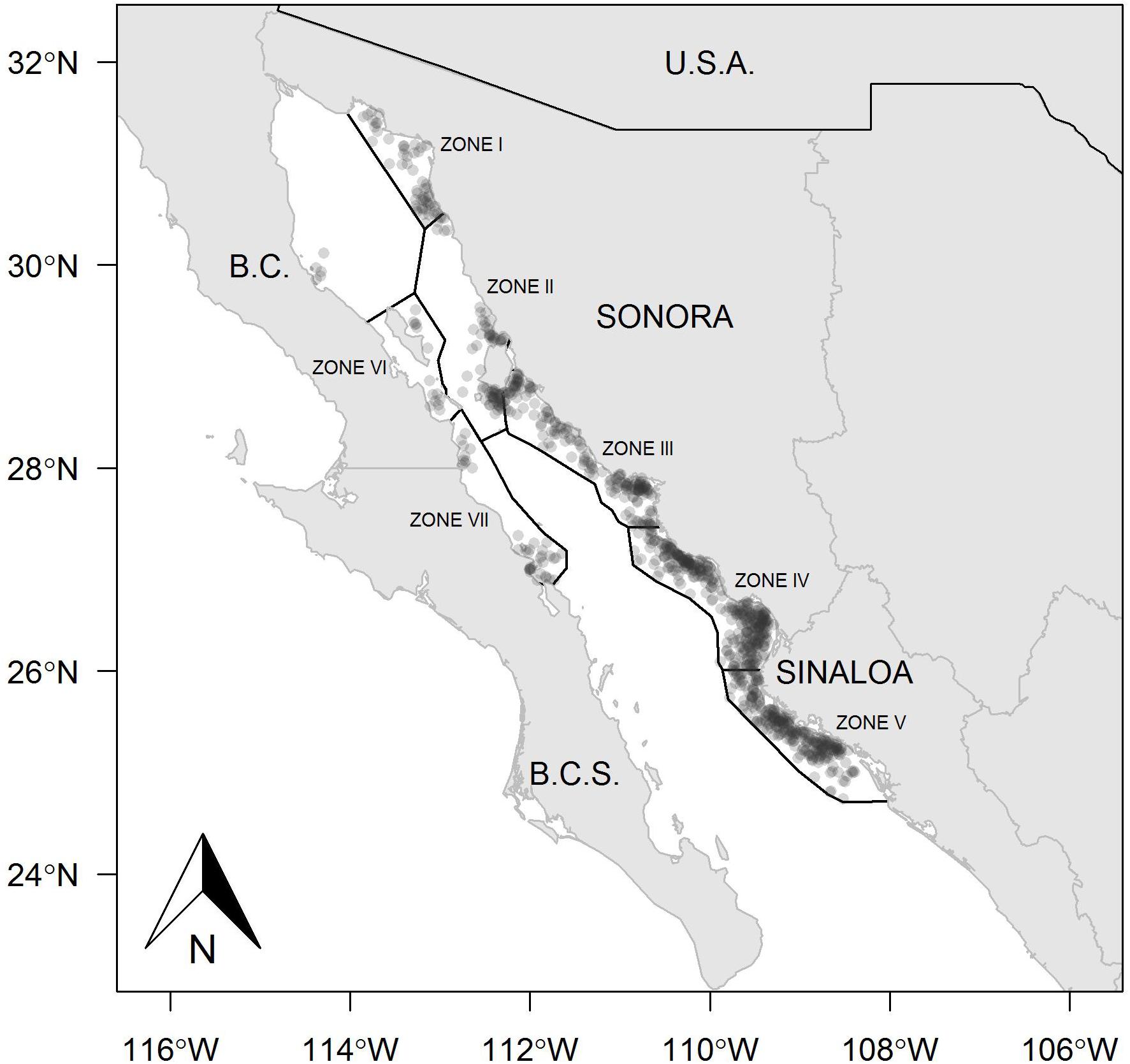
Figure 2. Zones and geographical locations of fishing sets in the small pelagic fishery in the Gulf of California, Mexico, during 2014.
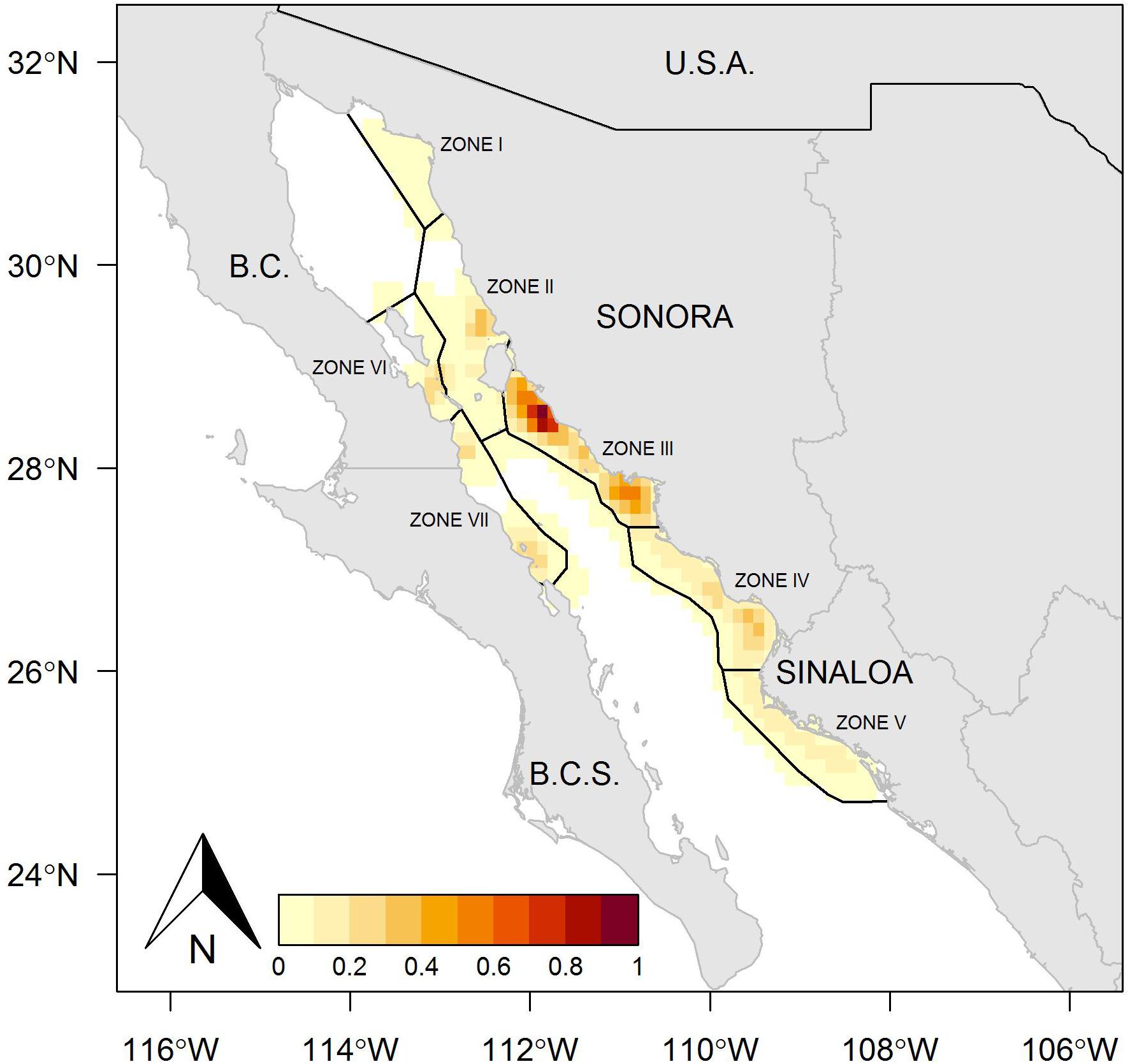
Figure 3. Study area showing the spatial distribution of fishing sets in the small pelagic fishery in the Gulf of California, Mexico, during 2013. The red contours indicate the geographical locations with a greater concentration of fishing effort.
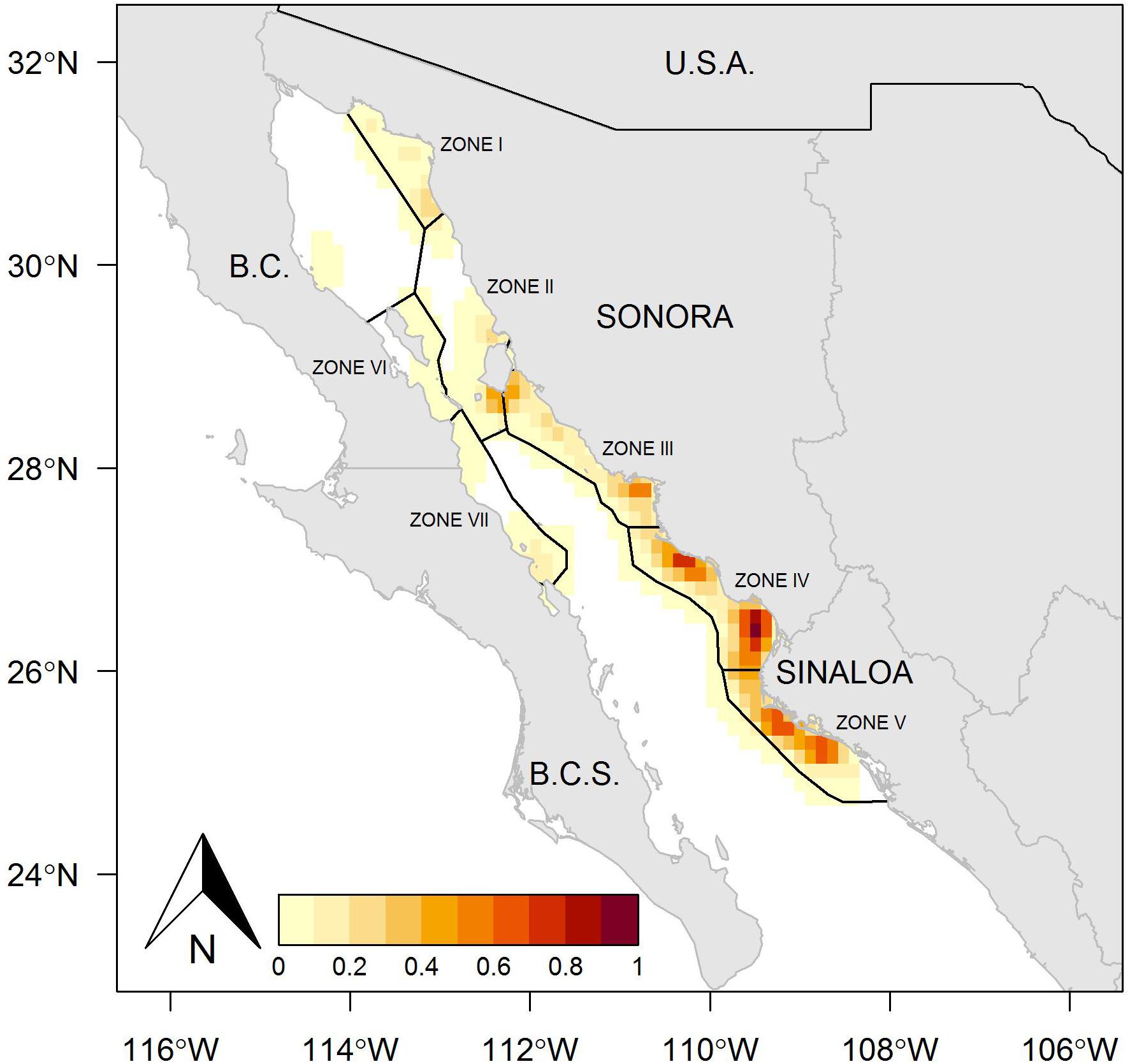
Figure 4. Study area showing the spatial distribution of fishing sets of the small pelagic fishery in the Gulf of California, Mexico, during 2014. The red contours indicate the geographical locations with a greater concentration of fishing effort.
Zone I corresponds to the upper Gulf of California, and showed a very low level of fishing activity, with only 204 fishing sets recorded across January, February, April, May, and June 2014 (Figures 5, 6); the baseline in this zone varied between φ = 52 and δ = 70 individuals attracted to the fishing set. For both statistical estimators, the monthly average number of individuals attracted by fishing sets varied from 104 (May) to 6 (June). Zone II showed baseline values (φ = 30 and δ = 33), estimated from 295 fishing sets from March 2013 to July 2014. The comparison of statistical estimators showed similar minimum and maximum values, which were observed during July 2013 (14 monthly average individuals attracted to the fishing set), and April 2014 (53 monthly average individuals attracted to the fishing set), respectively. This zone had a spatial coverage for the east coast in the Gulf of California (Figures 5, 6).
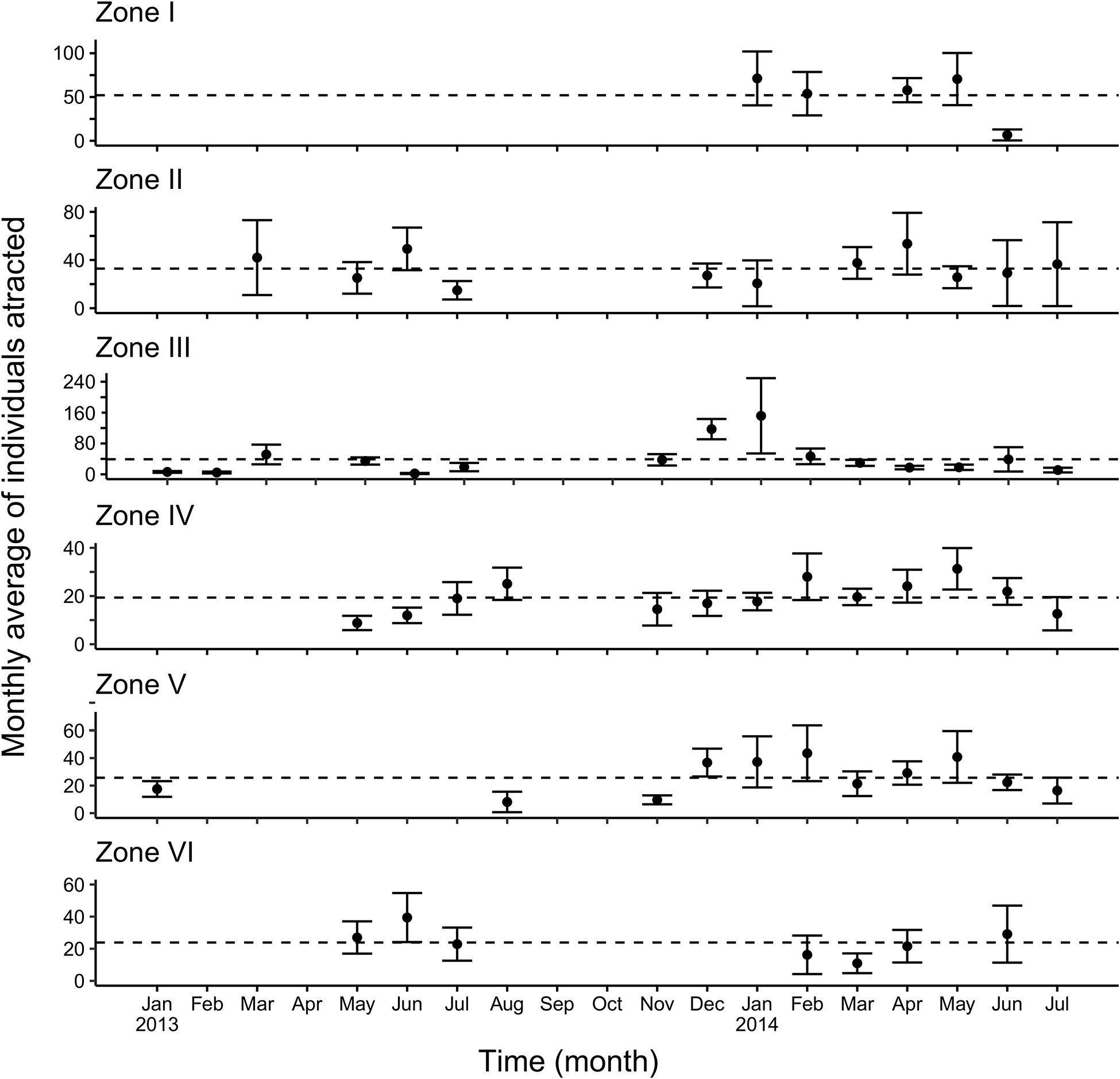
Figure 5. Monthly average values estimated for endangered and threatened marine fauna attracted by fishing sets based on the Kappenman estimator. The discontinuous line represents the expected value of individuals released alive by fishing set, defined by the φ value.
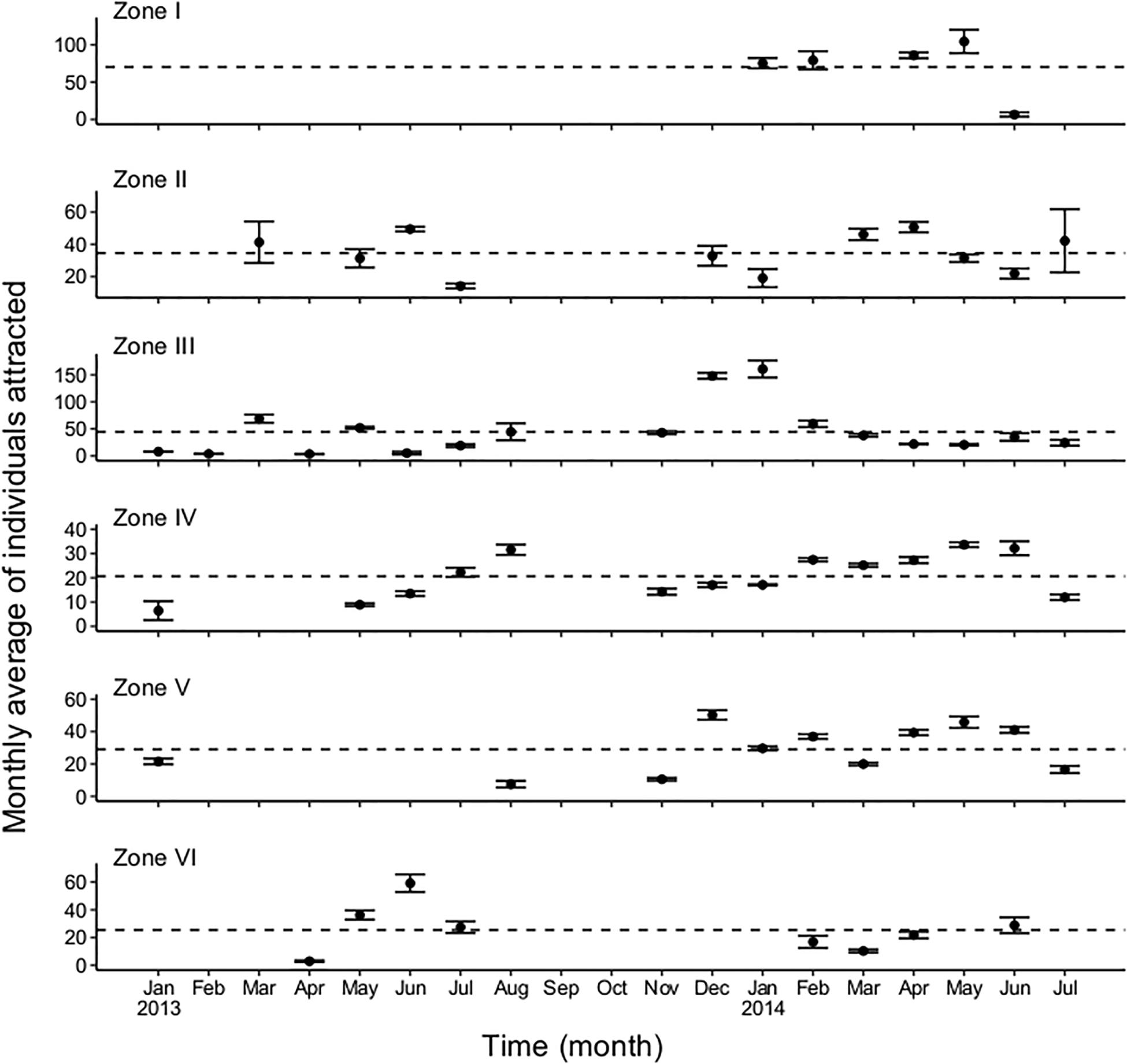
Figure 6. Monthly average values estimated for endangered and threatened marine fauna attracted by fishing sets based on the Pennington estimator. The discontinuous line represents the expected value of individuals released alive by fishing set, defined by the δ value.
Zone III was found to be a suitable area for the small pelagic fishing fleet, 802 fishing sets being recorded here. The baselines for both statistical estimators were φ = 39 and δ = 44. There was an estimated increase in the monthly average of individuals attracted between November 2013 and January 2014. Monthly averages of 39 and 160 individuals were thus estimated during November 2013 and January 2014, respectively. A low presence of organisms was recorded for several months (January, February, and April 2013), varying from 3 to 7 individuals attracted to the fishing set (Figures 5, 6). There were also a large number of fishing sets (867) in Zone IV. The baselines for the statistical estimators were φ = 19 and δ = 20, indicating a similar performance for both. Nevertheless, the monthly average values from February to June were relatively stable, showing only a slight increment for the Pennington estimator (from 27 to 32 individuals attracted to the fishing set), in contrast to the Kappenman estimator. An increase in the average number of individuals attracted was observed from May to August 2013. Considering the variation between the two statistical estimators, the monthly average of individuals attracted varied from 5 to 31 (Figures 5, 6). In comparison, the number of individuals attracted in this zone was less than that observed in Zone I.
In Zone V, 595 fishing sets were recorded; the baselines were φ = 26 and δ = 29. The fishing sets during 2013 were reduced (145) in this zone, and only occurred during January, August, November and December, in comparison to 2014 when there were 450 fishing sets from January to July. In Zone VI, 106 fishing sets were analyzed; the baselines were φ = 37 and δ = 49. Only 3 months were sampled in this zone, January 2013, and March and May 2014. In comparison, Zone VI showed the lowest harvest in the Gulf of California (Figures 5, 6). Finally, Zone VII, located on the west coast of the Gulf of California, exhibited similar baselines of φ = 24 and δ = 25, both estimated from 182 fishing sets, which were recorded from April to July 2013, and from February to April, and June 2014 for the Pennington estimator. The Kappenman estimator showed the same temporality, except in April 2013, when the data did not provide a monthly average of individuals attracted due to the limited fishing sets (10) (Figures 5, 6).
During the study period, 3,051 fishing sets were sampled, and the mortality of endangered and threatened marine fauna from the Gulf of California was 260 individuals. They were identified as follows: (a) sharks Carcharodon carcharias (1), and Sphyrna lewini (1); (b) fishes Hippocampus ingens (4), Totoaba macdonaldi (3), and Pomacanthus zonipectus (2); (c) reptiles Chelonia agassizii (2); (d) birds Pelecanus occidentalis (107), and Sula nebouxii (100); and (e) marine mammals Zalophus californianus (2), Tursiops truncatus (24), and Delphinus capensis (14). Of the 260 organisms that died, the highest mortality proportions were estimated for brown pelican P. occidentalis (41%), blue-footed booby S. nebouxii (38%), and common bottlenose dolphin T. truncatus (9%). The species of whale shark Rhincodon typus and olive ridley sea turtle Lepidochelys olivacea were also sighted by onboard observers, although these species did not exhibit interaction with the fishing sets during the study period (Table 1). Other species frequently attracted by fishing sets were members of the cnidarian, gastropoda, crustacean, cephalopoda, osteichthyes and chondrichthyes taxonomic groups, grouping approximately 13 species, which are not classified as endangered or threatened marine fauna (Table 2).
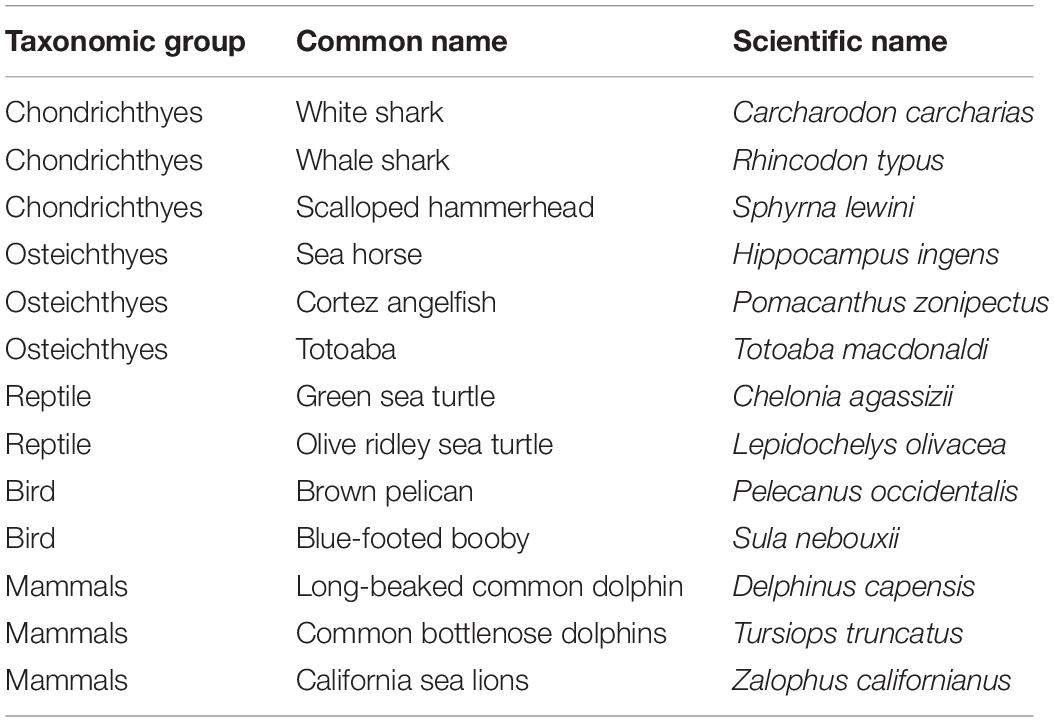
Table 1. Endangered and threatened marine fauna attracted by fishing sets in the small pelagic fishery in the Gulf of California, Mexico.

Table 2. Species most frequently attracted by fishing sets in the small pelagic fishery in the Gulf of California, Mexico.
The interaction between marine fauna and fishing sets included the number of organisms for whatever species was attracted to the fishing maneuver, as well as the number of organisms released and dead by species. In this study, the species released included individuals that interacted with the fishing gear and freely moved away from the fishing maneuver; this was mainly birds and marine mammals, so that few individuals were released for the fishers, and the low impact on these species of endangered and threatened marine fauna was observed in the high values of organisms released, which was greater than 88% (Table 3). In contrast, the taxonomic groups of sharks, fishes and reptiles exhibited a lower percentage of individuals released; although the number of organisms attracted to the fishing sets were also low; for example, the number of individuals attracted in these species varied between 3 (reptiles) and 12 (sharks). The fishes were characterized by Hippocampus ingens, Totoaba macdonaldi, and Pomacanthus zonipectus; if a fish species is captured it is difficult to release it alive, although the frequency of these fishes was extremely low, only nine individuals were recorded as bycatch from January 2013 to July 2014 (Table 3). The incidental catch of these fishes is not strictly conditioned as species attracted to the fishing sets, and their capture is an exceptional event in the small pelagic fishery.
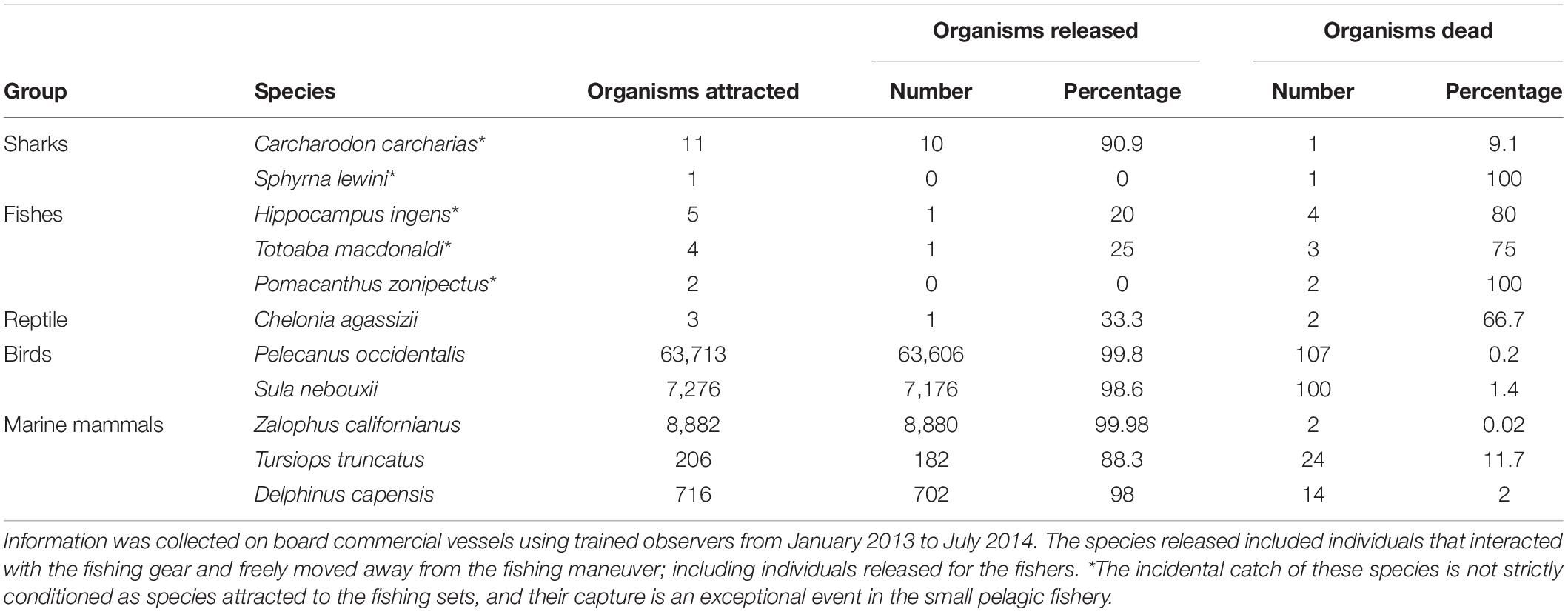
Table 3. Number of organisms of endangered and threatened marine fauna attracted, released, and killed by fishing sets in the small pelagic fishery from the Gulf of California, Mexico.
This study is the first analysis of endangered and threatened marine fauna attracted by fishing sets in a small pelagic fishery, including different species attracted to bait, offal, and discards from fishing vessels, and therefore, these species may have been temporally vulnerable to mortality. According to Gislason et al. (2000), incidental mortality is unavoidable in several marine fisheries. The estimates of this study, obtained using Kappenman and Pennington estimators provided two different average values of individuals attracted during each fishing set realized; both statistical distributions exhibited a high level of performance when the biological data was skewed. The presence and number of organisms varied greatly across zones, and depended entirely on the ability of the onboard observers to identify and count individuals.
It was not possible in this study to compare the Kappenman and Pennington estimators. Both estimators are useful for computing average values, but they are not goodness-of-fit functions, and therefore cannot be statistically discriminated (Hilborn and Walters, 1992). Nevertheless, both estimators indicated similar average values based on different statistical assumptions (Pennington, 1996; Kappenman, 1999). The application of both estimators was thus necessary because the information was collected in terms of animals sighted rather than dead, increasing the degree of uncertainty in the observations, including misidentifications and unaccounted numbers of animals, as well as the experience of the onboard observers (Uhlmann and Jeschke, 2011). The probability of identifying whether an individual is alive or dead may also be associated with specific environmental (e.g., sea state, visibility), technical (e.g., gear type, fishing strategy), or biological (e.g., body size, behavior, abundance) conditions (Uhlmann and Jeschke, 2011).
Cisneros-Mata et al. (1995) and Nevárez-Martínez et al. (2001) explained that the east coast of the Gulf of California is the main fishing marine area of the small pelagic harvest, mainly Sardinops sagax. Zones I and II along this coast in the northwestern part of the Gulf of California showed reduced fishing activity, as they were important sites for the spawning, mating and nursing of numerous species of commercial and ecological importance, including areas where fishing activity is limited and/or forbidden (Aragón-Noriega and Calderon-Aguilera, 2000). Despite the low presence of the fishing fleet in these regions, however, their average values were relatively high. Zones III, IV, and V were preferred by the fishing fleet, and the average number of individuals attracted by fishing set varied from 19 to 44. In comparison, the west coast did not vary notably in terms of the average number of individuals attracted into Zone VII (φ = 24, δ = 25 organisms). The average values estimated for each fishing zone from the Gulf of California may prove useful management criteria in order to avoid high rates of incidental mortality. According to Pitcher and Pauly (1998), baselines must meet three basic assumptions: (1) that there is a natural baseline which can be identified and agreed upon; (2) baselines can be described accurately; and (3) baselines can be restored. The actions implemented to improve the survival of non-target species caught by the Mexican small pelagic fishery, and their effects on endangered and threatened marine fauna require baselines, and a conservative baseline is recommended. The Pennington estimator may be useful for this purpose, considering the precision of its confidence intervals and the nature of the uncertainty involved in data collection regarding endangered and threatened marine fauna.
Undoubtedly, the number of organisms interacting with this fishery was very high and the mortality due to bycatch was extremely reduced (260 organisms). This result was similar to that reported for the sardine purse seine fishery on the southern coast of Western Australia, where only 30% of the fishing trips caused bycatch (Norriss et al., 2020). In comparison, the estimated bycatch of the purse seine fleets in south-central Chile affects the survival of birds such as the sooty shearwater (Ardenna grisea), Magellanic penguin (Spheniscus magellanicus), and Guanay cormorant (Phalacrocorax boungainvillii). Although specific studies are required to analyze the effect of the bycatch on the sardine fishery in the Gulf of California, the results suggest that the fishing gear used to catch small pelagic species has little impact on the ecosystem; the incidental catch is low, and even negligible for some taxonomic groups, such as birds, sharks, fishes, and reptiles. Management actions must be implemented to this end, and the incidental mortality rate in this fishery must be continuously monitored, particularly for the current status of species analyzed in this study.
The sardine fishery provides an abundant, easy and predictable food source for many opportunistic species, such as seabirds or marine mammals; however, this modified feeding behavior also exposes them to threats, such as accidental bycatch (Le Bota et al., 2019; Wise et al., 2019). The mortality caused by bycatch in the Mexican sardine fishery (purse-seine fleet) is less than that reported for fishing gear such as longlines, gillnets, trammel net or bottom trawl, where there is a severe impact for bycatch on several marine populations and the marine ecosystem (Hamilton and Baker, 2019; Holland and Martin, 2019; Zhou et al., 2019; Calado et al., 2021; Simeone et al., 2021). Thus, this study was focused on the estimation of regional baselines for the bycatch of endangered and threatened marine species attracted to a purse seine in the Mexican small pelagic fishery; these baselines quantify the number of organisms attracted to the fishing gear, which could potentially be killed due to accidental bycatch, although the results suggested the high survival of organisms successfully released by the fishers.
The fishing effort was concentrated on the east coast of the Gulf of California (Zones III, IV, and V), the traditional fishing grounds reported for Sardinops sagax (Cisneros-Mata et al., 1995), indicating that the fishing fleet had a very low harvest in the Biosphere Reserve of the upper Gulf of California and the spawning and nursery area for Pacific sardine. The effects of the annual bycatch on the pool of endangered and threatened marine species showed low mortality proportions; a similar situation was reported by Norriss et al. (2020) where zero mortalities for seabirds on at least 70% of trips were recorded during 2009–2018. Conversely, Simeone et al. (2021) reported high mortalities of seabirds; approximately 19,000 individuals of 16 species were killed over 14 years (2005–2019).
The framework for the spatial analysis of the bycatch for the pool of endangered and threatened marine species incidentally harvested by the Mexican fishing fleet requires adequate hypotheses. First, the effects of the fishing effort must be quantified for the total number of species incidentally caught; the pool of endangered and threatened marine species is particularly relevant for conservation goals, however, there are more species susceptible to incidental harvest. According to Gordoa et al. (2006), the importance of the fishing effort can be assessed through the changes in catchability for the target species; they also found a strong negative relationship between monthly catch rates for Namibian Cape hake and bycatch; when the catch rate diminished, the bycatch increased. Second, the bycatch analytically estimated using Pennington and Kappenman estimators indicates its temporal variability in two hypothetical distributions, and the selection of the best estimator requires further analysis, such that it can be used for a spatial analysis with the lowest uncertainty possible.
Studies of incidental mortality are relevant because they allow an understanding of the potential effects on non-target species and their populations; indeed, the objective is to reduce the vulnerability of these species (Lawrence and Bazhin, 1998). In this study, the bycatch in the Mexican small pelagic fishery was not strictly analyzed, because the field data of incidental mortality showed a very low frequency and was only useful for descriptive purposes. A total of 3,051 fishing sets were sampled during the fishing seasons in 2013 and 2014, with a mortality of 260 individuals. The contribution to the implementation of actions to improve the survival of non-target species should be monitored from the specific baseline for each zone, so that the impacts on the populations constituting the non-target species can be analyzed in the future.
In the small pelagic fishery, a conservative baseline based on the Pennington estimator was estimated for each zone in the Gulf of California, considering the precision of the confidence intervals and the nature of the uncertainty in the data collection based on sightings. The upper Gulf of California region showed a low presence of individuals, and the east coast of the Gulf of California recorded more sightings than the west coast; consequently the fishing effort is mainly distributed in the east coast. The number of organisms interacting with this fishery (seabirds, fishes, reptiles, marine mammals, and sharks) was very high, and the mortality due to bycatch was extremely reduced.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was reviewed and approved by the Comisión Nacional de Acuacultura y Pesca.
DA-R, MÁM-Z, and SG-P contributed to the data acquisition and analysis and data interpretation. EM-B and MN-M wrote the first draft of the manuscript. EM-B, MN-M, and HA-V contributed to the preparation of figures and drafting of the manuscript. EM-B, MN-M, GG-A, HV, HA-V, and EL-C contributed to the data analysis and figure preparation. DA-R, MÁM-Z, and MN-M contributed to the design and conception of the sampling. All authors reviewed and approved the manuscript.
Funding was provided by the Instituto Nacional de Pesca y Acuacultura; Conservacion y Biodiversidad, AC; Cámara Nacional de la Industria Pesquera y Acuícola, Sonora; the Fundación Carlos Slim, and the World Wildlife Fund La Paz, Mexico.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Fisheries observers onboard the commercial Pacific sardine fleet are acknowledged for their valuable assistance, as well as Ernesto Gastélum Nava for providing support with the database. HV would like to thank the support from COFAA-IPN and EDI-IPN. We thank two reviewers who offered valuable suggestions that greatly improved the presentation and readability of this scientific report.
Alverson, D. L., Freeberg, M. H., Murawski, S. A., and Pope, J. G. (1994). A Global Assessment of Fisheries Bycatch and Discards. FAO Fish. Tech. Pap. 339. Rome: FAO.
Aragón-Noriega, E. A., and Calderon-Aguilera, L. E. (2000). Does damming the Colorado River affect the nursery area of blue shrimp Litopenaeus stylirostris (Decapoda:Penaeidae) in the Upper Gulf of California? Rev. Biol. Trop. 48, 867–871.
Blackhart, K., Stanton, D. G., and Shimada, A. M. (2006). NOAA Fisheries Glossary. In: NOAA Technical Memorandum NMFS-F/SPO-69, MD. Silver Spring, MD: NOAA.
Burgoni, L., Neves, T. S., Leite, N. O. Jr., Carvalho, D., Sales, R., Furness, R. W., et al. (2008). Potential bycatch of seabirds and turtles in hook-and-line fisheries of the Itaipava fleet. Brazil. Fish. Res. 90, 217–224. doi: 10.1016/j.fishres.2007.10.013
Calado, J. G., Ramos, J. A., Almeida, A., Oliveira, N., and Paiva, V. H. (2021). Seabird-fishery interactions and bycatch at multiple gears in the Atlantic Iberian coast. Ocean Coast. Manage. 200:105306. doi: 10.1016/j.ocecoaman.2020.105306
Casey, J. M., and Myers, R. A. (1998). Near extinction of a large, widely distributed fish. Science 281, 690–691. doi: 10.1126/science.281.5377.690
Cisneros-Mata, M. A., Nevárez-Martínez, M. O., and Hammann, M. G. (1995). The rise and fall of the Pacific sardine, Sardinops sagax caeruleus Girard, in the Gulf of California, Mexico. Calif. Coop. Oceanic Fish. Invest. Rep. 36, 136–143.
Darby, J. T., and Dawson, S. M. (2000). Bycatch of yellow-eyed penguins (Megadyptes antipodes) in gillnets in New Zealand waters 1979-1997. Biol. Cons. 93, 327–332. doi: 10.1016/s0006-3207(99)00148-2
Dillingham, P., and Fletcher, D. (2008). Estimating the ability of birds to sustain additional human-caused mortalities using a simple decision rule and allometric relationships. Biol. Cons. 141, 1783–1792. doi: 10.1016/j.biocon.2008.04.022
Espino-Barr, E., Macías-Zamora, R., Cruz-Romero, M., and García-Boa, A. (1997). Catch per unit effort trends in the coastal fishery of Manzanillo, Colima, Mexico. Fish. Manag. Ecol. 4, 255–261. doi: 10.1046/j.1365-2400.1997.00123.x
Folmer, O., and Pennington, M. (2000). A statistical evaluation of the design and precision of the shrimp trawl survey off West Greenland. Fish. Res. 49, 165–178. doi: 10.1016/s0165-7836(00)00196-x
Gislason, H., Sinclair, M., Sainsbury, K., and O’Boyle, R. (2000). Symposium overview: incorporating ecosystem objectives within fisheries management. ICES J. Mar. Sci. 57, 468–475. doi: 10.1006/jmsc.2000.0741
Gordoa, A., Lesch, H., and Rodergas, S. (2006). Bycatch: complementary information for understanding fish behaviour. Namibian Cape hake (M. capensis and M. paradoxus) as a case study. ICES J. Mar. Sci. 63, 1513–1519. doi: 10.1016/j.icesjms.2006.05.007
Guy, T. J., Jennings, S. L., Suryan, R. M., Melvin, E. F., Bellman, M. A., Ballance, L. T., et al. (2013). Overlap of North Pacific albatrosses with the U.S. west coast groundfish and shrimp fisheries. Fish. Res. 147, 222–234. doi: 10.1016/j.fishres.2013.06.009
Hall, S. J. (1999). The Effect of Fishing on Marine Ecosystems and Communities. Fish Biology and Aquatic Resources Series. Oxford: Blackwell.
Hamilton, S., and Baker, G. B. (2019). Population growth of an endangered pinniped -the New Zealand sea lion (Phocarctos hookeri) -is limited more by high pup mortality than fisheries bycatch. ICES J. Mar. Sci. 76, 1794–1806. doi: 10.1093/icesjms/fsz039
Hilborn, R., and Walters, C. J. (1992). Quantitative Fisheries Stock Assessment: Choice, Dynamics and Uncertainty. New York, NY: Chapman and Hall.
Holland, D. S., and Martin, C. (2019). Bycatch quotas, risk pools, and cooperation in the Pacific whiting fishery. Front. Mar. Sci. 6:600. doi: 10.3389/fmars.2019.00600
Hsiang-Wen, H., and Kwang-Ming, L. (2010). Bycatch and discards by Taiwanese large-scale tuna longline fleets in the Indian Ocean. Fish. Res. 106, 261–270. doi: 10.1016/j.fishres.2010.08.005
Izquierdo, V., Lluch-Cota, S., Lluch-Cota, D., Ponce, G., Morales-Bojórquez, E., and Chávez, F. (2020). Is there a future in the sustainability certification of sardine and anchovy fisheries? Fisheries 45, 554–560. doi: 10.1002/fsh.10461
Jennings, S., and Kaiser, M. J. (1998). The effects of fishing on marine ecosystems. Adv. Mar. Biol. 34, 201–352. doi: 10.1016/s0065-2881(08)60212-6
Kappenman, R. F. (1999). Trawl survey based abundance estimation using data sets with unusually large catches. ICES J. Mar. Sci. 56, 28–35. doi: 10.1006/jmsc.1998.0422
Kimberly, S. D., Parrish, J. K., and Melvin, E. F. (2009). Understanding and addressing seabird bycatch in Alaska demersal longline fisheries. Biol. Cons. 142, 2642–2656. doi: 10.1016/j.biocon.2009.06.013
Lanz, E., López-Martínez, J., Nevárez-Martínez, M. O., and Dworak, J. A. (2009). Small pelagic fish catches in the Gulf of California associated with sea surface temperature and chlorophyll. Calif. Coop. Oceanic Fish. Invest. Rep. 50, 134–146.
Larios-Castro, E., Morales-Bojórquez, E., Díaz-Uribe, J. G., Vargas-López, V. G., and Massó-Rojas, J. A. (2021). Historical reconstruction of biomass of Panopea globosa from the southwest coast of the Baja California Peninsula, Mexico: a new perspective for geoduck fishery management. J. Shellfish Res. 40, 49–61. doi: 10.2983/035.040.0106
Lavín, M. F., Beier, E., and Badan, A. (1997). “Estructura hidrográfica y circulación del Golfo de California: Escalas Estacional e Interanual,” in Contribuciones a la Oceanografía Física en México. Monografía No. 3, ed. M. F. Lavín (Ensenada: Unión Geofísica Mexicana), 141–117.
Lawrence, J. M., and Bazhin, A. (1998). Life-history strategies and the potential of sea urchins for aquaculture. J. Shellfish Res. 17, 1515–1522.
Le Bota, T., Lescroël, A., Fort, J., Péron, C., Gimenez, O., Provost, P., et al. (2019). Fishery discards do not compensate natural prey shortage in Northern gannets from the English Channel. Biol. Conserv. 236, 375–384. doi: 10.1016/j.biocon.2019.05.040
Lescrauwaet, A. K., Torreele, E., Vincx, M., Polet, H., and Mees, J. (2013). Invisible catch: a century of bycatch and unreported removals in sea fisheries, Belgium 1929–2010 2010. Fish. Res. 147, 161–174. doi: 10.1016/j.fishres.2013.05.007
Lluch-Belda, D., Magallón, F. J., and Schwartzlose, R. A. (1986). Large fluctuations in the sardine fishery in the Gulf of California: possible causes. Calif. Coop. Oceanic Fish. Invest. Rep. 27, 136–140.
Løkkeborg, S., and Robertson, G. (2002). Seabird and longline interactions: effects of a bird-scaring streamer line and line shooter on the incidental capture of northern fulmars Fulmarus glacialis. Biol. Cons. 106, 359–364. doi: 10.1016/s0006-3207(01)00262-2
Madrid-Vera, J., Amezcua, F., and Morales-Bojórquez, E. (2007). An assessment approach to estimate biomass of fish communities from bycatch data in a tropical shrimp-trawl fishery. Fish. Res. 83, 81–89. doi: 10.1016/j.fishres.2006.08.026
Majluf, P., Babcock, E. A., Riveros, J. C., Schreiber, M. A., and Alderete, W. (2002). Catch and bycatch of sea birds and marine mammals in the small-scale fishery of Punta San Juan, Peru. Cons. Biol. 15, 1333–1343. doi: 10.1046/j.1523-1739.2002.00564.x
Melvin, E. F., Guy, T. J., and Read, L. B. (2014). Best practice seabird bycatch mitigation for pelagic longline fisheries targeting tuna and related species. Fish. Res. 149, 5–18. doi: 10.1016/j.fishres.2013.07.012
Morales-Bojórquez, E., and Nevárez-Martínez, M. O. (2005). Spawner–recruit patterns and investigation of Allee effect in Pacific sardine Sardinops sagax caeruleus in the Gulf of California, Mexico. Calif. Coop. Oceanic Fish. Invest. Rep. 46, 161–174.
Morales-Bojórquez, E., Orieta Muciño-Díaz, M., and Vélez-Barajas, J. A. (2008). Analysis of the decline of the abalone fishery (Haliotis fulgens and H. corrugata) along the westcentral coast of the Baja California Peninsula, Mexico. J. Shellfish Res. 27, 865–870. doi: 10.2983/0730-8000(2008)27[865:aotdot]2.0.co;2
Nelson, G. A. (2019). Fishmethods: Fishery Science Methods and Models. Available online at: rdrr.io/cran/fishmethods/ (accessed December 16, 2019).
Nevárez-Martínez, M. O., Lluch-Belda, D., Cisneros-Mata, M. A., Santos-Molina, J. P., Martínez-Zavala, M. A., and Lluch-Cota, S. E. (2001). Distribution and abundance of the Pacific sardine (Sardinops sagax) in the Gulf of California and their relation with the environment. Progr. Oceanogr. 49, 565–580. doi: 10.1016/s0079-6611(01)00041-6
Nevárez-Martínez, M. O., Martínez-Zavala, M. A., Jacob-Cervantes, M. L., Cotero-Altamirano, C. E., Santos-Molina, J. P., and Valdez-Pelayo, A. (2014). “Peces pelágicos menores (Sardinops sagax, Opisthonema spp., Scomber japonicus, Engraulis mordax, Cetengraulis mysticetus, Etrumeus teres, Trachurus symmetricus, Oligoplites spp.),” in Sustentabilidad y Pesca Responsable en México. Evaluación y Manejo, eds L. F. Beléndez-Moreno, E. Espino-Barr, G. Galindo-Cortes, M. T. Gaspar-Dillanes, L. Huidobro-Campos, and E. Morales Bojórquez (Guadalajara: Instituto Nacional de la Pesca), 85–140.
Norman, F. I. (2000). Preliminary investigation of the bycatch of marine birds and mammals in inshore commercial fisheries, Victoria, Australia. Biol. Cons. 92, 217–226. doi: 10.1016/s0006-3207(99)00055-5
Norriss, J. V., Fisher, E. A., and Denham, A. M. (2020). Seabird bycatch in a sardine purse seine fishery. ICES J. Mar. Sci. 77, 2971–2983. doi: 10.1093/icesjms/fsaa179
Ortiz, M., Legault, C. M., and Ehrhardt, N. (2000). An alternative method for estimating bycatch from the U.S. shrimp trawl fishery in the Gulf of Mexico, 1972–1995. Fish. Bull. 98, 583–599.
Pedrín, O. A., and Ancheita, A. (1976). Estadística Básica de la Explotación de Sardina en el Noroeste de México. Maxico: Serie Información. Instituto Nacional de Pesca, Vol. 76, 52
Pennington, M. (1983). Efficient estimators of abundance for fish and plankton surveys. Biometrics 39, 281–286. doi: 10.2307/2530830
Pennington, M. (1986). Some statistical techniques for estimating abundance indices from trawl surveys. Fish. Bull. 84, 519–525.
Pennington, M. (1996). Estimating the mean and variance from highly skewed marine data. Fish. Bull. 94, 498–505.
Pennington, M., and Stromme, T. (1998). Surveys as a research tool for managing dynamics stocks. Fish. Res. 37, 97–106. doi: 10.1016/s0165-7836(98)00129-5
Pitcher, T. J., and Pauly, D. (1998). “Rebuilding ecosystems, not sustainability, as the proper goal of fishery management,” in Reinventing Fisheries Management, eds T. Pitcher, D. Pauly, and P. Hart (London: Kluwer academic publishers), 311–325. doi: 10.1007/978-94-011-4433-9_24
Pope, J. G., MacDonald, D. S., Dann, N., Rynolds, J. D., and Jennings, S. (2000). Gauging the impact of fishing mortality on nontarget species. ICES J. Mar. Sci. 57, 689–696. doi: 10.1006/jmsc.2000.0729
R Core Team (2020). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Sheather, S. J., and Jones, M. C. (1991). A reliable data-based bandwidth selection method for kernel density estimation. J. Royal Stat. Soc. Ser. B. 53, 683–690.
Simeone, A., Anguita, C., Daigre, M., Arce, P., Vega, R., Luna-Jorquera, G., et al. (2021). Spatial and temporal patterns of beached seabirds along the Chilean coast: Linking mortalities with commercial fisheries. Biol. Conserv. 256:109026. doi: 10.1016/j.biocon.2021.109026
Uhlmann, S. S., and Jeschke, J. M. (2011). Comparing factors associated with total and dead sooty shearwater bycatch in New Zealand trawl fisheries. Biol. Cons. 144, 1859–1865. doi: 10.1016/j.biocon.2011.02.025
Venables, W. N., and Ripley, B. D. (2002). Modern Applied Statistics with S. New York, NY: Springer.
Wise, L., Galego, C., Katara, I., Marçalo, A., Meirinho, A., Monteiro, S. S., et al. (2019). Portuguese purse seine fishery spatial and resource overlap with top predators. Mar. Ecol. Prog. Ser. 617-618, 183–198. doi: 10.3354/meps12656
Zador, S., Punt, A. E., and Parrish, J. K. (2008). Population impacts of endangered short-tailed albatross bycatch in the Alaskan trawl fishery. Biol. Cons. 141, 872–882. doi: 10.1016/j.biocon.2008.01.001
Zhou, S., Daley, R. M., Fuller, M., Bulman, C. M., and Hobday, A. J. (2019). A data-limited method for assessing cumulative fishing risk on by-catch. ICES J. Mar. Sci. 76, 837–847. doi: 10.1093/icesjms/fsy206
Keywords: fishery, non-target species, mortality, sardine, endangered and threatened species
Citation: Morales-Bojórquez E, Nevárez-Martínez MO, García-Alberto G, Villalobos H, Aguirre-Villaseñor H, Larios-Castro E, González-Peláez SS, Arizmendi-Rodríguez DI and Martínez-Zavala MÁ (2021) Interaction Between Marine Fauna and the Small Pelagic Fishery in the Coastal Environment of the Gulf of California, Mexico. Front. Mar. Sci. 8:669176. doi: 10.3389/fmars.2021.669176
Received: 18 February 2021; Accepted: 17 May 2021;
Published: 21 June 2021.
Edited by:
Andrés M. Cisneros-Montemayor, University of British Columbia, CanadaReviewed by:
Diego Lercari Bernier, Universidad de la República, UruguayCopyright © 2021 Morales-Bojórquez, Nevárez-Martínez, García-Alberto, Villalobos, Aguirre-Villaseñor, Larios-Castro, González-Peláez, Arizmendi-Rodríguez and Martínez-Zavala. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Manuel O. Nevárez-Martínez, bWFudWVsLm5ldmFyZXpAaW5hcGVzY2EuZ29iLm14
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.