
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 16 December 2020
Sec. Aquatic Physiology
Volume 7 - 2020 | https://doi.org/10.3389/fmars.2020.589822
This article is part of the Research TopicAdvances in Ecotoxicology Research of Harmful Algal Blooms Using OMICsView all 3 articles
 José Carlos Martins1†
José Carlos Martins1† Dany Domínguez-Pérez1†Catarina Azevedo1Ana Catarina Braga2
Dany Domínguez-Pérez1†Catarina Azevedo1Ana Catarina Braga2 Pedro Reis Costa2
Pedro Reis Costa2 Hugo Osório3,4,5
Hugo Osório3,4,5 Vitor Vasconcelos1,6
Vitor Vasconcelos1,6 Alexandre Campos1*
Alexandre Campos1*The molecular pathways behind the toxicity of diarrheic shellfish toxins (DSTs) in bivalves have been scarcely studied. Thus, a shotgun proteomics approach was applied in this work to understand bivalves’ molecular responses to the dinoflagellate Prorocentrum lima (1.0 × 106 cells/L). Protein expression along with toxins levels were analyzed in the gills and digestive gland of the mussel Mytilus galloprovincialis during and after exposure to this toxic strain. Results revealed an accumulation of OA and DTX1 only in the digestive gland with maximum amounts attained at the end of uptake phase (day 5; 2819.2 ± 522.2 μg OA/kg and 1107.1 ± 267.9 μg DTX1/kg). At the end of the depuration phase (day 20), 16% and 47% of total OA and DTX1 concentrations remained in the digestive gland tissues, respectively. The shotgun proteomic analyses yielded 3051 proteins in both organs. A total of 56 and 54 differentially expressed proteins (DEPs) were revealed in the digestive gland and gills, respectively. Both organs presented the same response dynamics along the experiment, although with tissue-specific features. The early response (3 days uptake) was characterized by a high number of DEPs, being more marked in gills, in relation to the latter time points (5 days uptake and depuration). Functional enrichment analysis revealed the up-regulation of carboxylic (GO:0046943) and organic acid transmembrane transporter activity (GO:0005342) pathways after 3 days uptake for digestive gland. Matching to these pathways are a group of proteins related to transmembrane transport and response to toxic substances and xenobiotics, namely P-glycoprotein (ABCB11), Sodium-dependent proline transporter (SLC6A7), and Sideroflexin-1 (SFXN1). According to Clusters of Orthologous Groups (GOs) categories, most of the DEPs found for digestive gland in all time-points were related with “cellular processes and signaling” and involving signal transduction mechanisms, cytoskeleton and post-translational modification, protein turnover, chaperone functions. In gills, the early uptake phase was marked by a balance between DEPs related with “cellular processes and signaling” and “metabolism.” Depuration is clearly marked by processes related with “metabolism,” mainly involving secondary metabolites biosynthesis, transport, and catabolism. Proteomic data are available via ProteomeXchange with identifier PXD022293.
The marine phycotoxins responsible for the diarrheic shellfish poisoning (DSP), along with paralytic shellfish poisoning (PSP), and amnesic shellfish poisoning (ASP) are the toxins produced by algal blooms with the highest incidence in the world, being found in the most varied marine environments, causing serious contamination problems in marine fauna, fishery and aquaculture products (Shumway, 1995; Torgersen et al., 2008). The main alga producing DSP toxins (DSTs) and responsible for the contamination of seafood belongs to the genus Dinophysis (Lindholm, 1985; Gómez, 2005). The genus Prorocentrum also produces DSTs, however, being a benthic microalga, its relevance in the overall transfer of these phycotoxins to the marine food chain is reduced (Nielsen et al., 2013). The consumption of contaminated food products has been rising serious public health concerns (Shumway, 1995; Torgersen et al., 2008). Symptoms associated with DSTs include diarrhea, nausea, vomiting, and abdominal pain. Animals that feed directly on microalgae, such as filter feeding bivalve mollusks (mussels, cockles, scallops, and oysters) are among those which accumulate highest amounts of phycotoxins (Shumway, 1995). Being widely used in the human diet, these marine invertebrates are also responsible for the majority of food poisoning in Humans. DSTs are included in a broad group of lipophilic toxins and are represented by okadaic acid (OA) and its analogs called dinophysistoxins (DTXs). The mode of action of OA and DTXs is the inhibition of serine/threonine protein phosphatases 1 (PP1) and 2A (PP2A) by direct molecular interaction (Konoki et al., 2013). So far, no effective mitigation or preventive strategies have been proposed and/or put into practice regarding the contamination of bivalve mollusks with DSTs.
The dynamics of accumulation and clearance of these toxins is different among bivalves. Several studies have shown that they quickly accumulate DSTs by feeding and ingesting toxic algae (Rossignoli et al., 2011; Mafra et al., 2015; Nielsen et al., 2016). Differences in the accumulation of OA and DTX have been found between bivalve species (Vale et al., 1999; Vale and Maria, 2002; Vale, 2004, 2006). According to several monitoring studies, the mussel Mytilus galloprovincialis is one of the species that shows a higher potential to accumulate OA homologues associated to a lower ability to clear this toxin in comparison with other bivalves (Suzuki and Mitsuya, 2001; Kacem et al., 2010). As a case in point, M. galloprovincialis from Bizerta lagoon in Tunisia had the capacity to accumulate 10–70 times greater levels of OA in their tissues than the oyster Crassostrea gigas (Kacem et al., 2010). Furthermore, results also showed that OA-group toxins appeared to be reduced at a faster rate in oysters (t1/2 = 9 days) compared with mussels (t1/2 = 18 days; Kacem et al., 2010). Similar results were observed on Mutso Bay (Japan) during a Dinophysis fortii bloom with M. galloprovincialis attaining significantly higher toxin contents than those in the scallops Patinopecten yessoensis (Suzuki and Mitsuya, 2001). Moreover, toxin profiles were very different between both mollusks with DTX1 and DTX3 as the dominant toxins in mussels and scallops, respectively, Suzuki and Mitsuya (2001). The highly variable toxins profile found among these mollusks suggests that different physiological and molecular processes are involved in OA-group toxins accumulation (Vale et al., 1999; Vale and Maria, 2002; Vale, 2004, 2006). Likely, many of these processes are related with bivalves’ feeding behavior (ingestion of microalgae), and toxin assimilation, excretion and biotransformation/detoxification. With respect to this latter case, it is known that the digestive gland is the main organ where DSTs are accumulated (Bauder et al., 2001). Several studies have shown that these toxins undergo transformations during the digestive process resulting in the conversion of the parent compound into less toxic derivatives (Blanco, 2018; Lopes et al., 2019). The main transformation route is the esterification with fatty acids of different chain lengths (Blanco, 2018). Multiple chemical forms, collectively called DTX3, are accumulated in bivalves and result from the process of acylation, i.e., the binding of fatty acids to OA, DTX1, and DTX2 (Yasumoto et al., 1985; Marr et al., 1992; Vale et al., 1999; Suzuki and Mitsuya, 2001; MacKenzie et al., 2002; Vale and Maria, 2002; García et al., 2004; JØrgensen et al., 2005). Esterification is species- and toxin-specific and is recognized as an important step in depuration since esterified forms are cleared faster than free toxins (Vale et al., 2008). In contrast with most of the studied bivalve species, it was reported that the mussel M. galloprovincialis and several others present low esterification rates for OA-group toxins (OA, DTX1-2) and consequent slower depuration rates of the larger proportion of non-esterified toxins (Vale et al., 2008).
Up to now, it is not yet clear whether DSTs represent a real threat to bivalve survival. Several studies showed that DST-producing microalgae or their purified toxins can promote histopathological alterations (Simães et al., 2015; Romero-Geraldo et al., 2016; Neves et al., 2019) along with immunotoxicity (Chi et al., 2016, 2017), cytotoxicity (Hanana et al., 2012; Prado-Alvarez et al., 2012, 2013), and genotoxicity (Carvalho Pinto-Silva et al., 2003, 2005; Florez-Barros et al., 2011; McCarthy et al., 2014; Prego-Faraldo et al., 2015, 2016) effects in these animals. At the same time, bivalves seem to tolerate the accumulation of high toxic levels of these toxins in their tissues and, sometimes, for long periods of time (Bauder et al., 2001). Some studies indicate that OA produces the most dramatic effects at low concentrations after short exposure duration, whereas high concentrations and longer exposures initiate resistance mechanisms (Prego-Faraldo et al., 2015). Cytochrome P450, Glutathione S-transferases, ABC transporters (P-glycoprotein) and antioxidant metabolism enzymes (such as superoxide dismutase and catalase) have been associated to DSTs resistance in bivalves (Huang et al., 2014; Zou et al., 2015; Prego-Faraldo et al., 2017; Wei et al., 2019).
During the last decade, an increase number of OMICs studies, mostly toxicogenomics, have been applied in bivalves to describe the molecular pathways behind the toxicity of DSTs. Almost all describe the early response (6 to 96 h) of mussels and scallops experimentally exposed to purified OA and to the OA-producing dinoflagellate P. lima. The only proteomic study was developed by Huang et al. (2015b) in which protein profiles in gills of Perna viridis were compared after exposure to P. lima using 2D-electrophoresis. Taking this into account, the present work intends to take a step further regarding the understanding of the molecular responses behind DSTs toxicity in bivalves by carrying out a shotgun proteomics approach in the commercial mussel M. galloprovincialis experimentally exposed to P. lima. The molecular events associated with the response of mussels’ gills and digestive gland were investigated during accumulation (3 and 5 days) and depuration (14 days) of OA and DTX. Moreover, the period of the stress exposure to P. lima with the recovery phase for both organs were compared aiming at achieving a more comprehensive picture concerning the physiological and molecular processes that participate in DSTs response in these mollusks.
The strain PL13v of P. lima isolated in 1996 in the Ria of Vigo, NW Iberian Peninsula, was kindly provided by Dr. Paulo Vale, from the Portuguese Institute of the Sea and Atmosphere (IPMA). P. lima is a benthic dinoflagellate species with world-wide distribution. Cell size ranging between 32–50 μm in length and 20–28 μm in width. P. lima was cultivated in saline f2 medium (35‰ salinity) at 16°C ± 1°C, photoperiod of 14 h/10 h (light/dark) and light intensity of 20 μmol m–2 s–1. Cultures were up-scaled to 5 L to increase the production of biomass. Cell density was estimated in a light microscope (Leica, model DM LB 305) with a Neubauer chamber. The magnification used was 100x. Two-month old cultures were then utilized for the exposition experiment. Toxin content of cells harvested for the experiment were determined as described in section “Toxin analysis” and were found to produce OA (222.6 μg/g dw P. lima cells) and DTX1 (81.8 μg/g dw P. lima cells).
Mytilus galloprovincialis (length 75 ± 5 mm) were obtained from a local commercial provider. Mussels were transported in thermal boxes, refrigerated, to the laboratory, and cleaned with seawater. Mussels were distributed randomly in 6 aquaria filled with 50 L filtered sea water, with a total of 62 mussels per aquarium. Mussels were maintained in aquaria for 15 days with continuous aeration, temperature at 17°C ± 1°C, salinity 35.3 ± 0.2‰, pH 7,90 ± 0,10, dissolved oxygen 9,20 ± 0,10 mg/L and with artificial light (16 h/8 h, light/dark periods) of low irradiance. During acclimation animals were fed twice a day with 1.0 × 107 cells/L of freeze dried Isochrysis sp. (PhytoBloom ref. AADISS003) and Tetraselmis sp. (PhytoBloom AADTES003; Necton, Olhão, Portugal) at a cell ratio of 1:1. The water from aquaria was renewed every 2 days.
After acclimation, an experiment was carried out consisting in two sequential phases, 1- Uptake and 2- Depuration, with duration of 5 and 15 days, respectively. Two experimental mussel groups with distinct diets were outlined. Moreover, experimental groups were replicated in 3 aquaria (n = 3). Control mussels were fed during the whole experiment (uptake and depuration) with the mixture of Isochrysis sp. and Tetraselmis sp. (I + T) as described in 2.2. Another group of mussels was fed, during the uptake phase, with I + T supplemented with the toxic microalga (DST producer) P. lima at 1.0 × 106 cells/L, and during depuration with I + T. Mussels were fed daily with 2 doses of the diets described. Ten mussels were collected from each aquarium during uptake, at days 3 (T1) and 5 (T2), and during depuration at days 8 (T3), 14 (T4), and 20 (T5). After sampling, mussels were placed in ice for 5 min and dissected for study. Mussel whole body, gills and digestive gland were dissected and stored at −80°C.
An aliquot of P. lima cell culture was obtained by centrifugation (5,000 × g, 5 min.). Toxins were extracted in 3 mL MeOH and sonicated for 2 min at 25 W, 50% pulse duty cycle (Vibracell, Sonic & Materials, Newtown, CT, United States) in an ice bath. After centrifugation (10 min at 2,000 × g), the extract was filtered (0.22 μm) and stored at −20°C until LC-MSMS analysis.
Extraction of DSTs from mussels and their determination via liquid chromatography with tandem mass spectrometry (LC-MS/MS) detection was carried out following the Standardized Operating Procedure of the European Reference Laboratory for Marine Biotoxins (EURLMB) for the determination of marine lipophilic biotoxins in bivalve mollusks (EURLMB, 2015). A 2 g aliquot of shellfish meat homogenate was extracted with 9.0 mL MeOH by vortexing for 2 min. After centrifugation at 2,000 × g for 10 min, the resulting supernatant was transferred to a 30 mL centrifuge tube. The remaining tissue was re-extracted with 9.0 mL MeOH and homogenized with a Polytron mixer (Kinematica, Switzerland). After centrifugation, supernatants were combined and the volume adjusted to 20 mL with MeOH. An alkaline hydrolysis was performed to determine the total content of OA-group of toxins by converting the 7-O-acyl ester derivatives (DTX3) into their parent toxin. The hydrolysis was started by adding 313 μl of 2.5 M NaOH to a 2.5 mL aliquot of the methanolic extract in a test tube, which was homogenized for 30 s in the vortex and heated at 76°C for 40 min in a heating block. The sample was let to cool down until reaching room temperature and neutralized with 313 μl of 2.5 M HCl. The sample was vortex for 30 s and an aliquot was filtered through 0.2 μm syringe filter and 10 μl injected to the LC-MS/MS system.
The LC-MS/MS equipment consisted of an Agilent 1290 Infinity chromatograph coupled to a triple quadrupole mass spectrometer Agilent 6470 (Agilent Technologies, Germany). The chromatographic separation was conducted with a Zorbax SB-C8 RRHT column (2.1 × 50 mm, 1.8 μm), protected with a guard column (2.1 × 5 mm, 1.8 μm). The mobile phase A was water with 2 mM ammonium formate and 50 mM formic acid, and the mobile phase B was 95% acetonitrile with 2 mM ammonium formate and 50 mM formic acid. An elution gradient at a flow rate of 0.4 mL/min was used as follows: 0–3 min, gradient from 88 to 50% eluent A; 3–6.5 min gradient 50 to 10% eluent A; 6.5–8.9 min 10% eluent A; 8.9–10 min, gradient 10 to 88% eluent A. The detection was carried out in Multiple Reaction Monitoring (MRM) acquisition mode. Two MRM transitions were monitored in negative polarity: m/z 803 > 255 and m/z 803 > 563 for OA and m/z 817 > 255 and m/z 817 > 563 for DTX1, quantification and confirmation, respectively. Five calibration standard solutions ranging in concentration levels from 2.0 to 22.0 ng/mL with a correlation >0.990 was set up for quantification using a matrix match calibration and a certified OA and DTX1 reference standards purchased from CIFGA (Lugo, Spain).
Three mussels were collected per aquarium at days 3 and 5 (uptake) and day 14 (depuration) for proteomic analysis. The digestive gland and gills were dissected and stored at −80°C for further processing. Digestive gland and gill samples (approx. 0.2 g fw) were homogenized in Tris (100 mM), SDS 2% (w/v), dithiothreitol (0.1 M), pH 7.6 and protease inhibitors (complete protease cocktail tablets, Roche, Basel, Switzerland) with sonication (6 cycles of 5 s at 60 Hz), and incubated overnight as described previously (Campos et al., 2016). Samples were denatured with heat (95°C, 3 min) and clarified at 16,000 g for 20 min. The supernatants were recovered and total protein estimated with Bradford method. Proteins were digested following the filter aided sample preparation method described by Wisniewski et al. (2009), using centrifugal filter units with nominal molecular weight limit (NMWL) of 30 kDa (MRCF0R030, Millipore, Billerica, MA, United States). Protein samples (40 μg protein) were alkylated with iodoacetamide and digested with trypsin (recombinant, proteomics grade, Roche) for 16 h at 37°C, at an enzyme to protein ratio of 1:100 (w/w). Protein digests were recovered by centrifugal filtration, acidified with formic acid (FA; 10%, v/v), desalted and concentrated by reversed phase extraction (C18 Top-Tips, TT2C18, Glygen, Columbia, MD, United States). Before LC–MS/MS the peptides were recovered in 0.1% (v/v) FA.
Protein identification and quantitation was performed by nanoLC-MS/MS. This equipment is composed by an Ultimate 3000 liquid chromatography system coupled to a Q-Exactive Hybrid Quadrupole-Orbitrap mass spectrometer (Thermo Scientific, Bremen, Germany). Samples were loaded onto a trapping cartridge (Acclaim PepMap C18 100Å, 5 mm × 300 μm i.d., 160454, Thermo Scientific) in a mobile phase of 2% ACN, 0.1% FA at 10 μL/min. After 3 min loading, the trap column was switched in-line to a 50 cm by 75 μm inner diameter EASY-Spray column (ES803, PepMap RSLC, C18, 2 μm, Thermo Scientific, Bremen, Germany) at 250 nL/min. Separation was generated by mixing A: 0.1% FA, and B: 80% ACN, with the following gradient for total protein fraction: 2 min (2.5% B to 10% B), 50 min (10% B to 35% B), 8 min (35% B to 99% B), 5 min (50% B to 99% B), and 10 min (hold 99% B). Subsequently, the column was equilibrated with 2.5% B for 17 min. Data acquisition was controlled by Xcalibur 4.0 and Tune 2.9 software (Thermo Scientific, Bremen, Germany).
The mass spectrometer was operated in data-dependent (dd) positive acquisition mode alternating between a full scan (m/z 380–1580) and subsequent HCD MS/MS of the 10 most intense peaks from full scan (normalized collision energy of 27%). ESI spray voltage was 1.9 kV. Global settings: use lock masses best (m/z 445.12003), lock mass injection Full MS, chrom. peak width (FWHM) 15 s. Full scan settings: 70 k resolution (m/z 200), AGC target 3e6, maximum injection time 120 ms. dd settings: minimum AGC target 7e3, intensity threshold 6.4e4, charge exclusion: unassigned, 1, 8, >8, peptide match preferred, exclude isotopes on, dynamic exclusion 20 s. MS2 settings: microscans 1, resolution 35 k (m/z 200), AGC target 2e5, maximum injection time 110 ms, isolation window 2.0 m/z, isolation offset 0.0 m/z, spectrum data type profile. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE (Perez-Riverol et al., 2019) partner repository with the dataset identifier PXD022293.
The raw data was processed using Proteome Discoverer 2.4.0.305 software (Thermo Scientific) and searched against a custom protein database (46,797 protein sequences) obtained from the M. galloprovincialis transcriptome database (Campos et al., 2015), and against the UniProt KnowledgeBase (Bateman, 2019) using the taxonomy section of the genus Mytilus (72,020 protein sequences, accessed on September 30, 2020). The Sequest HT search engine was used to identify tryptic peptides. The ion mass tolerance was 10 ppm for precursor ions and 0.02 Da for fragment ions. Maximum allowed missing cleavage sites was set to 2. Cysteine carbamidomethylation was defined as constant modification. Methionine oxidation, protein N-terminus acetylation, Met-loss and Met-loss + acetyl were defined as variable modifications. Peptide confidence was set to high. The processing node Percolator was enabled with the following settings: maximum delta Cn 0.05; decoy database search target FDR 1%, validation based on q-value. Protein label free quantitation was performed with the Minora feature detector node at the processing step. Precursor ions quantification was performed at the processing step with the following parameters: unique plus razor peptides were considered for quantification and precursor abundance was based on intensity.
Protein functional classification (gene ontology) was carried out using the OmicsBox software v 1.4.111. Proteins were firstly blasted by running the CloudBlast option using BLASTp program against the non-redundant protein database at NCBI (nr database: ftp://ftp.ncbi.nlm.nih.gov/blast/db; accessed October 5th, 2020) and against the UniProt KnowledgeBase (accessed October 4th, 2020), setting a cut-off e-value of 1e–3. Proteins signatures were also annotated from the InterPro member databases (e.g., Pfam, PROSITE, PRINTS, ProDom, SMART, TIGRFAMs, PIRSF, SUPERFAMILY, Gene3D, and PANTHER) using the InterProScan software (Mitchell et al., 2019) integrated into the OmicsBox tool. Functional annotation of genes, and uncharacterized protein were also performed with eggNOG-Mapper 1.0.3 with EggNOG 5.0.0 (Huerta-Cepas et al., 2019). Genes names, COG/KOG (Clusters of Orthologous Groups of proteins/EuKaryotic Orthologous Groups)2 (Galperin et al., 2015), KEGG (Kyoto Encyclopedia of Genes and Genomes) pathways (Kanehisa et al., 2017), and KEGG KO (KEGG Ortholog database, https://www.kegg.jp/kegg/ko.html) annotations were derived from the eggNOG-mapper results. All annotation and the corresponding GO terms were then transferred to the sequences and eventually, merged with existing GO terms resulting from blast search.
All experiments were conducted with three replicates (n = 3). Toxin accumulation data is expressed as mean values ± standard deviation. The differences between treatments were evaluated for all data sets by a one-way ANOVA followed by a Tukey’s test. Data normality (Shapiro–Wilk’s test) and homoscedasticity (Levene’s test) was previously tested. A probability P value of less than 0.05 was considered significant. Statistical calculations and graph were performed using SPSS 25.0 (SPSS, Chicago, IL, United States).
Statistical analyses applied to the proteomic data was performed using R version 4.0.0. (R: a language and environment for statistical computing; Dessau and Pipper, 2008). The differentially expressed protein (DEP) and/or gene enrichment was assessed using the “DEP” R package (R Core Team, 2018; Zhang et al., 2018), based on the limma algorithm (Ritchie et al., 2015), considering those proteins identified in 2 out of 3 replicates of at least one condition, using “MinProb” as imputation function. Significant differences between exposed and control conditions for each tissue and sampled says were paired tested by a protein-wise linear models and empirical Bayes statistics. Finally, adjusted P value (alpha = 0.05), and the log2 fold change [lfc = log2(1.5), log2 fold change > 0.6] were used as a filter threshold to classify them as DEPs or enriched genes. Additional functional analyses to determine up/down regulated pathway was performed to the categorical GO terms corresponding to DEPs, using the hypergeometric distribution significance test (Fisher’s exact test, FDR < 0.05). Venn diagrams were used to identify the shared proteins using an online free tool, available at the webserver of the Bioinformatics and Evolutionary Genomics Center (BEG/Van de Peer Lab site, Ghent University, Belgium, http://bioinformatics.psb.ugent.be/webtools/Venn/).
The analysis of DSTs in the digestive gland and gills revealed accumulation of OA and DTX1 only in the former. A significant increase (p < 0.05) of OA concentration in mussels was detected between days 3 and 5 of the uptake phase. No significant differences were detected for DTX1 between the same days. The maximum amount recorded was 2819.2 ± 522.2 μg OA/kg and 1107.1 ± 267.9 μg DTX1/kg at the end of the uptake phase to the microalga P. lima. A significant decrease (p < 0.05) in the amount of OA and DTX1 accumulated in digestive gland with regard to day 5 (end of uptake period) was detected at days 14 and 20 of the experiment, during depuration (Figure 1). At the end of the depuration phase, only 16% and 47% of the total OA and DTX1, respectively, accumulated during the exposure to P. lima, remained in the digestive gland tissues. These results suggest that the elimination of OA is more efficient than DTX1 in this mussel species. Attending to the levels observed in the digestive gland and assuming that it generally represents4 12–14% of the mussels flesh (Blanco et al., 2007), it is estimated that the amounts in the whole body have exceeded more than 3 times the maximum limit of 160 μg OA eq./kg, allowed in shellfish for consumption.
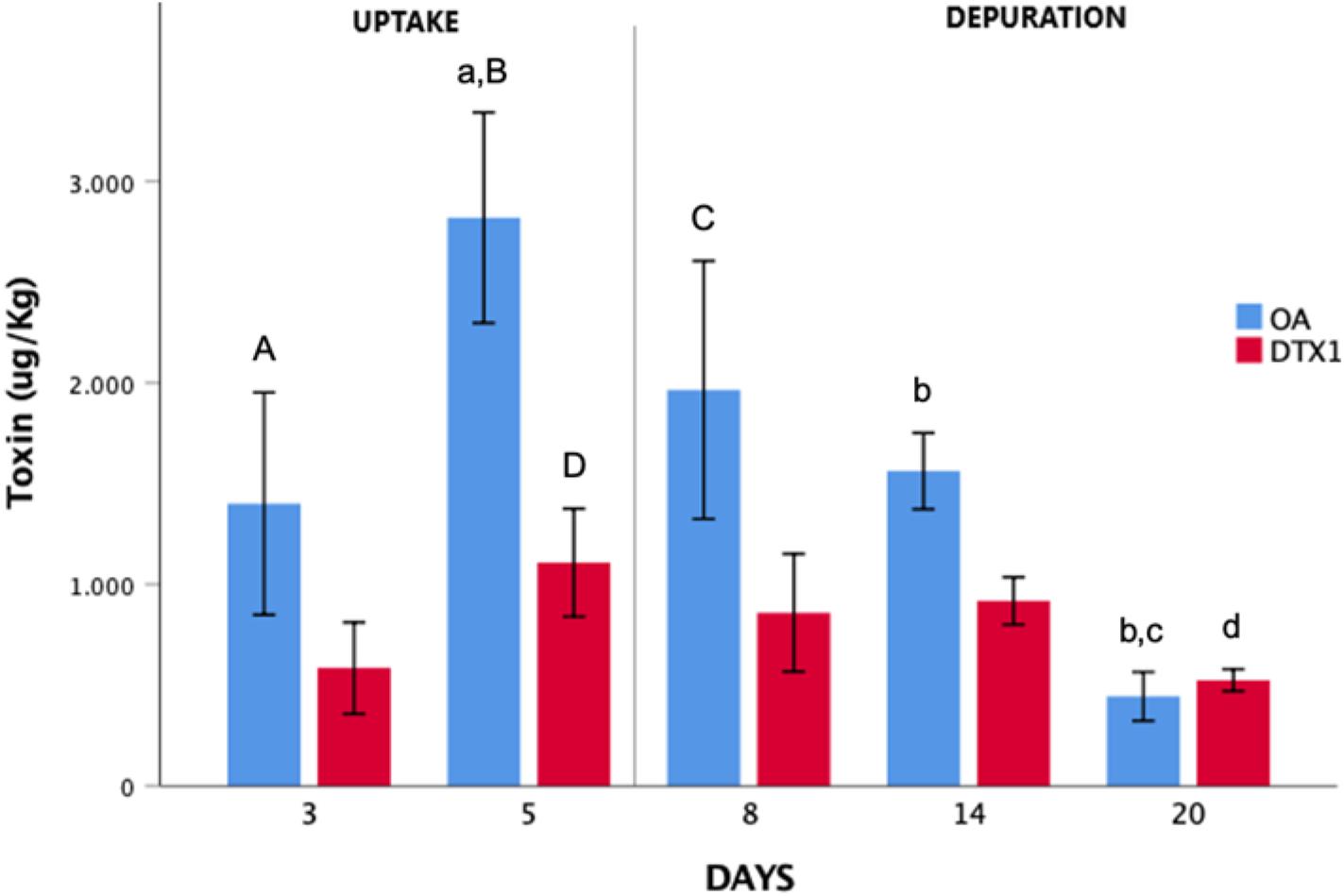
Figure 1. Time course of OA and DTX1 accumulation in digestive gland of mussels, during the uptake and depuration phases of the feeding experiment. Data are expressed as mean ± SD (n = 3). Significant differences between the same capital and lowercase letters (p < 0.05).
The shotgun proteomic analysis of 18 digestive gland and gill samples resulted in the identification of 2687 and 2717 proteins, respectively (Supplementary Table 1), considering the detection in at least one replicate. Protein relative abundances were subsequently analyzed using the “DEP” R package (Zhang et al., 2018), for detecting proteomic variations between control and mussels exposed to P. lima for 3 and 5 days, and after 9 days of depuration (14 days total length of experiment). This analysis revealed a total of 56 DEPs for the digestive gland, of which 34 were found after 3 days of exposure, whereas 18 and 9 proteins DEPs were revealed on day 5 of uptake and 9 days depuration, respectively. The overlapping DEPs are reported in the Venn diagram revealing no overlapped DEPs between all phases analyzed (3 and 5 days of uptake and 9 days depuration), four proteins shared between the uptake phases and only one DEP shared between 5 days and depuration (Figure 2A). One of the four DEPs overlapped in the uptake phases corresponded to p-glycoprotein (with NCBI accession A0SXG2), which possess ATPase activity and an ABC transporter domain. Another’s overlapped DEPs were an uncharacterized protein LOC111131611 (XP_022334930.1), related to VWA (von Willebrand factor type A) domain-containing protein, the hypothetical protein AM593_08637 (OPL20891.1) and the ras-related protein R-Ras2-like (XP_021348159.1). Noteworthy, the last-mentioned protein (OPL20891.1), likely belong to Ependymin super family, as well as the only protein shared between the 5-day uptake and the depuration phase (XP_013404178.1).
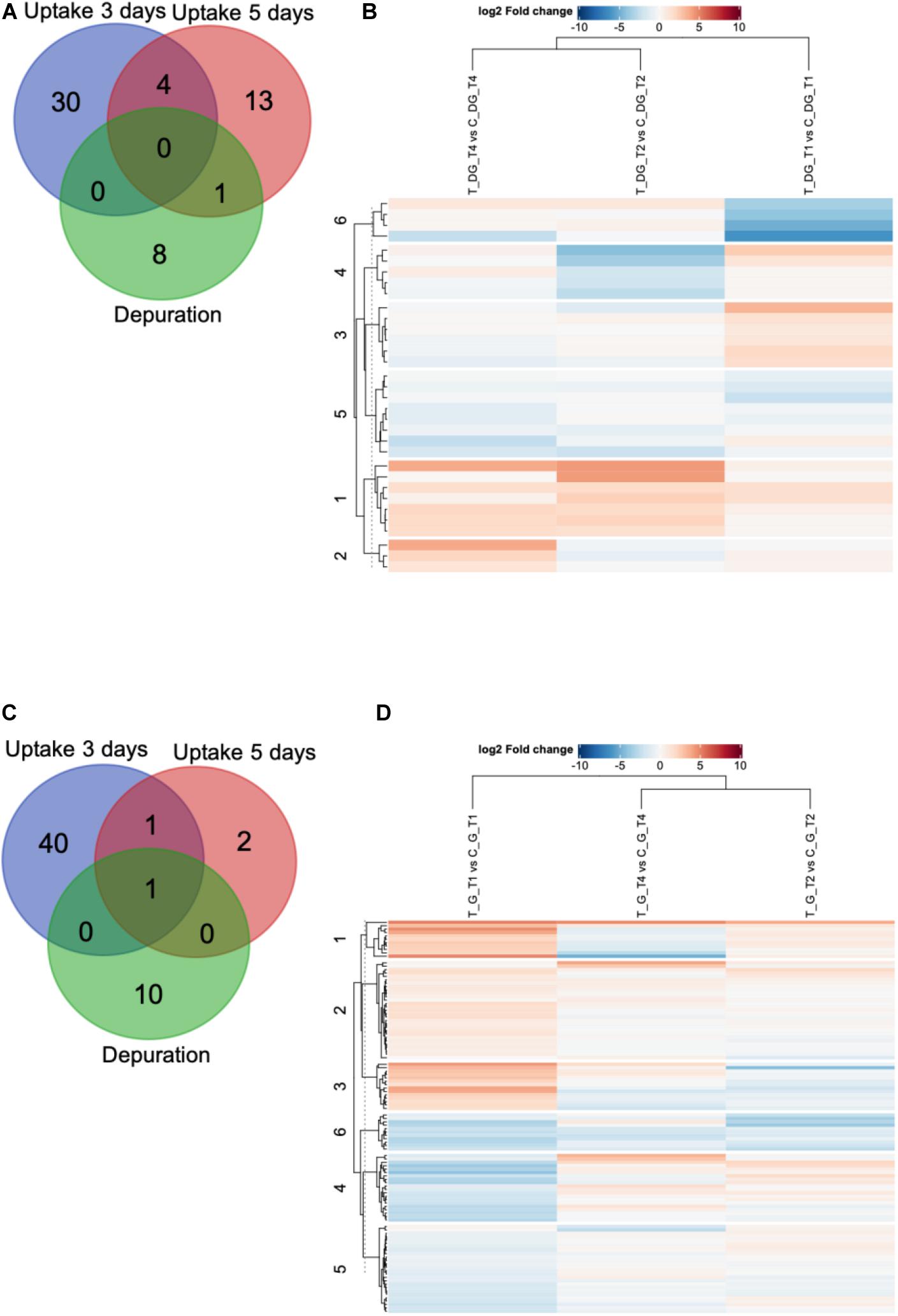
Figure 2. Venn diagram and heat map displaying the overlapping differentially expressed proteins (DEPs) found in mussel digestive glands (top panel) and gill (bottom panel). (A,C) Venn diagrams showing unique and shared DEPs identified for the uptake phase (3 days and 5 days uptake) and depuration (9 days depuration), and (B,D) heatmap depicting all DEPs between each comparison showing the differences as a log2 fold-changes expressed in a color map (red: upregulated; blue: downregulated). The rows represent DEPs which are clustered in six groups by k-means clustering (numbers on the left). The columns represent the tested groups 3 days uptake (T_DG/G_T1_vs_C_DG/G_T1), 5 days uptake (T_DG/G_T1_vs_C_DG/G_T1), and 9 days – depuration (T_DG/G_T4_vs_C_DG/G_T4).
In regard to gill tissues, 54 DEPs were detected from which 42, 4 and 11 were found after 3 and 5 uptake days and 9 days depuration, respectively. Like the Venn diagram shows (Figure 2C), one protein is shared between all phases analyzed (3 and 5 days of uptake and 9 days depuration) and corresponds to the predicted protein cubilin isoform X2 (XP_019922397.1). In the same way, the predicted UDP-glucose:glycoprotein glucosyltransferase 2 (XP_014777012.1) is shared by the uptake phases (3 and 5 days). No overlapped DEPs were found between the uptake phases individually (3 and 5 days) and 9 days depuration. The overall proteomics changes in the 3 analyzed phases of the experiment (3 and 5 days of uptake and 9 days depuration) showed that mussels experienced greater changes in the metabolism after 3 days exposure to P. lima. The high number of DEPs observed during this early phase for both organs highlights the sensitivity of mussels to this microalga. This early response, as well as all the protein changes dynamics through the subsequent phases could indeed be related, not only with the increase in the accumulation of toxins OA and DTX1 in tissues (especially for digestive gland), but also with the role of each organ as sites of interaction with bioavailable contaminants. Interestingly, and taking in account all the DEPs found for both organs in all phases, only one protein is shared by gills and digestive glands. This corresponds to sperm flagellar protein 2-like isoform X4 (XP_021376939.1) found for both tissues after 3 days uptake.
The overall proteomics changes of all DEPs found for both gill and digestive gland tissues regarding each tested group (exposed versus control group: T_DG/G_T1_vs_C_DG/G_T1: 3 days uptake; T_DG/G_T2_vs_C_DG/G_T2: 5 days uptake, T_DG/G_T4_vs_C_DG/G_T4: 9 days – depuration) are represented as log2 fold-changes in a heat-map, which are clustered in six groups by k-means clustering (Figures 2B,D). Coincidentally, the profile of expression of the DEPs in the early uptake phase (3 days) diverge from the latter time-points investigated (5 days uptake and 9 days depuration) for both organs. This result suggests that mussels show a response in the early up-take phase (3 days) increasingly different from the response in the 5 days uptake and 9 days depuration involving, putatively, different proteins and mechanisms.
The obtained contrast of the DEPs was graphically represented into a volcano plot depicting the adjust P values (- log10) against the fold changes (log2) for all phases and both organs (Figure 3). The gene enrichment analyses was performed with the “DEP” R package (Zhang et al., 2018), based on the limma algorithm (Ritchie et al., 2015). The corresponding sequences for each DEPs found were retrieved using the eggNOG-Mapper 1.0.3 with EggNOG 5.0.0 (Huerta-Cepas et al., 2019; see Supplementary Table 2). The complete list of DEPs and corresponding log2 fold-changes found for digestive gland and gills are shown in Supplementary Tables 3, 4, respectively. A hypergeometric distribution significance test (Fisher’s exact test, FDR < 0.05) was applied to the categorical GO terms of the corresponding DEPs for both digestive glands and gills. Among all phases analyzed, only two pathways in the 3 days uptake for the former organ were found to be enriched. These pathways are involved in carboxylic (GO:0046943) and organic acid transmembrane transporter activity (GO:0005342; Figure 4). Three proteins, ABCB11, SFXN1 and SLC6A7, matching to the up-regulated pathways, are indeed related to transmembrane transport, response to toxic substances and xenobiotics (Supplementary Table 3). More details about the function of these DEPs can be found in Supplementary Tables 2–4.
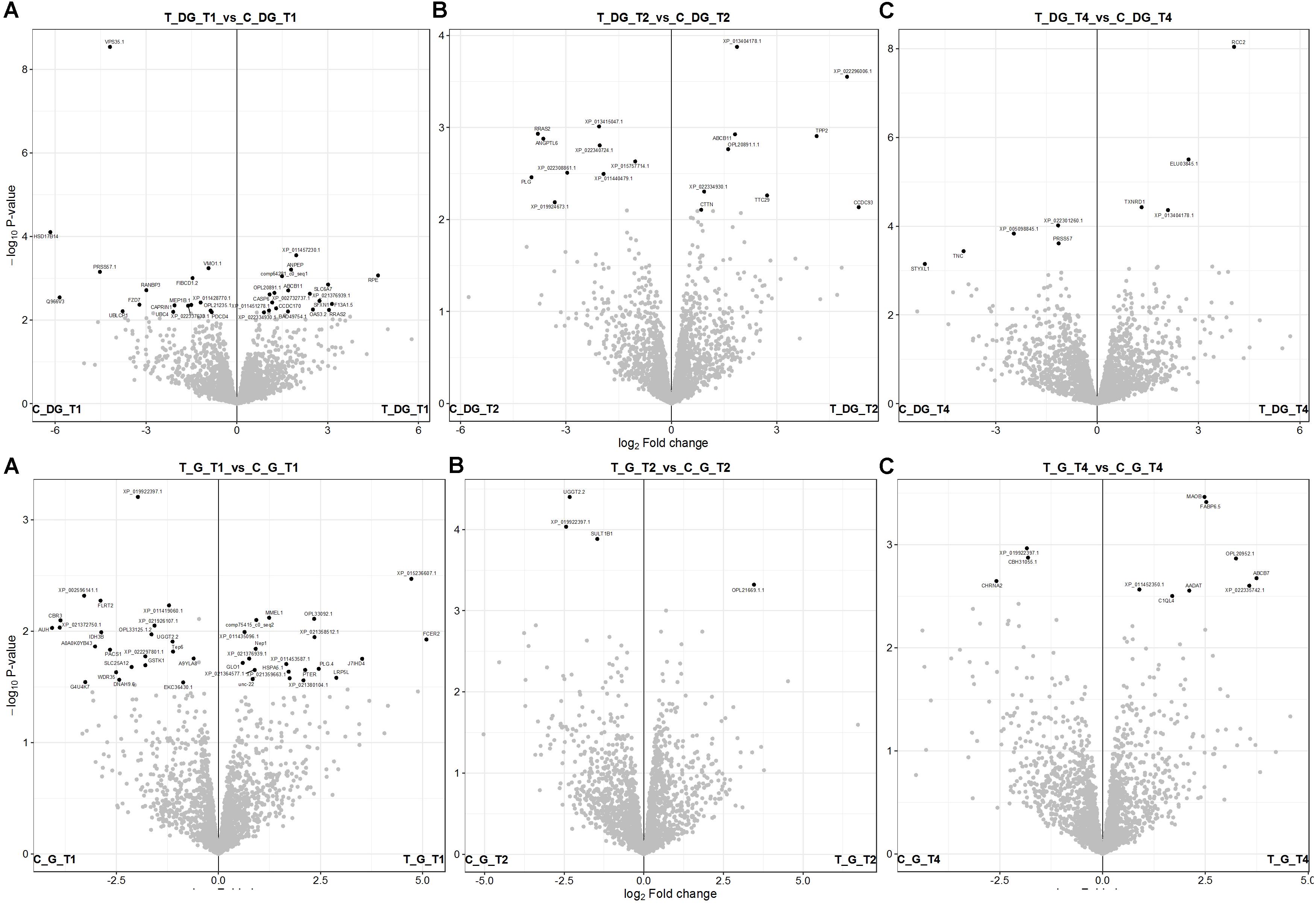
Figure 3. Volcano plot of differentialy expressed proteins (DEPs) found in the mussel digestive gland (top panel) and gill (bottom panel). Black points represent the DEPs showing the corresponding gene symbol, or the protein accession number when lacking the corresponding gene annotation. The axis depicted the P values (-log10) plotted against the fold changes (log2) between exposed group versus control for each tested condition: (A/a) 3 days uptake (T_DG/G_T1_vs_C_DG/G_T1), (B/b) 5 days uptake (T_DG/G_T1_vs_C_DG/G_T1), and (C/c) 9 days – depuration (T_DG/G_T4_vs_C_DG/G_T4).
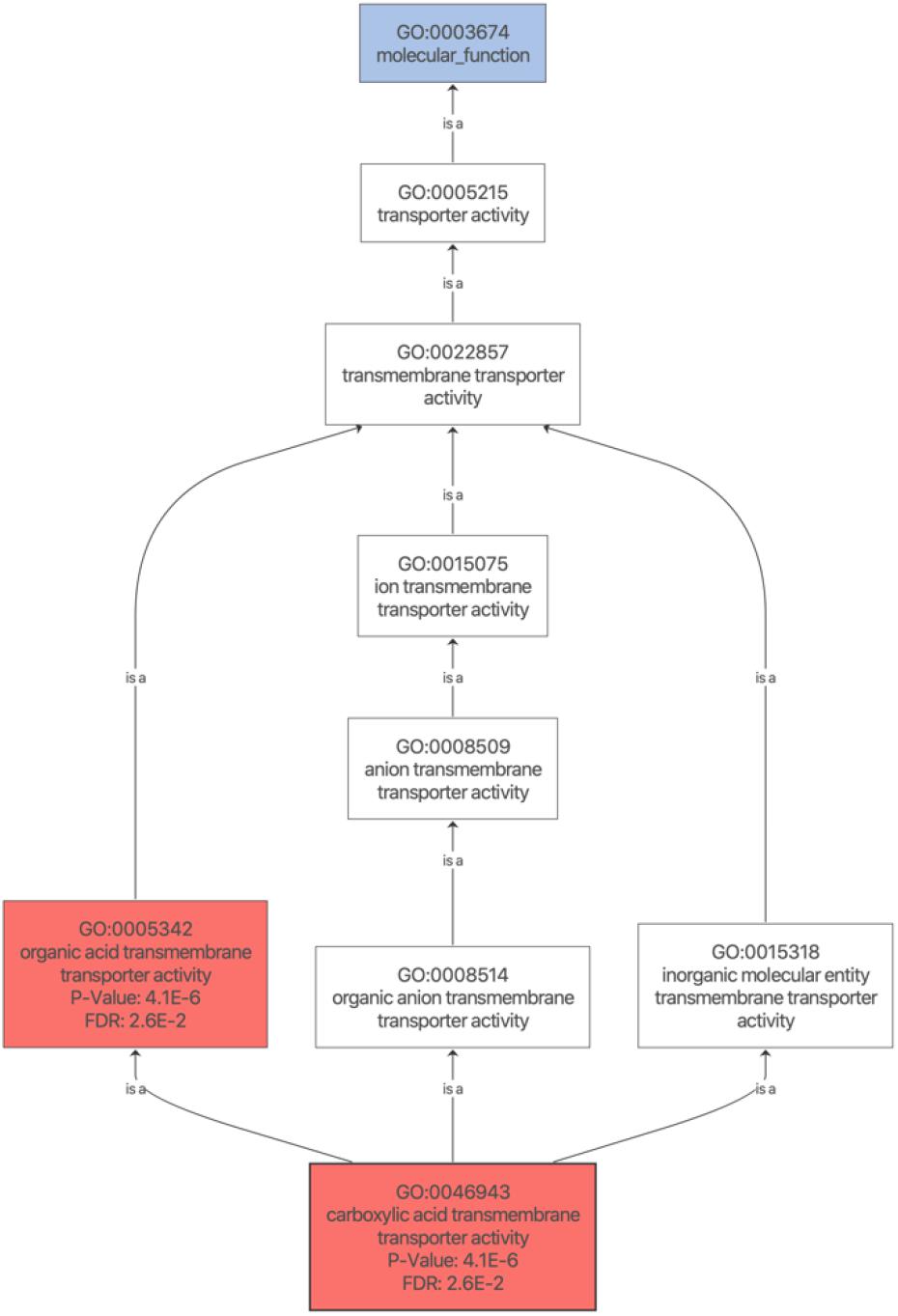
Figure 4. Enriched graph of the DEPs revealed by the hypergeometric distribution significance test. The enriched pathways (in red) and their corresponding categorical annotation (GO terms and GO description) found by applying the Fisher’s exact test (FDR < 0.05) to the DEPs found in the 3 days uptake for digestive gland tissues.
According to eggNOG annotation, the DEPs are functionally classified in Information Storage and Processing, Cellular Processes and Signaling, Metabolism and Poorly Characterized (Figure 5 and Supplementary Tables 2–4). The corresponding subcategories revealed a high number of DEPs poorly characterized (Figure 5). The category Cellular Processes and Signaling and Metabolism accounted for most of the sequences. Among this, the subcategories of “Signal transduction mechanisms,” “Posttranslational modification, protein turnover, chaperones,” and “Cytoskeleton” were more affected in the former, whereas “Secondary metabolites biosynthesis, transport and catabolism,” “Carbohydrate transport and metabolism,” and “Amino acid transport and metabolism” were more represented in the Metabolism category. The DEPs found in the gills showed a wider distribution among COG/KOG categories than the digestive glands. Likewise, some of the DEPs found in the gills appear to be functionally segregated by tissues and phases, given by their exclusive detection in “Replication, recombination and repair,” “Lipid transport and metabolism,” “Extracellular structures,” “Defense mechanisms,” and “Cell wall/membrane/envelope biogenesis.” Similarly occurred in the digestive glands with “Transcription” and “RNA processing and modification,” and “Cell cycle control, cell division, chromosome partitioning.” Besides, the distribution and relative abundances of the DEPs in some COG/KOG categories shows an antagonic pattern between up/down regulated DEPs of different tissues but in the same phase (Figure 5).
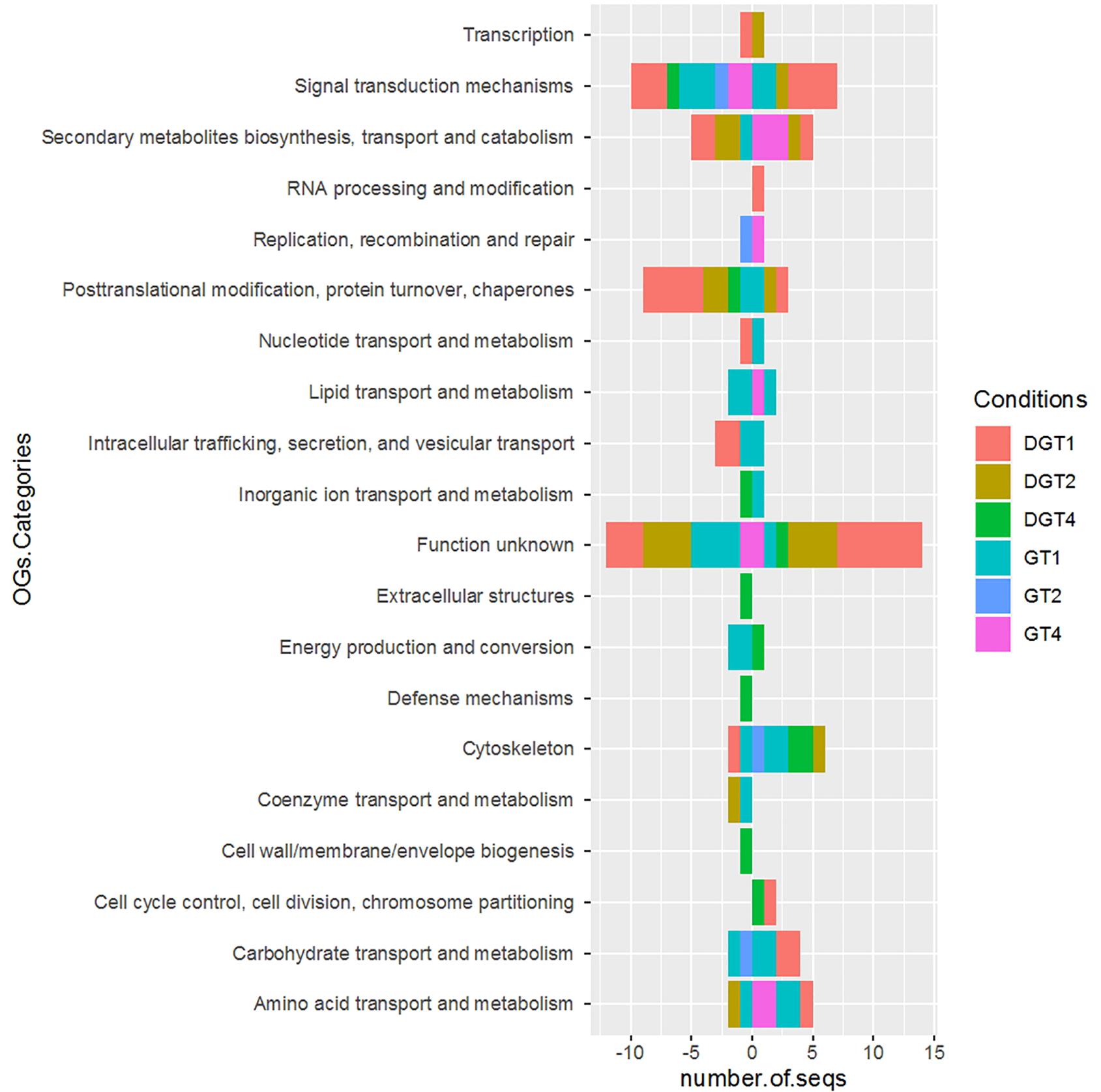
Figure 5. Functional annotation of the DEPs according to Orthologous Groups (OGs) categories obtained from eggNOG database. The divergent stacked bar represent the number of sequences (number.of.seqs) in the x-axis, corresponding to the up (positive values) and down (negative values) regulated DEPs matching the COG/KOG categories (y-axis). The phases studied for both tissues are represented with different colors bars, corresponding to the conditions (Conditions) in the right panel. The abbreviation of the conditions for the digestive gland are: 3 days uptake (DGT1), 5 days uptake (DGT2), and 9 days – depuration (DGT4), whereas for the gills are 3 days uptake (GT1), 5 days uptake (GT2), and 9 days – depuration (GT4).
The main aim of this study was to clarify the molecular mechanisms involved in bivalve‘s response to the toxic dinoflagellate P. lima. In this way, a shotgun proteomics approach was used to investigate the effect of DSTs accumulation in gill and digestive gland tissues of the Mediterranean mussel M. galloprovincialis during toxin uptake and depuration phases.
Such analyses were paired with the assessment of toxin accumulation in the two tissues, which revealed a substantial bioaccumulation of the toxins in the digestive gland. The toxin quantification results obtained in this work do not differ from previous ones that show rapid accumulation of OA and DTX by bivalves and the preferential accumulation of DSTs in this organ (Mafra et al., 2015; Blanco et al., 2017). Indeed, the accumulation of DSTs in organs such as gills and mantle has been shown to have little significance in the overall toxin accumulated in whole flesh, and to the estimation of the total toxicity of the animal. Differences of 7 to 10 orders were reported in total DSTs estimated in digestive gland and other organs (Bauder et al., 2001; Mafra et al., 2015; Kameneva et al., 2017). The rapid accumulation of DSTs is achieved, in laboratorial experiments, when bivalves keep constant filtration activity of the water and ingestion of toxic microalga (Bauder et al., 2001; Nielsen et al., 2020). Although rejection of live toxic cells as pseudofeces is an important pre-ingestion defense mechanism that enable bivalves to regulate the amount of toxin accumulated in the body, this response was not observed in the present work, i.e., mussels producing and excreting pseudofeces were not observed. The high amount of DSTs found in this study for digestive gland is consistent with other reports. For instance, scallops (Argopecten irradians) fed with 80 cells/ml P. lima for 2 days accumulated 1 μg/g DSTs (Bauder et al., 2001) and 6.8 μg/g DSTs were determined in the digestive gland from the mussel Crenomytilus grayanus fed in laboratory conditions with Prorocentrum foraminosum for 12 days (Kameneva et al., 2017). Also, 2.4 μg/g of DSTs were reported in naturally contaminated mussels Perna perna (Mafra et al., 2015). The maximum amount of DSTs accumulated in the digestive gland and the steady-state kinetics was characterized in scallops in a prolonged contamination study (Bauder et al., 2001). The authors demonstrated that the steady-state of OA and DTX accumulation can be achieved in a relatively short time, when scallops are constantly feeding on toxic microalga (Bauder et al., 2001). Moreover, the depuration rate of DSTs has been shown to vary considerably between bivalve species. Toxin half-life times of 1 day (P. perna), 2 days (C. gigas), and 5 days (C. virginica) have been estimated (Mafra et al., 2015). In mussels (M. edulis), half-life times of 5–6 days were reported for OA and DTX by Nielsen et al. (2016) and 11–20 days for total OA by Marcaillou et al. (2010). Such differences may reflect species-specific differences in feeding physiology and differential capacity of bivalve species to excrete and likely to metabolize DSTs. On the other hand, other variables such as temperature, and availability of food can also affect DSTs detoxification rates, since affect directly the metabolic activity of bivalves (Svensson, 2003; Marcaillou et al., 2010; Mafra et al., 2015).
The shotgun proteomic analysis applied in this study highlighted significant differences, both in terms of temporal response and biological processes involved, between gill and digestive gland tissues. The used time points included 3 and 5 days for the uptake phase and 14 days for depuration, as opposed to most previous works carried out in bivalves, which only took into account acute phase response. In one of those most recent studies, Prego-Faraldo et al. (2018) applied an RNA-Seq approach in the mussel M. galloprovincialis to analyze tissue-specific mussel transcriptome after early exposure (48 h) to a P. lima toxic strain (1 × 105 cell/L). Results were characterized by a higher number of differential expressed genes (DGEs) in gills compared to digestive gland and included transcripts involved in defense, immunity and metabolism. In the present study, similar effects were obtained after 3 days exposure but, in contrast, the number of DEPs was higher in digestive gland in comparison with gills after 5 days exposure and balanced between both tissues during depuration. Furthermore, and as expected, the total number of DEPs for both organs revealed to be much lower during this second phase in comparison with the exposure period. However, an unexpected fall of the number of DEPs was observed between day 3 and day 5 uptake for digestive gland and especially for gills, in which a near complete lack of changes occurred. The differences in DEPs strength between these two time points could be explained to some extent by the potential noise linked with the extreme levels of intraspecific genetic diversity, which leads to massive gene presence/absence variation (PAV; Gerdol et al., 2020). This phenomenon, which may affect up to 1/3rd of protein-coding genes, leads to misinterpretations of gene expression profiles, both in RNA-seq/microarray and proteomic analyses (Gerdol et al., 2020). However, in order to mitigate this issue, the experimental design of the present study comprehended pools of individuals (n = 3) and the use of statistical methods including FDR analysis to balance the random effect PAV is expected to have on the sampling of a few individuals and variability in the expression levels of proteins. Moreover, similar results on DEPs strength tendency had recently been observed in the digestive gland of P. viridis after P. lima exposure (2 × 106 cells/L; Dou et al., 2020). RNA-seq analysis showed a higher number of DEGs upon 6 h exposure to the dinoflagellate in comparison to 96 h along with no significant differences in mussels’ OA content in digestive glands between both time points (Dou et al., 2020). This apparent molecular adaptation to DSTs exposure and accumulation needs to be further explored in the future.
This proteomics work revealed responses in mussels during the uptake and depuration phases involving metabolism, cellular processes and signaling and information storage and processing for both gill and digestive gland tissues. Nevertheless, almost no common DEPs were found between organs in each functional category. This might indicate specific molecular processes associated to the role of each organ. In bivalve molluscs, the gills represent the primary sites of interaction with the bioavailable contaminants from the surrounding medium (water column, sediment) and/or the food ingested (Lemaire-Gony and Boudou, 1997). These biological barriers are involved in fundamental and physiological functions (respiration, osmoregulation, and nutrition; Lemaire-Gony and Boudou, 1997). The digestive gland is the major site of intracellular digestion (lysosomal) and absorption (Gosling, 2015). The exception among the identified DEPs was the sperm flagellar protein 2-like isoform X4 (XP_021376939.1) found for both tissues after 3 days uptake. It is known that natural spawning events in echinoderms are often correlated with other environmental conditions than temperature, such as phytoplankton blooms (Mercier and Hamel, 2009). A similar event could take place during the actual experiment, with P. lima acting as spawning trigger in mussels leading to the release of gametes into the water column and filtered again by mussels’ belonging to the same experimental group.
The functional annotation of DEPs from both tissues revealed an enrichment in functions related to carboxylic (GO:0046943) and organic acid transmembrane transporter activity (GO:0005342) up-regulated after 3 days uptake for digestive gland. Matching to the up-regulated pathways are three genes, namely ABCB11, SFXN1, and SLC6A7. The ABCB11 gene encodes for a membrane-associated protein (P-glycoprotein) member of the superfamily of ATP-binding cassette (ABC) transporters involved in the transport of drugs and xenobiotic substances. These proteins are expressed in different organs of bivalves (Huang et al., 2015a) and are known to respond to the presence of DST-producing P. lima (Huang et al., 2015b; Dou et al., 2020) and OA (Chi et al., 2018), suggesting a role in the elimination of DSTs. The Sideroflexin 1 protein encoded by SFXN1 gene is a mitochondrial serine transporter that mediates transport of serine into mitochondria, an important step of the one-carbon metabolism pathway. The protein encoded by SLC6A7 gene is known as Solute Carrier Family 6 Member 7, belonging to SLC gene superfamily and involved as membrane-bound transporters of amino acids and/or oligopeptides (He et al., 2009). In this way, functions related to transmembrane transport of carboxylic and organic acids assumed an extreme importance in the response of digestive gland to DSTs after 3 days uptake. Moreover, besides P-glycoprotein, other known DST-producing microalga and/or OA toxin gene/protein markers (Rey-Campos et al., 2020) were identified at this time point. Those include Caspase-2-like (XP_022316669.1) from antioxidant system and Fibrinogen-related protein (E5KXE9) from the immune system. Several cytoskeleton related proteins were also identified (OPL20891.1, XP_002732737.1, Q966V3). As an PP inhibitor, OA toxicity leads to the hyper-phosphorylation of cytoskeleton proteins and consequent cell morphological rearrangement and other adverse outcomes such as apoptosis. Among cytoskeleton proteins, the up-regulation of predicted ependymin-related protein 1-like (XP_002732737.1) has been related with cells intestine regeneration (Suárez-Castillo et al., 2004). In the same way, other relevant proteins were up-regulated at this time point for digestive gland. The coagulation factor XIII (F13A1) and 2 3-5-oligoadenylate synthase (OAS3) from the immune system, and with roles in other cellular processes like apoptosis, cell growth, differentiation and gene regulation. The ras-related protein R-Ras2-like (RRAS2), which is a GTP-binding protein with GTPase activity involved in the regulation of MAPK signaling pathway, promoting cell survival and inhibition of apoptosis (Bonni et al., 1999). The ribulose-phosphate 3-epimerase-like (RPE) and aminopeptidase N-like isoform X1 (ANPEP), both involved in carbohydrate and amino acid metabolism and transport. Responses related to these last proteins suggest that nutrient intake and digestion/transformation of nutrients were altered in this phase. Furthermore, the changes observed are more likely to be related with the composition of the diet than the presence of OA and DTX1. Indeed, the toxic diet offered to mussels during the uptake phase was constituted by non-toxic Isochrysis sp. and Tetraselmis sp. cells (freeze dried) and toxic P. lima cells. In contrast, the control diet was constituted only by the non-toxic Isochrysis sp. and Tetraselmis sp. Despite P. lima representing 1/10 of the diet (in terms of cell number), the differences in the nutrient composition between diets, due to the addition of the toxic microalga, may have led mussels to need to adjust the metabolism related to nutrition. It cannot be discarded also the hypothesis of the influence of OA and DTX1 in this response, since digestive gland accumulated high levels of these toxins. Most of the down-regulated proteins at 3 days uptake in digestive gland have functions related with signal transduction mechanisms and post-translational modification, protein turnover, chaperone functions. Among those are included programmed cell death protein 4-like (PDCD4), predicted frizzled-7-A (FZD7), predicted astacin-like (MEP1B), and putative ubiquitin-conjugating enzyme E2 4(UBC4), this last involved in ubiquitin mediated proteolysis. Interestingly, the ubiquitin-like domain-containing CTD phosphatase 1 (UBLCP1) with protein serine/threonine phosphatase activity is also down-regulated, which may be directly related to OA inhibition effects.
The following time point, 5 days uptake, was characterized by a much smaller number of DEPs in comparison with the previous one. Several proteins, including P-glycoprotein (ABCB11) and hypothetical protein AM593_08637, Ependymin domain (OPL20891.1) kept to be up-regulated. Adding up to these is the up-regulation of the predicted mammalian ependymin-related protein 1-like (XP_013404178.1) and cortactin isoform C (CTTN), both related with cytoskeleton organization and regeneration. In the same way, tripeptidyl-peptidase 2-like (TPP2) involved in ubiquitin-proteasome pathway, which is essential for many cellular processes, including the cell cycle, the regulation of gene expression, and responses to oxidative stress was also up-regulated. Likewise, the uncharacterized protein LOC111105848 (XP_022296006.1), known to be involved in gene transcription mechanisms which may be related to a positive regulation of these processes, consistent with a change in the metabolic condition of mussels and the restoration of protein synthesis processes after the initial impact (3 days uptake). The down-regulation of proteins including ras-related protein R-Ras2-like (RRAS2; previously up-regulated at 3 day uptake), predicted protein usf-like (XP_015757714.1), predicted cathepsin L1-like (XP_013415047.1), and Apolipoprotein (A0A3L5TUH3) denotes a clear disinvestment in regulation, signaling processes, and metabolism (carbohydrates and amino acids).
Depuration is characterized by the up-regulation of proteins associated to cytoskeleton organization, namely predicted mammalian ependymin-related protein 1-like (XP_013404178.1), previously also up-regulated in the end of the uptake phase and protein RCC2-like (RCC2), required for normal organization of the microtubule cytoskeleton in interphase cells. This phase is also characterized by responses in digestive gland related with oxidative stress and macromolecule transport. Thioredoxin reductase 1 (TXNRD1) was up-regulated, indicating oxidative stress in digestive gland cells induced, most likely, by DSTs. Surprisingly, this response occurs during depuration and not at the end of the uptake phase when DSTs reached highest concentration in this organ.
In gills, and after 3 days uptake, among the proteins of the antioxidant system that showed to be sensitive to DSTs are glutathione S-transferase sigma 1 (J7IHD4) and kappa 1-like isoform X1 (GSTK1). GSTs are phase II detoxifying enzymes that catalyze the covalent binding of glutathione to xenobiotics, reducing the toxicity of these compounds. Moreover, GSTs hold peroxidase activity and participate in the antioxidant defense (Birben et al., 2012; Martins et al., 2014). Although this family of enzymes does not seem to be directly involved in the metabolism of DSTs, bivalve GSTs showed to be sensitive to P. lima exposure (Zou et al., 2015). The modulation of these detoxification enzymes suggests the increase of ROS which may result in significant damage to cell structures. Severe molecular effects by OA in gills at this time point seems to be denoted by the modulation of the cytoskeleton proteins dynein gamma chain -flagellar outer arm-like isoform X2 (XP_021380104.1), major vault protein-like isoform X2 (XP_021364577.1) and predicted dynein beta chain – ciliary (DNAH9). This can lead to the impairment of respiratory cilia movement functions that can compromise the normal function of gills. Heat shock proteins are other known DST markers which are rapidly up-regulated in the presence of stressful stimuli. A byssal HSP-like protein 1 (HSPA5), a member of the heat shock protein 70 (HSP70) family was up-regulated in gills at 3 days uptake. Several proteins associated to signal transduction mechanisms have been also up-regulated. Those include Fc-e cd23 receptor (FCER2), which has essential roles in the regulation of IgE production and in the differentiation of B-cells; Low-density lipoprotein receptor-related protein 5-like protein (LRP5L), presenting Wnt-activated receptor activity; Twitchin (unc-22), a regulator of muscle contraction and relaxation; sodium/calcium exchanger 2-like isoform X8 (XP_021358512.1), presenting calmodulin binding activity. On the other hand, down-regulated proteins include predicted leucine-rich repeat-containing G-protein coupled receptor 4 (FLRT2) which presents functions in cell-cell adhesion, cell migration and axon guidance. Among proteins modulated with post-translational modification, protein turnover, chaperone functions are Kringle domain-containing protein (PLG), a plasminogen protein acting as a proteolytic factor in a variety of processes including inflammation and Cathepsin L (EKC36430.1), a lysosomal cysteine proteinase that plays a major role in intracellular protein catabolism. Many of the DEPs found for gills at day 3 uptake comprehend metabolic functions, namely carbohydrate, amino acid and lipid metabolism. Among these stand out the up-regulated lactoylglutathione lyase-like (GLO1), endothelin-converting enzyme homolog (Nep1), membrane metallo-endopeptidase-like 1 isoform X1 (MMEL1) and predicted carnitine O-acetyltransferase isoform X1 (XP_011435096.1). In contrast, predicted methylglutaconyl-CoA hydratase. mitochondrial isoform X2 (AUH), WD repeat-containing protein 35-like isoform X1 (WDR35) and predicted UDP-glucose:glycoprotein glucosyltransferase 1-like (UGGT2) are among the most down-regulated. Energy production and conversion is also affected according to the identified down-regulated predicted calcium-binding mitochondrial carrier protein Aralar2-like isoform X2 (SLC25A12) and predicted isocitrate dehydrogenase [NAD] subunit beta. mitochondrial-like (IDH3B).
The following time point, 5 days uptake, was characterized by a near lack of DEPs in comparison with the previous one. Among the four DEPs identified, tubulin chain beta-1 (OPL21669.1), a common cell structure marker of DSTs exposure in shellfish was the only up-regulated protein. The down-regulated proteins are associated with signal transduction mechanisms [predicted cubilin isoform X2 (XP_019922397.1)], carbohydrate metabolism and transport [predicted UDP-glucose:glycoprotein glucosyltransferase 1-like (UGGT2)] and replication, recombination and repair processes [sulfotransferase 1A1-like (SULT1B1)]. This last protein mediates sulfonation by increasing the water solubility of most compounds and consequent excretion, including xenobiotics as well as endogenous molecules.
Depuration is characterized by the up-regulation of mostly metabolic proteins. Among these stand out several proteins, including the predicted ATP-binding cassette sub-family B member 7, mitochondrial (ABCB7). This protein is a member of the MDR/TAP subfamily which are involved in multidrug resistance as well as antigen presentation. However, they are not often associated in previous studies to elimination of DSTs. Since OA was not detected in gills in this phase, probably their function might be more related to other activities, such as heme transporter activity. Among the found metabolic proteins are also included gastrotropin (FABP6), associated to lipid metabolism; the aminotransferase predicted kynurenine/alpha-aminoadipate, mitochondrial (AADAT) and the amine oxidase [flavin-containing]-like isoform X1 (MAOB). Another important protein up-regulated and associated to signal transduction mechanisms is Putative C1q domain containing protein MgC1q8 (C1QL4). It can present negative regulation of ERK1 and ERK2 cascade, which is a central signaling pathway that regulates a wide variety of stimulated cellular processes. These mainly include proliferation, differentiation, and survival, but apoptosis and stress response as well (Wortzel and Seger, 2011).
In this work, a shotgun proteomics approach was used to investigate the molecular mechanisms involved in the response of gills and digestive gland of the mussel M. galloprovincialis to a DST-producing P. lima strain. The assessment of toxin accumulation in the two tissues revealed a substantial bioaccumulation of OA and DTX1 only in the digestive gland. The maximum amount recorded was at the end of the uptake phase and, after depuration, toxin was still detected in the digestive gland tissues. Proteomic results for both organs presented similar dynamics which was characterized by a higher number of DEPs during the exposure phase in comparison with depuration. Surprisingly, the early uptake phase accounted for most responses during the exposure period. Furthermore, the profile of expression of the DEPs in the early uptake phase diverged from the latter time-points investigated. Results revealed tissue-specific responses with almost no common DEPs between both tissues. The hypergeometric distribution significance test revealed the up-regulation of carboxylic (GO:0046943) and organic acid transmembrane transporter activity (GO:0005342) pathways after 3 days uptake for digestive gland. In this last organ, most of the DEPs found for all phases were related with “cellular processes and signaling” and involving signal transduction mechanisms, cytoskeleton and post-translational modification, protein turnover, chaperone functions. In gills, the early uptake phase is characterized by a balance between DEPs related with “cellular processes and signaling” and “metabolism.” Regarding the former category, most of DEPs are also involved in the same subcategories found for digestive gland. At this time point, carbohydrate, amino acid and lipid metabolism assume a similar importance. During depuration, most of the DEPs found for gills stand for “metabolism,” involving mainly secondary metabolites biosynthesis, transport, and catabolism.
Despite the elevated accumulation of OA and DTX1 in mussel’s digestive gland, the proteomic results did not reveal significant molecular alterations linked to acute toxicity or severe molecular damage effects. With this in mind, it can be inferred that mussels show considerable tolerance to P. lima exposure and to accumulation of SDTs, and likely experienced only sub-toxic effects from these toxins.
The original contributions presented in the study are publicly available. This data can be found here: ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD022293.
JM, DD-P, AC, and PC: conceptualization. JM, DD-P, AC, PC, and VV: writing—original draft preparation. JM, DD-P, AC, PC, CA, AB, VV, and HO: writing—review and editing. JM, DD-P, AC, PC, CA, AB, and HO: methodology. JM, DD-P, AC, PC, CA, AB, HO, and VV: research. AC and PC: funding acquisition. All authors have read and agreed to the published version of the manuscript.
This work was funded by Portuguese Science Foundation (Fundação para a Ciência e a Tecnologia, FCT) and under the Projects MOREBIVALVES (PTDC/ASP-PES/31762/2017) and by the Strategic Funding UIDB/04423/2020 and UIDP/04423/2020 through national funds provided by FCT and European Regional Development Fund (ERDF), in the framework of the program PT2020. This work had also support from the Portuguese Mass Spectrometry Network, integrated in the National Roadmap of Research Infrastructures of Strategic Relevance (ROTEIRO/0028/2013 and LISBOA-01-0145-FEDER-022125).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2020.589822/full#supplementary-material
Bateman, A. (2019). UniProt: a worldwide hub of protein knowledge. Nucleic Acids Res. 47, D506–D515. doi: 10.1093/nar/gky1049
Bauder, A., Cembella, A., Bricelj, V., and Quilliam, M. (2001). Uptake and fate of diarrhetic shellfish poisoning toxins from the dinoflagellate Prorocentrum lima in the bay scallop Argopecten irradians. Mar. Ecol. Prog. Ser. 213, 39–52. doi: 10.3354/meps213039
Birben, E., Sahiner, U. M., Sackesen, C., Erzurum, S., and Kalayci, O. (2012). Oxidative stress and antioxidant defense. World Allergy Organ. J. 5, 9–19. doi: 10.1097/WOX.0b013e3182439613
Blanco, J. (2018). Accumulation of Dinophysis Toxins in bivalve molluscs. Toxins (Basel) 10:453. doi: 10.3390/toxins10110453
Blanco, J., Mariño, C., Martín, H., and Acosta, C. P. (2007). Anatomical distribution of diarrhetic shellfish poisoning (DSP) toxins in the mussel Mytilus galloprovincialis. Toxicon 50, 1011–1018. doi: 10.1016/j.toxicon.2007.09.002
Blanco, J., Martín, H., and Mariño, C. (2017). Reduction of diarrhetic shellfish poisoning (DSP) toxins accumulation in cultured mussels by means of rope clustering and hydrodynamic barriers. Aquaculture 479, 120–124. doi: 10.1016/j.aquaculture.2017.05.034
Bonni, A., Brunet, A., West, A. E., Datta, S. R., Takasu, M. A., and Greenberg, M. E. (1999). Cell survival promoted by the Ras-MAPK signaling pathway by transcription-dependent and -independent mechanisms. Science 286, 1358–1362. doi: 10.1126/science.286.5443.1358
Campos, A., Apraiz, I., Fonseca, R. R., Cristobal, S., da Fonseca, R. R., and Cristobal, S. (2015). Shotgun analysis of the marine mussel Mytilus edulis haemolymph proteome and mapping the innate immunity elements. Proteom. J. 15, 4021–4029. doi: 10.1002/pmic.201500118
Campos, A., Danielsson, G., Farinha, A. P., Kuruvilla, J., Warholm, P., and Cristobal, S. (2016). Shotgun proteomics to unravel marine mussel (Mytilus edulis) response to long-term exposure to low salinity and propranolol in a Baltic Sea microcosm. J. Proteom. 137, 97–106. doi: 10.1016/j.jprot.2016.01.010
Carvalho Pinto-Silva, C. R., Creppy, E. E., and Matias, W. G. (2005). Micronucleus test in mussels Perna perna fed with the toxic dinoflagellate Prorocentrum lima. Arch. Toxicol. 79, 422–426. doi: 10.1007/s00204-004-0645-1
Carvalho Pinto-Silva, C. R., Ferreira, J. F., Costa, R. H. R., Belli Filho, P., Creppy, E. E., and Matias, W. G. (2003). Micronucleus induction in mussels exposed to okadaic acid. Toxicon 41, 93–97. doi: 10.1016/S0041-0101(02)00214-213
Chi, C., Giri, S., Jun, J., Kim, S., Kim, H., Kang, J., et al. (2018). Detoxification- and immune-related transcriptomic analysis of gills from bay scallops (argopecten irradians) in response to algal toxin okadaic acid. Toxins (Basel) 10:308. doi: 10.3390/toxins10080308
Chi, C., Giri, S. S., Jun, J. W., Kim, H. J., Kim, S. W., Yun, S., et al. (2017). Effects of algal toxin okadaic acid on the non-specific immune and antioxidant response of bay scallop (Argopecten irradians). Fish Shellfish Immunol. 65, 111–117. doi: 10.1016/j.fsi.2017.03.031
Chi, C., Giri, S. S., Jun, J. W., Kim, H. J., Yun, S., Kim, S. G., et al. (2016). Marine toxin okadaic acid affects the immune function of bay scallop (Argopecten irradians). Molecules 21:1108. doi: 10.3390/molecules21091108
Dessau, R. B., and Pipper, C. B. (2008). R - En Programpakke til Statistisk Databehandling og Grafik. Ugeskr. Laeger 170, 328–330. Available online at: http://europepmc.org/article/med/18252159 (accessed November 7, 2020).
Dou, M., Jiao, Y. H., Zheng, J.-W., Zhang, G., Li, H.-Y., Liu, J. S., et al. (2020). De novo transcriptome analysis of the mussel Perna viridis after exposure to the toxic dinoflagellate Prorocentrum lima. Ecotoxicol. Environ. Saf. 192:110265. doi: 10.1016/j.ecoenv.2020.110265
EURLMB, (2015). EU-Harmonised Standard Operating Procedure for Determination of Lipophilic Marine Biotoxins in Molluscs by LC-MS/MS. Vigo: EURLMB.
Florez-Barros, F., Prado-Alvarez, M., Mendez, J., and Fernandez-Tajes, J. (2011). Evaluation of genotoxicity in gills and hemolymph of clam ruditapes decussatus fed with the toxic dinoflagellate Prorocentrum lima. J. Toxicol. Environ. Heal. A Curr. Issues 74, 971–979. doi: 10.1080/15287394.2011.582025
Galperin, M. Y., Makarova, K. S., Wolf, Y. I., and Koonin, E. V. (2015). Expanded Microbial genome coverage and improved protein family annotation in the COG database. Nucleic Acids Res. 43, D261–D269. doi: 10.1093/nar/gku1223
García, C., González, V., Cornejo, C., Palma-Fleming, H., and Lagos, N. (2004). First evidence of Dinophysistoxin-1 ester and carcinogenic polycyclic aromatic hydrocarbons in smoked bivalves collected in the Patagonia fjords. Toxicon 43, 121–131. doi: 10.1016/j.toxicon.2003.10.028
Gerdol, M., Moreira, R., Cruz, F., Gómez-Garrido, J., Vlasova, A., Rosani, U., et al. (2020). Massive gene presence-absence variation shapes an open pan-genome in the Mediterranean mussel. Genome Biol. 21:275. doi: 10.1186/s13059-020-02180-3
Gómez, F. (2005). A list of free-living dinoflagellate species in the world’s oceans. Acta Bot. Croat. 64, 129–212.
Gosling, E. (2015). Marine Bivalve Molluscs: Second Edition. Hoboken, NJ: Wiley Blackwell. doi: 10.1002/9781119045212
Hanana, H., Talarmin, H., Pennec, J. P., Droguet, M., Morel, J., and Dorange, G. (2012). Effect of okadaic acid on cultured clam heart cells: involvement of MAPkinase pathways. Biol. Open 1, 1192–1199. doi: 10.1242/bio.20122170
He, L., Vasiliou, K., and Nebert, D. W. (2009). Analysis and update of the human solute carrier (SLC) gene superfamily. Hum. Genomics 3, 195–206. doi: 10.1186/1479-7364-3-2-195
Huang, L., Liu, S. L., Zheng, J. W., Li, H. Y., Liu, J. S., and Yang, W. D. (2015a). P-glycoprotein and its inducible expression in three bivalve species after exposure to Prorocentrum lima. Aquat. Toxicol. 169, 123–132. doi: 10.1016/j.aquatox.2015.10.012
Huang, L., Zou, Y., Weng, H. W., Li, H. Y., Liu, J. S., and Yang, W. D. (2015b). Proteomic profile in Perna viridis after exposed to Prorocentrum lima, a dinoflagellate producing DSP toxins. Environ. Pollut. 196, 350–357. doi: 10.1016/j.envpol.2014.10.019
Huang, L., Wang, J., Chen, W. C., Li, H. Y., Liu, J. S., Jiang, T., et al. (2014). P-glycoprotein expression in Perna viridis after exposure to Prorocentrum lima, a dinoflagellate producing DSP toxins. Fish Shellfish Immunol. 39, 254–262. doi: 10.1016/j.fsi.2014.04.020
Huerta-Cepas, J., Szklarczyk, D., Heller, D., Hernández-Plaza, A., Forslund, S. K., Cook, H., et al. (2019). EggNOG 5.0: a hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 47, D309–D314. doi: 10.1093/nar/gky1085
JØrgensen, K., Scanlon, S., and Jensen, L. B. (2005). Diarrhetic shellfish poisoning toxin esters in Danish blue mussels and surf clams. Food Addit. Contam. 22, 743–751. doi: 10.1080/02652030500136928
Kacem, I., Bouaïcha, N., and Hajjem, B. (2010). Comparison of okadaic acid profiles in mussels and oysters collected in mediterranean lagoon. Tunisia. Int. J. Biol. 2:238. doi: 10.5539/ijb.v2n2p238
Kameneva, P. A., Krasheninina, E. A., Slobodskova, V. V., Kukla, S. P., and Orlova, T. Y. (2017). Accumulation and tissue distribution of dinophysitoxin-1 and dinophysitoxin-3 in the mussel crenomytilus grayanus feeding on the benthic dinoflagellate prorocentrum foraminosum. Mar. Drugs 15:330. doi: 10.3390/md15100330
Kanehisa, M., Furumichi, M., Tanabe, M., Sato, Y., and Morishima, K. (2017). KEGG: new perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 45, D353–D361. doi: 10.1093/nar/gkw1092
Konoki, K., Onoda, T., Watanabe, R., Cho, Y., Kaga, S., Suzuki, T., et al. (2013). In Vitro acylation of okadaic acid in the presence of various bivalves’ extracts. Mar. Drugs 11, 300–315. doi: 10.3390/md11020300
Lemaire-Gony, S., and Boudou, A. (1997). Mantle and gill fine structure in the freshwater Asiatic clam, Corbicula fluminea (Muller). Ann. Limnol. 33, 163–178. doi: 10.1051/limn/1997016
Lindholm, T. (1985). Mesodinium rubrum-a unique photosynthetic ciliate. Adv. Aquat. Microbiol. 3, 1–48.
Lopes, V. M., Costa, P. R., and Rosa, R. (2019). “Effects of harmful algal bloom toxins on marine organisms,” in Ecotoxicology of Marine Organisms. B Duarte, M Isabel, and V Caçador, (Boca Raton, FL: CRC Press), 42–88. doi: 10.1201/b22000-4
MacKenzie, L., Holland, P., McNabb, P., Beuzenberg, V., Selwood, A., and Suzuki, T. (2002). Complex toxin profiles in phytoplankton and Greenshell mussels (Perna canaliculus), revealed by LC-MS/MS analysis. Toxicon 40, 1321–1330. doi: 10.1016/S0041-0101(02)00143-5
Mafra, L. L., Ribas, T., Alves, T. P., Proença, L. A. O., Schramm, M. A., Uchida, H., et al. (2015). Differential okadaic acid accumulation and detoxification by oysters and mussels during natural and simulated Dinophysis blooms. Fish. Sci. 81, 749–762. doi: 10.1007/s12562-015-0882-7
Marcaillou, C., Haure, J., Mondeguer, F., Courcoux, A., Dupuy, B., and Pénisson, C. (2010). Aquatic living resources effect of food supply on the detoxification in the blue mussel, Mytilus edulis, contaminated by diarrhetic shellfish toxins. Aquat. Living Resour. 23, 255–266. doi: 10.1051/alr/2010026
Marr, J. C., Hu, T., Pleasance, S., Quilliam, M. A., and Wright, J. L. C. (1992). Detection of new 7-O-acyl derivatives of diarrhetic shellfish poisoning toxins by liquid chromatography-mass spectrometry. Toxicon 30, 1621–1630. doi: 10.1016/0041-0101(92)90034-3
Martins, J. C., Campos, A., Osório, H., da Fonseca, R., and Vasconcelos, V. (2014). Proteomic profiling of cytosolic glutathione transferases from three bivalve species: corbicula fluminea, Mytilus galloprovincialis and Anodonta cygnea. Int. J. Mol. Sci. 15, 1887–1900. doi: 10.3390/ijms15021887
McCarthy, M., O’Halloran, J., O’Brien, N. M., and van Pelt, F. F. N. A. M. (2014). Does the marine biotoxin okadaic acid cause DNA fragmentation in the blue mussel and the pacific oyster? Mar. Environ. Res. 101, 153–160. doi: 10.1016/j.marenvres.2014.09.009
Mercier, A., and Hamel, J. F. (2009). Endogenous and exogenous control of gametogenesis and spawning in echinoderms. Adv. Mar. Biol. 55, xi–xii, 1–291. doi: 10.1016/S0065-2881(09)55001-8
Mitchell, A. L., Attwood, T. K., Babbitt, P. C., Blum, M., Bork, P., Bridge, A., et al. (2019). InterPro in 2019: improving coverage, classification and access to protein sequence annotations. Nucleic Acids Res. 47, D351–D360. doi: 10.1093/nar/gky1100
Neves, R. A. F., Santiago, T. C., Carvalho, W. F., Silva, E., dos, S., da Silva, P. M., et al. (2019). Impacts of the toxic benthic dinoflagellate Prorocentrum lima on the brown mussel Perna perna: shell-valve closure response, immunology, and histopathology. Mar. Environ. Res. 146, 35–45. doi: 10.1016/j.marenvres.2019.03.006
Nielsen, L. T., Hansen, P. J., Krock, B., and Vismann, B. (2016). Accumulation, transformation and breakdown of DSP toxins from the toxic dinoflagellate Dinophysis acuta in blue mussels. Mytilus edulis. Toxicon 117, 84–93. doi: 10.1016/j.toxicon.2016.03.021
Nielsen, L. T., Krock, B., and Hansen, P. J. (2013). Production and excretion of okadaic acid, pectenotoxin-2 and a novel dinophysistoxin from the DSP-causing marine dinoflagellate Dinophysis acuta – effects of light, food availability and growth phase. Harmful Algae 23, 34–45. doi: 10.1016/j.hal.2012.12.004
Nielsen, P., Krock, B., Hansen, P. J., and Vismann, B. (2020). Effects of the DSP-toxic dinoflagellate Dinophysis acuta on clearance and respiration rate of the blue mussel, Mytilus edulis. PLoS One 15:e0230176. doi: 10.1371/journal.pone.0230176
Perez-Riverol, Y., Csordas, A., Bai, J., Bernal-Llinares, M., Hewapathirana, S., Kundu, D. J., et al. (2019). The PRIDE database and related tools and resources in 2019: improving support for quantification data. Nucleic Acids Res. 47, D442–D450. doi: 10.1093/nar/gky1106
Prado-Alvarez, M., Flórez-Barrós, F., Méndez, J., and Fernandez-Tajes, J. (2013). Effect of okadaic acid on carpet shell clam (Ruditapes decussatus) haemocytes by in vitro exposure and harmful algal bloom simulation assays. Cell Biol. Toxicol. 29, 189–197. doi: 10.1007/s10565-013-9246-1
Prado-Alvarez, M., Flórez-Barrós, F., Sexto-Iglesias, A., Méndez, J., and Fernandez-Tajes, J. (2012). Effects of okadaic acid on haemocytes from Mytilus galloprovincialis: a comparison between field and laboratory studies. Mar. Environ. Res. 81, 90–93. doi: 10.1016/j.marenvres.2012.08.011
Prego-Faraldo, M. V., Martínez, L., and Méndez, J. (2018). RNA-Seq analysis for assessing the early response to DSP Toxins in Mytilus galloprovincialis Digestive Gland and Gill. Toxins (Basel) 10:417. doi: 10.3390/toxins10100417
Prego-Faraldo, M. V., Valdiglesias, V., Laffon, B., Eirín-López, J. M., and Méndez, J. (2015). In vitro analysis of early genotoxic and cytotoxic effects of okadaic acid in different cell types of the mussel Mytilus galloprovincialis. J. Toxicol. Environ. Health A 78, 814–824. doi: 10.1080/15287394.2015.1051173
Prego-Faraldo, M. V., Valdiglesias, V., Laffon, B., Mendez, J., and Eirin Lopez, J. M. (2016). Early genotoxic and cytotoxic effects of the toxic dinoflagellate Prorocentrum lima in the mussel Mytilus galloprovincialis. Toxins (Basel) 8:159. doi: 10.3390/toxins8060159
Prego-Faraldo, M. V., Vieira, L. R., Eirin-Lopez, J. M., Méndez, J., and Guilhermino, L. (2017). Transcriptional and biochemical analysis of antioxidant enzymes in the mussel Mytilus galloprovincialis during experimental exposures to the toxic dinoflagellate Prorocentrum lima. Mar. Environ. Res. 129, 304–315. doi: 10.1016/j.marenvres.2017.06.009
Rey-Campos, M., Novoa, B., Pallavicini, A., Gerdol, M., and Figueras, A. (2020). Comparative genomics reveals a significant sequence variability of myticin genes in Mytilus galloprovincialis. Biomolecules 10, 1–18. doi: 10.3390/biom10060943
R Core Team, (2018). R: A Language and Environment for Statistical Computing. Available online at: https://www.gbif.org/pt/tool/81287/r-a-language-and-environment-for-statistical-computing (accessed November 7, 2020).
Ritchie, M. E., Phipson, B., Wu, D., Hu, Y., Law, C. W., Shi, W., et al. (2015). Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 43:e47. doi: 10.1093/nar/gkv007
Romero-Geraldo, R., de, J., García-Lagunas, N., and Hernández-Saavedra, N. Y. (2016). Crassostrea gigas exposure to the dinoflagellate Prorocentrum lima: histological and gene expression effects on the digestive gland. Mar. Environ. Res. 120, 93–102. doi: 10.1016/j.marenvres.2016.07.011
Rossignoli, A. E., Fernández, D., Regueiro, J., Mariño, C., and Blanco, J. (2011). Esterification of okadaic acid in the mussel Mytilus galloprovincialis. Toxicon 57, 712–720. doi: 10.1016/j.toxicon.2011.02.003
Shumway, S. E. (1995). Phycotoxin-related shellfish poisoning: bivalve molluscs are not the only vectors. Rev. Fish. Sci. 3, 1–31. doi: 10.1080/10641269509388565
Simães, E., Vieira, R. C., Schramm, M. A., Mello, D. F., De Almeida Pontinha, V., Da Silva, P. M., et al. (2015). Impact of harmful algal blooms (Dinophysis acuminata) on the immune system of oysters and mussels from Santa Catarina. Brazil. J. Mar. Biol. Assoc. U. K. 95, 773–781. doi: 10.1017/S0025315414001702
Suárez-Castillo, E. C., Medina-Ortíz, W. E., Roig-López, J. L., and García-Arrarás, J. E. (2004). Ependymin, a gene involved in regeneration and neuroplasticity in vertebrates, is overexpressed during regeneration in the echinoderm Holothuria glaberrima. Gene 334, 133–143. doi: 10.1016/j.gene.2004.03.023
Suzuki, T., and Mitsuya, T. (2001). Comparison of dinophysistoxin-1 and esterified dinophysistoxin-1 (dinophysistoxin-3) contents in the scallop patinopecten yessoensis and the mussel Mytilus galloprovincialis. Toxicon 39, 905–908. doi: 10.1016/S0041-0101(00)00205-1
Svensson, S. (2003). Depuration of okadaic acid (Diarrhetic Shellfish Toxin) in mussels, Mytilus edulis (Linnaeus), feeding on different quantities of nontoxic algae. Aquaculture 218, 277–291. doi: 10.1016/S0044-8486(02)00504-5
Torgersen, T., Wilkins, A. L., Rundberget, T., and Miles, C. O. (2008). Characterization of fatty acid esters of okadaic acid and related toxins in blue mussels (Mytilus edulis) from Norway. Rapid Commun. Mass Spectrom. 22, 1127–1136. doi: 10.1002/rcm.3490
Vale, P. (2004). Differential dynamics of dinophysistoxins and pectenotoxins between blue mussel and common cockle: a phenomenon originating from the complex toxin profile of Dinophysis acuta. Toxicon 44, 123–134. doi: 10.1016/j.toxicon.2004.04.002
Vale, P. (2006). Differential dynamics of dinophysistoxins and pectenotoxins, part II: offshore bivalve species. Toxicon 47, 163–173. doi: 10.1016/j.toxicon.2005.10.009
Vale, P., Antónia, M., and Sampayo, M. (1999). Esters of okadaic acid and dinophysistoxin-2 in Portuguese bivalves related to human poisonings. Toxicon 37, 1109–1121. doi: 10.1016/S0041-0101(98)00247-5
Vale, P., Botelho, M. J., Rodrigues, S. M., Gomes, S. S., Sampayo, M. A., and de, M. (2008). Two decades of marine biotoxin monitoring in bivalves from Portugal (1986-2006): a review of exposure assessment. Harmful Algae 7, 11–25. doi: 10.1016/j.hal.2007.05.002
Vale, P., and Maria, M. A. (2002). Esterification of DSP toxins by Portuguese bivalves from the Northwest coast determined by LC-MS – a widespread phenomenon. Toxicon 40, 33–42. doi: 10.1016/S0041-0101(01)00183-0
Wei, X. M., Lu, M. Y., Duan, G. F., Li, H. Y., Liu, J. S., and Yang, W. D. (2019). Responses of CYP450 in the mussel Perna viridis after short-term exposure to the DSP toxins-producing dinoflagellate Prorocentrum lima. Ecotoxicol. Environ. Saf. 176, 178–185. doi: 10.1016/j.ecoenv.2019.03.073
Wisniewski, J. R., Zougman, A., Nagaraj, N., and Mann, M. (2009). Universal sample preparation method for proteome analysis. Nat. Methods 6, 359–362. doi: 10.1038/nmeth.1322
Wortzel, I., and Seger, R. (2011). The ERK cascade: distinct functions within various subcellular organelles. Genes Cancer 2, 195–209. doi: 10.1177/1947601911407328
Yasumoto, T., Murata, M., Oshima, Y., Sano, M., Matsumoto, G. K., and Clardy, J. (1985). Diarrhetic shellfish toxins. Tetrahedron 41, 1019–1025. doi: 10.1016/S0040-4020(01)96469-5
Zhang, X., Smits, A. H., Van Tilburg, G. B. A., Ovaa, H., Huber, W., and Vermeulen, M. (2018). Proteome-wide identification of ubiquitin interactions using UbIA-MS. Nat. Protoc. 13, 530–550. doi: 10.1038/nprot.2017.147
Keywords: okadaic acid, dinophysistoxins, Prorocentrum lima, Mytilus galloprovincialis, quantitative proteomics, molecular markers
Citation: Martins JC, Domínguez-Pérez D, Azevedo C, Braga AC, Costa PR, Osório H, Vasconcelos V and Campos A (2020) Molecular Responses of Mussel Mytilus galloprovincialis Associated to Accumulation and Depuration of Marine Biotoxins Okadaic Acid and Dinophysistoxin-1 Revealed by Shotgun Proteomics. Front. Mar. Sci. 7:589822. doi: 10.3389/fmars.2020.589822
Received: 06 August 2020; Accepted: 23 November 2020;
Published: 16 December 2020.
Edited by:
Valerio Matozzo, University of Padua, ItalyReviewed by:
Marco Gerdol, University of Trieste, ItalyCopyright © 2020 Martins, Domínguez-Pérez, Azevedo, Braga, Costa, Osório, Vasconcelos and Campos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alexandre Campos, YWNhbXBvc0BjaWltYXIudXAucHQ=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.