 Thomas A. Jefferson
Thomas A. Jefferson Jeffrey E. Moore
Jeffrey E. Moore- 1Clymene Enterprises LLC, Lakeside, CA, United States
- 2Southwest Fisheries Science Center, NOAA Fisheries, La Jolla, CA, United States
The Indo-Pacific finless porpoise (Neophocaena phocaenoides) occurs year-round in waters of southern and eastern Hong Kong. The population biology of the population was studied in the late 1990s and early 2000s, but has not been examined recently. There are several threats facing the animals, and there has been recent concern about this species’ status, due to an increase in the number of strandings in Hong Kong. Line-transect surveys using medium-size vessels have been conducted for small cetaceans in most of Hong Kong’s waters since 1996. We examined the density and abundance of finless porpoises in this area, using a Bayesian hierarchical framework, consisting of a Markov process model used to describe population dynamics and a distance-sampling observation model that links the line-transect data to the latent biological process. Time-series analyses for both dry- and wet-season data from 1996 to 2019 showed annual fluctuations in abundance indices, but no evidence of a significant long-term trend in finless porpoise abundance in Hong Kong. This indicates that the increase in strandings may be related to some factor other than changing population size of Hong Kong finless porpoises. In 2019, there were approximately 176 finless porpoises in Hong Kong’s waters in the dry season (winter/spring), the season of highest use. Highest densities (0.535 porpoises/km2) were recorded in South Lantau in the dry season. Based on the results of the current study, we recommend further survey work, improvements in data collection, possible tagging and molecular work to elucidate population structure, cumulative impact assessment, and the establishment of several marine parks for the conservation of finless porpoises in Hong Kong.
Introduction
The Indo-Pacific finless porpoise (Neophocaena phocaenoides) is widely distributed throughout coastal waters from the Taiwan Strait and Indonesia westwards to the Persian Gulf (Jefferson and Hung, 2004; Jefferson and Wang, 2011). These animals are highly vulnerable to human impacts, especially fisheries bycatch. The species as a whole is thought to be declining in numbers and is listed as Vulnerable on the IUCN Red List (Wang and Reeves, 2017), making it a high priority for conservation.
Hong Kong1 occurs at the eastern edge of the Pearl River Estuary (PRE), the second largest estuary in Chinese waters (Figure 1). Although 20 marine mammal species have been recorded from Hong Kong waters, there are only two resident species: the Indo-Pacific humpback dolphin (Sousa chinensis, also locally called the Chinese white dolphin) and the Indo-Pacific finless porpoise (Jefferson and Hung, 2007; W. C. Ng, Hong Kong Agriculture, Fisheries and Conservation Department [AFCD], unpublished data). Whereas a great deal has been learned about the Indo-Pacific humpback dolphin in Hong Kong in the last 25 years, the Indo-Pacific finless porpoise is much less understood. The latter is more difficult to detect and observe and thus studying its population biology is notoriously challenging.

Figure 1. Hong Kong, showing the survey strata, and the transect lines for those strata in which finless porpoises have traditionally been found. SW and SE Lantau strata are analyzed together in this study as South Lantau. Dolphin and porpoise use of the East Lantau area has dropped to near zero, and so this area is no longer surveyed. Areas to the west and north of Lantau Island are not used by finless porpoises, but are habitat for Indo-Pacific humpback dolphins. Mirs Bay was only surveyed by helicopter, due to its remote location.
The only previous analysis of abundance and population status of finless porpoises in Hong Kong is from dedicated work conducted in the late 1990s through 2000 (Jefferson and Braulik, 1999; Jefferson, 2000; Jefferson et al., 2002). This work showed that finless porpoises occur year-round in Hong Kong, with seasonal influxes in cooler dry-season months, bringing the local numbers to a maximum of about 152 porpoises in spring (Jefferson et al., 2002). Porpoises also occur in adjacent mainland Chinese waters to the south and west of Hong Kong, and taking some of these areas into account, the overall population may have numbered at least 217 porpoises in 2000/2001. There is some evidence suggesting that the PRE contains a distinct population of finless porpoises (Jia et al., 2014), though little is known about individual movements into and out of Hong Kong waters (Jefferson et al., 2002). However, it is clear that Hong Kong itself does not contain a separate population of finless porpoises.
A recent analysis of population dynamics of finless porpoises in the PRE region (which includes Hong Kong) suggested that the population declined from 1996 to 2005, and then slightly increased from 2006 to 2014 (Lin et al., 2019). It was suggested that these changes were due to fluctuations in fishing effort in the area. However, this analysis was based only on stranded carcasses, and not on survey data of live animals. Since there are a number of potential biases that could affect stranding access and recovery (see Lin et al., 2019), this study should be considered preliminary in nature, and findings need to be confirmed from more direct studies sampling the living population.
In recent years, the AFCD and their contractors have studied some aspects of finless porpoise biology in Hong Kong. Contract reports have included general information and summaries on finless porpoise distribution, encounter rates, group size, and habitat utilization, making use of both small-scale grid density analysis and habitat modeling (see Hung, 2019 and references therein). Many of these analyses are updated annually. However, there has been no detailed analysis of the line transect survey data to estimate large-scale density and abundance in Hong Kong, nor any evaluation of long-term trends.
Other than the study from carcasses discussed above, no evaluation of Hong Kong finless porpoise population status has been undertaken in the last 18 years (since 2002), although data relevant to this issue have been collected each year since 1996 under studies conducted by AFCD. A recent dramatic increase in stranding rates of finless porpoises in Hong Kong (see Figure 2) has prompted new concern about the status of the species and the health of the local Hong Kong population. The present study was initiated in response to this concern, and a major goal was to use the available data from past surveys (1996–2019) in order to estimate current density and abundance and in particular to examine long-term trends in abundance.
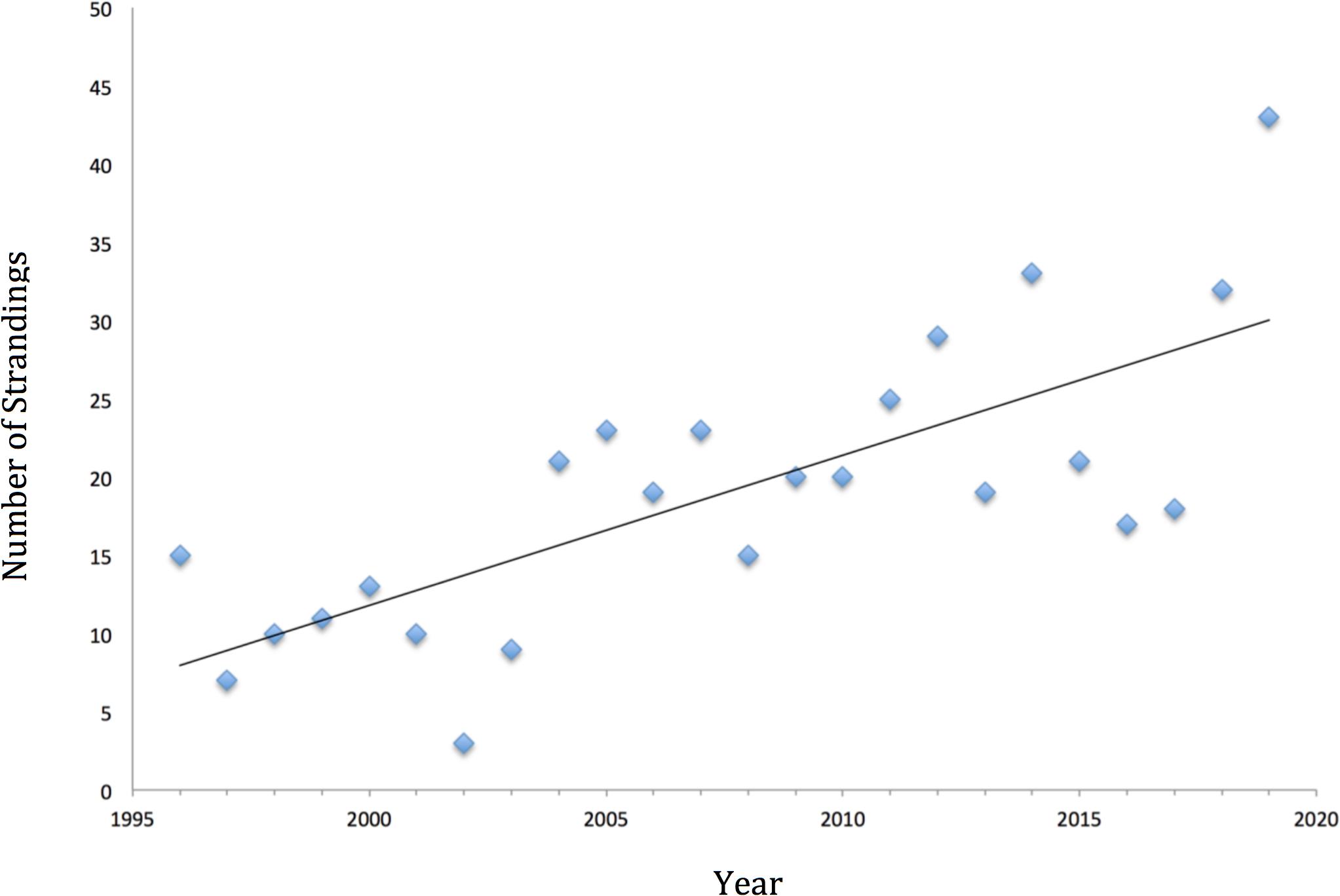
Figure 2. Strandings of finless porpoises in Hong Kong, by year of study (1996–2019). The line was fit by linear regression.
Materials and Methods
Study Area
The Hong Kong Special Administrative Region (HK SAR) is a semi-autonomous region in southern China (surrounded by the PRC’s Guangdong Province). Hong Kong waters comprise about 1800 km2 of inshore waters, most partially surrounded by land, but some of the southern waters face the open South China Sea. Vessel surveys have been conducted in all of Hong Kong’s marine waters, with the exception of Victoria Harbor and Tolo Harbor (which are both highly modified with heavy boat traffic, and are not considered suitable habitat for cetaceans) and Mirs Bay (which was too remote to survey by vessel and has only been surveyed using occasional helicopter surveys – see Jefferson et al., 2002).
Hong Kong’s waters are relatively shallow, all less than 40–45 m deep, and the western waters are estuarine, influenced by freshwater input from the Pearl River, one of China’s largest rivers. The southern and eastern waters are more marine and less turbid. Vessel traffic is heavy in the western area (North Lantau, West Lantau, and South Lantau), moderate in the central area (East Lantau and Lamma), and light in the eastern area (Po Toi, Ninepins, Sai Kung, and Mirs Bay).
Hong Kong was divided into nine main survey areas (some areas were later subdivided for finer-scale resolution of Indo-Pacific humpback dolphin habitat – Table 1 and Figure 1). Finless porpoises only occur in the southern, central, and eastern areas (Jefferson et al., 2002); the study area for this project was therefore defined to include the survey areas of South Lantau, Lamma, Po Toi, Ninepins, and Sai Kung. These are the areas that were identified as most consistently used by finless porpoises in the intensive study of Jefferson et al. (2002). Survey lines were devised to provide relatively even coverage of each survey area, with transect lines running north/south or east/west. Primary lines ran perpendicular to the coast, and were spaced 1–4 km apart, with the objective that each survey area could be effectively surveyed in a single day’s effort.
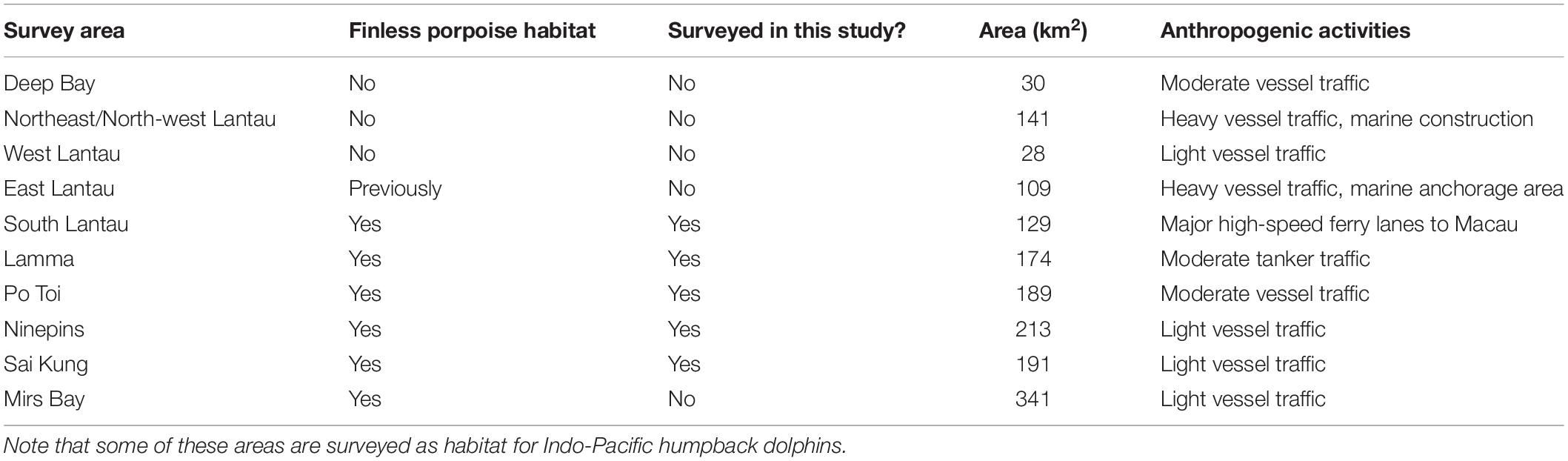
Table 1. Summary of all survey strata in Hong Kong and their characteristics.
Stranding Documentation
Marine mammal strandings in Hong Kong have been systematically documented since the early 1990s. A 24-hour stranding reporting hotline was established by the AFCD in the early 1990s. Most strandings are reported by the public, lifeguards, commercial vessel operators, or government workers (for example, marine police vessels) and each stranding report is investigated by a team from Ocean Park Conservation Foundation (an NGO associated with an oceanarium in Hong Kong). Basic data on the events surrounding each stranding are collected, and necropsies are conducted on those carcasses that are in accessible locations. Stranding data are considered to provide a reasonable indicator of trends in mortality (which could vary with changes in population size or with per capita mortality rate), as effort to document strandings has been generally consistent for the past several decades.
Vessel Surveys
Surveys for cetaceans were conducted from one of several inboard research vessels, all with similar structure and ranging in size from 12 to 15 m. Most of the vessels were junk-style boats with an elevated open upper deck area, and all vessels had a full 180° unobstructed view ahead of the vessel. Observers conducted searches from a sitting position on the front of the upper flying bridge area, which yielded an eye height of 4–5 m above the water’s surface.
The on-effort search team consisted of two observers. The primary observer searched for porpoises continuously through 7 × 35 Fujinon FMTRC (or similar) marine binoculars, and the data recorder searched with unaided eye (only using binoculars occasionally to check potential sighting cues) and recorded data. Survey data were recorded on paper sheets for later transcription to a computer database. Both observers searched the full 180° arc in front of the vessel, from 270° to 90° (with straight off the bow defined as 0°). There were usually 3–6 observers on the boat, and in order to reduce fatigue, observers rotated into and out of the on-effort team for 30 min at each position, allowing 30–120 min of rest for each hour of intensive searching. Most observers had previous experience in small cetacean line-transect surveys and all had taken at least one training course consisting of 2 days of classroom and 1 day of at-sea training.
Surveys were planned as much as possible to avoid poor weather conditions, and acceptable survey conditions were defined as winds less than or equal to Beaufort sea state 5, no heavy rain or fog, and visibility greater than or equal to 1200 m. On-effort searches were conducted as the vessel transited the transect lines at a constant speed of 13–15 km/h. Effort data consisted of description of overall sighting conditions, time and position for the start and end of each transect line, vessel speed, Beaufort sea state, visibility (in meters), and distance traveled in each search leg. Position, distance traveled, and vessel speed were determined from a handheld GPS unit (various models).
When porpoises were sighted, the search team was taken off effort (after the collection of data on initial sighting location and position) and the vessel was diverted to approach the group for detailed observations on behavior, better group size estimation, and the collection of photographs (when possible). Sighting data consisted of date and time of sighting, initial position, angle and distance to sighting, sea state, group size (best, high, low) and composition (e.g., presence of calves), response to survey vessel, and any vessel associations. When detailed observations were complete, the observers went back on effort and the vessel continued on-effort searching from a point just past where the sighting occurred (to avoid re-counting the same porpoise group again).
Sighting angles were determined with the aid of marine binoculars with built-in compass and reticles. Upon sighting porpoises, a compass reading was taken on the angle of the group from the bow of the boat, and then immediately after a second angle reading was taken along the bow of the vessel. Subtracting these two angle readings, provided a sighting angle to the nearest degree (this helped to reduce the tendency to round angles to the nearest 5° or 10°). Distances were estimated by eye by experienced observers, many of whom had undergone extensive training involving blind estimation of distances to objects that were then measured accurately using laser rangefinder binoculars (see Jefferson et al., 2002). This allowed for more accurate estimation of sighting distance (which was usually < 1 km), since the inability to see the horizon in the inshore survey areas prevented us from using reticles in the binoculars to determine distances.
Line Transect and Modeling Analyses
Estimates of abundance were calculated using Multiple Covariate Distance Sampling (MCDS), with Beaufort as a covariate for estimating the detection function. This was based on earlier work using Conventional Distance Sampling (CDS), which showed that Beaufort had a significant effect on detectability of finless porpoises (see Jefferson et al., 2002). Trend estimation generally followed the approach used by Moore and Barlow (2011, 2013, 2014) for using line-transect distance-sampling data to estimate population size and trends for cetaceans off the United States West Coast. The framework is hierarchical Bayesian, consisting of a Markov process model used to describe population dynamics and a distance-sampling observation model that links the line-transect data to the latent biological process. The population is spatially open (not all animals in the population occur simultaneously in the study area) and seasonally variable (the population is not migratory, but habitat varies seasonally so that different parts of the population occur in different areas in different seasons). Therefore, inter-annual dynamics were modeled separately for the dry season (winter and spring, December to May) and wet season (summer and fall, June to November), as the absolute numbers differ for these two seasonal categories, and trends could hypothetically differ as well (for example, if wintering habitats changed differently than summering habitats). Parameters were estimated using Bayesian MCMC methods with OpenBUGS software version 3.2.3 rev 1012. Truncation distance was set at 1000 m, which is beyond the maximum perpendicular sighting distance of 732 m, so effectively no data were truncated.
Process Model
Let Ns,a,y, be the population size for season s (1 = wet, 2 = dry), spatial stratum a, and year y (1996–2019). Change in seasonal abundance is modeled using a simple exponential stochastic growth model:
where rs is the annual rate of change parameter for season s, and εs,y is normally distributed (on the log scale) annual stochastic error within the seasonal model. This model assumes a fixed stratum effect within season (but not across seasons), i.e., each initial population value, Ns,a,1996, is estimated as a separate parameter. The data would not support estimating separate trends for each stratum within season; thus it is assumed that the relative proportions of animals using the different strata (within a given seasonal model) is constant across years.
The stratum densities (needed for estimating the model parameters; see below) are Ds,a,y = Ns,a,y/Aa, where Aa are the stratum area sizes. The total seasonal abundance across all K strata is . Population change for each year is λs,y = exp(rs + εs,y), or Ns,a,y + 1/Ns,a,y. The geometric mean of the yearly changes was taken as the mean rate of change for the population (by season); this was the primary metric of interest for the study.
Observation Model
The number of survey sightings was modeled as a Poisson random variable:
where b is the Beaufort sea state (b = 0, 1, 2, 3) (note we excluded sightings from higher Beaufort states from the analysis). The expected number of sightings is:
where hs,y is the mean group (herd) size of sightings (season and year-dependent, but not stratum-dependent); vb is the effective strip half-width (Buckland et al., 2001, 2004) when surveying in Beaufort state b; Ls,a,y,b is the amount of effort, i.e., length of transect conducted; and g(0) is the probability of detecting a group that is on the trackline.
Our estimate of trackline detection probability, g(0) = 0.607, is a new calculation from the same basic set of data used to produce our earlier estimate of 0.72 (Jefferson et al., 2002). However, while the earlier estimate only used POD (acoustic) data, the new estimate was computed using both POD data and surface and dive time (visual behavior observation) data. Thus our new estimate gives a more reliable estimate of g(0) that incorporates both perception bias and availability bias components, and also is relatively precise (with a CV of 0.161).
Estimating vb is the distance-sampling problem. Let g(x) describe the probability of detecting a group that is at perpendicular distance x from the trackline. We assume a half-normal model for g(x), which has a single parameter. This parameter is estimated as a function of Beaufort sea state, i.e., σb = exp(β0 + β1b). The detection function is integrated from x = 0 to a truncation distance of W = 1 km to find vb. The decision to model detection as a function of Beaufort sea state is supported by DIC for this model being 20 less than for the null model of constant detection.
Group size data were treated as Poisson random variables with expectations dependent on season (fixed effect) and year (random effect), so that:
where zi is the group size for sighting i and
where γs is the season-specific long-term mean and is inter-annual variance (assumed to be the same for both seasons). The decision to model season-specific group size was made a priori, given that population density is known to differ by season. Including random year effects was supported by DIC for this model being 90 less than a model of fixed seasonal group size.
Priors and Bayesian Model Fitting
Normal flat priors were specified for detection function parameters (β0,β1) and growth rate parameters (rs). Uniform priors were specified for initial population sizes (Ns,a,1996 ∼ Unif(0, 1000)), seasonal group size means (γs ∼ Unif(1, 10)) and the random effect parameters for stochastic variance in population change and group size (σs ∼ Unif(0, 2); σh ∼ Unif(0, 5)). An informative prior was used for trackline detection probability, i.e., g(0) ∼ Beta(14.55, 9.42).
Two MCMC chains were each run for a length of 2,000,000 samples, including a burn-in of 1,000,000. Samples were thinned by 1000, so that posterior distributions consisted of 20,000 samples (10,000 per chain). MCMC convergence of model parameters was assessed using the statistic, available as an output value of the function “bugs” within the R2OpenBugs package (these outputs are included in the Supplementary Material).
Results
Strandings
A total of 456 finless porpoises are known have stranded in Hong Kong from 1996 to 2019. During the 24-year period, there was an average of 19.0 ± SD 8.94 finless porpoise strandings per year (Figure 2). The median was 19, and the mode was 15. Despite annual fluctuations, finless porpoise strandings in Hong Kong have shown an overall increasing trend (Figure 2). However, the pattern may be more nuanced; it appears that following relatively stable or even declining strandings during the 1990s and early 2000s, there was a sudden jump in numbers in 2004, after which numbers were relatively stable but variable through 2017. In the last 2 years, the numbers spiked, with the most recent year of data (2019) also being the year with the highest number of strandings. These inflection points do not correspond with any known factors that might affect porpoise numbers.
Survey Data Summary
A total of 54,709 km of survey effort was conducted from 1996 to 2019, of which 49,126 km was conducted in Beaufort states 0–3 and thus considered useable for analysis. Beaufort did not vary significantly throughout the study. Mean Beaufort sea state for the data used in the analysis was consistently about 2.0 (1.8–2.2) during every year of the time series. Effort was relatively even throughout all months of the year (Figure 3), but highly variable across years and strata (Figure 4). Strata that were not well sampled, particularly over the most recent decade, were excluded from the analysis (i.e., Mirs Bay, Sai Kung). Thus, for the wet-season model, the analysis was based on data only from South Lantau, Po Toi, and Ninepins; these included 20,032 km of effort. For the dry-season model, the analysis was based on data from South Lantau and Lamma, which included 20,640 km of effort. Thus, analyses were based on a total of 40,672 km of effort, representing 83% of the total Beaufort 0–3 effort collected. Abundance estimates from the model thus represent abundance for the included strata only, and they are thus used as indices upon which to base inferences about abundance trends in the region.
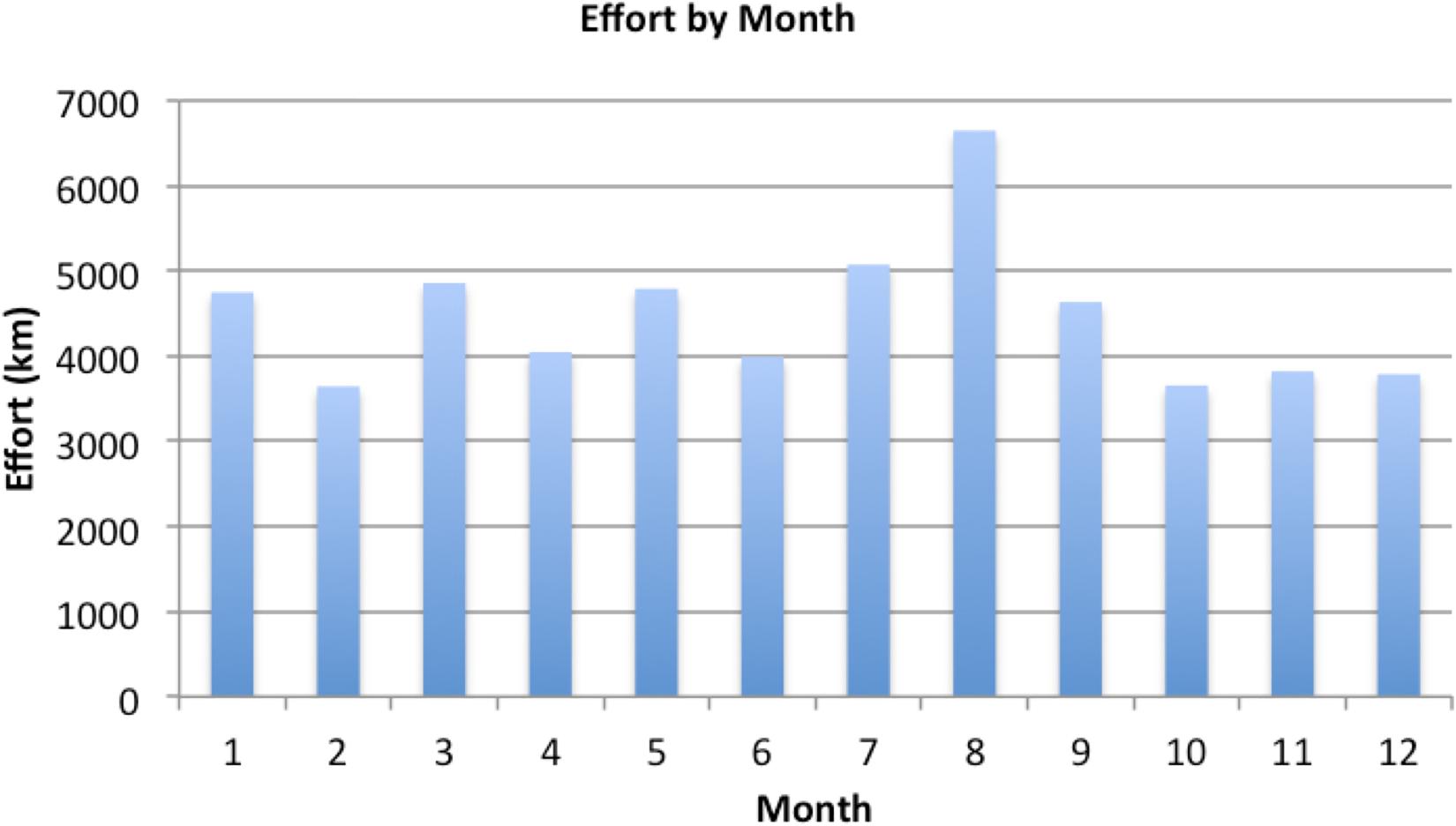
Figure 3. Survey effort by month, showing relatively consistent effort throughout the year. The dry season is winter to spring (December to May) and the wet season is summer to fall (June to November).
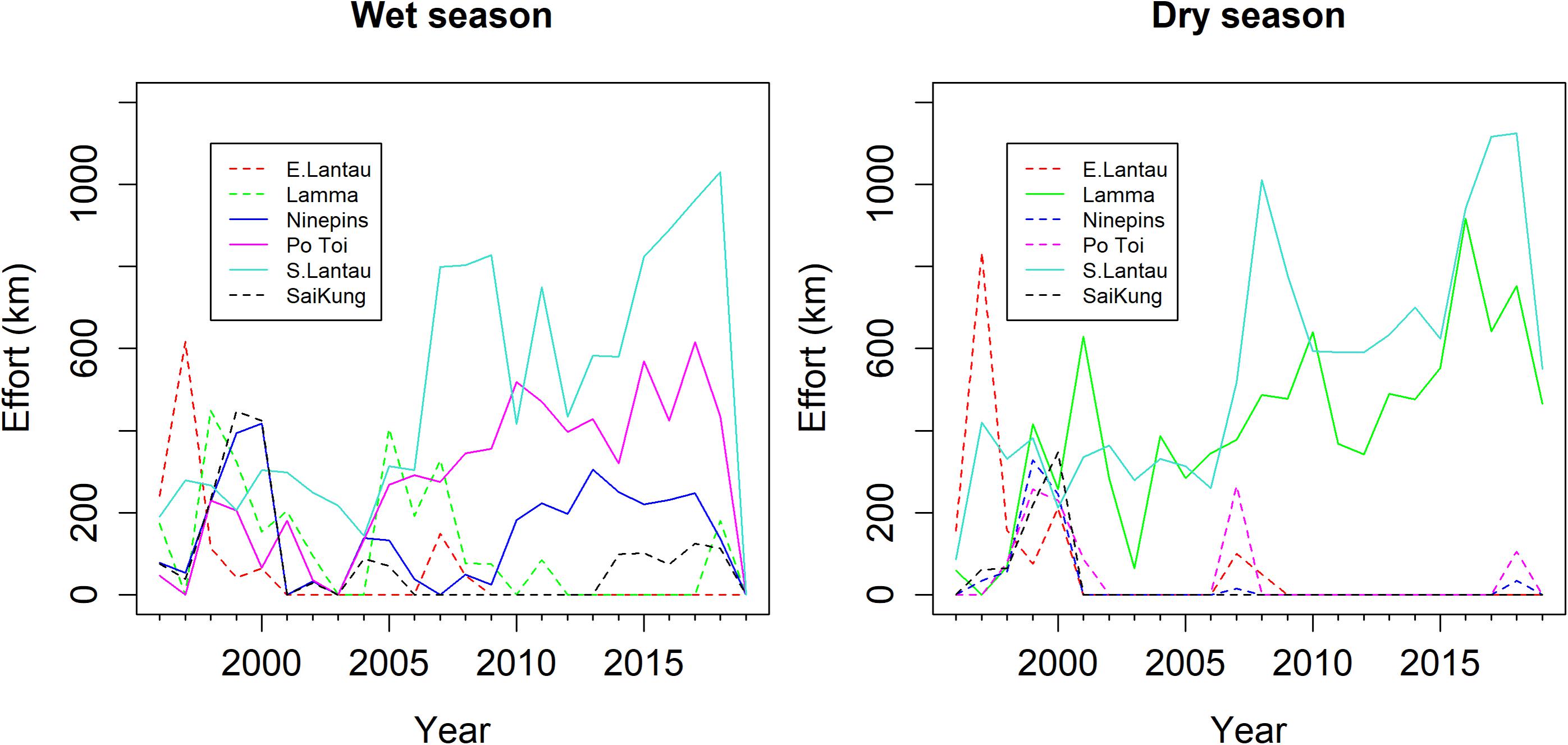
Figure 4. Effort conducted (Beaufort states 0–3) by year and strata. Solid and dotted lines show effort for sites included and excluded from the analysis, respectively.
There were a total of 1582 sightings of finless porpoise groups, of which 1277 were made while on effort across all strata and Beaufort states; the remainder were recorded while “off-effort.” Of these, 1231 (96%) occurred in Beaufort states 0–3. And of these, 1181 sightings (96%) were retained for conducting the seasonal analyses after omitting effort from some strata (see previous paragraph). Most (880, or 74.5%) of the 1181 sightings were recorded during the dry season, even though effort used in the analysis was approximately equal across wet and dry seasons.
Detection Function
Figure 5 shows the probability density functions for g(x) (i.e., f(x)), fit to the sighting-distance data from different Beaufort states. Detection probability decreases with increasing Beaufort state, as evidenced by the steepening decline in f(x) and higher proportion of sightings occurring closer to the vessel. The shape parameter estimates (posterior means and 95% CRIs) were β0 = -1.12 (-1.22 to -1.02) and β1 = -0.33 (-0.38 to -0.29). The latter (slope parameter) provides strong evidence of the effect of Beaufort on the shape parameter of the half-normal detection function. Corresponding estimates of effective strip half-width for each Beaufort state are 0.41, 0.29, 0.21, and 0.15 km for Beaufort states 0–3. Posterior distributions for detection function and other analysis parameters are in the Supplementary Material, along with MCMC convergence summaries. Detection function parameters converged well.
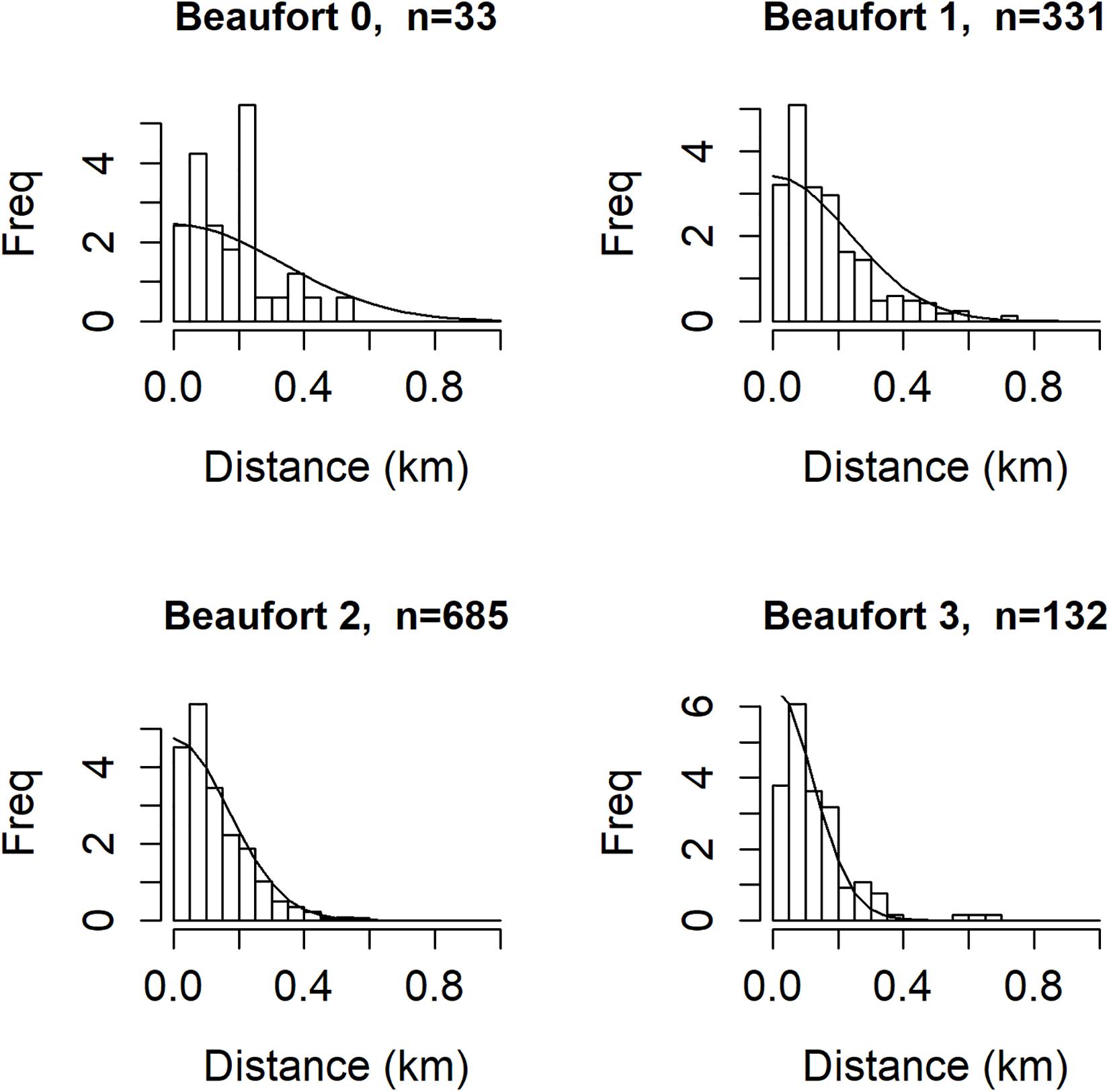
Figure 5. Histograms of sighting distances for n = 1181 sightings included in the analysis, by Beaufort state. Smooth lines are the probability density functions (half normal model) for g(x), i.e., f(x).
Group Size
Long-term mean group size was 2.37 (SE = 0.24; 95% CRI: 1.83–2.78) during the wet season and 2.84 (SE = 0.18; 95% CRI: 2.46–3.18) during the dry season. Annual estimates were variable (σh = 0.71), ranging from 1.5 to 3.3 across wet seasons and 2.1 to 4.6 across dry seasons.
Population Index and Trends
Based on the population indices, finless porpoise numbers appear to have been fairly stable, on average, from 1996 to 2019, although numbers within the study area vary considerably from year to year (Figure 6). During this time period, the average annual population change was λs = 1.022 (SE = 0.038; 95% CRI: 0.950–1.10) across wet seasons and 1.016 (SE = 0.022; 95% CRI: 0.975–1.06) across dry seasons. If anything, our trend estimates are overly precise, given Poisson assumptions in our model (encounter rate and group size) that may underestimate variance. Finless porpoises are more abundant during the dry season than wet season. This is evident from the raw sightings data (most sightings occurring during the dry season, even though effort was similar for the two season categories) and in the population estimates. Average (geometric mean) population abundance index for South Lantau, Po Toi, and Ninepins during the wet season was 67 animals with a SE of 14 and 95% CRI of 45–101. Inter-annual estimates ranged from 31 to 201. On average, the proportions of animals occurring in each of the three study areas was 0.15, 0.51, 0.34, respectively. Average abundance index for South Lantau and Lamma during the dry season was 117, with a SE of 22 and 95% CRI of 85–171. Inter-annual estimates ranged from 57 to 242. On average, the proportions of animals occurring in each of the two study areas was 0.58, 0.42, respectively.
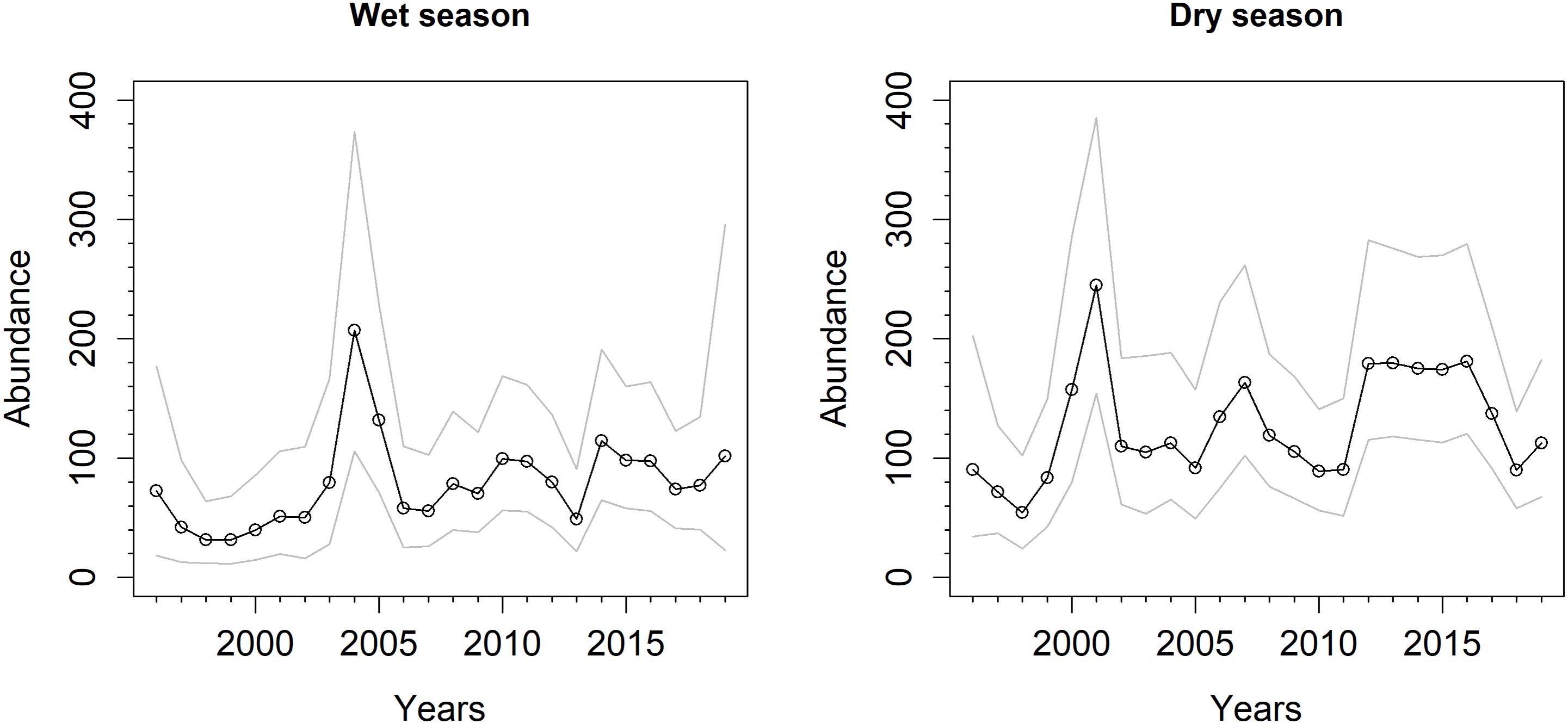
Figure 6. Time series of population index estimates from 1996 to 2019 during the wet and dry seasons. Values are Bayesian posterior means (points) and 95% credible intervals (gray lines). For the wet season, the index represents the abundance for South Lantau, Po Toi, and Ninepins strata. For the dry season, the index represents the abundance for South Lantau and Lamma strata.
While overall population trends have been relatively stable during the survey period, the last several years do coincide with both a spike in strandings (2018 and 2019) and a sharp estimated decrease in the dry-season abundance index (2016–2018).
We cannot directly estimate total abundance of finless porpoises in Hong Kong, due to survey effort being restricted to only certain survey areas in particular seasons. However, we can provide approximate abundance estimates if we assume that the proportions of porpoises in different areas is the same as it was in the early 2000s. This information comes from Jefferson et al. (2002), who conducted surveys in all survey areas inhabited by finless porpoises in 1998–2000. With this assumption, the abundance indices estimated here can be multiplied by a correction factor of 1.263 for the wet season, and 1.560 for the dry season, thereby providing approximate overall abundance for the 24 years of the study (Table 2).
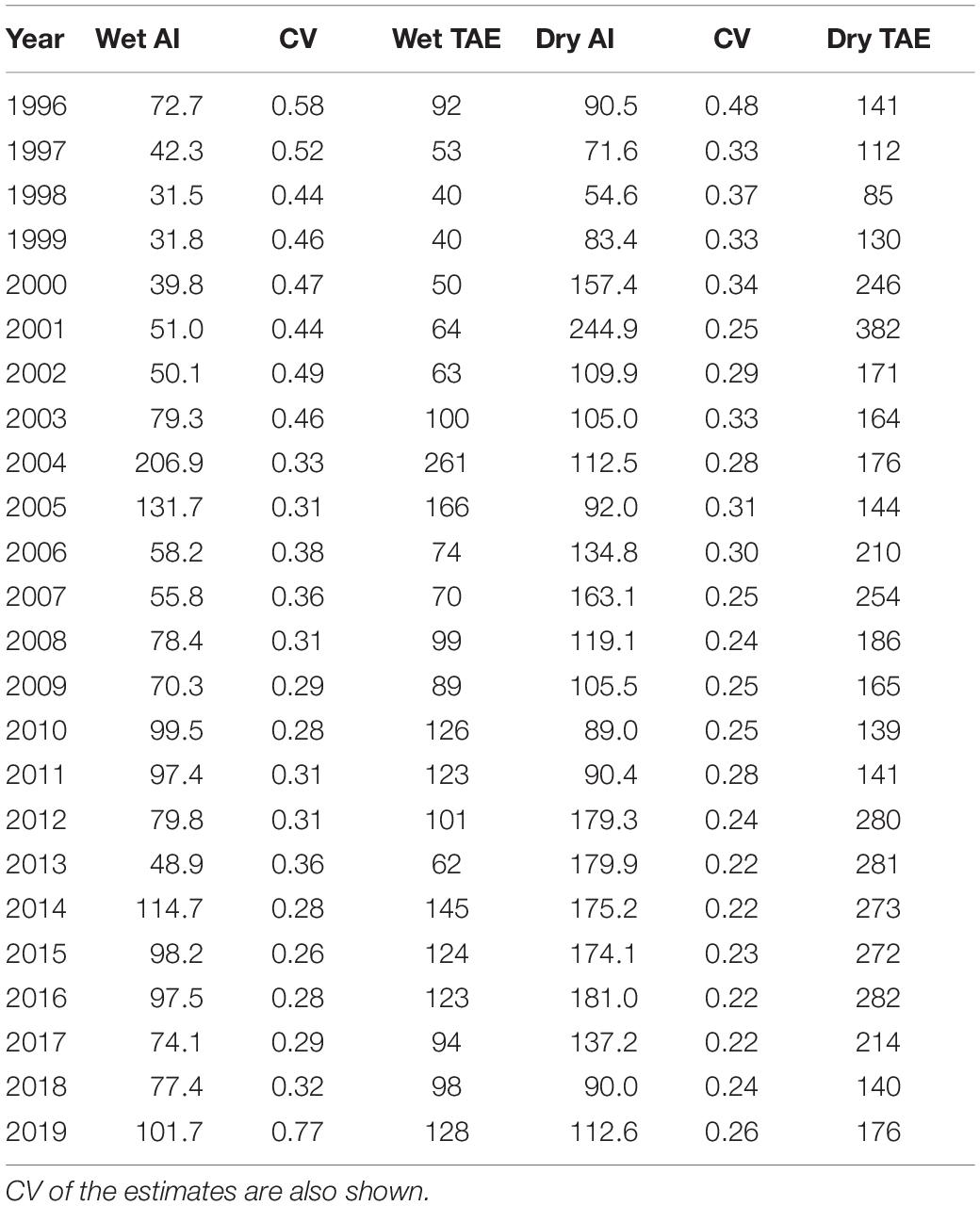
Table 2. Annual abundance indices (Al) by season, and their associated total abundance estimates (TAE).
Average (geometric mean) densities for the various strata ranged from a low of 0.084 porpoises/km2 for the wet season in South Lantau to a high of 0.535 porpoises/km2 for the dry season in South Lantau. The dry season South Lantau density estimate was more than twice that for Lamma (0.291 porpoises/km2), which had the highest densities recorded in all of Hong Kong for the years 1998–2000 (Jefferson et al., 2002). This suggests either that Lamma has become less important for finless porpoises in the last 20 years, that South Lantau has become more important, or both. Other strata (Ninepins and Po Toi) had intermediate densities in the current study.
Although there is no evidence of a major change in abundance over the course of the study, the results provide more evidence for an increase than for a decrease. For the parameter (geometric mean lambda from 1996 to 2019), the proportion of posterior MCMC samples with values > 1.0 gives the probability that the population index has increased over the duration of the study. For the wet season, this probability is 0.71, and for the dry season, this probability is 0.75. Taking 1.0 minus these values, the probability that the wet-season numbers have declined is 0.29, and the probability that the dry-season numbers have declined is 0.25.
Discussion
Comparisons to Previous Estimates
There are two species of finless porpoises, N. phocaenoides, the Indo-Pacific finless porpoise, and Neophocaena asiaeorientalis, the narrow-ridged finless porpoise (Jefferson and Wang, 2011). Only the former regularly occurs in Hong Kong, although there are a small number of extralimital records of narrow-ridged specimens from strandings in Hong Kong (Ocean Park Conservation Foundation, unpublished data). Both species can be a challenging to work with when conducting visual surveys at sea, due to their small size, dark color, and cryptic behavior of the animals, which make them difficult to detect, unless the sighting condition are very good. Due to these difficulties, some have doubted whether distance-sampling methods could be applied to these cryptic species. Nonetheless, good success has been achieved by careful planning of studies. Line transect methods have been used to reliably estimate density and abundance of finless porpoises in Japanese waters from vessel and aerial surveys for over 25 years (Shirakihara et al., 1994, 2007; Yoshida et al., 1997, 1998; Amano et al., 2003; Taguchi et al., 2007). The first use of this method for finless porpoises outside of Japan was the work by Jefferson and Braulik (1999) and Jefferson et al. (2002) in Hong Kong in the late 1990s and early 2000s. More recently, line transect methods have been used to assess finless porpoise abundance from vessel surveys in South Korea (Zhang et al., 2004; Park et al., 2007), Bangladesh (Smith et al., 2008), Malaysia (Minton et al., 2013), and the Yangtze River of China (Zhao et al., 2008).
Taken together, the above listed studies show that, despite the challenges, finless porpoises are suitable targets for line-transect surveys, and reliable estimates of density and abundance can be obtained. Most of the studies, however, have been one-off projects, which collected data only over a year or two, and thus have not been able to provide much information on long-term trends or population status. Our work presented here represents the longest-running study of consistent line-transect survey work for this species anywhere in the world. Our dataset, covering a period of 24 years, provides us with the ability to quantitatively determine trends in the number of porpoises using Hong Kong waters over this time period.
The uneven sampling throughout the whole study area is a shortcoming of this study, as it prevented us from making complete estimates of abundance for the whole of Hong Kong directly from empirical data, as was done in the Jefferson et al. (2002) study. It is hoped that in the future, all survey areas inhabited by the species will be surveyed in each of the wet and dry seasons, so that we can make complete estimates of abundance from empirical data.
Population Structure and Status of Hong Kong Finless Porpoises
There is still much unknown about the population status of the finless porpoises surrounding Hong Kong. There is little doubt that the range of the population extends well beyond Hong Kong’s waters, and that there are regular movements across the border between Hong Kong the surrounding waters of Guangdong Province. But, the true range of the population remains largely unknown. Recently, Jia et al. (2014) conducted a study of molecular variation in the South China Sea, which included samples from Hong Kong and surrounding waters, Taiwan, the Yangtze River, and the Yellow Sea. Their analyses showed that samples from the PRE (or Pearl River Delta, as they called it) were distinct from those in the other regions. They concluded from this that there is a Pearl River Delta population of finless porpoises, which is somewhat questionable. Furthermore, their study did little to clarify Hong Kong porpoises’ population boundaries, nor even to provide any real insight into population structure. Finless porpoises in the Yellow Sea, Yangtze River, and other areas north of the Taiwan Strait are already known to be from separate populations, as these animals are of a completely different species, the narrow-ridged finless porpoise (Jefferson and Wang, 2011).
Taking the results of Jia et al. (2014) as a starting point, Lin et al. (2019) conducted a demographic analysis of stranded carcasses of finless porpoises from the PRE during the period 2006–2014. They found evidence that the “population” was overall relatively stable, with a moderate decreasing trend from 1996 to 2005, and a slight increasing trend from 2006 to 2014. They credited this largely to the overall reduction in fishing effort in the region in the last two decades, suggesting that net entanglement and other fisheries-related issues may be the major human impacts that these animals face. If evaluated as a distinct population or stock for assessment (the IUCN Red List authority uses the term “subpopulation” to refer to such units of assessment), the PRE “subpopulation” would therefore qualify as Near Threatened (NT), using the IUCN Red List criteria (Lin et al., 2019). However, as we have argued, it is highly questionable whether there really is a PRE stock or population of finless porpoises. Finless porpoises apparently avoid the estuarine waters of the Pearl River (see Jefferson et al., 2002), and thus it seems very unlikely that the population would be centered around this geographic feature. It seems to us more likely that the range of the population that inhabits Hong Kong waters may extend to the north and offshore, areas quite distant from the freshwater influence of the Pearl River.
Geographic Variation in Presence and Density
Previous work specifically in Hong Kong (using 2014–2019 data from the same set of line-transect surveys that we have analyzed in this report) has found that in recent years the highest density areas used by Hong Kong finless porpoises in the dry season are between the Soko Islands and Shek Kwu Chau (in the South Lantau area), and in the wet season mainly around Po Toi Island (in the Po Toi area) and the far eastern waters of the Ninepins and Sai Kung survey areas (Hung, 2019). This is in contrast to earlier work, which showed the highest-density area for the animals to be a nearshore zone to the west of Lamma Island (Jefferson et al., 2002). While short-term (within a 3–4 years period) fluctuations in where porpoises aggregate from year to year were evident, examination of encounter rates and small-scale densities showed no strong evidence of any significant long-term trends in numbers, other than a potential increasing trend in the region of Southwest Lantau from 2004 to 2018 (Hung, 2019).
One interesting finding from the previous work was the possible acoustic evidence of finless porpoises in areas along West Lantau and Northwest Lantau in recent years, both areas in which they have never been detected in almost 25 years of intensive visual line-transect surveys (Hung, 2019). Whether this represents some kind of false positive (such as from humpback dolphin vocalizations, as Hung, 2019 suggests may be the case) or results from the actual expansion of finless porpoises into these waters traditionally used only by humpback dolphins remains unknown, but is worthy of further investigation.
Implications for Management
The current work, as well as the studies by Hung (2019) and Lin et al. (2019) seem to suggest that the population of finless porpoises that inhabits Hong Kong may not be facing the serious threats that have resulted in declines for the humpback dolphins that inhabit Hong Kong’s western waters (see Huang et al., 2012; Jefferson, 2018). This might seem surprising in light of the vulnerability of porpoises in general to human impacts and high human population density in the region, but when one looks at where the heaviest fishing and industrial activity in Hong Kong is located, it makes more sense. The western waters used by humpback dolphins are being intensively developed and support extensive fisheries (at least they did until recently) and very heavy vessel traffic of all sorts. By contrast, the southern and eastern waters of Hong Kong, where finless porpoise predominate, have much less human activity, although there are heavily used high-speed ferry lanes south of Lantau Island, and development there is on the increase. However, our conclusions must be viewed cautiously, given high uncertainty in the population monitoring time series. Spatial and annual variation in encounter rates was substantial (and likely higher than estimated), attributable to a combination of sampling variation, animal movement in and out of the study area, and true changes in population size. The relative importance of these different factors is unknown. For these reasons, short-term changes in estimates, such as the rapid decrease observed for the past couple years, are difficult to interpret and require additional monitoring and investigation to understand whether these are biologically important.
Expansions of coastal development west of Lamma (waste managementfacility) south of Lantau Island (waste management facility) and inHong Kong’s eastern waters (potential wind farm development) are being pursued, or at least considered, and such changes could potentially influence Hong Kong’s finless porpoise populations. We hope that a lesson can be learned from the humpback dolphin situation (which has been declining as its habitat in northern Lantau waters has degraded in recent years – see Jefferson, 2018), and that any development in finless porpoise habitat, if it is approved, should only proceed after careful environmental impact assessment has deemed such work compatible with a healthy finless porpoise population. Impacts should be assessed using the best available data, and uncertainties must be acknowledged. Above all, effective protection of finless porpoise prime habitat must be given, or the animals may suffer ill effects.
Recommendations
Based on insights from this study, we offer the following recommendations to AFCD related to future monitoring and management of finless porpoises in Hong Kong:
1.) Vessel survey work directed toward finless porpoises should continue, using the same methods described here, so that the database can be further extended into the future, facilitating future trends analyses,
2.) Survey effort should be allocated such that all important habitat areas for finless porpoises in Hong Kong are surveyed in every year (or at least every other year), i.e., survey effort in South Lantau, Lamma, Po Toi, Ninepins, and Sai Kung should be conducted in both wet and dry seasons,
3.) Better information on trackline availability should be collected – this could involve land-based or aerial surveys using UAVs (drones) to collect group dive and surface time data or tagging (TDR) studies,
4.) In order to evaluate the degree to which Hong Kong finless porpoises range into adjacent mainland Chinese waters, finless porpoise satellite tagging work should be seriously considered,
5.) The geographic range of the population that inhabits Hong Kong waters should also be investigated, through the use of molecular methods comparing Hong Kong and mainland specimens from both east and west of Hong Kong,
6.) Cumulative impacts of any future development in finless porpoise habitat should be adequately considered in EIA studies, and given serious consideration, and
7.) Management efforts should be directed toward developing marine parks in areas identified as critical habitat for finless porpoises in Hong Kong:
a. The region between the Soko Islands and Shek Kwu Chau (which is currently the highest-density area for the species in Hong Kong), and
b. The area to the south and west of Lamma Island, which previously had been identified as a high-density area and potential calving ground (see Jefferson et al., 2002).
Data Availability Statement
The data analyzed in this study are subject to the following licenses/restrictions: We do not own the data. Requests to access these datasets should be directed to ZGlja19rY19jaG9pQGFmY2QuZ292Lmhr.
Ethics Statement
The finless porpoise surveys were conducted with the permission and approval of the Hong Kong Agriculture, Fisheries, and Conservation Department.
Author Contributions
TJ conceived the study, carried out fieldwork, analyzed data, and wrote the majority of the manuscript. JM conducted Bayesian line-transect analyses, and assisted with writing of the manuscript. Both authors contributed to the article and approved the submitted version.
Funding
The Hong Kong Agriculture, Fisheries, and Conservation Department (AFCD) funded this study. JM is employed by NOAA Fisheries.
Conflict of Interest
TJ was employed by company Clymene Enterprises LLC.
The remaining author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank our AFCD colleagues Dick Choi and Wai Chuen Ng for their help with providing data and logistics. Much of the data were collected by survey teams from Ocean Park Conservation Foundation and the Hong Kong Cetacean Research Project, the latter led and overseen by Samuel Hung. The data we used in this report were collected by a large crew of field observers, too numerous to mention by name, but we appreciate their dedicated efforts, sometimes in challenging field conditions. To all these individuals and organizations, we express our heartfelt thanks.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2020.574381/full#supplementary-material
Footnotes
- ^ Hong Kong is a Special Administrative Region within China, giving it a high degree of autonomy and distinct international status. Thus, while Hong Kong is technically part of the China, it is treated as a distinct entity in this paper.
References
Amano, M., Nakahara, F., Hayano, A., and Shirakihara, K. (2003). Abundance estimate of finless porpoises off the Pacific coast of eastern Japan based on aerial surveys. Mammal Stud. 28, 103–110. doi: 10.3106/mammalstudy.28.103
Buckland, S. T., Anderson, D. R., Burnham, K. P., Laake, J. L., Borchers, D. L., and Thomas, L. (2001). Introduction to Distance Sampling: Estimating Abundance of Biological Populations. Oxford: Oxford University Press.
Buckland, S. T., Anderson, D. R., Burnham, K. P., Laake, J. L., Borchers, D. L., and Thomas, L. (2004). Advanced Distance Sampling. Oxford: Oxford University Press.
Huang, S. L., Karczmarski, L., Chen, J., Zhou, R., Zhang, H., Li, H. Y., et al. (2012). Demography and population trends of the largest population of Indo-Pacific humpback dolphins. Biol. Conserv. 147, 234–242. doi: 10.1016/j.biocon.2012.01.004
Hung, S. K. (2019). Monitoring of Marine Mammals in Hong Kong Waters (2018-19): Final Report. Hong Kong Cetacean Research Project Report to the Agriculture, Fisheries and Conservation Department. Hong Kong: Hong Kong Cetacean Research.
Jefferson, T. A. (2018). Hong Kong’s Indo-Pacific humpback dolphins (Sousa chinensis): assessing past and future anthropogenic impacts and working toward sustainability. Aquat. Mamm. 44, 711–728. doi: 10.1578/am.44.6.2018.711
Jefferson, T. A., and Braulik, G. T. (1999). Preliminary report on the ecology of the finless porpoise in Hong Kong waters. IBI Rep. 9, 41–54.
Jefferson, T. A. (ed.) (2000). Conservation Biology of the Finless Porpoise (Neophocaena phocaenoides) in Hong Kong Waters: Final Report. Kowloon: The Agriculture, Fisheries and Conservation Department, 285.
Jefferson, T. A., and Hung, S. K. (2004). Neophocaena phocaenoides. Mammal. Spec. 746:12. doi: 10.1644/746
Jefferson, T. A., and Hung, S. K. (2007). An updated, annotated checklist of the marine mammals of Hong Kong. Mammalia 71, 105–114.
Jefferson, T. A., Hung, S. K., Law, L., Torey, M., and Tregenza, N. (2002). Distribution and abundance of finless porpoises in Hong Kong and adjacent waters of China. Raffles Bull. Zool. Suppl. 10, 43–55.
Jefferson, T. A., and Wang, J. Y. (2011). Revision of the taxonomy of finless porpoises (genus Neophocaena): the existence of two species. J. Mar. Anim. Ecol. 4, 3–16.
Jia, K., Lin, W., Gui, D., Karczmarski, L., and Wu, Y. (2014). Molecular evidence reveals the distinctiveness of Indo-Pacific finless porpoises (Neophocaena phocaenoides) in the Pearl River Estuary and insights into genus Neophocaena’s origin. Mar. Biol. 161, 1919–1930. doi: 10.1007/s00227-014-2474-y
Lin, W., Karczmarski, L., Li, J., Chan, S. C. Y., Guo, L., and Wu, Y. (2019). Differential population dynamics of a coastal porpoise correspond to the fishing effort in a large estuarine system. Aquat. Conserv. Mar. Freshw. Ecosys. 2019, 1–12.
Minton, G., Peter, C., Zulkifli Poh, A. N., Ngeian, J., Braulik, G., Hammond, P. S., et al. (2013). Population estimates and distribution patterns of Irrawaddy dolphins (Orcaella brevirostris) and Indo-Pacific finless porpoises (Neophocaena phocaenoides) in the Kuching Bay, Sarawak. Raffles Bull. Zool. 61, 877–888.
Moore, J. E., and Barlow, J. (2011). Bayesian state-space model of fin whale abundance trends from a 1991-2008 time series of line-transect surveys in the California Current. J. Appl. Ecol. 48, 1195–1205. doi: 10.1111/j.1365-2664.2011.02018.x
Moore, J. E., and Barlow, J. (2013). Declining abundance of beaked whales (family Ziphiidae) in the California current large marine ecosystem. PLoS One 8:e52770. doi: 10.1371/journal.pone.0052770
Moore, J. E., and Barlow, J. (2014). Improved abundance and trend estimates for sperm whales in the eastern North Pacific from Bayesian hierarchical modeling. Endang. Spec. Res. 25, 141–150. doi: 10.3354/esr00633
Park, K. J., Kim, Z. G., and Zhang, C. I. (2007). Abundance estimation of the finless porpoise, Neophocaena phocaenoides, using models of the detection function in a line transect. J. Korean Fish. Soc. 40, 201–209. doi: 10.5657/kfas.2007.40.4.201
Shirakihara, K., Shirakihara, M., and Yamamoto, Y. (2007). Distribution and abundance of finless porpoise in the Inland Sea of Japan. Mar. Biol. 150, 1025–1032. doi: 10.1007/s00227-006-0363-8
Shirakihara, M., Shirakihara, K., and Takemura, A. (1994). Distribution and seasonal density of the finless porpoise Neophocaena phocaenoides in the coastal waters of western Kyushu, Japan. Fish. Sci. 60, 41–46. doi: 10.2331/fishsci.60.41
Smith, B. D., Ahmed, B., Mansur Mowgli, R., and Strindberg, S. (2008). Species occurrence and distributional ecology of nearshore cetaceans in the Bay of Bengal, Bangladesh, with abundance estimates for Irrawaddy dolphins Orcaella brevirostris and finless porpoises Neophocaena phocaenoides. J. Cet. Res. Manage. 10, 45–58.
Taguchi, M., Yoshioka, M., and Kashiwagi, M. (2007). Seasonal changes in density of the finless porpoise (Neophocaena phocaenoides) around the mouth of Mikawa Bay, Japan. Mammal. Sci. 47, 11–17.
Wang, J. Y., and Reeves, R. R. (2017). Neophocaena phocaenoides. IUCN Red List Threat. Spec. 2017:e.T198920A50386795.
Yoshida, H., Shirakihara, K., Kishino, H., and Shirakihara, M. (1997). A population size estimate of the finless porpoise, Neophocaena phocaenoides, from aerial sighting surveys in Ariake Sound and Tachibana Bay, Japan. Res. Pop. Ecol. 39, 239–247. doi: 10.1007/bf02765270
Yoshida, H., Shirakihara, K., Kishino, H., Shirakihara, M., and Takemura, A. (1998). Finless porpoise abundance in Omura Bay, Japan: estimation from aerial sighting surveys. J. Wildl. Manage. 62, 286–291. doi: 10.2307/3802290
Zhang, C. I., Park, K. J., Kim, Z. G., and Sohn, H. (2004). Distribution and abundance of finless porpoise (Neophocaena phocaenoides) in the west coast of Korea. J. Korean Fish. Soc. 37, 129–136. doi: 10.5657/kfas.2004.37.2.129
Keywords: Bayesian statistics, cetaceans, conservation, line-transect analysis, management, marine mammals, population dynamics
Citation: Jefferson TA and Moore JE (2020) Abundance and Trends of Indo-Pacific Finless Porpoises (Neophocaena phocaenoides) in Hong Kong Waters, 1996–2019. Front. Mar. Sci. 7:574381. doi: 10.3389/fmars.2020.574381
Received: 19 June 2020; Accepted: 01 December 2020;
Published: 22 December 2020.
Edited by:
Jeremy Kiszka, Florida International University, United StatesReviewed by:
Len Thomas, University of St Andrews, United KingdomBernd Wuersig, Texas A&M University-Commerce, United States
Copyright © 2020 Jefferson and Moore. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Thomas A. Jefferson, c2NseW1lbmVAYW9sLmNvbQ==