 David J. López-Cortés1†
David J. López-Cortés1† Erick J. Núñez Vázquez1,2*
Erick J. Núñez Vázquez1,2* Juan J. Dorantes-Aranda3
Juan J. Dorantes-Aranda3 Christine J. Band-Schmidt4
Christine J. Band-Schmidt4 Francisco E. Hernández-Sandoval1
Francisco E. Hernández-Sandoval1 José J. Bustillos-Guzmán1
José J. Bustillos-Guzmán1 Ignacio Leyva-Valencia5
Ignacio Leyva-Valencia5 Leyberth J. Fernández-Herrera4
Leyberth J. Fernández-Herrera4- 1Centro de Investigaciones Biológicas del Noroeste, La Paz, Mexico
- 2Investigación para la Conservación y el Desarrollo, La Paz, Mexico
- 3Institute for Marine and Antarctic Studies, University of Tasmania, Hobart, TAS, Australia
- 4Instituto Politécnico Nacional-Centro Interdisciplinario de Ciencias Marinas, La Paz, Mexico
- 5CONACyT, Instituto Politécnico Nacional-Centro Interdisciplinario de Ciencias Marinas, La Paz, Mexico
The marine dinoflagellate Margalefidinium polykrikoides is a harmful species that has affected aquaculture, fisheries and tourism activities. It produces reactive oxygen species (ROS) as well as hemolytic and neurotoxic-like substances that have been associated with mass mortalities of marine organisms. It has a tropical and subtropical distribution that has mainly affected Asia and North America. The economic impacts for aquaculture industries have been estimated to be up to US$140M. In Latin America, no economic estimates have been performed. Harmful algal blooms by M. polykrikoides are more frequent in Mexico and Central America. Proliferations of this dinoflagellate are associated with winds, upwelling, convergence areas, local convection of seawater, and eutrophication of coastal areas by nitrogen and phosphorus compounds from rainwater runoff, as well as agricultural and aquaculture activities, into coastal waters. Eco-physiological and toxicological studies have provided detailed descriptions of the growth of algal strains from these regions and the harmful effects on fish and shrimp, as well as the role the production of ROS and polyunsaturated fatty acids in their toxicity. It is also possible that M. polykrikoides has an ecological role in the regulation of blooms of other harmful algae. In this contribution, we review the records of harmful algal blooms of M. polykrikoides in Latin America and the research that has been conducted with this species.
Introduction
Global Harmful Algal Blooms of M. polykrikoides
Margalefidinium polykrikoides (=Cochlodinium polykrikoides) is a photosynthetic and mixotrophic marine dinoflagellate that forms cysts. It forms harmful algal blooms (HABs), causing economic losses in fish farming areas (US$140M in world wide; Dorantes-Aranda, 2012) mainly in Asia (Kim, 1998; Kim et al., 1999; Zhong and Gobler, 2009). M. polykrikoides was first identified in Phosphorescent Bay, Puerto Rico by Margalef (1961), and since then Margalefidinium blooms have been reported across Asia, Europe and North America. In the last three decades, Margalefidinium HABs have impacted coastal regions of Australia, Canada, China, Croatia, India, Iran, Italy, Japan, Korea, Malaysia, Oman, Philippines, Russia, Saudi Arabia, United Arab Emirates, and the United States (Kudela and Gobler, 2012). The largest bloom recorded occurred in the Arabian Gulf from August to October 2008 affecting 1,200 km of coastline, killing wild, and farmed fish as well as damaging coral reefs (Richlen et al., 2010). A wide variety of harmful substances have been reported for this species (Figure 1), including reactive oxygen species (ROS) (superoxide anion, hydrogen peroxide, hydroxyl radical), hemolytic and neurotoxic-like substances, hemagglutinins, and free polyunsaturated fatty acids (Onoue and Nozawa, 1989; Hallegraeff, 1992; Lee, 1996; Landsberg, 2002; Jeong et al., 2004; Dorantes-Aranda et al., 2009, 2010). The genus Cochlodinium was established in the late nineteenth century with the identification of C. strangulatum (Schütt, 1895), and the first record of this genus in the Gulf of California occurred in 1942 (Osorio-Tafall, 1943). Recently, C. polykrikoides was re-assigned to the new genus Margalefidinium and renamed as M. polykrikoides (Gómez et al., 2017). In North America, HABs of M. polykrikoides have been observed in Canada and the US with impacts on fish and shellfish industries (Whyte et al., 2001; Gobler et al., 2008, 2012; Tomas and Smayda, 2008; Mulholland et al., 2009; Zhong and Gobler, 2009; Griffith et al., 2019). The damage by a M. polykrikoides bloom in Canada was estimated in a loss of US$2M in 1999 (Whyte et al., 2001). In Latin America (LA) there are no estimates of the economic losses caused by this dinoflagellate. The purpose of this work is to review the HABs of M. polykrikoides and the impacts they have caused in LA by country.
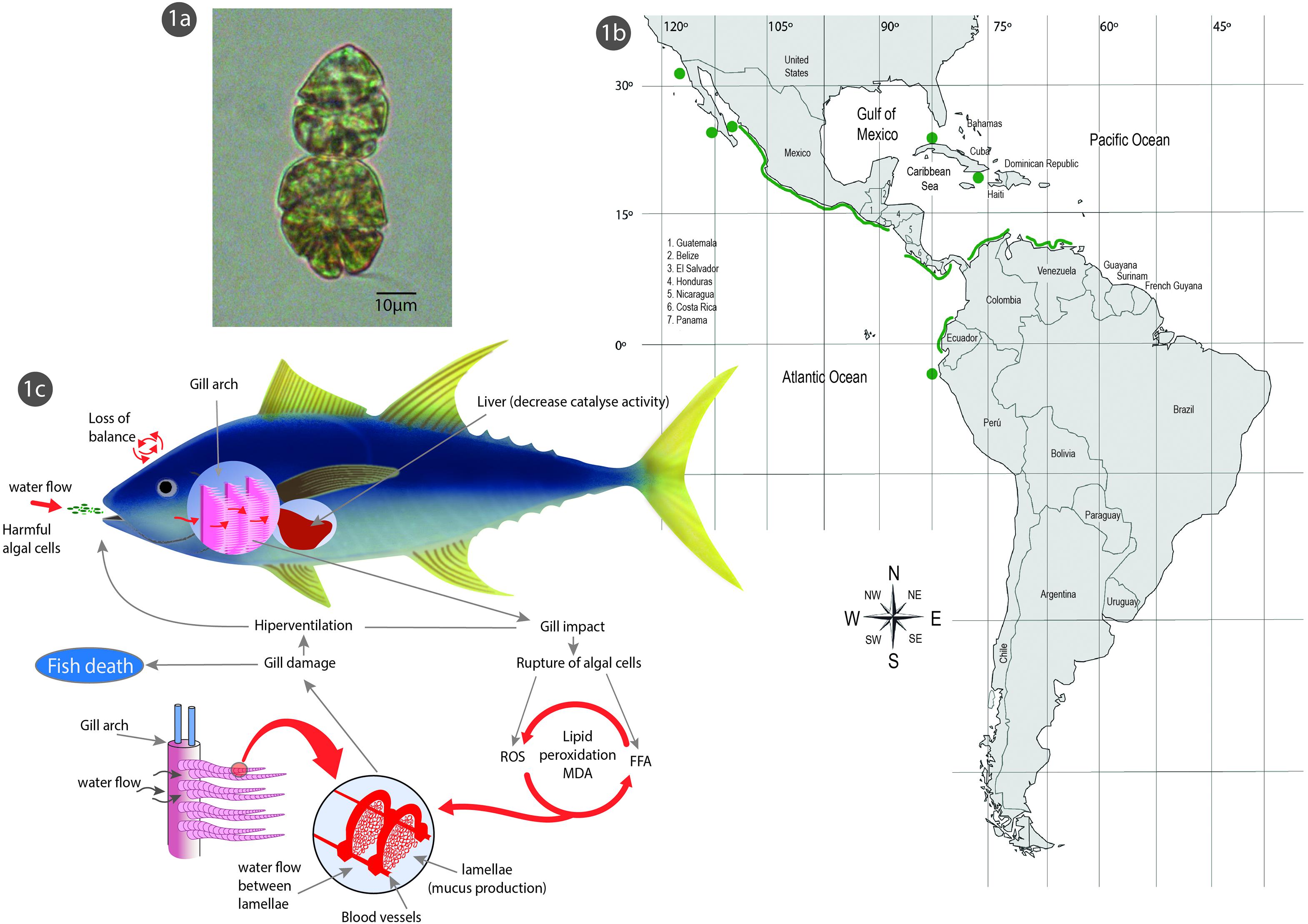
Figure 1. (a) Microscopic image (63×) of Margalefidinium polykrikoides in culture (chain of two cells), isolated from Bahía de La Paz, Gulf of California, Mexico. Scale bar = 10 μm. (b) HABs records of M. polykrikoides in coastal waters of Latin America. (c) Suggested route of ichthyotoxicity caused by M. polykrikoides: fish gills are impacted when algal cells in the water lyse, releasing reactive oxygen species (ROS) and free polyunsaturated fatty acids that cause lipid peroxidation and physiological disturbances leading to fish mortality (modified after Hallegraeff et al., 2017).
Mexico
Gulf of California
M. polykrikoides was first observed in Bahía de Mazatlán, Sinaloa, in spring of 1979 proliferating together with Gymnodinium catenatum. It was suggested that M. polykrikoides HABs are commonly formed in Bahía de Mazatlán during winter and early spring coinciding with the upwelling of cold waters in the region (Morey-Gaines, 1982; Cortés-Altamirano, 1987). From September to October 2000, M. polykrikoides was present during 22 days causing a notable odor and fish mortalities (Cortés-Altamirano and Gómez-Aguirre, 2001; Cortés-Altamirano et al., 2002; Table 1). Fish kills were also reported in 2003 and 2012 (Cortés-Altamirano et al., 2019). A record of HABs in Bahía de Mazatlán between 1979 and 2014 by Cortés-Altamirano et al. (2019) revealed that this species forms blooms during summer and fall when El Niño exerts its inhibitory effect. This dinoflagellate has also been reported forming blooms in shrimp farming ponds and coastal lagoons of Sinaloa with adverse effects such has fish, oysters and octopus mortality, as well as skin hyperpigmentation in swimmers (Alonso-Rodríguez et al., 2004, 2008). Despite the occurrence of mortalities, the economic impacts of these blooms have not been determined.
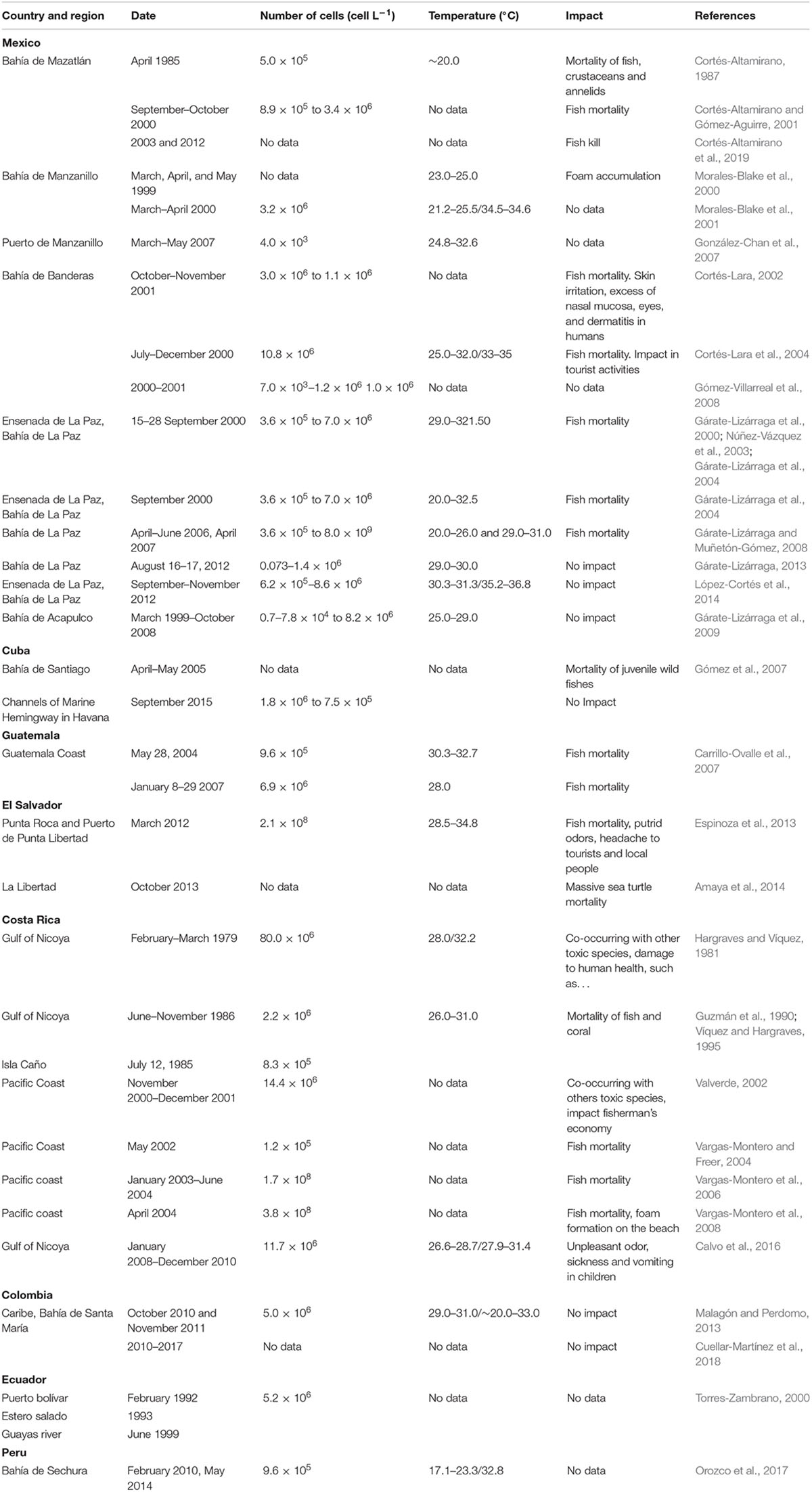
Table 1. Events of harmful algal blooms by the ichthyotoxic dinoflagellate Margalefidinium polykrikoides in Latin America.
An outbreak of this dinoflagellate was reported for the first time in Bahía de La Paz, in the southern part of the Gulf of California, in September 2000 (Gárate-Lizárraga et al., 2000). Chlorophyll a concentrations ranged from 2.7 to 56.8 mg L−1. Gárate-Lizárraga et al. (2004) described another bloom in this bay in November 2000. The bloom emerged after 2 days of heavy rain and wind coinciding with an increase in nutrient concentrations (Table 1). Another bloom occurred in September-November 2001, which extended outside the bay probably a result of Hurricane Juliette; mortalities of 166 fish (76 adults and 90 sub-adults fish) were observed in ponds, including the species Lutjanus peru (n = 102), Pomadasys macracanthus (n = 60), and L. argentiventris (n = 4), with abundant cells of M. polykrikoides observed in the gills (Núñez-Vázquez et al., 2003; Gárate-Lizárraga et al., 2004; Figure 2). Muciño-Márquez (2010) also reported M. polykrikoides near tuna pens in Bahía de la Paz in September 2006, however, no adverse effects were reported.
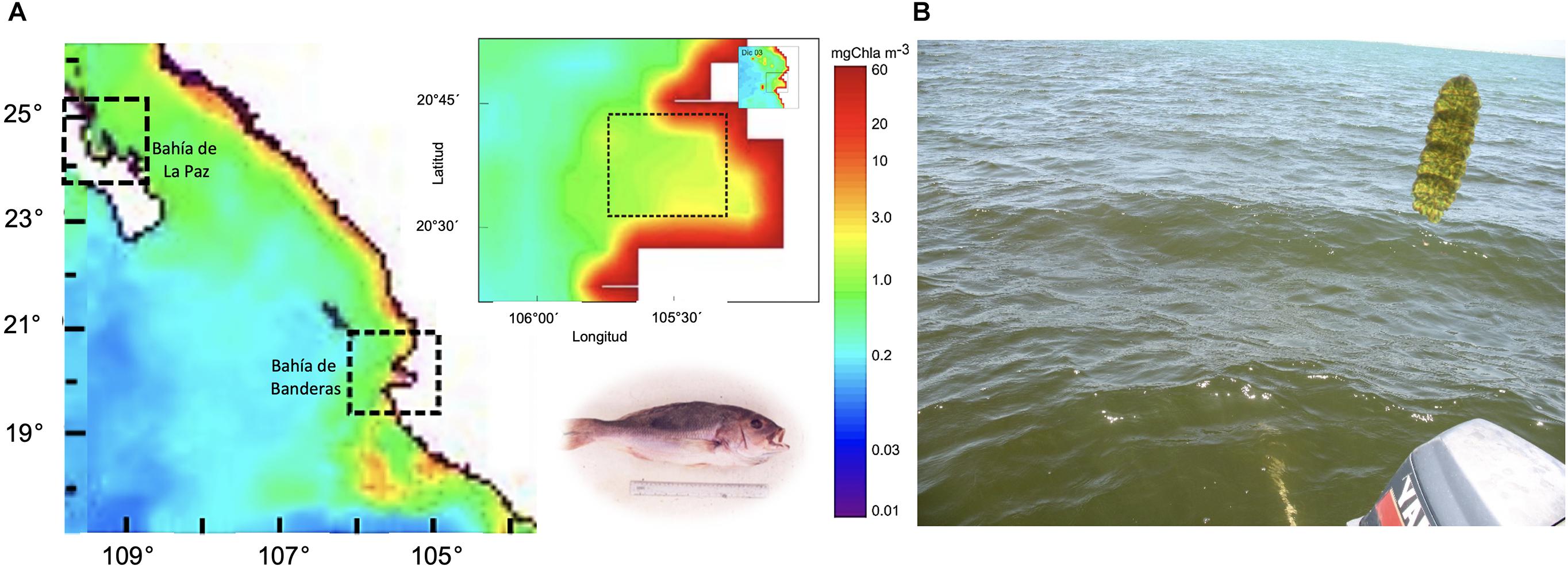
Figure 2. (A) Satellite images showing chlorophyll a concentration in mgChla m–3 in December 2001. Dotted box corresponds to Bahía de La Paz and Bahía de Banderas, some of these blooms could have been formed by M. polykrikoides. Maps from SeaWiFS level 3 images. The bottom left image shows fish kills of Pacific red snappers (Lutjanus peru) in culture ponds caused by M. polykrikoides in Bahía de La Paz in 2001–2002. (B) Brown coloration of the sea surface created by M. polykrikoides in Bahía de la Paz (2016).
López-Cortés et al. (2014) followed the development of a bloom of M. polykrikoides in Bahía de La Paz from the beginning (September) to its decay (October) in 2012. This event reached a maximum cell density of 8.6 × 106 cell L–1, with a chlorophyll a content of 121.2 mg m–3 and peridinin of 40.2 mg m–3, coinciding with a N:P (nitrogen:phosphorus) ratio of 2:3. This event was associated with NNE winds and rain, which may have contributed to the enrichment of nutrients in the water column; the direction of the wind changed to the SE, and its intensity decreased to <1.3 m s–1, which was when the bloom appeared. No mortality of marine organisms were observed during this period. According to Gárate-Lizárraga (2013) and López-Cortés et al. (2014), recurrent blooms of M. polykrikoides in this bay are associated with the mixing of the water column and upwelling of deep waters.
Mexican Pacific
The first record of M. polykrikoides was recorded in Bahía de Manzanillo, Colima in 2000 (López, 2000; Morales-Blake et al., 2001). Another extensive bloom by Margalefidinium sp. was reported during late winter and early spring of 1999, where the paralytic toxin producing dinoflagellate G. catenatum was also found. However, no impacts in wildlife were reported (Morales-Blake et al., 2000). Another event was recorded in this bay between March and May in 2007, involving again several HAB species such as Akashiwo sanguinea, Karenia mikimotoi, G. catenatum as well as M. polykrikoides. This event was associated with upwelling of deep waters (González-Chan et al., 2007).
The most extensive bloom by M. polykrikoides (1.1 × 106–3.0 × 106 cell L–1) reported in Mexico lasted 12 weeks (September to November 2000), affecting 63 km of coastline in Bahía de Banderas. This event impacted tourism activities in Puerto Vallarta and Nuevo Vallarta in the states of Jalisco and Nayarit, respectively (Cortés-Lara et al., 2004). Another M. polykrikoides bloom was reported in the area the following year (Cortés-Lara, 2002). Cortés-Lara (2002) mentioned that the phenomenon remained for 5 months in Puerto Vallarta causing mortality of eels, octopuses, and 13 species of fish, including flounders, sardines, snappers and puffers. Gómez-Villarreal et al. (2008) monitored HABs in Bahía Banderas using satellite images during 2000 and 2001 (Figure 2). High chlorophyll a levels were associated with blooms of M. polykrikoides during the summer-fall season, which were more intense in 2000 than in 2001, with a range in cell abundances from 7 × 103 to 1.2 × 106 cell L–1 in 2000, and an average cell abundance of 1 × 106 cell L–1 in 2001.
Gárate-Lizárraga and Muñetón-Gómez (2008) reported mortalities of farmed tuna in Bahía Magdalena, B.C.S associated with M. polykrikoides and reported that this dinoflagellate is distributed from Ensenada, B.C. to the coasts of Oaxaca. Gárate-Lizárraga et al. (2009) also reported HAB events of M. polykrikoides and G. catenatum in Bahía de Acapulco. Both species were recorded from December 2005 to December 2007. The abundance of M. polykrikoides ranged from 39 × 103 cell L–1 in January 2008 to 8,228 × 103 cell L–1 in December 2005, within a thermal range of 25–29°C; however, no mortalities of marine organisms were observed. Maciel-Baltazar and Hernández-Becerril (2013) reported this species in 2009 for the first time in the Gulf of Tehuantepec.
Recently, Fimbres-Martínez et al. (2018), Fimbres-Martínez (2019) described the presence of Margalefidinium sp. and raphidophytes of the genera Chattonella, Heterosigma and Fibrocapsa in Bahía de Todos Santos, in the northern Pacific of Mexico, which were the suspected cause of mortalities of farmed tuna (Thunnus thynnus) in this region (García-Mendoza et al., 2018).
Experimental Studies
During the last decade, efforts have been directed toward the isolation of strains from Bahía de La Paz to study their ecology and toxicology. Growth rates calculated for these strains varied between 0.11 and 0.39 div day–1 with maximum biomasses of 7.1 ± 0.5 and 9.4–11.0 × 106 cell L–1 using modified GSe and GSe media (Dorantes-Aranda et al., 2009, 2010; Zumaya-Higuera, 2017). Some authors have suggested that M. polykrikoides is an invasive species that has been transported by seawater currents and ship ballast water (Sierra-Beltrán et al., 2001; Cortés-Altamirano et al., 2006; Meave del Castillo, 2014; Páez-Osuna et al., 2017). However, the sequences of strains isolated from Bahía de La Paz were identical to that of American and Malaysian strains (Zumaya-Higuera, 2017).
Núñez-Vázquez et al. (2003) observed that fish juveniles of Mugil sp. died when exposed to M. polykrikoides at a cell abundance of 4.1 × 106 cell L–1. However, cell extracts showed no toxicity by mouse bioassay and did not have adverse effects on juvenile shrimp Litopenaeus vannamei. However, Pérez-Morales et al. (2017) reported 100% mortality of L. vannamei zoea larvae after exposing them for 120 h to cells of M. polykrikoides (3.0 × 106 cell L–1). Dorantes-Aranda et al. (2010), also observed 100% mortality of the spotted rose snapper Lutjanus guttatus after exposure to ≥3.0 × 106 cell L–1 of M. polykrikoides. Fish showed loss of balance, breathing difficulty, oxidative stress in gill lamellae and liver, abnormal production of mucus and asphyxiation, suggesting that the production of ROS by the dinoflagellate caused oxidative damage that lead to their death. In a complimentary study, the same authors observed that extracts from 5.2 × 106 M. polykrikoides cell L–1 and 27.0 × 106 cell L–1 caused 50% hemolysis in L. guttatus and human erythrocytes, respectively. The authors also reported hexadecaenoic (16:0), docosahexaenoic (22:6n3), and octadecapentanoic (18:5n3) as the most abundant fatty acids (Dorantes-Aranda et al., 2009). The latter fatty acid has also been found in other nocive microalgae showing fish cell toxicity, suggesting that it plays a key role in the ichthyotoxicity caused by microalgae (Dorantes-Aranda et al., 2009; Mooney et al., 2011). The possible ichthyotoxic pathway of M. polykrikoides is shown in Figure 1c, as suggested by Hallegraeff et al. (2017).
Allelopathy of M. polykrikoides has been demonstrated by exposing cells and filtrates of M. polykrikoides to live cells of G. catenatum. M. polykrikoides caused cell damage to G. catenatum, such as detachment of the membrane, deformation, prominent nuclei, loss of flagella, and lysis, suggesting that M. polykrikoides could inhibit or regulate the growth of G. catenatum in the natural environment (Zumaya-Higuera, 2017).
Caribbean and Central America
Cuba
M. polykrikoides proliferated in Bahía de Santiago in April and May 2005 covering an area of 0.8 km2. The HAB was short (4 days), however, it caused mortality of wild juvenile fish (Mujil curema, Opisthonema oglinum, Acanthurus chirurgus, Haemulon spp.) and crabs (Callinectes sapidus). During the bloom, cysts and high cell concentrations were recorded (Table 1; Gómez et al., 2007). Another bloom occurred in the channels of the Marina Hemingway in Havana in September 2015. A cell concentration of 1.8 × 106 cell L–1 was reported, and the bloom declined after 2 days of torrential rain, decreasing to 7.5 × 105 cell L–1. No impact on wildlife was reported (Delgado et al., 2016).
Guatemala to Nicaragua
A HAB of M. catenatum was recorded for the first time in Guatemala in 2004, which lasted 32 days and occurred from May to June. Toward the end of the bloom, on the 23rd of June, samples for species identification and cell density were obtained that contained 9.6 × 105 cell L–1 which explained the abnormal values of chlorophyll a (>30 mg m–3) observed in satellite images (Carrillo-Ovalle et al., 2007). High chlorophyll a levels (10–39 mg m–3) were also found off the coast of Honduras, El Salvador and the Gulf of Fonseca (gulf shared between El Salvador, Honduras and Nicaragua). Rain contributed to high nutrient concentrations (Table 1), and the water temperature was 30.3°C. A second event was recorded in January 2007 with a higher cell concentration (6.9 × 106 cell L–1). This bloom occurred at a lower temperature (28°C) during the influence of coastal water upwelling. Wild fish mortalities were observed during both events (Carrillo-Ovalle et al., 2007).
El Salvador
Espinoza et al. (2013), documented a bloom of M. polykrikoides off the coast of El Salvador in March 2012, with a maximum cell density of 2.1 × 108 cell L–1. The bloom was 3 km wide and 13 km long, and mortality of benthic fish, bad odor and severe headaches in tourists and local residents were reported. The major tourist season in El Salvador is from March to April, however, no economic losses were estimated for this event. Mass sea turtle mortalities occurred in La Libertad in October 2013 (Amaya et al., 2014). PSP toxins were detected in turtle tissues as well as in oysters. Although the most abundant species during this event was G. catenatum, other species such as P. bahamense, A. monilatum, and M. polykrikoides were also reported. In July 2017 a mixed bloom caused by M. polykrikoides and Scripsiella trochoidea affected several coastal areas of El Salvador (Ochoa-Arguello, 2017). During this event, a sanitary closure was applied.
Costa Rica and Panama
A bloom of M. polykrikoides reported as M. catenatum occurred in the Gulf of Nicoya, Costa Rica in February-March 1979 with a width of 200 m and length of 2–3 km. A strain was isolated from this event and growth rate of 0.3 div day–1 was estimated. The maximum abundance of cells during the event was 80.0 × 106 cell L–1 (Hargraves and Víquez, 1981; Table 1).
An annual monitoring program of harmful dinoflagellates was conducted in the middle and upper Gulf of Nicoya between January 1985 and March of 1986. HABs of M. polykrikoides were commonly observed in the lower gulf between June and October, mainly during the rainy season. The highest abundance of M. polykrikoides was 5.0 × 106 cell L–1 during this period (Víquez and Hargraves, 1995). Ramírez et al. (1989) suggested that the decline of anchovy eggs in the estuary of Punta Morales recorded was caused by a red tide near the estuary in August 1985 (Table 1).
In the second half of 1985, in Panama (Caño Island, Costa Rica, and Uva Island), mass mortalities of reef fish, invertebrates and corals were associated with a bloom of M. catenatum and Gonyaulax monilata. In Caño Island, a coral mortality up to 100% was observed between the surface and 3 m of depth. The species most affected were Pocillopora elegans and the zooxantella Tubastrea coccinea. Additionally, hundreds of fish of the family Scarideae, Balistidae, Acanthuridae, Pomancentridae, and Tetraodontidae; hermit crabs, brachyuran crabs and gastropods. In Uva Island, 13% mortality of Pocillopora spp. occurred. Mucus adhesion to polyps and interference with the expansion of polyps appeared to be the cause of coral mortality (Guzmán et al., 1990). Valverde (2002) reported a toxic bloom of Pyrodinium bahamense var. compressum, G. catenatum, and M. polykrikoides, on the Pacific Coast of Costa Rica from late November 2000 to December 2001 (Table 1). No impact was observed when the abundance of cells of M. polykrikoides reached 14.4 × 106 cell L–1. Vargas-Montero and Freer (2004) reported a bloom in Puntarenas and Caldera beach in the Gulf of Nicoya in May 2002 (1.2 × 105 cell L–1). This event was dominated by M. polykrikoides and the cyanobacterium Trichodesmium erythraeum. Large fish died, and deformity of fish larvae was also observed.
M. polykrikoides bloomed again on the Pacific region of Costa Rica between January 2003 and June 2004. Cell abundance was 1.7 × 108 cell L–1 (Table 1), and cysts were also observed in October 2003 (Vargas-Montero et al., 2006). The following bloom, in January 2004, covered an area of 50 km2 and in April of the same year a high number of cells were also found (3.8 × 108 cell L–1). During these events, corals died and fish mortalities of the families Carangidae, Lutjanidae, Muraenidae, Engraulidae, and the Solidae occurred (Vargas-Montero et al., 2006). M. polykrikoides commonly bloomed in this region with low abundance of other dinoflagellate species such as Ceratium dens, Gonyaulax spinifera, Heterocapsa sp., Mesodinium rubrum, P. bahamense var. compressum and the cyanobacterium T. erytraeum. Vargas-Montero et al. (2006, 2008) concluded that northeaster winds may be the most influential factor in the formation of HABs of M. polykrikoides in Costa Rica.
A total of 11 HABs of different species of dinoflagellates were reported in the Gulf of Nicoya from 2008 to 2010. Of these, eight were observed during the rainy season (May to November) and three during the dry season (December to April) (Calvo et al., 2016). Authors emphasize that the most impacting HAB occurred in September 2010, when 16 phytoplankton species were observed, with M. polykrikoides as the dominant species, occurring during a La Niña event. Heavy rain contributed to the enrichment of nutrients in the Gulf of Nicoya, as a result of runoff discharges of the rivers Tempisque and Grande de Tárcoles.
South America
Venezuela
Only one report exists on a bloom of Margalefidinium sp. on the coast of Sucre, Venezuela, which occurred in August 1977. Cells of Margalefidinium were found in the digestive tract of the mussel Perna perna. During this bloom, Gonyaulax tamarensis var. excavate (=Alexandrium catenella) and Noctiluca miliaris were also observed, and the presence of paralytic shellfish toxins were detected in mussels, which were associated with human intoxications (Reyes-Vásquez et al., 1979).
Colombia
A bloom of M. polykrikoides was recorded for the first time in Bahía de Santa María, in the Colombian Caribbean in October and November 2010. The maximum density was 5.0 × 106 cell L–1, and the event covered an area of 6 km2, however, no impact on fauna was observed (Malagón and Perdomo, 2013). The key factor for the proliferation was the contribution of terrestrial nutrients during the rainy season. This bloom coincided with SSE and SE winds during the day, and NNW and NW during the night. Cuellar-Martínez et al. (2018) reported two more blooms in the same bay between 2010 and 2017 with no harmful effects.
Ecuador
HABs are frequent in Ecuador, more commonly in the Gulf of Guayaquil. There have also been reports of blooms in the southern part of the Santa Elena peninsula, Manglar alto, Manta, Cojimies and in the Galápagos Islands. One of the most frequent and abundant species is Margalefidinium sp., which bloomed in Puerto Bolívar (March 1992), Estero Salado (March 1993), and Guayas river in June 1999, reaching a maximum cell abundance in Guayas River of 5.2 × 106 cell L–1 (Torres-Zambrano, 2000). The high frequency of HABs in the Gulf of Guayaquil is associated with poor water quality due to mangrove removal, ship traffic, shrimp farming, population increase, combined with El Niño events.
Peru
Individual blooms of A. sanguinea, M. rubrum, Protoceratium minimum, and M. polykrikoides occurred in Bahía de Pisco in February 2010 and May 2014. These events were associated with upwelling of deep waters, with a temperature range from 17.1 to 23.3°C. The bacteria Vibrio alginolyticus, V. metschmicovich, V. vulfenicus, and V. parahaemolyticus were isolated during the bloom. A strain of V. parahaemolyticus was virulent and a public health event with diarrhetic symptoms was associated with the bacteria (Orozco et al., 2017).
Conclusion
The toxic dinoflagellate M. polykrikoides is wide spread in Latin American coastal waters. It has formed recurrent blooms that have been documented mainly in Mexico and Central American countries. M. polykrikoides has been able to proliferate in a wide temperature range (17–32°C), and has affected several marine organisms, including fish, crabs, corals, shrimp, eels, octopuses, and gastropods. Blooms of M. polykrikoides in Latin America have been associated with periods of heavy rain that cause an increase in nutrients in coastal waters due to runoff from rivers. Moreover, coastal upwellings associated with wind patterns, mixing of the water column and rain, all of which can create nutrient enrichment of surface waters, seem to favor proliferation and recurrence of this dinoflagellate. Although significant economic losses have been reported in other countries due to the negative impacts of M. polykrikoides, this information is still lacking in Latin America, possibly due to scarce socioeconomic studies. A continuous monitoring program is required to obtain information, during and after bloom occurrence. Also, data of cyst beds should be included to generate ecological information that can provide the opportunity to evaluate the potential impacts on fisheries and aquaculture. Especially to address the economic and social repercussions that HABs may have, given the recent experience of two continuous blooms created by Pseudochattonella verruculosa and Alexandrium catenella in Chile in 2016 that caused economic losses of US$800M, creating unemployment and social riots triggered by protests (Mascareño et al., 2018). Mitigation approaches for HABs of this dinoflagellate are also a subject not yet investigated in Latin America. The higher amount of reports from Mexico might be because this country possesses a longer coastal area, as well as having a high number of experts on harmful algae, or simply because M. polykrikoides has formed more blooms in the Mexican coastline than in other Latin America countries. The presence of Margalefidinium as well as other species of marine fish-killing microalgae in Latin America will require constant monitoring mainly in fish-growing areas to avoid severe economic impacts, such as the recent cases in Chile and Mexico (Clement et al., 2016; García-Mendoza et al., 2018; León-Muñoz et al., 2018; Mardones et al., 2019).
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the institutional projects PAC (Planeación Ambiental y Conservación) of the CIBNOR, to G. Hernández-García end edition of figures (CIBNOR), Dr. J. L. Peña-Manjarrez (CETMAR-DGECyTM, SEP, Ensenada) for the satellite image HAB of the M. polykrikoides, Instituto Politécnico Nacional (IPN Grant SIP 2019–5649), and Don Johnson, Ph.D. for improving the language part of the manuscript. We are grateful to the Consejo Nacional de Ciencia y Tecnología (FORDECyT Grant 260040), and RedFAN-CONACyT. CB-S is a COFFA-IPN and EDI fellow. We also thank the reviewers for the suggestions provided during editing of this manuscript.
References
Alonso-Rodríguez, R., Hernández-Becerril, D. U., and Gárate-Lizárraga, I. (2008). “Catálogo de microalgas de las lagunas costeras de Sinaloa,” in Serie Lagunas Costeras de Sinaloa 4, ed. F. P. áez-Osuna (Sinaloa, ME: Universidad Nacional Autónoma de México).
Alonso-Rodríguez, R., Páez-Osuna, F., and Gárate-Lizárraga, I. (2004). El fitoplancton en la camaronicultura y larvicultura: Importancia de un buen manejo. México, ME: Comité Estatal de Sanidad Acuícola de Sinaloa.
Amaya, O., Ruiz, G., Espinoza, J., and Rivera, W. (2014). Saxitoxin analyses with a receptor binding assay (RBA) suggest PSP intoxication of sea turtles in El Salvador. Harmful Algae News 48, 6–7. doi: 10.3389/fmars.2018.00409
Calvo, V. E., Berrocal, A. K., and Boza, A. J. (2016). Floraciones algales nocivas durante el periodo 2008-2010 en el Golfo de Nicoya. Costa Rica. Rev. Mar. Coast. 8, 129–149. doi: 10.15359/revmar.8-1.9
Carrillo-Ovalle, L., Cortés-Altamirano, R., Leiva-Cerezo, V., and Ramírez, V. (2007). Cochlodinium catenatum on Guatemala coast (2004 & 2007). Harmful Algae News 33, 15–17.
Clement, A., Lincoqueo, L., Saldivia, M., Brito, C. G., Muñoz, F., Fernández, C., et al. (2016). Exceptional summer conditions and HABs Pseudochattonella in southern Chile create record impacts on salmon farms. Harmful Algae News 53, 1–3.
Cortés-Altamirano, R. (1987). Observaciones de mareas rojas en la Bahía de Mazatlán. Sinaloa, México. Cienc. Mar.. 13, 1–19. doi: 10.7773/cm.v13i4.557
Cortés-Altamirano, R., Alonso-Rodríguez, A., and Salas-de León, D. A. (2019). Historical observations of algal blooms in Mazatlán bay, Sinaloa, Mexico (1979-2014). PLoS One 14:e0210631. doi: 10.1371/journal.pone.0210631
Cortés-Altamirano, R., and Gómez-Aguirre, S. (2001). “Florecimientos algales nocivos en la Bahía de Mazatlán, México (2000),” in Proceedings of the Memoria in Extenso del Congreso Internacional COLACMAR 16-20 de septiembre, San Andrés.
Cortés-Altamirano, R., Cifuentes, J. L., and Gaxiola López, J. (2002). “Mareas rojas: biodiversidad de microbios que pintan el mar,” in Atlas de Biodiversidad de Sinaloa, (México, ME: Colegio de Sinaloa), 29–41.
Cortés-Altamirano, R., Lavín, M. F., Sierra-Beltrán, A., and Cortés-Lara, M. C. (2006). Una hipótesis sobre el transporte de microalgas invasoras desde el Pacífico oeste tropical hasta el Golfo de California por las corrientes marinas. Cienc. Del. Mar. U. A. S. 18, 19–26. doi: 10.1371/journal.pone.0210631
Cortés-Lara, M. C. (2002). Informe del fenómeno de marea roja en Bahía de Banderas Jalisco-Nayarit, octubre-noviembre de 2001. Rev. Biomed. 13, 73–75.
Cortés-Lara, M. C., Cortés-Altamirano, R., and Sierra-Beltrán, A. P. (2004). Presencia de Cochlodinium catenatum (Gymnodiniales: Gymnodiniaceae) en mareas rojas de Bahía de Banderas, Pacífico Mexicano. Rev. Biol. Trop. 52(Suppl.1), 35–49.
Cuellar-Martínez, T., Ruiz-Fernández, A. C., Alonso-Hernández, C., Amaya-Monterrosa, O., Quintanilla, R., Carrillo-Ovalle, H. L., et al. (2018). Addressing the problem of harmful algal blooms in Latin America and the Caribbean- a regional network for early warning and response. Front. Mar. Sci. 5:409. doi: 10.3389/fmars.2018.00409
Delgado, G., Arencibia, G., Lucas, F., and Rosquete, C. M. A. (2016). Cochlodinium polykrikoides Margalef bloom in the channels of marine Hemingway, La Habana, Cuba. Harmful Algae News 53:8.
Dorantes-Aranda, J. J. (2012). Development and novel application of an in vitro fish gill cell assay to elucidate the ichthyotoxic mechanism of the microalga Chattonella marina (Raphidophyceae). Ph.D. Thesis, University of Tasmania: Burnie, TAS
Dorantes-Aranda, J. J., García-de la Parra, L. M., Alonso-Rodríguez, R., and Morquecho, L. (2009). Hemolytic activity and fatty acids composition in the ichthyotoxic dinoflagellate Cochlodinium polykrikoides isolated from Bahía de la Paz, Gulf of California. Mar. Poll. Bull. 58, 1401–1405. doi: 10.1016/j.marpolbul.2009.06.007
Dorantes-Aranda, J. J., García-de la Parra, L. M., Alonso-Rodríguez, R., Morquecho, L., and Voltolina, D. (2010). Toxic effect of the harmful dinoflagellate Cochlodinium polykrikoides on the spotted rose snapper Lutjanus guttatus. Environm. Toxicol. 25, 319–326. doi: 10.1002/tox.20507
Espinoza, J., Amaya, O., Reguera, B., Alvarado, Y., Ruiz, G., and Escobar, D. (2013). A Cochlodinium polykrikoides bloom in El Salvador. Harmful Algae News 47:13.
Fimbres-Martínez, M. (2019). Distribución y abundancia de microalgas ictiotóxicas en la costa Noroeste de Baja California, México. MSc. Thesis, Centro de Investigación Científica y de Educación Superior de Ensenada: Mexico, ME.
Fimbres-Martínez, M., García-Mendoza, E., Olivos-Ortiz, A., Rivas-Camargo, D., and Helenes-Escamilla, J. (2018). “Spatial and temporal abundance of ichthyotoxic microalgae in Todos Santos bay, Baja California, Mexico. abstract book ICHA 2018,” in Proceedings of the 18th International Conference on Harmful Algae, (Nantes).
Gárate-Lizárraga, I. (2013). Bloom of Cochlodinium polykrikoides (dinophyceae): gymnodiniales) in Bahía de La Paz. Gulf of California. Mar. Poll. Bull. 67, 217–222.
Gárate-Lizárraga, I., and Muñetón-Gómez, M. C. (2008). Los riesgos de las mareas rojas en el Pacífico Mexicano. Conversus 3, 20–23.
Gárate-Lizárraga, I., Bustillos-Guzmán, J. J., Morquecho, L. M., and Lechuga-Devéze, C. H. (2000). First outbreak of Cochlodinium polykrikoides in the Gulf of California. Harmful Algae News 21:7.
Gárate-Lizárraga, I., Díaz-Ortiz, J., Pérez-Cruz, B., Alarcón-Tacuba, M., Torres-Jaramillo, A., Alarcón-Romero, M. A., et al. (2009). Cochlodinium polykrikoides and Gymnodinium catenatum in Bahía de Acapulco, Mexico (2005-2008). Harmful Algae News 40, 8–9.
Gárate-Lizárraga, I., López-Cortés, D. J., Bustillos-Guzmán, J. J., and Hernández-Sandoval, F. E. (2004). Blooms of Cochlodinium polykrikoides (Gymnodiniaceae) in the Gulf of California, Mexico. Rev. Biol. Trop. 52(Suppl.1), 51–58.
García-Mendoza, E., Cáceres-Martínez, J., Rivas, D., Fimbres-Martínez, M., Sánchez-Bravo, Y., Vásquez-Yeomans, R., et al. (2018). Mass mortality of cultivated northern bluefin tuna Thunnus thynnus orientalis associated with Chattonella species in Baja California, Mexico. Front. Mar. Sci. 5:454. doi: 10.3389/fmars.2018.00454
Gobler, C., Berry, J., Anderson, D., Burson, O. R., Koch, A., Rodgers, F., et al. (2008). Characterization, dynamics, and ecological impacts of harmful Cochlodinium polykrikoides blooms on eastern long island NY, USA. Harmful Algae 17, 293–307. doi: 10.1016/j.hal.2007.12.006
Gobler, C., Burson, J., Koch, A., Tang, F., and Mulholland, M. R. (2012). The role of nitrogenous nutrients in the occurrence of harmful algal blooms caused by Cochlodinium polykrikoides in New York estuaries (USA). Harmful Algae 17, 64–74. doi: 10.1016/j.hal.2012.03.001
Gómez, F., Richlen, M. L., and Anderson, S. M. (2017). Molecular characterization and morphology of Cochlodinium strangulatum, the type species of Cochlodinium, and Margalefidinium gen. nov. for C. polykrikoides and allied species (Gymnodiniales, Dinophyceae). Harmful Algae 63, 32–44. doi: 10.1016/j.hal.2017.01.008
Gómez, L., Licea, Y., Díaz, U., and Álvarez, I. (2007). Primer reporte de Cochlodinium polykrikoides (Margalef, 1961) en aguas Cubanas. Rev. Invest. Mar. 28, 87–89.
Gómez-Villarreal, M. C., Martínez-Gaxiola, M. D., and Peña-Manjarrez, J. L. (2008). Proliferaciones algales 2000-2001 en Bahía de Banderas, México según el sensor SeaWiFS. Rev. Biol. Trop. 56, 1653–1664.
González-Chan, R. B., Hernández-Silva, L., Navarro-Ornelas, J. G., and Blanco-Padilla, M. A. (2007). Proliferación Algal Nociva en Las Bahías del Puerto de Manzanillo, Colima (Marzo-Mayo 2007). Nuevo Vallarta: Memorias del XIV congreso de Ciencia y Tecnología del Mar.
Griffith, A. W., Shumway, S. E., and Gobler, C. J. (2019). Differential mortality of north atlantic bivalve molluscs during harmful algal blooms caused by the dinoflagellate, Cochlodinium (a.k.a. Margalefidinium) polykrikoides. Estuaries Coasts 42, 190–203. doi: 10.1007/s12237-018-0445-0
Guzmán, M. H., Cortés, J., Glynn, W. P., and Richmond, H. R. (1990). Coral mortality associated with dinoflagellate bloom in the eastern Pacific (Costa Rica and Panama). Mar. Ecol. Prog. Ser. 60, 299–303.
Hallegraeff, G. M. (1992). Harmful algal blooms in the Australian region. Mar. Pollut. Bull. 25, 186–190. doi: 10.1016/0025-326X(92)90223-S
Hallegraeff, G., Dorantes-Aranda, J. J., Mardones, J., and Seger, A. (2017). “Review of progress in our understanding of fish-killing microalgae: implications for management and mitigation,” in Proceedings of the 17th International Conference on Harmful Algae. International Society for the Study of Harmful Algae and Intergovernmental Oceanographic Commission of UNESCO 2017, eds L. A. O. Proença and G. Hallegraeff (Brasil).
Hargraves, E. P., and Víquez, R. (1981). The dinoflagellate red tide in Golfo de Nicoya. Costa Rica. Rev. Biol. Trop. 29, 31–38.
Jeong, H. J., Yoo, Y. D., Kim, J. S., Kim, T. H., Kim, J. H., Kang, N. S., et al. (2004). Mixotrophy in the phototrophic harmful alga Cochlodinium polykrikoides (Dinophycean): prey species, the effects of prey concentration, and grazing impact. J. Eukaryot. Microbiol. 51, 563–569.
Kim, C. S., Lee, S. G., Kim, H. G., and Jung, J. (1999). Reactive oxygen species as causative agents in the ichthyotoxicity of the red tide dinoflagellate Cochlodinium polykrikoides. J. Plankton Res. 21, 2105–2115.
Kim, H. G. (1998). “Cochlodinium polykrikoides blooms in Korean coastal waters and their mitigation,” in Harmful Algae, eds B. Reguera, J. Blanco, T. Fernández, and T. Wyatt (Vigo: Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO), 227–228.
Kudela, R. M., and Gobler, C. J. (2012). Harmful dinoflagellate blooms caused by Cochodinium sp.: global expansion and ecological strategies facilitating blooms formation. Harmful Algae 14, 71–86. doi: 10.1016/j.hal.2011.10.015
Landsberg, J. H. (2002). The effects of harmful algal blooms on aquatic organisms. Rev. Fish. Sci. 10, 113–390. doi: 10.1080/20026491051695
Lee, J. S. (1996). Bioactive components from the red tide plankton Cochlodinium polykrikoides. J. Korean Fish. Soc. 29, 165–173.
León-Muñoz, J., Urbina, M. A., Garreaud, R., and Iriarte, J. L. (2018). Hydroclimatic conditions trigger record harmful algal bloom in western Patagonia (summer 2016). Sci. Rep. 8:1330. doi: 10.1038/s41598-018-19461-4
López-Cortés, D. J., Band-Schmidt, C. J., Bustillos-Guzmán, J. J., Hernández-Sandoval, F. E., Mendoza-Flores, A., and Núñez-Vázquez, E. J. (2014). Condiciones ambientales durante un florecimiento de Cochlodinium polykrikoides (Gymnodiniales, Dinophyceae) en la Ensenada de La Paz, Golfo de California. Rev. Biol. Mar. Oceanog. 40, 97–110. doi: 10.4067/S0718-19572014000100011
Maciel-Baltazar, E., and Hernández-Becerril, D. U. (2013). Especies de dinoflagelados atecados (Dinophyta) de la costa de Chiapas, sur del Pacífico Mexicano. Rev. Bio. Mar. Oceanog. 48, 245–259. doi: 10.4067/S0718-19572013000200005
Malagón, L. A., and Perdomo, L. V. (2013). Cochlodinium polykrikoides bloom in the Colombian caribbean. Harmful Algae News 47, 14–15.
Mardones, J. I., Fuenzalida, G., Zenteno, K., Alves-de-Souza, C., Astuya, A., and Dorantes-Aranda, J. J. (2019). Salinity-growth response and ichthyotoxic potency of the Chilean Pseudochattonella verruculosa. Front. Mar. Sci. 6:24. doi: 10.3389/fmars.2019.00024
Margalef, R. (1961). Hidrografía y fitoplancton de un área marina de la costa meridional de Puerto Rico. Invest. Pesq. 18, 33–96.
Mascareño, A., Cordero, R., Azócar, G., Billi, M., Henríquez, P. A., and Ruz, G. A. (2018). Controversies in social-ecological systems: lessons from a major red tide crisis on Chiloe Island, Chile. Ecol. Soc. 23:art15. doi: 10.5751/ES-10300-230415
Meave del Castillo, M. E. (2014). “Plancton marino introducido por agua de lastre,” in Especies Acuáticas Invasoras En México, eds R. Mendoza and P. Kolef (México, ME: Comisión Nacional para el conocimiento y Uso de la Biodiversidad), 289–308.
Mooney, B. D., Dorantes-Aranda, J. J., Place, A. R., and Hallegraeff, G. M. (2011). Ichthyotoxicity of gymnodinioid dinoflagellates: PUFA and superoxide effects in sheepshead minnow larvae and rainbow trout gill cells. Mar. Ecol. Prog. Ser. 426, 213–224.
Morales-Blake, A., Cavazos-Guerra, C., and Hernández-Becerril, D. U. (2001). Unusual HABs in Manzanillo bay. Colima, Mexico. Harmful Algae News 22:6.
Morales-Blake, A., Hernández-Becerril, D. U., and Cavazos-Guerra, C. (2000). “Registros de mareas rojas en la Bahía de Manzanillo, Colima, México,” in Estudios sobre plancton en México y el Caribe, eds E. Ríos-Jara, E. Juárez-Carrillo, M. Pérez-Peña, E. López-Uriarte, E. G. Robles-Jarero, D. U. Hernández-Becerril, et al. (México, ME: Universidad de Guadalajara), 81–82.
Morey-Gaines, G. (1982). Gymnodinium catenatum Graham (Dinophyceae): morphology and affinities with armoured forms. Phycologia 21, 151–163.
Muciño-Márquez, R.E. (2010). Variación estacional de la comunidad fitoplanctónica en las granjas atuneras en la Bahía de La Paz, Baja California Sur. Master Thesis, Centro Interdisciplinario de Ciencias Marinas-IPN: La Paz.
Mulholland, M. R., Morse, R. E., Boneillo, G. E., Bernhardt, P. W., Filipinno, K. C., Procise, L. A., et al. (2009). Understanding causes and impacts of the dinoflagellate Cochlodinium polykrikoides blooms in the Chesapeake. Estuaries Coasts 32, 734–747.
Núñez-Vázquez, E. J., Heredia-Tapia, A., Pérez-Urbiola, J. C., Alonso-Rodríguez, R., Arellano-Blanco, J., Cordera-Tapia, A., et al. (2003). “Evaluation of dinoflagellates toxicity in recent HAB event in the Gulf of California, Mexico,” in Proceedings of the HABTECH, Workshop, eds P. Holland, L. Rhodes, and L. Brown, Nelson.
Ochoa-Arguello, I. (2017). Marea roja en Centroamérica avanza hacia costas de Chiapas. El Orbe. Available at: https://elorbe.com/seccion-politica/local/2017/07/10/marea-roja-en-centroamerica-avanza-hacia-costas-de-chiapas.html (accessed July 10, 2017).
Onoue, Y., and Nozawa, K. (1989). “Separation of toxins from harmful red tides occurring along the coast of Kagoshima prefecture,” in Red tides: Biology, Environmental, Science, and Toxicology, eds T. Okaichi, D. M. T. Anderson, and T. Nemoto (New York, NY: Elsevier), 371–374.
Orozco, R., Quispe, Y., Lorenzo, A., and Zamudio, M. L. (2017). Asociación de floraciones de algas nocivas y Vibrio spp., en áreas de pesca y acuicultura de bivalvos de moluscos en las bahías de Sechura y Pisco. Perú. Rev. Per. Biol. 24, 111–116. doi: 10.15381/rpb.v24i1.13111
Osorio-Tafall, B. F. (1943). El mar de Cortés y la productividad fitoplanctónica de sus aguas. An. Esc. Nal. Cienc. Biolog. Mex. 3, 73–118.
Páez-Osuna, F., Álvares-Borrego, S., Ruiz-Fernández, A. C., and García-Hernández, J. (2017). Environmental status of Gulf of California: a pollution review. Earth Sci. Rev. 166, 181–2005. doi: 10.1016/j.earscirev.2017.01.014
Pérez-Morales, A., Band-Schmidt, C. J., and Martínez-Díaz, S. F. (2017). Mortality on zoea stage of the pacific white shrimp Litopenaeus vannamei caused by M. polykrikoides (Dinophyceae) and Chattonella spp. (Raphidophyceae). Mar. Biol. 164:57. doi: 10.1007/s00227-017-3083-3
Ramírez, A., Szelistowski, W., and Lopez, M. (1989). Spawning pattern and larval recruitment in Gulf of Nicoya anchovies (Pisces: Engraulidae). Rev. Biol. Trop. 37, 55–62.
Reyes-Vásquez, G., Ferraz-Reyes, E., and Vásquez, E. (1979). “Toxic dinoflagellate blooms in northeastern Venezuela during 1977,” in Toxic Dinoflagellate Blooms, eds D. L. Taylor and H. H. Seliger (Amsterdam, NE: Elsevier-North Holland, Inc:).
Richlen, M. L., Morton, S. L., Jamali, E. A., Rajan, A., and Anderson, D. M. (2010). The catastrophic 2008-2009 red tide in the Arabian Gulf region, with observations on the identification and phylogeny of the fish-killing dinoflagellate Cochlodinium polykrikoides. Harmful Algae 9, 163–172. doi: 10.1016/j.hal.2009.08.013
Schütt, F. (1895). Die Peridineen der Plankton-Expedition. I. Theil Studien Über die Zellen der Peridineen. Ergebnisse der Plankton-Expedition der Humboldt-Stiftung, IV. M.a.A. Kiel, Leipzig: Lipsius und Tischler.
Sierra-Beltrán, A., Lluch-Cota, D. B., Cortés-Altamirano, R., Castillo-Chávez, M., Carrillo, L., Pacas, L., et al. (2001). “Possibilities for environmental and biological monitoring in the Pacific coast of central and north america as a strategy for prevention and mitigation of harmful algal blooms (HABs). an example of small budget cooperative efforts leading to a mayor benefit,” in Proceeding of the Second International Conference on Harmful Algae Management and Mitigation, eds Z. Mingyuan, Z. Yinglin, L. Cheong, and S. Hall, Brasil.
Tomas, C. R., and Smayda, T. J. (2008). Red tides blooms of Cochlodinium polykrikoides in a coastal cove. Harmful Algae 7, 308–317. doi: 10.1016/j.hal.2007.12.005
Torres-Zambrano, G. (2000). Mareas rojas durante 1989-1999, en aguas ecuatorianas. Acta Oceanog. Pacific, INOCAR, Ecuador. 10, 127–136.
Valverde, J. O. (2002). The Longest and Recent Red Tide Event in Costa Rica. Book of Abstracts, Molluscan Shellfish Safety. Galicia: Santiago de Compostela.
Vargas-Montero, M., and Freer, E. (2004). Proliferaciones algales nocivas de cianobacterias (Oscillatoriaceae) y dinoflagelados (Gymnodiniaceae) en el Golfo de Nicoya, Costa Rica. Rev. Biol. Trop. 52(Suppl.1), 121–125.
Vargas-Montero, M., Freer, B. E., Guzmán, J. C., and Vargas, J. C. (2008). Florecimientos de dinoflagelados nocivos en la costa Pacífico de Costa Rica. Hidrobiológica 18(Suppl.1), 15–23.
Vargas-Montero, M., Freer, E., Jiménez-Montealegre, R., and Guzmán, J. C. (2006). Occurrence and predominance of the fish killer Cochlodinium polykrikoides on the Pacific coast of Costa Rica. Afr. J. Mar. Sci. 28, 215–217. doi: 10.2989/18142320609504150
Víquez, R., and Hargraves, E. P. (1995). Annual cycle of potentially harmful dinoflagellate in the Gulf of Nicoya. Costa Rica. Bull. Mar. Sci. 57,0 467–475.
Whyte, J. N. C., Haigh, N., Ghinter, N. G., and Keddy, L. (2001). First record of blooms of Cochlodinium sp. (Gymnodiniales, Dinophyceae) causing mortality to aquacultured salmon on the west coast of Canada. Phycologia 40, 298–304. doi: 10.2216/i0031-8884-40-3-298.1
Zhong, T. Y., and Gobler, C. J. (2009). Characterization of the toxicity of Cochlodinium polykrikoides isolates from Northeast US estuaries to finfish and shellfish. Harmful Algae 8, 454–462. doi: 10.1016/j.hal.2008.10.001
Keywords: dinoflagellates, Margalefidinium polykrikoides, Cochlodinium polykrikoides, HAB, Latin America
Citation: López-Cortés DJ, Núñez-Vázquez EJ, Dorantes-Aranda JJ, Band-Schmidt CJ, Hernández-Sandoval FE, Bustillos-Guzmán JJ, Leyva-Valencia I and Fernández-Herrera LJ (2019) The State of Knowledge of Harmful Algal Blooms of Margalefidinium polykrikoides (a.k.a. Cochlodinium polykrikoides) in Latin America. Front. Mar. Sci. 6:463. doi: 10.3389/fmars.2019.00463
Received: 05 July 2018; Accepted: 10 July 2019;
Published: 20 August 2019.
Edited by:
Angel Borja, Centro Tecnológico Experto en Innovación Marina y Alimentaria (AZTI), SpainReviewed by:
Maximo Jorge Frangopulos, University of Magallanes, ChileKyoko Yarimizu, National Research Institute of Fisheries Science, Japan Fisheries Research and Education Agency, Japan
Copyright © 2019 López-Cortés, Núñez-Vázquez, Dorantes-Aranda, Band-Schmidt, Hernández-Sandoval, Bustillos-Guzmán, Leyva-Valencia and Fernández-Herrera. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Erick J. Núñez-Vázquez, ZW51bmV6MDRAY2libm9yLm14
†Deceased