 Marina Arregui1
Marina Arregui1 Yara Bernaldo de Quirós1*Pedro Saavedra2
Yara Bernaldo de Quirós1*Pedro Saavedra2 Eva Sierra1Cristian M. Suárez-Santana1
Eva Sierra1Cristian M. Suárez-Santana1 Manuel Arbelo1
Manuel Arbelo1 Josué Díaz-Delgado1,3
Josué Díaz-Delgado1,3 Raquel Puig-Lozano1Marisa Andrada1
Raquel Puig-Lozano1Marisa Andrada1 Antonio Fernández1
Antonio Fernández1- 1Atlantic Center for Cetacean Research, Institute of Animal Health and Food Safety (IUSA), Veterinary School, University of Las Palmas de Gran Canaria, Las Palmas, Spain
- 2Department of Mathematics, University of Las Palmas de Gran Canaria, Las Palmas, Spain
- 3Laboratory of Wildlife Comparative Pathology, Department of Pathology, School of Veterinary Medicine and Animal Science, University of São Paulo, São Paulo, Brazil
Strikes between vessels and cetaceans have significantly increased worldwide in the last decades. The Canary Islands archipelago is a geographical area with an important overlap of high cetacean diversity and maritime traffic, including high-speed ferries. Sperm whales (Physeter macrocephalus), currently listed as a vulnerable species, are severely impacted by ship strikes. Nearly 60% of sperm whales’ deaths are due to ship strikes in the Canary Islands. In such cases, subcutaneous, muscular and visceral extensive hemorrhages and hematomas, indicate unequivocal antemortem trauma. However, when carcasses are highly autolyzed, it is challenging to distinguish whether the trauma occurred ante- or post-mortem. The presence of fat emboli within the lung microvasculature is used to determine a severe “in vivo” trauma in other species. We hypothesized fat emboli detection could be a feasible, reliable and accurate forensic tool to determine ante-mortem ship strikes in stranded sperm whales, even in decomposed carcasses. In this study, we evaluated the presence of fat emboli by using an osmium tetroxide (OsO4)-based histochemical technique in lung tissue of 24 sperm whales, 16 of them with evidence of ship strike, stranded and necropsied in the Canaries between 2000 and 2017. About 70% of them presented an advanced autolysis. Histological examination revealed the presence of OsO4-positive fat emboli in 13 out of the 16 sperm whales with signs of ship strike, and two out of eight of the “control” group, with varying degrees of abundance and distribution. A classification and regression tree was developed to assess the cut off of fat emboli area determining the high or low probability for diagnosing ship-strikes, with a sensitivity of 89% and a specificity of 100%. The results demonstrated: (1) the usefulness of fat detection as a diagnostic tool for “in vivo” trauma, even in decomposed tissues kept in formaldehyde for long periods of time; and (2) that, during this 18-year period, at least, 81% of the sperm whales with signs of ship strike were alive at the moment of the strike and died subsequently. This information is highly valuable in order to implement proper mitigation measures in this area.
Introduction
Strikes between vessels and cetaceans (“ship strikes”) have become an issue of concern in the last decades due to an increase of the number and speed of ships (Laist et al., 2001). Reports of ship strikes have been published worldwide with fin whales (Balaenoptera physalus), humpback whales (Megaptera novaeangliae), North Atlantic right whales (Eubalaena glacialis) and sperm whales (Physeter macrocephalus) being the most affected species (Van Waerebeek and Leaper, 2008). Areas with high cetacean diversity and high maritime traffic overlap have been identified as hot spots as ship strikes may compromise the population status of some cetacean species in those areas. In Europe, these areas include the Mediterranean Sea (Panigada et al., 2006; Frantzis et al., 2019), the Strait of Gibraltar (de Stephanis and Urquiola, 2006) and the Canary Islands (Carrillo and Ritter, 2010).
The Canary Islands form a Spanish archipelago of seven main volcanic islands, located in the north-west of Africa. It is one of the richest areas for cetacean biodiversity in the Northeast Atlantic, with 30 species identified, the sperm whale among them (Tejedor and Martín, 2013). Sperm whales are present year round in Canarian waters, with higher numbers in spring and autumn due to seasonal migrations (André, 1997). They are listed as vulnerable by the International Union for Conservation of Nature (Taylor et al., 2008), and are the most affected species by ship strikes in Canarian waters (Arbelo et al., 2013; Díaz-Delgado et al., 2018). Some factors proposed to explain the susceptibility of sperm whales to ship strikes are: (1) long periods at the surface for socialization or resting after prolonged dives (Whitehead and Weilgart, 1991; André, 1997; Watkins et al., 1999; Watwood et al., 2006); (2) drift-dives, performed at a low-activity state, which will allow them to perform bi-hemispheric sleep, being unaware of approaching ships until being touched (Miller et al., 2008); or (3) possible loss of sensitivity to low-frequency sounds produced by ship engines in Canarian waters (André, 1997).
International but mainly inter-island ferry traffic in the Canarian waters has increased considerably in the last years including: normal ferries (15–20 knots), fast ferries (21–29 knots), and high-speed ferries (≥30 knots) (Aguilar et al., 2000; de Stephanis and Urquiola, 2006; Ritter, 2010). Vanderlaan and Taggart (2007), used North Atlantic right whale ship strike data to develop a model of the probability of mortality based on strikes occurring at different speeds, regardless of vessel size. The authors suggested that strikes at speeds over 18 knots were fatal almost 100% of the time.
When whales get hit by a vessel, they can present: sharp trauma lesions, generated by the propeller or the keel of the vessel, and/or, blunt trauma lesions, caused by a non-rotating feature of the vessel, like the hull or the skeg (Campbell-Malone et al., 2008; Moore et al., 2013). Injuries within the first category, usually involve the presence of one or more linear to curvilinear laminar incising wounds, that usually cause damage to the underlying soft tissue. Extreme injuries, frequently lethal, involve damage to the axial musculature or the vertebral column affecting locomotion, or even the complete separation of part of the body, with severe central nervous system (CNS) injury. In blunt traumas, areas of hemorrhage and edema in the blubber, subcutaneous tissue, and skeletal muscle are common features, as well as luxations and/or fractures, usually concomitant. In more severe cases, rupture of internal organs can be observed (Campbell-Malone et al., 2008; Moore et al., 2013). Full necropsies should be carried out as some injuries, especially those related with blunt trauma, may not be apparent externally. In very decomposed carcasses, differentiation between ante-mortem lesions and post-mortem changes can be very challenging (Campbell-Malone et al., 2008; Moore et al., 2013).
Fat embolism is defined as the mechanical obstruction of blood vessels by circulating fat particles (Watson, 1970; Hulman, 1995). In humans it is usually related with traumas involving long and pelvic bones (Watson, 1970; Fulde and Harrison, 1991). After trauma, fat cells from the bone marrow of fractured bones or from damaged soft tissues, enter the bloodstream through torn venules in the injury or fracture site, and typically first arrive to the lungs where they may get trapped within the pulmonary microvasculature (<20 μm in diameter) (Watson, 1970). For this reason, the lung is considered a target organ for fat emboli detection (Levy, 1990). The presence of fat emboli within the lungs constitutes evidence of antemortem injury, as cardiac function is needed, even for a short time, to allow the circulation of fat droplets to the lungs (Armstrong et al., 1955; Mason, 1968; Saukko and Knight, 2004). It is a common and usually asymptomatic finding (Watson, 1970; Fulde and Harrison, 1991), that infrequently leads to a clinical disorder known as fat embolism syndrome (Glover and Worthley, 1999). Its severity has been related to the multiplicity of the fractures, and it occurs very rapidly after severe trauma (Tanner et al., 1990), being also present in those cases in which the death occurs immediately after the trauma (Emson, 1958).
In the Canary Islands, over 57% of the sperm whales stranded since 2000 presented evidence of ship strike, and over 70% of them were in an advanced or very advanced decomposition state (Arbelo et al., 2013; Díaz-Delgado et al., 2018). Thus, we aimed to analyze lungs from sperm whales dead in Canarian waters between 2000 and 2017 with signs of ship strike to determine: (1) if fat embolism is a common finding in sperm whale’s lung tissue, (2) if the presence of fat emboli within the lung vessels is a useful diagnostic tool to assess ante-mortem ship strikes, and (3) if lung fat emboli density relates to the severity or location of the traumatic injuries.
Materials and Methods
Animals Included in the Study
Between January 2000 and December 2017, 35 sperm whales encountered dead, floating or stranded, in the Canary Islands (28°N, 16°W; Spain) were necropsied, following standardized protocols (Kuiken and García Hartmann, 1991), to find out the cause of death. Required permission for the management of stranded cetaceans was issued by the environmental department of the Canary Islands’ Government and the Spanish Ministry of Environment. No experiments were performed on live animals.
Age categories were established based on total body length (Perrin et al., 2009) and histologic gonadal examination, including: neonate, calf, juvenile, subadult and adult (Geraci and Lounsbury, 2005). Decomposition code was established according to Kuiken and García Hartmann (1991) classification, with a modification for code 1: code 1 for “very fresh” was assigned to an animal that has recently died. The other codes remained the same: code 2 for “fresh dead animals” (no bloating nor changes in coloration, eatable meat), code 3 for “moderate autolysis” (may present with some skin desquamation, the carcass might have started to swollen, and organs may have changed coloration and more friable), code 4 for “advanced autolysis” (skin desquamation, swollen carcass, organs difficult to recognize), and code 5 for “very advanced autolysis” (the skin may be absent, some or all organs may be liquefied, also mummification or adipocera may be observed in some carcasses). Body condition was determined based on anatomical parameters such as the presence of certain prominent bones, the dorso-axial muscular mass, and the presence or absence of fat deposits (Joblon et al., 2014).
The lungs of 16 sperm whales with evidence of ship strike and eight sperm whales without (control group), were studied to detect fat emboli (Table 1) using the osmium tetroxide (OsO4) technique.
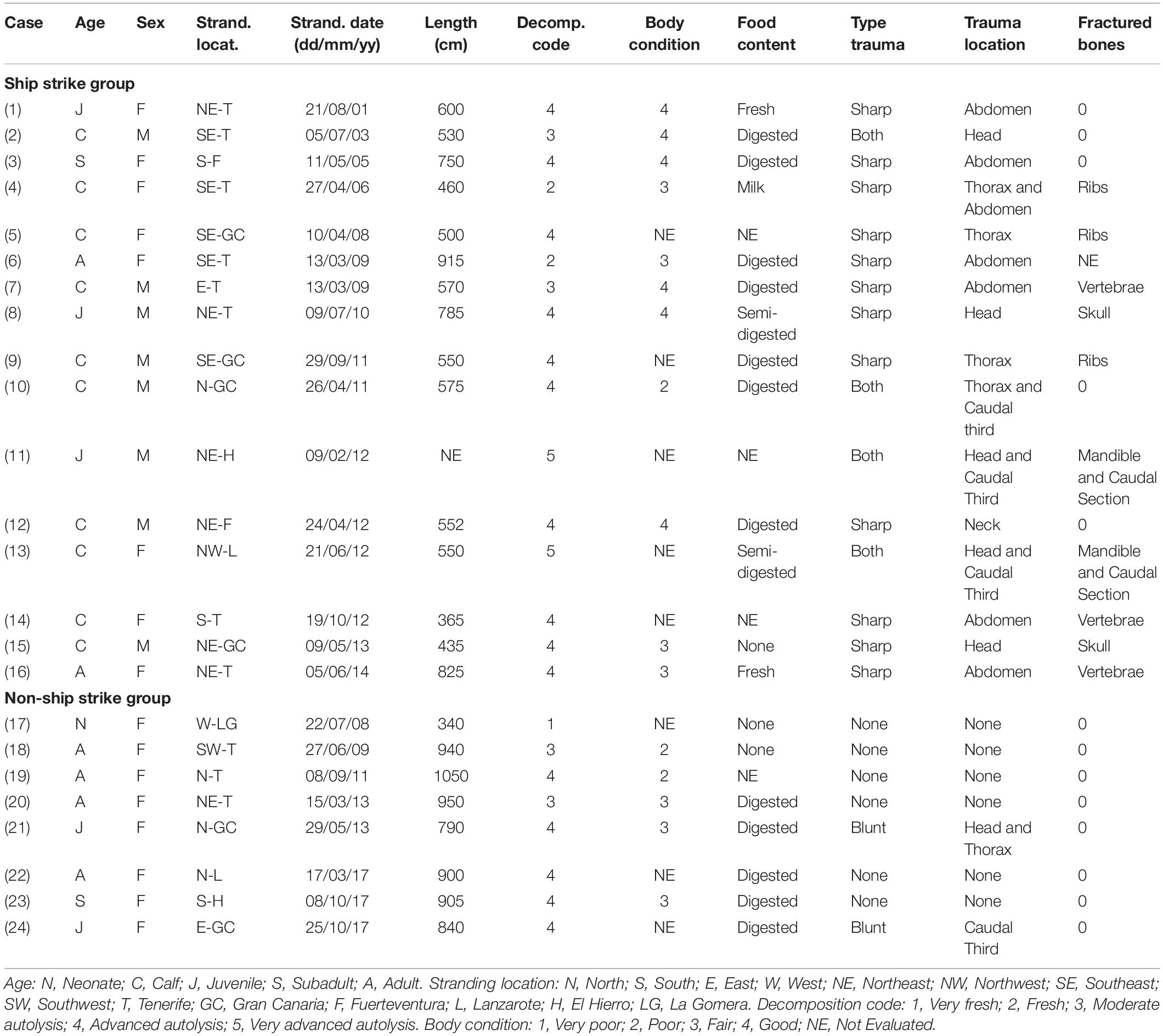
Table 1. Epidemiological and biological data of the sperm whales included in the present study with evidence of ship strike.
Osmium Tetroxide Technique
A retrospective study was carried out using lung tissue samples fixed in 10% buffered formalin between 2000 and 2017 and kept in the Institute of Animal Health Tissue Bank. Formalin-fixed lung samples were cut into thin sections (≤1 mm) to ensure the proper penetration of OsO4. Post-fixation with OsO4 is needed as lipids are soluble in the processing solvents used to embed the tissues in paraffin. The sections were then washed with running tap water for 20 min followed by 10 min in distilled water. Next, the sections were immersed in 1% OsO4 aqueous solution (sonication was previously used to dissolve the commercial crystalline OsO4) within hermetically sealed bottles on a shaker inside a chemical hood. Then, the sections were rinsed in running tap water for 30 min and immersed in 1% periodic acid until the dark osmicated tissues were uniformly cleared (Abramowsky et al., 1981). Samples were washed for 30 min with tap water, and rinsed three times with distilled water. Then, the samples were routinely processed and embedded in paraffin-wax, sectioned at 5 μm-thick, treated with picric acid (1% in ethanol 96%) for 24 h to remove excess formalin pigment (Abramowsky et al., 1981), and counterstained with hematoxylin and eosin (HE). Finally, slides were mounted in DPX mounting medium. Tissue sections as blubber and rete mirabile (which have abundant adipocytes), were used as positive controls (treated with OsO4) and negative controls (non-treated with OsO4) to validate the technique.
Microscopic Analysis
All lung sections were evaluated for the presence/absence of fat emboli, as well as the area occupied by those emboli within lung vessels using light microscopy (Olympus BX51).
Each lung tissue section was divided in “N” number of 100× magnified microscopic fields (MF) (Ocular: 10× and Objective: 10×). A Bootstrap analysis was carried out to determine the number of 100 MFs (“n”) that needed to be studied for each tissue section (Table 2). We considered a good estimation of the true value if the total bound error was below 8%. Depending on the total tissue section area the Bootstrap analysis yielded results between 14 and 20 100 MFs. These fields were randomly selected and photographed using an Olympus XC30 camera (Olympus Soft Imaging Solutions GmbH©, Johann-Krane-Weg 39, D-48149 Münster) (Table 2). Fields containing pleura (adipocytes are normally present in the pleura of sperm whales) (Figure 1B), and/or large-diameter bronchi/bronchioles (empty spaces) were discarded to ensure a similar parenchyma size comparison between the different fields.
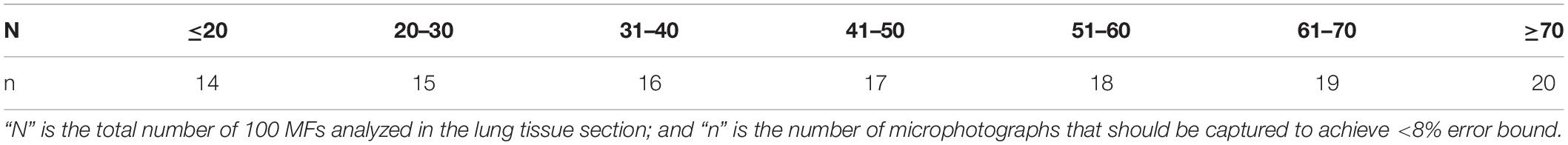
Table 2. Number of microphotographs captured based on the total number of 100 magnifications microscopic fields (100 MFs) of the tissue section.
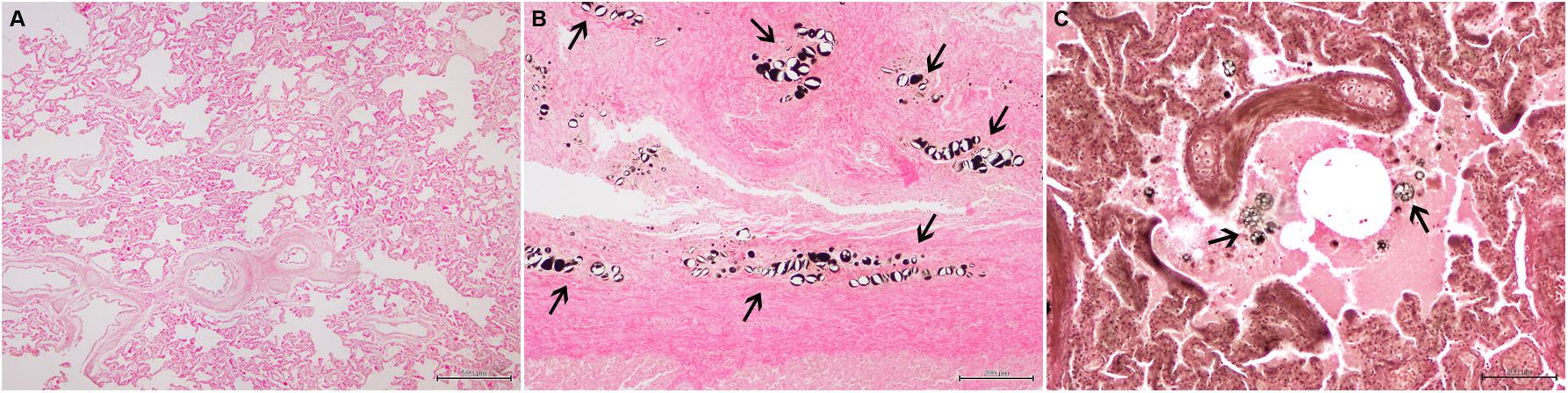
Figure 1. Lung sections treated with OsO4. (A) Appearance of a lung parenchyma negative to fat emboli (Bar = 500 μm). (B) Black-stained adipocytes in the pleura (arrows) (Bar = 200 μm). (C) Black-stained fat droplets within a bronchiole (arrows) (Bar = 200 μm).
The software ImageJ (1.48v, Wayne Rasband, National Institute of Health, United States) was used to determine the area occupied by fat emboli, in pixels, in each of the photographs. Each of the 100 MFs’ photographs has a total area of 1,920,000 pixels. Fat emboli are recognized as black droplets primarily in the lumen of capillaries and small- and medium-size arteries. The software ImageJ allows the quantification of areas of a certain color automatically, or the quantification of selected areas manually. We manually selected the fat emboli areas in the lung parenchyma, as not all black areas were fat emboli [e.g., various artifacts, fat in bronchioli and/or alveoli (Figure 1C)], and the emboli were not homogenously stained. As a result, for each animal, we ended up having an “n” number of 100 MFs (photographs), each of them with an area in pixels occupied by fat emboli.
Analysis for the Validation of the Osmium Tetroxide Technique as a Complementary Diagnostic Tool for Ship-Strikes
The 25th, 50th, 75th, and 90th percentiles of the areas (pixels) occupied by fat emboli in the “n” 100 MFs studied were calculated for each animal (Table 3). As fat emboli were also present in a few lungs of the “control group,” a classification and regression tree (CART) was developed to obtain a cut-off value from which the probability of association with ship-strike is high. This procedure classifies data using a sequence of if–then rules. The basis of the decision tree algorithms is the binary recursive partitioning of the data. The most discriminative variable is first selected to partition the data set into child nodes. The splitting continues until some stopping criterion is reached. The tree was constructed according to the following algorithm: in the first stage, the tree grows until all cases are correctly classified, and in the second stage, we used the tenfold cross-validation method of successive pruning (Breiman et al., 1984). Finally, the tree that minimized the error measurement (deviance) was chosen. Then, the low and high-probability categories obtained were compared using the exact Fisher test. The sensitivity and specificity were estimated by means of 95% confidence intervals (95% CI).
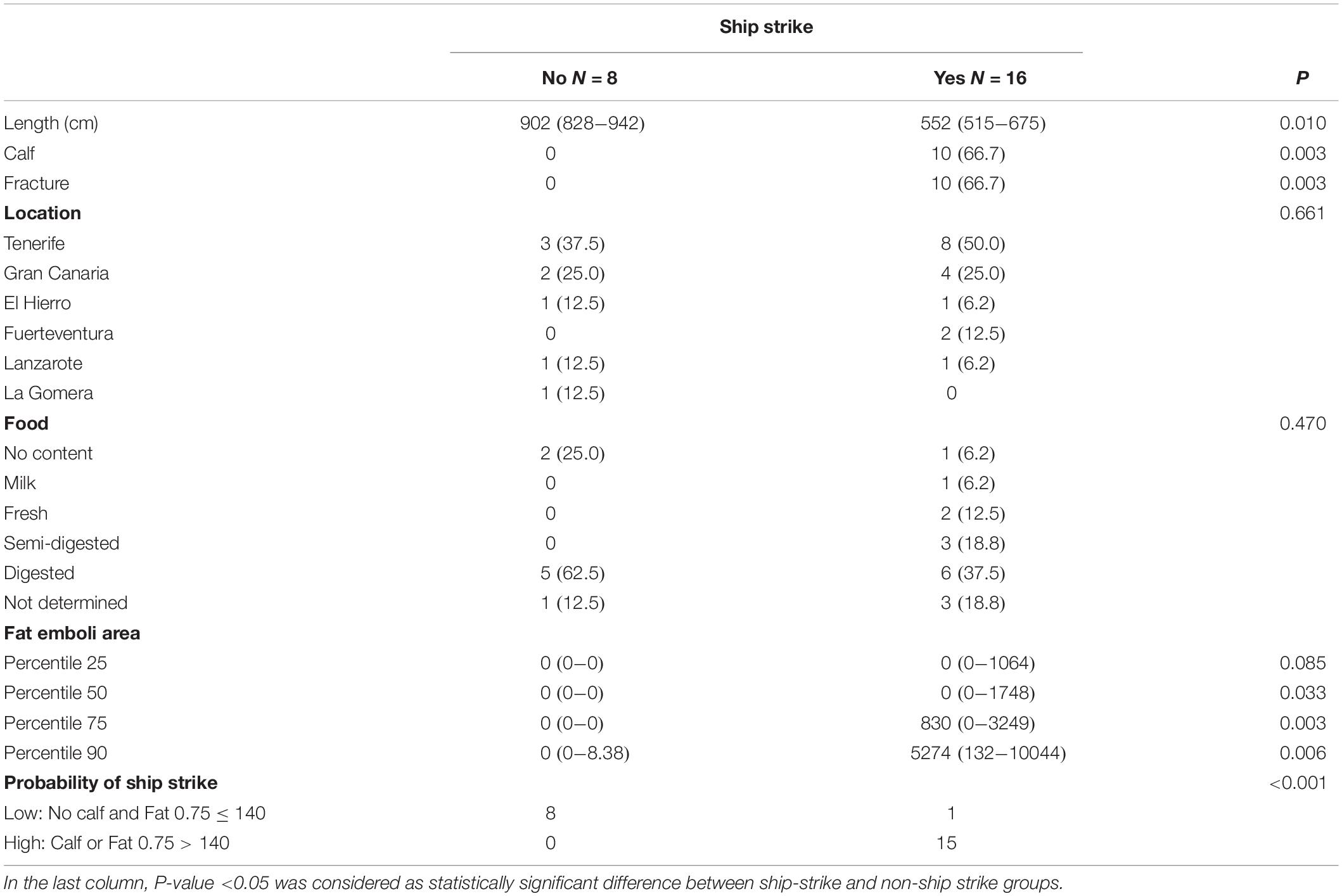
Table 3. Categorical variables studied expressed as frequencies and percentages or continuous, expressed as medians and interquartile ranges (IQR = 25th–75th percentile).
Exploration of Association Between Trauma-Related Variables and Fat Emboli Severity
Categorical variables were expressed as frequencies and percentages, and continuous variables, like fat emboli areas, were expressed as medians and interquartile ranges (IQR = 25th–75th percentile) (Table 3).
The variables age, presence/absence stomach food content and degree of digestion of the ingesta, presence of fractures and stranding location were compared between both groups using the Chi-square (χ2) test or the exact Fisher test for percentages; and the Wilcoxon test for independent data for the medians (Table 3).
The variables age, sex, body condition, presence/absence and degree of digestion of the stomach food content, trauma location and presence/absence of fractures were analyzed within the group with evidence of ship strike to assess potential associations with fat emboli severity (lung area occupied by fat emboli) (Table 1). For this aim, a linear analysis was carried out. The variables introduced in the model were age (calf/not calf), sex, body condition, presence/absence and degree of digestion of the stomach food content, trauma location and presence/absence of fractures. Then, a selection of variables based on the Akaike information criteria was performed.
Data were analyzed using the R package, version 3.3.1 (R Development Core Team, 2016).
Results
Presence of Fat Embolism
A total of 83% (13/16) sperm whales with evidence of ship strike (Figure 2) had intravascular OsO4-positive fat emboli. Fat emboli ranged from 67 to 59773 pixels, and were seen circulating in medium and small caliber intrapulmonary arteries and/or obliterating arterioles and capillaries, both in fresh and decomposed specimens (Figure 3). None but two of eight “control” sperm whales had detectable fat emboli (Figure 1A). Those two animals (cases 21 and 24) had rare isolated OsO4-positive fat emboli (<650 pixels) in arterioles (Table 3).
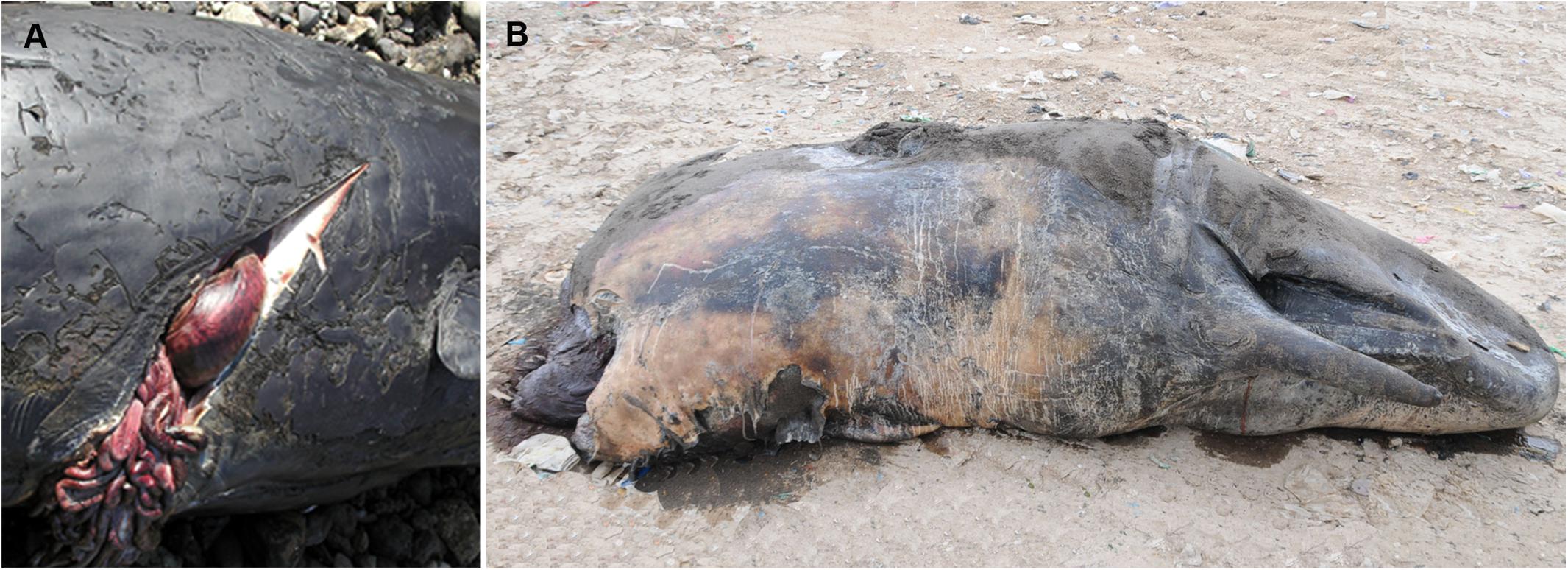
Figure 2. Vessel strike-related injuries in sperm whales (Physeter microcephalus) stranded in the Canary Islands. (A) Presence of a deep incision in the right flank of the animal, caudal to the pectoral fin, with soft tissue exposure, abdominal perforation and evisceration and costal fractures, case 4. (B) Complete amputation of the vertebral column at the level of the last thoracic vertebrae, case 14.
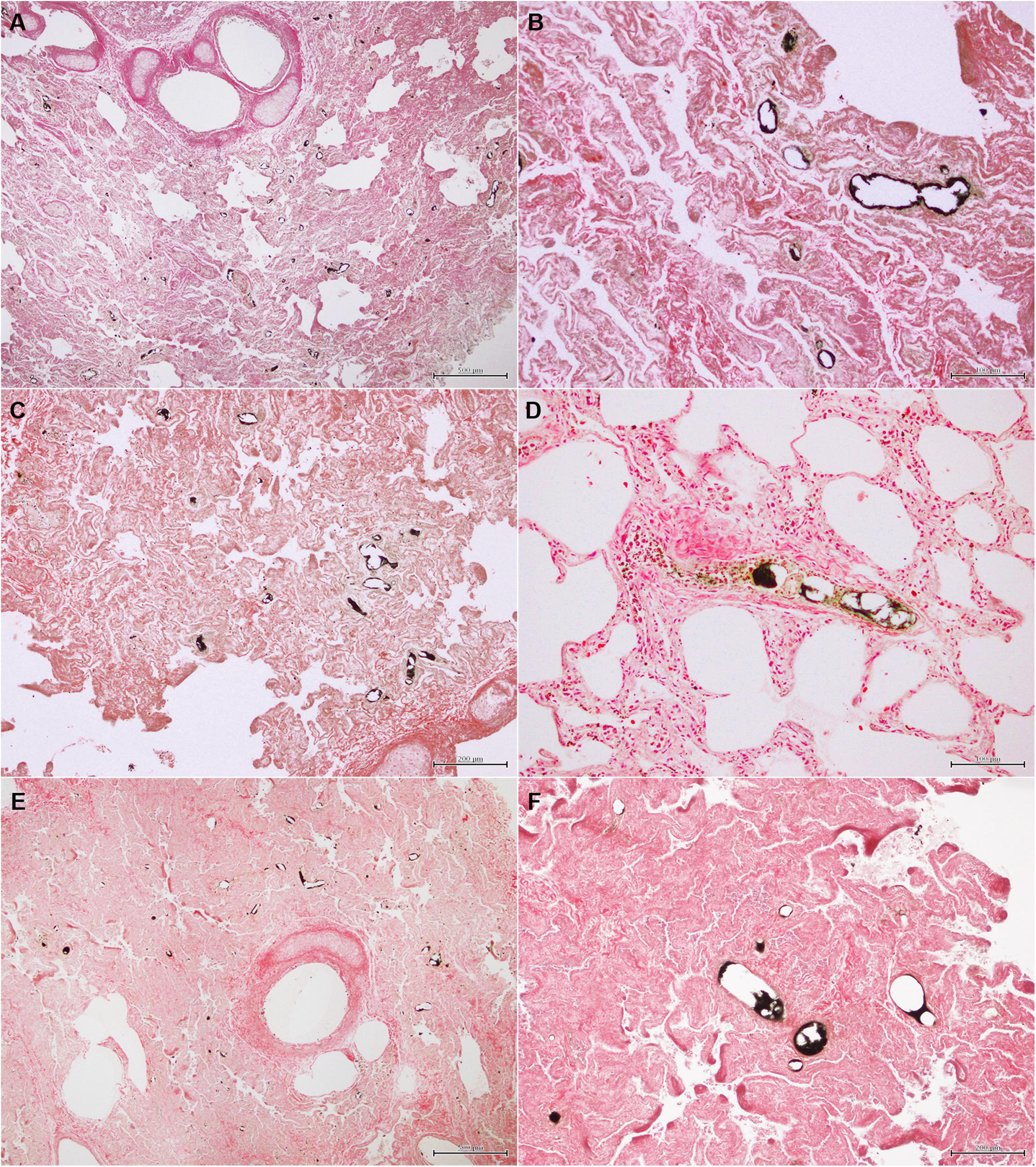
Figure 3. Black stained (OsO4-positive) fat emboli in the lung microvasculature, mainly arterioles, of some of the sperm whales stranded in the Canary Islands with signs of ship strike (A–F). Bar: A = 500 μm; B = 100 μm; C = 200 μm; D = 100 μm; E = 500 μm; F = 200 μm.
Calves were “significantly” (p = 0.003) more likely to be involved in ship-strikes than other age categories. As well, the presence of fractures was “significantly” associated to ship-strikes p = 0.003. Other variables studied like the stranding location (island) or the presence/absence and degree of digestion of the ingesta, were not significantly different between both groups (Table 3). An association between trauma-related variables and fat emboli severity could not be established.
When assessing the probability of ship strike based on the fat emboli area, significant differences between non-strike and strike groups started to be seen in the 50th-percentile values, but the highest discriminant power between both groups corresponded to the fat emboli area’s value of the 75th-percentile (Table 3). The CART indicates that if the value of the 75th-percentile fat emboli area is greater than 140 pixels in the animal studied (cut-off value), the probability of having been hit by a vessel is high, and so the animal is assigned to the strike group. If the value of the area occupied by fat emboli in the 75th-percentile is lower than 140 pixels, and the animal is a calf, it is also assigned to the strike group. If none of the previous conditions are met, the animals are assigned to the non-strike group (p < 0.001) (Table 3 and Figure 4). The sensitivity and the specifity were 89% (52–100; 95% CI) and 100% (78–100; 95% CI), respectively.
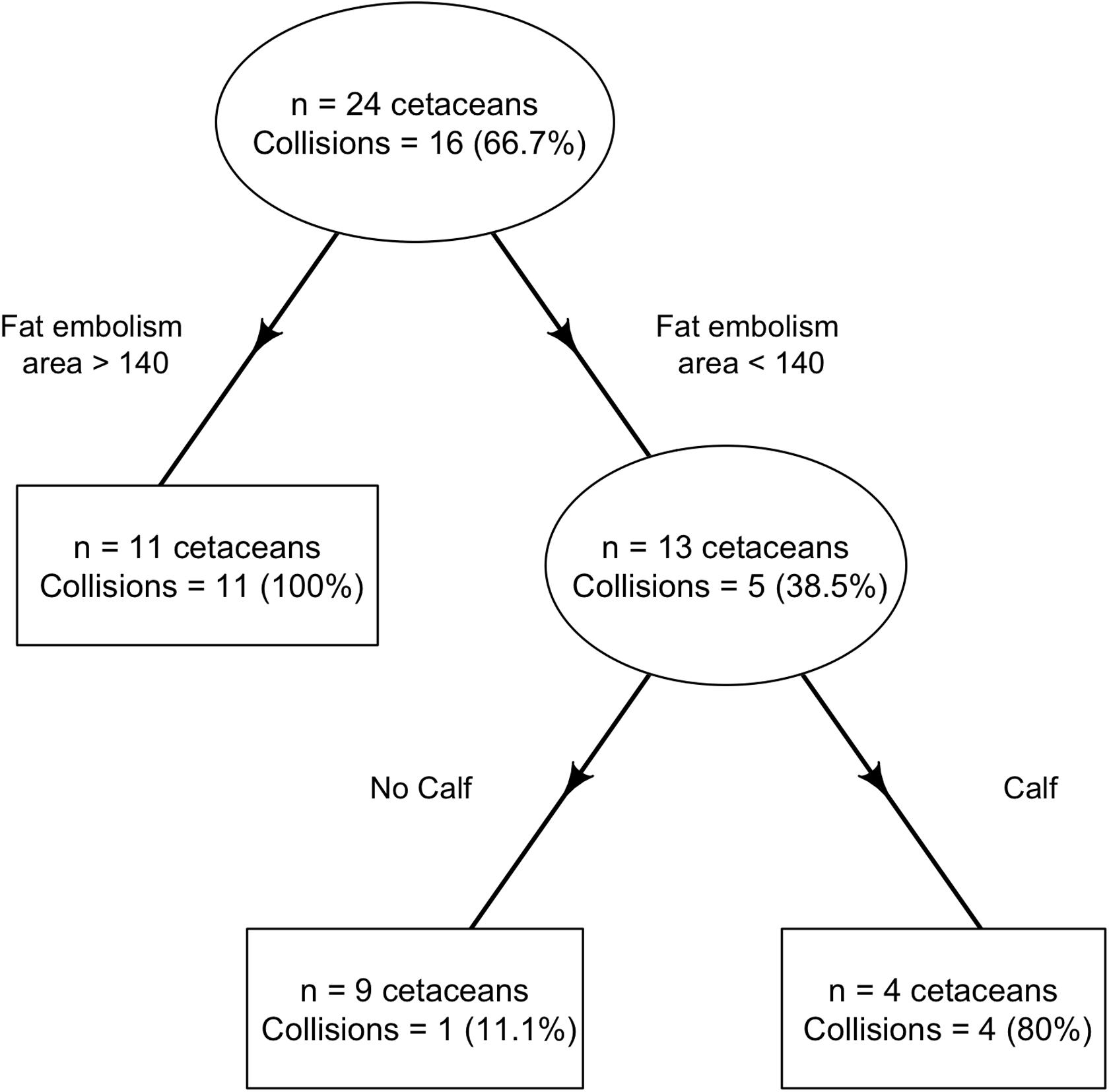
Figure 4. Result of the classification and regression tree to determine the cut off for ship-strike probability of the studied sperm whales. A high or low probability of ship strike was established according to: first, the fat emboli-P75 value and secondly, being a calf or not a calf. If fat emboli area <140 and Not a Calf → Non-strike group; If fat embolism area <140 and Calf → Strike group; and if fat embolism area >140 → Strike group. Its sensitivity is 89% (95% CI = 52–100%) and its specificity 100% (95% CI = 78–100%).
Factors Related to Ship Strikes in the Canary Islands
Most sperm whales with evidence of ship-strike were calves (10/16; 62.5%), followed by juveniles (3/16; 18.75%), adults (2/16; 12.5%) and subadults (1/16; 6.25%). All the adults/subadults included in this study were females.
Most of the animals with evidence of ship strike appeared floating or stranded along the east coast of Tenerife (56.25%), followed by the east coast of Gran Canaria (18.75%), east coast of Fuerteventura (12.5%), and finally east coast of El Hierro (6.25%) and west coast of Lanzarote (6.25%) (Figure 5).
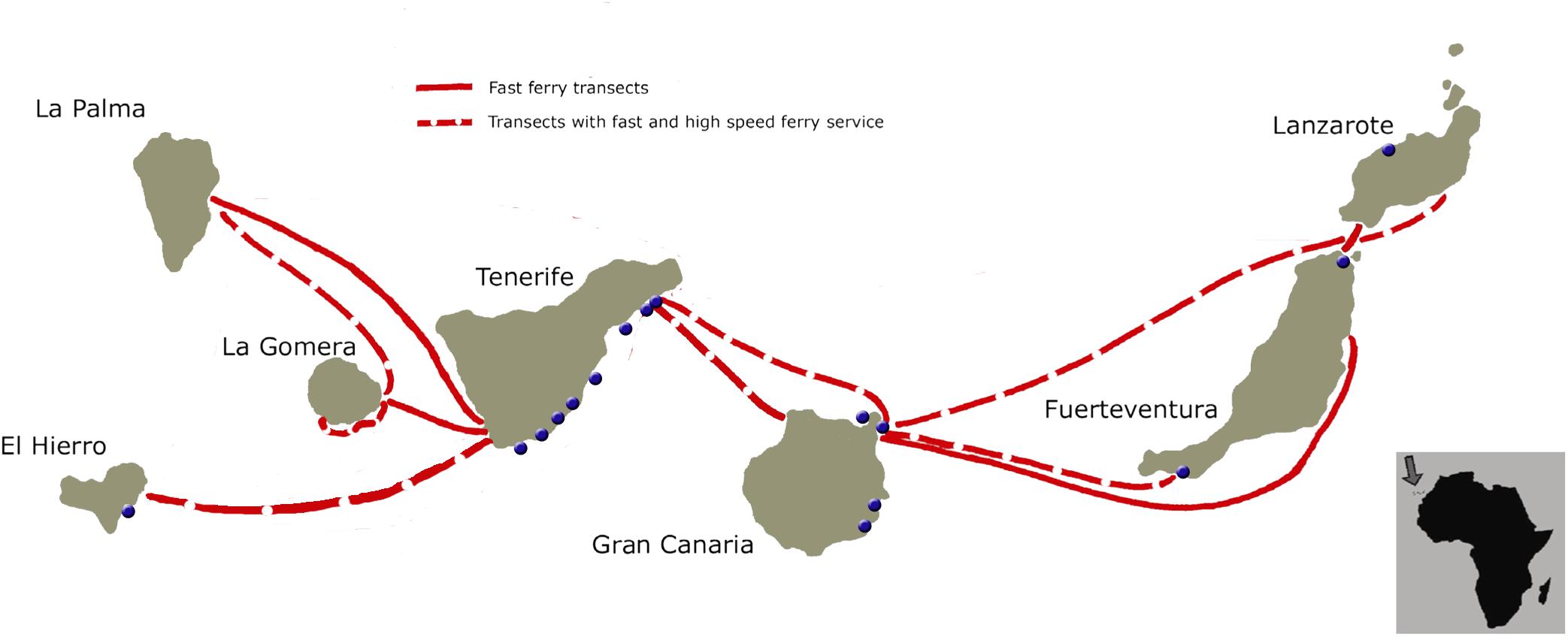
Figure 5. Inter-island ferry transects in the Canary Islands (different speed-ferries are represented with different line types), and stranding locations (blue dots) of the sperm whales with evidence of ship strikes in the Canary Islands [modified from Ritter (2010)].
Discussion
Presence of Fat Embolism and Its Significance in Ship Strikes
The lack of detectable fat emboli in lung tissue of most control sperm whales suggests that fat embolism is not a physiological or common finding in lungs of stranded sperm whales. In addition, the presence of abundant OsO4-positive fat emboli in most sperm whales with evidence of ship-strike indicates an association with trauma. The etiology of fat emboli in the blood stream may be trauma- or non-trauma related (Glover and Worthley, 1999). Trauma conditions may include marrow-containing bone fractures or adipocyte-rich soft tissue injuries (Watson, 1970; Fulde and Harrison, 1991; Gupta and Reilly, 2007). Both possibilities likely coexisted in our cases with evidence of ship-strike. The presence of fat emboli in the lung microvasculature indicates that the animal was alive at the moment of the strike and that cardiovascular collapse did not ensue immediately with successful pulmonary irrigation for an unknown period of time.
In addition to fat emboli, other typical findings of antemortem ship strike in cetaceans include subcutaneous, muscular and/or internal hemorrhage with hematoma formation, organ contusion and/or rupture with bleeding, e.g., in airways, in gastrointestinal tract, and edema in various organs mainly due to increased hydrostatic pressures, increased permeability due to hypoxia and vascular disruption (Campbell-Malone et al., 2008; Moore et al., 2013). Traumatic injuries on the dorsum are usually considered as antemortem or perimortem as carcasses tend to float with the ventral or lateral side upward, making a dorsal strike of a carcass unlikely (Laist et al., 2001; Campbell-Malone et al., 2008). On histopathologic examination, the presence of inflammatory response, hemorrhage or edema (Campbell-Malone et al., 2008; Moore et al., 2013), as well as acute, monophasic myocyte (segmentary, discoid) degeneration, contraction band necrosis, and/or fragmentation of the myofibers in the skeletal muscle (Sierra et al., 2014) support antemortem trauma.
Nevertheless, when working with decomposed carcasses it is not always feasible to assess many of the ship strike evidences described above. It is in these cases, where the detection of fat emboli in the lungs has proven to be a valuable and reliable confirmatory diagnostic tool, allowing us to conclude that, at least, 83% of the studied stranded sperm whales in the Canary Islands with evidence of ship strike were alive at the moment of the strike.
All the same, the methodology for fat emboli detection is not devoid of certain limitations that may lead to an under-estimation of fatal vessel strikes, such as the slanted and arbitrary sampling of lung tissue. Kinra and Kudesia (2004), suggested that fat emboli are not homogenously distributed along lung tissue. Although different lung areas, including cranial, medial, and caudal samples from both lungs should be routinely collected during the necropsy, there is not a specific sampling protocol to accurately assess lung fat emboli. Thus, results from small lung portions taken arbitrarily should be carefully interpreted as they may not be representative of the whole tissue. Future anatomical and topographical studies of pulmonary blood circulation and fat emboli distribution, respectively, are necessary to assess which lung areas should be sampled for an accurate fat emboli detection.
Microscopically, the severity of fat embolism has been traditionally assessed using a simple scale based on the number of emboli encountered in the tissue section studied (Saukko and Knight, 2004). Here we proposed and evaluated fat emboli area as a better estimator to assess fat emboli severity since the same number of emboli in two different lung histological sections may occupy different areas, and the one with the largest area occupied, would be a more severe case.
The rule developed for our samples based on the area occupied by fat emboli allowed us to discriminate between sperm whales that suffered strikes from those that died due to other causes, even when fat emboli was present in some animals of the control group (Cases 21 and 24). In these two cases, there was a blunt trauma of unknown origin. Possible etiologies included intra- or interspecific interactions or a potential live-stranding event (Díaz-Delgado et al., 2018). Intra-/interspecific traumatic interactions are frequent among cetaceans, and may result in blunt traumas where internal hemorrhages and/or bone fractures may occur (Arbelo et al., 2013; Díaz-Delgado et al., 2018). In the case of sperm whales, they have been observed being attacked by killer whales (Orcinus orca) or male sperm whales fighting with each other (Whitehead, 2009). These interactions have been observed to occasionally cause fat emboli (Díaz-Delgado et al., 2018).
When relating the severity of fat embolism and the severity of the trauma a general positive correlation was established by Emson (1958) based on the type and number of bones fractured of 100 patients who died after injury. On the contrary, we did not find any association between variables related to trauma (i.e., presence of fractures or location of the trauma) with fat embolism severity. This could be due to the low sample size, heterogeneous distribution of fat emboli, methodological bias, immediate cardiovascular collapse with none or little pulmonary irrigation post-trauma (sudden death), or to the fact that the strike occurred post-mortem. Fatal lesions, often involving direct cardiovascular trauma with or without rupture of large vessels (hypovolemic shock) and/or severe neurogenic dysfunction, may cause an abrupt death, leading to immediate cease of the blood, and could explain the absence of fat emboli in different body organs, including the lung. This has been documented in aircraft fatalities, were extensive injuries were associated with lower grades of fat embolism (Mason, 1962), or no emboli at all in disintegration cases (Kinra and Kudesia, 2004), showing the importance of intact circulation for the formation of fat emboli. We surmise rapid cardiovascular collapse could explain lack of detectable fat emboli in three of our cases (cases 7, 10, and 13). These animals presented severe injuries, including abdominal evisceration or caudal amputation (Díaz-Delgado et al., 2018).
Factors Related to Ship Strikes in the Canary Islands
All the sperm whales included in this study, were either female adults/subadults, or juveniles or calves, of any sex. This is in agreement with the fact that sperm whale females and their progeny composed the main groups present all year round in Canarian waters, which are considered nursery and breeding areas (André, 1997).
The fact that young animals are not fully adapted to dive and need to spend more time at surface, together with their relatively slow swimming speed compared to adults, may explain their higher vulnerability to ship strikes (Papastavrou et al., 1989; Laist et al., 2001; Whitehead, 2009). Mothers with recent calves (cases 6 and 7) may be also at higher risk as they will spend more time in the surface with their offspring.
A previous study estimated the absolute abundance and density of sperm whales in Canarian waters, and concluded that the species would not be able to sustain the current level of strikes (Fais et al., 2016). This impact is aggravated by the female philopatry in the Canaries, as they are not genetically connected to west North Atlantic populations (Alexander et al., 2016), and by the number of calves and reproductive females affected by strikes.
Most struck sperm whales appeared in the east coast of Tenerife, in agreement with previous studies (André, 1997; Carrillo and Ritter, 2010; Ritter, 2010). A major explanation for this is that the channel between Tenerife and Gran Canaria is a prime habitat for sperm whales in the Canaries (André, 1997; Fais et al., 2016), as well as an area with a high maritime traffic density, dominated by fast and high speed ferries (Ritter, 2010). An overlap between most of the sperm whales’ stranding locations and fast-ferry transects was also observed in the present study.
To conclude, this study provided compelling histochemical evidence of fat emboli as a reliable confirmatory diagnostic tool of ante-mortem ship-strike even in decomposed sperm whale carcasses. Our results demonstrated that most of the sperm whales with evidence of ship-strike and stranded in the Canaries were alive at the moment of the strike. However, this may be an underestimation as cases where immediate cardiovascular collapse (sudden death) occur, may lack detectable fat emboli in the lungs. A final diagnosis of antemortem ship-strike may considerably benefit from fat emboli detection in lung tissue, particularly when other trauma-related gross and microscopic findings are not evident.
Some future directions may include the study of fat emboli distribution within the lungs to determine if some areas are more affected by fat emboli, and based on the results, the development of a homogenized lung sampling protocol to detect fat emboli. Alternative techniques to osmium tetroxide, which is extremely toxic, should be developed to study lipids histologically. Lipid composition analyses of fat emboli may contribute to a better understanding of its pathogenesis in these animals.
Some mitigation measures have been implemented in other locations and have proven to be effective, such as a mandatory vessel-speed restriction in the United States East Coast (Conn and Silber, 2013), the establishment of Traffic Separation Schemes (TSS) in the Bay of Fundy (Vanderlaan et al., 2008) or the proposal of recommended Areas To Be Avoided (ATBA) like the Roseway Basin Area (Vanderlaan and Taggart, 2009). Similarly, mitigation measures to reduce ship-strike mortalities and guarantee the survival of the sperm whales’ population in Canarian waters should be further explored and implemented.
Data Availability
All datasets generated for this study are included in the manuscript and/or the supplementary files.
Ethics Statement
All animals included in the present study were dead, either floating offshore or stranded in the coast. Required permission for the management of stranded cetaceans was issued by the environmental department of the Canary Islands’ Government and the Spanish Ministry of Environment. No experiments were performed on live animals.
Author Contributions
AF: conceptualization. MArb, ES, YBdQ, JD-D, CS-S, RP-L, and MArr: sampling. MArr: laboratory analyses. PS, MArr, and YBdQ: data analyses. AF: funding. MArr: writing. All authors: review and editing. YBdQ and AF: supervision.
Funding
This study was funded by the National Project CGL2015/71498P and the Canary Islands Government, which has funded and provided support to the stranding network. MArr was funded by the University Professor Formation fellowship from the Spanish Ministry of Education (FPU; 15/02265). YBdQ was funded by a postdoctoral fellowship from the University of Las Palmas de Gran Canaria. JD-D was the recipient of a postdoctoral fellowship by the São Paulo Research Foundation (FAPESP; Grant #2017/02223-8).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank the Electronic Microscopy Service in the Faculty of Health Sciences (University of Las Palmas de Gran Canaria), all the members and volunteers of the Cetacean Stranding Network, Marisa Tejedor and associated nongovernmental organizations: SECAC and Canary Conservation. The authors also thank Ana María Alfonso for technical assistance. MArb is also a co-corresponding author.
References
Abramowsky, C. R., Pickett, J. P., Goodfellow, B. C., and Bradford, W. D. (1981). Comparative demonstration of pulmonary fat emboli by “en bloc” osmium tetroxide and oil red O methods. Hum. Pathol. 12, 753–755. doi: 10.1016/s0046-8177(81)80179-7
Aguilar, N., Carillo, M., Delgado, I., Díaz, F., and Brito, A. (2000). “Fast ferries impact on cetaceans in the canary islands: strikes and displacement,” in Proceedings of the. 14th Annaul Conference, (St Louis: ECS, Cork), 164.
Alexander, A., Steel, D., Hoekzema, K., Mesnick, S. L., Engelhaupt, D., Kerr, I., et al. (2016). What influences the worldwide genetic structure of sperm whales (Physeter macrocephalus)? Mol. Ecol. 25, 2754–2772. doi: 10.1111/mec.13638
André, M. (1997). Distribución y Conservación Del Cachalote (Physeter macrocephalus) en las Islas Canarias. Ph.D. Thesis University of Las Palmas de Gran Canaria: Tafira.
Arbelo, M., Espinosa de los Monteros, A., Herráez, P., Andrada, M., Sierra, E., Rodríguez, F., et al. (2013). Pathology and causes of death of stranded cetaceans in the canary islands (1999-2005). Dis. Aquat. Org. 103, 87–99. doi: 10.3354/dao02558
Armstrong, J. A., Fryer, D. I., Stewart, W. K., and Whittingham, H. E. (1955). Interpretation of injuries in the comet aircraft disasters: an experimental approach. Lancet 265, 1135–1143.
Breiman, L., Friedman, J., Olshen, R. A., and Stone, C. J. (1984). Classification and Regression Trees. Belmont: Wadsworth International Group.
Campbell-Malone, R., Barco, S. G., Daoust, P.-Y., Knowlton, A. R., McLellan, W. A., Rotstein, D. S., et al. (2008). Gross and histologic evidence of sharp and blunt trauma in north atlantic right whales (Eubalaena glacialis) killed by vessels. J. Zoo Wildl. Med. 39, 37–55. doi: 10.1638/2006-0057.1
Carrillo, M., and Ritter, F. (2010). Increasing numbers of ship strikes in the canary Islands: proposals for immediate action to reduce risk of vessel-whale collisions. J. Cetacean Res. Manag. 11, 131–138.
Conn, P. B., and Silber, G. K. (2013). Vessel speed restrictions reduce risk of collision-related mortality for North Atlantic right whales. Ecosphere 4, 1–16. doi: 10.1890/ES13-00004.1
de Stephanis, R., and Urquiola, E. (2006). “Collisions between ships and cetaceans in Spain,” in Proceedings of the International Whaling Commission Scientific Commitee SC/58/BC5, (Cambridge).
Díaz-Delgado, J., Fernández, A., Sierra, E., Sacchini, S., Andrada, M., Vela, A. I., et al. (2018). Pathologic findings and causes of death of stranded cetaceans in the Canary Islands (2006-2012). PLoS One 13:e0204444. doi: 10.1371/journal.pone.0204444
Emson, H. E. (1958). Fat embolism studied in 100 patients dying after injury. J. Clin. Pathol. 11, 28–35. doi: 10.1136/jcp.11.1.28
Fais, A., Lewis, T. P., Zitterbart, D. P., Álvarez, O., Tejedor, A., and Aguilar Soto, N. (2016). Abundance and distribution of sperm whales in the Canary Islands: can sperm whales in the archipelago sustain the current level of ship-strike mortalities? PLoS One 11:e0150660. doi: 10.1371/journal.pone.0150660
Frantzis, A., Leaper, R., Alexiadou, P., Prospathopoulos, A., and Lekkas, D. (2019). Shipping routes through core habitat of endangered sperm whales along the hellenic trench, greece: can we reduce collision risks? PLoS One 14:e0212016. doi: 10.1371/journal.pone.0212016
Geraci, J. R., and Lounsbury, V. J. (2005). Marine Mammals Ashore: A Field Guide for Strandings. Baltimore, MD: National Aquarium in Baltimore.
Gupta, A., and Reilly, C. S. (2007). Fat embolism. Contin. Educ. Anesth. Crit. Care Pain 7, 148–151. doi: 10.1093/bjaceaccp/mkm027
Hulman, G. (1995). The pathogenesis of fat embolism. J. Pathol. 176, 3–9. doi: 10.1002/path.1711760103
Joblon, M. J., Pokras, M. A., Morse, B., Harry, C. T., Rose, K. S., Sharp, S. M., et al. (2014). Body condition scoring system for delphinids based on short-beaked common dolphins (Delphinus delphis). J. Mar. Anim. Their Ecol. 7, 5–13.
Kinra, S. L. P., and Kudesia, W. C. S. (2004). Pulmonary fat embolism in IAF aircraft accident fatalities: a 33 year retrospective study. Indian J. Aerosp. Med. 48, 57–62.
Kuiken, T., and García Hartmann, M. (1991). “Dissection techniques and tissue sampling,” in Proceedings of the 1st ECS Workshop on Cetacean Pathology, (Saskatoon, SK).
Laist, D. W., Knowlton, A. R., Mead, J. G., Collet, A. S., and Podesta, M. (2001). Collisions between ships ad whales. Mar. Mammal Sci. 17, 35–75. doi: 10.1111/j.1748-7692.2001.tb00980.x
Mason, J. K. (1968). Pulmonary fat and bone marrow embolism as an indication of ante-mortem violence. Med. Sci. Law 8, 200–206. doi: 10.1177/002580246800800311
Mason, J. K. (ed.) (1962). “Timing of injuries,” in Aviation Accident Pathology: A Study of Fatalities, (London: Butterworths), 239–268.
Miller, P. J. O., Aoki, K., Rendell, L. E., and Amano, M. (2008). Stereotypical resting behavior of the sperm whale. Curr. Biol. 18, R21–R23. doi: 10.1016/j.cub.2007.11.003
Moore, M. J., Van Der Hoop, J., Barco, S. G., Costidis, A. M., Gulland, F. M., Jepson, P. D., et al. (2013). Criteria and case definitions for serious injury and death of pinnipeds and cetaceans caused by anthropogenic trauma. Dis. Aquat. Org. 103, 229–264. doi: 10.3354/dao02566
Panigada, S., Pesante, G., Zanardelli, M., Capoulade, F., Gannier, A., and Weinrich, M. T. (2006). Mediterranean fin whales at risk from fatal ship strikes. Mar. Pollut. Bull. 52, 1287–1298. doi: 10.1016/j.marpolbul.2006.03.014
Papastavrou, V., Smith, S. C., and Whitehead’, H. (1989). Diving behaviour of the sperm whale, Physeter macrocephalus, off the galapagos islands. Can. J. Zool. 67, 839–846. doi: 10.1139/z89-124
Perrin, W. F., Würsig, B., and Thewissen, J. G. M. (eds) (2009). Encyclopedia of Marine Mammals. Cambridge, MA: Academic Press.
R Development Core Team (2016). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available at: https://www.rproject.org/Google Scholar
Ritter, F. (2010). A quantification of ferry traffic in the Canary Islands (Spain) and its significance for collisions with cetaceans. J. Cetacean Res. Manag. 11, 139–146.
Saukko, P., and Knight, B. (2004). “Complications of injury,” in Knight′s Forensic Pathology, eds P. Saukko and B. Knight (Boca Raton: CRC Press), 343–345.
Sierra, E., Fernández, A., Espinosa De Los Monteros, A., Arbelo, M., Díaz-Delgado, J., Andrada, M., et al. (2014). Histopathological muscle findings may be essential for a definitive diagnosis of suspected sharp trauma associated with ship strikes in stranded cetaceans. PLoS One 9:e88780. doi: 10.1371/journal.pone.0088780
Tanner, B., Jones, J., Cople, S., and Davison, P. (1990). Fat embolization in trauma facilities. Annu. Emerg. Med. 19:57.
Taylor, B. L., Baird, R., Barlow, J., Dawson, S. M., Ford, J., Mead, J. G., et al. (2008). Physeter macrocephalus. IUCN Red List of Threat. Species 2008:e.T41755A10554884. doi: 10.2305/IUCN.UK.2008.RLTS.T41755A10554884.en
Tejedor, M., and Martín, V. (2013). “Colección de referencia de cetáceos de las Islas canarias: una herramienta para la investigación y conservación en la macaronesia,” in Poster at the VI Congreso de la Sociedad Española de Cetáceos, Tarifa.
Van Waerebeek, K., and Leaper, R. (2008). “Second report of the IWC vessel strike data standardisation working group,” in proceeding of the 60th IWC Scientific Committee Annual Meeting, (Santiago), doi: 10.13140/RG.2.1.2020.2001
Vanderlaan, A. S. M., and Taggart, C. T. (2007). Vessel collisions with whales: the probability of lethal injury based on vessel speed. Mar. Mamm. Sci. 23, 144–156. doi: 10.1111/j.1748-7692.2006.00098.x
Vanderlaan, A. S. M., and Taggart, C. T. (2009). Efficacy of a voluntary area to be avoided to reduce risk of lethal vessel strikes to endangered whales. Conserv. Biol. 23, 1467–1474. doi: 10.1111/j.1523-1739.2009.01329.x
Vanderlaan, A. S. M., Taggart, C. T., Serdynska, A. R., Kenney, R. D., and Brown, M. W. (2008). Reducing the risk of lethal encounters: vessels and right whales in the Bay of Fundy and on the Scotian Shelf. Endanger. Species Res. 4, 283–297. doi: 10.3354/esr00083
Watkins, W. A., Daher, M. A., DiMarzio, N. A., Samuels, A., Wartzok, D., Fristrup, K. M., et al. (1999). Sperm whale surface activity from tracking by radio and satellite tags. Mar. Mamm. Sci. 15, 1158–1180. doi: 10.1111/j.1748-7692.1999.tb00883.x
Watson, A. J. (1970). Genesis of fat emboli. J. Clin. Pathol. 23, 132–142. doi: 10.1136/jcp.s3-4.1.132
Watwood, S. L., Miller, P. J. O., Johnson, M., Madsen, P. T., and Tyack, P. L. (2006). Deep-diving foraging behaviour of sperm whales (Physeter macrocephalus). J. Anim. Ecol. 75, 814–825. doi: 10.1111/j.1365-2656.2006.01101.x
Whitehead, H. (2009). “Sperm whale,” in Encyclopedia of Marine Mammals, eds W. F. Perrin, B. Würsig, and H. Thewissen (San Diego, CA: Academic Press), 1091–1097.
Keywords: sperm whale, antemortem, fat embolism, Canary Islands, ship strike
Citation: Arregui M, Bernaldo de Quirós Y, Saavedra P, Sierra E, Suárez-Santana CM, Arbelo M, Díaz-Delgado J, Puig-Lozano R, Andrada M and Fernández A (2019) Fat Embolism and Sperm Whale Ship Strikes. Front. Mar. Sci. 6:379. doi: 10.3389/fmars.2019.00379
Received: 17 March 2019; Accepted: 18 June 2019;
Published: 03 July 2019.
Edited by:
Jessica Redfern, Southwest Fisheries Science Center (NOAA), United StatesReviewed by:
Frances Gulland, University of California, Davis, United StatesAmy Richardson Knowlton, New England Aquarium, United States
Copyright © 2019 Arregui, Bernaldo de Quirós, Saavedra, Sierra, Suárez-Santana, Arbelo, Díaz-Delgado, Puig-Lozano, Andrada and Fernández. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yara Bernaldo de Quirós, eWFyYS5iZXJuYWxkb0B1bHBnYy5lcw==