 Heather Heenehan1
Heather Heenehan1 Joy E. Stanistreet2
Joy E. Stanistreet2 Peter J. Corkeron3
Peter J. Corkeron3 Laurent Bouveret4Julien Chalifour5Genevieve E. Davis1,6
Laurent Bouveret4Julien Chalifour5Genevieve E. Davis1,6 Angiolina Henriquez7
Angiolina Henriquez7 Jeremy J. Kiszka8
Jeremy J. Kiszka8 Logan Kline1
Logan Kline1 Caroline Reed9Omar Shamir-Reynoso10Fabien Védie11Wijnand De Wolf12
Caroline Reed9Omar Shamir-Reynoso10Fabien Védie11Wijnand De Wolf12 Paul Hoetjes13
Paul Hoetjes13 Sofie M. Van Parijs3*
Sofie M. Van Parijs3*- 1Integrated Statistics, Northeast Fisheries Science Center, National Marine Fisheries Service, National Oceanic and Atmospheric Administration, Woods Hole, MA, United States
- 2Bedford Institute of Oceanography, Fisheries and Oceans Canada, Dartmouth, NS, Canada
- 3Northeast Fisheries Science Center, National Marine Fisheries Service, National Oceanic and Atmospheric Administration, Woods Hole, MA, United States
- 4Observatoire des Mammifères Marins de l’Archipel Guadeloupéen, Port-Louis, Guadeloupe
- 5Association de Gestion de la Réserve Naturelle de Saint Martin, Anse Marcel, Saint Martin
- 6Biology Department, University of Massachusetts Boston, Boston, MA, United States
- 7Aruba Marine Mammal Foundation, Savaneta, Aruba
- 8Department of Biological Science, Florida International University, North Miami, FL, United States
- 9Ossining High School, Ossining, NY, United States
- 10National Authority for Marine Affairs, Santo Domingo, Dominican Republic
- 11DEAL Martinique, Schœlcher, Martinique
- 12STINAPA Bonaire, Bonaire, Sint Eustatius and Saba
- 13Nature and Food Quality Unit, Ministry of Agriculture, Bonaire, Sint Eustatius and Saba
Assessing marine soundscapes provides an understanding of the biological, geological and anthropogenic composition of a habitat, including species diversity, community composition, and human impacts. For this study, nine acoustic recorders were deployed between December 2016 and June 2017 off six Caribbean islands in several Marine Parks: the Dominican Republic (DR), St. Martin (SM), Guadeloupe east and west (GE, GW), Martinique (MA), Aruba (AR), and Bonaire (BO). Humpback whale song was recorded at five sites on four islands (DR, SM, GE, GW, and MA) and occurred on 49–93% of recording days. Song appeared first at the DR site and began 4–6 weeks later at GE, GW, and MA. No song was heard in AR and BO, the southernmost islands. A 2-week period was examined for the hourly presence of vessel noise and the number and duration of ship passages. Hourly vessel presence ranged from low (20% – DR, 30% – SM), medium (52% – MA, 54% – BO, 77% – GE) to near continuous (99% – GW; 100% – AR). Diurnal patterns were observed at BO, GE, and MA with few to no vessels present during night time hours, possibly reflecting the activity of recreational craft and fishing vessels. At the DR and GW sites, vessel traffic was ubiquitous for most of the day, likely reflecting heavy cruise ship and container ship presence. Soundscapes were diverse across islands with persistent fish choruses, sporadic sperm whale (Physeter macrocephalus) and dolphin (Delphinidae) presence at BO, minke whales (Balaenoptera acutorostrata) from late December to late February at MA and an earthquake recorded across all sites. These analyses provide an important first step in characterizing the health and species richness in Caribbean marine parks and demonstrate a surprising high anthropogenic foot print. Vessel traffic in particular contributes adversely to marine soundscapes, masking marine mammal sounds, potentially changing typical animal behavior and raising the risk of ship strike.
Introduction
Sound has low attenuation and travels effectively in sea water, moving approximately five times faster than it does in air. Where vision fails (at night, at depths where sunlight does not penetrate, and in turbid water), sound production and hearing function as efficient communication and sensory mechanisms. Since sound is efficient in light-limited habitats, marine animals have evolved to rely heavily on their use of sound for communication, foraging, and navigation (e.g., Benoit-Bird and Au, 2009; Radford et al., 2011; Janik and Sayigh, 2013, respectively). Passive acoustic monitoring (PAM) lets us exploit this key sensory modality and overcome some of the challenges of traditional visual surveys to learn about marine animals and their environment simply by listening. When we listen, we have the opportunity to determine species presence (e.g., Heenehan et al., 2016), distribution (e.g., Davis et al., 2017), migration (e.g., Risch et al., 2014), and abundance (e.g., Van Parijs et al., 2002). We can also characterize the acoustic environment or soundscape of an area. A soundscape is considered to be all sounds present in a given place over a certain period of time (Krause and Gage, 2003; Pijanowski et al., 2011).
The types of sounds that comprise a soundscape vary between sites and vary on multiple timescales (for examples see McWilliam and Hawkins, 2013; Erbe et al., 2015; Haver et al., 2017; Heenehan et al., 2017) but usually include a combination of sounds from three broad categories: (1) sounds from geological or physical processes (e.g., from earthquakes, wind, and rain), sometimes referred to as the geophony; (2) sounds from non-human living things (e.g., sounds from marine mammals and fish), sometimes referred to as the biophony; (3) the sounds produced by humans (e.g., vessel noise, seismic surveying, and sonar), sometimes referred to as the anthrophony (Krause and Gage, 2003; Pijanowski et al., 2011). Identifying the different components that comprise a soundscape can provide insights into the composition of a given marine environment. This soundscape characterization can prove a useful way for monitoring long term changes within a given environment, such as increases or decreases in species composition and/or anthropogenic noise impacts that may affect the ecological diversity and health of a given site (e.g., Haver et al., 2017).
Sounds within these three soundscape categories overlap in time, space, and frequency. Therefore, acoustic recordings capture multiple sound sources within and across these categories (Van Opzeeland and Boebel, 2018). Although the specific components of a soundscape preserved in a recording depends on many factors including the location of the recorder, the location of the sound source(s), and the sampling rate, recordings may be used to characterize spectral and temporal overlap between sounds and explore potential masking (Van Opzeeland and Boebel, 2018). Masking occurs when a sound not only overlaps with but actually affects or interferes with the ability to receive another sound (American National Standards Institute, 1994; Clark et al., 2009; Erbe et al., 2016). For example, a well-studied masking relationship is the one between shipping noise and baleen whale calls in Stellwagen Bank National Marine Sanctuary off the coast of Massachusetts, United States. The temporal, spatial, and spectral overlap of these sounds has resulted in high levels of masking (Cholewiak et al., 2018) and a large loss of communication space for these animals (Hatch et al., 2012, 2016).
In recent years, the importance of managing acoustic habitats, such as marine parks, sanctuaries or areas of biological importance, in order to minimize anthropogenic impacts has become increasingly recognized (e.g., Hatch and Fristrup, 2009; Hatch et al., 2012; Williams et al., 2014, 2015; Merchant et al., 2015; McKenna et al., 2017). Several designated marine parks exist within Caribbean waters, established primarily for the protection habitat such as coral reefs, mangroves and sea grass, as well as marine protected species or species of local importance, such as humpback whales, Megaptera novaeangliae (Knowles et al., 2015; di Sciara et al., 2016).
Marine shipping, particularly large ocean container ships, hydrocarbon transport, and cruise ships, is a recognized and persistent anthropogenic source of low-frequency ocean noise, contributing to the masking of essential sounds produced and heard by marine animals and fish (e.g., Weilgart, 2007; Hatch et al., 2008; Hildebrand, 2009; Erbe et al., 2012; McKenna et al., 2012; Merchant et al., 2014; Williams et al., 2015). The Caribbean region is largely made up of small and specialized open island economies, which import a large proportion of their consumer goods. In addition, any local production of goods and services is heavily dependent on the import of raw materials and unfinished parts. In 1996, the Caribbean region’s level of foreign trade, as a proportion of GDP, was 78%, compared with 25% for Latin America for the same period (Hoffmann, 1997). Imports arrive either by air or by sea, resulting in heavy marine traffic throughout the Greater Caribbean Sea. As a result ship-generated noise presents a significant threat to the regions’ marine ecosystems and their underwater soundscapes.
For this study acoustic recordings were collected across seven sites throughout the Caribbean, of which six sites were situated in marine parks. The primary aim was to identify the main soundscape contributors in order to establish an acoustic baseline and understand levels of anthropogenic noise and evaluate potential impacts on marine mammals both within and between sites. As humpback whales are an identified species of importance across most marine parks, we used the extensive research on humpback whale utilization of the Caribbean breeding grounds to provide a basis for determining the locations of our study sites (Kennedy, 2018). Male humpback whales sing on breeding grounds (Herman, 2017), and their song is loud and persistent (e.g., Payne and McVay, 1971; Vu et al., 2012), making it an excellent acoustic indicator of these whales’ occurrence in an area. The extent to which vessel noise overlaps with humpback whales’ communication space can provide insight into potential masking and interference of noise with breeding behavior. Besides impacting whales’ communication space, vessel noise can also affect the stress hormone levels of whales (Rolland et al., 2012), which is of conservation concern, especially in an area where whales are calving (e.g., Bejder et al., 2019)1.
A secondary focus of this study was on the timing of arrival and departure of humpback whales at different sites in the Caribbean. Several thousand humpback whales migrate from feeding grounds in the northern North Atlantic to breed in the Caribbean in winter and spring (Stevick et al., 1998; Bettridge et al., 2015). The only other known breeding ground for these whales in the North Atlantic is in the waters of the Cape Verdes Islands, off West Africa (Ryan et al., 2014), used by less than 300 individuals. It appears that humpback whales that migrate to the southeastern Caribbean (Stevick et al., 2018) mostly do so late in the breeding season (mid-March to May), and are distinct from those occurring earlier (January – early March, Stevick et al., 2018). Some individual whales have been photographically identified in separate years off the Cape Verdes islands and in the southeastern Caribbean (Stevick et al., 2016). Further, it appears that the feeding grounds of the humpbacks occurring later in Caribbean waters are off Europe and Scandinavia, rather than in the western North Atlantic (Stevick et al., 2018). The abundance of this later-migrating population of humpback whales is likely much smaller than the earlier migrating whales, and so is of greater conservation concern, hence our interest in their overlap with vessel noise.
Materials and Methods
Nine passive acoustic recorders were deployed between December 2016 and January 2017, and recovered between May and June 2017 at seven recording sites in waters throughout the Caribbean island chain from Dominican Republic (DR), St. Martin (SM), Guadeloupe east and west (GE and GW) and Martinique (MA), to Bonaire (BO) and Aruba (AR) (Figure 1 and Table 1). All sites, with the exception of AR, were located within marine park waters, with four sites on the ocean side of the islands and three on the leeward side within the Caribbean Sea. The DR site was located within the Silver and Navidad Bank Sanctuary (Mattila et al., 1989). The GW, GE, and MA sites were all located within the Agoa Sanctuary, with the GW site also located within the National Park of Guadeloupe. The SM site was located within the Natural Reserve of Saint Martin, and the BO site was located within the Stichting Nationale Parken (STINAPA) Bonaire National Marine Park. Two types of autonomous, bottom-mounted recording devices were used, Marine Autonomous Recording Units (MARUs; Cornell University, Ithaca, NY, United States) and SoundTraps (Ocean Instruments, Auckland, New Zealand2). MARUs were deployed at six locations (DR, GE, GW, MA, BO, and AR; Figure 1 and Table 1) and programmed to collect continuous recordings at a sampling rate of 2 kHz, suitable for recording low-frequency sounds including baleen whale calls and vessel noise. SoundTraps were deployed at three locations (SM, GE, and BO; Figure 1 and Table 1), and programmed to record at a sampling rate of 48 kHz, allowing a broader characterization of the soundscape and the detection of higher-frequency sounds, such as dolphin whistles and sperm whale echolocation clicks. In order to obtain recordings across the full deployment period at this higher sampling rate, the SoundTrap recordings were collected using a duty-cycled recording schedule of 1 h of recording every 4 h for two sites (SM and BO). A less frequent recording schedule of 1 h every 4 days occurred for the SoundTrap at GE, due to a programming error (Table 1). Sites were chosen based on a variety of factors including past observations of humpback whales, marine park waters, oceanographic conditions, and human activities. All units were anchored at depths ranging from 16 to 59 m with the hydrophone suspended or mounted 1–2 m above the sea floor (Table 1). The MARUs were anchored with metal weights and contain a built-in acoustic release mechanism for remote release (Calupca et al., 2000), thus were used at deeper sites such as DR and MA. SoundTraps were strapped to a cement or metal-based anchor, with deployments and retrievals assisted by divers, and thus were used at shallower sites such as GE and BO.
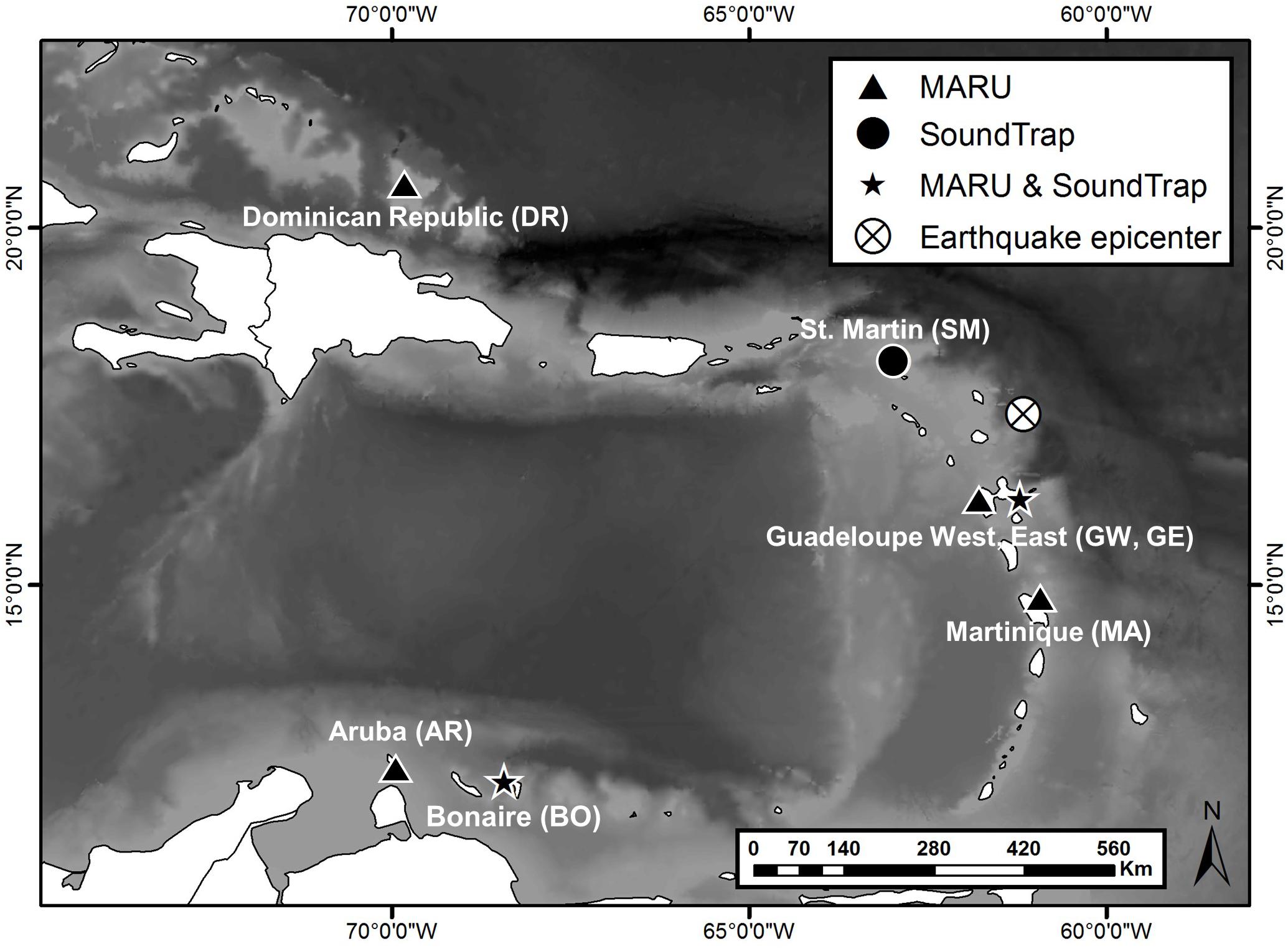
Figure 1. A map of the seven passive acoustic recording sites and equipment deployed in the Caribbean, December 2016 through June 2017. The recorder type, Marine Acoustic Recording unit (MARU) or SoundTrap acoustic recorder are marked as well. X marks the epicenter for a large earthquake that occurred on 17 April 2017 at 17.450 N 61.142 W.
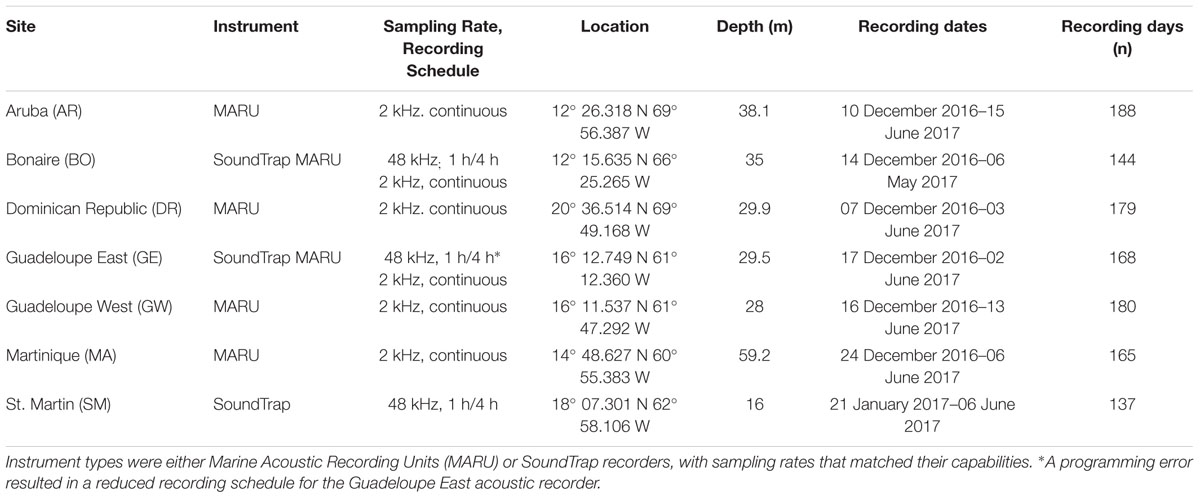
Table 1. Summary of passive acoustic recording effort across seven sites in the Caribbean Sea from December 2016 to June 2017.
All recordings were visually reviewed by an acoustic analyst for the daily presence of marine mammals and vessel noise using the sound analysis software Raven Pro 1.5 and 2.0 (Bioacoustics Research Program, Cornell University, Ithaca, NY, United States). For low-frequency analyses of humpback whale song, minke whale pulse trains, vessel noise and an earthquake event, spectrograms were viewed across a frequency range of 0–1000 Hz and a time window of 3 min.
Low Frequency Analyses – Baleen Whales
In order to identify low frequency sounds across all sites, the MARU data was preferentially used, rather than the SoundTrap data, since these data consisted of continuous, rather than duty cycled, recordings. This was possible for all sites except for SM, where no MARU was deployed. SoundTrap recordings for SM were decimated to a sampling rate of 2 kHz to match the sampling rate of the MARU recordings and standardize the analyses across recorder types. For each day, daily presence of humpback whale song was marked at the first observed occurrence of song, and absence was marked if no song was found in the entire day. During the humpback whale analysis, minke whale (Balaenoptera acutorostrata) pulse trains (Risch et al., 2013) were observed in the MA recordings and an identical analysis for daily presence of these pulses was further conducted at that site. Each clear pulse train detected was then categorized by pulse train type. These different types of minke whale pulse trains included speed-up trains, in which the inter-pulse-interval decreases across the duration of the pulse train; slow-down trains, in which the inter-pulse interval increases across the duration of the pulse train; and constant trains with no clear change in inter-pulse-interval as described in Risch et al. (2013).
Low Frequency Analyses – Vessel Noise
A detailed analysis of the presence of vessel noise was carried out over a 2 weeks period between 12 March 2017 and 25 March 2017 across all seven sites. This period was selected to correspond with a period in which humpback whale song was consistently present across five recording sites. Each recording hour was marked as containing either vessel noise, humpback whale song, both or neither sound. The percentage of hours with vessel noise and humpback whale song is presented for each site. Next, selection boxes were made around all discernible vessel noise events and the duration of each vessel passages was calculated. For the sites where it was possible to identify discrete vessel passages, the number of vessel passages per hour were compared with night and day times in order to look for diel patterns in vessel activity.
High Frequency Analyses – Odontocetes
Long-term spectral averages (LTSAs) were created for all high-frequency data using the Triton software (Scripps Whale Acoustic Lab, Scripps Institution of Oceanography, La Jolla, CA, United States) developed in MATLAB (The Mathworks, Inc., Natick, MA, United States). LTSAs provide a compressed spectrogram view with a 5 s time resolution and 100 Hz frequency resolution, and facilitate efficient visual review of large datasets. The prevalence of snapping shrimp across all sites in the high-frequency recordings complicated this analysis and tended to mask other sounds in the LTSAs. Odontocete sounds such as clicks, echolocation click trains, whistles and burst pulses were clearly visible and distinguishable from the snapping shrimp in the background as their frequency range, click patterns and sound levels were distinctive. The daily presence of dolphin (Delphinidae) whistles and sperm whale (Physeter macrocephalus) clicks was analyzed at sites where they were present.
Acoustic Analysis of an Earthquake
The acoustic presence of an earthquake was discovered on the acoustic recordings and examined across all seven sites. The distance from the epicenter of the earthquake to each recording site where it was heard was measured using ArcGIS 10.3.13. The duration, frequency, and timing of the earthquake and the number of aftershocks were measured using Raven Pro 1.5. Earthquake acoustic signatures were analyzed using a time window of 10 min and only events occurring between the hours of 00:45 and 03:00 UTC-4 were included.
Results
Recorders were deployed between 7th December 2016 and 13th June 2017, in waters between 16 and 59 m depth, for 137 – 188 days in total. Details of deployments at each individual site are given in Table 1.
Low Frequency Analyses – Baleen Whales
Humpback whale song was present across five of the seven recording sites (DR, SM, GE, GW, and MA; Figure 2). Song was not present in the recordings from the AR or BO sites.
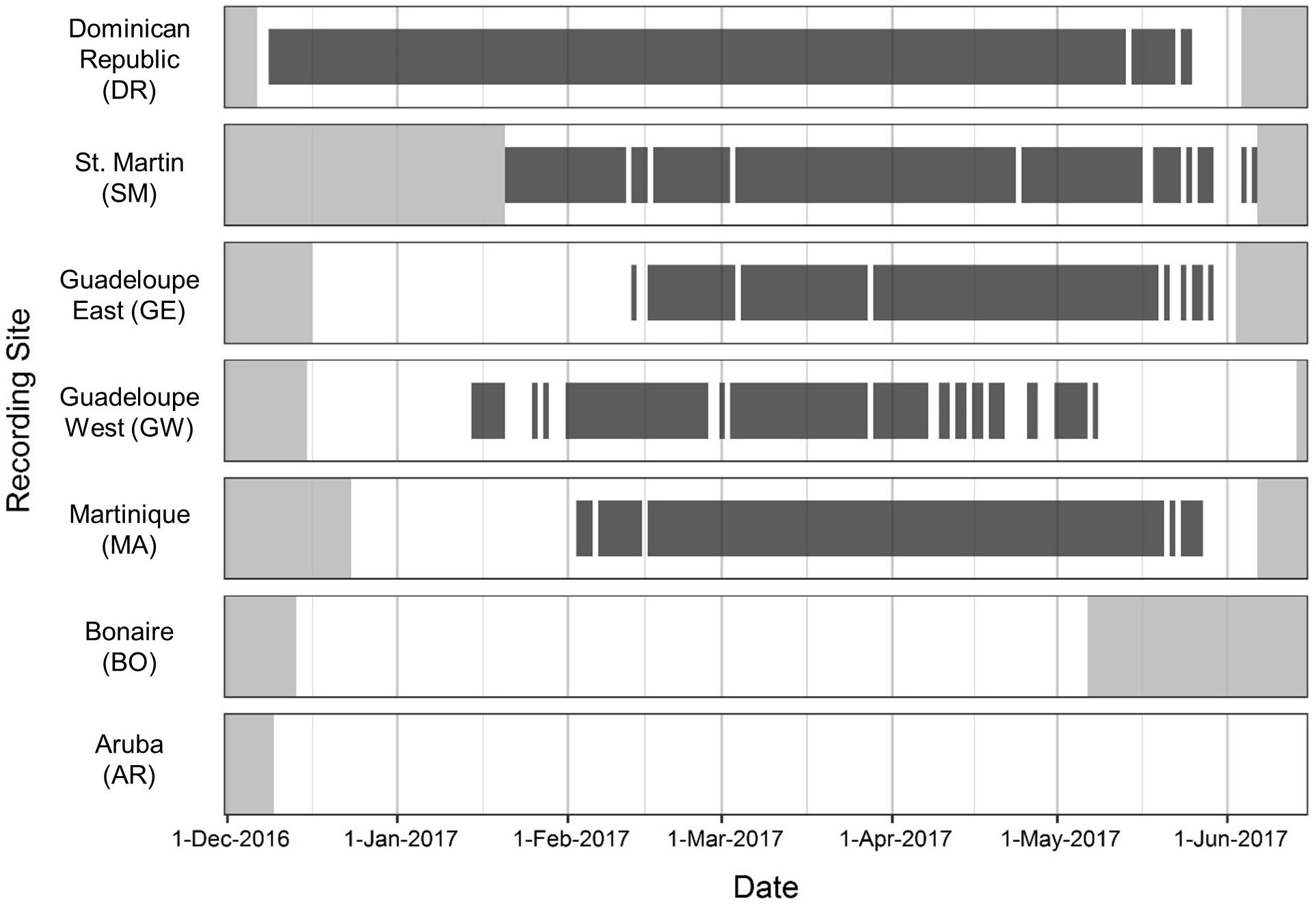
Figure 2. Seasonal occurrence of humpback whale song from December 2016 to Jun 2017 across all recording sites, shown from north (top) to south (bottom). Dark gray bars indicate recording dates with humpback song present; light gray shading indicates periods with no recording effort. Recordings were collected continuously at all sites except for St. Martin, where a 25% duty-cycle (1 h/4 h) was used.
There was variation among the sites in the seasonal timing (beginning and cessation) of song presence as well as the percentage of days with song. Humpback whale song first began on 09 December 2016 at the DR site, our northernmost Caribbean recording site. Song was then present at the DR every day until 13 May 2017, then continued more sporadically until 25 May 2017 (Figure 2). Singing activity was often intense, with multiple singers recorded simultaneously. Song was present in the DR for a total of 166 (93%) of the 179 recording days.
Song was recorded next in GW, beginning on 15 January 2017 and occurring intermittently through 05 May 2017. Humpback whale song was present on 88 (49%) of the 180 recording days. Song was detected at GW a full month earlier than EG, and was present more intermittently at GW than at the other sites. At SM, song was recorded throughout the deployment period including the first and last recording day from 21 January 2017 through 06 June 2017 (Figure 2). Humpback whale song was present on 123 (90%) of the 137 (duty-cycled) recording days and occurred sporadically in June, which was the latest occurrence of song recorded anywhere in the study. However, it is highly probable that song may have started earlier and ended later at the SM site as the SoundTrap recorder could not be deployed sooner due to poor weather conditions and the recordings ended prior to the end of song presence. Song started latest in MA and EG, starting on the 03 and 13 of February 2017 respectively, and ending 27 May 2017 in MA and 29 May 2017 in EG. For MA, humpback whale song was present in 110 (67%) of the 165 recording days, and for EG, on 97 (58%) of the 168 recording days.
Minke whale pulse trains were detected almost every day throughout the MA recordings from 24 December 2016 to 20 February 2017, and on 3 days between 12 and 19 March 2017 (Figure 3A). Three different types of pulse trains produced by minke whales (Balaenoptera acutorostrata) were seen over the 56 days when minke whales were recorded. Of the 3,265 pulse trains, 71% were speed-up trains, reflecting similar vocal trends reported by Risch et al. (2014) in the broader Atlantic, who also reported a preponderance of speed-up trains. The pulse trains consisted of 25% slow-down trains, and 4% constant trains.
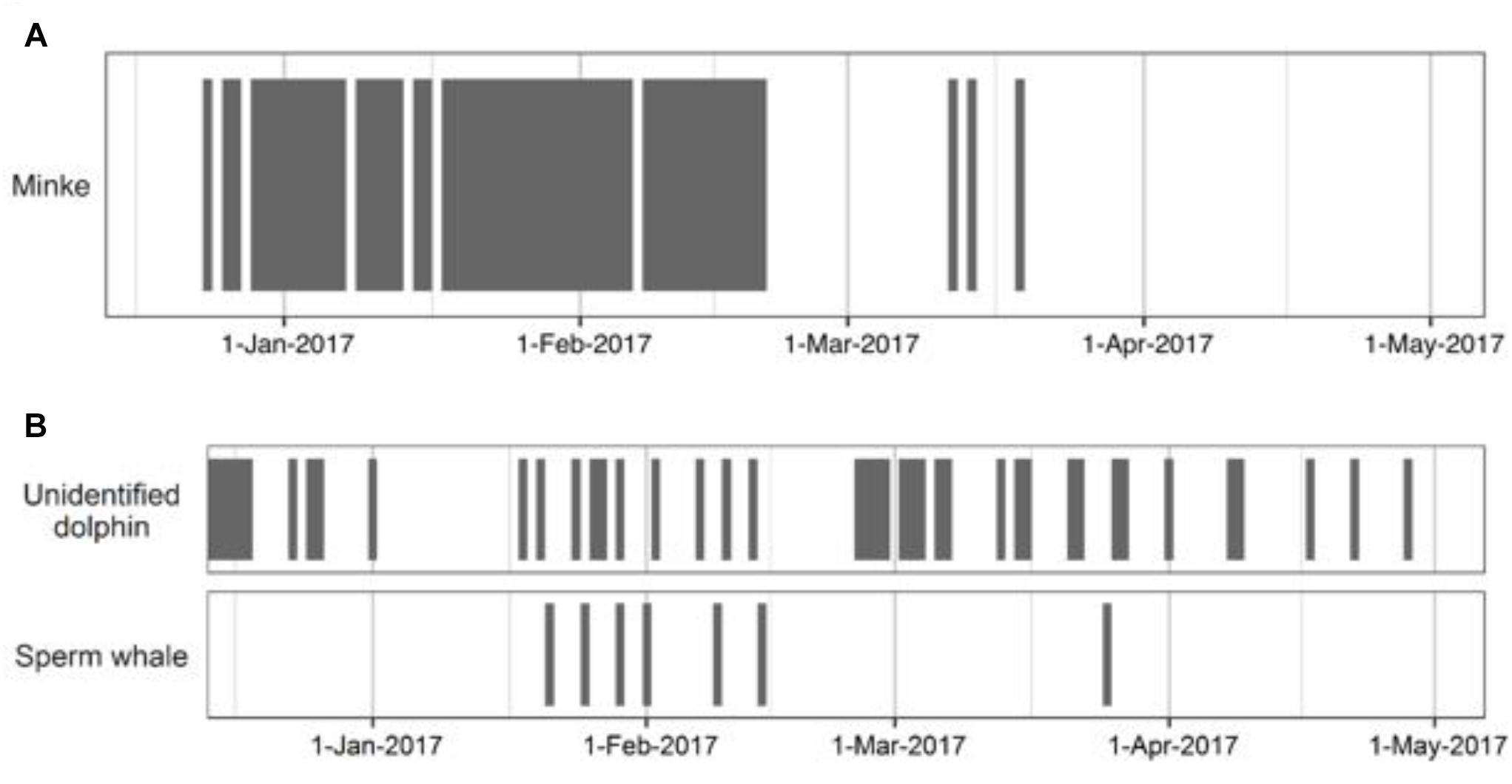
Figure 3. Daily presence of (A) minke whale pulse trains in Martinique and (B) unidentified dolphin whistles and sperm whale clicks and social sounds in Bonaire.
Low Frequency Analyses – Vessel Noise
A total of 336 h of data were analyzed for this 2 weeks period. The two sites that were most dominated by vessel noise during the 2 weeks detailed analysis were the AR and GW sites with vessel noise present on 100 and 99% of hours at these sites, respectively. These were followed by GE (77%), BO (54%), and MA (52%) with the lowest hourly presence of vessel noise at the SM (30%) and DR (20%) sites (Table 2 and Figure 4). The AR recording site was the only site not located in a marine park, sanctuary or protected area and was in a very heavy vessel traffic zone near a port with constant vessel passage and no discernible humpback whale song (Supplementary Figure S1). It was not possible to calculate the number or duration of vessel passages at this site, since vessel presence was so constant that it was difficult to distinguish when one vessel passage started and another one ended.
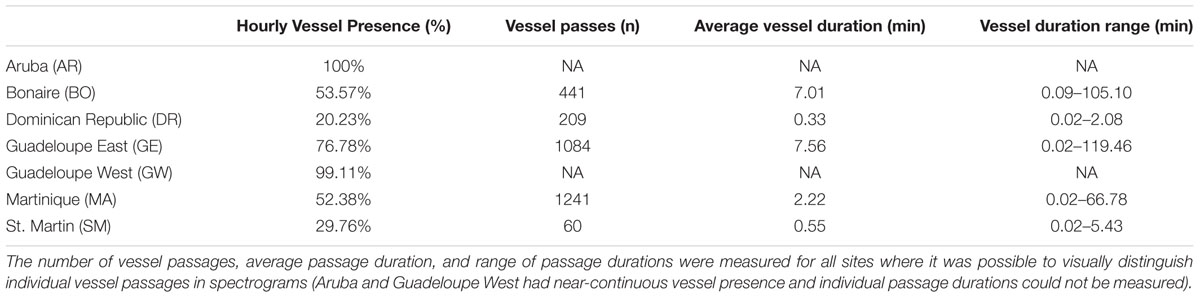
Table 2. A summary of the hourly vessel presence at each recording site over a 2-week period between 12 March 2017 and 25 March 2017.
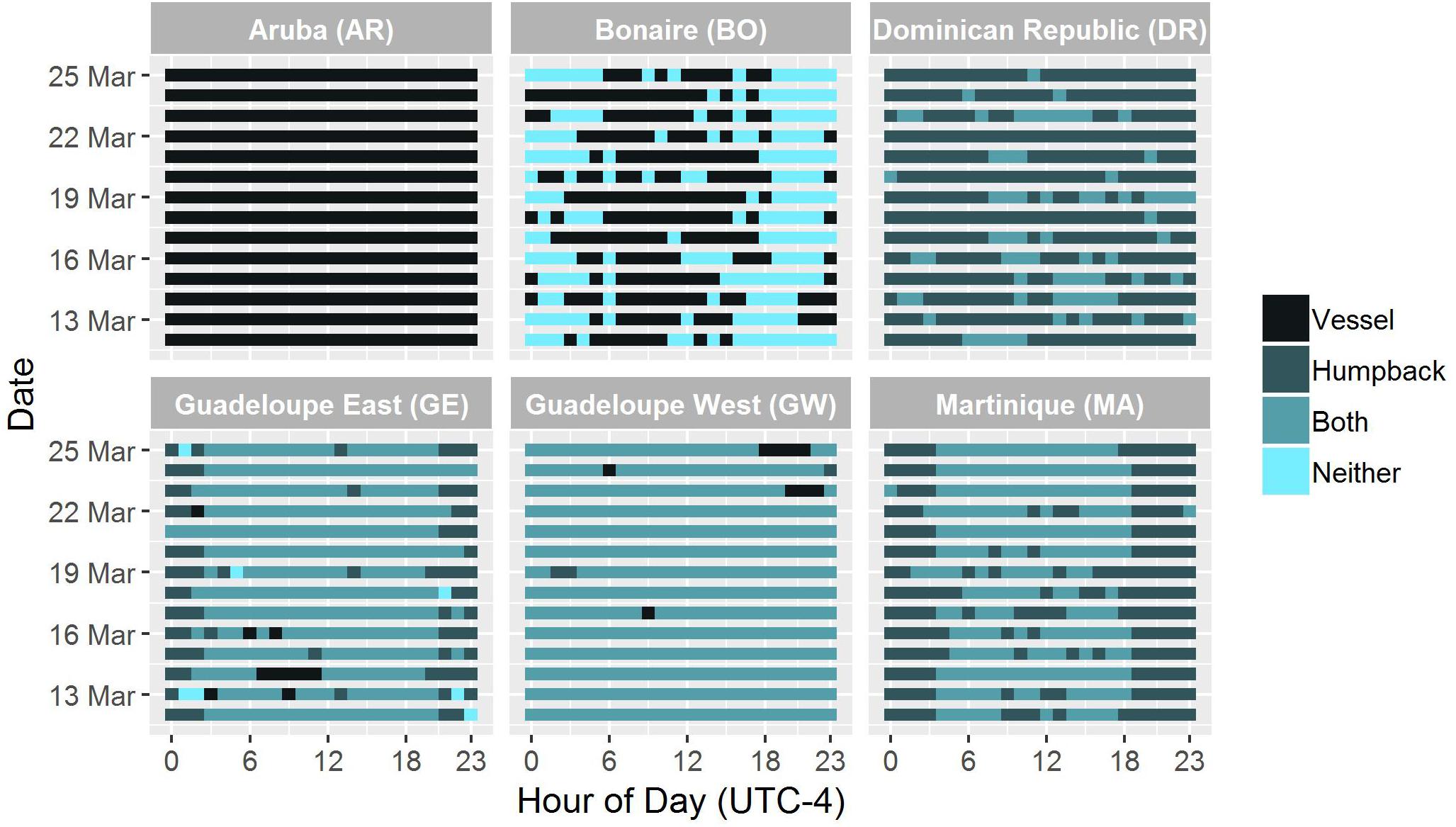
Figure 4. Hourly presence of vessel noise, humpback whale song, both or neither at each recording site from 12 March 2017 to 25 March 2017.
At the GW site, both humpback song and persistent vessel noise were present each day. The majority of recording hours were characterized by overlapping humpback song and vessel noise characterized the majority of the hours recorded (96%). Only 12 h across 5 different days, included bouts of only vessel noise or only whale song. There were 9 h without whale song and only 3 h without vessel noise at the GW site (Figure 4). Similar to the AR site, it was not possible to estimate the number of vessel passages or their duration at GW due to heavy ship traffic and continuous vessel noise (Supplementary Figure S1).
Vessel activity and song presence was more variable at the GE site, with overlapping whale song and vessel noise being predominant (vessel noise 3% of time), humpback song (21.1% of time), both sounds (73.8% of time), and neither sound (2.1% of time) (Figure 4). This eastern site was further removed from the ports reflecting the slight decrease in vessel noise for this area (Supplementary Figure S1). Diel patterns in the number of vessel passages were examined by plotting the number of vessel passages without humpback song for the four sites, GE, BO, MA, and GE (Figure 5). SM was excluded from the diel pattern analysis because of the duty-cycled nature of the data. At GE, a diel pattern in the number of vessel passages was observed with vessel noise receding late at night between the hours of 21:00 and 3:00 UTC-4 (Figure 5). The greatest density of vessels passages occurred during daylight hours.
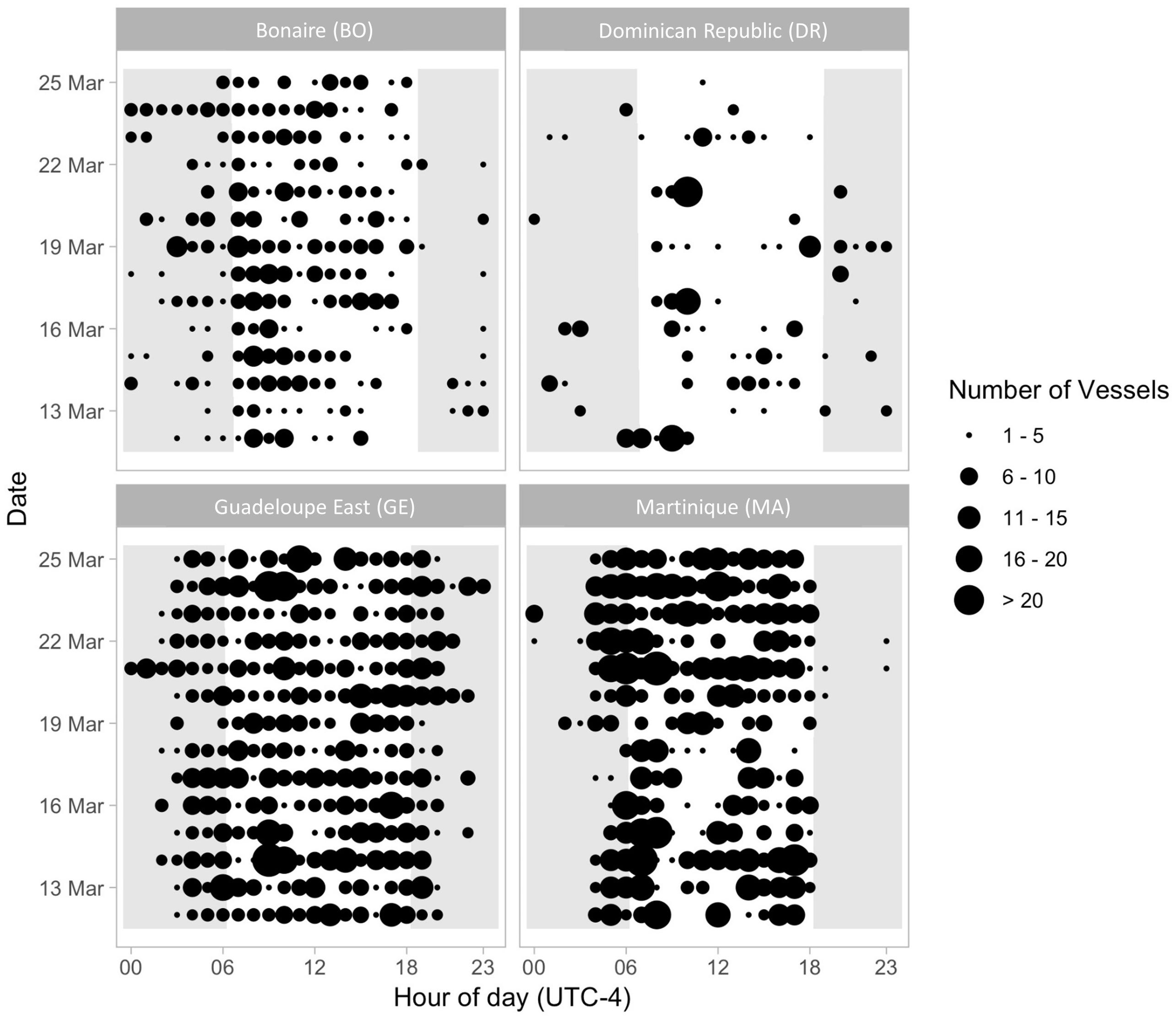
Figure 5. Diel patterns in the hourly number of vessel passages at each recording site from 12 March 2017 to 25 March 2017. Dot size indicates the number of vessel passages acoustically detected for per hour on each day. Light gray shading indicates hours before sunrise and after sunset.
Similar to AR, humpback whale song was not detected at the BO site. Instead, the recordings at BO consisted of hours with only vessel noise (53.6% of time) or neither humpback song nor vessel sound (46.4%) (Table 2 and Figure 4). The BO site was located in the marine park and away from the main ports (Supplementary Figure S1). A less clear diel pattern was present in the number of vessel passages at BO compared to GE. Although the change in number of vessel passages was less distinct, fewer vessels were present during night between the hours of 18:00 and 06:00 (Figure 5).
MA had both hours with humpback song only (47.6%) and both humpback song and vessel noise (52.4%, Figure 4). The MA location was in deeper water and on the opposite side of the island to the main port (Supplementary Figure S1). There was a very distinct diurnal pattern with vessel passages being almost completely absent between 18:00:00 and 05:00 (Figure 5).
SM data were duty-cycled sampling for 1 h every 4 h, therefore only 6 h of recordings were available each day (n = 84 h) for analysis. Humpback song presence was notable at this site, with 67.8% of the sampled data containing only humpback song. Vessel noise was also present and overlapped with whale song for 29.8% of the time. The location of this recorder was also far from the islands main ports (Supplementary Figure S1). Due to the duty cycle of the recorder and the limited number of hours for analysis a diel comparison was not attempted.
Lastly, the DR site had no hours with solely vessel presence while hours with humpback song made up the majority of the soundscape (79.8%); humpback song and vessel noise did overlap occasionally (20.2%) (Table 2 and Figure 4). Humpback whale song was present in every hour of the 2-week sample period and was often present without vessel noise in those hours. The DR site was located the furthest offshore compared to all the sites, likely reflecting the decreased vessel noise present on these recordings (Supplementary Figure S1). Although the total number of vessel passages was greatest during daylight hours, there were vessels present at night as well, and no clear diel pattern was apparent due to sparse vessel presence at this site (Figure 5).
High Frequency Analyses – Odontocetes
At the BO site, sounds from odontocetes, namely unknown delphinid species and sperm whales (Physeter macrocephalus) detected sporadically throughout the dataset (Figure 3B). Dolphin sounds usually consisted of whistles. Sperm whale detections included echolocation clicks as well as sounds used in social communication, including slow clicks and a trumpet-like sound. Fish sounds were constantly present but were not analyzed for this study. At the SM site, possible dolphin sounds were noted only once in the SoundTrap recordings on 30 April 2017. Snapping shrimp dominated all high-frequency recordings.
Acoustic Analysis of an Earthquake
A suspected earthquake was initially discovered in the GE recordings and was subsequently detected at all other recording sites. A United States Geological Survey (USGS) website4 and local news posts5,6 were used to verify that a 5.6 magnitude earthquake had occurred at 05:23 UTC on 17 April 2017. The epicenter of this 5.6 magnitude earthquake was in the Flinn Engdahl region of Antigua and Barbuda, Leeward Islands (Figure 1). The sound from the earthquake was recorded within an 8.75 min window at all recording sites (Table 3). The relative sound energy and duration of acoustic signal of the earthquake was highest near to the epicenter and decreased as it was received on recorders further away (Supplementary Figure S2). The number of aftershocks recorded varied across sites, with the highest numbers recorded at GE and GW and fewer at MA, SM, DR, AR, and BO.
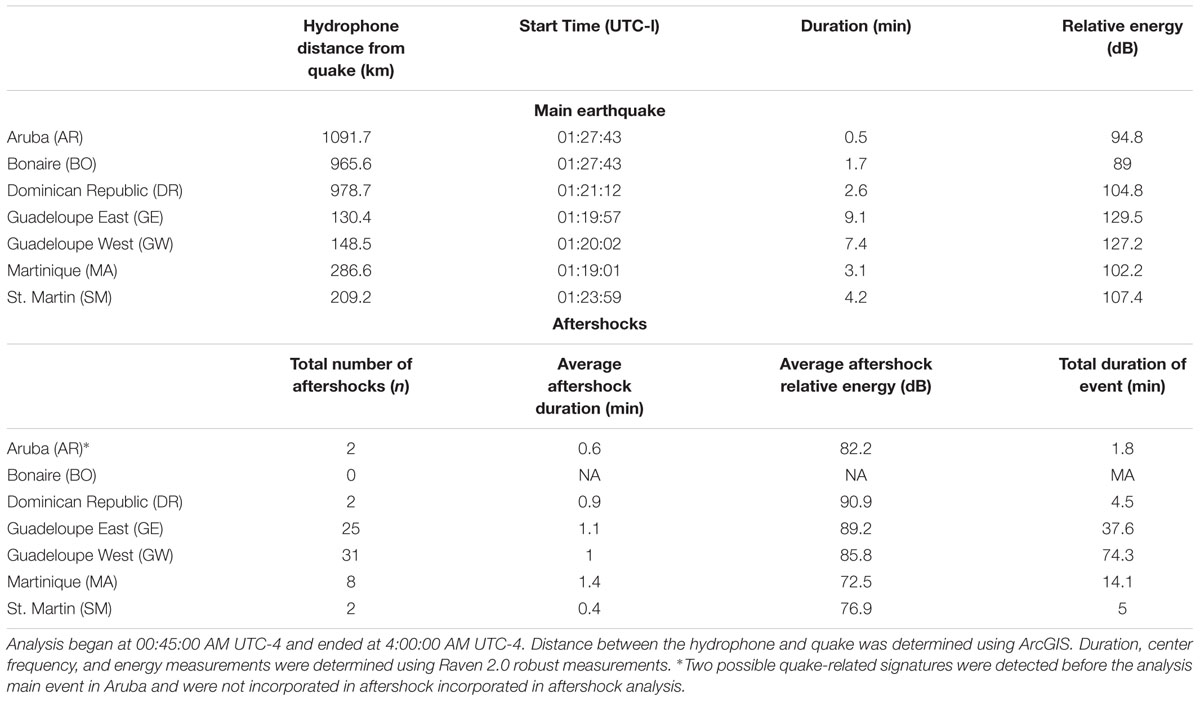
Table 3. Summary of 17 April 2017 earthquake and aftershock analysis.
Discussion
This study provides a broad scale view of the composition of marine soundscapes across seven sites located throughout the Caribbean region. All but one site was located within marine park waters and for most of these areas, this study provides the first comprehensive look at the acoustic environment, species composition and anthropogenic footprint within each park. By quantifying the soundscape at each site, this study demonstrates the value of this approach and the importance of continued long-term monitoring of each distinctive marine environment. Continued data collection would support the capacity to track changes in species presence and increases or decreases in anthropogenic noise, as well as provide an index for evaluating and understanding the health of each underwater environment over time (Dumyahn and Pijanowski, 2011; Nedelec et al., 2015; Schmeller et al., 2017).
Humpback whale sound was present across all islands other than BO and AR. There were clear differences in the timing of song presence at the five sites where humpback whale song was recorded, with singers arriving later in the year at the eastern Caribbean sites (GW, EG, and MA). Song was detected at all five sites until mid-to late-May, greatly extending the time over which humpbacks are known to occur at DR and SM. A detailed analysis of song patterns was beyond the scope of this study, and further analyses of song recorded at all sites would be informative. So, the data collected from this study demonstrates that the preponderance of whale photo-identifications made at the eastern Caribbean sites is not a sampling artifact, but is a true indication of the timing of these whales’ presence in the region. This reinforces Stevick et al.’s (2018) contention that this is a second breeding population of humpback whales occurring in Caribbean waters.
Minke whales, sperm whales, dolphins, snapping shrimp and fish choruses made up the rest of the biological composition of the soundscapes at these sites. Minke whales in the Caribbean were documented previously in one PAM study off Saba (Risch et al., 2014) and visually confirmed by Debrot et al. (2011, 2013) before the 6-month acoustic monitoring effort off MA in the present study. The seasonal presence of minke whales around MA was similar to that previously reported, with pulse trains occurring from the beginning of the deployment in late December through mid- March, further supporting a likely wintering ground for minke whales in Caribbean waters.
The BO site, in particular, had a rich and diverse presence of fish sounds, and future research into chorusing patterns and spatial and temporal variation of fish sounds in these recordings would provide further insight into this important biological component of the soundscape (e.g., Staaterman et al., 2013; Nedelec et al., 2015). Sperm whales and dolphins were also present off BO, a coral reef island and is surrounded by fringing reef. The Caribbean Sea basin plunges to depths of more than 5000 m not far from BO, which is likely why sperm whales, a deep-diving species, could be heard at this recording site. Sperm whales have been included in many descriptions of marine mammals throughout the Caribbean (Debrot and Barros, 1994; Jefferson and Lynn, 1994; Debrot et al., 2011, 2013; Luksenburg, 2014), and at least six different species of dolphins have been sighted in the waters around Aruba, Bonaire, and Curacao (Luksenburg, 2014; Geelhoed et al., 2014).
AR was located on the northern side of the island and appeared to be our least biologically rich site, as the location of the site was near the main shipping channel for vessels coming to and past the island (Figure 1). No marine mammals were detected on these recordings and further efforts should be made to evaluate where areas of higher biodiversity, and less shipping noise, may occur around the island.
Our geological finding of the 17 April 2017 earthquake was an unexpected discovery and demonstrates the ability of soundscape monitoring to also record major geological and weather events in an area (Speeth, 1961; Locascio and Mann, 2005). The acoustic signal of the earthquake traveled to all our recordings sites over a distance of more than 1000 km. Earthquakes have been shown to travel underwater for at least 1000–5000 km (e.g., Astafyeva and Afraimovich, 2006).
Vessel traffic in the Caribbean was a considerable contributor to the soundscape of each site, recorded on 20–100% of hours during the 2-week recording period that was analyzed in detail. Since the early 1960s, there has been a dramatic increase in worldwide ship traffic, both commercial and recreational (Eyring et al., 2005; McDonald et al., 2006). Vessel noise levels and spectral signatures vary considerably depending in vessel characteristics and operations (McKenna et al., 2012, 2013). Noise produced by large commercial vessels has repeatedly been shown to impact the ability of endangered whales, fish and other marine animals to maintain acoustic contact, especially in protected areas such as designated areas of special biological interest such as sanctuaries and marine parks (e.g., Hatch et al., 2008, 2016; Stanley et al., 2017). In addition, smaller vessels such as fishing boats and tourism such as whale watching have been shown to contribute to a significant proportion of masking and in situations where both large and smaller vessels exist the effect is cumulative (Cholewiak et al., 2018). As a result, the significantly high levels of vessel noise as well as the number of vessel passages especially for AR, GE, GW, and MA ought to raise concern for the managers in charge of the marine parks in and around these sites.
Global shipping transit maps show clear routes passing through the southern Caribbean islands and the leeward part of the Caribbean sea (e.g., Halpern et al., 2008). Cruise ship tourism in the Caribbean is a large industry that has grown substantially over the past three decades (Sprague-Silgado, 2017). One way to understand the differences in vessel noise and passages across our study sites is to look at the location of the ports on each of the islands (Supplementary Figure S1). Where the recorders were further from ports, such as at the DR, less vessel noise was recorded (Figure 5). Similarly GE and GW differed, with less vessel noise in GE. In this case GW was located on the leeward side of the island, closer to a port than was GE.
GE and MA had very distinctive diurnal patterns with few to no vessel passages during the night, while vessel passages at BO and the DR were more broadly dispersed throughout both day and night. These diurnal patterns could provide an indication of the types of vessels and activities occurring at each site. Clear diurnal patterns in vessel presence suggests traffic consisting of smaller recreational vessels or fishing vessel activity, while the lack of a diurnal pattern may reflect the presence of large container ships and cruise liners. In the DR, the distant location of the recorder makes is almost certain that a considerable amount of the vessel noise heard was produced by the illegal fishing fleet which converges in this area. Members of this fleet consist of one large mother ship from which 25–50 smaller vessels are dispersed to engage in fishing throughout Silver Bank (Salas et al., 2011). For this situation, PAM offers an opportunity to monitor the level of timing of illegal fishing within the marine park.
All recorders, with the exception of the one off Aruba, were sited in designated Marine Protected Areas. The GE, GW, MA, and SM recorders were within the Agoa Sanctuary. This MPA, created in 2012, includes all the waters of the Exclusive Economic Zone of the French West Indies7. The BO recorder was in the Bonaire National Marine Park, established in 1979, primarily for the protection of coral reefs. This MPA comprises the coastal waters of the islands of Bonaire and Klein Bonaire to a depth of 60 m8. The Silver and Navidad Bank Sanctuary, now the Sanctuary for Marine Mammals of the Dominican Republic was first declared in 1986 (Mattila et al., 1989). Of the MPAs where recorders were placed, the DR recorder was the only one in an MPA that includes measures specifically established to manage human use of the area for marine mammal protection.
Apart from masking humpback song, vessel traffic risks masking the very quiet calls between female whales and their calves on the calving ground (Videsen et al., 2017). While the Sanctuary for Marine Mammals of the Dominican Republic is removed from shipping lanes, the amount of shipping into the Agoa Sanctuary and the Bonaire National Marine Park is more substantial. Recent work has considered how MPAs can include mitigating anthropogenic sound into management planning (e.g., Erbe et al., 2012; Williams et al., 2014, 2015). Zoning planning for the Agoa Sanctuary could consider ways to mitigate the impact of anthropogenic noise on humpback whales.
As well as acoustic effects, shipping traffic also poses the risk of vessel strike on whales. Humpback females with newborn calves, given their behavior, are a particular concern (Bejder et al., 2019). Spatially explicit management to reduce the likelihood of shipping collision with large whales is well developed (e.g., Conn and Silber, 2013), and tools to notify mariners of the presence of whales are also available (Baumgartner et al., 2019). Again, we encourage when planners are zoning the Agoa Sanctuary, they consider these management options.
Overall, this study has shown that vessel traffic is as much part of the soundscapes of many Caribbean marine park sites as marine megafauna, including humpback whales. It is difficult to assess how the presence of anthropogenic noise may interfere with the essential mating activities of humpback whales, however, we have documented significant overlap in humpback whale song and vessel noise at several Caribbean sites throughout the breeding season. In addition, impacts of masking on other marine animals is likely, especially at the sites with higher noise levels. Passive acoustics provides an invaluable tool for monitoring long term composition of a marine animals and anthropogenic contributors in marine parks. Marine park managers should consider this approach when designing long term monitoring and management strategies for their sites.
Data Availability
The datasets generated for this study are available on request to the corresponding author.
Author Contributions
HH was the postdoctoral candidate who organized, planned, and ran the study. GD, JS, LK, and CR were all involved in the analyses of the data from this study. PC and SVP provided the senior scientist oversight, funding, and conceptual ideas for the study. LB, JC, JK, AH, OS-R, FV, WDW, and PH were all linked to one or more of the marine parks or deployment sites and provided logistical support, divers, helped obtain permits and facilitated the collection of the data in each of their respective regions.
Funding
This project was funded through a National Marine Fisheries Service, The Office of Science and Technology, International Science Grant.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Many thanks to the entire field team for their hard work to make all of this possible. We would specifically like to thank Danielle Cholewiak, Leah Crowe, Julianne Gurnee, Pascual Prota, Yamil Rodríguez Asilis, Hinya De Peña, Werner Leo Varela, Gus Torreira, Kerenza Rannou, Sabine Engel, Luciano Mazzeo, Castro Perez, Nicolas Maslach, Franck Roncuzzi, Nelly Pélisson, Thibaud Rossard, Alain Goyeau, Marlene, Dany Moussa, Hervé Magnin, Axel Priouzeau, Jeffrey Bernus, Denis Etienne, and M. Thibaut Kalbe. In addition, we would also like to thank the Bioacoustics Research Program team at Cornell Lab of Ornithology especially Chris Tessaglia-Hymes, Edward James Moore III, Daniel Patrick Salisbury, and Mark Renkawitz from NOAA NEFSC. Without Mark’s shipping expertise we never would have gotten our equipment to the sites. CHAMP was supported by funding from NOAA as well as in-kind support from the Dominican Republic’s National Authority for Maritime Affairs, the Aruba Marine Mammal Foundation, the Bonaire National Marine Park and National Office for the Caribbean Netherlands, the Dutch Caribbean Nature Alliance, the Observatoire des Mammifères Marins de l’Archipel Guadeloupéen (OMMAG), the National Park of Guadeloupe, the Natural Reserve of Saint Martin, and Martinique’s Authority for Maritime Affairs. All research was conducted with permission from the appropriate authorities. Research in the Dominican Republic was conducted under authorization from the Ministro de Medio Ambiente y Recursos Naturales. Research in Aruba was conducted under authorization from the Directorate of Shipping Aruba. Research in Bonaire was conducted under authorization from the Ministerie van Infrastructuur en Milieu. Research in St. Martin was conducted under authorization from the Reserve Naturelle de Nationale de Saint Martin. Research in Guadeloupe was conducted under authorization from the Parc National de la Guadeloupe and the Direction de la Mer de la Guadeloupe. Research in Martinique was conducted under authorization from the Direction de la Mer de la Martinique. We are grateful to the two reviewers, and the manuscript’s editor, whose comments greatly improved this work.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2019.00347/full#supplementary-material
FIGURE S1 | Detailed maps of the recording sites located near each island, with symbols indicating the type of recorder deployed at each site [Marine Acoustic Recording Unit (MARU), SoundTrap, or both], as well as the locations of all major ports.
FIGURE S2 | The acoustic signature of an earthquake detected at 01:23 UTC-4 on 17 April, 2017 off the Antigua Islands. The MARU at Guadeloupe East was closest to the epicenter of the quake at 130 km while the St. Martin SoundTrap was approximately 209 km from the epicenter. The MARU at Aruba was the furthest from the epicenter at 1091 km.
Footnotes
- ^ https://www.nature.com/articles/s41598-018-36870-7
- ^ https://www.oceaninstruments.co.nz/
- ^ http://www.arcGIS.com
- ^ https://earthquake.usgs.gov/earthquakes/eventpage/us10008iaa/executive#executive
- ^ https://www.themontserratreporter.com/leeward-islands-jolted-by-strong-tremor/
- ^ http://dominicanewsonline.com/news/homepage/news/general/src-records-series-of-earthquakes-off-antigua/
- ^ http://www.sanctuaire-agoa.com/
- ^ https://stinapabonaire.org/bonaire-national/
References
American National Standards Institute (1994). Acoustical Terminology. ANSI S1.1-1994. New York, NY: ANSI.
Astafyeva, E. I., and Afraimovich, E. L. (2006). Long-distance traveling ionospheric disturbances caused by the great Sumatra-Andaman earthquake on 26 December 2004. Earth Planets Space. 58, 1025–1031. doi: 10.1186/bf03352607
Baumgartner, M., Bonnell, J., Van Parijs, S. M., Corkeron, P. J., Hotchkin, C., Ball, K., et al. (2019). Persistent near real-time passive acoustic monitoring for baleen whales from a moored buoy: system description and evaluation. Methods Ecol. Evol.
Bejder, L., Videsen, S., Hermannsen, L., Simon, M., Hanf, D., and Madsen, P. T. (2019). Low energy expenditure and resting behaviour of humpback whale mother-calf pairs highlights conservation importance of sheltered breeding areas. Sci. Rep. 9:771. doi: 10.1038/s41598-018-36870-7
Benoit-Bird, K. J., and Au, W. W. (2009). Phonation behavior of cooperatively foraging spinner dolphins. J. Acoust. Soc. Am. 125, 539–546. doi: 10.1121/1.2967477
Bettridge, S., Baker, C. S., Barlow, J., Clapham, P. J., Ford, M., Gouveia, D., et al. (2015). Status review of the humpback whale (Megaptera novaeangliae) under the Endangered Species Act NOAA NMFS. [Online]. Available at www.nmfs.noaa.gov/pr/species/Status%20Reviews/humpback_whale_sr_2015.pdf (accessed February 20, 2019).
Calupca, T. A., Fristrup, K. M., and Clark, C. W. (2000). A compact digital recording system for autonomous bioacoustic monitoring. J. Acoust. Soc. Am. 108:2582. doi: 10.1121/1.4743595
Cholewiak, D., Clark, C. W., Ponirakis, D., Frankel, A., Hatch, L. T., Risch, D., et al. (2018). Communicating amidst the noise: Modeling the aggregate influence of ambient and vessel noise on baleen whale communication space in a national marine sanctuary. Endang. Species Res. 36, 59–75. doi: 10.3354/esr00875
Clark, C. W., Ellison, W. T., Southall, B. L., Hatch, L., Van Parijs, S. M., Frankel, A., et al. (2009). Acoustic masking in marine ecosystems: Intuitions, analysis, and implication. Mar. Ecol. Prog. Ser. 395, 201–222. doi: 10.3354/meps08402
Conn, P. B., and Silber, G. (2013). Vessel speed restrictions reduce risk of collision-related mortality for North Atlantic right whales. Ecosphere 4:43. doi: 10.1890/ES13-00004
Davis, G. E., Baumgartner, M. F., Bonnell, J. M., Bell, J., Berchock, C., Bort Thornton, J., et al. (2017). Long-term passive acoustic recordings track the changing distribution of North Atlantic right whales (Eubalaena glacialis) from 2004 to 2014. Sci Rep. 7:13460. doi: 10.1038/s41598-017-13359-3
Debrot, A., Witte, R., and Scheidat, M. (2011). The Marine Mammals of the Dutch Caribbean: A Comparison Between EEZ Sectors, Contrasts and Concerns International Whaling Commission Working. Paper Presented SC/63/E9. Impington
Debrot, A. O., and Barros, N. B. (1994). Additional cetacean records for the leeward Dutch Antilles. Mar. Mammal Sci. 10, 359–368. doi: 10.1111/j.1748-7692.1994.tb00491.x
Debrot, A. O., Esteban, N., Bervoets, T., Hoetjes, P. C., and Scheidat, M. (2013). Marine mammals of the northeastern Caribbean windward Dutch Islands: Saba, St. Eustatius, St. Maarten, and the Saba Bank. Car. J. Sci. 47, 159–172. doi: 10.18475/cjos.v47i3.a5
di Sciara, G. N., Hoyt, E., Reeves, R., Ardron, J., Marsh, H., Vongraven, D., et al. (2016). Place-based approaches to marine mammal conservation.”. aquatic conservation: Mar. Freshwater Ecosyst. 26, 85–100. doi: 10.1002/aqc.2642
Dumyahn, S. L., and Pijanowski, B. C. (2011). Soundscape conservation. Lands. Ecol. 26, 1327–1344. doi: 10.1007/s10980-011-9635-x
Erbe, C., Liong, S., Koessler, M. W., Duncan, A. J., and Gourlay, T. (2016). Underwater sound of rigid-hulled inflatable boats. J. Acoust. Soc. Am. 139, EL223–EL227. doi: 10.1121/1.4954411
Erbe, C., MacGillivray, A., and Williams, R. (2012). Mapping cumulative noise from shipping to inform marine spatial planning. J. Acoust. Soc. Am. 132, EL423–EL428. doi: 10.1121/1.4758779
Erbe, C., Verma, A., Mccauley, R., Gavrilov, A., and Parnum, I. (2015). The marine soundscape of the Perth Canyon. Progr. Oceanogr. 137, 38–51. doi: 10.1016/j.pocean.2015.05.015
Eyring, V., Köhler, H. W., Van Aardenne, J., and Lauer, A. (2005). Emissions from international shipping: 1. The last 50 years. J. Geophys. Res. 110:D17.
Geelhoed, S. C. V., Janinhoff, N., Verdaat, J. P., Bemmelen, R. S. A. V., and Scheidat, M. (2014). Aerial surveys of marine mammals and other fauna around Aruba, Curacao and Bonaire, November 2013. Institute for Marine Resources & Ecosystem Studies Report C012/14. http://edepot.wur.nl/292240. (accessed January 28, 2014).
Halpern, B. S., Walbridge, S., Selkoe, K. A., Kappel, C. V., Micheli, F., D’agrosa, C., et al. (2008). A global map of human impact on marine ecosystems. Science 319, 948–952.
Hatch, L., Clark, C., Merrick, R., Van Parijs, S. M., Ponikaris, D., Schwehr, K., et al. (2008). Characterizing the relative contributions of large vessels to total ocean noise fields: a case study using the Gerry E. Studds Stellwagen Bank National Marine Sanctuary. Environ. Manage. 42, 735–752. doi: 10.1007/s00267-008-9169-4
Hatch, L. T., Clark, C. W., Van Parijs, S. M., Frankel, A. S., and Ponirakis, D. W. (2012). Quantifying loss of acoustic communication space for right whales in and around a U.S. National Marine Sanctuary. Conserv. Biol. 26, 983–994. doi: 10.1111/j.1523-1739.2012.01908.x
Hatch, L. T., and Fristrup, K. M. (2009). No barrier at the boundaries: implementing regional frameworks for noise management in protected natural areas.”. Mar. Ecol. Progr. Series 395, 223–244. doi: 10.3354/meps07945
Hatch, L. T., Wahle, C. M., Gedamke, J., Harrison, J., Laws, B., Moore, S. E., et al. (2016). Can you hear me here? Managing acoustic habitat in US waters. Endang. Species Res. 30, 171–186. doi: 10.3354/esr00722
Haver, S. M., Klinck, H., Nieukirk, S. L., Matsumoto, H., Dziak, R. P., and Miksis-Olds, J. L. (2017). The not-so-silent world: Measuring Arctic, Equatorial, and Antarctic soundscapes in the Atlantic Ocean. Deep-Sea Res. Part I. 122, 95–104. doi: 10.1016/j.dsr.2017.03.002
Heenehan, H. L., Tyne, J. A., Bejder, L., Van Parijs, S. M., and Johnston, D. W. (2016). Passive acoustic monitoring of coastally associated Hawaiian spinner dolphins, Stenella longirostris, ground-truthed through visual surveys. J. Acoust. Soc. Am. 140, 206–215. doi: 10.1121/1.4955094
Heenehan, H. L., Van Parijs, S. M., Bejder, L., Tyne, J. A., Southall, B. L., Southall, H., et al. (2017). Natural and anthropogenic events influence the soundscapes of four bays on Hawaii Island. Mar. Pollut. Bull. 124, 9–20. doi: 10.1016/j.marpolbul.2017.06.065
Herman, L. M. (2017). The multiple functions of male song within the humpback whale (Megaptera novaeangliae) mating system: Review, evaluation, and synthesis. Biol Rev. 92, 1795–1818. doi: 10.1111/brv.12309
Hildebrand, J. A. (2009). Anthropogenic and natural sources of ambient noise in the ocean. Mar. Ecol. Prog. Ser. 395, 5–20. doi: 10.1146/annurev-marine-121211-172423
Hoffmann, J. (1997). Facilitation of Trade and Transport in Latin America and the Caribbean. FAL Bulletin # 136. http://www.cepal.org/Transporte/noticias/bolfall/3/5123/fal136e.html (accessed March 14, 2019).
Janik, V. M., and Sayigh, L. S. (2013). Communication in bottlenose dolphins: 50 years of signature whistle research. J. Compar. Physiol. A 199, 479–489. doi: 10.1007/s00359-013-0817-7
Jefferson, T. A., and Lynn, S. K. (1994). Marine mammal sightings in the Caribbean Sea and Gulf of Mexico, summer 1991. Car. J. Sci. 30, 83–89.
Kennedy, A. S. (2018). From whaling to tagging: The evolution of North Atlantic humpback whale research in the West Indies. Mar. Fish. Rev. 79, 23–37. doi: 10.7755/mfr.79.2.2
Knowles, J. E., Doyle, E., Schill, S. R., Roth, L. M., Milam, A., and Raber, G. T. (2015). Establishing a marine conservation baseline for the insular Caribbean.”. Mar. Policy 60, 84–97. doi: 10.1016/j.marpol.2015.05.005
Krause, B. L., and Gage, S. H. (2003). Testing Biophony as an Indicator of Habitat Fitness and Dynamics. Natural Soundscape Vital Signs Pilot Program Report. Sequoia National Park: Wild Sanctuary, Inc.
Locascio, J. V., and Mann, D. A. (2005). Effects of hurricane charley on fish chorusing. Biol. Lett. 1, 362–365. doi: 10.1098/rsbl.2005.0309
Luksenburg, J. A. (2014). The cetaceans of Aruba, southern Caribbean. J. Mar. Biol. Assoc. U.K. 94, 1161–1174. doi: 10.1017/S0025315413000337
Mattila, D. K., Clapham, P. J., Katona, S. K., and Stone, G. S. (1989). Population composition of humpback whales, Megaptera novaeangliae, on Silver Bank, 1984. Can. J. Zool. 67, 281–285. doi: 10.1139/z89-041
McDonald, M. A., Hildebrand, J. A., and Wiggins, S. M. (2006). Increases in deep ocean ambient noise in the Northeast Pacific west of San Nicolas Island, California. J. Acous. Soc. Am. 120, 711–718. doi: 10.1121/1.2216565
McKenna, M. F., Gabriele, C., and Kipple, B. (2017). Effects of marine vessel management on the underwater acoustic environment of Glacier Bay National Park, AK. Ocean Coast. Manag. 139, 102–112. doi: 10.1016/j.ocecoaman.2017.01.015
McKenna, M. F., Ross, D., Wiggins, S. M., and Hildebrand, J. A. (2012). Underwater radiated noise from modern commercial ships. J. Acous. Soc. Am. 131, 92–103. doi: 10.1121/1.3664100
McKenna, M. F., Wiggins, S. M., and Hildebrand, J. A. (2013). Relationship between container ship underwater noise levels and ship design, operational and oceanographic conditions. Sci. Rep. 3:1760.
McWilliam, J. N., and Hawkins, A. D. (2013). A comparison of inshore marine soundscapes. J. Exp. Mar. Biol. Ecol. 446, 166–176. doi: 10.1016/j.jembe.2013.05.012
Merchant, N. D., Fristrup, K. M., Johnson, M. P., Tyack, P. L., Witt, M. J., Blondel, P., et al. (2015). Measuring acoustic habitats. Methods Ecol. Evol. 6, 257–265.
Merchant, N. D., Pirotta, E., Barton, T. R., and Thompson, P. M. (2014). Monitoring ship noise to assess the impact of coastal developments on marine mammals. Mar. Poll. Bull. 78, 85–95. doi: 10.1016/j.marpolbul.2013.10.058
Nedelec, S. L., Simpson, S. D., Holderied, M., Radford, A. N., Lecellier, G., Radford, C., et al. (2015). Soundscapes and living communities in coral reefs: temporal and spatial variation. Mar. Ecol. Prog. Ser. 524, 125–135. doi: 10.3354/meps11175
Payne, R. S., and McVay, S. (1971). Songs of humpback whales. Science 173, 585–597. doi: 10.1126/science.173.3997.585
Pijanowski, B. C., Farina, A., Gage, S. H., Dumyahn, S. L., and Krause, B. L. (2011). What is soundscape ecology? An introduction and overview of an emerging new science. Lands. Ecol. 26, 1213–1232. doi: 10.1007/s10980-011-9600-8
Radford, C. A., Stanley, J. A., Simpson, S. D., and Jeffs, A. G. (2011). Juvenile coral reef fish use sound to locate habitats. Coral Reefs 30, 295–305. doi: 10.1007/978-1-4939-2981-8_129
Risch, D., Castellote, M., Clark, C. W., Davis, G. E., Dugan, P. J., Hodge, L. E. W., et al. (2014). Seasonal migrations of North Atlantic minke whales: novel insights from large-scale passive acoustic monitoring networks. Move. Ecol. 2:24. doi: 10.1186/s40462-014-0024-3
Risch, D., Clark, C. W., Dugan, P. J., Popescu, M., Siebert, U., and Van Parijs, S. M. (2013). Minke whale acoustic behavior and multi-year seasonal and diel vocalization patterns in Massachusetts Bay. USA. Mar. Ecol. Progr. Series 489, 279–295. doi: 10.3354/meps10426
Rolland, R. M., Parks, S. E., Hunt, K. E., Castellote, M., Corkeron, P. J., Nowacek, D. P., et al. (2012). Evidence that ship noise increases stress in right whales. Proc. R Soc. B Biol. Sci. 279, 2363–2368. doi: 10.1098/rspb.2011.2429
Ryan, C., Wenzel, F., Suarez, P. L., and Berrow, S. D. (2014). An abundance estimate for Megaptera novaeangliae breeding around Boa Vista. Cape Verde Islands. Zoological Caboverdiana 5, 20–28.
Salas, S., Chuenpagdee, R., Charles, A. T., and Seijo, J. C. (eds) (2011). Coastal Fisheries of Latin America and the Caribbean. Rome: Food and Agriculture Organization of the United Nations.
Schmeller, D. S., Böhm, M., Arvanitidis, C., Barber-Meyer, S., Brummitt, N., Chandler, M., et al. (2017). Building capacity in biodiversity monitoring at the global scale. Biodiver. Conser. 26, 2765–2790. doi: 10.1007/s00267-017-0866-8
Sprague-Silgado, J. (2017). The Caribbean cruise ship business and the emergence of a transnational capitalist class. J. World Sys. Res. 23, 93–125. doi: 10.5195/jwsr.2017.623
Staaterman, E., Rice, A. N., Mann, D. A., and Paris-Limouzy, C. B. (2013). Soundscapes from a Tropical Eastern Pacific reef and a Caribbean Sea reef. Coral Reefs 32, 553–557. doi: 10.1007/s00338-012-1007-8
Stanley, J. A., Van Parijs, S. M., and Hatch, L. T. (2017). Underwater sound from vessel traffic reduces the effective communication range in Atlantic cod and haddock. Sci. Rep. 7:14633. doi: 10.1038/s41598-017-14743-9
Stevick, P. T., Berrow, S. D., Bérubé, M., Bouveret, L., Broms, F., Jann, B., et al. (2016). There and back again: Multiple and return exchange of humpback whales between breeding habitats separated by an ocean basin. J. Mar. Biol. Assoc. U.K. 96, 885–890. doi: 10.1017/s0025315416000321
Stevick, P. T., Bouveret, L., Gandilhon, N., Rinaldi, C., Rinaldi, R., Broms, F., et al. (2018). Migratory destinations and timing of humpback whales in the southeastern Caribbean differ from those off the Dominican Republic. J. Cetacean Res. Manag. 18, 127–133.
Stevick, P. T., Øien, N., and Mattila, D. K. (1998). Migration of a humpback whale (Megaptera novaeangliae) between Norway and the West Indies. Mar. Mammal Sci. 14, 162–166. doi: 10.1111/j.1748-7692.1998.tb00701.x
Van Opzeeland, I., and Boebel, O. (2018). Marine soundscape planning: seeking acoustic niches for anthropogenic sound. J. Ecoacoust. 2:5GSNT8. doi: 10.22261/jea.5gsnt8
Van Parijs, S. M., Smith, J., and Corkeron, P. J. (2002). Using calls to estimate the abundance of inshore dolphins: A case study with Pacific humpback dolphins Sousa chinensis. J. Appl. Ecol. 39, 853–864. doi: 10.1046/j.1365-2664.2002.00756.x
Videsen, S. K., Bejder, L., Johnson, M., and Madsen, P. T. (2017). High suckling rates and acoustic crypsis of humpback whale neonates maximise potential for mother–calf energy transfer. Funct. Ecol. 31, 1561–1573. doi: 10.1111/1365-2435.12871
Vu, E. T., Risch, D., Clark, C. W., Gaylord, S., Hatch, L. T., Thompson, M. A., et al. (2012). Humpback whale song occurs extensively on feeding grounds in the western North Atlantic Ocean. Aquatic Biol. 14, 175–183. doi: 10.3354/ab00390
Weilgart, L. S. (2007). The impacts of anthropogenic ocean noise on cetaceans and implications for management. Can. J. Zool. 85, 1091–1116.
Williams, R., Clark, C. W., Ponirakis, D., and Ashe, E. (2014). Acoustic quality of critical habitats for three threatened whale populations.”. Animal Conser. 17, 174–185. doi: 10.1111/acv.12076
Williams, R., Wright, A. J., Ashe, E., Blight, L. K., Bruintjes, R., Canessa, R., et al. (2015). Impacts of anthropogenic noise on marine life: publication patterns, new discoveries, and future directions in research and management. Ocean Coastal Manag. 115, 17–24. doi: 10.1016/j.ocecoaman.2015.05.021
Keywords: passive acoustic monitoring, soundscape, marine mammal, humpback whale, anthropogenic noise, Caribbean
Citation: Heenehan H, Stanistreet JE, Corkeron PJ, Bouveret L, Chalifour J, Davis GE, Henriquez A, Kiszka JJ, Kline L, Reed C, Shamir-Reynoso O, Védie F, De Wolf W, Hoetjes P and Van Parijs SM (2019) Caribbean Sea Soundscapes: Monitoring Humpback Whales, Biological Sounds, Geological Events, and Anthropogenic Impacts of Vessel Noise. Front. Mar. Sci. 6:347. doi: 10.3389/fmars.2019.00347
Received: 27 February 2019; Accepted: 06 June 2019;
Published: 02 July 2019.
Edited by:
Christine Erbe, Curtin University, AustraliaReviewed by:
Francine Kershaw, Natural Resources Defense Council, United StatesLaura J. May-Collado, The University of Vermont, United States
Copyright © 2019 Heenehan, Stanistreet, Corkeron, Bouveret, Chalifour, Davis, Henriquez, Kiszka, Kline, Reed, Shamir-Reynoso, Védie, De Wolf, Hoetjes and Van Parijs. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sofie M. Van Parijs, c29maWUudmFucGFyaWpzQG5vYWEuZ292